micrornas in viral replication and pathogenesis
TRANSCRIPT

MicroRNAs in Viral Replication and Pathogenesis
DEREK M. DYKXHOORN
ABSTRACT
MicroRNAs (miRNAs) are an important class of small, noncoding, regulatory RNAs found to be involved in
regulating a wide variety of important cellular processes by the sequence-specific inhibition of gene expres-
sion. Viruses have evolved a number of mechanisms to take advantage of the regulatory potential of this highly
conserved, ubiquitous pathway known as RNA interference (RNAi). This review will focus on the recent efforts
to understand the complex relationship between vertebrate viruses and the RNAi pathway, as well as the role
of silencing pathways in the inhibition of pathogenic genetic elements, including transposons and retrotrans-
posons.
INTRODUCTION
M ICRORNAS (MIRNAS) REPRESENT A CLASS OF small RNA
molecules that serve important regulatory functions in a
variety of cellular processes, including differentiation, develop-
ment, and metabolism (Ambros, 2004; Bartel, 2004; Chen et al.,
2004; Poy et al., 2004; Bentwich et al., 2005). In recent years,
there has been considerable interest in the roles they play in the
regulation of gene expression. RNA interference (RNAi) is a
highly conserved, ubiquitous gene regulatory mechanism that
uses small, noncoding RNA species to direct the sequence-spe-
cific silencing of gene expression. RNAi-mediated silencing has
been shown to play an important role in the antiviral defense
systems of plants and insects (e.g., Drosophila melanogaster)
(Waterhouse et al., 2001; Keene et al., 2004; Sanchez-Vargas
et al., 2004; Voinnet, 2005; Galiana-Arnoux et al., 2006; van Rij
et al., 2006; Wang et al., 2006). This defense system is based on
the recognition and endonucleolytic cleavage of long double-
stranded RNAs (dsRNAs), produced as a consequence of the
replication of many viral pathogens, into small interfering RNAs
(siRNAs) by the RNase III-type enzyme Dicer. These virus-
specific siRNAs are incorporated into the RNA-induced silencing
complex (RISC) and direct the cleavage of the cognate viral mes-
senger RNAs (mRNAs), resulting in the suppression of viral in-
fection. As the vertebrate immune system can effectively respond
to virally encoded long dsRNAs, a much more complex rela-
tionship between vertebrate viruses and the RNAi pathway has
evolved.
Most miRNAs are transcribed from RNA polymerase II
promoters, allowing their expression to be regulated in a cell-
and tissue-specific manner, similar to protein-coding mRNAs.
miRNAs can be expressed individually (monocistronic ex-
pression) or as part of a cluster (di- or polycistronic expres-
sion), and can be encoded in the introns of protein-coding or
noncoding genes or as independent transcriptional units (Lagos-
Quintana et al., 2001; Lau et al., 2001; Lee et al., 2002, 2004;
Cai et al., 2004; Rodriguez et al., 2004). These small RNA
molecules are expressed from highly structured precursor tran-
scripts, termed primary miRNAs (pri-miRNAs), that undergo
a series of cleavage reactions to produce the mature, *22-
nucleotide (nt) miRNAs (Fig. 1) (Lee et al., 2002; Kim, 2005).
The miRNA-encoding stem-loop structure is recognized and
cleaved by the RNase III enzyme Drosha, in conjunction with
its binding partner DiGeorge syndrome critical region gene 8
(DGCR8, or Pasha in Drosophila and C. elegans) (Lee et al.,
2003; Zeng et al., 2005; Han et al., 2006; Yeom et al., 2006).
This cleavage produces a short stem-loop RNA of *60 nts,
termed the precursor miRNA (pre-miRNA), which is trans-
ported out of the nucleus and into the cytoplasm by exportin 5
(Yi et al., 2003; Lund et al., 2004). Once in the cytoplasm, the
pre-miRNA is recognized by Dicer, which works in concert
with human immunodeficiency virus (HIV-1) trans-activating
response (TAR) RNA-binding protein (TRBP, or Loquacious
in Drosophila) to cleave the pre-miRNA into a short dsRNA
molecule containing the miRNA and the accompanying pas-
senger strand (Tomari et al., 2004; Chendrimada et al., 2005;
Forstemann et al., 2005; Gregory et al., 2005; Haase et al., 2005;
Saito et al., 2005; Tomari and Zamore, 2005). In most cases,
the passenger strand is rapidly dissociated and degraded while
the mature miRNA is taken up into the effector complex, the
The CBR Institute for Biomedical Research, Harvard Medical School, Boston, Massachusetts.
DNA AND CELL BIOLOGYVolume 26, Number 4, 2007# Mary Ann Liebert, Inc.Pp. 239–249DOI: 10.1089/dna.2006.0559
239

miRNA-containing RISC (miRISC) (Khvorova et al., 2003;
Schwarz et al., 2003; Matranga et al., 2005). The miRISC di-
rects the silencing of mRNAs containing sequences comple-
mentary to the miRNA through 2 possible mechanisms, mRNA
cleavage or translational inhibition (Ambros, 2004; Bartel,
2004; Meister et al., 2004). The choice of mechanism depends
on the overall level of complementarity between the miRNA
and the target site on the mRNA. Early evidence suggested
that target sequences with imperfect complementarity were si-
lenced by the inhibition of translation while those with perfect
complementarity led to the cleavage and degradation of the
target mRNA (Doench et al., 2003). However, more recent
studies have found that even miRNAs with limited homol-
ogy could direct the cleavage of their target mRNAs (Bagga
et al., 2005; Pasquinelli et al., 2005; Massirer and Pasquinelli,
2006).
Transcription
Drosha/DGCR8
Exportin 5
(A)N(A)N
(A)N
(A)N
(A)N
Dicer/TRBP
A
B
pri-miRNA
pre-miRNA
miRNA
transient doublestranded miRNA
FIG. 1. The miRNA pathway. The miRNAs are expressed as long primary transcripts called pri-miRNAs containing a char-acteristic hairpin RNA secondary structure that is cleaved by the RNase III-type endonuclease Drosha with the dsRNA-bindingprotein DGCR8, forming a short hairpin RNA termed the pre-miRNA (Lee et al., 2005). The pre-miRNA is translocated to thecytoplasm by exportin 5, where it is cleaved by Dicer (partnered with TRBP), producing a short-lived double-stranded miRNAintermediate containing the mature miRNA and the accompanying passenger strand (Bohnsack et al., 2004). The passenger strandis rapidly dissociated from the mature miRNA, which is incorporated into the ribonucleoprotein effector complex miRISC. Thesequence of the mature miRNA serves to determine the specificity of the silencing complex. The miRISC can silence through oneof two mechanisms: (A) the inhibition of translation or (B) mRNA cleavage. In general, target sites with a limited degree ofcomplementarity induce translational inhibition, while those with a high degree of complementarity lead to mRNA cleavage anddegradation.
240 DYKXHOORN

miRNAs have several characteristics that make them an at-
tractive option for viruses to utilize in the regulation of gene
expression. Due to their small size, miRNAs can be easily
encoded in viral genomes, where space is of a premium. In
addition, miRNAs are nonimmunogenic and take advantage
of a ubiquitous, endogenous host regulatory mechanism. Fur-
ther, a single miRNA has the potential to alter the expression
pattern of a large number of genes (Lim et al., 2005). miRNAs
could be envisioned to function in viral pathogenesis in sev-
eral ways, including the regulation of viral gene expression by
host miRNAs, the regulation of viral gene expression by virus-
encoded miRNAs, and the regulation of host genes by virus-
encoded miRNAs. In addition, some viruses, particularly in
plants and lower eukaryotes, have been shown to encode fac-
tors (RNAs and proteins) that can inhibit the endogenous RNAi
pathway. This review will focus on the complex relationship
between vertebrate viruses and the RNAi pathway, as well as
the role of silencing pathways in the inhibition of pathogenic
genetic elements including transposons and retrotransposons.
