making sense of dlx1 antisense rna - core · making sense of dlx1 antisense rna petra kraus a , v...
TRANSCRIPT

Developmental Biology 376 (2013) 224–235
Contents lists available at SciVerse ScienceDirect
Developmental Biology
0012-16
http://d
n Corr
E-m
journal homepage: www.elsevier.com/locate/developmentalbiology
Genomes and Developmental Control
Making sense of Dlx1 antisense RNA
Petra Kraus a, V Sivakamasundari a, Siew Lan Lim a, Xing Xing a, Leonard Lipovich b, Thomas Lufkin a,n
a Stem Cell and Developmental Biology, Genome Institute of Singapore, Singapore 138672, Singaporeb Center for Molecular Medicine & Genetics and the Department of Neurology, Wayne State University School of Medicine, Detroit, MI, USA
a r t i c l e i n f o
Article history:
Received 17 August 2012
Received in revised form
27 January 2013
Accepted 29 January 2013Available online 8 February 2013
Keywords:
Long non-coding RNA
Dlx
Gene-targeting
Antisense RNA
Natural antisense transcripts
NAT
06/$ - see front matter & 2013 Elsevier Inc. A
x.doi.org/10.1016/j.ydbio.2013.01.035
esponding author. Fax: þ65 6808 8307.
ail address: [email protected] (T. Lufkin
a b s t r a c t
Long non-coding RNAs (lncRNAs) have been recently recognized as a major class of regulators in
mammalian systems. LncRNAs function by diverse and heterogeneous mechanisms in gene regulation,
and are key contributors to development, neurological disorders, and cancer. This emerging importance
of lncRNAs, along with recent reports of a functional lncRNA encoded by the mouse Dlx5-Dlx6 locus, led
us to interrogate the biological significance of another distal-less antisense lncRNA, the previously
uncharacterized Dlx1 antisense (Dlx1as) transcript. We have functionally ablated this antisense RNA via
a highly customized gene targeting approach in vivo. Mice devoid of Dlx1as RNA are viable and fertile,
and display a mild skeletal and neurological phenotype reminiscent of a Dlx1 gain-of function
phenotype, suggesting a role for this non-coding antisense RNA in modulating Dlx1 transcript levels
and stability. The reciprocal relationship between Dlx1as and Dlx1 places this sense–antisense pair into
a growing class of mammalian lncRNA-mRNA pairs characterized by inverse regulation.
& 2013 Elsevier Inc. All rights reserved.
Introduction
Increasing evidence points to a role for non-coding RNAs (ncRNAs)in development and disease, a field rapidly gaining importance(Babajko et al., 2009; Berdal et al., 2002; Blin-Wakkach et al., 2001;Brockdorff et al., 1991; Chen and Carmichael, 2010; Clemson et al.,1996; Costa, 2010; Eberhart et al., 2008; Faghihi and Wahlestedt,2009; Feng et al., 2006; Ginger et al., 2006; Gupta et al., 2010;Lipovich et al., 2010; Lyle et al., 2000; Nagano et al., 2008; Qureshiet al., 2010; Rinn et al., 2007; Sana et al., 2012; St Laurent et al., 2009;Wang and Chang, 2011; Wang et al., 2011; Young et al., 2005).NcRNAs include microRNAs (miRNAs) and long non-coding RNAs(lncRNAs), with an emerging class of lncRNAs that act as endogenousriboregulators of transcription factors (TFs), including lncRNAs thatdirectly bind TF proteins to regulate them (Chen and Carmichael,2010; Costa, 2010; Hung and Chang, 2010; Hung et al., 2011; Kinoet al., 2010; Lipovich et al., 2010; Martianov et al., 2007; Redon et al.,2010; Sana et al., 2012; Wang and Chang, 2011). Intriguing examplesof these regulators include the lncRNA GAS5, a ribo-mimic of thegenomic-DNA glucocorticoid receptor binding site and hence a directsuppressor of endogenous glucocorticoid receptor binding to genomictargets (Kino et al., 2010), and SRA, whose lncRNA isoform activatesestrogen receptor alpha through a shared ribonucleoprotein complex(Chooniedass-Kothari et al., 2010). Amongst the lncRNAs we find alarge group of endogenous antisense RNAs (also known as natural
ll rights reserved.
