mir-1246 promotes siha cervical cancer cell proliferation, invasion, and migration through...
TRANSCRIPT

1 3
Arch Gynecol ObstetDOI 10.1007/s00404-014-3260-2
GynecOlOGIc OncOlOGy
MiR‑1246 promotes SiHa cervical cancer cell proliferation, invasion, and migration through suppression of its target gene thrombospondin 2
Junying Chen · Desheng Yao · Shan Zhao · Chanjuan He · Nan Ding · Li Li · Fengyi Long
Received: 26 July 2013 / Accepted: 16 April 2014 © Springer-Verlag Berlin Heidelberg 2014
with the miR-1246 antagonist had significantly increased THBS2 expression (gray value = 12.90 ± 19.81; P = 0.037). Moreover, SiHa cells co-transfected with miR-1246 and the THBS2 3′-UTR-containing plasmid exhibited decreased luciferase enzyme activity compared with the control.Conclusion MiR-1246 induced cScc SiHa cell pro-liferation, invasion and migration. Preliminary evidence suggests that miR-1246 might promote cScc tumorigen-esis and progression by the suppression of its target gene THBS2.
Keywords cervical squamous cell carcinoma · miRnA-1246 · Thrombospondin-2 · Tumorigenesis · Tumor progression
Introduction
MicroRnAs (miRnA) are small non-coding RnAs with a length of 21–23 nucleotides. They can degrade or inhibit the expression of target genes through binding to 3′UTRs in a non-fully or fully complementary manner to prevent transcription. The identification of miRnA has been a sig-nificant breakthrough in our understanding of gene expres-sion regulation, especially in disease [1]. Studies have reported that miR-1246 is an oncogenic miRnA [2–8]. Moreover, our previous studies have shown that miR-1246 is specifically elevated in both serum and tumor tissues of cervical squamous cell carcinoma (cScc) patients with lymph node metastasis. This indicates that it could be used as a serum marker for lymph node metastasis during cScc [9]. However, the role of miR-1246 is not well understood. In this study, the biological functions of miR-1246 and its effects on a predicted target gene, thrombospondin-2
Abstract Purpose To investigate the effects of miR-1246 on prolif-eration, invasion, and migration in the human (cScc) cell line SiHa.Methods SiHa cells were assigned into three groups: miR-1246 analog; miR-1246 antagonist; and control. The MTT, transwell, and wound healing assays were performed to evaluate the proliferation, invasion, and migration abili-ties of SiHa cells, respectively. Western blot was carried out to detect protein expression of thrombospondin-2 (THBS2) before and after transfection with miR-1246 analog, antag-onist, or control. In addition, a THBS2 3′-UTR-containing dual luciferase plasmid was generated and co-transfected with miR-1246, the inhibitor, or non-specific miRnA, into SiHa cells to observe its effects on THBS2-driven lucif-erase enzyme activity.Results MTT, transwell, and wound healing assays revealed that proliferation, migration, and invasion were all significantly enhanced (P < 0.01) in SiHa cells transfected with miR-1246 analog, but were suppressed in those trans-fected with the miR-1246 antagonist. Western blot data showed that miR-1246 analog-transfected SiHa cells had significantly decreased THBS2 expression when compared with control-transfected cells (gray value = 6.28 ± 10.22 vs. 9.58 ± 17.58; P = 0.013) while those transfected
J. chen · S. Zhao · F. long Department of Gynecology, The First Affiliated Hospital of Guangxi Medical University, nanning, People’s Republic of china
D. yao (*) · S. Zhao · c. He · n. Ding · l. li Department of Gynecological Oncology, Affiliated cancer Hospital of Guangxi Medical University, nanning 530022, Guangxi, People’s Republic of chinae-mail: [email protected]

Arch Gynecol Obstet
1 3
(THBS2), were studied using cytological tests, bioinfor-matic predictions, and target protein identification. This preliminary exploration found that miR-1246 promotes the development of cervical squamous carcinoma and that this is likely mediated through regulation of its target gene, THBS2.