VIRUS-ENCODED MIRNAS
Herpesviruses and miRNAs
Although the prototypical miRNA, lin-4, was discovered in
the early 1980s as a noncoding RNA whose loss could lead to
distinct developmental and morphological abnormalities, it was
the landmark discovery of the silencing potential of siRNAs that
helped elucidate the silencing capacity of small RNA molecules
(Horvitz and Sulston, 1980; Chalfie et al., 1981; Sulston and
Horvitz, 1981; Tuschl et al., 1999; Zamore et al., 2000). These
siRNAs were shown to have distinct structural features, a du-
plexed RNA of *19–21 nts with a phosphate group at each 50
terminus and a 30 overhang of 2–3 nts, reminiscent of cleavage
products from RNase III-type enzymes (Elbashir et al., 2001).
The biochemical characterization of siRNAs led to the rapid
identification of Dicer as the RNase III-type enzyme responsi-
ble for their production (Bernstein et al., 2001). This began the
elucidation of the RNAi pathway through which both miRNAs
and siRNAs, produced by Dicer cleavage, induced posttran-
scriptional gene silencing. Taking advantage of the unique struc-
tural features of siRNAs and miRNAs, methods were developed
for the cloning of small regulatory RNAs from a variety of or-
ganisms, cells, and tissues (Djikeng et al., 2001; Elbashir et al.,
2001; Lagos-Quintana et al., 2001, 2002, 2003; Aravin et al.,
2003; Houbaviy et al., 2003; Ambros and Lee, 2004). These
cloning procedures demonstrated the complexity of the small
regulatory RNA world and facilitated the development of
computer algorithms for the in silico identification of miRNAs.
The same cloning protocols were used to identify and charac-
terize miRNAs, repeat-associated siRNAs (rasiRNAs, small
RNA molecules that are produced from repetitive DNA ele-
ments including retrotransposons and transposons), and, more
recently, Piwi-interacting RNAs (piRNAs, small RNAs com-
monly found in germline cells that associate with members of
the Piwi family of Argonaute proteins) (Djikeng et al., 2001;
Elbashir et al., 2001; Aravin et al., 2003).
It did not take long before these cloning protocols were used
to identify miRNAs produced from virus-encoded sequences.
In the first published report, Pfeffer et al. (2004) identified
miRNAs from Epstein Barr virus (EBV, also called human
herpesvirus-4 (HHV-4)), a member of the g–herpesvirus family
that preferentially infects B cells. These miRNAs were found
in 2 clusters within the EBV genome. The first and largest
cluster, containing 14 miRNAs, was mapped to the intronic
regions of the BamHI-A region rightward transcript (BART)
gene (BART miRNA-1 to miRNA-14). The second cluster,
containing the 3 remaining miRNA sequences, was located
within the 50 and 30 untranslated regions (UTRs) of the BamHI
fragment H rightward open reading frame 1 (BHRF1) gene
(Pfeffer et al., 2004; Cai et al., 2006). The function of these
EBV-encoded miRNAs remains unknown. Scanning the hu-
man genome for potential target sites for the EBV-encoded
miRNAs has given a list of genes involved in the regulation of
B cell proliferation and apoptosis, B cell–specific chemokines,
transcriptional regulators, and components of signal trans-
duction pathways. The analysis of cell lines latently infected with
EBV has shown specific patterns of EBV miRNA expression.
These cell lines can be differentiated based on the 3 different
stages of latency (stages I to III), as defined by the expression of
specific sets of viral proteins. The profiling of miRNA expression
from cell lines representing different stages of latency showed the
expression of high levels of BART miRNAs in all stages of la-
tency, as well as lytic infections. On the other hand, the BHRF1
miRNAs were highly expressed in latency stage III cell lines
but were almost undetectable in latency stages I and II (Cai
et al., 2006). Interestingly, 1 of the EBV-encoded miRNAs, miR-
BART2, can target the cleavage of the EBV BALF5gene that
encodes the viral DNApolymerase (Furnari et al., 1993; Pfeffer et
al., 2005). MiR-BART2 overlaps with, and is expressed in the
antisense orientation to, the 30 UTR of the EBVBALF5gene, and
a truncated BALF5mRNAhas been identified that corresponds to
the product of amiR-BART2-directed cleavage reaction. The role
that this autoregulatory mechanism plays in the life cycle of EBV
remains to be determined.
Using both biochemical and computational approaches, miR-
NAs have been identified in a number of other members of the
herpesvirus family. Ten miRNAs have been identified by cloning
of small RNA molecules and northern blot analysis in Kaposi’s
sarcoma-associated virus (KSHV, or HHV-8) (g-herpesvirus), 9miRNAs frommurine herpesvirus-68 (MHV-68) (g-herpesvirus),and 9 miRNAs from human cytomegalovirus (HCMV, or HHV-
5) (b-herpesvirus) (Cai et al., 2005; Grey et al., 2005; Pfeffer
et al., 2005; Samols et al., 2005). However, computational
analysis failed to identify miRNAs from the genomes of several
other herpesviruses; the �-herpesvirus HHV-3 (varicella-zoster
virus) or the b-herpesvirus HHV-6 and HHV-7. Interestingly,
there was a decided lack of conservation in both sequence and
position in themiRNAs identified by Pfeffer et al. (2005) from the
different herpesvirus genomes. This suggests that the miRNAs
expressed from these different herpesviruses are not involved in
core functions of the virus (viral replication, viral gene expres-
sion, etc.) but have evolved independently to allow each virus to
adapt to the specific cell types in which the virus persists. How-
ever, the comparison of 2 more closely related herpesviruses,
EBV and Rhesus lymphocryptovirus (rLCV), both members of
the lymphocryptovirus genus of herpesviruses, led to the bio-
chemical identification of 7 highly conserved miRNAs between
them (Cai et al., 2006). These conserved miRNAs were present
MICRORNAS IN VIRAL REPLICATION AND PATHOGENESIS 241

in both the BHRF1 and BART1 miRNA clusters of the viral
genomes. The high degree of miRNA conservation found be-
tween these 2 viruses may be reflective of the similar cellular
environments in which these viruses replicate.
Viral miRNAs that regulate host gene
Every virus is dependent, to one degree or other, on the
cellular machinery to complete the viral life cycle. Therefore,
complex viral-host interactions have evolved to ensure the
fitness and survival of the virus. The discovery of the regula-
tory potential of small RNA molecules has added another facet
to this complex relationship. The inhibition of apoptosis is a
common strategy used by viruses to ensure their survival. The
expression of a noncoding RNA in herpes simplex virus-1
(HSV-1) has been associated with the maintenance of viral
latency in neuronal cells (Ahmed et al., 2002; Gupta et al.,
2006). In fact, this transcript, appropriately termed the latency-
associated transcript (LAT), is the only viral gene expressed
during latency ( Jones, 2003). Computational analysis predicted
that a stem-loop structure, reminiscent of the structure of pre-
miRNAs, could be formed in exon 1 of the LAT gene (Fig. 2)
(Inman et al., 2001; Ahmed et al., 2002; Gupta et al., 2006).