).
antisense transcripts, NATs), transcribed from complex genetic loci onthe opposite strand of protein-coding transcripts (Feng et al., 2006;Katayama et al., 2005; Rinn et al., 2007; Sleutels et al., 2002; Thakuret al., 2004). LncRNA NATs are frequently related to TF-encodinggenes (Werner, 2005). Several mechanisms of these RNAs have beensuggested (Vanhee-Brossollet and Vaquero, 1998). NATs, includinglncRNAs, may overlap exons and/or introns of their associated sensetranscripts, and can mediate or regulate gene expression in cis and/orin trans (for reviews see (Costa, 2010; Lipovich et al., 2010; Sana et al.,2012; Wang and Chang, 2011). Recently, a connection between NATsand potential pathological changes in the CNS has been establishedfor Bdnf mRNA and Bdnf-as lncRNA by employing a siRNA knockdownstrategy (Modarresi et al., 2012).
While Dlx1 antisense (Dlx1as) RNA has been described pre-viously (Dinger et al., 2008; Jeong et al., 2008; Liu et al., 1997;McGuinness et al., 1996) to date it remains to be defined whetherthe naturally occurring Dlx1as transcript has any function orshould be considered as mere transcriptional noise. The genomicregion encoding the Dlx1as RNA is embedded between two well-characterized genes: Dlx1 and Dlx2 (McGuinness et al., 1996),homeobox-containing transcription factors that are part of distal-
less, an evolutionarily conserved gene family (Depew et al., 2005;Kraus and Lufkin, 1999; McGuinness et al., 1996). While theDlx homeoproteins are key transcriptional regulators of down-stream target genes with crucial roles in craniofacial and appen-dicular skeleton as well as neurological development (Cobos et al.,2005; Depew et al., 2005; Jeong et al., 2008; Jones et al., 2011;Mao et al., 2009; Petryniak et al., 2007; Robledo et al., 2002), andthe Evf2 lncRNA, a NAT of the Dlx5-Dlx6 locus, has been identified

P. Kraus et al. / Developmental Biology 376 (2013) 224–235 225
as an in-trans regulator of Dlx2 expression (Bond et al., 2009;Feng et al., 2006), little is known about the Dlx1as RNA (Dingeret al., 2008; Liu et al., 1997; McGuinness et al., 1996). Recentfindings regarding a role of Dlx1 in behavioral abnormalities inmice such as hyperactivity, reduced fear response and epilepsy(Cobos et al., 2005; Jones et al., 2011; Mao et al., 2009; Petryniaket al., 2007) alongside implications of lncRNAs (reviewed in(Qureshi et al., 2010)) and miRNAs (reviewed in (Meza-Sosaet al., 2012)) in the development of the central nervous system(CNS) and their potential role as causal agents or modulator indifferent types of neurodegenerative diseases, led us to investi-gate whether the Dlx1as RNA had a function.
Mice devoid of Dlx1 die within 1 month after birth whenmaintained on a C57BL/6 genetic background (Qiu et al., 1997;Wonders and Anderson, 2005). In outbred mice, expression ofDlx1 is crucial for the survival of somatostatin (somþ) andneuropeptideY (NPYþ) producing GABAeric interneurons thatundergo increased apoptotic cell death in adult Dlx1 loss-of-function mice (Cobos et al., 2005). The observed behavioralphenotypes in these mice have been attributed to this loss ofsubtype-specific interneurons as a consequence of the introduc-tion of a gene targeted 2.8 kb deletion in the Dlx1 locus, removingall of exons 2 and 3 (Cobos et al., 2005; Qiu et al., 1997). Such adeletion inadvertently would have also affected if not deleted thegenomic region encoding the Dlx1as RNA (McGuinness et al.,1996). Exons of the Dlx1 gene, including specifically a region thatencodes the homeobox domain portion highly conserved at theamino acid level relative to Dlx2, are overlapped in the oppositedirection by exons of the Dlx1as RNA. Here, to investigate anypossible function of the Dlx1as RNA, we truncated the Dlx1as RNAusing gene targeting to abolish Dlx1as expression and hence anypotential hybridization between the Dlx1as and Dlx1 sensetranscripts, providing an in vivo analysis of an endogenousantisense lncRNA by a gene-targeted, loss-of-function approach.To date our study is the second to target an lncRNA that regulatesdistal-less homeobox genes (Bond et al., 2009). Our gene targetingwas carefully customized based on bioinformatic evidence toensure a truncation of the Dlx1as RNA upstream of the sequenceallowing for potential hybridization while leaving the surround-ing Dlx1 and Dlx2 locus unaffected. The mice homozygous for thetargeted allele are viable and fertile with mild skeletal andneurological phenotypes demonstrating the biological relevanceof the Dlx1as RNA as a modulator of the Dlx1 transcript levels.Functionally ablating the Dlx1as RNA essentially replicates a Dlx1gain-of-function phenotype. We therefore suggest that the non-coding Dlx1as RNA has an endogenous role as a negative regulatorof Dlx1 mRNA and may fine-tune Dlx1 transcript availability.