Materials and methods
cell culture, reagents, and equipment
The human cervical carcinoma cell line SiHa was pro-vided by the laboratory of Guangxi Medical University. It was grown in RPMI-1640 medium with 10 % fetal bovine serum (FBS) at 37 °c in a 5 % cO2 incubator. RPMI-1640 medium, FBS, glycerol, and 0.25 % trypsin were purchased from Hyclone (logan, UT, USA). MMT and DMSO were purchased from Beijing Solarbio Science & Technology co., Beijing, china. lipofectamine 2000 was purchased from Invitrogen (carlsbad, cA, USA). luciferase Assay Reagent, Stop & Glo buffer, and PlB solution were pur-chased from Promega (Madison, WI, USA). The pMD18-T vector, T4 DnA ligase, Xho1, Xbal I, exTaq polymer-ase, and DnA markers were purchased from Takara (Otsu, Japan). SuperScript II reverse transcriptase was purchased from Invitrogen. Tryptone and yeast extract were purchased from Sigma (St louis, MO, USA). Rabbit anti-human THBS2 polyclonal antibody was purchased from Beijing Biosynthesis Biotechnology co., Beijing, china. Mouse β-actin monoclonal antibody was purchased from Beyotime Institute of Biotechnology, Jiangsu, china. Fluorescence-labeled goat anti-mouse monoclonal antibody (cat. no. 926-68020) was purchased from lI-cOR (lincoln, ne, USA). Protein markers were purchased from Fermentas (Burling-ton, canada). Polyvinylidene fluoride (PVDF) membranes were purchased from Millipore (Schwalbach, Germany). The miR-1246-mimics (5′-UGUAUccUUGAAUGGA UUUUUGGAGcAGGAGUGGAcAccUGAcccAAA GGAAAUcAAUccAUAGGcUAGcAAU-3′) and miR-1246 inhibitor (5′-ccTGcTccAAAAATccATT-3′) were pur-chased from Guangzhou RiboBioco., ltd., Guangzhou, china.
cell transfection
SiHa cells were seeded in six-well plates at a density of 2 × 105 cells/well and cultured at 37 °c until they reached 60–80 % confluence. Two hours before transfection, the culture medium was changed to serum-free RPMI-1640. For each transfection reaction, RPMI-1640 culture medium (serum- and antibiotic-free) was gently mixed with the appropriate oligo at a ratio of 50:1 and incubated at RT
for 5 min. For each transfection reaction, a separate tube containing 500 μl serum- and antibiotic-free RPMI-1640 was gently mixed with 10 μl lipo2000 and incubated at RT for 5 min. next, the diluted oligo was mixed gently with the diluted lipo2000 and incubated at RT for 20 min. The transfection mix was then added to the cells and the plates were placed in the culture incubator. Six hours later, the transfection medium mixture was aspirated from the wells and replaced with serum- and antibiotic-containing medium for a further 12 h of culture in the dark. The SiHa cells were then washed with PBS three times, detached and diluted to a single-cell suspension, and analyzed by flow cytometry to assess transfection efficiency.
MTT proliferation assay
SiHa cells were trypsinized and made into single-cell sus-pensions by gentle pipetting. The cells were counted and adjusted to a concentration of 2.5 × 105 cells/ml. They were then seeded in a 96-well culture plate at 200 μl/well. cells were seeded in triplicate for each transfection group: miR-1246 mimics; miR-1246 inhibitor; and blank control, and incubated under conditions of 37 °c, 5 % cO2. After 24, 48, 72 and 96 h, 20 μl of MTT solution (5 mg/ml) was added to each well. After a further 4 h of incubation, the culture medium was aspirated carefully, to avoid washing away the blue crystals, and 150 μl DMSO was added to each well. After gentle shaking of the plate for 10 min, to help the crystals dissolve, the absorbance of each well was measured (OD490) on a microplate reader. The experiment was repeated three times and the values obtained were used to generate mean growth curves and to calculate the cell proliferation rate by the following formula.
cell proliferation rate (%) = mean OD value of experi-mental group/mean OD value of control × 100.
Scratch assay
SiHa cells were seeded in 24-well plates at 70 % conflu-ence and cultured until they formed a monolayer that occu-pied 100 % of the surface area. next, a linear wound was made by scratching the monolayer with a 200 μl pipette tip. The cells were cultured for a further 24 h before the medium was removed and hematoxylin-and-eosin (H&e) staining was performed. Images were captured and the gap size was measured. The experiment was repeated three times.
cell invasion assay
Matrigel was melted at 4 °c overnight; it was then mixed with RPMI-1640 culture medium at a ratio of 1:6. The inserts of 24-well Transwell plates were coated with

Arch Gynecol Obstet
1 3
60 μl of the diluted Matrigel and then dried completely in the incubator. Seventy-two hours after transfection with mimics or the antagonist, SiHa cells were detached and adjusted to a concentration of 5 × 104/ml and 200 μl of this cell suspension was seeded into the top chamber of the Matrigel-coated Transwells. RPMI-1640 complete medium (containing 20 % FBS) was added to the bottom chamber. After 15 h, cells were removed from the membrane and fixed with 95 % ethanol for 30 min. next, they were H&e-stained and photographed, and the number of invading cells was counted. The experiment was repeated three times for each group.
cell migration assay
Apart from not adding Matrigel over the insert membrane in the Transwell, all other procedures were the same as those described for the cell invasion assay.