The cloning of small RNA molecules from SY5Y cells trans-
fected with a complementary DNA (cDNA) expressing the
HSV-1 LAT gene demonstrated that this region encoded a
miRNA (Gupta et al., 2006). Expression of these small RNA
molecules was confirmed by northern blot analysis. Cells that
express the LAT gene have been shown to be refractory to
cisplatin-induced apoptosis but only in the context of a func-
tioning RNAi pathway. The siRNA-mediated silencing of
Dicer, which is necessary for the formation of mature miRNAs,
RLRS
RS
U S
RL
UL
LA
T
LAT
HSV-1 genome
LAT
5'-CCGUGGCGGCCCGGCCCGGGGCCCC
5'-UGGCGGCCCGGCCCGGGGCC-3'
GG C G
GA
CCCA
A3'-AACACCCCGGGGCCCGGCCCCGGGG
AN7MeGppp SMAD3
7MeGppp ANTGF-β
7MeGpppSMAD
3
AN
7MeGpppTGF-β
AN
Apoptosis
Dicer processing
Drosha processing
mRNA recognition
mRNA cleavage
pre-miRNA-LAT
miRNA-LAT
A B
FIG. 2. Antiapoptotic effects of the HSV-1 LAT miRNA. (A) The HSV-1 genome is a linear double-stranded DNAmolecule thatrapidly circularizes upon infection (Roizman and Sears, 1996). Over 100 genes are expressed from this complex genome, which canbe divided into 2 unique regions: the long unique (UL) and short unique (US) regions, flanked by long repeat (RL) and short repeat(RS) regions. Although the majority of HSV-1 genes are encoded in the unique regions, LAT, the sole viral transcript expressed inlatently infected cells, is located within the RL region (Ahmed et al., 2002; Jones, 2003; Gupta et al., 2006). (B) The LAT forms astem-loop structure, which is sequentially cleaved by Drosha (producing pre-miR-LAT, sequence shown) and Dicer into the maturemiRNA, miR-LAT. The miR-LAT recognizes complementary sites in the 30 UTR of 2 genes important for the induction ofapoptosis, TGF-b and SMAD3. The miR-LAT-directed cleavage of these mRNAs leads to the inhibition of apoptosis in latentlyinfected cells, ensuring prolonged survival of the virus (Cole, 1996).
242 DYKXHOORN

inhibited the antiapoptotic effects of the LAT gene. These ef-
fects were also attenuated in HSV-1-infected cells treated with
a noncleavable oligonucleotide complementary to miR-LAT.
Computational analysis of the host genome for potential miR-
LAT target sites found sites within the 30UTR of 2 genes as-
sociated with apoptosis: transforming growth factor-b (TGF-b)and mothers against decapentaplegic homolog 3 (SMAD3).
The expression of both these genes was down-regulated in cells
infected with wild-type HSV-1 but not in cells infected with a
mutant HSV-1 strain containing a deletion in exon 1 of the LAT
gene. As expected, this mutant virus was unable to protect the
cells from apoptosis. The loss of TGF-b and SMAD3 mRNA
in HSV-1-infected cells indicated that miR-LAT-mediated si-
lencing was occurring by the cleavage and degradation of the
target mRNAs. This is not unexpected, as miR-LAT has a high
degree of complementarity with the target sites. These results
clearly indicate that miR-LAT can protect HSV-1-infected cells
from apoptosis induced by TGF-b signaling and contribute to
the maintenance of HSV-1 latency. In addition, as miR-LAT is
sufficient for the maintenance of viral latency, HSV-1-infected
cells can evade detection by the host immune system, which
often accompanies the production of potentially immunogenic
viral proteins.
Viral miRNAs targeting viral messages
As an alternative to the silencing of host genes, virus-encoded
miRNAs may carry out their regulatory function by targeting
viral messages. This was found to be the case in simian virus 40
(SV40). Computational analysis of SV40 identified a potential
pre-miRNA stem loop that mapped to the 30 UTR of the late pre-
mRNA (Fig. 3) (Sullivan et al., 2005). This predicted stem loop
was highly conserved across related primate polyomaviruses.
Interestingly, this region is downstream of the polyadenylation
cleavage site. Northern blot and RNase protection assay (RPA)
analysis confirmed that 2 miRNAs were expressed from this
region, 1 from each strand of the predicted stem loop. These
miRNAs accumulated late in SV40 infection, concomitant with
the expression of the late viral mRNAs. The SV miRNAs over-
lap with, and are completely complementary to, the 30 UTR of
the early SV40 transcripts that encode the T antigens. This
would imply that the SV miRNAs should facilitate the silencing
of early viral gene products. The identification of truncated early
viral transcripts whose ends correspond to the cleavage site of
the 2 miRNAs confirmed that the early viral mRNAs were the
targets of the SV miRNAs (Sullivan et al., 2005). Cells infected
with a mutant virus that lacked the region encoding the SV
miRNAs showed no decrease in early viral mRNAs and failed
to accumulate the truncated early viral mRNAs. As expected,
the mutant virus produced higher levels of both the large and
small T antigens. However, the mutant virus was able to produce
comparable levels of infectious progeny virus, demonstrating
that the loss of the SV miRNAs had no effect on viral replica-
tion. Because T antigens are potent targets of cytotoxic T lym-
phocytes (CTLs), the down-modulation of T antigen expression
may allow the SV40-infected cells to evade immune detection.
Mutant virus–infected cells showed a significantly enhanced
susceptibility to lysis by T antigen–specific CTL clones and re-
leased higher amounts of interferon-g compared to wild-type
SV40-infected cells (Sullivan et al., 2005). Therefore, it appears
that the SV40miRNAs are used to regulate T antigen expression
to reduce the susceptibility of SV40-infected cells to CTL attack
and decrease cytokine release, thereby enabling the infected
cells to evade detection by the host immune system.
As noted above, a similar autoregulatory mechanism has been
predicted for an EBV miRNA (miR-BART2), which is com-
pletely complementary to the 30 UTR of the viral BALF5 gene
that encodes the EBV DNA polymerase. EBV-infected cells
produce a truncated BALF5 mRNA, whose cleavage site corre-
sponds precisely to the predicted cleavage site of miR-BART2.
No functional consequence has been associated with miRNA-
mediated silencing of BALF5.
miRNAs are encoded in a variety of primary transcripts, in-
cluding the introns of protein-coding or noncoding genes and
independent transcriptional units. The recent identification of
a Kaposi’s sarcoma associated virus miRNA (miR-K12-10),
which is expressed from within the open-reading frame (ORF)
of a protein-coding gene (the kaposin gene, which is involved
in cellular transformation), may serve as a novel regulatory
mechanism (Cai et al., 2005; Pfeffer et al., 2005). Instead of
the targeting of mRNA by the miRNA, which leads to the
inhibition of gene expression, the processing of the pri-miRNA
by Drosha would cleave the mRNA, causing its degradation.
The mechanism used by the virus to regulate this process to
ensure adequate expression of the viral gene product (kaposin)
remains unknown. Interestingly, the pre-miRNA that yields
miR-K12-10 is partially edited by a double-stranded RNA-
specific adenosine deaminase that converts the adenosine at
position 2 of the miRNA into an inosine, leading to a glycine
to serine change in the kaposin protein (Pfeffer et al., 2005).
Similar editing events have been found for both the human and
mousemiR-22 (Luciano et al., 2004). Interestingly, a recent study
has shown that a single adenosine-to-inosine modification in
miR376 was able to alter the target specificity of the miRNA,
demonstrating that adenosine deamination of miRNAs can play
an important role in their regulatory function (Kawahara et al.,
2007).
Host miRNA regulation of viral genes
Just as viruses can alter the expression of host genes, viral-
host interactions have coevolved so that viruses can subvert host
miRNAs to regulate their own gene expression. One such ex-
ample involves the primate foamy virus type 1 (PFV-1) (Le-
cellier et al., 2005). The suppression of the RNAi response in
cells infected with PFV-1 stimulated the accumulation of prog-
eny virus. Fragments of the viral genome were fused to the green
fluorescent protein (GFP) reporter gene to identify regions of the
genome that contained potential miRNA binding sites. One ge-
nomic fragment that led to the suppression of GFP expression in
293T cells was isolated. Computational analysis of this region of
the viral genome identified a potential miR-32 binding site. The
introduction of mutations into this site led to the increased ac-
cumulation of progeny virus, similar to that seen when the en-
dogenous RNAi pathway was inhibited. This same effect was
seen when PFV-1-infected cells were treated with a noncleav-
able oligonucleotide complementary to miR-32, confirming that
miR-32 was the effector molecule silencing viral gene expres-
sion and replication. How can the PFV-1 virus be expressed in
the context of this host-encoded suppressive miRNA?