Material and methods
Construct design and generation of Dlx1as4xPA/4xPA mice
A 13149 bp long homology arm (LHA) and a 1204 bp shorthomology arm (SHA) were used to target the endogenous Dlx1as
locus by homologous recombination in R1 ES cells. A loxPpgkGb2Neo-
loxP selection cassette followed by four polyadenylation sites (4xPA)
between Exon 1 and Exon 2 of the sequence encoding for the Dlx1as
RNA or 30 to Exon 3 of the Dlx1 gene respectively, precisely by deleting160 bp between position 102651 and 102811 of AL928931 (seeFig. 1). This design will leave the Dlx1 sense coding region as wellas known conserved enhancer elements intact (Ghanem et al., 2003;McGuinness et al., 1996).
Dlx1as RNA validation by RT–PCR and quantification of embryonic
gene expression levels in Dlx1as4xPA/4xPA mice
Total RNA was isolated from fresh brains of P28 adult mice orfrom fresh E13.5 or formalin-fixed and paraffin embedded (FFPE)E13.5 embryos using either TRIzol reagent (Invitrogen) or HighPure FFPE RNA Kit (Roche) according to manufacturer’s instruc-tions. After the final RNA precipitation step, RNA samples fromfresh tissue were treated with TurboTM DNase (Ambion) before thefinal purification by passing through RNeasy columns (QIAGEN).Yield and quality of RNA was determined with a NanoDropspectrometer (NanoDrop Technology). First strand cDNA wassynthesized from 5 mg (fresh) to 0.5 mg (FFPE) of total RNA byreverse transcription (RT) PCR at 50 1C for 30 min in the presenceof 200 ng/ml random hexamers and 10 mM each of dNTPs andRevertAidTM Premium Enzyme mix (Fermentas). cDNA sampleswere diluted in DNAse-free water 1:5 prior to reverse transcrip-tion (RT)–PCR or 1:10 prior to real-time PCR (qPCR). RT–PCR andqPCR were performed to confirm Dlx1as RNA expression wasabrogated in Dlx1as4xPA/4xPA mice. cDNA reverse transcribed frommRNA isolated from Dlx1as4xPA/4xPA and Dlx1asþ /þ mice wereamplified by PCR using a primer pair designed to amplify theDlx1as transcript. The quality of cDNA samples was assessed bytheir amplification in the presence or absence of reverse tran-scriptase to confirm that the absence of the Dlx1as RT–PCRproduct in Dlx1as4xPA/4xPA mice was not an artificial result ofdeterioration of RNA samples or DNA contaminiation. RT-PCRreactions were carried out in total volumes of 50 ml and included400 nM of each primer and 5 ml of diluted cDNA template contain-ing 250 ng cDNA. PCR thermocycling conditions for the Dlx1as
transcript were 93 1C for 3 min, 93 1C for 15 s, 51 1C for 30 s, 68 1Cfor 1 min 30 s; for 35 cycles using LongRange PCR enzyme mix(Qiagen). B2m was amplified at 93 1C for 3 min, 93 1C for 15 s,58 1C for 30 s, 68 1C for 1 min; for 35 cycles using LongRange PCRenzyme mix. Gel electrophoresis analysis of the RT–PCR productsindicated a nucleic acid band of 1184 bp, the size expected for theDlx1as cDNA PCR product. Control reactions without reversetranscriptase produced no such DNA fragment, indicating theobserved RT–PCR product was not a result of amplification ofcontaminating genomic DNA. For qPCR the transcript of either thehousekeeping gene b2-microglobulin (B2m), Hprt or b-Actin wasused as a calibrator. For qPCR validation of expression levels ofDlx1, Dlx2, Olig2, Mash1 and Gad67 total RNA was extracted fromheads of three biological replicates of Dlx1as4xPA/4xPA and Dlx1asþ /þ
E13.5 FFPE embryos each. Samples were run as technical triplicates.qPCR was carried out in a volume of 25 ml on a ABI7500 thermo-cycler using SYBRgreen (Invitrogen) with primers as listed in thetable below. For statistical significance a T-test three analysis withone tail was performed in Excel.