MiR-1246 target gene selection
MicroRnA target genes were selected using a microRnA database and websites. Several target prediction programs, including DIAnAmT, miRanda, miRDB, miRWalk, PIc-TAR4, PIcTAR5, PITA, RnA22, and Targetscan, were used. Also, GenBank was searched for target gene candi-dates that are particularly related to cancer cell invasion and metastasis. Taking into account the changes in biologi-cal activity observed in SiHa cells transfected with miR-1246, THBS2 was selected for further investigation.
Western blotting
Seventy-two hours after transfection, protein samples were extracted from SiHa cells and stored at −20 °c until use. To separate proteins, 8 % separating/5 % stacking gels were prepared. electrophoresis was performed at 80 V until the samples entered the separating gels and then continued at 100–120 V for approximately 60–90 min. Samples were then transferred to a PVDF membrane which was subse-quently blocked in 5–7 % skim milk for 1.5 h. The gel por-tion containing the reference protein was also transferred to a PVDF membrane and blocked. The PVDF membrane containing the target protein was then immersed in primary antibody diluted 1:200 with skim milk in a total volume of 5 ml, according to the manufacturer’s instructions, and incubated overnight at 4 °c. The dilution of the primary antibody for the reference PVDF membrane was 1:10,000 in a total volume of 10 ml. next, the target and reference membranes were incubated with a secondary antibody diluted at 1:10,000 in a total volume of 5 ml at RT for 1 h. The membranes were then washed with TBST for 5 min and immunoreactive bands were visualized with an infrared
image processing system to analyze the molecular weight and optical density.
construction of the double fluorescein enzyme plasmid containing the THBS2 3′-UTR
Based on the calculations of the target prediction soft-ware, possible binding sites of miR-1246 on THBS2 (nM_003247, 5,826 bp in length) included the 291, 1,030, 1,225, 1,322, 1,523, 1,559, 1,570, 1,577 and 1,620 regions. The design and synthesis of reporter constructs containing the 3′-UTR nucleic acid sequence of THBS2 were performed by adding Xho1 restriction endonuclease sites at the 5′-end of the sequence (forward primer: AAA GAGccGGATGTAcGTGG; reverse primer: ccTTTG cAATTcTTGccccc) to produce a fragment of 876 bp that covered the sequence 806–1,681, and contained eight potential miR-1246 binding sites.
A clear, specific band was observed from the PcR-amplified fragment which was subsequently ligated into the pMDl8T vector and inserted into the pmirGlO plasmid. The recombinant plasmid sequencing results were then compared with BlAST. The resulting sequence was the 3′-UTR sequence representing the region 806–1,681 of the THBS2 gene.
luciferase reporter gene detection
SiHa cells were co-transfected with the expression vector and miR-1246 mimics (experimental) or the expression vector and a non-specific miRnA, miR-16 mimics (con-trol). Ten replicates were prepared for each test condition. Forty-eight hours after transfection, the activity of luc2 was determined using the dual luciferase assay reagents where Renilla luciferase activity was measured as an internal reference.
Statistical analysis
All data were processed and analyzed using the SPSS soft-ware (ver. 16.0). P values of <0.05 were considered to indi-cate statistical significance. The t test was used to compare the mean values between two samples. Analysis of vari-ance (AnOVA) was used to compare the means of multiple samples.