MICRORNAS IN VIRAL REPLICATION AND PATHOGENESIS 243

Studying the RNAi response in plants and insects may pro-
vide a potential answer to this question. Unlike mammals that
have an adaptive immune system that allows for the recognition
and elimination of viral pathogens, plants lack such a system.
Instead, plants take advantage of the fact that the majority of
plant viruses go through a dsRNA intermediate during their
replication cycle (Waterhouse et al., 2001; Dunoyer et al.,
2004; Dunoyer and Voinnet, 2005; Brodersen and Voinnet,
2006). These long dsRNAs serve as substrates for Dicer
cleavage, producing siRNAs that can feed back and silence
viral gene expression. In fact, plants use existing siRNAs to
protect themselves from infection by a more pathogenic, but
related, plant virus. To circumvent this silencing, many plant
viruses have evolved strategies to inhibit the RNAi response.
This is achieved by targeting different stages in the RNAi
pathway, including inhibiting the processing of long dsRNAs
into siRNAs (e.g., P1-HcPro from Turnip Crinkle virus) and
sequestering siRNAs and thereby preventing their entry into
effector complexes (e.g., p19 from Tomato bushy stunt virus)
(Dunoyer et al., 2004). A similar strategy has been suggested
for PFV-1. The ectopic expression of PFV-1 Tas protein, a
transcriptional activator expressed early in the viral life cycle,
can inhibit miRNA activity in tissue culture cells. In addition,
the transgenic expression of PFV-1 Tas in plant cells was able
AN
AN
AN
AN
AN
AN
5'-pU U GGA GA C
U3'-U UCUG -
CGACGAACA UG GAGGACU GGGGC UGAAAU GC
CGACUUUGUCCCGCUUCUGAUUGGAAUUCUUGU - - - AC -
A CU
AA GUG
GA GA C
UCUG -
C5'-pU GGGGC UGAAAU GC
CGp-5'ACUUUGUCCCG3'-UGAUU-3'
Earlytranscripts
Latetranscripts
Large Tantigen VP1
SV40 genome
Early infection
Late infection
Large T antigen
Large T antigen
VP1
Cytotoxic T lymphocyte response
ORI
Transcriptionalregulatory/promoter
region
polyadenylation sitespre-SV40miRNA
Dicer processing
Drosha processing
mRNA recognition
mRNA cleavage
A B
FIG. 3. SV40 miRNAs and immune evasion. (A) The SV40 genome is a circular double-stranded DNA molecule from whichearly and late transcripts are sequentially expressed from a noncoding region of the genome that contains the origin of replication(ORI) and the transcriptional regulation/promoter elements (Cole, 1996). A miRNA, SV40 miRNA, is encoded in the 30 UTR ofthe late transcripts. Interestingly, the pre-SV40 miRNA stem loop is located downstream of the polyadenylation site of the latetranscripts. (B) Although necessary for viral replication, the T antigens, which are expressed early in the SV40 infection cycle, arepotent targets of the host immune system, particularly CTLs. The expression of 2 miRNAs, one from each strand of the pre-SV40miRNA (sequence shown), which recognize complementary sequences in the 30 UTR of the early transcripts, regulate the level ofT-Ag expression. This silencing of T-Ag expression helps SV40 to evade immune detection by inhibiting the recognition andattack of SV40-infected cells by CTLs.
244 DYKXHOORN

to suppress an RNAi response. It is noteworthy, that persistent
PFV-1 infection has been found to result in the accumulation
of defective viruses containing deletions in the Tas gene (Saib
et al., 1993, 1995; Meiering et al., 2000).
MiRNA-directed enhancement of hepatitisC virus replication
A unique relationship has been found between the hepatitis C
virus (HCV) and the highly expressed, liver-specific miR-122
( Jopling et al., 2005). Instead of directing the silencing of viral
gene expression and replication, miR-122 appears to be required
for the efficient viral RNA expression. The first clue that miR-
122 has a positive effect on HCV replication came from the
observation that HCV replicons were able to replicate in miR-
122-expressing liver cell lines (e.g., Huh7 cells) but not in cells
that lacked miR-122 expression (e.g., HepG2). The sequestra-
tion of miR-122 by complementary 20O-methyl oligonucleo-
tides impaired the ability of HCV to replicate in the normally
permissive Huh7 cells. Computational analysis of the HCV ge-
nome identified 2 potential binding sites for miR-122, one in the
30 noncoding region (NCR) and the other in the 50 NCR (Jopling
et al., 2005). Mutational analysis of these binding sites dem-
onstrated that the site in the 50 NCR, but not in the 30 NCR, wasimportant for facilitating the miR-122-dependent accumulation
of viral RNA. This effect was reversible as the accumulation of
viral RNAs could be restored by introducing exogenous miR-
122-containing complementary mutations into the cells. The
positive effect of miR-122 on HCV appears to be at the level of
replication and not by causing alterations in the translation or
stability of the viral RNA (Jopling et al., 2005). The mechanism
underlying this positive effect of miR-122 on HCV remains to
be elucidated.
Viral suppressors of RNAi
The RNAi pathway in plants and insects serves as a primi-
tive, nucleic acid–based adaptive immune system that recog-
nizes long dsRNA molecules produced during viral replication
and cleaves them into siRNAs that inhibit further viral gene
expression and replication. In response to this, several plant
and insect viruses have evolved mechanisms to suppress the
antiviral effects of the RNAi pathway (Dunoyer et al., 2004;
Dunoyer and Voinnet, 2005; Brodersen and Voinnet, 2006; van
Rij et al., 2006). Although the relationship of vertebrate viruses
with the RNAi pathway differs significantly from that of plant
and insect viruses, there are examples of specific viruses in-
fecting vertebrate cells that have developed mechanisms for the
inhibition of the RNAi pathway to facilitate their replication.
As described above, the PFV-1 Tas protein inhibits miRNA
expression, protecting the virus from silencing by the host miR-
32 (Lecellier et al., 2005).