Transcript
Primer name Primer sequence ReferenceDlx1as–Exon1
Dlx1as-qPCR-F4 50-ttgctacatcaaatcaaaagggtcg-30 n/a Dlx1as-qPCR-R4 50-ggacctctttcctccactctgttct-30Dlx1as–Exon2
Dlx1as-qPCR-F3 50-atttattccagtttgcagttgcagg-30 n/a Dlx1as-qPCR-R3 50-gactgtgacacctagaaccagaggc-30
P. Kraus et al. / Developmental Biology 376 (2013) 224–235226
Dlx1as–Exon2
Dlx1asE2-SYBRF3 50-ccgagctagggcaaacagtt-30 n/a Dlx1asE2-SYBRR3 5’-gcctctggttctaggtgtcacagt-30Dlx1as
Dlx1as1184-F 50-ttttcttttgctttcctgtc-30 n/a Dlx1as1184-R 50-tctaaaagttgccaacagcg-30Dlx1
Dlx1-RT-F 50-agagaggaccaatgagcctt-30 (Mu et al., 2004) Dlx1-RT-R 50-cacggtggatttcaatcggt-30Dlx2
Dlx2-RT-F 50-cttcatccattgccagtgga-30 (Mu et al., 2004) Dlx2-RT-R 50-aaacatagggactgctgagg-30Olig2
Olig2-RT-F 50-ctggtgtctagtcgcccatc-30 (Stecca and Ruiz i Altaba, 2009) Olig2-RT-R 50-aggaggtgctggaggaagat-30Mash1
Mash1-RT-F2 50-tctcctgggaatggactttg -30 n/a Mash1-RT-R2 50-ggttggctgtctggtttgtt-30Gad67
Gad67-RT-F 50-agatagccctgagcgacgag-30 (Sinclair et al., 2010) Gad67-RT-R 50-atggccgatgattctggttc-30B2M
B2M-F 50-ggagaatgggaagccgaaca-30 n/a B2M-R 50-gcattgggcacagtgacaga-30Hprt
qPcr-Hprt-F 50-gcagtacagccccaaaatgg-30 (Lengacher et al., 2004) qPcr-Hprt-R 50-aacaaagtctggcctgtatccaa-30bActin
Actb-RT-F 50-caccctgtgctgctcacc-30 (Sinclair et al., 2010)Actb-RT-R
50-gcacgatttccctctcag-30Skull morphology assessment by alizarin red and alcian blue staining
Dlx1as4xPA/þ animals were intermated to generate wild typeand homozygous P0 offspring. The P0 neonates were decapitatedfollowing strict IACUC guidelines for euthanasia and genotyped.After overnight fixation in 100% EtOH the skull was dissected freeof all other tissue types and subjected to alcian blue stainingrepresenting chondrogenic tissue and alizarin red staining char-acteristic for mineralized tissue as described in (Kraus et al., 2001).
Histology
P27 to P30 mice were deeply anesthetized with Avertin(0.2 mg/10 g body weight) and perfused with cold 4% PFA inDEPC/PBS and the brains carefully removed from the skull.Embryos were removed from the uterus of timed pregnant femalesat E12.5, E13.5 and decapitated at E15.5, then washed in 1� PBS.All samples were fixed in an excess volume of 4% PFA in DEPC/PBSat 4 1C under agitation over night, processed via an automatictissue processor (Leica) for paraffin embedding and sectioned at10 um. In situ hybridization was essentially carried out asdescribed in (Kraus et al., 2012) using the following probes: Dlx1(IMAGE clone: 30360273) was linearized with EcoRI and tran-scribed from T3 to generate the antisense probe and linearizedwith NotI and transcribed from T7 for the sense probe. The senseprobe was used to detect Dlx1as expression. Mash1/Ascl1 (IMAGEclone: 6415061), Olig2 (IMAGE clone: 5695592) and Somatostatin
(IMAGE clone: 476246) were linearized with EcoRI and transcribedfrom T3, Lhx6 (a donation from Vassilis Pachnis) was linearizedwith NotI and transcribed from T3, while Gad67 (IMAGE clone:5358787) was linearized with SalI and transcribed from T7 togenerate the antisense probe using the DIG RNA labeling kit(Roche). It is of importance to note that SISH to detect theexpression of Gad67, Somatostatin, Olig2, Mash1, Lhx6, Dlx1 senseand Dlx1 antisense for several replicates of each genotype withinan age group have been carried out within the same experiment toallow for comparability. For cell-counts comparable size-definedareas of horizontal/coronal sections through the hippocampus ofadult brains were selected and neurons positive for a given probewithin this area were accounted for manually similar to what hasbeen described by Petryniak et al. (2007). The number of neuronsper section was averaged for the sections analyzed.