Results
Determination of cell transfection efficiency
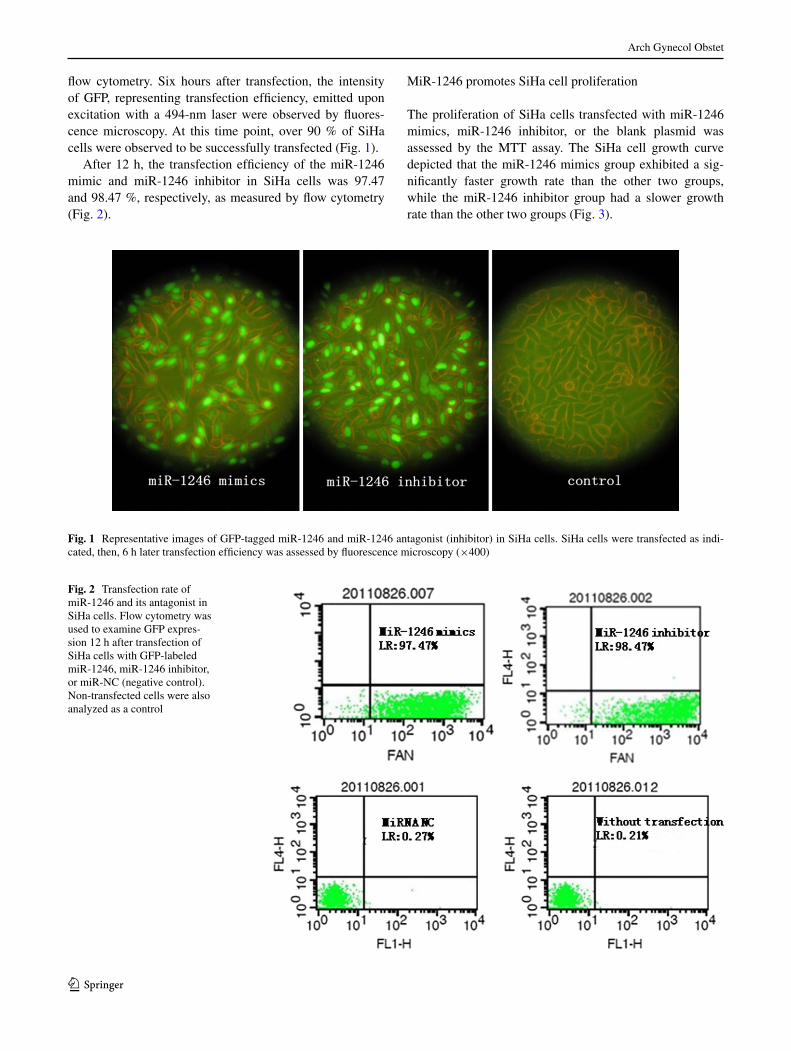
The transfection efficiency of GFP-tagged miRnA mim-ics and antagonist were first assessed by microscopy and
Arch Gynecol Obstet
1 3
flow cytometry. Six hours after transfection, the intensity of GFP, representing transfection efficiency, emitted upon excitation with a 494-nm laser were observed by fluores-cence microscopy. At this time point, over 90 % of SiHa cells were observed to be successfully transfected (Fig. 1).
After 12 h, the transfection efficiency of the miR-1246 mimic and miR-1246 inhibitor in SiHa cells was 97.47 and 98.47 %, respectively, as measured by flow cytometry (Fig. 2).
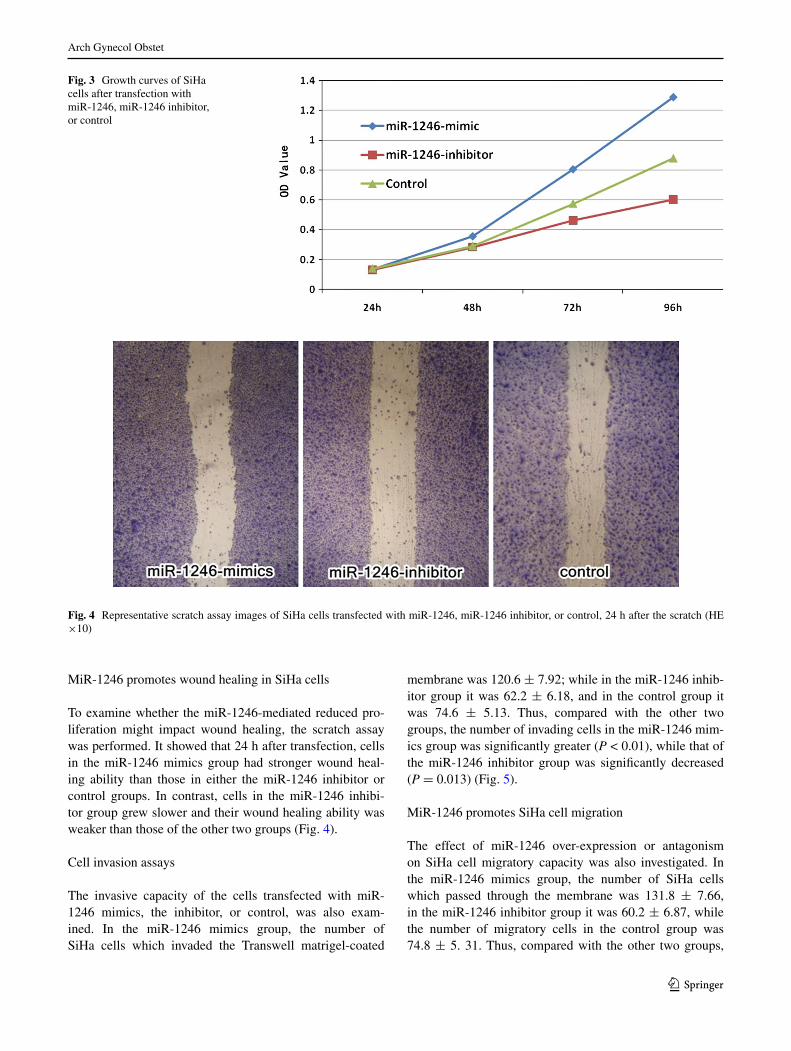
MiR-1246 promotes SiHa cell proliferation
The proliferation of SiHa cells transfected with miR-1246 mimics, miR-1246 inhibitor, or the blank plasmid was assessed by the MTT assay. The SiHa cell growth curve depicted that the miR-1246 mimics group exhibited a sig-nificantly faster growth rate than the other two groups, while the miR-1246 inhibitor group had a slower growth rate than the other two groups (Fig. 3).
Fig. 1 Representative images of GFP-tagged miR-1246 and miR-1246 antagonist (inhibitor) in SiHa cells. SiHa cells were transfected as indi-cated, then, 6 h later transfection efficiency was assessed by fluorescence microscopy (×400)
Fig. 2 Transfection rate of miR-1246 and its antagonist in SiHa cells. Flow cytometry was used to examine GFP expres-sion 12 h after transfection of SiHa cells with GFP-labeled miR-1246, miR-1246 inhibitor, or miR-nc (negative control). non-transfected cells were also analyzed as a control
Arch Gynecol Obstet
1 3
MiR-1246 promotes wound healing in SiHa cells
To examine whether the miR-1246-mediated reduced pro-liferation might impact wound healing, the scratch assay was performed. It showed that 24 h after transfection, cells in the miR-1246 mimics group had stronger wound heal-ing ability than those in either the miR-1246 inhibitor or control groups. In contrast, cells in the miR-1246 inhibi-tor group grew slower and their wound healing ability was weaker than those of the other two groups (Fig. 4).
