microbial adaptations and controlling mechanisms of ...857707/fulltext01.pdf · 2015 digital...
TRANSCRIPT

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2015
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1296
Microbial adaptations andcontrolling mechanisms ofsurface-associated microhabitatheterogeneity in aquatic systems
JAN TORSTEN JESKE
ISSN 1651-6214ISBN 978-91-554-9351-6urn:nbn:se:uu:diva-263206

Dissertation presented at Uppsala University to be publicly examined in Ekman Salen,Norbyvägen 14, Uppsala, Friday, 20 November 2015 at 13:15 for the degree of Doctor ofPhilosophy. The examination will be conducted in English. Faculty examiner: ProfessorAndreas Schramm (Aarhus University).
AbstractJeske, J. T. 2015. Microbial adaptations and controlling mechanisms of surface-associatedmicrohabitat heterogeneity in aquatic systems. Digital Comprehensive Summaries of UppsalaDissertations from the Faculty of Science and Technology 1296. 69 pp. Uppsala: ActaUniversitatis Upsaliensis. ISBN 978-91-554-9351-6.
Habitat heterogeneity is a driving factor for speciation and ecosystem functioning and iswell studied in macro-ecology. Yet our understanding of microbial adaptations, and governingprocesses is incomplete. The here presented thesis aims at giving us a better understanding ofpatterns in micro-heterogeneity, and microbial adaptations to such heterogeneity with particularfocus on surface-dominated, aquatic habitats. The most prominent microbial adaptation tosurface associated mode of life is biofilm formation. Biofilms rely heavily on type IV pili.These pili systems are well studied in Bacteria, but largely unknown in Archaea. Therefore,the first part of this thesis focuses on resolving genetic and structural feature of the type IVlike aap-pilus of the thermo-acidophilic Sulfolobus acidocaldarius. We found the aap-pilus tobe indispensible for biofilm formation, and to be unparalleled in variability of its quaternarystructure and cross regulation with other filaments. The second part of this thesis investigatesparticle colonization in the water column, focusing on diatoms as a model system, allowing anin situ assessment of different stages of particle colonization, and potential particle-specificityof the associated bacterial community. Opposing reports from marine systems, we did notobserve diatom-specificity in the associated bacterial community. Instead we found bacterialcommunity subsets, one likely originating from sediment resuspension, and the other beingcontrolled by biofilm-forming populations (e.g. Flexibacter), able to attach to newly formedparticle surfaces and subsequently facilitate secondary colonization by other bacteria. Finally,the habitat heterogeneity in top-layers of lake sediments were investigated in experimentalmicrocosms. Cell-specific oxygen consumption rates were determined, to assess microbialactivity across different scales. Individual activity rates differed strongly across all investigatedscales, likely due to spatially heterogeneous distribution of nutrients with differing quality.Vice versa, the influence of microbial activity on micro-habitat-heterogeneity was investigated.We correlated sediment redox-state with bacterial community composition and populations.Our results indicate that habitat heterogeneity is generally beneficial for microorganism, andgreater heterogeneity results in greater bacterial diversity. However, this heterogeneity-diversityrelationship is limited and microorganisms actively stabilize their immediate redox environmentto a preferred, community-specific, stable state, if cell abundances exceed a minimum threshold.
Keywords: microbial habitat heterogeneity
Jan Torsten Jeske, Department of Ecology and Genetics, Limnology, Norbyv 18 D, UppsalaUniversity, SE-75236 Uppsala, Sweden.
© Jan Torsten Jeske 2015
ISSN 1651-6214ISBN 978-91-554-9351-6urn:nbn:se:uu:diva-263206 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-263206)

To my parents and for Skadi


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Henche, A.L., Ghosh, A., Yu, X., Jeske, T., Egelman, E. Albers, S.V. (2012) Structure and function of the adhesive type IV pilus of Sulfolobus acidocaldarius. Environmental Microbiology, 14; 3188–3202
II Jeske, J.T., Centler, F., Ahmed Osman, O., Wendeberg, A., Bertils-son, S. Population-linkages in freshwater bacterial communities as-sociated with individual diatoms and particles (manuscript)
III Jeske, J.T., Müller, R.A., Wendeberg, A., Bertilsson, S., (2015) Mi-croscale decoupling of sediment oxygen consumption and microbial biomass in an oligotrophic lake (submitted manuscript)
IV Jeske, J.T., Wendeberg, A., Bertilsson, S. Bacterial communities shaping the redox environment in top-layer sediments of an oligo-trophic high latitude lake (manuscript)
Reprints were made with permission from the respective publishers.


Contents
Introduction ..................................................................................................... 9 Freshwater microbial ecology .................................................................... 9 Microhabitat heterogeneity ....................................................................... 10 Biofilms .................................................................................................... 12 Type IV pili .............................................................................................. 14 Sulfolobus acidocaldarius as a model organisms ..................................... 16 Oxygenation and Redox-state of freshwater sediments ........................... 17 Diatom bacterial interactions – Diatoms as a model system for bacteria-particle interactions ................................................................................... 20
Aims of Thesis .............................................................................................. 23
Methods ......................................................................................................... 24 Strains and study sites ............................................................................... 24 Molecular tools to assess type IV pili and diversity ................................. 26 Measuring environments .......................................................................... 28 Measuring micro-organisms in (micro) environments; a focus on Microscopy ............................................................................................... 29
Results ........................................................................................................... 31 Paper I ....................................................................................................... 31 Paper II ..................................................................................................... 35 Paper III .................................................................................................... 37 Paper IV .................................................................................................... 40
Conclusion and Perspectives ......................................................................... 45
Summary in English ...................................................................................... 49
Svensk sammanfattning ................................................................................. 52
Acknowledgments ......................................................................................... 56
References ..................................................................................................... 61

Abbreviations
BCC Bacterial community composition DNA Deoxyribonucleic acid RNA Ribonucleic acid rRNA Ribosomal RNA OM Organic matter ORP Oxidative reductive potential POM Particulate organic matter bp Base-pair EPS Extracellular polymeric substance GHG Greenhouse gas PCR Polymerase chain reaction 5-FOA 5-fluoro-orotic-acid SEM Scanning electron microscopy TEM Transmission electron microscopy GOI Gene of interest MO Microorganism Cryo-EM Cryogenic electron microscopy

9
Introduction
Freshwater microbial ecology Fresh water microbial ecology is the ecology of microorganisms in freshwa-ter habitats, such as lakes and rivers. This encompasses the biogeography, the dispersion of microorganisms in time and space, as well as processes, mechanisms, and interactions between microorganisms, and how they affect other microorganisms, trophic levels, and even whole ecosystems. Microor-ganisms are defined as organisms that cannot (with some notable exceptions) be observed without technical aids; typical they are smaller than 100 !m. By this definition Archaea, Bacteria, most unicellular Eukaryotes, e.g. yeasts, and some multicellular Eukaryotes are defined as microorganisms.
Figure 1. Conceptual visualization of carbon flow, and trophic interactions in limnic systems, together with a schematic representation of the dominant electron acceptors in the sediment. Producers are represented as green boxes, while consumers are represented as red boxes. Arrows represent general direction of carbon flow.

10
With a gross primary production estimated at 0.65 Pg C yr-1 (Pace & Prairie 2005) lakes are among the systems with the highest production per area on earth (Wetzel 2001) with large portions of this production being attributed to microorganisms. Recent research on the significance of stream ecosystems is showing similar high microbial productivity for streams and rivers. Acting as either sinks or sources of greenhouse gases (GHG), based on their geographic location, prevailing climate, nutrient inputs, morpholo-gy, and a number of other factors, lakes clearly play a central role in the global carbon cycle (Tranvik et al. 2009).
Yet, despite the growing amount of data and emergent conceptual frame-works, uncertainty and lack of mechanistic insight is often faced in research aiming at understanding the ecology of freshwater microorganisms. Hence, the microbiome is often handled as an operational “black box” with uniform features and functionality. There is clearly a need to open this frequently encountered black box if we are to have a better understanding of the micro-biology underpinning the ecosystem-scale processes characteristic for fresh-water systems and subsequently use this information in predictive models.
Microhabitat heterogeneity Habitat heterogeneity is a widely accepted concept in macro-ecology (Hughes et al. 2001; Vadeboncoeur, Zanden & Lodge 2002; Jessup et al. 2004), soil environmental microbiology (Jessup et al. 2004; Ritz et al. 2004; Jonge & Moldrup 2009; Bienhold, Boetius & Ramette 2012; Kofoed et al. 2012) and has also been highlighted and recognized in marine microbial ecology (e.g. Martiny et al. 2006; Pedro 2006; Malfatti, Samo & Azam 2010; Stocker 2012; Otto et al. 2014). Heterogeneity is caused by fragmenta-tion of different habitats, by e.g. counter-gradients of nutrients, physical disruption, turbulence, or interfaces of various sorts. In opposition to other fields of ecology, only few studies have so far specifically addressed effects and concepts of microhabitat heterogeneity in limnic systems (e.g. Vadeboncoeur et al. 2002; Maerki et al. 2004, 2009; Logue & Lindström 2008; Soininen et al. 2011).
In the water column, microhabitats are mostly shaped by turbulent flow leading to discrete and constantly changing patches of nutrients. These patches often attract, within certain limits, microorganisms capable of active motility using chemosensing mechanisms. Additionally, some microorgan-isms will enlarge their surface using filamentous appendages, such as type IV pili, or matrices of extracellular polymeric substances (EPS) (Malfatti et al. 2010), to more efficiently acquire and assimilate nutrients from the sur-rounding medium. Alternatively, some bacteria achieve close associations with particulate sources of nutrients and other resources, such as phytoplank-ton and particulate organic matter. Additional heterogeneity is introduced by

11
temperature gradients that control water density, and can lead to water mass separation during e.g. summer lake stratification, operating in parallel to depth-dependent gradients in irradiance. Other strong environmental gradi-ents are also frequently observed in lakes, e.g. haloclines and oxyclines, all of which contribute to fragmentation and patchiness of the ecosystem into distinct habitats at both the macroscopic and microscopic scales.
Figure 2. Marine microenvironments and gradients. (Top left) DOM exudation of phytoplankton, (right) cell lysis, (center bottom) stationary or sinking detritus and marine snow (Stocker 2012), original image credits: R. Stocker, J.R. Seymour and G. Gorick
Soils and sediments, although more static, often feature even greater het-erogeneity with steeper gradients and greater levels of physical fragmenta-tion, with a multitude of surfaces. They retain gas bubbles, which would quickly dissolve into, or travel through the water column. Thus, in soils and sediments, the combination of strong counter-gradients, and spatial variabil-ity of ions, and greater physical stability with constraints including: grain size, water content and porosity, create an even broader range of nutrient sources and specific niches than in the typically more uniform water column

12
(Maerki et al. 2004; Maerki, Müller & Wehrli 2006; Maerki et al. 2009; Røy, Huettel & Jørgensen 2004).
The Baas Becking hypothesis on biogeography of microorganisms states that “everything is everywhere but the environment selects” (Baas Becking 1934). This idea is questioned and challenged as often as it is promoted, but the idea of heterogeneity at microscopic scales gives a new twist. While observations in microbial biogeography often seem to contradict the Baas Becking hypothesis, they mostly do not account for microhabitat heterogene-ity on all scales relevant for microorganisms. This lack of consideration for heterogeneity at the appropriate scale becomes more poignant as microscop-ic processes potentially have significant, unaccounted effects on ecosystem scales (Green & Bohannan 2006; Benton et al. 2007).
Biofilms By definition, prokaryotes are single celled organisms. They do not contain a nucleus, and typically, but with some exceptions have no inner compartmen-talization, and they reproduce via binary cell division. This was elegantly formulated by Andrews (1998):
"At first glance, bacteria do not appear to be modular for the following rea-sons: (a) Their unit of iteration is an individual cell, not a multicellular entity, and thus is not a conventional module; (b) many bacteria are at least poten-tially motile and most are unbranched; and (c) their haploid nature and ir-regular means of sexual reproduction, including a proclivity to acquire genes by horizontal transfer mechanisms, implies that a strictly defined origin and endpoint to a genetic individual (i.e. clonal lineage) is conceptual at best."
In contrast, multicellular eukaryotic organisms typically contain a multitude of cells, highly organized, and spatially and functionally differentiated as united cell structures (tissues). Different tissues form different organs, and different organs fulfill different, specific roles for the benefit of the organism. However, this hierarchical clustering and cooperation of cells to fulfill specialized func-tions is, often mirrored in unicellular eukaryotic organisms such as algae and fungi, and in prokaryotic organisms, with with pertinent examples including filamentous Cyanobacteria, Myxococcus fruiting bodies, and Streptomyces mycelia. In close cooperation, united cell structures allow otherwise single-celled organisms to act in unity and fulfill a variety of different metabolic and physiologic tasks that require spatial separation and differentiation. Often, these multicellular bacterial assemblages that depend on a subset of properties, are referred to as biofilms, as they were first termed in 1978 (Costerton, Geesey & Cheng 1978). This concept of biofilms contradicts the theoretical framework of prokaryotes as purely unicellular organisms. Biofilms have been shown to be abundant in virtually every environment and often surpass plank-

13
tonic counterparts by orders of magnitude both in numbers and biomass (Torsvik, Øvreås & Thingstad 2002). Furthermore biofilm constituents may differ entirely from their planktonic counterparts in terms of transcribed genes (Donlan 2002). In fact consortia of bacteria are very abundant in nature, either as single species associations ranging from diplococcoid cell pairs to entire complex biofilms. On greater scales these associations even include complex interspecies relationships encompassing multiple species and entire food webs, often enclosed in volumes as little as few mm³, e.g. microbial mats on intertid-al mudflats (Olff et al. 2009).
But what really is a biofilm? The crudest definition would be that a biofilm is any assembly of multiple microbial cells, embedded in a self-produced, extracellular matrix often referred to as EPS, attached to a surface (Whitfield & Keenleyside 1995). These surface-associations are typically mediated by filamentous appendages such as flagella and type IV pili (Donlan 2002; Vu et al. 2009). Additionally, EPS matrices typically contain polysaccharides, pro-teins, DNA and RNA, and filamentous appendages that facilitate adherence. Secretion of such matrices is also a very effective means to avoid desiccation, retain extracellular enzymes, increase resistance to antibiotics and toxic com-pounds, and improve bacterial resistance to environmental stress (Costerton 1995; James, Holmström & Kjelleberg 1996; Watnick & Kolter 2000; Hall-Stoodley, Costerton & Stoodley 2004; Vu et al. 2009).
Adaptation of biofilm structure for survival in varyingenvironments. Intriguingly, the visual characteristicsof biofilms growing in diverse environments arestrikingly similar, indicating there are important con-vergent survival strategies that are conferred in part bystructural specialization (FIG. 2).
Biofilms growing in fast-moving water tend to formfilamentous STREAMERS regardless of whether they occurin the drainage run-off from acid mines12, in hydro-thermal photosynthetic mats (algal or bacterial)6 or asPERIPHYTON in rivers (FIG. 2). In quiescent waters, biofilms
the Korarchaeota and Aquificales respectively8,9.Taken together, the data indicate that the ability toform biofilms is an ancient and integral characteristicof prokaryotes. In the context of evolution and adap-tation it is likely that biofilms provided homeostasisin the face of the fluctuating and harsh conditions ofthe primitive earth (extreme temperatures, pH andexposure to ultraviolet (UV) light), thereby facilitatingthe development of complex interactions betweenindividual cells and providing an environment whichwas sufficient for the development of signalling
R E V I EW S
Propelled by shear forces, aggregatedcells can become detached, or roll or ripple along a surface in sheets and remain in their protected biofilm state.
‘Wall formers’
‘Dispersers’
‘Persisters’
Starvation can induce bacteriato shrink and adopt a spore-likestate, known as ultramicrobacteria,which wait in water, soil, rock ortissue until conditions aresuitable for active growth.
1
Chemical gradients createmicroenvironments fordifferent microbial speciesor levels of activity.
6
8
Although antimicrobials damageouter cell layers, the biofilmcommunity is resistant.
7
The close proximity of cellsin the matrix facilitates the exchange of molecular signals that regulate behaviour.
5Nutrients diffuse into the matrix.
4
Active bacteria can attach to almostany surface. Changes in gene expression transform ‘swimmers’ to ‘stickers’ within minutes.
2
Attached bacteria multiply andencase the colonies with a slimy matrix.
3
Figure 1 | Conceptualization of biofilm development and dynamic behaviours. The figure was compiled from laboratory and natural observations of pureculture (both Gram-positive and Gram-negative organisms), and mixed-culture biofilms. (For an interactive web-based version of Figure 1 and biofilm movies showingdynamic processes of growth and detachment, rolling and rippling, see the Online links). Image courtesy of P. Dirckx, Center for Biofilm Engineering, USA.
Figure 3. Conceptual representation of biofilm formation, compiled from observa-tions of single-species cultures (Gram positive and gram negative bacteria), and mixed biofilms. Reprint with permission from (Hall-Stoodley et al. 2004).