Ethics statement, mouse husbandry and tissue collection
All animal procedures were performed according to the Singa-pore AnSTAR Biological Resource Center (BRC) Institutional Ani-mal Care and Use Committee (IACUC) guidelines and the IACUCprotocols employed were reviewed and approved by the afore-mentioned committee before any animal procedures were under-taken for this study described here (IACUC Protocols 080348/110689 and 080377).
Results
High sequence conservation is observed not only between theorthologous human and mouse Dlx1 proteins, but also between theparalogous, tandemly encoded murine Dlx1 and Dlx2 proteins(McGuinness et al., 1996). The Dlx1as NAT lncRNA is hence tran-scribed from an evolutionarily conserved region. Furthermore, thefirst-intron GT splice donor of Dlx1as is highly conserved throughoutplacental mammals, marsupials, and monotremes, according to the30-species MultiZ alignment in the UCSC Genome Browser. The firstantisense intron’s splice acceptor, located inside the Dlx1 30UTR(Fig. 1) is conserved in eutherian mammals, as are the splice donorsand acceptors of all additional introns located upstream of Dlx1along the genome, based on the four Genbank accession numbers(AK132348, BF469576, AK139461, and BB647803). Interestingly,despite the sequence conservation, the gene structure landscape ofthe human DLX1 locus is less well-conserved relative to mouse:there is no evidence of spliced human cDNAs or ESTs equivalent toDlx1as, although this might be merely an outcome of the limitednature of human cDNA libraries available, compared to the exten-sive FANTOM3 mouse libraries. While there have been reports offunctional NAT lncRNAs in mouse (for review see (Faghihi andWahlestedt, 2009)) in this study, using a highly customized genetargeting approach, we for the first, time demonstrate a Dlx1as loss-of-function phenotype, making ‘‘sense’’ of the existence of the Dlx1antisense RNA.
Effective targeting by homologous recombination of the Dlx1as RNA
without disruption of the Dlx1 sense transcript.
We have generated, characterized and validated a functionalablation of a NAT lncRNA by gene targeting. To generate this allele
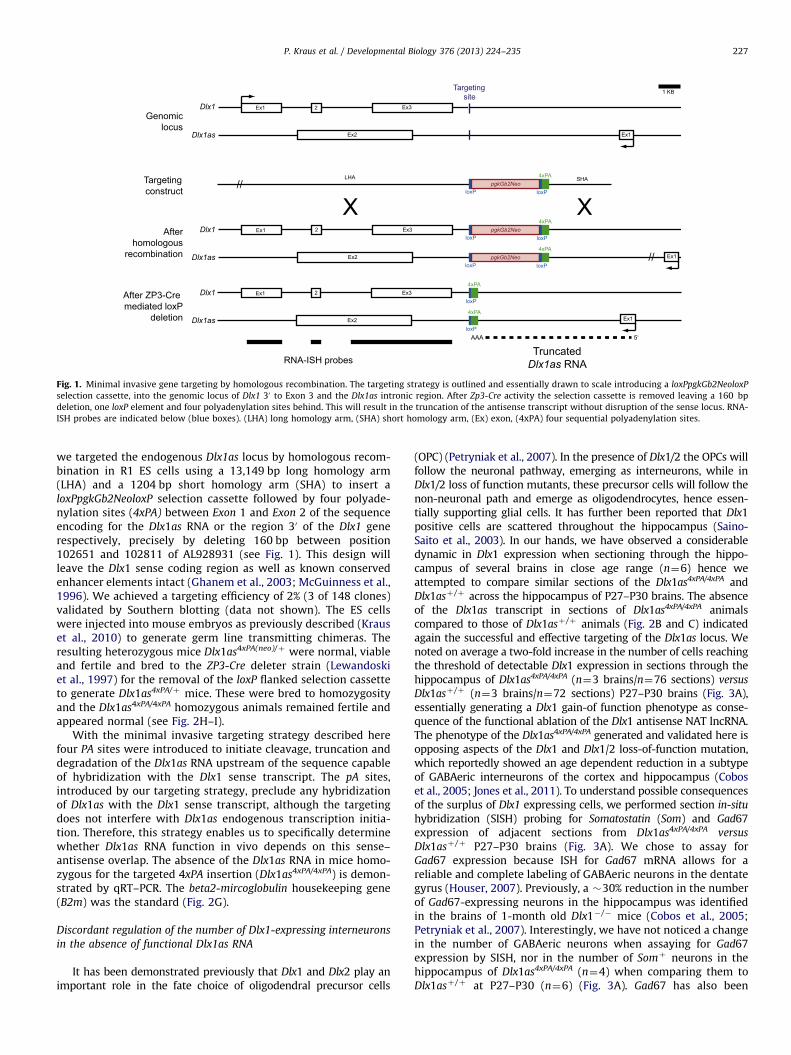
Fig. 1. Minimal invasive gene targeting by homologous recombination. The targeting strategy is outlined and essentially drawn to scale introducing a loxPpgkGb2NeoloxP
selection cassette, into the genomic locus of Dlx1 30 to Exon 3 and the Dlx1as intronic region. After Zp3-Cre activity the selection cassette is removed leaving a 160 bp
deletion, one loxP element and four polyadenylation sites behind. This will result in the truncation of the antisense transcript without disruption of the sense locus. RNA-
ISH probes are indicated below (blue boxes). (LHA) long homology arm, (SHA) short homology arm, (Ex) exon, (4xPA) four sequential polyadenylation sites.