cell invasion assays
The invasive capacity of the cells transfected with miR-1246 mimics, the inhibitor, or control, was also exam-ined. In the miR-1246 mimics group, the number of SiHa cells which invaded the Transwell matrigel-coated
membrane was 120.6 ± 7.92; while in the miR-1246 inhib-itor group it was 62.2 ± 6.18, and in the control group it was 74.6 ± 5.13. Thus, compared with the other two groups, the number of invading cells in the miR-1246 mim-ics group was significantly greater (P < 0.01), while that of the miR-1246 inhibitor group was significantly decreased (P = 0.013) (Fig. 5).
MiR-1246 promotes SiHa cell migration
The effect of miR-1246 over-expression or antagonism on SiHa cell migratory capacity was also investigated. In the miR-1246 mimics group, the number of SiHa cells which passed through the membrane was 131.8 ± 7.66, in the miR-1246 inhibitor group it was 60.2 ± 6.87, while the number of migratory cells in the control group was 74.8 ± 5. 31. Thus, compared with the other two groups,
Fig. 3 Growth curves of SiHa cells after transfection with miR-1246, miR-1246 inhibitor, or control
Fig. 4 Representative scratch assay images of SiHa cells transfected with miR-1246, miR-1246 inhibitor, or control, 24 h after the scratch (He ×10)

Arch Gynecol Obstet
1 3
the migratory capacity of the miR-1246 mimics group was significantly greater (P < 0.01), while that of the miR-1246 inhibitor group was significantly lower (P < 0.01).
MiR-1246 impairs THBS2 protein expression
To begin to investigate how miR-1246 affects SiHa cells proliferation, migration and invasion, the expression of its predicted target gene, THBS2, was examined by western blot and quantified by densitometry. The protein expres-sion level of THBS2 in the miR-1246 inhibitor group was significantly lower than that of the control group (gray value = 6.28 ± 10.22 vs. 9.58 ± 17.58; P = 0.013), while that of the miR-1246 inhibitor group was significantly greater (gray value = 12.9 ± 19.81; P = 0.037). These data suggest that the miR-1246 analog may down-regulate the expression of THBS2 protein in SiHa cells (Fig. 6).