14
The abundance of microbial biofilms in natural systems has long been un-derestimated, but advances in methodology have shown that even on the smallest particles, e.g. marine snow and other microscopic particulate organ-ic matter, biofilms are frequent (Malfatti et al. 2010; Khandeparker et al. 2013) and have great impact on biogeochemical processes of ecosystem scale relevance (e.g. Battin et al. 2001; Donlan 2002; Besemer 2015).
Type IV pili Bacteria possess a multitude of proteinaceous surface structures allowing them to interact with their surrounding environment. Well studied microbial surface structures include the bacterial flagella and its proposed predecessor the bacterial type III secretion system (Aizawa 2001; Blocker, Komoriya & Aizawa 2003), type II secretion system (Peabody et al. 2003), type IV pili and their archaeal homolog the archaellum (Albers & Jarrell 2015). Bacterial type IV pili have been proposed to be the bacterial “Swiss army knife”, ful-filling a multitude of functions, ranging from surface attachment, with func-tional and structural roles in biofilm formation (O’Toole & Kolter 1998; Shime-Hattori et al. 2006; Varga et al. 2006), to cell-cell contact, exchange of nutrients or secondary metabolites, DNA uptake (Aas, Løvold & Koomey 2002; Averhoff & Friedrich 2003; Averhoff 2004) to functions as complex as predation (Lambert et al. 2008; Mahmoud & Koval 2010), virulence (Paranjpye et al. 2007) or motility (Wall & Kaiser 1999; Mattick 2002). It was recently shown that archaeal type IV-like pili (archaellum) are involved in swimming motility, gliding motility (Albers & Jarrell 2015), surface at-tachment and biofilm formation (Henche et al. 2012). All of these functions are pivotal for the respective microorganisms to interact with their surround-ing environment and other organisms. However, pili and filamentous ap-pendages are often costly in terms of gene expression and protein synthesis, and are often equally easily lost as acquired. Type IV bacterial pili are typi-cally thin (5-8 nm in diameter) (Soto & Hultgren 1999), proteinaceous fila-ments that can extend from barely protruding the bacterial cell envelope to several micrometers (Soto & Hultgren 1999). Reference values for the max-imum length of type IV pili remain to be defined, but filaments exceeding several µm in length were frequently observed in environmental samples (Soto & Hultgren 1999; Craig et al. 2006).
Due to its unique machinery of assembly, the bacterial and archaeal type IV pili can be produced and disassembled by the cell at high speeds allowing rapid protrusion and retraction often used for gliding, swarming, and twitch-ing motility (Kohler et al. 2000; Mattick 2002; Anyan et al. 2014). Type IV piliation systems are widely distributed amongst all bacterial phyla, and their similarity to archaeal homologs suggests an early evolutionary origin. How-ever, precise genetic dating of its origin is hampered by the frequent in-

15
volvement of type IV pili in horizontal gene transfer. The evolutionary origin of the type IV piliation system is unknown but several of the key components in the type IV pilus assembly systems also have homologs in type II secretion, and archaellum assembly systems, suggesting a common origin is likely (Albers, Szabó & Arnold 2003; Craig, Pique & Tainer 2004; Johnson et al. 2006; Albers & Meyer 2011)
Outer membrane
Peptidoglycan
Innermembrane
1
2
3
6
7
5
84
Figure 4. Schematic type IV pili assembly machinery in gram negative bacteria. 1) Prepilin with class III signal peptide and α-helical domain for membrane insertion. 2) Membrane inserted Prepilin Peptidase, which cleaves the membrane insertion site at the class III signal sequence of the prepilin. 3) Pilin subunit to be inserted into the pilus, with N-terminal α-helical domain facing into the core of the growing pilus (4), and the globular C-terminal domain facing outwards. 5) Membrane anchoring pro-tein. 6) and 7) ATPases for assembly/disassembly of the pilus. 8) Outer membrane porin.
The pilus consists of polymers of amphipathic pilin proteins, with a hydro-philic C-terminal part facing outwards, and a hydrophobic N-terminus facing inwards, which is important for assembly and structural integrity of the pilus. Pilins are inserted into the growing pilus as prepilins, by means of a class III signal sequence (Szabó et al. 2007) that is cleaved of after membrane inser-tion by a peptidase (Albers et al. 2003). Energy for the protrusion of the growing pilus is provided by membrane associated ATPases. The pilus is anchored to the membrane via an integral membrane protein. The pilin,

16
membrane anchor protein and ATPase form the core assembly mechanism. Additional chaperones and outer membrane proteins are typically required for pilus assembly (Figure 4). And the total number of proteins involved in pilus formation typically ranges between 10 and 18 different proteins (Peabody et al. 2003). Opposed to the bacterial flagella with its hollow cen-ter, the inner body of the bacterial pilus is densely packed, so transport of nutrients has to occur along the C-terminal parts of the pilin proteins, which are, unlike their N-terminal parts and the type III insertion sequences highly variable.
The most recent addition to the list of type IV pili functions is the pro-posed function as electron-conductive, extracellular filaments, i.e. nanowires that allow electron shuttling between bacteria and their environment (Lovley 2012a). Geobacter metallireducens has repeatedly been shown to produce electrically conductive type IV like pili, nanowires, that when in contact with insoluble Fe(III) compounds, can reduce these to Fe(II) and harvest energy in the process (Lovley 2012a b). It moreover allows them to have an excep-tional versatility in the variety of metallic compounds that they are capable of reducing, ranging from Fe(III) to Mn(IV), even allowing them to precipi-tate uranium which could make them a key candidate for bioremediation of heavy metal contaminated aquatic habitats (Lovley et al. 2011). Geobacter species, including Geobacter sulfurreducens are widely used in microbial fuel cells based on their exceptional performance in metal reduction and their unique cell membrane characteristics, that enable easy passage of elec-trons in and out of their cell envelope (Srikanth et al. 2008).
Sulfolobus acidocaldarius as a model organisms Model organisms are organisms with a set of properties that make them es-pecially well suited for being studied under controlled conditions. In micro-biology, this typically means organisms that are susceptible to genetic modi-fication, which allows manipulating genes and testing their respective func-tion in an environment that allows monitoring of how these dele-tions/insertions/modifications affect their phenotype. There is an abundance of bacterial, and even eukaryotic model organisms, but archaeal model or-ganisms are heavily underrepresented compared to their abundance in na-ture. Often archaea are genetically inaccessible because of our incomplete understanding of their unique transcriptional characteristics, i.e. translational and transcriptional machinery that possesses characteristics of both Eukary-otes and Bacteria. Notably in this context is their RNA-polymerase II appa-ratus, which is similar to eukaryotic homologs, but regulated in a bacterial fashion, including a TATA-box binding protein and a transcription factor B being similar to bacterial ones (Bell and Jackson 2001), to name just a few.

17
First description of Sulfolobus was in 1966 from acidic ponds in the Yel-lowstone national park (Dworkin et al. 2006), and was fully described in 1973 (Brierley & Brierley 1973) – consequentially the strain was named Sulfolobus acidocaldarius. In short succession to that, a similar species was described, found in solfataric craters from Naples, Italy, first called Caldari-ella acidophila (de Rosa et al. 1974; de Rosa, Gambacorta & Bu’lock 1975), but later renamed to Sulfolobus solfataricus (Zillig et al. 1980). Those two species are highly similar, in terms of their habitat, ecological niche and metabolic properties, although S. solfataricus can utilize a wider array of nutrient sources. They both live at an optimal growth temperature between 65°C and 90°C, with an environmental pH of between pH 1 and pH 6. In fact their environmental niche is so similar, that they were often found coex-isting in the same habitat. Yet, later sampling efforts for S. acidocaldarius were often unsuccessful. As it turns out, this lack of success in sampling was directly attributed to the sampling methods, which, at the time, typically were to sample for planktonic organisms in the water, which only harvested S. solfataricus (S. Albers, personal communication). In contrast, S. acidocal-darius was later found, when sampling the, biofilm-rich, near-shore areas of the same craters and ponds, indicating preference for a much less planktonic way of life. Fortunately, both those thermo-acidophilic crenarchaeotes (S. solfataricus and S. acidocaldarius) have properties suitable for a model or-ganism, and have in the past allowed researchers to have a closer look on some of the unique, properties that differ in archaea, or those they have in common with eukaryotes or bacteria, enlarging our understanding of all three domains of life (Woese & Fox 1977).
As Sulfolobales are Archaea and live in hot and highly acidic environ-ments, antibiotics, which are typically used for genetic modification, and screening for the generated mutants, cannot be employed as a tool for work-ing with Sulfolobus. In order to cope with this methodological speed bump, and in order to make Sulfolobus accessible to genetic modification a shuttle vector based on the S. solfataricus-specific virus SSV1 was established, to allow expression in Escherichia coli, and S. solfataricus and S. acidocaldar-ius. Employing this plasmid, a ∆pyrE-Sulfolobus-strain, carrying a uracil auxotrophy, was created (Jonuscheit et al. 2003; Schelert et al. 2004; Berkner et al. 2007; Albers & Driessen 2008; Berkner & Lipps 2008). In-frame deletion mutants are then selected utilizing an introduced uracil auxo-trophy in combination with 5-FOA-addition in a two step selection process (Grogan & Gunsalus 1993; Wagner et al. 2009, 2012).
Oxygenation and Redox-state of freshwater sediments Sediments have been recognized for their importance in biogeochemical processes since the early days of limnology (Lindeman 1942). They are of

18
crucial importance for lake carbon turnover, as well as burial, and long time storage and resuspension of nutrients. Their contribution to GHG emissions often depends on water depth and nutrient loading of the overlying water column and in fully oxygenated and shallow lakes, carbon mineralization is often favored over burial (Tranvik et al. 2009).
Unlike the water column, sediments can physically retain gas bubbles and store nutrients. In combination with the generally higher physical heteroge-neity and patchiness of sediments and spatial variability in oxygen, ion, or nutrient concentrations, grain size or porosity multitude of unique niches is created for microorganisms to exploit (Maerki et al. 2004, 2006, 2009; Røy et al. 2004). Another key aspect of sediments, differentiating them from the more uniform water column is their relative abundance of surfaces. This abundance of niches, and richness of nutrients in comparison to the water column, results in cell densities in sediments exceeding those found in the water column by orders of magnitude. Individual metabolic activities how-ever, may differ significantly between water column and sediment, and are typically higher in planktonic microorganisms. Yet, the shear abundance of biomass and nutrients typically makes sediments a hotspot for whole lake processes and carbon turnover. Biomass of microorganisms in sediments is often as high as combined biomass of all other benthic organisms (Meyerreil 1994; Haglund et al. 2002; Torsvik et al. 2002).
To gain energy from the abundant nutrient sources in sediments, bacteria need electron acceptors, with the most thermodynamically favorable one being oxygen (Fenchel & Finlay 2008). Yet, with the gradual depletion of oxygen over depth in the sediment, the availability of alternative electron acceptors, in a since long recognized sequential order, becomes an additional selective factor for microbial community structure (Nealson 1997)
Alternative electron acceptors are, as the name suggests, necessary for microorganisms to harvest (accept) the surplus in electrons, arising from the oxidation of reduced organic and inorganic compounds, while generating protons to drive the concentration-gradient based proton pumps. Thermody-namically, the chemical reactions microorganisms can utilize to store energy can only occur when the resulting products are energetically more feasible. This difference in energy between the educt and the product can be ex-pressed as electric potential differences, the redox potential, typically given as ∆E0´(V) between the oxidized and reduced terminal electron acceptor. One of these environmentally important alternative electron acceptor pairs is SO4
2-→HS- ~ -0,22V. Other electron acceptor pairs, from the most oxidized to the most reduced, and thus increasing in potentially available energy are CO2 / CH4, ~ -0,25V, SO4
2- / HS-, ~ -0,22V, NO2-, N2O, N2 / NO3
-, ~ +0,4V and Fe3+ / Fe2+, ~ +0,75. Fe(III) reduction stands out with its comparatively high potential to accept electrons which is close to that of oxygen (H2O/O2 +0,82V) and is typically the first alternative electron acceptor to be metabo-lized along the oxic-anoxic continuum. Furthermore, dissimilatory reduction

19
of Fe(III) to Fe(II), and the closely related reduction of Mn(IV) to Mn(III), is possibly the most ancient biologically driven reductive process with global significance (Lovley 1991). However, opposed to for example NO3
- or CO2, Fe(III) is almost insoluble, and easily precipitates in aquatic environments, and thus necessitates adaptive mechanisms to utilize Fe(III) and make it bioavailable as an electron acceptor.
The frequent insolubility of electron acceptors, as exemplified above for Fe(III) requires mechanisms to access and utilize solid matter, which is often done by microbial cells via direct contact with the surface, also termed sur-face attachment. Consequently, most organisms found in sediments will readily attach to surfaces. Mechanism of surface attachment often employ filamentous appendages, such as type IV pili or flagella, as well as matrix-based mechanisms of attachment using EPS (McBride 2001, 2004; Klausen et al. 2003; Burrows 2012).
With growing knowledge and understanding of sediment biogeochemistry and filamentous appendages, a conceptual framework was built and termed electromicrobiology (Lovley 2012a). Electromicrobiology has lead to the dis-covery of bacteria being capable to access extracellular sources of protons and electrons via physical contact. This allows utilizing even insoluble electron acceptors readily, and over long distances. Simultaneously filaments and fila-mentous appendages allow transport of electrons in dense networks, e.g. in Geobacter, (Lovley 2012b) or as frequently observed in the world wide abun-dant, filamentous cable bacteria (Pfeffer et al. 2012; Malkin et al. 2014; Vasquez-Cardenas et al. 2015). Electron shuttling is a third alternative for electron transport, employing secondary metabolites to transport electrons from one point in space to another, which is of special importance in biofilms (Straub & Schink 2003; Kappler et al. 2004; Paquete et al. 2014). Especially those secondary metabolites would have a visible effect on the redox envi-ronment, apart from depletion of reduced compounds, secondary effect on the redox environment, detectable in pH- and redox-fingerprinting of sediments. Redox and pH fingerprinting have also been shown to be suitable methods to detect cable bacteria (Larsen, Nielsen & Schramm 2015; Nielsen & Risgaard-Petersen 2015).
Such direct correlations of oxidative reductive potential (ORP) with bio-logical activity have been verified for laboratory cultures of single organisms and mixed communities (Escalante-Minakata et al. 2009; Rosu, Murguía & Ibarra-Junquera 2010; Hunting & Kampfraath 2013; Hunting et al. 2015), but the respective behavior in sediments is still not fully understood. On larger spatial scales however, it is still generally assumed that environmental properties and respiratory depletion of electron acceptor reservoirs, controls ORP (Jacob 1970; Revsbech et al. 1983; Vorenhout et al. 2004).

20
Diatom bacterial interactions – Diatoms as a model system for bacteria-particle interactions Diatoms (Baccilariophyceae) are a major group of algae, known since the beginning of the “age of microscopy” due to their unique morphology, and quite simple beauty when observed under the microscope. This beauty lies in their distinctive, transparent, SiO2-based, two-partite, shell (Figure 5.), the frustule, which allows to identify species, and serves as a room for diatoms to interact with bacteria (Kaczmarska, Ehrman & Harris 2005; Armbrust 2009). This space for diatom-bacteria-interactions is called the phycosphere. Like all algae, diatoms are photoautotrophs. Yet, despite their ability to gain energy directly from sunlight, they often rely on syntrophic bacterial partners to supply them with essential vitamins, e.g. vitamin B12 (Carlucci & Silbernagel 1969; Guillard & Kilham 1977; Droop 2007; Amin, Parker & Armbrust 2012).
Diatoms are abundant in virtually every aquatic system, but estimating their biomass is often hampered due to high levels of patchiness and season-ality. It is estimated that they are responsible for up to one fifth of the global primary production and between 20 and 40% of the earth’s oxygen-production (Falkowski 1998). In light of the massive amounts of carbon fixed in and by diatoms, their influence on nutrient cycling, GHG-emissions and carbon-fixation is undeniable. It is thus of pivotal importance to better understand diatoms in the future (Schindler et al. 1997). However, their im-pact on humans is not reduced to GHG dynamics, and they directly influence factors as diverse as economics and human health, as well as tourism and fisheries. One example for this socio-economic impact of diatoms is their affiliation with amnesic shellfish poisoning via the eco-toxins they excrete, causing diseases after direct or indirect ingestion (Granéli & Turner 2006).
Planktonic diatoms depend on water mixing to stay suspended in the pho-tic zone, and generally sink at low turbulence. This results in short blooms followed by rapid declines in biomass, which makes them an ideal model to study patterns of colonization and short term effects on primary production, GHG emission, as well as nutrient transport and sedimentation processes. This is particularly true for dimictic lakes, characterized by periods of sea-sonal mixing and stratification. In such systems, their rapid growth rate often makes them the first abundant primary producer in early spring, or enables them to outcompete other phytoplankton in during autumn mixing. This rapid primary production has tremendous ecological impact, as it represents the “kick-off” for all processes linked on higher trophic levels in such sys-tems. This seasonality is not only apparent at the whole lake level. Species succession between different species can also be observed (Stoermer 1993; Köster & Pienitz 2006). In combination with environmental data, this makes it possible to study detailed succession patterns and patterns of response to environmental disturbances.

21
FIG 1 Micrographs of representative diatom species. (A) Light micrographs of diatoms. Clockwise from the top left corner: Striatella unipunctata, Odontella sp.,Stephanopyxis turris, Pseudo-nitzschia sp., Thalassiosira sp., Cylindrotheca sp., Asterionellopsis glacialis, Skeletonema costatum, Grammatophora oceanica, and Chaetocerossp. Images are courtesy of Colleen Durkin. (B) Scanning electron microscopy (SEM) images of diatoms. Clockwise from the top left corner: Didymosphenia geminate(Lyngbye), valve view; Lauderia annulata with bacteria, girdle view showing attachment of two cells; Thalassionema nitzschioides (Grunow), valve view showing therounded valve end; Coscinodiscus sp. after sexual reproduction in a culture restored large cells (top) from small gametangial cells (bottom); Chaetoceros didymus withbacteria; Asterionellopsis glacialis with bacteria; Actinoptychus senarius; Lithodesmium undulatum, girdle view; and Stephanopyxis turris (Greville) (center). Images arecourtesy of Julie Koester (Coscinodiscus sp.) and Mark Webber (the rest).
668 mmbr.asm.org Microbiology and Molecular Biology Reviews
on July 16, 2015 by Uppsala U
niv BMC
http://mm
br.asm.org/
Figure 5. Scanning electron microscopy images of diatoms. Top left: Didy-mosphenia geminate, top middle, Lauderia annulata, top right: Thalassionema nitzschoides, middle left: Lithodesmium undulatum (girdle View), center: Stephano-pyxis turris, middle right Cosciodiscus sp., bottom left: Actynoptichus senarius, bottom middle: Asterionellopsis glacialis, bottom right: Chaetoceros didymus. (Amin et al. 2012). Notably, on L. annulata, A. glacialis, and C. didyemus associat-ed bacteria are clearly visible.
Often this succession in phytoplankton community, including diatoms, is matched by seasonally reoccurring changes in bacterial community composi-tion, associated to the phycosphere (Bell & Mitchell 1972; Kirschner & Velimirov 1997; Grossart et al. 2005; Rösel & Grossart 2012). This implies close functional interactions between bacteria and phytoplankton; yet, the nature of many of these interactions remains undisclosed. There are evidenc-es that many of them, are commensal or mutually beneficial (Sapp, Wichels & Gerdts 2007; Sapp et al. 2015; Bruckner et al. 2008; Beier & Bertilsson 2011). Additional evidence for how common diatom-bacteria interactions are, comes from the diatom genome itself, which can consist of as much as 5% of bacterial sequences. Such a large fraction of genetic material originat-ing in another organism indicates frequent DNA exchange between diatoms and their associated microbiome (Armbrust et al. 2004; Bowler et al. 2008). Furthermore, it could be demonstrated that diatoms specifically react to cer-tain bacteria associated to them, e.g. by changing the type of exudates they produce (Myklestad 1995). This encompasses severe side effects for hu-mans, as it includes toxin production (Sison-Mangus et al. 2014). Simulta-

22
neously, diatom-bacterial interactions can be disadvantageous for the dia-toms, due to effects like infection, predation, or even simple physical effects like increased sedimentation (Gärdes 2010; Gärdes et al. 2011).
The bacterial community associated with diatoms has typically been de-scribed based on non-axenic cultures (Schäfer et al. 2002; Grossart et al. 2005; Kaczmarska et al. 2005; Bruckner et al. 2008), with some exceptions trying to resolve bacteria-particle associations on a single diatom (particle) level (Malfatti et al. 2010; Cai et al. 2014; Amin et al. 2015). These studies typically represent the diatom associated bacterial community to consist mostly of heterotrophic Alpha-, Beta- and Gammaproteobacteria, with large fractions of Bacteroidetes, and Verrucomicrobia. Such a bacterial communi-ty composition is in contrast with what is frequently described from bulk water samples, that are typically dominated by Actinobacteria as well a Al-pha and Gammaproteobacteria. Yet, Actinobacteria were also found in fre-quent association to diatoms, however not closely attached, but seemingly more in a lose, commensal mode described as `cheating´ (Beier & Bertilsson 2011), whereas Bacteroidetes and Proteobacteria are not only typically closely associated, but were demonstrably shown to be capable of directly affecting which exudates are being produced by the diatoms.

