membrane specializations in the developmentally transient perireticular nucleus of the rat
TRANSCRIPT

Membrane Specializationsin the Developmentally TransientPerireticular Nucleus of the Rat
EION J. RAMCHARAN* AND R.W. GUILLERY
Department of HumanAnatomy, University of Oxford,Oxford OX1 3QX, United Kingdom
ABSTRACTThe perireticular nucleus lies among the fibres of the internal capsule and in the rat is
relatively large at birth. Like the subplate of the neocortex, it is dramatically reduced in theadult (Mitrofanis, 1992). On embryonic day 17 (E17), cells of the perireticular nucleus projectto the cortex (Adams and Baker, 1995); at birth there is also a projection to the thalamus(Mitrofanis and Baker, 1993). When corticofugal axons reach the perireticular nucleus, theyseparate into the descending tracts and the corticothalamic pathway. The axons then enterthe internal capsule on their way to the thalamus, defasciculate and contribute to a complexzone of interweaving fibres seen in this region (Adams and Guillery, 1994). We have found thatthe cells of the perireticular nucleus are immunopositive for an antibody (3A10) to aphosphorylated neurofilament protein (Yamada et al., 1991), and we have shown that these3A10-positive perireticular cells extend their dendrites across the path of cortical fibres at theearliest age studied (E17). We have confirmed this dendritic orientation by Golgi staining at alater stage (P3). Electron micrographs show that axo-somatic and axo-dendritic synapses firstappear in the perireticular nucleus at about the day of birth. Prenatally, between E14and E19, the major specialized membrane interrelationships seen are omega formationsbetween adjacent profiles that often contain vesicles. Omega formations are rarely seen atlater stages. Puncta adhaerentia typified by opposing membrane densities with no associatedvesicles were seen at all ages studied. We suggest that early membrane contacts (omegaformations) may represent transient organisational or guidance influences since they occurduring a period when axonal pathways are being defined. J. Comp. Neurol. 380:435–448,1997. r 1997 Wiley-Liss, Inc.
Indexing terms: junctions; synapses; Golgi staining; omega formations; 3A10
The corticothalamic and the thalamocortical fibres turnabruptly toward their eventual targets in the area of arecently described group of cells, the perireticular nucleus(Clemence and Mitrofanis, 1992; Mitrofanis, 1992). Thisnucleus, which lies medial to the globus pallidus andlateral to the thalamic reticular nucleus, is also the regionof a complex interweaving of fibres (Mitrofanis and Guil-lery, 1993). On encountering the cells of the perireticularnucleus, the corticofugal axons separate into two distinctfibre tracts: One continues past these cells and goes on toform the corticospinal and corticobulbar pathways, whilethe axons destined to terminate in the thalamus turnsharply toward their target and defasciculate in the area ofthe perireticular nucleus before entering the dorsal thala-mus (Mitrofanis and Guillery, 1993).The corticothalamic fibres develop during a period when
the perireticular nucleus is growing to its largest size.After the pathways between the cortex and thalamus have
been established, the size of the nucleus is markedlyreduced (Mitrofanis, 1992). This developmental transienceof the nucleus, which is comparable to that of the subplatecells of the neocortex (Kostovic and Rakic, 1980), has led tothe suggestion that the perireticular nucleus plays asignificant role in directing the formation of connectionsbetween the cortex and the thalamus. The perireticular
Grant sponsor: Wellcome Trust.Eion J. Ramcharan’s current address is Department of Neurobiology,
State University of NewYork, Stony Brook, NewYork 11794-5230.R.W. Guillery’s current address is Department of Anatomy, University of
Wisconsin School of Medicine, 1300UniversityAvenue,Madison,Wisconsin53706.*Correspondence to: Eion J. Ramcharan, Department of Neurobiology,
State University of NewYork, Stony Brook, NY 11794-5230.Received 11 December 1995; Revised 26 August 1996; Accepted 7
November 1996.
THE JOURNAL OF COMPARATIVE NEUROLOGY 380:435–448 (1997)
r 1997 WILEY-LISS, INC.

cells project to the cortex around a time (E17 in the rat)when the thalamic axons first reach their cortical targets,and cortical fibres are already on their way through theinternal capsule (Adams and Baker, 1995). This suggestseither that thalamic fibres may be guided toward theirtargets by the axons of the perireticular cells or thatgrowing corticofugal axonsmay use the perireticular fibresto navigate their passage at least as far as the internalcapsule. Perireticular cells also send axons to the thala-mus, but this connection is not made until the day of birth(Mitrofanis and Baker, 1993).In order to look further at how the axons that pass
through the perireticular nucleus may relate to these cells,we have made a light and electron microscopic study ofthis region. We have looked with the electron microscopeat membrane specializations between the cellular profilesin the region of the perireticular cells as an indication ofthe influence one neuron may exert on another in thisregion; andwe have concentrated particularly on determin-ing the first appearance of synapses in order to correlatethe onset of synaptogenesis with the time at which axonsturn toward their target. Perireticular cells in early em-bryos were identified both by their position in the develop-ing internal capsule and by immunohistochemical label-ling with an antibody (3A10, Yamada et al., 1991) to aphosphorylated neurofilament protein, which was found toimmunostain cells specifically of the diencephalon. Golgiimpregnation of perireticular cells also served to show theorientation of the cells and their relationship to axonsentering the internal capsule.