MiR-1246 impairs the transcriptional activity THBS2
To further examine whether miR-1246 affects the transcrip-tional activity of THBS2, luciferase reporter assays were performed using a THBS2 reporter construct (pmirGlO). In SiHa cells co-transfected with miR-1246 and pmirGlO, the relative luciferase expression was 0.51 ± 0.08, that of a non-specific miRnA, miR-16, and pmirGlO was 0.92 ± 0.04, and that of the pmirGlO alone group was 0.97 ± 0.03. In short, significantly lower expression of luciferase was observed in SiHa cells co-transfected with miR-1246 com-pared with that of either the non-specific miRnA or the con-trol (P < 0.01). This result suggests that specific targets are present in the 3′-UTR region (806–1,681) of THBS2 which might be specifically bound by miR-1246 to negatively affect its transcriptional activity (Fig. 7).
Discussion
MiR-1246 (sequence: AAUGGAUUUUUGGAGcAGG) was first identified during the sequencing of human stem
Fig. 5 Representative images of invasive SiHa cells transfected as indicated (He ×400)
Fig. 6 Protein expression of THBS2 in SiHa cells 72 h after transfec-tion as indicated (WB)
Fig. 7 effect of miR-1246 on THBS2-driven luc2 relative expres-sion. luciferase activity was measured in SiHA cells transfected with a THBS2 luciferase reporter construct (pmirGlO) alone or together with either miR1246 or miR-16. Data represent mean luciferase activity normalized to that of Renilla ± SD; **P < 0.01

Arch Gynecol Obstet
1 3
cells in 2008 [10]. Though only a few studies have reported an association between miR-1246 and tumorigenesis, it has been recognized as an oncogenic miRnA. Baraniskin [11] suggested that miR-1246 is a serum tumor marker for pancreatic ductal adenocarcinoma and colon cancer, while Pigati and colleagues [2] reported that miR-1246 is associated with breast cancer. However, how miR-1246 is involved in cancer development and progression has been largely unexplained. Recently, the oncogenic role of miR-1246 during tumorigenesis has been further explored. In 2012, liao and Zhang each reported that miR-1246 triggers the activation of VeGF (vascular endothelial growth factor) signaling via the regulation of p53 and DyRK1A (Down’s syndrome-associated protein kinase) and thereby induces tumor angiogenesis and metastasis [4, 7] [12]. Pigati [2] further found that miR-1246 is selectively released from breast cancer cells, and this selective release is highly associated with the degree of cell malignancy. Addition-ally, Baraniskin [11] proposed that serum miR-1246 might come from RnU2-1f (U2 small nuclear RnA fragment), a tumor-associated nucleotide that is very stable in serum. These findings suggest that the level of serum miR-1246 might not directly reflect variations of intracellular miRnA level. They also highlight that the role of miR-1246 in tumor metastasis deserves further investigation.
To this end, our previous study showed that miR-1246 is specifically elevated in both serum and tumor tissues in cScc patients with lymph node metastasis [9], consistent with that of Takeshita [13] who reported that miR-1246 in identify-ing patients with esophageal squamous cell carcinoma from healthy individuals, indicating that it could be used as a serum marker for lymph node metastasis during cScc.
In this study, to further clarify the role of miR-1246 in promoting cScc tumorigenesis and progression, we trans-fected the cScc cell line SiHa with miR-1246 analog, or an antagonist, and assessed cell proliferation, migration and invasion using the MTT, transwell, and wound heal-ing assays, respectively. Our results showed that, compared with control cells, miR-1246 analog-transfected SiHa cells had a significantly increased growth rate as well as elevated migratory and invasive abilities. In contrast, the growth rate of cells transfected with miR-1246 antagonist was reduced, as was their invasive and migratory capacities. These find-ings suggest that miR-1246 enhances SiHa cell growth, invasion and migration, and may thus function as an onco-gene in cScc development and progression.