23
Aims of Thesis
The main goal of my thesis is to advance our appreciation and understanding of microbial responses to micro-scale habitat heterogeneity. This encom-passes the attempt to better understand mechanisms and effects of especially biofilm formation and redox and oxygen interfaces in aquatic systems. In order to do so, questions were asked concerning some of the molecular mechanisms involved in microbial surface interactions, but also species-species, and species-environment interactions.
In Paper I, the specific aim was to resolve the assembly machinery, func-tion and structure of the type IV like pilus, involved in surface attachment, in the thermo-acidophilic model crenarchaeon Sulfolobus acidocaldarius. In Paper II, the main goal was to identify and characterize, with a focus on diatoms, the particle-associated microbiome in a dimictic lake and resolve what the driving factors for bacterial community composition are with re-gards to the nature of the particle. In Paper III oxygen consumption rates, as a proxy for total metabolic activity of a bacterial community, were assessed in sediment microcosms representative for a high-latitude, oligotrophic Lake. This was done to evaluate individual activities of bacteria, how they change with cell densities and community composition, and which environ-mental factors are the main drivers. Finally, in Paper IV the focus was on identifying to what degree bacterial community composition in the top layer sediments correlated with the prevalent redox-state of said sediment, and to which degree biotic or abiotic factors influence both observations. Addition-al focus was set on how the bacterial community composition is shaped in relation to habitat heterogeneity, and whether the redox state could be caus-ally linked to individual parts of the BCC.
My strategy was "bottom up" looking first at the molecular underpinning to biofilm in a “single” microorganism, then increase the scale to look at how multiple bacterial cells interact on a “single” particle, then to eventually trace these particles en route to the sediment, where spatially resolved asso-ciations between the microscale environment and microbial communities were studied further. This research gives an into habitat heterogeneity and what defines a habitat on scales relevant for microorganisms, allowing better predictions of microbial processes and adaptations resulting from a hetero-geneous world.

24
Methods
Strains and study sites Sulfolobus acidocaldarius Type IV piliation systems are well studied in bacteria, but have only recently been discovered in Archaea. The work presented in Paper I used Sulfolobus acidocaldarius MW001 to study archaeal piliation, not exclusively, but es-pecially, from the perspective of surface adhesion, motility and biofilm for-mation. Strain MW001 is a uracil auxotrophic mutant of the S. acidocaldari-us wild type, containing a disrupted pyrE gene, responsible for de novo ura-cil synthesis. This uracil auxotrophy allows screening for successful integra-tion of a plasmid containing a pyrEF cassette. A two-step selection method is used, wherein the first step the plasmid is integrated into the genome via homologues recombination after electroporation. Subsequently. The pyrEF genes within the cassette are flanked by inverse repeated regions, which are mirrored on both ends of the plasmid. Incubation in media containing (5-fluoro-orotic-acid), but no additional uracil, forces a second recombination event that will lead to a deletion of the gene of interest, or a reversal to the wild type, which is tested using PCR. Lake Erken For my attempt to unravel microbial interactions on particles, including dia-toms, and their associated bacterial communities (Paper II), I selected Lake Erken (Lat: 59.83653 Long: 18.64212), Stockholms Län, Sweden as my study system. The main reason for selecting this lake was the already estab-lished environmental monitoring program at Lake Erken and the later inte-gration in SITES (Swedish Infrastructure for ecosystem services). It is one of the best-studied lakes in the world, with vast amounts of data available on biotic, as well as abiotic factors over extended periods of time. This in turns allows for easy comparison with other studies, among others, trying to assess planktonic communities across all three domains of life and how such com-munities and populations react to environmental change. Lake Erken is also dimictic with pronounced seasonality and for temperate lakes representative thermal stratification during summer and winter while completely mixing during spring and autumn. It is thus a good model to study species succes-sion in microbial communities, and here specifically bloom forming diatoms and their associated bacteria.

25
Lake Ånnsjön Lake Ånnsjön sediments were used as a model lake to study cell-number correlated oxygen consumption rates (Paper III), and the relationship be-tween bacterial community composition, and cell abundance and the redox state of their environment (Paper IV). Clearwater Lake Ånnsjön was select-ed because of the geomorphological interesting features and its oligotrophic state, as well as availability of environmental data. Unlike eutrophic sedi-ments, the oligotrophic sediments of Lake Ånnsjön feature an oxic-anoxic continuum that is stretched across longer spatial scales enabling acquisition of highly spatially resolved cell abundances and BCC trends, which could not be readily assessed in more productive systems. Lake Ånnsjön is located in Jämtlands Lan, Sweden, 63°16’N, 12°33’E, 525 m above sea level, em-bedded in the mountainous and alpine, border-region between Sweden and Norway (Figure 6). It is fed by the two mountain-rivers Enan and Handölan, connecting the lake with its 1.564 km2 catchment area. The average yearly discharge is 40 to 60 m3 s-1, peaking after snowmelt, and a theoretical reten-tion time of 176 days. The catchment area comprises 14.3% wetlands, 21.3% forested areas, and 61.8% open land. Wide outcrops of crystalline bedrocks and thin patches of soil over quaternary moraine, and more recent debris are found in abundance across the catchment, including large areas of ultrabasic, iron-rich rock-formations, (e.g. Amphibolite), as well as quartz-feldspar, and mica rich regions in the south.
Figure 6. Map of Lake Ånnsjön with dominant habitats indicated by different color-ations. Sampling sites are marked as bottom (B), shore (C), and inflow (D).

26
The Ånnsjön setting allowed us to simultaneously study and compare highly dissimilar, yet connected lake habitats. These habitats included the benthic shore-region, the inflow and the bottom. The shore region, comprising 31.5km2, thus making up more than half of the lake, was defined as a shal-low areas (<2m water depth), with intermitted macrophyte growth, especial-ly in the north. The 3.4 km2 inflow was similar to the shore in mostly featur-ing shallow areas, yet with much higher densities of island-like accumula-tions of macrophytes, and unlike the typically sandy shallow areas, having elevated amounts of sedimented particulate organic matter. The bottom area, making up the remaining 22.7km2 of the lake, was characterized as being deeper than 2 m, with a maximum depth greater than 15 m water depth at its maximum in the southern parts of the lake.
Molecular tools to assess type IV pili and diversity Polymerase chain reaction Polymerase chain reaction (PCR), is a standard molecular tool widely used in all fields of biology and so also in all four papers included in this thesis (Paper I-IV). The principle of PCR relies on synthesis of new DNA strands using a (typically) DNA-Polymerase, together with a buffer solution and nucleotides, in combination with a pre-synthesized pair of forward and re-verse primers, which are needed to initiate the amplification. These short oligonucleotides (primers) that frame the ROI (region of interest) will specif-ically bind to homologous sequences on the target template. Aligning is temperature sensitive, and the specificity of the amplification relies on ap-propriate temperatures ensuring, that only the GOI is amplified. The result-ing amplified DNA fragments can then be recovered and characterized in down-stream applications. Functional assessment of archaeal type IV pili Similar to PCR, cloning is now a standard method widely used in biological research. Cloning in microorganisms, such as archaea and bacteria, relies on the capacity of cells to take up and incorporate external DNA in the cell. Such material is typically introduced in plasmids, via the same mechanisms exploited in PCR. Plasmids are small circular DNA elements that contain selection factors, origin of transcription, terminator sequences and several restriction sites. These plasmids can be transcribed individually, or, if de-sired, integrated into the target organism’s genome based on homologous recombination.
In the herein presented study (Paper I) a suicide vector plasmid was used, which could be amplified and methylated in Escherichia coli, prior to inser-tion via electroporation (Berkner 2007) into S. acidocaldarius MW001. There it would integrate into the genome based on two homologous recom-

27
bination events, thus deleting the GOI (Wagner 2012). Due to the nature of the marker-less in-frame deletions in S. acidocaldarius recombination events were checked using PCR.
By deleting the other two, archaeal filaments from the S. acidocaldarius genome, the aap-pilus could be harvested from liquid cultures and thus be used for characterization with Cryo-EM. Extraction of the pilus was done, by removing the pilus from the cell by vigorous shaking and consecutive re-peated centrifugation, and density-gradient centrifugation. Sequencing, Statistical analysis, Phylogeny, and data handling Bacterial community analyses Bacterial communities were analyzed from PCR-amplified universal markers (a variable portion of the 16S rRNA gene) using parallelized next generation Illumina sequencing for the readout (Paper II-IV). Illumina-sequencing is a high throughput sequencing method, which, in its core, is a next generation Sanger sequencing. Fluorescently labeled nucleotides are incorporated into a single stranded, matrix-attached template. After each deoxynucleoside tri-phosphate (dNTP) addition, the fluorescence signal is recorded, thus deter-mining which type of dNTP was added to the growing strand. The nucleotide label itself serves as a terminator for polymerization, which is consecutively cleaved of enzymatically to allow incorporation of the next nucleotide. For the sequencing performed here, I used a two-step PCR reaction, with and without barcoded primers, prior to sequencing. The primers used were standard PCR primers targeting 16SrRNA gene from the entire bacterial domain. The 16SrRNA gene is an often used, well-suited phylogenetic marker, as it is present in all organisms, and contains both variable and high-ly conserved regions. Using phylogenetic analysis of the obtained sequences makes it possible to perform taxonomic identification and reconstruct the evolutionary relationships between organisms. Phylogenetic analyses were performed using publically available online tools, such as clustalW, selected for their robustness and wide use in working with 16SrRNA data.
Similar to the other applied methods, I tried to use conservative ap-proaches in data handling and statistical analysis, to ensure better compara-bility with other, published and future, studies. The general treatment of the sequencing data was done, following the procedures of Sinclair et al. (2015). We used Bray Curtis dissimilarity distance measures for all (permuted mul-tivariate) analysis of variance, as well as the non-metric multidimensional scaling analysis. Co-occurrence network analysis were done on presence absence and Raup-Crick dissimilarity, weighing the observed occurrence of an OTU versus the probabilistic occurrence of that OTU in the dataset.
Diversity indices and analysis of diversity, as well as all statistical ap-proaches were done using PAST, and the R-environment. The α-diversity indices used were Fisher’s α-diversity, Simpson’s index, Shannon-Wiener index, as well as Pielou’s index. The combination, and internal comparison,

28
of these indices allowed to best account for bias introduced to the diversity models by differing and/or low species numbers (Shannon 1948; Hammer, Harper & Ryan 2009; Oksanen 2015). Estimates of beta diversity used Eu-clidean distance models, as a robust method to determine difference in vari-ance between the samples and their group centroid (Oksanen 2015).
Measuring environments Measuring microenvironments A large share of this work (Paper III and IV) is centered on the ability to resolve microenvironments and habitat heterogeneity. Such an attempt bears with it a common problem in ecology: processes taking place and manifest-ing at different scales. I used microsensors to assess redox potential and ox-ygen concentrations in the sediments ensuring minimal possible disturbance and maximal spatial resolution. Due to the minimal destructive nature of this approach it also allowed for parallel measurement of biological aspects such as cell numbers and bacterial community composition. Due to their common use, and applicability, the methods used can be readily compared to contem-porary and future studies.
Oxygen levels and redox state of the sediments were assessed with com-mercially availably microsensors. These microsensors allowed for a real-time resolution at micrometer scale, without prior manipulation of the sam-ples. The main problem, however, is that it does not allow for repeated measures at later times, as the microsensors introduce channels into the sed-iment which offsets oxygen saturation levels and redox state, once the sensor is retracted again. Typically such measurements would be made in undis-turbed sediments cores, yet, I decided to use microcosms of artificially re-constructed sediments. The rationale for this was that only single point measurements are possible in sediment cores because of spatial limitations caused by the upper diameter of the microsensors and attached machinery. Hence it is then virtually impossible to identify within- and between -variation in environmental parameters. The microcosms, on the other hand, allow for multiple measurements in close proximity, thus finding averages for larger parts of sediments, and simultaneously identifying the amount of individual variance. Measuring macroenvironments A similarly generalized approach was taken to measure remaining environ-mental properties, using standard techniques that can easily be reproduced, attempting to get as high vertical resolution as practically possible. I decided to measure total Carbon, Nitrogen, and Phosphorus concentrations from freeze-dried sediment sub cores. This allowed resolving those basic nutrient levels at a resolution of 0.5 cm, same being true for water content of the

29
sediment. Finally, inorganic nutrients, as well as TOC, TIC and TN were measured from pore-water. Extraction methods for these sediment character-istics did unfortunately not allow for a resolution similar to the other men-tioned environmental properties. In summary, the overall combination of applied methods, and the results they produced, allows for simple reproduc-tion and application, and testing of the here presented hypothesis in future studies, trying to evaluate if the principles presented hold true for different systems.
Measuring micro-organisms in (micro) environments; a focus on Microscopy The ability to resolve structural features of microbial communities, single microorganisms and subcellular features was central to this thesis. In order to do so, different microscopic techniques were applied, each with its own strengths and limitations. To better understand the choice of methods, it is necessary to have a basic understanding of resolution and magnification. Magnification is how much an image can be enlarged, and thus how the ob-server will perceive it. In this sense it is really more about personal prefer-ences, than of actual value for qualitative assessments. Resolution on the other hand, describes what the smallest distance between two points that can be distinguished.
The microscopic methods used here, were transmission, scanning, and cryogenic electron microscopy, confocal laser-scanning microscopy, and automated epifluorescence microscopy. The two types of electron microsco-py have, by far, with the highest potential resolution, being in the sub-nanometer range (0.4nm) after optimization of the electron beam. However, these are destructive microscopic technique and require intense specimen preparation, and operate under vacuum, which might change cell morpholo-gy. Additionally, even if specific staining methods such as gold-coating are available, differentiation between microorganisms and other particles can sometimes be challenging and mainly based on subjective interpretation. Environmental screening with such methods, as required for Paper III and IV, was not feasible. However, SEM and TEM, as well as Cryo-EM were needed to resolve structural features of S. acidocaldarius and its type IV-like pili (Paper I). It could be used for this due to the used extraction methods used, which ensured that either only Sulfolobus acidocaldarius or the respec-tive filament-of-interest (knock-out mutants) were inspected. Cryo-EM was used to identify structural feature of the individual filaments. Freeze-cuts were made at nanometer scale and filaments were then reconstituted 3-dimensionally from snapshots of individual cross-sections, obtained from different angles (Craig et al. 2006).

30
Confocal laser-scanning microscopy is an optical type of microscopy. Similar to conventional microscopy it cannot provide spatial resolution be-low the diffraction (Claxton). Using fluorophores, this limitation can to some extent be avoided, as done for the analysis of the S. acidocaldarius biofilms. This, in combination with Z-stacking, allows to not only resolve the individ-ual cells in a biofilm, each in its respective vertical layer, but also partially resolve cellular features such as type IV pili or similar appendages (Paper I). Confocal laser-scanning microscopy has its own set of limitations, such as the limited area and vertical depth that can be characterized. Because of this limitation it was not used for the screening efforts of Paper III and IV, despite superiority in terms of resolution and identical sample treatment. An often encountered, but often too little considered problem is chromatic aber-ration. At higher magnification, where no corrective lenses can be used, it leads to a signal detection shift between different wavelengths. This was a problem often encountered in the epifluorescence microscopy used in Paper III and Paper IV. In particular, due to the automated counting and automat-ed focus alignment, one of the detection channels was often out of focus, which was accounted for with thorough quality control of the obtained mi-crographs. Among the microscopic methods described here, epifluorescence has the lowest resolution. On the other hand, it has the benefit of not being destructive, easy sample preparation, high throughput, and can accordingly be used to scan large areas, a feature needed for assessing cell-abundance profiles across vertical sediment profiles.

31
Results
Paper I Rationale, background and aim: Type IV like pili are of crucial im-portance for microorganisms (Archaea and Bacteria) to physically interact with their environment. Their roles include, virulence, DNA-uptake, cell-cell and cell-surface interactions, biofilm formation and more. Bacterial type IV pili are well studied across a wide range of model organisms, however, our knowledge of similar filaments in Archaea is lacking behind. With the emer-gence of new analytical techniques, and improved cultivation methods, we now know that Archaea are not only abundant in all environments, but also that they have a multitude of unique surface structures to interact with their respective environment. Using computational analysis of class III signal sequences, type IV-like piliation related genes were identified in Archaea, which led to the discovery of the archaellum, the UV-pilus and the aap-pilus. Consecutively, aim in this study was to investigate and resolve structure and function of the aap-pilus, a hitherto unknown type IV pilus in the thermo-acidophilic crenarchaeon Sulfolobus acidocaldarius. Within the greater scope of this thesis, this was done to assess one of the putative mechanisms of one microorganism to adapt to habitat heterogeneity, and transition be-tween planktonic life and surface association. Results and Conclusions: We found that the aap-pilus of S. acidocaldarius, despite its similar genetic makeup, is entirely different from known bacterial type IV pili, or other archaeal filaments, with its 3 strand helical structure, with 138° rotation and a 5.7 Å rise per subunit, and a 110 Å diameter. We demonstrated it to be indispensible for biofilm formation. The core assembly machinery of the aap-pilus consists of five genes, three of which (membrane protein, assembly ATPase, and a putative iron-sulfur-oxido-reductase) are encoded in one operon, with two putative pilin subunits, encoded in differ-ent, loci. Unlike other, more typical type IV pili, the aap-pilus of S. acido-caldarius was found to possess great variability in its helical parameters, likely caused by variations in the hydrophobic N-terminal domains of the pilin, facing inwards into the filament (Figure 7).