MATERIALS AND METHODS
Lister-Hooded rats ranging in age from embryonic day14 (E14) to postnatal day 3 (P3) were used in this study.Animal care protocols and procedures were carried out inaccordance with Home Office regulations. All fetal ratswere obtained from timedmatings. The first appearance ofa seminal plug after a female and male had been puttogether was taken as E0 of the pregnancy. The day ofbirth was taken as P0. Pregnant females were anaesthe-tized with an overdose of sodium pentobarbitone adminis-tered intraperitoneally, and fetuses were removed andperfused with one of the following fixatives: 1) a mixturecontaining 2% paraformaldehyde, 2.5% glutaraldehyde,and 15% of a saturated solution of picric acid; or 2) amixture containing 4% paraformaldehyde and 1% glutaral-dehyde. All fixatives were buffered with 0.1M phosphatebuffer pH 7.4. Postnatal animals were similarly anaesthe-tized and first perfused with phosphate-buffered saline(PBS), followed by perfusion fixation.
Processing for electron microscopy
Hemisected brains were rinsed in phosphate buffer,postfixed in 1–2% osmium tetroxide (in phosphate bufferat pH 7.4) for one to five hours, washed in the same buffer,and stained en bloc in 1% uranyl acetate in 70% ethanol forone hour. Brains were dehydrated in increasing concentra-tions of ethanol, immersed in propylene oxide, left over-night in a mixture of Araldite and propylene oxide (1:1),then embedded in fresh Araldite and baked at 60°C for 48hours. Coronal semithin sections (1 µm) were cut, thenstained with 1% methylene blue, and the area of interestwas identified and drawn with the aid of a camera lucida.Thin sections (silver to gold) of the chosen area were
collected, stained with 1% aqueous uranyl acetate followedby lead citrate, and viewed under a Jeol 1010 transmissionelectron microscope.
3A10 Immunohistochemistry
Embryonic brains for immunohistochemistry were per-fused with a mixture of 4% paraformaldehyde, 0.05%glutaraldehyde, and 15% of a saturated solution of picricacid buffered in 0.1M phosphate buffer, pH 7.4. 100 µmVibratome sections were incubated in 10% rabbit serumand then left in a mouse monoclonal antibody raisedagainst 3A10 (1:100 in PBS; collected from hybridoma cellsobtained from the Developmental Studies HybridomaBank, Iowa) overnight at 4°C. After a two-hour incubationin a biotinylated rabbit anti-mouse secondary antibody,the sections were left in avidin conjugated to horseradishperoxidaseABC solution (Vectastain Kit) for one hour. TheABC-bound peroxidase was revealed by a 3,3 diaminoben-zidine (0.05%) 0.006%H2O2mixture. Sections were washedin PBS between incubations. The sections were osmicatedin 1% osmium tetroxide (buffered with 0.1% phosphatebuffer) and processed for electron microscopy as describedabove. Although the tissue preservation of material pre-pared in this manner was poor, the Vibratome sectionswere useful for showing the orientation of perireticularcells and their dendrites.
Single-section Golgi preparation
100 µm Vibratome sections were immersed in 1% os-mium tetroxide for 40 minutes, washed, and then left in3.5% potassium dichromate overnight. The sections wereplaced between two glass slides and left in 2% silvernitrate solution for six hours (see Bolam, 1992). Sectionswere then mounted in glycerine and coverslipped.
RESULTS
Perireticular cellsare immunopositive for 3A10
At E17 the cells of the perireticular nucleus are clearlyimmunoreactive for the antibody 3A10 (Fig. 1). In 100 µmVibratome sections that were immunohistochemicallytreated for 3A10, the cell bodies and the proximal den-drites of perireticular cells are heavily labelled. However,no fine dendritic branches are stained (Fig. 1b). Most of theimmunopositive cells have two major dendrites giving thecells a bipolar appearance (Fig. 1b). The orientation of thecells is in an oblique dorsoventral plane roughly perpen-dicular to the path of corticofugal fibres entering theperireticular nucleus on their way to the thalamus orlower centres. In the plane of the coronal sections, two tothree of these cells in a row can span the whole area of theincoming axons. Themorphology and the orientation of theperireticular cells in relation to the incoming axons sug-gests, therefore, that the relatively small perireticularnucleus, which lies in the path of most of the axons passingthrough the internal capsule, has individual cells lying inthe path of a great many of the axons.