The identification of miR-1246 target genes provides new insight to study the mechanisms underlying its role in cancer development. Using bioinformatics tools and biological experiments, we identified THBS2 as a miR-1246 target gene. To confirm this result, we compared the expression of THBS2 in SiHa cells transfected with either the miR-1246 analog or antagonist. The results showed that
when compared with the control, the expression level of THBS2 was significantly decreased in miR-1246 analog-transfected SiHa cells, while it was significantly increased in miR-1246 antagonist-transfected SiHa cells. This indi-cates that miR-1246 inhibits THBS2 expression. The dual luciferase enzyme assay further revealed that miR-1246 interfered with the transcriptional activity of the THBS2 3′-UTR (806–1,681), suggesting that the miR-1246 binding site may be located in the 3′-UTR of THBS2. These results therefore confirm that THBS2 is a miR-1246 target gene.
THBS2 is a member of the thrombospondin family, which consists of various cell–cell or cell–interstitial gly-coproteins that are capable of regulating cell adhesion and migration. The THBS family can be divided into two subgroups, A and B, based on molecular structure. It is generally believed that THBS1 can both inhibit and pro-mote angiogenesis, while THBS2 is a tumor suppressor with high expression in fibroblasts and low expression in endothelial cells; it has also been shown to inhibit tumor cell growth and angiogenesis [14, 15]. Some studies have reported that THBS2 is specifically expressed in various cancers including breast cancer, colon cancer and ovarian cancer. Moreover, since it can regulate the hydrolysis of the ecM, THBS2 could be used as an indicator of tumor invasion as well as of the outcome of neoadjuvant therapy [16, 17]. Indeed, the mechanisms by which THBS2 inhib-its angiogenesis are mainly associated with matrix metal-loproteinases (MMP) and the ecM [15–18]. yang [19] has reported that the lack of THBS2 induces MMP-2 overex-pression and cell–matrigel degradation. Hirose [20] also demonstrated that decreased THBS2 affects the levels of MMP-2 and MMP-9, and that the THBS2–MMP interac-tion directly modulates the metabolism of ecM. currently, only a few studies have reported an association between THBS2 and cScc. We hypothesize that THBS2 inhibits tumor invasion and angiogenesis in cScc through regula-tion of the ecM receptor pathway, whereas over-expres-sion of miR-1246 may counteract these effects through THBS2 inhibition, but this shall be verified in further stud-ies. In this study, we confirm that miR-1246 induces SiHa cell growth, enhances cell invasion and migration, and sup-presses THBS2 expression. Thus, we speculate that inhibi-tion of THBS2 expression is a mechanism by which miR-1246 promotes cScc tumorigenesis and progression.
Acknowledgments The present study was supported by grants from the natural Science Foundation of Guangxi Zhuang Autonomous Region, china (2011GXnSFA018184, 2013GXnSFBA019130, 2013GXnSFBA019132. www.gxsti.net), and the Public Health self-financing project research project of Guangxi Zhuang Autonomous Region (Z2012071).
Conflict of interest The authors declare that they have no conflict of interest.

Arch Gynecol Obstet
1 3
References
1. Medina PP, nolde M, Slack FJ (2010) OncomiR addiction in an in vivo model of microRnA-21-induced pre-B-cell lymphoma. nature 467:86–90
2. Pigati l, yaddanapudi Sc, Iyengar R, Kim DJ, Hearn SA, Dan-forth D, Hastings Ml, Duelli DM (2010) Selective release of microRnA species from normal and malignant mammary epithe-lial cells. PloS One 5:e13515
3. Gillen Ae, Gosalia n, leir SH, Harris A (2011) MicroRnA reg-ulation of expression of the cystic fibrosis transmembrane con-ductance regulator gene. Biochem J 438:25–32
4. Zhang y, liao JM, Zeng SX, lu H (2011) p53 downregulates Down syndrome-associated DyRK1A through miR-1246. eMBO Rep 12:811–817
5. Jaiswal R, luk F, Gong J, Mathys JM, Grau Ge, Bebawy M (2012) Microparticle conferred microRnA profiles––implications in the transfer and dominance of cancer traits. Mol cancer 11:37