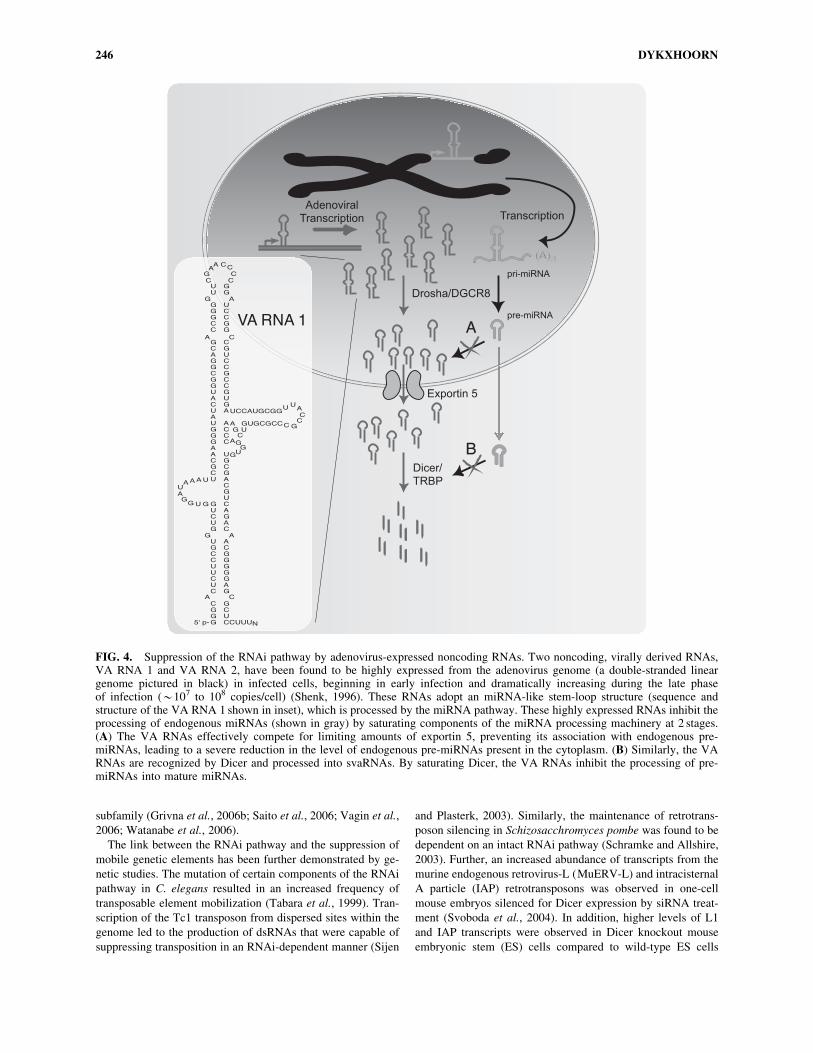
An alternative mechanism for the suppression of the RNAi
machinery has been identified in adenovirus-infected cells. Two
inhibitory, noncoding RNAs (termed viral associated RNAs
(VA RNA 1 and VA RNA 2)) have been shown to be highly
expressed in adenovirus-infected cells (Fig. 4) (Mathews and
Shenk, 1991). Analysis of the structure of these RNAs has pre-
dicted that they adopt an imperfect stem-loop secondary struc-
ture similar to that found in pre-miRNAs. The VA RNAs appear
to inhibit the RNAi pathway at 2 points. Similar to pre-miRNAs,
they use the nuclear-export receptor exportin 5 for their trans-
location from the nucleus into the cytoplasm. The VA RNAs are
so highly expressed that they effectively compete with the en-
dogenous miRNAs for exportin 5, leading to the saturation of the
nuclear translocation activity (Lu and Cullen, 2004). Once in the
cytoplasm, the VA RNAs are recognized and processed by Dicer
into small VA RNAs (svaRNAs), which have been shown to
associate with RISC during lytic viral infections. These highly
abundant RNAs appear to competitively bind to Dicer and
prevent the processing of endogenous Dicer substrates (e.g.,
miRNAs) (Lu and Cullen, 2004; Andersson et al., 2005). Inter-
estingly, VA RNAs can bind to protein kinase R (PKR), the
dsRNA recognition protein that activates the innate immune
response to long dsRNA species (like those seen during the rep-
lication cycle of many viruses), impairing the activation of the
antiviral interferon response (Mathews and Shenk, 1991). The
treatment of adenovirus-infected cells with 20O-methyl oligo-
nucleotides that bind and sequester the svaRNAs has been shown
to impair progeny virus production and reduce viral titers. The
VA RNAs have also been shown to be potent inhibitors of the
interferon response, which acts to recognize long double-stran-
ded RNAs usually associated with the replication cycle of certain
viruses. The VA RNAs accomplish this by binding to PKR and
inhibiting its activation (Kitajewski et al., 1986).
RNAI-MEDIATED SUPPRESSION OF MOBILEGENETIC ELEMENTS
Biochemical and computational approaches have been used
to gain a better understanding of the complexity of the small
regulatory RNA coding capacity from a wide variety of cell
types, organisms, and development stages. The large-scale
analysis of the small RNA component of the Arabidopsis
thaliana genome found that many regions of the genome that
were devoid of protein-coding genes encoded a significant
amount of small RNA molecules (Lu et al., 2005). Although
miRNAs represented the most abundant species of nonredun-
dant small RNAs in this analysis, the largest proportion of the
identified small RNAs matched sequences from repetitive ge-
nomic sequences (e.g., retrotransposons and transposons) and
intergenic regions. The isolation of small RNAs encoded from
repetitive sequence elements is not limited to plants. The anal-
ysis of small RNA molecules from Drosophila melanogaster
(Aravin et al., 2003, 2006; Saito et al., 2006; Vagin et al., 2006),
Trypanosoma brucei (Djikeng et al., 2001), mice (Grivna et al.,
2006a, 2006b; Watanabe et al., 2006), and human cells (Yang
and Kazazian, 2006) identified small RNAs that corresponded to
a variety of repetitive elements, including LTR and non-LTR
retrotransposons, transposons, satellite and microsatellite DNA,
and heterochromatic regions. These small RNAs are termed
rasiRNAs (Aravin et al., 2003). Interestingly, they appear to
have unique characteristics that distinguish them from miRNAs
and siRNAs. The majority of rasiRNAs appear to be expressed
in germline tissues, they are longer than siRNAs and miRNAs,
and accumulate from only 1 strand (antisense strand) (Grivna et
al., 2006a; Saito et al., 2006; Vagin et al., 2006;Watanabe et al.,
2006). Unlike miRNAs and siRNAs that interact with members
of the Ago subfamily of Argonaute proteins for their effector
functions, rasiRNAs are associated with members of the Piwi
MICRORNAS IN VIRAL REPLICATION AND PATHOGENESIS 245
subfamily (Grivna et al., 2006b; Saito et al., 2006; Vagin et al.,
2006; Watanabe et al., 2006).
The link between the RNAi pathway and the suppression of
mobile genetic elements has been further demonstrated by ge-
netic studies. The mutation of certain components of the RNAi
pathway in C. elegans resulted in an increased frequency of
transposable element mobilization (Tabara et al., 1999). Tran-
scription of the Tc1 transposon from dispersed sites within the
genome led to the production of dsRNAs that were capable of
suppressing transposition in an RNAi-dependent manner (Sijen
and Plasterk, 2003). Similarly, the maintenance of retrotrans-
poson silencing in Schizosacchromyces pombe was found to be
dependent on an intact RNAi pathway (Schramke and Allshire,
2003). Further, an increased abundance of transcripts from the
murine endogenous retrovirus-L (MuERV-L) and intracisternal
A particle (IAP) retrotransposons was observed in one-cell
mouse embryos silenced for Dicer expression by siRNA treat-
ment (Svoboda et al., 2004). In addition, higher levels of L1
and IAP transcripts were observed in Dicer knockout mouse
embryonic stem (ES) cells compared to wild-type ES cells
A
B
UU
GGGCC
GCAGGCGGUACUAUGGGAACGCU
GUCUG
UGCCUUCUC
CGGG
GG
UCCGG
CGUCCGCCGUGA
AC CC
UGCGACGUCAGAC
ACGGGGGAG
GCUCCUUUN
A
G
C
UCCAUGCGG
A GUGCGCC
A
A
G
C
A
CG
AA CC
CC
U UAC
CGC
GC
AGG
UG
U
UAAAUAG
G U G
5' p-
VA RNA 1
Adenoviral
Transcription
Drosha/DGCR8
Exportin 5
Dicer/
TRBP
Transcription
(A)N(A)N
pri-miRNA
pre-miRNA
FIG. 4. Suppression of the RNAi pathway by adenovirus-expressed noncoding RNAs. Two noncoding, virally derived RNAs,VA RNA 1 and VA RNA 2, have been found to be highly expressed from the adenovirus genome (a double-stranded lineargenome pictured in black) in infected cells, beginning in early infection and dramatically increasing during the late phaseof infection (*107 to 108 copies/cell) (Shenk, 1996). These RNAs adopt an miRNA-like stem-loop structure (sequence andstructure of the VA RNA 1 shown in inset), which is processed by the miRNA pathway. These highly expressed RNAs inhibit theprocessing of endogenous miRNAs (shown in gray) by saturating components of the miRNA processing machinery at 2 stages.(A) The VA RNAs effectively compete for limiting amounts of exportin 5, preventing its association with endogenous pre-miRNAs, leading to a severe reduction in the level of endogenous pre-miRNAs present in the cytoplasm. (B) Similarly, the VARNAs are recognized by Dicer and processed into svaRNAs. By saturating Dicer, the VA RNAs inhibit the processing of pre-miRNAs into mature miRNAs.
246 DYKXHOORN

(Kanellopoulou et al., 2005). These results indicate that the
RNAimachinery plays an important role in protecting cells from
the negative effects of mutagenic genetic elements.