Golgi preparation of perireticular cells
Golgi preparations of 100 µm Vibratome sections fromP3 rat brains confirm that perireticular cells extend theirproximal dendrites perpendicularly across the path offibres of the internal capsule (Fig. 2a–c).
436 E.J. RAMCHARAN AND R.W. GUILLERY

Fig. 1. a:A low-power line drawing of a coronal, 100 µm Vibratomesection from an E17 rat brain, processed for 3A10 immunoreactivity.PRN, perireticular nucleus*; TRN, thalamic reticular nucleus; GP,globus pallidus. The arrows indicate the path of cortical axons
entering the perireticular nucleus. b: Higher-power photomicrographof the cells in the PRN. The cells are roughly in the same orientation asin the low-power drawing. The arrows show the course of axonsentering the PRN. Scale bar 5 50 µm.
PERIRETICULAR MEMBRANE CONTACTS 437

Fig. 2. a:A low-power line drawing of a coronal, 100 µm Vibratomesection from a P3 rat brain, processed for Golgi staining. The positionsof a perireticular cell and axons entering the perireticular nucleusshown in Fig. 2b and 2c aremarked by the arrow. The related areas areshown for orientation. PRN, perireticular nucleus; ST, striatum; GP,globus pallidus; TRN, thalamic reticular nucleus; DT, dorsal thala-
mus; FIM, fimbria. b: A camera lucida drawing showing the orienta-tion of a perireticular cell relative to the axons entering the nucleus. c:A photomicrograph of the same perireticular cell as in Fig. 2bemphasizing the relationship between the dendritic alignment and thecourse of the axons. Scale bars 5 500 µm in a, 50 µm in b, c.
438 E.J. RAMCHARAN AND R.W. GUILLERY

Electron microscopy of junctionalspecializations
Axons invading the internal capsule form various typesof membrane contacts with the cells of the perireticularnucleus at all ages studied (E14–P3). In the neonatal andearly postnatal rats the area of the perireticular nucleusconsists mainly of neuronal cell bodies, unmyelinatedaxons, growth cones, and glial elements. Glial processescan often be identified by the presence of ribosomes andintermediate glial filaments in the cytoplasm. Growthcones are recognised by their occasional filopodial exten-sions and their clear, uniform cytoplasm, lacking ribo-somes. Axons with microtubules in their axoplasm and noribosomes can be relatively easily distinguished from otherprofiles. Synapses can be identified by the criteria definedby Peters et al. (1976): If they exhibited typical synapticdensities and clustering of clear synaptic vesicles in thepresynaptic terminal, and if synaptic clefts were foundbetween the pre- and postsynaptic membranes, they wererecorded as synaptic junctions (Fig. 3a). However, not all ofthese features were clearly identifiable in all junctionalspecializations, and a wide range of synapse-like junctionswere seen (Fig. 3b–f). The variation tended to be due to: 1)the apparent thickness of the densities, which is often theresult of the obliquity of the cut; 2) the absence of a clearlyidentifiable synaptic cleft, which again is often dependenton the obliquity of the cut; and 3) the relatively sparsedistribution of identifiable vesicles around the active zoneof the synapse (Fig. 3b–f).At the earliest stages studied (E14–E19), there was no
evidence of synapse-like junctions in the perireticularnucleus or anywhere in the region of the internal capsule.The first clearly identifiable synapses were seen on postna-tal day 1 (P1). Although synapses were not easy to find,and first impressions suggested that there were only a few,a survey of an area approximately 10,000 µm2, using theX-Y plotting device of the electronmicroscope, showed that95 synapse-like junctions were present in a central part ofa caudal portion of the nucleus at P1 in one section. In acentral zone from the rostral part of the nucleus, 27synapse-like junctions were found in a 10,000 µm2 area atthis stage. About 10% of the junctions were classified asdoubtful and were not included in the above counts. Whena similar area of the perireticular nucleus of an E19 brainwas scanned, no synaptic junctions were found.Synaptic junctions at P1 were found between growing
axons and dendrites (axo-dendritic) (Fig. 4a, b, large star)or between axons and cell bodies (axo-somatic) (Fig. 4d).Sometimes synapses were also seen between two profiles,one of which could be identified as axonal whereas theidentity of the other profile was often unclear. Figure 4 (a,c, small star) shows one such synapse where one element isan axon but the other could be either an axon or a dendrite.Although synapses were most clearly seen at P1 and later,some synapse-like junctions were seen at P0.The length of the synaptic density in the clearly identifi-
able synapses was relatively short. However, in some casesthere were relatively longer, darker membrane zones,between adjacent profiles, compared to membranes in theimmediate vicinity. Although they did not have the samethickness seen in obviously synaptic junctions, they wereoccasionally associated with a small cluster of vesicles.Figure 5 shows one such junction in the perireticularnucleus of a P1 brain. The characteristics of this form of
membrane specialization suggest that these opposingmem-brane interfaces with their associated vesicles may repre-sent early stages of synapse formation. In sections from aP3 brain, synapses were more clearly mature (Fig. 6a–d);none of the immature forms were seen at this age. Theprofiles that appeared to be immature synapses were notincluded in the counts of synapses in the perireticularnucleus.