6. Jones cI, Zabolotskaya MV, King AJ, Stewart HJ, Horne GA, chevassut TJ, newbury SF (2012) Identification of circulating microRnAs as diagnostic biomarkers for use in multiple mye-loma. Br J cancer 107:1987–1996
7. liao JM, Zhou X, Zhang y, lu H (2012) MiR-1246: a new link of the p53 family with cancer and Down syndrome. cell cycle 11:2624–2630
8. Piepoli A, Tavano F, copetti M, Mazza T, Palumbo O, Panza A, di MFF, Pazienza V, Mazzoccoli G, Biscaglia G, Gentile A, Mas-trodonato n, carella M, Pellegrini F, di SP, Andriulli A (2012) Mirna expression profiles identify drivers in colorectal and pan-creatic cancers. PlOS One 7:e33663
9. chen J, yao D, li y, chen H, He c, Ding n, lu y, Ou T, Zhao S, li l, long F (2013) Serum microRnA expression levels can pre-dict lymph node metastasis in patients with early-stage cervical squamous cell carcinoma. Int J Mol Med 32:557–567
10. Morin RD, O’connor MD, Griffith M, Kuchenbauer F, Delaney A, Prabhu Al, Zhao y, McDonald H, Zeng T, Hirst M, eaves cJ, Marra MA (2008) Application of massively parallel sequencing to microRnA profiling and discovery in human embryonic stem cells. Genome Res 18:610–621
11. Baraniskin A, nopel-Dunnebacke S, Ahrens M, Jensen SG, Zoll-ner H, Maghnouj A, Wos A, Mayerle J, Munding J, Kost D, Rein-acher-Schick A, liffers S, Schroers R, chromik AM, Meyer He, Uhl W, Klein-Scory S, Weiss FU, Stephan c, Schwarte-Waldhoff I, lerch MM, Tannapfel A, Schmiegel W, Andersen cl, Hahn SA (2013) circulating U2 small nuclear RnA fragments as a novel diagnostic biomarker for pancreatic and colorectal adenocarci-noma. Int J cancer 132:e48–e57
12. Baek KH, Zaslavsky A, lynch Rc, Britt c, Okada y, Siarey RJ, lensch MW, Park IH, yoon SS, Minami T, Korenberg JR, Folkman J, Daley GQ, Aird Wc, Galdzicki Z, Ryeom S (2009) Down’s syndrome suppression of tumour growth and the role of the calcineurin inhibitor DScR1. nature 459:1126–1130
13. Takeshita n, Hoshino I, Mori M, Akutsu y, Hanari n, yoney-ama y, Ikeda n, Isozaki y, Maruyama T, Akanuma n, Komatsu A, Jitsukawa M, Matsubara H (2013) Serum microRnA expres-sion profile: miR-1246 as a novel diagnostic and prognostic bio-marker for oesophageal squamous cell carcinoma. Br J cancer 108:644–652
14. czekierdowski A, czekierdowska S, Danilos J, czuba B, Sodo-wski K, Sodowska H, Szymanski M, Kotarski J (2008) Microves-sel density and cpG island methylation of the THBS2 gene in malignant ovarian tumors. J Physiol Pharmacol 59(Suppl 4):53–65
15. Hirose y, chiba K, Karasugi T, nakajima M, Kawaguchi y, Mikami y, Furuichi T, Mio F, Miyake A, Miyamoto T, Ozaki K, Takahashi A, Mizuta H, Kubo T, Kimura T, Tanaka T, Toyama y, Ikegawa S (2008) A functional polymorphism in THBS2 that affects alternative splicing and MMP binding is associated with lumbar-disc herniation. Am J Hum Genet 82:1122–1129
16. Kim H, Watkinson J, Varadan V, Anastassiou D (2010) Multi-can-cer computational analysis reveals invasion-associated variant of desmoplastic reaction involving InHBA, THBS2 and cOl11A1. BMc Med Genomics 3:51
17. Wang y, Fu W, Xie F, Wang y, chu X, Wang H, Shen M, Wang y, Wang y, Sun W, lei R, yang l, Wu H, Foo J, liu J, Jin l, Huang W (2010) common polymorphisms in ITGA2, POn1 and THBS2 are associated with coronary atherosclerosis in a can-didate gene association study of the chinese Han population. J Hum Genet 55:490–494
18. Muth cA, Steinl c, Klein G, lee-Thedieck c (2013) Regulation of hematopoietic stem cell behavior by the nanostructured pres-entation of extracellular matrix components. PloS One 8:e54778
19. yang Z, Kyriakides TR, Bornstein P (2000) Matricellular pro-teins as modulators of cell-matrix interactions: adhesive defect in thrombospondin 2-null fibroblasts is a consequence of increased levels of matrix metalloproteinase-2. Mol Biol cell 11:3353–3364
20. Rodder S, Scherer A, Korner M, eisenberger U, Hertig A, Raulf F, Rondeau e, Marti HP (2010) Meta-analyses qualify metzincins and related genes as acute rejection markers in renal transplant patients. Am J Transplant 10:286–297