CONCLUSIONS
Only several years have passed since the first demonstration
that RNAi functions in mammalian systems, yet a substantial
amount of research has gone into understanding the roles that
small RNA regulatory molecules play in diverse cellular pro-
cesses. It should come as no surprise that viruses, which are
particularly adept at commandeering the cellular machinery to
ensure their own fitness and survival, have evolved mechanisms
that allow them to take advantage of these ubiquitous regulatory
mechanisms. It is clear that we are only just beginning to plumb
the depths of the relationship between viruses and other patho-
genic mobile genetic elements and the RNAi pathway.
REFERENCES
AHMED, M., LOCK, M., MILLER, C.G., and FRASER, N.W. (2002).Regions of the herpes simplex virus type 1 latency-associated tran-script that protect cells from apoptosis in vitro and protect neuronalcells in vivo. J. Virol. 76, 717–729.
AMBROS, V. (2004). The functions of animal microRNAs. Nature431, 350–355.
AMBROS, V., and LEE, R.C. (2004). Identification of microRNAs andother tiny noncoding RNAs by cDNA cloning. Methods Mol. Biol.265, 131–158.
ANDERSSON, M.G., HAASNOOT, P.C., XU, N., BERENJIAN, S.,BERKHOUT, B., and AKUSJARVI, G. (2005). Suppression of RNAinterference by adenovirus virus-associated RNA. J. Virol. 79, 9556–9565.
ARAVIN, A., GAIDATZIS, D., PFEFFER, S., LAGOS-QUINTANA,M., LANDGRAF, P., IOVINO, N., et al. (2006). A novel class ofsmall RNAs bind to MILI protein in mouse testes. Nature 442, 203–207.
ARAVIN, A.A., LAGOS-QUINTANA, M., YALCIN, A., ZAVOLAN,M., MARKS, D., SNYDER, B., et al. (2003). The small RNA profileduring Drosophila melanogaster development. Dev. Cell 5, 337–350.
BAGGA, S., BRACHT, J., HUNTER, S., MASSIRER, K., HOLTZ, J.,EACHUS, R., et al. (2005). Regulation by let-7 and lin-4 miRNAsresults in target mRNA degradation. Cell 122, 553–563.
BARTEL, D.P. (2004). MicroRNAs: genomics, biogenesis, mecha-nism, and function. Cell 116, 281–297.
BENTWICH, I., AVNIEL, A., KAROV, Y., AHARONOV, R., GI-LAD, S., BARAD, O., et al. (2005). Identification of hundreds ofconserved and nonconserved human microRNAs. Nat. Genet. 37,766–770.
BERNSTEIN, E., CAUDY, A.A., HAMMOND, S.M., and HANNON,G.J. (2001). Role for a bidentate ribonuclease in the initiation step ofRNA interference. Nature 409, 363–366.
BOHNSACK, M.T., CZAPLINSKI, K., and GORLICH, D. (2004).Exportin 5 is a RanGTP-dependent dsRNA-binding protein thatmediates nuclear export of pre-miRNAs. RNA 10, 185–191.
BRODERSEN, P., and VOINNET, O. (2006). The diversity of RNAsilencing pathways in plants. Trends Genet. 22, 268–280.
CAI, X., HAGEDORN, C.H., and CULLEN, B.R. (2004). HumanmicroRNAs are processed from capped, polyadenylated transcriptsthat can also function as mRNAs. RNA 10, 1957–1966.
CAI, X., LU, S., ZHANG, Z., GONZALEZ, C.M., DAMANIA, B., andCULLEN, B.R. (2005). Kaposi’s sarcoma-associated herpesvirusexpresses an array of viral microRNAs in latently infected cells.Proc. Natl. Acad. Sci. USA 102, 5570–5575.
CAI, X., SCHAFER, A., LU, S., BILELLO, J.P., DESROSIERS, R.C.,EDWARDS, R., et al. (2006). Epstein-Barr virus microRNAs are
evolutionarily conserved and differentially expressed. PLoS Pathog.2, e23.
CHALFIE, M., HORVITZ, H.R., and SULSTON, J.E. (1981). Muta-tions that lead to reiterations in the cell lineages of C. elegans. Cell24, 59–69.
CHEN, C.Z., LI, L., LODISH, H.F., and BARTEL, D.P. (2004). Mi-croRNAs modulate hematopoietic lineage differentiation. Science303, 83–86.
CHENDRIMADA, T.P., GREGORY, R.I., KUMARASWAMY, E.,NORMAN, J., COOCH, N., NISHIKURA, K., et al. (2005). TRBPrecruits the Dicer complex to Ago2 for microRNA processing andgene silencing. Nature 436, 740–744.
COLE, C.N. (1996). Polyomavirinae: the viruses and their replication.In Fundamental Virology, 3rd edition. B.N. FIELDS, D.M. KNIPE,P.M. HOWLEY, et al., eds. (Lippincott-Raven Publishers, Phila-delphia, PA), pp. 917–945.
DJIKENG, A., SHI, H., TSCHUDI, C., and ULLU, E. (2001). RNAinterference in Trypanosoma brucei: cloning of small interferingRNAs provides evidence for retroposon-derived 24–26-nucleotideRNAs. RNA 7, 1522–1530.
DOENCH, J.G., PETERSEN, C.P., and SHARP, P.A. (2003). siRNAscan function as miRNAs. Genes Dev. 17, 438–442.
DUNOYER, P., LECELLIER, C.H., PARIZOTTO, E.A., HIMBER,C., and VOINNET, O. (2004). Probing the microRNA and smallinterfering RNA pathways with virus-encoded suppressors of RNAsilencing. Plant Cell 16, 1235–1250.
DUNOYER, P., and VOINNET, O. (2005). The complex interplaybetween plant viruses and host RNA-silencing pathways. Curr. Opin.Plant Biol. 8, 415–423.
ELBASHIR, S.M., LENDECKEL, W., and TUSCHL, T. (2001). RNAinterference is mediated by 21- and 22-nucleotide RNAs. GenesDev. 15, 188–200.
FORSTEMANN, K., TOMARI, Y., DU, T., VAGIN, V.V., DENLI,A.M., BRATU, D.P., et al. (2005). Normal microRNA maturationand germ-line stem cell maintenance requires Loquacious, a double-stranded RNA-binding domain protein. PLoS Biol. 3, e236.
FURNARI, F.B., ADAMS, M.D., and PAGANO, J.S. (1993). Uncon-ventional processing of the 30 termini of the Epstein-Barr virus DNApolymerase mRNA. Proc. Natl. Acad. Sci. USA 90, 378–382.
GALIANA-ARNOUX, D., DOSTERT, C., SCHNEEMANN, A.,HOFFMANN, J.A., and IMLER, J.L. (2006). Essential functionin vivo for Dicer-2 in host defense against RNA viruses in dro-sophila. Nat. Immunol. 7, 590–597.
GREGORY, R.I., CHENDRIMADA, T.P., COOCH, N., and SHIE-KHATTAR, R. (2005). Human RISC couples microRNA biogenesisand posttranscriptional gene silencing. Cell 123, 631–640.
GREY, F., ANTONIEWICZ, A., ALLEN, E., SAUGSTAD, J.,MCSHEA, A., CARRINGTON, J.C., et al. (2005). Identification andcharacterization of human cytomegalovirus-encoded microRNAs. J.Virol. 79, 12095–12099.
GRIVNA, S.T., BEYRET, E., WANG, Z., and LIN, H. (2006a). Anovel class of small RNAs in mouse spermatogenic cells. GenesDev. 20, 1709–1714.
GRIVNA, S.T., PYHTILA, B., and LIN, H. (2006b). MIWI associateswith translational machinery and PIWI-interacting RNAs (piRNAs)in regulating spermatogenesis. Proc. Natl. Acad. Sci. USA 103,13415–13420.