Other types of membrane specializations
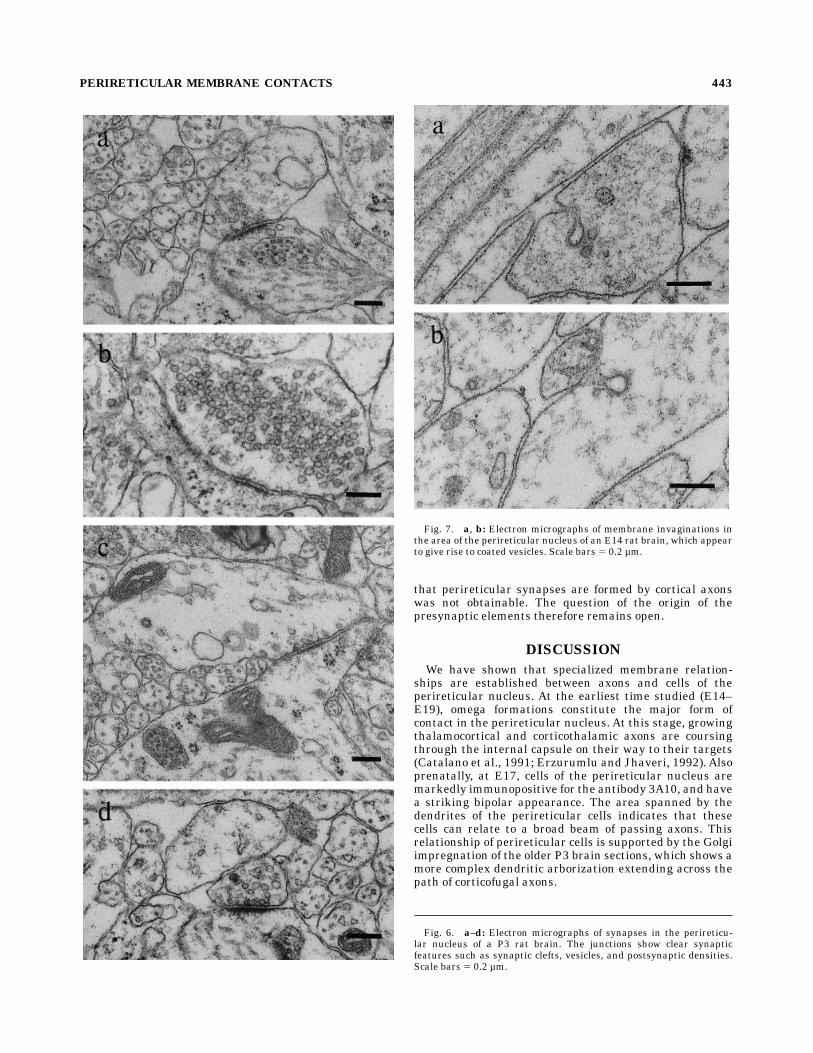
There is a high incidence of omega formations (Figs.7a,b, 8a–d) between neighbouringmembranes in the perire-ticular nucleus and throughout the internal capsule at theearliest ages studied (E14–E19). The presence of omegaformations in early development does not appear to in-volve the membranes of cell bodies. They are found be-tween abutting membranes of axons, dendrites, and glia.Figures 7a and 7b show two such membrane invaginationsfrom the internal capsule of an E14 embryonic brain. Thecellular elements that contribute to omega formations orlie adjacent to them often contain vesicles of variousshapes and sizes, and these may have been the result ofsimilar membrane invaginations (Fig. 8d). These mem-brane specializations, which first occur several days beforesynapse formation, appear to be the result of a cellularelement either internalizing a portion of its own mem-brane (single-membrane omega) (Fig. 8a) or endocytosinga part of the membrane of an adjacent profile (Fig. 8b).These membrane contacts were most commonly foundbetween E14 and E19 (the earliest ages studied); this is atime when cortical and thalamic axons that have probablyentered the internal capsule (Catalano et al., 1991; Erzu-rumlu and Jhaveri, 1992) change their course.The membrane specializations described above are
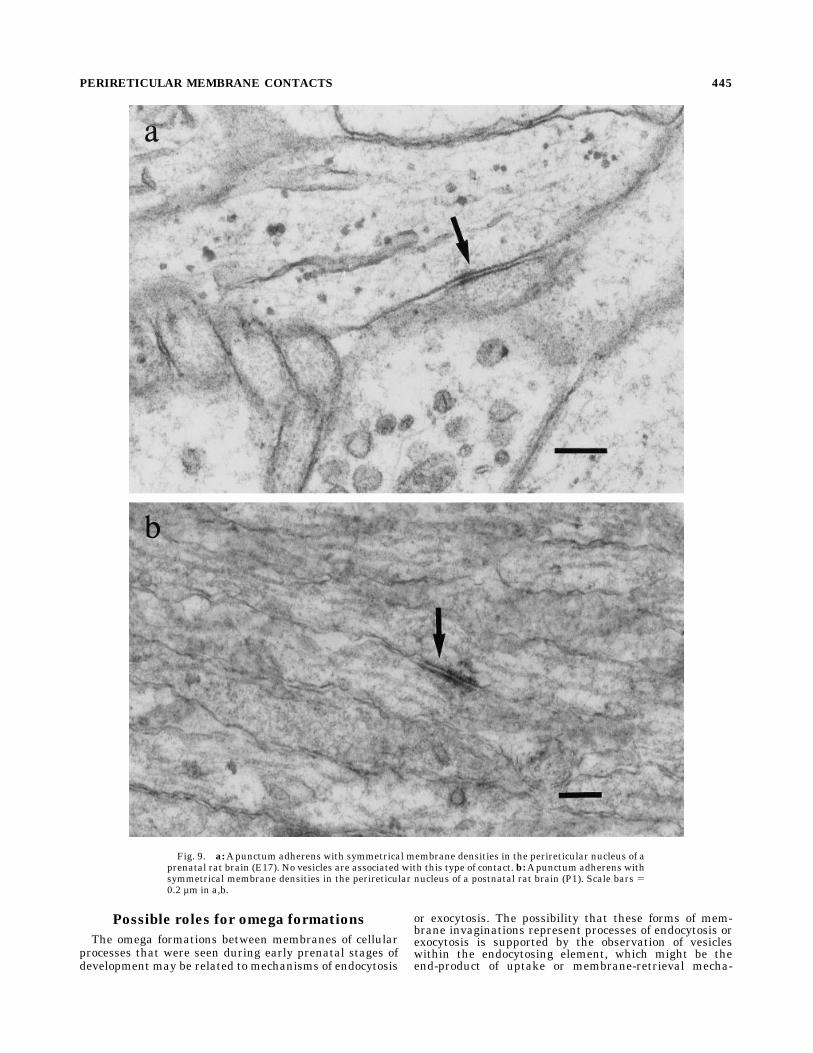
clearly distinct from the puncta adhaerentia, which wereseen in the perireticular nucleus at all ages studied (Fig.9). These junctions, which are characterized by symmetri-cal densities, are never associated with clusters of vesicles.Although these membrane contacts are thought to sub-serve adhesion properties, early prenatal forms may beassociated with developmentally significant cellular inter-actions.