32
C D
Fig. 3. Electron microscopic analysis of the aap pili.
scale bars are 100 nm.
layer line, identified as |n| = 3, that is at a spacing of ~
reconstructions, and contained 4613 segments. The near equatorial layer line would therefore be n = -would be n = +
micrograph. This produces the very visible Thon rings in the power spectra.
the structure and with the hydrophobic N-terminal a-helices in the centre. A single aseen in the inner core (d).D. A histogram of mass per unit length measurements of the pili determined by STEM.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd,
Figure 7. Side (a) and top (b) view of the Cryo-EM 3D reconstructed aap-pilus of S. acidocaldarius. With raised density threshold (c), it becomes apparent that the core of the aap-pilus is considerably less dense indicating the hydrophobic N-terminal regions to face inwards, while the hydrophilic (C-terminal), globular do-mains of the pilin subunits face outwards. (d) Hydrophobic, N-terminal α-helices are indicated in the inner core
In-frame deletion of the genes involved into the aap-pilus assembly led to hypermotility (Figure 9), while the archaellum is down regulated upon pilus expression (Figure 8). This cross regulation could be interpreted as a direct adaptation to shifting between motile and sessile modes of life, and adapta-tion to different habitats. Especially deletion of the aapF gene resulted in visible hyperarchaellation. Furthermore it demonstrated that each of the in-volved genes is indispensible for assembly and function of the aap-pilus.
Furthermore, deletion of the aap-pilus, led to flat, carpet like biofilms. This indicates that the aap-pilus also plays an important role in initial attachment, and structural integrity of the S. acidocaldarius biofilm. This carpet-like bio-film phenotype was consistent across all aap-pilus deletion mutants. Addi-

33
tionally, the ∆aapX and ∆aapE showed a strong increase in EPS production (Figure 10). This increase in EPS secretion might partially be responsible for the increased swarming motility in these two strains (Figure 9).
S. solfataricus in the aapF homologue SSO2386 two anti-sense RNAs were described (Wurtzel et al., 2010). If thatis the case for S. acidocaldarius too, the qRT-PCR datamight just be the result of a high expression and accumu-lation of the antisense RNA. Although primers werechosen in such a way that they should bind outside of thepossible transcribed antisense RNAs, the transcription ofthese might not be perfectly terminated leading to a wrongsignal in the qRT-PCR suggesting higher expressionlevels.
Deletion mutant analysis
To dissect which subunits of the aap pilus operon areessential for assembly, single in frame deletion mutantswere obtained of all five aap subunit genes. Using apop-in/pop-out method as described recently by Wagnerand colleagues (2012), in frame deletion mutants weregenerated in a uracil-auxotrophic background strain ofS. acidocaldarius MW001. Additionally, a double mutantlacking both potential pilin genes was constructed. Allmutants were analysed in terms of their growth kinetics;however, no significant difference compared with the wild-type growth behaviour was observed.
Transmission electron microscopic images of all aapdeletion mutants were compared with the wild-type cells,which exhibits archaella, aap pili and threads (Fig. 5B). Innone of the aap deletion strains aap pili could be detected,indicating that all five genes are indispensible for pilus
strain. The deletion of
all aap
DaapX, E and Fmotility, whereas D
operon.
biofilm (Henche et al
B
and aap
Figure 8. TEM micrographs of the S. acidocaldarius wild type (MW001) and the aap-pilus deletion mutants. The archaellum is indicated with black arrows, while the aap pilus is indicated with white ones. All aap-pilus deletion mutants were aap-pilus deficient and only expressed the S. acidocaldarius archaellum. Scale bars at 100nm
Figure 9. Semi-solid-gelrite Swarming motility assay of the aap-pilus deletion mutants, in comparison to the wild type (MW001) and an archaellum deficient, non-motile, mutant (∆flaJ)
In summary the aap-pilus was characterized as a unique archaeal surface structure, similar to bacterial type IV pili, with unique structural features and a yet unreported varia-bility in pilus helicality. It was demonstrated to be involved in regu-latory crosstalk with the archaellum, responsible for swimming motility, and biofilm formation and stability. My personal involvement in this project was construction of the ini-
tial aapF and flaJ deletion mutants, establishing the motility assays, and building the theoretical framework of the biofilm phenotype of S. acidocal-darius as well as the regulatory cross talk with the archaellum.
As previously shown, the DaapF deletion mutant showeda flat biofilm with a high density of cells possibly causedby the overexpression of the archaella (Fig. 7). Thedouble pili deletion strain DaapADaapB and the DaapBwere most similar to this phenotype, whereas the DaapAmutant was comparable to the wild type. Surprisingly, theDaapX and DaapE strains showed a strong increase in
surface attachment and shaping the S. acidocaldariusbiofilm (Henche et al., 2012). During this study, we couldshow that the pilus is very stable against different dena-turing conditions including hot phenol and detergenttreatment. Considering the fact that S. acidocaldariusthrives at temperatures ranging from 70–90°C and lowpH in its natural habitat (Brock et al., 1972), the highrigidity of the filament seemed to be an adaptation to thisenvironment. For bacteria, a high stability of type IV piliwas observed which was linked to their biological func-tions in these organisms. In N. gonorrhoeae and Myxo-coccus xanthus, it was demonstrated that the type IVpilus requires a high stability to withstand mechanicalforces when retracting the pilus to perform twitchingmotility (Merz et al., 2000; Clausen et al., 2009).Recently, thermal and proteolytic stability could also belinked to the GC pilus of N. gonorrhoeae which was sug-gested to stem from aromatic residues present in thepilin PilE (Li et al., 2012). The C-terminal region ofthe two pilins AapA and AapB of the aap pilus containsphenylalanine and tyrosine residues which could thencontribute to a more firm binding of the pilins in the fila-ment. This would suggest that not only hydrophobic inter-actions of the N-terminal a-helices of the pilins areforming the filament, but also the C-terminus of the pilinsmight be involved in forming a rigid structure as well.Additionally, numerous asparagines are present inthe pilins which are possible target residues of theN-glycosylation machinery. The high stability observedfor the aap structure is reflected in the biological role ofthis pilus which is predominantly to promote adhesion toa surface (Henche et al., 2012). Especially, the establish-ment of sufficient binding in an environment character-ized by extreme acidity, heat and consequently highBrownian motion, demands a stable connection of thecell with a surface. In nature, S. acidocaldarius is pre-dominantly found in the crusty edges of geothermalsprings, indicating that this organism prefers the biofilmlifestyle. In contrast to this, other Sulfolobaceae do notform this filament and show a rather simple biofilm devel-
Fig. 6. Swarming motility assay of aap deletion mutants. The cellswere grown to an OD600 of 1 in Brock medium containing 0.1%dextrin and 0.1% tryptone and spotted on a Brock plate containingonly 0.15% gelrite. The tryptone concentration in the plates wasreduced to 0.005% to induce the production of archaella andincubated at 75°C in special metal boxes to prevent evaporation ofmedium. After 7 days, the swarming of the different strains wasanalysed by scanning the plates.
8 A.-L. Henche et al.

34
Fig. 7. Static biofilm formation of all aap deletion mutants in comparison to the wild-type MW001 analysed by confocal laser scanningmicroscopy. Cells were incubated for 3, 6 and 8 days in small Petri dishes at 75°C in special metal boxes to prevent evaporation of medium.The cells were visualized using DAPI (blue channel) and EPS was stained using the two fluorescently labelled lectins ConA and IB4. Thegreen channel represents ConA which binds to glucose and mannose residues, whereas IB4 binds to a-galactosyl residues (yellow channel).Overlay of all three channels are shown. Scale bars are 40 mm.
The adhesive type IV pilus of S. acidocaldarius 9
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
Figure 10. Time series static biofilm assay of the aap-pilus deficient mutants in comparison with the S. acidocaldarius wild type (MW001). Biofilm was visualized using confocal laser-scanning microscopy. Two fluorescently labeled lectins were used to visualize the EPS on the biofilm, green indicating glucose and mannose residues, yellow indicating α-galactosyl residues. Cells are visualized using DAPI (blue channel). Aap-pilus deficient mutants express a flat, carpet like biofilm with raised cell densities. Additionally, ∆aapX and ∆aapE show increased EPS produc-tion.

35
Paper II Rationale, background and aim: Particles are hotspots for biological activ-ity in aquatic systems. They are rich in nutrients in comparison to their aquatic surroundings, and present a stable environment with discrete, well-organized niches. Due to these unique properties, particles are usually rapid-ly colonized by bacteria from the surrounding environment. We used dia-toms as a model system to study these direct contact bacteria-particle inter-actions on single particle level. Diatoms are good model system, because of their tight interactions with bacterial communities, and their strong seasonal-ity. This enables studies on succession patterns in particle colonization. The aim of this study within the framework of this thesis was to assess and iden-tify driving mechanisms in bacterial community composition associated with particles and whether these are controlled intrinsically within the bacterial community or depend primarily on the nature of the colonized particle Results: Bacterial communities on individual particles from a freshwater lake were mainly composed of lineages affiliated with Proteobacteria and Bacteroidetes. This is in agreement with earlier bulk studies of particles col-lected by filtration, and surveys of non-axenic cultures of diatoms, that har-bored similar bacterial communities in opposition to bulk water, which typi-cally comprises much higher abundances of Actinobacteria (Beier & Bertilsson 2011; Eiler, Heinrich & Bertilsson 2012).
No significant species-specificity of the bacterial community composition could be detected, as proposed for most marine systems (Grossart et al. 2005). Instead, two large co-occurrence network modules were observed, which were almost mutually exclusive (Figure 12), similar to findings from pacific deep chlorophyll layers (Baker & Kemp 2014). Correlation of α-diversity with the sequencing data revealed that likely one of the modules, and thus one set of particles, is dominated by bacteria of the genus Flexibac-ter, absent in the study from Baker and Kemp (2014). We assume this group prepares a gateway, or serve as keystone species for primary colonization (Zhang et al. 2006). These pioneer species then determine the assembly of the emerging bacterial community, in a single-particle biofilm tha likely feature srong functional and metabolic interactions (Figure 13). Interesting-ly, it was found that the abundant freshwater LD12 clade as well as several members of the Flavobacteria, were positively correlated with α-diversity, despite being gernerally numerically abundant. This indicates a late entry into the particle associated community. The second, smaller module was almost devoid of Flexibacter and had a much wider array of putative gate-way species, including members of the Planctomycetes, Acidobacteria, Ver-rucomicrobia, and Lentisphaera. Several of these are typically more associ-ated with sediments or soils and it can be argued that these particles might

36
origin from sediment resuspension, explaining the contracting bacterial community composition in the two identified modules.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Ast
erionella
Aula
cose
irace
ae
Baci
llario
phyta
Chlo
rella
ceae
Cym
bella s
ubturg
idula
Desm
idia
ceae
Melo
sira
varia
ns
Navi
cula
les
Thala
ssio
sira
ceae
unclass
ified P
roch
lora
les
Unkn
own
Verrucomicrobia
Proteobacteria
Planctomycetes
OD1
Gemmatimonadetes
Fusobacteria
Firmicutes
Elusicmicrobia
Cyanobacteria
Chloroflexi
Chlamydiae
Bacteroidetes
Actinobacteria
Acidobacteria
AC1
49 2 1 3 1 4 1 1 16 1 4 4
Bacillario
phyceae
Figure 11. Dominant bacterial phyla as observed on the different diatom species and particles, with the sample size for each particle notated on top of each bar. Especial-ly Bacteroidetes and Proteobacteria are dominant on all particles.
í� í� í� í� 0 � �
í�0
��
NMDS1
10'6�
Desmidiaceae
unclassified Rhodophyta
Aulacoseiraceae
Thalassiosiraceae
Chlorellaceae
AulacoseiraceaeDinophysis norvegicaMelosira varians
AsterionellaNaviculalesUnknownCymbella subturgidulaunknown ProchloralesBacillariophyta
0RGXOH��
Module 1
Figure 12. Non metric multidimensional scaling of the bacterial community compo-sition found on the particles. Two clusters are found, correlating to the two co-occurrence network Modules (encircled)

37
A A
A
A
A
A
AA
A
A
A
A
Figure 13. Schematic model of diatom-particle colonization by a pioneer species (brown) using filamentous appendages to attach to the particle surface. EPS is then secreted onto the diatom (green), which in combination with the pili of the pioneer species mediates recruitment of secondary colonizers (blue). Commensal bacteria, or bacteria that are not recruited or mutually excluded are represented in red.
We conclude, that the bacterial community associated with single planktonic freshwater particles do not depend, at least not strongly, on the nature of the particle itself, and instead seems to be controlled by biotic interactions with-in the particle community and keystone-species shaping the colonization.
Paper III Rationale, background and aim: Freshwater sediments are hotspots of biological activity and greatly contribute to the carbon budget of lakes and oceans alike. Sediments feature steep chemical gradients and high heteroge-neity, making them suitable systems for application of high-resolution meth-ods to study microbial microscale interactions. In the present study, mi-croscale variation in microbial oxygen consumption rates was compared to spatial organization of the microbial community to identify linkages between organisms and process. Results and Conclusions: Prior research indicated, that the specific activity of sediment microbiota is overall lower than that of pelagic microorganisms (Haglund et al. 2003). The

38
reason for this discrepancy is the focus of Paper III, where the cell-specific metabolic activity was assessed over depth in top layer sediments. To do this, modeling based on oxygen microprofiles were used to estimate oxygen consumption rates. This was done in parallel to measurements of microbial cell densities with single cell resolution. Additionally I tried to resolve for habitat heterogeneity and the effects of biogeochemical and physical proper-ties of the respective lake sediments used as models. This was done using environmental descriptors, such as carbon quantity, porosity and water con-tent as well as Nitrogen and Phosphorus concentrations.
-0.002
dept
h (c
m)
0
0.2
0.4
0.6
0 200 400 -0.002 0 200 400 -0.002 0 200 400
dept
h (c
m)
0
0.2
0.4
0.6
-0.00015
dept
h (c
m)
0
0.2
0.4
0.6
0 200 400 -0.00015 0 200 400 -0.00015 0 200 400
dept
h (c
m)
0
0.2
0.4
0.6
-0.02
dept
h (c
m)
0
0.2
0.4
0.6
0.8
1
1.2
O2, prod
(nmol cm-3 s-1)
0 200 400
[C] O2 (µmol L-1)
-0.02
cells (per unit area)
0 200 400 -0.02 0 200 400
dept
h (c
m)
0
0.2
0.4
0.6
0.8
1
1.2
A B C
D E F
G H J
INFL
OW
SHORE
BOTT
OM
Figure 14. Oxygen concentration (blue line with standard error represented in light blue), oxygen consumption (grey bars) and cell numbers (black lines) as averages over depth from the three different microcosms.
In opposition to our initial hypothesis, there was no linear correlation be-tween cell densities and cell numbers across either horizontal or vertical scales. Instead, we found the habitat with the least overall density of micro-organisms to be the most active. Interestingly, this was also the habitat with the highest relative water content and porosity indicating similarities to pe-lagic systems (Haglund et al. 2002, 2003). Sediments from the shore-zone, representingr half the area of the lake, featured extremely low oxygen con-

39
sumption rates, despite comparatively high cell numbers. This is likely due to shortage of bioavailable phosphorus caused by high calcium concentra-tions and co-precipitation of phosphate (House 1999). Sediments from the lake inflow had high cell numbers, and intermediate, but more variable oxy-gen consumption rates. Despite the habitat-specific differences, two general observations were consistent among all three sites: (i) there was a sediment top layer with a oxygen consumption peak that was most likely biologically driven, and (ii) a deep oxygen consumption peak appearing even with low cell numbers. We interpret the latter as abiotic oxygen consumption by oxi-dation of metal ions diffusing up from deeper parts of the sediment (Berg, Risgaard-Petersen & Rysgaard 1998; Lalonde et al. 2012).
ï2 ï1 0 1 2
ï2ï1
01
2
CCA1
CC
A2
depth
C
CN
cell
O2cons
10
bottom
in ow
shore
alpha div
Figure 15. Non-metrical multidimensional scaling (NMDS) of the bacterial commu-nities observed in the three sediment microcosms, with environmental descriptors plotted on top of them in a canonical correspondence analysis (CCA).
The bacterial community composition and its relationship to environmental factors were assessed using non-metric multidimensional scaling and canon-ical correspondence analysis (Figure 15). We found that bacterial communi-ty composition was well explained by the C:N-ratio, organic carbon concen-tration as well as depth, cell numbers and α-diversity. We suspect that the

40
shore community to a large degree derives from primary colonizers that are at least partially inactive because of carbon and phosphorus limitation.
Importanly, there was no linear relationship between oxygen consump-tion and cell abundances. Based on this, it was hypothesized that the micro-heterogeneity originating from physical and chemical sediment properties are the main factors controlling metabolic rates within the sediment microbi-ome (Meyerreil 1994; Fenchel & Finlay 2008). This results in great spatial variance in metabolic rates across the lake ecosystem. In assessing micro-habitat heterogeneity using microsensors, microscopy, biogeochemical char-acterization of sediments, and sequencing, it was possible to directly link habitat heterogeneity and microbial activity and define patterns that can be used as a foundation for better predictions of processes at the lake ecosystem scale.
Paper IV Rationale, background and aim: Early paradigms in soil and sediment biogeochemistry assumed that the redox state of an environment is largely dependent on concentrations of electron donors and acceptors, largely un-knowing of the impact of biological activity on the redox state (ZoBell 1946). Subsequent research showed, that biological activity not only shifts the availability and concentrations of these electron donors and acceptors as primary defining parameters for the oxidative reductive potential (ORP) of a sediment, but that also secondary metabolites excreted from microorganism shape the specific redox environment, and that bacterial communities in single organism or mixed cultures actively shape, define, and stabilize the ORP. The interplay of those environmental gradients with microbial activity forms unique niches on microscopic scales. To evaluate the impact of bacte-rial communities on such redox-niche formation was the main goal of this study. Additionally it was evaluated to what degree BCC is explained by, or conversely, explain variation and state of the measured redox conditions in different sediments. Results and Conclusions: In this study we examined the redox environment in three sediment microcosms, representing major habitats in Lake Ånnsjön, using redox microsensors. Our results demonstrate, that the ORP in the sed-iments is stabilized in the top layers of the sediment, when microbial abun-dances are above a minimum cell number threshold, identified to ~1.8 × 10-2 cells per µm2. Cell numbers steeply declined with depth in the sediment, and below this threshold, thus in deeper zones of the sediment. At such depths no significant biotic influence in the ORP could be detected, and the redox envi-ronment is most likely defined by purely geochemical properties of the sed-iment, as suggested by earlier paradigms.
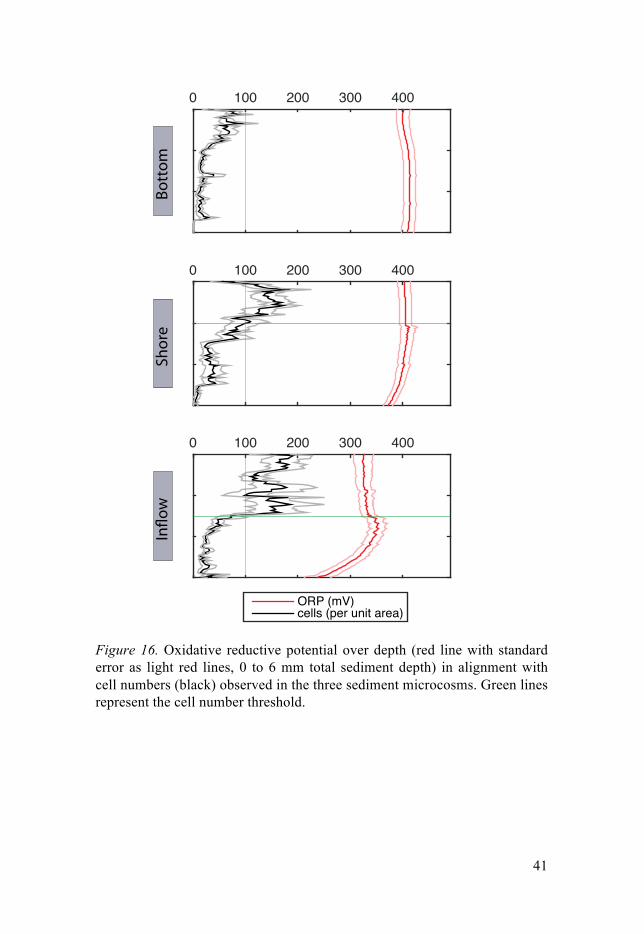
41
Figure 16. Oxidative reductive potential over depth (red line with standard error as light red lines, 0 to 6 mm total sediment depth) in alignment with cell numbers (black) observed in the three sediment microcosms. Green lines represent the cell number threshold.
0 100 200 300 400
0 100 200 300 400
0 100 200 300 400
ORP (mV)
cells (per unit area)
Bott
omSh
ore
In!o
w