GUPTA, A., GARTNER, J.J., SETHUPATHY, P., HATZI-GEORGIOU, A.G., and FRASER, N.W. (2006). Anti-apoptoticfunction of a microRNA encoded by the HSV-1 latency-associatedtranscript. Nature 442, 82–85.
HAASE, A.D., JASKIEWICZ, L., ZHANG, H., LAINE, S., SACK, R.,GATIGNOL, A., et al. (2005). TRBP, a regulator of cellular PKRand HIV-1 virus expression, interacts with Dicer and functions inRNA silencing. EMBO Rep. 6, 961–967.
HAN, J., LEE, Y., YEOM, K.H., NAM, J.W., HEO, I., RHEE, J.K.,et al. (2006). Molecular basis for the recognition of primary mi-croRNAs by the Drosha-DGCR8 complex. Cell 125, 887–901.
HORVITZ, H.R., and SULSTON, J.E. (1980). Isolation and geneticcharacterization of cell-lineage mutants of the nematode Caenor-habditis elegans. Genetics 96, 435–454.
HOUBAVIY, H.B., MURRAY, M.F., and SHARP, P.A. (2003). Em-bryonic stem cell-specific microRNAs. Dev. Cell 5, 351–358.
MICRORNAS IN VIRAL REPLICATION AND PATHOGENESIS 247

INMAN, M., PERNG, G.C., HENDERSON, G., GHIASI, H., NES-BURN, A.B., WECHSLER, S.L., et al. (2001). Region of herpessimplex virus type 1 latency-associated transcript sufficient for wild-type spontaneous reactivation promotes cell survival in tissue cul-ture. J. Virol. 75, 3636–3646.
JONES, C. (2003). Herpes simplex virus type 1 and bovine herpesvirus1 latency. Clin. Microbiol. Rev. 16, 79–95.
JOPLING, C.L., YI, M., LANCASTER, A.M., LEMON, S.M., andSARNOW, P. (2005). Modulation of hepatitis C virus RNA abun-dance by a liver-specific microRNA. Science 309, 1577–1581.
KANELLOPOULOU, C., MULJO, S.A., KUNG, A.L., GANESAN, S.,DRAPKIN, R., JENUWEIN, T., et al. (2005). Dicer-deficient mouseembryonic stem cells are defective in differentiation and centromericsilencing. Genes Dev. 19, 489–501.
KAWAHARA, Y., ZINSHTEYN, B., SETHUPATHY, P., IIZASA, H.,HATZIGEORGIOU, A.G., and NISHIKURA, K. (2007). Redirec-tion of silencing targets by adenosine-to-inosine editing of miRNAs.Science 315, 1137–1140.
KEENE, K.M., FOY, B.D., SANCHEZ-VARGAS, I., BEATY, B.J.,BLAIR, C.D., and OLSON, K.E. (2004). RNA interference acts as anatural antiviral response to O’nyong-nyong virus (Alphavirus; To-gaviridae) infection of Anopheles gambiae. Proc. Natl. Acad. Sci.USA 101, 17240–17245.
KHVOROVA, A., REYNOLDS, A., and JAYASENA, S.D. (2003).Functional siRNAs and miRNAs exhibit strand bias. Cell 115, 209–216.
KIM, V.N. (2005). MicroRNA biogenesis: coordinated cropping anddicing. Nat. Rev. Mol. Cell Biol. 6, 376–385.
KITAJEWSKI, J., SCHNEIDER, R.J., SAFER, B., MUNEMITSU,S.M., SAMUEL, C.E., THIMMAPPAYA, B., et al. (1986). Adeno-virus VAI RNA antagonizes the antiviral action of interferon bypreventing activation of the interferon-induced eIF-2 alpha kinase.Cell 45, 195–200.
LAGOS-QUINTANA, M., RAUHUT, R., LENDECKEL, W., andTUSCHL, T. (2001). Identification of novel genes coding for smallexpressed RNAs. Science 294, 853–858.
LAGOS-QUINTANA, M., RAUHUT, R., MEYER, J., BORKHARDT,A., and TUSCHL, T. (2003). New microRNAs from mouse andhuman. RNA 9, 175–179.
LAGOS-QUINTANA, M., RAUHUT, R., YALCIN, A., MEYER, J.,LENDECKEL, W., and TUSCHL, T. (2002). Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739.
LAU, N.C., LIM, L.P., WEINSTEIN, E.G., and BARTEL, D.P. (2001).An abundant class of tiny RNAs with probable regulatory roles inCaenorhabditis elegans. Science 294, 858–862.
LECELLIER, C.H., DUNOYER, P., ARAR, K., LEHMANN-CHE, J.,EYQUEM, S., HIMBER, C., et al. (2005). A cellular microRNAmediates antiviral defense in human cells. Science 308, 557–560.
LEE, Y., AHN, C., HAN, J., CHOI, H., KIM, J., YIM, J., et al. (2003).The nuclear RNase III Drosha initiates microRNA processing. Na-ture 425, 415–419.
LEE, Y., JEON, K., LEE, J.T., KIM, S., and KIM, V.N. (2002). Mi-croRNA maturation: stepwise processing and subcellular localiza-tion. EMBO J. 21, 4663–4670.
LEE, Y., KIM, M., HAN, J., YEOM, K.H., LEE, S., BAEK, S.H., et al.(2004). MicroRNA genes are transcribed by RNA polymerase II.EMBO J. 23, 4051–4060.
LIM, L.P., LAU, N.C., GARRETT-ENGELE, P., GRIMSON, A.,SCHELTER, J.M., CASTLE, J., et al. (2005). Microarray analysisshows that some microRNAs downregulate large numbers of targetmRNAs. Nature 433, 769–773.
LU, C., TEJ, S.S., LUO, S., HAUDENSCHILD, C.D., MEYERS, B.C.,and GREEN, P.J. (2005). Elucidation of the small RNA componentof the transcriptome. Science 309, 1567–1569.
LU, S., and CULLEN, B.R. (2004). Adenovirus VA1 noncoding RNAcan inhibit small interfering RNA and microRNA biogenesis. J.Virol. 78, 12868–12876.
LUCIANO, D.J., MIRSKY, H., VENDETTI, N.J., and MAAS, S.(2004). RNA editing of a miRNA precursor. RNA 10, 1174–1177.
LUND, E., GUTTINGER, S., CALADO, A., DAHLBERG, J.E., andKUTAY, U. (2004). Nuclear export of microRNA precursors. Sci-ence 303, 95–98.
MASSIRER, K.B., and PASQUINELLI, A.E. (2006). The evolving roleof microRNAs in animal gene expression. Bioessays 28, 449–452.
MATHEWS, M.B., and SHENK, T. (1991). Adenovirus virus-associ-ated RNA and translation control. J. Virol. 65, 5657–5662.
MATRANGA, C., TOMARI, Y., SHIN, C., BARTEL, D.P., and ZA-MORE, P.D. (2005). Passenger-strand cleavage facilitates assemblyof siRNA into Ago2-containing RNAi enzyme complexes. Cell 123,607–620.
MEIERING, C.D., COMSTOCK, K.E., and LINIAL, M.L. (2000).Multiple integrations of human foamy virus in persistently infectedhuman erythroleukemia cells. J. Virol. 74, 1718–1726.
MEISTER, G., LANDTHALER, M., PATKANIOWSKA, A., DOR-SETT, Y., TENG, G., and TUSCHL, T. (2004). Human Argonaute2mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell15, 185–197.
PASQUINELLI, A.E., HUNTER, S., and BRACHT, J. (2005). Mi-croRNAs: a developing story. Curr. Opin. Genet. Dev. 15, 200–205.
PFEFFER, S., SEWER, A., LAGOS-QUINTANA, M., SHERIDAN,R., SANDER, C., GRASSER, F.A., et al. (2005). Identification ofmicroRNAs of the herpesvirus family. Nat. Methods 2, 269–276.