What is the origin of the axons contributingto the membrane specializations?
Biocytin, a predominantly anterograde tracer, was in-jected into the cortex of newborn rat pups (P0–P2) todetermine the origin of the axons that form membranespecializations with cells of the perireticular nucleus.Light microscopy of labelled axons showed beaded axonsfrom the cortex entering the internal capsule. The axonswere beaded over their entire visible course, not only in theinternal capsule (Fig. 10). Figure 10a shows beaded axonsof the corticothalamic tract (CT) and the descending tracts(DT). Figure 10b shows beaded axons in the intermediatezones of the neocortex. Retrogradely labelled cells wereseen only rarely in the dorsal thalamus. The beads on theaxons were interpreted as possibly representing sites ofjunctional interactions (Beaudet and Descarries, 1978; seediscussion). Electron microscopy showed that the earlysynaptic junctions were but poorly preserved in the biocy-tinmaterial. Clearly identifiable synapses thatwere readilyvisible in sections from conventional processing for elec-tron microscopy of material at the same stage (see above)were extremely rare in the biocytin material. There were
PERIRETICULAR MEMBRANE CONTACTS 439

Fig. 3. a: An electron micrograph of a synapse in the perireticularnucleus of a P1 rat brain. At this magnification all the features of aclassical synapse can be seen, i.e., synaptic vesicles, a postsynapticdensity, and a synaptic cleft. b–f:A range of the synapse-like junctionsthat were commonly seen in the perireticular nucleus of P1 rat brains.
The synaptic clefts were often not seen because of the obliquity of thecut. However, the presence of synaptic vesicles and densities were themost obvious identifying features of synapses. Scale bars 5 0.2 µm ina, 0.5 µm in b, 1 µm in c–f.