42
Figure 17. Non-metrical multidimensional scaling analysis of the bacterial commu-nity composition of samples retrieved from the sediment microcosms. Samples 1, 4, 7, 10, 13, 16 represent the top 2 mm of the respective microcosm (bottom = red, shore = blue, inflow = green), samples 2, 5, 8, 11, 14, and 17 were sampled from mid depth (2 to 4 mm in the sediment), and samples 3, 6, 9, 12, 15, and 18 from the respective deepest sampling point (4 to 6 mm sediment depth). Environmental de-scriptors were plotted on top of the NMDS.
This bacterial influence was most prominent in the inflow sediment micro-cosm (Figure 16), where cell numbers were highest, but simultaneously met with steep ORP gradients in the deeper zones. Additionally we observed the widest array of average ORP in the top zones in the inflow sediments. This in turns is paralleled by the most diverse and variable bacterial community composition, indicating that the ORP profiles could serve as a proxy for different microbial communities (Hunting & Kampfraath 2013). Although the cell number threshold was not met in the bottom sediments, we observed the most stable redox profiles at this site. We believe this is caused by the higher water content and porosity in this sediment, resulting in greater diffu-sive velocities, and a practical equilibrium of ORP-defining compounds. This implies that reduced habitat heterogeneity would be positively correlat-ed with lowered biomass. Shore sediments were an intermediate between the two other microcosms. Patterns for this habitat were similar to those found in the inflow, but less pronounced. This is likely due to lowered metabolic ac-tivity caused by nutrient limitation, as already described in Paper III. Non-metrical multidimensional scaling analysis revealed that at the microcosm
M./% ./. ./% ./F ./L ./0 &/.
M./F
M./%
./.
./%
./F
!"#$&
!"
#$
%
&
%O
F
'
L
E
0
N
&.
&& &%
&O
&F
&'
&L
&E&0
#3JHK
777=>?7254@5)837A5BCD
12345637=>?
I3++7)9:;347254@5)83
1234563783++7)9:;34<
=>?7254@5)837A:@8DG,HH,:
()*+,-
$K,43

43
level, ORP could not explain differences in bacterial community composi-tion (Figure 17). This is in agreement with our initial hypothesis, that the bacterial communities actively shape the redox environment.
Ultimately, driving factors for the bacterial community composition ap-peared to be depth, cell number and average ORP. We found that bacterial community composition in deeper zones of all sediments, were highly simi-lar, with reduced diversity, likely representing a type of deeper sediment community distributed across the entire lake bottom area. Again, this loss of diversity is correlated to reduced heterogeneity, with deeper sediments being less influenced by turbulences and variations in the sediment-water boundary zone (Bryant et al. 2010a b) The top layer bacterial communities were highly dissimilar, with higher α- and β-diversity, indicating that they are highly specific to the individual sediment type.
An in depth analysis of the bacterial community composition and the most prominent members revealed an abundance of Caulobacter in all sedi-ments (Figure 18). Runner-up species in abundance were Streptococcus, Massilia and Bdellovibrio, as well as the LD12 freshwater alphaproteobacte-ria lineage. From this data we infer that predation (Bdellovibrio) could be an important factor in regulating the bacterial community composition. Nitro-gen fixation may also be of importance in sediments as indicated by the presence of the known diazotroph Massilia.
In summary, sediment ORP seems to be shaped by bacterial activity, as long as a minimum cell abundance threshold is met. Below this threshold, geochemical and other abiotic parameters seem to define the redox environ-ment. A sediment community type appeared to be distributed across the deeper sediment zones across the entire lake ecosystem. This community type would potentially be adapted for a life in low-energy habitats, while more energy rich top layers harbor bacterial communities better adapted to the more heterogeneous top layer sediments, and are unique to their respec-tive micro-environment. In conclusion, increased habitat heterogeneity ap-pears to be beneficial for bacterial diversity and biomass. However, there appears to be a fragile balance between too much and too little heterogenei-ty, as indicated by the observed stabilizing effect of sediment microbiota on ORP.

44
0.01
Se
diminib
acteriu
m
Sedim
inibact
erium
Se
diminib
acteriu
m
Clo
acibac
terium
Fla
vobact
erium
Fla
vobact
erium
Fla
vobact
erium
SAR1
1 clad
e LD
12 fre
shwate
r grou
p
Brad
yrhizo
biacea
e
R
hodoba
cterac
eae
Sp
hingom
onas
Ca
ulobac
ter
Azosp
irillum
Ro
seomo
nas
A
cetoba
cterac
eae
Sp
haerot
ilus
Lim
nohabi
tans
Lim
nohabi
tans
Lim
nohabi
tans
Com
amona
daceae
Tep
idimona
s
C
omam
onadac
eae
Ma
ssilia
Po
lynucl
eobact
er
Ha
emoph
ilus
En
hydrob
acter
Pe
rlucidib
aca
Ha
lomona
s phoc
eae
Sh
ewane
lla
Psych
romona
s
Ne
vskia
Un
known
Chtho
niobac
terale
s famil
yVe
rrucom
icrobia
OP
B35 s
oil gro
upVe
rrucom
icrobia
OPB3
5 soil
group
uncu
ltured
Carno
bacter
iaceae
Str
eptoco
ccus
Str
eptoco
ccus
En
teroco
ccus
Sta
phyloc
occus
Acid
imicro
biacea
e
CL500
-29 ma
rine gr
oup
Coryn
ebacte
rium
Propio
nibact
erium
Spor
ichthy
aceae
- hgcI
clade
uncu
ltured
Sporic
hthyac
eae
u
ncultur
ed Sp
orichth
yaceae
Spor
ichthy
aceae
- hgcI
clade
Gem
matim
onadac
eae
Candi
date d
ivision
TM6
Ba
cteriov
orax
Ba
cteriov
orax
Bottom
Shore
Inflow
Legend
: Bottom
0 mm
Bottom
- 2 mm
Bottom
-4 mm
Shore
0 mm
Shore
-2 mm
Shore
-4 mm
Inflow
0 mm
Inflow
-2 mm
Inflow
-4 mm
Deltap
rotoba
cteria
Actino
bacter
ia
Verru
comicro
bia
Firmic
utes
Bacte
roidete
s
Alphap
roteob
acteria
Gamm
aprote
obacte
ria
Betap
roteob
acteria
Figure 18. Phylogenetic tree of the 50 most dominant OTUs found in the three sed-iment microcosms. Bars represent relative abundance for each depth, together with the respective sediment microcosm.

45
Conclusion and Perspectives
Habitat heterogeneity is one of the main parameters shaping and controlling microbial diversity. This has both direct and indirect effects on small-scale processes with impacts on also at the ecosystem level and even beyond the ecosystem boundaries to regional and global scales. My thesis sheds light on the adaptive mechanisms of microorganisms, such as biofilm formation and active control of redox environments, to cope with such habitat heterogenei-ty. Patterns of functional interactions between microorganisms are also stud-ied as they represent additional and often critical means by which individual microbial populations can cope with an environment where resources vary across temporal and spatial scales. The scientific approach is bottom up in moving from an in depth study revealing the significance and multifunction-ality of archaeal type IV pilus system in defining central functional traits of the thermo-acidophilic crenarchaeon Sulfolobus acidocaldarius; i.e. being responsible for surface attachment and biofilm formation as well as motility (Paper I). Bacterial colonization patterns on individual particles in the water column were analyzed in the second study, also considering functional inter-actions within the bacterial community (Paper II). The bacterial community composition and populations were also assessed in the final parts of this work (Paper III & IV), yet in the context of steep chemical gradients form-ing in lake sediments and the interplay between metabolic rates, key envi-ronmental properties (ORP, oxygen availability) and bacterial community structure in sediment microcosms from an oligotrophic high-latitude lake.
The characterization of the S. acidocaldarius type IV pilus (Paper I), shows that, despite being homologous, the archaeal type IV pilus is unique in its structure compared to other bacterial type IV pili studied so far. Its resilience and variability in quaternary structure have not been reported ear-lier in similar systems and underline the importance and impact even minute changes in primary structure can have on quaternary structure and function. Similarly, the cross regulation with the archaellum has shed a new light on adaptation of archaea to differing modes of life and biofilm formation. The observation that archaella are down regulated when pili are expressed during biofilm formation and surface attachment has direct impact for our under-standing of stability and control of archaeal biofilms. Similarly, these obser-vations strengthen our understanding of biofilm formation in general, and, how transitions from one mode of life to another might occur and be regulat-ed. This is shown by upregulation of one pilus, responsible for swimming

46
motility during planktonic mode of life, while another involved in surface attachment and biofilm formation is downregulated, and vice versa. The aap-pilus is missing in the closest relative of S. acidocaldarius, S. solfatari-cus, which is accordingly mainly planktonic. As expected S. acidocaldarius feature superior growth in biofilms, pointing to ecological partitioning of the two lineages. This type of pilus related regulatory mechanisms, loss, and acquisition of genes in response, e.g. pilus mediated natural competence, to different habitats are important processes driving processes in microbial adaptation to habitat heterogeneity (Mattick 2002; Averhoff & Friedrich 2003; Luke et al. 2004). The structural importance of archaeal type IV pili for S. acidocaldarius biofilm formation has the potential to strongly affect our understanding of archaeal biofilms, functioning, and niche adaptation in the future.
The mechanism of early surface adhesion followed by biofilm formation and excretion of EPS, initialized, regulated and mediated by the aap-pilus in S. acidocaldarius is found also in biofilm forming bacteria (e.g. Watnick & Kolter 2000; Donlan 2002). This could for example be the case for the pio-neer lineage Flexibacter, identified in the single particle microbiome analy-sis (Paper II). In species closely related to the here identified Flexibacter, such as Flavobacterium psychrophilum and Flavobacterium columnare (Flexibacter columnare), biofilm formation was for example found to be directly correlated with a loss of gliding motility (Decostere et al. 1999; Cai, De La Fuente & Arias 2013). Type IV like pili have previously been report-ed for these bacteria, (McBride 2004), and similar regulatory mechanisms as observed for S. acidocaldarius are likely to be work here.
Several abundant planktonic bacteria, such as the alphaproteobacterial lineage LD12 and Actinobacteria acI, have not been reported to contain fil-amentous appendages. The underlying reason for this is not known, but it can be suspected, that it is linked to reduced genome-size and small volume as adaptations to evade predation and cope with a nutrient-scarce environ-ment (Andersson, Larsson & Hagström 1986; Corno & Jürgens 2006; Corno et al. 2008). Such typical planktonic freshwater lineages are likely to depend on primary colonizers, or pioneer species, to have the ability to adhere and exploit particle surfaces. The mechanism behind this could be that primary colonizers such as Flexibacter, or functionally equivalent populations pos-sessing filaments and being capable of forming EPS matrices, first colonize a given surface such as the diatoms studied in paper II. These pioneer spe-cies then produce a matrix and conditions the surface and thus enables im-migration of other bacteria, that do not have to possess the functional attrib-utes to produce biofilms de novo (Zhang et al. 2006; Honda 2011). Such a process with a few specialists present in the community to induce primary colonization of particles, followed by sequential integration of the bulk of community members positively associated to the pioneer species, could be described as a whole community response to habitat heterogeneity. Concep-

47
tually, this sets a theoretical framework for whole-community symbiosis, where few members are highly specialized on a key function and the whole community benefits, while the specialists also benefit from other type of interactions when environmental conditions may change. Moreover, such a framework is already observable in tissues of multicellular organisms, yet this concept is not often applied to unicellular microorganism. One of these proposed community-level symbiosis modules was related to microbial community network Module 1, found on the majority of the diatoms studied in paper II. Likely, this module represents the typical bacterial community on aquatic particles. Module 2 on the other hand is likely to originate from sediment resuspension. The clear differences in the particle-associated BCC between those two Modules indicate that the microbiome undergoes a com-munity change upon contact with the sediment. This is suggested by the missing particle specificity of the two distinct particle-associated community subsets (Modules), and the more frequent association of bacteria found in Module 2 with sediment and soils.
Such habitat heterogeneity becomes even more apparent in the sediments, where gradients become steeper, and more often defined by physical frag-mentation Turbulence is also eliminated in sediments, making it more stable in time and space. I used oxygen consumption rates as a proxy for metabolic activity, to address patterns in spatial heterogeneity, and cell-specific activi-ty, and which drivers are defining them (Paper III). Cell specific oxygen consumption rates were highly heterogeneous across all scales, and most likely defined by carbon quantity and quality. In an ideal system, all cells would, be at similar activity levels, and a linear response between cell num-ber and oxygen consumption should be apparent. This as a null hypothesis, falsified in this work, suggesting that heterogeneity in environmental factors has a strong impact on biological systems, not only over large spatial scales, but also across much smaller scales in the micrometer range. Thorough stud-ies of the mechanisms behind this heterogeneity, and a profound understand-ing of their magnitude, and respective responses of the microbiome, would have the potential to greatly improve modeling of central microbial process-es, and help to reduce uncertainties and error rates in such models.
In this last study, (Paper IV) the relationship between cell abundances, bacterial community composition and the ORP of the Lake Ånnsjön sedi-ment microcosms were investigated. This was done to better resolve not only how microhabitat heterogeneity affects bacteria, but also how bacteria affect their habitat and especially habitat heterogeneity. It is known that bacteria stabilize ORP in liquid cultures, but not if this effect also applies to spatially resolved naturally complex environments (Escalante-Minakata et al. 2009; Hunting & Kampfraath 2013). Vertical and partially horizontal variability of one environmental descriptor (ORP) was assessed in its relationship to mi-crobiome descriptors (cell abundance, BCC). Indeed it was observed that bacteria stabilize the ORP, and reduce heterogeneity, if possible (above

48
threshold) as a way of active niche formation. An alternative hypothesis, would be, that a stable, less heterogeneous environment is more beneficial to colonization and to support bacterial life. This really is asking the question: was there a stable redox environment that then was colonized, or was coloni-zation the reason for the environment to stabilize? The more redox-stable bottom microcosm, with its low cell numbers despite stable ORP, suggests that the latter is more likely. Unlike in the other sediments, physical proper-ties and associated increased diffusive velocity and equilibrium of redox-active compounds in this sediment could explain the stable redox state. In the inflow and shore microcosms such processes would be less significant and microbial processes would play a more significant role in counteracting physical habitat heterogeneity. In a nutshell, this last part of my work is re-turning to the Baas Becking hypothesis of everything being everywhere and the environment selecting. In the system I studied, this hypothesis does not seem to hold true, seeing that the community actively transforms the habitat.
A second conceptual framework is apparent throughout my thesis. This second concept is that habitat heterogeneity is generally beneficial for mi-crobial communities. It was repeatedly observed that increased heterogeneity of the habitat leads to greater biomass, or more diversity. Hence, more heter-ogeneous systems harbor greater diversity, with enhanced resilience (Allison & Martiny 2008; Peter et al. 2011) and greater metabolic potential (Hunting et al. 2015). However, too much heterogeneity may be equally detrimental, and this could be the explanation for why microbial driven processes active-ly counteract heterogeneity and stabilize the environment. These processes reach from actively searching and infiltrating a niche using type IV pili and biofilm formation, to recruitment of potential syntrophic partners on parti-cles, to stabilizing the environment by controlling the redox environment. Stated in a simple question that remains open: Is life not always about walk-ing, or better gliding, on the edge between chaos and order, only where ex-actly is this edge?