PFEFFER, S., ZAVOLAN, M., GRASSER, F.A., CHIEN, M., RUSSO,J.J., JU, J., et al. (2004). Identification of virus-encoded microRNAs.Science 304, 734–736.
POY, M.N., ELIASSON, L., KRUTZFELDT, J., KUWAJIMA, S., MA,X., MACDONALD, P.E., et al. (2004). A pancreatic islet-specificmicroRNA regulates insulin secretion. Nature 432, 226–230.
RODRIGUEZ, A., GRIFFITHS-JONES, S., ASHURST, J.L., andBRADLEY, A. (2004). Identification of mammalian microRNA hostgenes and transcription units. Genome Res. 14, 1902–1910.
ROIZMAN, B., and SEARS, A.E. (1996). Herpes simplex viruses andtheir replication. In Fundamental Virology, 3rd edition. B.N.FIELDS, D.M. KNIPE, P.M. HOWLEY, et al., eds. (Lippincott-Raven Publishers, Philadelphia, PA), pp. 1043–1107.
SAIB, A., KOKEN, M.H., VAN DER SPEK, P., PERIES, J., and DETHE, H. (1995). Involvement of a spliced and defective humanfoamy virus in the establishment of chronic infection. J. Virol. 69,5261–5268.
SAIB, A., PERIES, J., and DE THE, H. (1993). A defective humanfoamy provirus generated by pregenome splicing. EMBO J. 12,4439–4444.
SAITO, K., ISHIZUKA, A., SIOMI, H., and SIOMI, M.C. (2005).Processing of pre-microRNAs by the Dicer-1-Loquacious complexin Drosophila cells. PLoS Biol. 3, e235.
SAITO, K., NISHIDA, K.M., MORI, T., KAWAMURA, Y., MIYOSHI,K., NAGAMI, T., et al. (2006). Specific association of Piwi withrasiRNAs derived from retrotransposon and heterochromatic regionsin the Drosophila genome. Genes Dev. 20, 2214–2222.
SAMOLS, M.A., HU, J., SKALSKY, R.L., and RENNE, R. (2005).Cloning and identification of a microRNA cluster within the latency-associated region of Kaposi’s sarcoma-associated herpesvirus. J.Virol. 79, 9301–9305.
SANCHEZ-VARGAS, I., TRAVANTY, E.A., KEENE, K.M.,FRANZ, A.W., BEATY, B.J., BLAIR, C.D., et al. (2004). RNAinterference, arthropod-borne viruses, and mosquitoes. Virus Res.102, 65–74.
SCHRAMKE, V., and ALLSHIRE, R. (2003). Hairpin RNAs andretrotransposon LTRs effect RNAi and chromatin-based gene si-lencing. Science 301, 1069–1074.
SCHWARZ, D.S., HUTVAGNER, G., DU, T., XU, Z., ARONIN, N.,and ZAMORE, P.D. (2003). Asymmetry in the assembly of theRNAi enzyme complex. Cell 115, 199–208.
SHENK, T. (1996). Adenoviridae: the viruses and their replication. InFundamental Virology, 3rd edition. B.N. FIELDS, D.M. KNIPE,P.M. HOWLEY, et al., eds. (Lippincott-Raven Publishers, Phila-delphia, PA), pp. 979–1016.
SIJEN, T., and PLASTERK, R.H. (2003). Transposon silencing in theCaenorhabditis elegans germ line by natural RNAi. Nature 426, 310–314.
SULLIVAN, C.S., GRUNDHOFF, A.T., TEVETHIA, S., PIPAS, J.M.,and GANEM, D. (2005). SV40-encoded microRNAs regulate viralgene expression and reduce susceptibility to cytotoxic T cells. Na-ture 435, 682–686.
248 DYKXHOORN

SULSTON, J.E., and HORVITZ, H.R. (1981). Abnormal cell lineagesin mutants of the nematode Caenorhabditis elegans. Dev. Biol. 82,41–55.
SVOBODA, P., STEIN, P., ANGER, M., BERNSTEIN, E., HANNON,G.J., and SCHULTZ, R.M. (2004). RNAi and expression ofretrotransposons MuERV-L and IAP in preimplantation mouse em-bryos. Dev. Biol. 269, 276–285.
TABARA, H., SARKISSIAN, M., KELLY, W.G., FLEENOR, J.,GRISHOK, A., TIMMONS, L., et al. (1999). The rde-1 gene, RNAinterference, and transposon silencing in C. elegans. Cell 99, 123–132.
TOMARI, Y., MATRANGA, C., HALEY, B., MARTINEZ, N., andZAMORE, P.D. (2004). A protein sensor for siRNA asymmetry.Science 306, 1377–1380.
TOMARI, Y., and ZAMORE, P.D. (2005). MicroRNA biogenesis:drosha can’t cut it without a partner. Curr. Biol. 15, R61–R64.
TUSCHL, T., ZAMORE, P.D., LEHMANN, R., BARTEL, D.P., andSHARP, P.A. (1999). Targeted mRNA degradation by double-stranded RNA in vitro. Genes Dev. 13, 3191–3197.
VAGIN, V.V., SIGOVA, A., LI, C., SEITZ, H., GVOZDEV, V., andZAMORE, P.D. (2006). A distinct small RNA pathway silencesselfish genetic elements in the germline. Science 313, 320–324.
VAN RIJ, R.P., SALEH, M.C., BERRY, B., FOO, C., HOUK, A.,ANTONIEWSKI, C., et al. (2006). The RNA silencing endonucleaseArgonaute 2 mediates specific antiviral immunity in Drosophilamelanogaster. Genes Dev. 20, 2985–2995.
VOINNET, O. (2005). Induction and suppression of RNA silencing:insights from viral infections. Nat. Rev. Genet. 6, 206–220.
WANG, X.H., ALIYARI, R., LI, W.X., LI, H.W., KIM, K., CAR-THEW, R., et al. (2006). RNA interference directs innate immunityagainst viruses in adult Drosophila. Science 312, 452–454.
WATANABE, T., TAKEDA, A., TSUKIYAMA, T., MISE, K.,OKUNO, T., SASAKI, H., et al. (2006). Identification and charac-terization of two novel classes of small RNAs in the mouse germline:
retrotransposon-derived siRNAs in oocytes and germline smallRNAs in testes. Genes Dev. 20, 1732–1743.
WATERHOUSE, P.M., WANG, M.B., and LOUGH, T. (2001). Genesilencing as an adaptive defence against viruses. Nature 411, 834–842.
YANG, N., and KAZAZIAN, H.H., JR. (2006). L1 retrotransposition issuppressed by endogenously encoded small interfering RNAs inhuman cultured cells. Nat. Struct. Mol. Biol. 13, 763–771.
YEOM, K.H., LEE, Y., HAN, J., SUH, M.R., and KIM, V.N. (2006).Characterization of DGCR8/Pasha, the essential cofactor for Droshain primary miRNA processing. Nucleic Acids Res. 34, 4622–4629.
YI, R., QIN, Y., MACARA, I.G., and CULLEN, B.R. (2003). Ex-portin-5 mediates the nuclear export of pre-microRNAs and shorthairpin RNAs. Genes Dev. 17, 3011–3016.
ZAMORE, P.D., TUSCHL, T., SHARP, P.A., and BARTEL, D.P.(2000). RNAi: double-stranded RNA directs the ATP-dependentcleavage of mRNA at 21 to 23 nucleotide intervals. Cell 101, 25–33.
ZENG, Y., YI, R., and CULLEN, B.R. (2005). Recognition and cleav-age of primary microRNA precursors by the nuclear processing en-zyme Drosha. EMBO J. 24, 138–148.
Address reprint requests to:
Derek M. Dykxhoorn
The CBR Institute for Biomedical Research
Harvard Medical School
Boston, MA 02115
E-mail: [email protected]
Received for publication December 7, 2006; received in re-
vised form January 16, 2007; accepted January 17, 2007.
MICRORNAS IN VIRAL REPLICATION AND PATHOGENESIS 249