440 E.J. RAMCHARAN AND R.W. GUILLERY

only two to three identifiable synapses in a 10,000 µm2
area of the internal capsule. This compares poorly with the95 synaptic junctions seen in a similar area of the internal
capsule of a brain treated for standard electron microscopyonly. Because of the lack of adequate preservation of thepresumably delicate early postnatal tissue, clear evidence
Fig. 4. a: Electron micrograph showing an axo-dendritic synapse(large star) and a synapse between an axon and a neurite (axon ordendrite) (small star). b: A higher-power electron micrograph of theaxo-dendritic synapse (large star) in Fig. 4a. c: A higher-power
electron micrograph of the axo-neuritic synapse (small star) in Fig. 4a.d: Electron micrograph of an axo-somatic synapse at P1. Scale bar 5 1µm in a, 0.5 µm in b–d.
PERIRETICULAR MEMBRANE CONTACTS 441

Fig. 5. Electron micrograph of an axon in the perireticular nucleus of a P1 brain forming a putativesynapse with the membrane of a cell body. The inset is a high-power micrograph of the junction showingthe cluster of vesicles and the beginnings of a synaptic density. Scale bars 5 1 µm, 0.5 µm in inset.
that perireticular synapses are formed by cortical axonswas not obtainable. The question of the origin of thepresynaptic elements therefore remains open.
DISCUSSION
We have shown that specialized membrane relation-ships are established between axons and cells of theperireticular nucleus. At the earliest time studied (E14–E19), omega formations constitute the major form ofcontact in the perireticular nucleus. At this stage, growingthalamocortical and corticothalamic axons are coursingthrough the internal capsule on their way to their targets(Catalano et al., 1991; Erzurumlu and Jhaveri, 1992). Alsoprenatally, at E17, cells of the perireticular nucleus aremarkedly immunopositive for the antibody 3A10, and havea striking bipolar appearance. The area spanned by thedendrites of the perireticular cells indicates that thesecells can relate to a broad beam of passing axons. Thisrelationship of perireticular cells is supported by the Golgiimpregnation of the older P3 brain sections, which shows amore complex dendritic arborization extending across thepath of corticofugal axons.
Fig. 6. a–d: Electron micrographs of synapses in the perireticu-lar nucleus of a P3 rat brain. The junctions show clear synapticfeatures such as synaptic clefts, vesicles, and postsynaptic densities.Scale bars 5 0.2 µm.
Fig. 7. a, b: Electron micrographs of membrane invaginations inthe area of the perireticular nucleus of an E14 rat brain, which appearto give rise to coated vesicles. Scale bars 5 0.2 µm.
PERIRETICULAR MEMBRANE CONTACTS 443

Fig. 8. All the electronmicrographs in this figure were taken in theperireticular nucleus of E17 rat brains. a: A few single membraneinvaginations were seen in the perireticular nucleus. This form ofmembrane specialization involves only one profile. b: This electronmicrograph shows an omega formation between two profiles in theperireticular nucleus. The double membrane of the omega indicates
the involvement of the membranes of both opposing elements. c: Ahigh-power micrograph of a commonly occurring omega-like mem-brane specialization between adjacent elements. d: This micrographshows profiles containing vesicular bodies of various sizes that wereprobably a result of omega-like invaginations. Scale bars 5 1 µm ina,b, 0.05 µm in c,d.
Possible roles for omega formations
The omega formations between membranes of cellularprocesses that were seen during early prenatal stages ofdevelopment may be related to mechanisms of endocytosis
or exocytosis. The possibility that these forms of mem-brane invaginations represent processes of endocytosis orexocytosis is supported by the observation of vesicleswithin the endocytosing element, which might be theend-product of uptake or membrane-retrieval mecha-
Fig. 9. a:Apunctum adherens with symmetrical membrane densities in the perireticular nucleus of aprenatal rat brain (E17). No vesicles are associated with this type of contact. b:Apunctum adherens withsymmetrical membrane densities in the perireticular nucleus of a postnatal rat brain (P1). Scale bars 50.2 µm in a,b.
PERIRETICULAR MEMBRANE CONTACTS 445

nisms. If the vesicles are transversely cut membrane-bound organelles such as smooth endoplasmic reticulum(SER), then the possibility that these membrane invagina-tions are related to retrieval of membrane or secretorymechanisms is highly likely (Rothman and Fine, 1980).There is some evidence to suggest that growth cones and
membranes of other cellular elements (glia) interact in a
similar manner (Gorgels, 1991). The functional conse-quence of endocytosis is unclear. It may be a means bywhich the membranes of growing axons are internalized,so that membrane-bound receptors are removed by theseprocesses and consequently the action mediated by thereceptors would be terminated. If these receptors are for afactor that is permissive to growth, then the internaliza-
Fig. 10. a: A photomicrograph of the internal capsule of a P1 brain showing beaded axons filled withbiocytin. The separation of corticofugal axons into the corticothalamic tract (CT) and the descendingtracts (DT) can be seen. b: Beaded axons in the intermediate zone of the neocortex in the same section asFig. 10a. Scale bars 5 10 µm in a,b.