49
Summary in English
Bacteria and Archaea are the smallest living things on the planet. With sizes often less than a few µm, but an enormous metabolic potential, their impact on whole ecosystem; on the whole planet; is undeniable. Yet, their minute size also means that their immediate environment differs entirely from the way we perceive nature, and something as small as a grain of sand can har-bor an entire microcosmos. Microbial ecology is the science of understand-ing microbial driven processes and how they interact and affect their envi-ronment, as well as their dispersion in time and space.
The Baas-Becking hypothesis states: – Everything is everywhere, but the environment selects – Being one of the most influential theories in microbial ecology, it is also one of the most often challenged. Yet, in order to under-stand its implications, and to challenge and improve on it, we have to first understand the term environment on a scale that is relevant for microorgan-isms. This means understanding the borders of a given environment or habi-tat, its interface with adjacent habitats, nutrient flows, how they are linked and how processes happening in one affect the other. Defining these limits and interfaces is a major task already in macro-ecology, but when going beyond what the human eye can resolve, not knowing the factors determin-ing habitat fragmentation, it becomes an even greater challenge. To further our knowledge of these systems we are often forced to restrict the scales at which we operate and investigate isolated model systems. In my thesis, I try to integrate observations across several scales to unravel individual process-es at single organism level, stepping up to small microbial communities and their interactions with single particles and finally proceed to resolve micro-bial interactions and processes taking place at the micrometer to mm scale, ultimately assessing heterogeneity in different habitats across entire ecosys-tems.
Different model systems, centered on surface-interactions and biofilm formation where chosen to accomplish this, i.e. shed a light on microbial interactions with their environment, and how heterogeneity of the environ-ment affects and is influenced by them. One decisive factor in the mentioned surface interactions, and biofilm formation, is the ability to physically inter-act with a surface. In Bacteria and Archaea, this is often accomplished via filamentous appendages, one of which are type IV pili. Not only are those type IV pili known to be basically the bacterial multifunctional "Swiss army knife", but also they were repeatedly shown to be involved and even indis-

50
pensible, for biofilm formation. A biofilm in this regard is an accumulation of cells, enclosed within a self produced matrix that is beneficial for its members by e.g. protecting them from desiccation, antibiotic compounds, predation, but also allowing for transport of nutrients and secondary metabo-lites, cell-cell communication and many more. Type IV pili in bacteria have previously been shown to be centrally involved in surface attachment as well as biofilm formation and individual interactions between its members and their environment. Yet, little is known about similar filamentous appendages in the archaeal domain. Here, we used the archaeal model organism Sulfolo-bus acidocaldarius to study such a type IV pilus homolog in Archaea. Simi-lar to bacteria, we found that this pilus, the aap-pilus, is indeed indispensible for surface attachment and biofilm formation. However, we also found it to differ strongly from bacterial type IV pili in features of quaternary structure, setting archaeal type IV pili apart from their bacterial counterparts. Further-more, we found it to be tightly linked in a cross-regulatory network with another S. acidocaldarius filament, the archaellum, responsible for swim-ming motility, indicating discrete roles of those two in adaptation to trans-gressing from one mode of life (planktonic) to another (sessile in biofilms).
When some of the cellular traits conferring the ability to form biofilms had been uncovered, I wanted to move from single organisms to probe inter-actions of bacterial populations and communities within the close spatial boundaries of single microscopic particles from a freshwater system. Both different diatoms and similar sized particles, were investigated with regards to the physically associated bacteria to better understand colonization pat-terns and microbe-environment interactions in a more ecological framework. Here it was observed, that in opposition to reports from marine systems, the bacterial communities were not specific to the observed diatom type. Instead we found that pioneer-, or keystone species, are shaping the bacterial com-munity associated to those particles, with especially Flexibacter taking a prominent role in early colonization. Simultaneously, we found that the ob-served bacterial community were split into two distinct subsets, one domi-nated by bacteria typically associated with the open water-column and parti-cles therein, and the other one likely originating from sediments with envi-ronmental conditions quite different from the pelagic habitats.
Organic and inorganic nutrients typically accumulate in sediments and this also leads to high concentration of microorganisms compared to the less nutrient-rich pelagic zone. A large part of the lake metabolism happens in these typically anoxic habitats and the accumulation of both particles and microorganisms and enhanced microbial metabolic processes create a highly heterogeneous environment characterized by steep chemical gradients and high habitat heterogeneity. The next aim was therefore to study how this spatial heterogeneity affected microbial distribution patterns, diversity and activity. In these environments, habitat heterogeneity is, at least in theory, easier to define, due to minimal turbulence and higher temporal stability.

51
Oxygen concentrations and cell numbers were measured and cell specific consumption rates were then calculated and correlated with bacterial com-munity composition. This was done to assess whether bacteria behave uni-formly along gradients in the sediment. No linear correlation could be found, indicating that factors not measured in the scope of this work likely control activity levels and thus the cell-specific metabolic capabilities of the com-munity members. Neither could bacterial diversity sensu richness explain much of the variation in oxygen consumption. We instead found porosity and water content, physically defining habitat heterogeneity, as well as var-iations in carbon content and carbon quality to be the most decisive meas-ured factors explaining these variations in overall metabolic activity of the sediment microbiota, on vertical scales bridging only mm, as well as on whole lake ecosystem scales.
It is known, that bacteria are capable to control redox environments in liquid cultures on temporal scales. With respect to habitat heterogeneity I wanted to test, whether this also applies to spatial scales. Thus, the Ånnsjön sediments were used to link the redox state of the sediment with the bacterial community composition and populations. Investigating their coupling, we found, that the bacterial community in the sediments appears to stabilize and actively shape the redox environment. However, this was only observed above a minimum cell density threshold. This active redox control counter-acts the intrinsic habitat heterogeneity and microorganisms actively form suitable niches to exploit, in a fragile balanced system between too much, and too little heterogeneity.
In conclusion, while our understanding of underlying mechanisms of dif-ferent adaptations to habitat heterogeneity, and different habitats per se, es-pecially in terms of biofilm formation and surface attachment, is generally well developed, we still lack a principal understanding of what defines those exact habitats and the respective boundaries in a natural setting.

52
Svensk sammanfattning
Mikroorganismer, såsom bakterier och arkéer, utgör de minsta levande org-anismerna på vår planet. Deras storlek är ofta mindre än en mikrometer men de har en oerhörd mångfald och förmåga att driva många olika metabola processer, som i sin tur kan påverka hela ekosystem och i förlängningen hela vår planet. Deras minimala storlek innebär också att den miljö som de erfar och utsätts för, till stora delar skiljer från vår uppfattning om deras habitat. I en synbart homogen vattenmassa kan till exempel ett enstaka sandkorn bidra med en stor mängd olika skilda livsmiljöer som i sin tur befolkas av en mångfald av olika mikroorganismer som i varierande utsträckning påverkar varandra; ett mikrokosmos! Inom den vetenskapliga disciplinen mikrobiell ekologi studeras mikroorganismernas mångfald och inbördes interaktioner, men samtidig även deras påverkan på den omgivande miljön genom på-gående metabola processer eller andra cellulära egenskaper. Olika mikroorg-anismers utbredning och spridningsmönster studeras också för att därigenom erhålla kunskap om deras egenskaper, begränsningar och påverkan på om-världen.
Den Holländske botanisten och mikrobiologen Lourens Baas Becking myntade redan 1934 uttrycket att "allting finns överallt men miljön särskil-jer" (fri översättning av "Everything is everywhere, but the environment se-lects"). Detta har blivit en av de mest inflytelserika teorierna inom den eko-logiska mikrobiologin, men har också blivit starkt ifrågasatt. För att till fullo förstå konsekvenserna av Baas Beckings teori och för att testa och vidareut-veckla densamma, måste man först beskriva och beakta termen "miljö" från en mikroorganisms perspektiv; e.g. identifiera den spatiella skala som är relevant och har betydelse för mikrometerstora bakterier och arkéer. I detta ingår även att definiera hur olika miljöer bör avgränsas, beskriva betydelse-fulla gränsskikt, flöden av energi och näringsämnen i naturen (också över mindre avstånd) samt beskriva hur olika samlokaliserade och separerade processer är kopplade och en mängd andra relevanta interaktioner. Att göra detta tillfredsställande är en betydande analytisk utmaning, kanske framför-allt inom den mikrobiella ekologin där det mänskliga ögat, i alla fall i från-varo av hjälpmedel, är till föga hjälp. Att man inom detta relativt unga forsk-ningsfält inte heller har något bra grepp om vilka specifika faktorer som har betydelse för skapandet och avgränsningarna i habitatens utbredning gör utmaningen än större.

53
För att få en förbättrad och fördjupad kunskap om hur dessa system fun-gerar är vi ofta nödgade att tillämpa olika begränsningar i vilka rumsliga och tidsmässiga skalor som utforskas och kanske använda förenklade eller slutna system som experimentella modeller. I min avhandling försöker jag koppla ihop processer, företeelser och egenskaper över både makroskopiska och mikroskopiska rumsliga skalor, för att på detta sätt beskriva och förstå pro-cesser och egenskaper hos enskilda mikrobiella celler och hur de interagerar med den omgivande miljön. Vidare utforskas olika mikroorganismers inbör-des kopplingar på enstaka partiklar som koloniseras av olika typer av bakte-rier och hur skarpa kemiska gradienter i sjösediment formar och formas av de mikroorganismer som lever där och hur dessa mikrobiologiska egenskap-er varierar inom hela akvatiska ekosystem.
För att åstadkomma detta studerades ett antal modellsystem av varierande komplexitet men som alla var kopplade till biofilmer och mikrobiella inter-aktioner med och på olika typer av ytor. Den generella målsättningen var att belysa och förstå mikrobiella interaktioner med den omgivande miljön och specifikt studera betydelsen av spatiell heterogenitet. En central egenskap som möjliggör bildandet av en biofilm är mikroorganismens förmåga att rent fysiskt kolonisera en yta. Hos bakterier och arkéer sker detta i stor utsträck-ning med hjälp av filamentösa utskott på cellens yta, och typ IV pili är ett utbrett exempel på denna typ av struktur. Dessa filamentösa utskott är känd som bakteriens "schweizerkniv" i sin multifunktionalitet och är också av kritisk betydelse vid cellernas bildande av biofilmer. I detta sammanhang är en biofilm definierad som en ansamling av mikrobiella celler som hålls samman av en egenproducerad organisk matris. Denna inbäddning gynnar cellerna på flera olika sätt, bland annat genom att erbjuda skydd mot uttork-ning, antibiotika exponering och predation. Även genom att bidra till kvar-hållande och effektivare upptag av både näringsämnen och sekundära meta-boliter samt genom förbättrade möjligheter till kommunikation mellan celler kan bildandet av en biofilm gynna de organismer som är delaktiga. Man vet att bakteriers typ IV pili är av central betydelse för cellernas inbindning till ytor och efterföljande bildning av biofilmer, men vår kunskap om dessa yt-strukturers funktion och betydelse hos arkéer är betydligt sämre. Min forsk-ning visar att hos en av få modellorganismer för arkéer (Sulfolobus aci-docaldarius) har motsvarigheten till dessa typ IV pili (aap-pilus), precis som hos bakterier, en nyckelroll för cellernas förmåga att binda in till ytor och bilda biofilmer. Detta trots att strukturen hos dessa utskott hos arkéerna var mycket avvikande jämfört med utskotten hos bakterier. Vidare så fanns det en stark koppling i regleringen av uttrycket av dessa aap-pili och ett annat viktig filamentöst utskott: flagellen som ger cellen förmåga att förflytta sig i en vätska. Denna regulatoriska koppling har sannolikt en viktig roll i cellens övergång från ett frisimmande till en ytassocierad livsstil.
När en del av de cellulära egenskaper som påverkar och styr förmågan att binda in till ytor klarlagts för denna modellorganisms ville jag gå vidare och

54
studera mikroorganismers kolonisering av ytor under mer realistiska förhål-landen där olika typer av mikroorganismer konkurrerar, samverkar och på andra sätt påverkar varandra i koloniseringen av ytor och utnyttjandet av substrat. Genom att analysera artsammansättningen hos de bakterier som återfanns på enstaka levande och döda partiklar/ytor i sjövatten kunde kopp-lingar mellan olika bakteriegrupper och arter beskrivas och utvärderas. En rad olika partiklar, inklusive olika arter av kiselalger och andra eukaryota växtplankton, inkluderades i studien för att svara på frågan om vad som av-gör artsammansättningen hos de bakteriesamhällen som växer fram i nära fysisk kontakt med varandra på mikroskopiskt små partiklar; är partiklarnas beskaffenhet viktigast eller har funktionella kopplingar inom bakteriesam-hället större betydelse? Till skillnad från tidigare studier i marina vatten kunde vi inte se någon större skillnad i bakteriernas artsammansättning på olika typer av kiselalger. Istället pekade resultaten på att vissa nyckelbakte-rier som sannolikt står för den initiala koloniseringen av ytorna (så kallade pionjär-arter) styr artsammansättningen hos de bakteriesamhällen som senare koloniserar partiklarna. Ett slående exempel på en sådan pionjär-art i det studerade sjövattnet var Flexibacter som redan tidigare beskrivits som en biofilmbildande bakterie. Genom att studera artsammansättningen på partik-larnas bakterieflora var det också möjligt att identifiera partiklar som sanno-likt härrör från omgivande landområden eller underliggande sediment där miljöförhållanden skiljer sig höggradigt från den öppna vattenmassan.
I sediment ansamlas organiska och oorganiska näringsämnen och detta får också till följd att halten mikroorganismer blir hög jämfört med den normalt mycket mindre näringsrika ovanliggande vattenmassan. Mycket av sjöns ämnesomsättning sker i dessa syrefattiga habitat och ansamlingen av både partiklar och mikroorganismer och de sistnämndas metabola aktivitet leder till en höggradigt heterogen miljö som kännetecknas av skarpa kemiska gra-dienter och betydande habitat-heterogenitet. Mot bakgrund av detta blev det en naturlig följd att studera hur denna heterogenitet påverkade bakteriers utbredning, mångfald och aktivitet. Till skillnad från partiklar som påverkas av turbulens och andra snabba förändringar i omgivande miljöförhållanden, när den till exempel färdas från ytvatten till syrefritt bottenvatten eller resus-penderas från mörka sediment till kraftigt solbelysta ytvatten, så är förhål-landen i sedimenten, trots betydande spatiell variation, mycket mer stabila över tid. Det är väl känt att syretillgång och syreförbrukningen som sker vid bakteriers cellandning har starkt styrande inverkan på en rad biogeokemiska processer. I en första studie studerades därför i vilken utsträckning bakterier-nas antal och biomassa i olika sediment och från olika djup kunde förklara observerad syreförbrukning. Inget linjärt samband kunde påvisas, utan den specifika cellandningen (mängd syrgas förbrukad per cell och tidsenhet) verkade istället vara väldigt olika i de olika typerna av sediment som stude-rades och vidare analyser indikerade att andra uppmätta faktorer, såsom se-dimentets fysiska beskaffenhet (porositet, vattenhalt) och halten och kvali-

55
teten på det organiska materialet, verkade ha en större påverkan på syrgas-förbrukningen. Inte heller bakteriernas artrikedom kunde förklara variation-en i syrgasförbrukning i någon större utsträckning.
Dessa sedimentsystem utnyttjades även för att studera kopplingen mellan sedimentens redoxförhållanden och bakteriesamhällets artasammansättning. Tidigare studier av renkulturer av olika bakterier har visat att bakterier besit-ter förmågan att skapa karaktäristiska redoxförhållanden genom sin egen metabola aktivitet. Erhållna djupprofiler av redoxpotential och bakteriesam-hällets sammansättning pekar på att detta även sker i naturligt komplexa mikrobiella habitat såsom sjösediment och därigenom bidrar till minskad lokal spatiell habitat-heterogenitet. Dock krävs en lägsta halt bakterier för att denna effekt ska bli märkbar.
Sammanfattningsvis så har vår kunskap om hur mikroorganismer anpas-sar sig till en variabel miljö ökat och de underliggande mekanismerna som leder fram till dessa anpassningar, framför allt beträffande bildandet av bio-filmer, har också kunnat påvisas och studeras. En utmaning som vi dock fortfarande har framför oss är att ta fram tydliga principer och modeller för hur mikrobiella habitat bör avgränsas och var gräns-skikten återfinns i vår heterogena naturmiljö.

56
Acknowledgments
I do not even know where to start with this, but I guess, it is always a good idea to start with a quote and there are two, both from the same guy, I could think of to describe the last years.
“If you're going to be crazy, you have to get paid for it or else you're going to be locked up.”
“Like most others, I was a seeker, a mover, a malcontent, and at times a stu-pid hell-raiser. I was never idle long enough to do much thinking, but I felt somehow that some of us were making real progress, that we had taken an honest road, and that the best of us would inevitably make it over the top. At the same time, I shared a dark suspicion that the life we were leading was a lost cause, that we were all actors, kidding ourselves along on a senseless od-yssey. It was the tension between these two poles - a restless idealism on one hand and a sense of impending doom on the other - that kept me going.”
Hunter S. Thompson
Now, on the first one, sometimes I see myself in it, and I want, in the first
place, and most of all, thank Annelie and Stefan, my two supervisors, and please excuse the expression, for being batsh**t crazy enough to really real-ly pay me, I still can’t believe they did that. But thank you so so much for allowing me to develop, for pushing me in this melting pot of crazy ideas incredibly intelligent, talented, creative, hardworking people, letting me simmer in there for a few years in the hope I would become a part of that soup. And even more than just that, to develop, develop my ideas, develop my attitude, develop my talents, learn to overcome adversaries, develop skills I never thought I had, and overcome others I wish I never had to begin with and in the end build a research philosophy. Annelie, I remember great talks, sushi on the lawn and feet dangling in the pond talking science and everything else that is important. Thank you for that. Stefan, the warm wel-come you gave me here in the cold of Sweden when I got here in January a few years ago will always be appreciated. So are the BBQs and the great food and get-togethers at your place. I much appreciated your abundance of great ideas and brainstorming in how to tackle scientific questions in the beginning of a project, and the clearness of thought and guidance in the end. And I also wanted to thank you for pushing me in the water when it came to

57
teaching and those hours of teaching, and the enjoyable chaos that came with it from teaching Eco-pop at Erken in my first year, to Infection biology and Microbial ecology in my last. Thank you two so much for making all that happen.
Lars, your crystal clear lines of thought, and guidance and scientific input, as well as the words of motivation and your last minute comments on my kappa were of great help and I wanted to thank you for them, you are what keeps this department together and make it the inspiring workplace it is. Alex, Sebastian, and Eva, your help in so many of the things, and the discus-sion we had over my research helped me a great deal in coming to where I stand now. Richard and Peter, teaching the Fish course with you guys, al-ways was one of the highlights of my year, and I enjoyed it endlessly, de-spite the odor, that just happens to come along with being locked into a room for a few days with some slowly thawing decade old fish. Anna Brunberg, you are the one person that makes all the teaching happen, and your positive attitude and friendliness is one of the cornerstones of the limno-department. Silke and Gesa, just having you around for all the fikas, and being able to ask questions out of and within my field, and your valuable comments and questions during presentations and discussions.
At this point I also want to thank the Malmens foundation, the Ohlsson Borg foundation and the Helmholtz Center for Environmental Research in Leipzig and the Helmholtz foundation for the financial support that made this work possible.
Janne and Christoffer, no Acknowledgement would be complete ever, be it in a thesis, or in one of the papers that are published out of the limno de-partment, without your names on it, and your guys’ humbleness, helpfulness and dedication astonishes me every day. Janne, those hours in organizing the garage, and getting things to work and improve in there, were not only fun, but a great escape for me to get some time to think when my office walls were closing in on me. Ute, without you I could not have done the work I did in Leipzig, and I thank you so much for it. Moritz, thanks for all the help with all those codes and sequences, guess there’s hope for me dealing with bioinformatics after all.
Then of course theres all the post-docs, collaborators and former PhDs I had the pleasure to work with. Florian, between Jazz evenings in Leipzig and Co-occurrence network analysis I had a lot of fun working with you and Paper II wouldn’t have happened without you. Omneya, the same is true for you, thanks for that. Martha and Claudia you guys are great fun and between nerdtalk on movies and comics, keeping an eye on Simcha and introducing students into the fun of working with microscopes I enjoyed your friendship and friendliness. Sari, Sarahi Erik, Simone, Kristin, Oona, Birgit, Heli and Johanna, you guys are an integral part and the think tank of this department, that I had so much fun being part of. Christian, you're an inspiration and one of the most driven and intelligent people I know. Pia, I thank you for giving