446 E.J. RAMCHARAN AND R.W. GUILLERY

tion of membranes containing these receptors may be astop signal for growth in a particular direction; whereas ifthe receptors provide a cue that is inhibitory to growth,then endocytosis of these membrane-bound receptors mayallow growth to proceed along a certain route (Luo andRaper, 1994; Serafini et al., 1994). Uptake of membrane-bound adhesion molecules is also likely. Because some ofthese membrane interactions appear to occur betweenprofiles within bundles, this might be a way in whichneighbouring axons lose their adhesive affinities for eachother, resulting in defasciculation; and this defascicula-tion, previously described in the region of the perireticularnucleus, may contribute to the complex lattice-work seenin this area. This could then represent the early axonalreorganization that occurs as fibres pass among the cells inthe perireticular nucleus.
Possible roles of synaptic junctions
The axo-somatic and the axo-dendritic synapses seen inthe perireticular nucleus suggest that the postsynapticneurons in this nucleus do interact directly with presynap-tic axons. The formation of synapses in the perireticularnucleus at the time of birth shows that these sites ofcommunication between cells may not be necessary for theguidance of the earliest thalamocortical or corticothalamicaxons that grow through the internal capsule to their finaltargets, since they first appear at a time when many ofthese early axons have already reached their destinations.However, these junctions in the perireticular nucleus mayplay a role in influencing the developmental course of theaxons passing through the nucleus.It has been shown that the synthesis of neurotrophins
may be regulated by neuronal activity (Thoenen, 1995).Synaptic junctionsmay therefore influence the release andactions of neurotrophins. Whatever action is mediated bythese junctional specializations, the transient nature ofthe nucleus and the consequential transience of the syn-apses indicate that the functional mechanism(s) mediatedby these junctions must be primarily of developmentalsignificance. Presumably, once connections between thecortex and the thalamus have been stabilized, the synapticjunctions would detach or be withdrawn as perireticularcells begin to die off.The assumption being made here is that the synapses
seen in this study are axo-neuronal, but the possibilityexists that the growing axons may be forming synapticcontacts with glial cells in the perireticular nucleus (see,e.g., Gorgels, 1991). Our material shows some clearlyneuro-neuronal junctions. For example, the dendrite con-tributing to the axo-dendritic synapse in Figure 3a (andunder high power in 3b) contains clearly identifiablemicrotubules that are distinct from the more slenderintermediate filaments seen in the cytoplasm of glialprocesses (Kostovic and Rakic, 1990). Furthermore, theneuronal cell body of the axo-somatic synapse seen inFigure 3d has the characteristics of a differentiated neu-rone. It has a complex Golgi apparatus, an elaborateendoplasmic reticulum with polyribosomes, and a roundednucleus (not shown). Some junctions may be on glial cells,but a significant number of these synapses are formedupon neuronal cells of the perireticular nucleus.
Where do the axons contributingto the synapses originate?
The origins of the axons forming synapses with theperireticular cells is not known. They are most likely toarise in the cortex or the thalamus. Our attempts toaddress this question have proved unsuccessful. The factthat axons, labelled with biocytin following cortical injec-tions of the tracer, are beaded suggests that these axonsare forming junctions. Since these axons are beaded alongtheir entire length, including their passage through theintermediate zone, there must be some question about thefunctional significance of the beads. However, junctionsin the subplate region, for instance, may represent localguidance and/or refinement cues, a role that passes tojunctions in the new environs of the perireticular nucleusas axons course through the internal capsule. The perire-ticular nucleus could therefore be seen as acting in concertwith the subplate cells in influencing axonal targeting.
Possible role of Puncta adherentia
Puncta adherentia are thought to be sites where cellsadhere to each other. We found these junctional specializa-tions prenatally and postnatally in the perireticularnucleus. These junctions are also known to be abundant inadults, suggesting that the functional significance of theseinteractions may not be specific to development. However,it has been shown that several proteins that control thefate and proliferation of Drosophila epithelial cells (e.g.,Boss, Sevenless, and Notch) are localised in adherensjunctions (see the review by Woods and Bryant, 1993).There is therefore a possibility that these junctions play amore than simple adhesive role in mammalian develop-ment, and later act to maintain neighbourhood relation-ships.