58
me a home in those first days in Sweden and for your genuine way of ex-pressing yourself, my beginnings here wouldn’t have been the same without you. Hannes and Philipp, you guys were a kind of role model when I got here, and I hope I somehow lived up to the task following your footprints every now and then. Both your guys’ laughter has been dearly missed on the hallway, and during fishing in those last years and I hope there will be a time we get together again at Ekoln, fishing in November, freezing like crazy, being all miserable, and disappointed with Hannes being the only one catch-ing – great memories, or listening to disco inferno as a party starter. Phillip, I have to admit I stole that technique, and repeat it every time somebody gives me responsibility over a playlist these days.
Andrea, Nuria, Pili and Merce, the Spanish and Catalan connection – you guys are awesome and I miss, and will miss you all. The energy and friend-liness, and the temperament you guys radiate is infectious and oh so much fun, I really enjoyed every minute. Andrea and Mercé I think we still have an open game of Monopoly. And Mercé, you are such a kind person, thanks for welcoming me to the department and being a friend. Maria Morales, thanks for all the moral support and for being a friend. Ina, those angry emails of yours – the pleasure of reading them, and the inspiration to write my own – priceless. Charles and Jérômé – one of my first off-time memories of Sweden and the time here is that little Easter trip to Öland and Kalmar with you guys. The disoriented roaming around in medieval castles, even more disoriented driving around on Öland in my tiny Skoda, roadside BBQ and that little Easter-bunny-incident are all memories I deeply cherish. François, sharing a room with you in Granada was a pleasure, despite the “little” downsides of this arrangement. You are a great guy, a pleasant com-pany and a person I would always work and ride with.
Now all the people in Leipzig, the Studentriders there, here in Uppsala, and all around the world, you guys were and are my escape route when the world was getting to small and I needed to break free for a bit. And the same is true for all of you guys in “the event group”. Salar, Rob and Axel, fishing in suits and midnight golfing, only in Sweden I guess. Giulia, thanks for all the motivations and being there and taking care that I don't go crazy.
Göran, Marghareta och Portos, Tack ska ni ha! De några timmar varje veckan jag kunde fly från jobbet till den liten paradiset ni byggt är precis vad förhindrade att jag gick galen inom senaste året. Alltså, ska jag tacka ni för att göra att jag pratar svenska, så tack igen. Och Portos, du är finaste Hästen i hela världen! Johnny, det gäller för dig också, inte saken med finaste Häs-ten, men att gör mig prata svenska. Också, att visa mig runt i Sverige, visa mig bästa ställen att plocka svamp och diskuterar hela natten lång, Tack för det.
My dear fellow PhD students, companions on my journey in the last few years, whether our ways just briefly crossed, or if I basically carried you around on my back (I’m talking to you Hannah), Rock on guys and don’t

59
give up, if I can do it you can do it. Please forgive me if I do not thank every one of you individually for being such good company and friends, but some of you I want to address a little more specific. Blaize and Monica, you guys…what shall I say? Midsommers were great, fikas were great, you guys are great. Your joyful natures always picked me up; keep on being who you are and never let anyone tell you anything else. Blaize, I’m still sorry that I chopped your machine into pieces, but you have to admit getting there and back again was quite a ride. Monica, there are so many memories, Denmark, mushroom hunting, boat rides to name just a few, when I started you were still doing your master thesis and look at you now, you are almost done and I believe in you. And of course, thank you for your help at Ånnsjön, I couldn’t have done it otherwise. Maria Marklund and Jovana, you too will do it, I don't think there’s a hinder big enough to keep you two from going where you want to, and you impress me. Valerie and Inga, my comrades in crime and the two people that can relate best to some of the aspects of my last years. All that moving around, and sitting in between the chairs, it was al-ways good to have somebody around that felt the same, somebody to talk to about those things, thank you.
Dolly, you are an inspiration, your guidance, help and calm mind at Ånnsjön, and talking about science and life and everything was always, and will always be invaluable, thank you so much, and never forget to give Eddy a hug from me on our Birthday. My two roomies at work. Lorena and Fred. Thanks for letting me have the office all for myself all that time - what an immense luxury that was. But seriously, I love you guys and our little en-clave of chaos is in my eyes probably the coziest place in all of EBC.
Maddy, there’s really just one word I can say, mornin’ – it was often enough the only reason to get up and tackle this whole thesis thing. Waschtl, the two of us playing havoc during all those lab courses, dude I couldn’t have done half of them without you being my benchbuddy. And later on, discussing science philosophy with you on the phone and all that, you know, you know. Franzi, you made Leipzig and the UFZ a home, thanks for lend-ing me an ear when I needed one and helping me break the circle, when that was in order, I wish you all the best. Lenalein. Was hätt ich nur gemacht ohne deine Couch? Aber im Ernst, ohne dich wär vieles nicht drin gewesen, und selbst wenn, dann nur halb so lustig, und Prinz Marten hüten, Schränke fehlbeschriften und Berghütten beinahe anzünden sind jetzt nur ein paar der Geschichten die mir da einfallen, Viel Glück mit deinem Doc in der Schweiz, ich drück die Daumen das du mindestens soviel Spaß dabei hast wie ich in den letzten Jahren hatte.
Anne, Hannah, Roger, Aodhán and Dermot. You guys are family and I have no clue how to begin with this. Hannah, I have no words, just a raised eyebrow and an astonished, slightly doubting look on my face. We started together, we almost finished together on the minute actually, that were all in all almost five years, and despite driving me crazy at times I wouldn’t want

60
to miss one second, and I think I will never be able to eat pasta with pesto without a smile on my face and a good memory in my mind. Just do me one favour, if you come visiting me, wherever I may be, give me a ten minutes heads up, so I can go and stock up on potato chips and chocolate.
Roger, working with you, out at Tämnaren on your projects, well, that was hardly work, or on our combined project taught me so much. You are a great friend and were always there when I needed to head downtown for a beer (it just kind of never stayed at one, except that one time). Thanks for having me at your wedding with your beautiful wife. All the best for your combined future, and I hope we will somehow manage to work together in the future (it could even involve some Moose).
Aodhán and Dermot. Thanks to you two I had a home here in Uppsala, and from you two I learned that a good cup of Barry’s tea cures everything, not to mention me now appreciating tea in the first place, and that slight accent I apparently incorporated. There are so many great hours I remember, and so many more I don’t. You guys are like brothers to me.
Anne, many people inspired me on the way here, many impressed me, but none touched my life as much as you did. There will always be a movie ready and a place for you in my Tiki bar, and if I wont have one of those I will build one for you. I thank you for proof reading my thesis, but most of all for putting my head in the right place over and over and over again. You are one of my best and dearest friends, and most of all you are family.
Das Beste kommt ja immer zum Schluss: meine Familie. Tina und Skadi, ich bin einfach nur überglücklich, dass es euch gibt. Mom und Dad, Eure nie endende Unterstützung ist die Säule auf der ich meine Welt aufgebaut habe und ich kann euch einfach gar nicht genug dafür danken. Ohne Euch Beide wäre das alles hier nicht möglich gewesen, von meinem ersten Mikroskop, über Nachhilfestunden in Mathe, Ihr habt so viel für mich getan. Ich sehe Euch heute noch auf dem Bahnsteig stehen und mir zuwinken als ich damals nach Marburg gezogen bin um Biologie zu studieren, und seitdem gab es keinen Moment an dem Ihr an mir gezweifelt habt und das hat mir unendlich viel Kraft gegeben. Danke, ich hab Euch so lieb. Selbst wenn ich ständig in der Welt unterwegs bin und meine eigenen Wege gehe, Ihr seid immer bei mir und meine große Konstante. Zum guten Schluss, Uli und Holgi, meine beiden Allerallerallerlieblingsgeschwisterkinder, ich spreche nun mittlerwei-le gebrochen drei Sprachen, aber die Worte um auszudrücken wie sehr ich Euch liebe hab ich noch in keiner davon gefunden, und ich glaube auch nicht, dass es die gibt. Danke, dass Ihr da seid!

61
References
Aas F.E., Løvold C. & Koomey M. (2002) An inhibitor of DNA binding and uptake events dictates the proficiency of genetic transformation in Neisseria gonorrhoeae: Mechanism of action and links to Type IV pilus expression. Molecular Microbiology 46, 1441–1450.
Aizawa S.I. (2001) Bacterial flagella and type III secretion systems. FEMS microbiology letters 202, 157–164.
Albers S., Szabó Z. & Arnold J.M. (2003) Archaeal Homolog of Bacterial Type IV Prepilin Signal Peptidases with Broad Substrate Specificity Archaeal Homolog of Bacterial Type IV Prepilin Signal Peptidases with Broad Substrate Specificity. 185, 3918–3925.
Albers S.-V. & Jarrell K.F. (2015) The archaellum: how archaea swim. Frontiers in Microbiology 6, 1–12.
Albers S.-V. & Meyer B.H. (2011) The archaeal cell envelope. Nature reviews. Microbiology 9, 414–426.
Albers S.V. & Driessen a. J.M. (2008) Conditions for gene disruption by homologous recombination of exogenous DNA into the Sulfolobus solfataricus genome. Archaea 2, 145–149.
Allison S.D. & Martiny J.B.H. (2008) Resistance, resilience, and redundancy in microbial communities. 105, 11512–11519.
Amin S. a., Hmelo L.R., van Tol H.M., Durham B.P., Carlson L.T., Heal K.R., et al. (2015) Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature.
Amin S. a., Parker M.S. & Armbrust E. V. (2012) Interactions between Diatoms and Bacteria. Microbiology and Molecular Biology Reviews 76, 667–684.
Andersson a, Larsson U. & Hagström Å. (1986) Size-selective grazing by a microflagellate on pelagic bacteria. Marine Ecology Progress Series 33, 51–57.
Anyan M.E., Amiri A., Harvey C.W., Tierra G., Morales-Soto N., Driscoll C.M., et al. (2014) Type IV pili interactions promote intercellular association and moderate swarming of Pseudomonas aeruginosa. Proceedings of the National Academy of Sciences 111, 18013–18018.
Armbrust E.V. (2009) The life of diatoms in the world’s oceans. Nature 459, 185–192.
Armbrust E.V., Berges J. a, Bowler C., Green B.R., Martinez D., Putnam N.H., et al. (2004) The genome of the diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science (New York, N.Y.) 306, 79–86.
Averhoff B. (2004) DNA Transport and Natural Transformation in Mesophilic and Thermophilic Bacteria. Journal of Bioenergetics and Biomembranes 36, 25–33.
Averhoff B. & Friedrich A. (2003) Type IV pili-related natural transformation systems: DNA transport in mesophilic and thermophilic bacteria. Archives of Microbiology 180, 385–393.
Baas Becking L.G.M. (1934) Geobiologie of inleiding tot de milieukunde. W.P. Van Stockum & Zoon, Den Haag.

62
Baker L. & Kemp P. (2014) Exploring bacteria–diatom associations using single-cell whole genome amplification. Aquatic Microbial Ecology 72, 73–88.
Battin T.O.M.J., Wille A., Sattler B. & Battin T.O.M.J. (2001) Phylogenetic and Functional Heterogeneity of Sediment Biofilms along Environmental Gradients in a Glacial Stream Phylogenetic and Functional Heterogeneity of Sediment Biofilms along Environmental Gradients in a Glacial Stream. Applied and Environmental Microbiology 67, 799–807.
Beier S. & Bertilsson S. (2011) Uncoupling of chitinase activity and uptake of hydrolysis products in freshwater bacterioplankton. Limnology and Oceanography 56, 1179–1188.
Bell W. & Mitchell R. (1972) Chemotactic and Growth Responses of Marine Bacteria to Algal Extracellular Products. Biol. Bull. 143, 265–277.
Benton T.G., Solan M., Travis J.M.J. & Sait S.M. (2007) Microcosm experiments can inform global ecological problems. 22.
Berg P., Risgaard-Petersen N. & Rysgaard S. (1998) Interpretation of measured concentration profiles in sediment pore water. Limnology and Oceanography 43, 1500–1510.
Berkner S., Grogan D., Albers S.V. & Lipps G. (2007) Small multicopy, non-integrative shuttle vectors based on the plasmid pRN1 for Sulfolobus acidocaldarius and Sulfolobus solfataricus, model organisms of the (cren-)archaea. Nucleic Acids Research 35, 1–12.
Berkner S. & Lipps G. (2008) Genetic tools for Sulfolobus spp.: Vectors and first applications. Archives of Microbiology 190, 217–230.
Besemer K. (2015) Biodiversity, community structure and function of biofilms in stream ecosystems. Research in Microbiology, 1–8.
Bienhold C., Boetius A. & Ramette A. (2012) The energy–diversity relationship of complex bacterial communities in Arctic deep-sea sediments. The ISME Journal 6, 724–732.
Blocker A., Komoriya K. & Aizawa S.-I. (2003) Type III secretion systems and bacterial flagella: insights into their function from structural similarities. Proceedings of the National Academy of Sciences of the United States of America 100, 3027–3030.
Bowler C., Allen A.E., Badger J.H., Grimwood J., Jabbari K., Kuo A., et al. (2008) The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456, 239–244.
Brierley C.L. & Brierley J.A. (1973) A chemoautotrophic and thermophilic microorganism isolated from an acid hot spring. Canadian journal of microbiology 19, 183–188.
Bruckner C.G., Bahulikar R., Rahalkar M., Schink B. & Kroth P.G. (2008) Bacteria Associated with Benthic Diatoms from Lake Constance : Phylogeny and Influences on Diatom Growth and Secretion of Extracellular Polymeric Substances. Applied and Environmental Microbiology 74, 7740–7749.
Bryant L.D., Lorrai C., McGinnis D.F., Brand A., Wüest A. & Little J.C. (2010a) Variable sediment oxygen uptake in response to dynamic forcing. Limnology and Oceanography 55, 950–964.
Bryant L.D., McGinnis D.F., Lorrai C., Brand A., Little J.C. & Wüest A. (2010b) Evaluating oxygen fluxes using microprofiles from both sides of the sediment–water interface. Limnology and Oceanography: Methods 8, 610–627.
Burrows L.L. (2012) Twitching Motility: Type IV Pili in Action. Annual Review of Microbiology 66, 493–520.

63
Cai H., Jiang H., Krumholz L.R. & Yang Z. (2014) Bacterial Community Composition of Size-Fractioned Aggregates within the Phycosphere of Cyanobacterial Blooms in a Eutrophic Freshwater Lake. PLoS ONE 9, e102879.
Cai W., De La Fuente L. & Arias C.R. (2013) Biofilm formation by the fish pathogen flavobacterium columnare: Development and parameters affecting surface attachment. Applied and Environmental Microbiology 79, 5633–5642.
Carlucci A.F. & Silbernagel S.B. (1969) Effect of vitamin concentrations on growth and development of vitamin-requiring Algae. Journal of Phycology 5, 64–67.
Corno G., Caravati E., Callieri C. & Bertoni R. (2008) Effects of predation pressure on bacterial abundance, diversity, and size-structure distribution in an oligotrophic system. Journal of Limnology 67, 107–119.
Corno G. & Jürgens K. (2006) Direct and indirect effects of protist predation on population size structure of a bacterial strain with high phenotypic plasticity. Applied and Environmental Microbiology 72, 78–86.
Costerton J.W. (1995) Overview of microbial biofilms. Journal of Industrial Microbiology 15, 137–140.
Costerton J.W., Geesey G.G. & Cheng K.J. (1978) How bacteria stick. Scientific American 238, 86–95.
Craig L., Pique M.E. & Tainer J. a (2004) Type IV pilus structure and bacterial pathogenicity. Nature reviews. Microbiology 2, 363–378.
Craig L., Volkmann N., Arvai A.S., Pique M.E., Yeager M., Egelman E.H., et al. (2006) Type IV Pilus Structure by Cryo-Electron Microscopy and Crystallography: Implications for Pilus Assembly and Functions. Molecular Cell 23, 651–662.
Decostere a., Haesebrouck F., Van Driessche E., Charlier G. & Ducatelle R. (1999) Characterization of the adhesion of Flavobacterium columnare (Flexibacter columnaris) to gill tissue. Journal of Fish Diseases 22, 465–474.
Donlan R.M. (2002) Biofilms: Microbial life on surfaces. Emerging Infectious Diseases 8, 881–890.
Droop M.R. (2007) Vitamins, phytoplankton and bacteria: Symbiosis or scavenging? Journal of Plankton Research 29, 107–113.
Dworkin M., Falkow S., Rosenberg E., Schleifer K.-H. & Stackebrandt E. (2006) The Prokaryotes: A Handbook on the Biology of Bacteria: Archaea. Bacteria: Firmicutes, Actinomycetes.
Eiler A., Heinrich F. & Bertilsson S. (2012) Coherent dynamics and association networks among lake bacterioplankton taxa. The ISME Journal 6, 330–342.
Escalante-Minakata P., Ibarra-Junquera V., Rosu H.C., De León-Rodríguez a. & González-García R. (2009) Online monitoring of Mezcal fermentation based on redox potential measurements. Bioprocess and Biosystems Engineering 32, 47–52.
Falkowski P.G. (1998) Biogeochemical Controls and Feedbacks on Ocean Primary Production. Science 281, 200–206.
Fenchel T. & Finlay B. (2008) Oxygen and the spatial structure of microbial communities. Biological Reviews 83, 553–569.
Gärdes A. (2010) A bilateral model system for the molecular investigation of diatom-bacteria interactions by.
Gärdes A., Iversen M.H., Grossart H.-P., Passow U. & Ullrich M.S. (2011) Diatom-associated bacteria are required for aggregation of Thalassiosira weissflogii. The ISME journal 5, 436–445.
Granéli E. & Turner J.T. (2006) Ecology of Harmful Algae. Springer Berlin Heidelberg.