CONCLUSIONS
In conclusion, we suggest that the early type of mem-brane contact—i.e., the omega formations, which may beendocytotic or exocytotic events—represent early tran-sient interactions related to axon guidance, growth, orsurvival, since they occur at times when axons are firstgrowing through the perireticular nucleus. Synapses inthe perireticular nucleus, on the other hand, may act in anactivity-dependent determination of pathway relation-ships as fibres pass between the thalamus and the cortex.This organisation, once set up, may be maintained byadhesion sites.
ACKNOWLEDGMENTS
Many thanks to Dr. Gary Baker for his advice. Thanksalso to David Stroud, Mohan Masih, Mary Walker, ZillahDeusen, FionaWatkins, and JulieWickson for their techni-cal support; and to BrianArcher and Colin Beesly for theirphotographic assistance.This work was supported by the Wellcome Trust.
LITERATURE CITED
Adams, N.C., and G.E. Baker (1995) Cells of the perireticular nucleusproject to the developing neocortex of the rat. J. Comp. Neurol.359:613–626.
PERIRETICULAR MEMBRANE CONTACTS 447

Adams, N.C., and R.W. Guillery (1994) Developing rat corticothalamicfibres defasciculate when they reach the perireticular nucleus. Soc.Neurosci. Abs. 20:1685.
Bolam, J.P. (1992) Preparation of central nervous system tissue for lightand electron microscopy. In J.P. Bolam (ed): Experimental Neuro-anatomy. APractical Approach. Oxford: IRL, pp. 1–29.
Beaudet, A., and L. Descarries (1978) The monoamine innervation of ratcerebellar cortex: Synaptic and nonsynaptic axon terminals. Neurosci-ence 3:851–860.
Catalano, S.M., R.T. Robertson, andH.P. Killackey (1991) Early ingrowth ofthalamocortical afferents to the neocortex of the prenatal rat. Proc.Natl. Acad. Sci. USA 88:2999–3003.
Clemence, A.E., and J. Mitrofanis (1992) Cytoarchitectonic heterogeneitiesin the thalamic reticular nucleus of cats and ferrets. J. Comp. Neurol.322:167–181.
Erzurumlu, R.S., and S. Jhaveri (1992) Emergence of connectivity in theembryonic rat parietal cortex. Cerebral Cortex 2:336–352.
Goodman, C., and C. Shatz (1993) Developmental mechanisms that gener-ate precise patterns of neuronal connectivity. Cell 72(Suppl.):77–98.
Gorgels, T.G.M.F. (1991) Junctional specializations between growth conesand glia in the developing rat pyramidal tract: Synapse-like contactsand invaginations. J. Comp. Neurol. 306:117–128.
Kostovic, I., and P. Rakic (1980) Cytology and time of origin of interstitialneurons in the white matter in infant and adult human and monkeytelencephalon. J. Neurocytol. 9:219–242.
Kostovic, I., and P. Rakic (1990) Developmental history of the transientsubplate zone in the visual and somatosensory cortex of the macaquemonkey and human brain. J. Comp. Neurol. 297:441–470.
Luo, Y., and J.A. Raper (1994) Inhibitory factors controlling growth conemotility and guidance. Curr. Opin. Neurobiol. 4:648–654.
Mitrofanis J. (1992) Patterns of antigenic expression in the thalamicreticular nucleus of developing rats. J. Comp. Neurol. 320:161–181.
Mitrofanis J., and G.E. Baker (1993) Development of the thalamic reticularand perireticular nuclei in rats and their relationship to growingcorticofugal and corticopetal axons. J. Comp. Neurol. 338:575–587.
Mitrofanis, J., and R.W. Guillery (1993) New views of the thalamic reticularnucleus in the adult and the developing brain. Trends Neurosci.16:240–245.
Peters, A., S.L. Palay, and H. deF. Webster (1976) The Fine Structure of theNervous System: The Neurons and Supporting Cells. Philadelphia:Saunders.
Rothman, J.E., and R.E. Fine (1980) Coated vesicles transport newlysynthesized membrane glycoproteins from endoplasmic reticulum toplasma membrane in two successive stages. Proc. Natl. Acad. Sci. USA77:780–784.
Serafini, T., T.E. Kennedy, M.J. Galko, C. Mirzayan, T.A. Jessell, and M.Tessier-Lavigne (1994) The netrins define a family of axon outgrowth-promoting proteins homologous to C. elegansUNC-6. Cell 78:409–424.
Thoenen, H. (1995) Neurotrophins and neuronal plasticity. Science 270:593–598.
Woods, D.F., and P.J. Bryant (1993) Apical junctions and cell signalling inepithelia. J. Cell Sci. Suppl. 17:171–181.
Yamada, T., M. Placzek, H. Tanaka, J. Dodd, and T.M. Jessell (1991) Controlof cell pattern in the developing nervous system: Polarizing activity ofthe floor plate and notocord. Cell 64:635–647.
448 E.J. RAMCHARAN AND R.W. GUILLERY