64
Green J. & Bohannan B.J.M. (2006) Spatial scaling of microbial biodiversity. Trends in Ecology and Evolution 21, 501–507.
Grogan D.W. & Gunsalus R.P. (1993) Sulfolobus acidocaldarius synthesizes UMP via a standard de novo pathway: Results of a biochemical-genetic study. Journal of Bacteriology 175, 1500–1507.
Grossart H.P., Levold F., Allgaier M., Simon M. & Brinkhoff T. (2005) Marine diatom species harbour distinct bacterial communities. Environmental Microbiology 7, 860–873.
Guillard R.R.L. & Kilham P. (1977) The ecology of marine planktonic diatoms. In: The biology of diatoms. pp. 372–469. Blackwell Oxford.
Haglund A.L., Lantz P., Törnblom E. & Tranvik L. (2003) Depth distribution of active bacteria and bacterial activity in lake sediment. FEMS Microbiology Ecology 46, 31–38.
Haglund A.L., Törnblom E., Boström B. & Tranvik L. (2002) Large differences in the fraction of active bacteria in plankton, sediments, and biofilm. Microbial Ecology 43, 232–241.
Hall-Stoodley L., Costerton J.W. & Stoodley P. (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nature reviews. Microbiology 2, 95–108.
Hammer Ø., Harper D. & Ryan P. (2009) PAST-PAlaeontological STatistics, ver. 1.89. University of Oslo, Oslo, 1–31.
Henche A.-L., Koerdt A., Ghosh A. & Albers S.-V. (2012) Influence of cell surface structures on crenarchaeal biofilm formation using a thermostable green fluorescent protein. Environmental microbiology 14, 779–93.
Honda K. (2011) Porphyromonas gingivalis sinks teeth into the Oral microbiota and periodontal disease. Cell Host and Microbe 10, 423–425.
House W. a. (1999) The Physico-Chemical Conditions for the Precipitation of Phosphate with Calcium. Environmental Technology 20, 727–733.
Hughes J.B., Hellmann J.J., Ricketts T.H. & Bohannan B.J.M. (2001) Counting the Uncountable : Statistical Approaches to Estimating Microbial Diversity MINIREVIEW Counting the Uncountable : Statistical Approaches to Estimating Microbial Diversity. Applied and environmental microbiology 67, 4399–4406.
Hunting E.R. & Kampfraath a. a. (2013) Contribution of bacteria to redox potential (Eh) measurements in sediments. International Journal of Environmental Science and Technology 10, 55–62.
Hunting E.R., Vijver M.G., van der Geest H.G., Mulder C., Kraak M.H.S., Breure A.M., et al. (2015) Resource niche overlap promotes stability of bacterial community metabolism in experimental microcosms. Frontiers in Microbiology 6, 1–7.
Jacob H.E. (1970) Redox Potential. Methods in microbiology, 91–123. James S.G., Holmström C. & Kjelleberg S. (1996) Purification and characterization
of a novel antibacterial protein from the marine bacterium D2. Applied and environmental microbiology 62, 2783–2788.
Jessup C.M., Kassen R., Forde S.E., Kerr B., Buckling A., Rainey P.B., et al. (2004) Big questions , small worlds : microbial model systems in ecology. 19.
Johnson T.L., Abendroth J., Hol W.G.J. & Sandkvist M. (2006) Type II secretion: From structure to function. FEMS Microbiology Letters 255, 175–186.
Jonge L.W. De & Moldrup P. (2009) Soil Infrastructure , Interfaces & Translocation Processes in Inner Space (“ Soil-it-is ”): towards a road map for the constraints and crossroads of soil architecture and biophysical processes. 1485–1502.

65
Jonuscheit M., Martusewitsch E., Stedman K.M. & Schleper C. (2003) A reporter gene system for the hyperthermophilic archaeon Sulfolobus solfataricus based on a selectable and integrative shuttle vector. Molecular Microbiology 48, 1241–1252.
Kaczmarska I., Ehrman J.M. & Harris J. (2005) Diversity and Distribution of Epibiotic Bacteria on Pseudo-nitzschia Multiseries (Bacillariophyceae) in Culture, and Comparison with Those on Diatoms in Native Seawater.
Kappler A., Benz M., Schink B. & Brune A. (2004) Electron shuttling via humic acids in microbial iron(III) reduction in a freshwater sediment. FEMS Microbiology Ecology 47, 85–92.
Khandeparker L., D’Costa P.M., Anil A.C. & Sawant S.S. (2013) Interactions of bacteria with diatoms: Influence on natural marine biofilms. Marine Ecology, 233–248.
Kirschner a. K.T. & Velimirov B. (1997) A seasonal study of bacterial community succession in a temperate backwater system, indicated by variation in morphotype numbers, biomass, and secondary production. Microbial Ecology 34, 27–38.
Klausen M., Heydorn A., Ragas P., Lambertsen L., Aaes-Jørgensen A., Molin S., et al. (2003) Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Molecular Microbiology 48, 1511–1524.
Kofoed M.V.W., Nielsen D.Å., Revsbech N.P. & Schramm A. (2012) Fluorescence in situ hybridization (FISH) detection of nitrite reductase transcripts (nirS mRNA) in Pseudomonas stutzeri biofilms relative to a microscale oxygen gradient. Systematic and Applied Microbiology 35, 513–517.
Kohler T., Curty L.K., Barja F., Van Delden C. & Pechere J.C. (2000) Swarming of Pseudomonas aeruginosa is dependent on cell-to-cell signaling and requires flagella and pili. Journal of Bacteriology 182, 5990–5996.
Köster D. & Pienitz R. (2006) Seasonal diatom variability and paleolimnological inferences - A case study. Journal of Paleolimnology 35, 395–416.
Lalonde K., Mucci A., Ouellet A. & Gélinas Y. (2012) Preservation of organic matter in sediments promoted by iron. Nature 483, 198–200.
Lambert C., Hobley L., Chang C.Y., Fenton A., Capeness M. & Sockett L. (2008) A Predatory Patchwork: Membrane and Surface Structures of Bdellovibrio bacteriovorus. Elsevier Masson SAS.
Larsen S., Nielsen L.P. & Schramm A. (2015) Cable bacteria associated with long-distance electron transport in New England salt marsh sediment. Environmental Microbiology Reports 7, n/a–n/a.
Logue J.B. & Lindström E.S. (2008) Biogeography of bacterioplankton in inland waters. 99–114.
Lovley D.R. (1991) Dissimilatory Fe ( III ) and Mn ( IV ) Reduction. 55, 259–287. Lovley D.R. (2012a) Electromicrobiology. Annual Review of Microbiology 66, 391–
409. Lovley D.R. (2012b) Long-range electron transport to Fe(III) oxide via pili with
metallic-like conductivity. Biochemical Society transactions 40, 1186–90. Lovley D.R., Ueki T., Zhang T., Malvankar N.S., Shrestha P.M., Flanagan K. a., et
al. (2011) Geobacter. The Microbe Electric’s Physiology, Ecology, and Practical Applications, 1st edn. Elsevier Ltd.
Luke N.R., Howlett A.J., Shao J. & Campagnari A. a (2004) Expression of Type IV Pili by. 72, 6262–6270.
Maerki M., Müller B., Dinkel C. & Wehrli B. (2009) Mineralization pathways in lake sediments with different oxygen and organic carbon supply. Limnology and Oceanography 54, 428–438.

66
Maerki M., Müller B. & Wehrli B. (2006) Microscale mineralization pathways in surface sediments: A chemical sensor study in Lake Baikal. Limnology and Oceanography 51, 1342–1354.
Maerki M., Wehrli B., Dinkel C. & Müller B. (2004) The influence of tortuosity on molecular diffusion in freshwater sediments of high porosity. Geochimica et Cosmochimica Acta 68, 1519–1528.
Mahmoud K.K. & Koval S.F. (2010) Characterization of type IV pili in the life cycle of the predator bacterium Bdellovibrio. Microbiology 156, 1040–1051.
Malfatti F., Samo T.J. & Azam F. (2010) High-resolution imaging of pelagic bacteria by Atomic Force Microscopy and implications for carbon cycling. The ISME journal 4, 427–439.
Malkin S.Y., Rao A.M., Seitaj D., Vasquez-Cardenas D., Zetsche E.-M., Hidalgo-Martinez S., et al. (2014) Natural occurrence of microbial sulphur oxidation by long-range electron transport in the seafloor. The ISME journal 8, 1843–1854.
Martiny J.B.H., Bohannan B.J.M., Brown J.H., Kane M., Krumins J.A., Kuske C.R., et al. (2006) Microbial biogeography : putting microorganisms on the map. 4, 102–112.
Mattick J.S. (2002) Type IV pili and twitching motility. Annual review of microbiology 56, 289–314.
McBride M.J. (2001) Bacterial gliding motility: multiple mechanisms for cell movement over surfaces. Annual review of microbiology 55, 49–75.
McBride M.J. (2004) Cytophaga-flavobacterium gliding motility. Journal of Molecular Microbiology and Biotechnology 7, 63–71.
Meyerreil L. a. (1994) Microbial life in sedimentary biofilms - The challenge to microbial ecologists. Marine Ecology Progress Series 112, 303–311.
Myklestad S.M. (1995) Release of extracellular products by phytoplankton with special emphasis on polysaccharides. Science of the Total Environment 165, 155–164.
Nealson K.H. (1997) SEDIMENT BACTERIA : Who ’ s There , What Are They Doing , and What ’ s New ? Earth.
Nielsen L.P. & Risgaard-Petersen N. (2015) Rethinking Sediment Biogeochemistry After the Discovery of Electric Currents. Annual Review of Marine Science 7, 425–442.
O’Toole G. a & Kolter R. (1998) Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol Microbiol 30, 295–304.
Oksanen J. (2015) Vegan : ecological diversity. Olff H., Alonso D., Berg M.P., Eriksson B.K., Loreau M., Piersma T., et al. (2009)
Parallel ecological networks in ecosystems. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 364, 1755–1779.
Otto S.A., Diekmann R., Flinkman J., Kornilovs G. & Mo C. (2014) Habitat Heterogeneity Determines Climate Impact on Zooplankton Community Structure and Dynamics. 9.
Pace M. & Prairie Y. (2005) Respiration in lakes. Respiration in aquatic ecosystems, 103–121.
Paquete C.M., Fonseca B.M., Cruz D.R., Pereira T.M., Pacheco I., Soares C.M., et al. (2014) Exploring the molecular mechanisms of electron shuttling across the microbe/metal space. Frontiers in Microbiology 5, 1–12.
Paranjpye R.N., Johnson A.B., Baxter A.E. & Strom M.S. (2007) Role of type IV pilins in persistence of Vibrio vulnificus in Crassostrea virginica oysters. Applied and Environmental Microbiology 73, 5041–5044.

67
Peabody C.R., Chung Y.J., Yen M.R., Vidal-Ingigliardi D., Pugsley A.P. & Saier M.H. (2003) Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. Microbiology 149, 3051–3072.
Pedro C. (2006) Marine microbial diversity : can it be determined ? 14. Peter H., Beier S., Bertilsson S., Lindström E.S., Langenheder S. & Tranvik L.J.
(2011) Function-specific response to depletion of microbial diversity. The ISME journal 5, 351–361.
Pfeffer C., Larsen S., Song J., Dong M., Besenbacher F., Meyer R.L., et al. (2012) Filamentous bacteria transport electrons over centimetre distances. Nature, 10–13.
Revsbech N.P., Jørgensen B.B., Blackburn T.H. & Cohen Y. (1983) Microelectrode studies of the photosynthesis and O2, H2S, and pH profiles of a microbial mat. Limnology and Oceanography 28, 1062–1074.
Ritz K., Mcnicol J.W., Nunan N., Grayston S., Millard P. & Atkinson D. (2004) Spatial structure in soil chemical and microbiological properties in an upland grassland. 49, 191–205.
De Rosa M., Gambacorta a., Millonig G. & Bu’Lock J.D. (1974) Convergent characters of extremely thermophilic acidophilic bacteria. Experientia 30, 866–868.
De Rosa M., Gambacorta A. & Bu’lock J.D. (1975) Extremely thermophilic acidophilic bacteria convergent with Sulfolobus acidocaldarius. Journal of general microbiology 86, 156–164.
Rösel S. & Grossart H.P. (2012) Contrasting dynamics in activity and community composition of free-living and particle-associated bacteria in spring. Aquatic Microbial Ecology 66, 169–181.
Rosu H.C., Murguía J.S. & Ibarra-Junquera V. (2010) Detection of mixed-culture growth in the total biomass data by wavelet transforms. Journal of Applied Research and Technology 8, 240–248.
Røy H., Huettel M. & Jørgensen B.B. (2004) Transmission of oxygen concentration fluctuations through the diffusive boundary layer overlying aquatic sediments. Limnology and Oceanography 49, 686–692.
Sapp M., Schwaderer A.S., Wiltshire K.H., Hoppe H., Gerdts G., Wicheis A., et al. (2015) Microbial Ecology ! Bacterial Communities in the Phycosphere of Microalgae ? 53, 683–699.
Sapp M., Wichels A. & Gerdts G. (2007) Impacts of cultivation of marine diatoms on the associated bacterial community. Applied and Environmental Microbiology 73, 3117–3120.
Schäfer H., Abbas B., Witte H. & Muyzer G. (2002) Genetic diversity of “satellite” bacteria present in cultures of marine diatoms. FEMS Microbiology Ecology 42, 25–35.
Schelert J., Dixit V., Hoang V., Drozda M., Blum P. & Simbahan J. (2004) Occurrence and Characterization of Mercury Resistance in the Hyperthermophilic Archaeon Sulfolobus solfataricus by Use of Gene Disruption Occurrence and Characterization of Mercury Resistance in the Hyperthermophilic Archaeon Sulfolobus solfataricus by Us. 186, 427–437.
Schindler D.W., Carpenter S.R., Cole J.J., Kitchell J.F. & Pace M.L. (1997) Influence of Food Web Structure on Carbon Exchange between Lakes and the Atmosphere. Science 277, 254–251.
Shannon C.E. (1948) A mathematical theory of communication. The Bell System Technical Journal 27, 379–423.

68
Shime-Hattori A., Iida T., Arita M., Park K.S., Kodama T. & Honda T. (2006) Two type IV pili of Vibrio parahaemolyticus play different roles in biofilm formation. FEMS Microbiology Letters 264, 89–97.
Sinclair L., Osman O.A., Bertilsson S. & Eiler A. (2015) Microbial Community Composition and Diversity via 16S rRNA Gene Amplicons: Evaluating the Illumina Platform. Plos One 10, e0116955.
Sison-Mangus M.P., Jiang S., Tran K.N. & Kudela R.M. (2014) Host-specific adaptation governs the interaction of the marine diatom, Pseudo-nitzschia and their microbiota. The ISME journal 8, 63–76.
Soininen J., Korhonen J.J., Karhu J. & Vetterli A. (2011) Disentangling the spatial patterns in community composition of prokaryotic and eukaryotic lake plankton. 56, 508–520.
Soto G.E. & Hultgren S.J. (1999) Bacterial adhesins: Common themes and variations in architecture and assembly. Journal of Bacteriology 181, 1059–1071.
Srikanth S., Marsili E., Flickinger M.C. & Bond D.R. (2008) Electrochemical characterization of Geobacter sulfurreducens cells immobilized on graphite paper electrodes. Biotechnology and Bioengineering 99, 1065–1073.
Stocker R. (2012) Marine Microbes See a Sea of Gradients. Science 338, 628–633. Stoermer E.F. (1993) Evaluating diatom succession: some pecularities of the Great
Lakes case. Journal of Paleolimnology 8, 71–83. Straub K.L. & Schink B. (2003) Evaluation of electron-shuttling compounds in
microbial ferric iron reduction. FEMS Microbiology Letters 220, 229–233. Szabó Z., Stahl A.O., Albers S. V., Kissinger J.C., Driessen a. J.M. & Pohlschröder
M. (2007) Identification of diverse archaeal proteins with class III signal peptides cleaved by distinct archaeal prepilin peptidases. Journal of Bacteriology 189, 772–778.
Torsvik V., Øvreås L. & Thingstad T.F. (2002) Prokaryotic diversity — magnitude , dynamics, and controlling factors. Science 296, 1064–6.
Tranvik L.J., Downing J. a., Cotner J.B., Loiselle S. a., Striegl R.G., Ballatore T.J., et al. (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnology and Oceanography 54, 2298–2314.
Vadeboncoeur Y., Zanden M.J. Vander & Lodge D.M. (2002) Putting the Lake Back Together : Reintegrating Benthic Pathways into Lake Food Web Models. 52.
Varga J.J., Nguyen V., O’Brien D.K., Rodgers K., Walker R. a. & Melville S.B. (2006) Type IV pili-dependent gliding motility in the Gram-positive pathogen Clostridium perfringens and other Clostridia. Molecular Microbiology 62, 680–694.
Vasquez-Cardenas D., van de Vossenberg J., Polerecky L., Malkin S.Y., Schauer R., Hidalgo-Martinez S., et al. (2015) Microbial carbon metabolism associated with electrogenic sulphur oxidation in coastal sediments. The ISME Journal, 1–13.
Vorenhout M., van der Geest H.G., van Marum D., Wattel K. & Eijsackers H.J.P. (2004) Automated and continuous redox potential measurements in soil. Journal of environmental quality 33, 1562–1567.
Vu B., Chen M., Crawford R.J. & Ivanova E.P. (2009) Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14, 2535–2554.
Wagner M., Berkner S., Ajon M., Driessen A.J.M., Lipps G. & Albers S.-V. (2009) Expanding and understanding the genetic toolbox of the hyperthermophilic genus Sulfolobus. Biochemical Society transactions 37, 97–101.

69
Wagner M., van Wolferen M., Wagner A., Lassak K., Meyer B.H., Reimann J., et al. (2012) Versatile genetic tool box for the crenarchaeote Sulfolobus acidocaldarius. Frontiers in Microbiology 3, 1–12.
Wall D. & Kaiser D. (1999) Type IV pili and cell motility. Molecular microbiology 32, 1–10.
Watnick P. & Kolter R. (2000) Biofilm, city of microbes. Journal of Bacteriology 182, 2675–2679.
Wetzel R.G. (2001) Limnology. Elsevier. Whitfield C. & Keenleyside W.J. (1995) Regulation of expression of group IA
capsular polysaccharides in Escherichia coli and related extracellular polysaccharides in other bacteria. Journal of Industrial Microbiology 15, 361–371.
Woese C.R. & Fox G.E. (1977) Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences of the United States of America 74, 5088–5090.
Zhang K., Choi H., Dionysiou D.D., Sorial G. a. & Oerther D.B. (2006) Identifying pioneer bacterial species responsible for biofouling membrane bioreactors. Environmental Microbiology 8, 433–440.
Zillig W., Stetter K.O., Wunderl S., Schulz W., Priess H. & Scholz I. (1980) The Sulfolobus-“Caldariella” group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases. Archives of Microbiology 125, 259–269.
ZoBell C.E. (1946) Studies on Redox Potential of Marine Sediments. AAPG Bulletin 4, 477–513.

Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1296
Editor: The Dean of the Faculty of Science and Technology
A doctoral dissertation from the Faculty of Science andTechnology, Uppsala University, is usually a summary of anumber of papers. A few copies of the complete dissertationare kept at major Swedish research libraries, while thesummary alone is distributed internationally throughthe series Digital Comprehensive Summaries of UppsalaDissertations from the Faculty of Science and Technology.(Prior to January, 2005, the series was published under thetitle “Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-263206
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2015