P. Kraus et al. / Developmental Biology 376 (2013) 224–235 227
we targeted the endogenous Dlx1as locus by homologous recom-bination in R1 ES cells using a 13,149 bp long homology arm(LHA) and a 1204 bp short homology arm (SHA) to insert aloxPpgkGb2NeoloxP selection cassette followed by four polyade-nylation sites (4xPA) between Exon 1 and Exon 2 of the sequenceencoding for the Dlx1as RNA or the region 30 of the Dlx1 generespectively, precisely by deleting 160 bp between position102651 and 102811 of AL928931 (see Fig. 1). This design willleave the Dlx1 sense coding region as well as known conservedenhancer elements intact (Ghanem et al., 2003; McGuinness et al.,1996). We achieved a targeting efficiency of 2% (3 of 148 clones)validated by Southern blotting (data not shown). The ES cellswere injected into mouse embryos as previously described (Krauset al., 2010) to generate germ line transmitting chimeras. Theresulting heterozygous mice Dlx1as4xPA(neo)/þ were normal, viableand fertile and bred to the ZP3-Cre deleter strain (Lewandoskiet al., 1997) for the removal of the loxP flanked selection cassetteto generate Dlx1as4xPA/þ mice. These were bred to homozygosityand the Dlx1as4xPA/4xPA homozygous animals remained fertile andappeared normal (see Fig. 2H–I).
With the minimal invasive targeting strategy described herefour PA sites were introduced to initiate cleavage, truncation anddegradation of the Dlx1as RNA upstream of the sequence capableof hybridization with the Dlx1 sense transcript. The pA sites,introduced by our targeting strategy, preclude any hybridizationof Dlx1as with the Dlx1 sense transcript, although the targetingdoes not interfere with Dlx1as endogenous transcription initia-tion. Therefore, this strategy enables us to specifically determinewhether Dlx1as RNA function in vivo depends on this sense–antisense overlap. The absence of the Dlx1as RNA in mice homo-zygous for the targeted 4xPA insertion (Dlx1as4xPA/4xPA) is demon-strated by qRT–PCR. The beta2-mircoglobulin housekeeping gene(B2m) was the standard (Fig. 2G).
Discordant regulation of the number of Dlx1-expressing interneurons
in the absence of functional Dlx1as RNA
It has been demonstrated previously that Dlx1 and Dlx2 play animportant role in the fate choice of oligodendral precursor cells
(OPC) (Petryniak et al., 2007). In the presence of Dlx1/2 the OPCs willfollow the neuronal pathway, emerging as interneurons, while inDlx1/2 loss of function mutants, these precursor cells will follow thenon-neuronal path and emerge as oligodendrocytes, hence essen-tially supporting glial cells. It has further been reported that Dlx1positive cells are scattered throughout the hippocampus (Saino-Saito et al., 2003). In our hands, we have observed a considerabledynamic in Dlx1 expression when sectioning through the hippo-campus of several brains in close age range (n¼6) hence weattempted to compare similar sections of the Dlx1as4xPA/4xPA andDlx1asþ /þ across the hippocampus of P27–P30 brains. The absenceof the Dlx1as transcript in sections of Dlx1as4xPA/4xPA animalscompared to those of Dlx1asþ /þ animals (Fig. 2B and C) indicatedagain the successful and effective targeting of the Dlx1as locus. Wenoted on average a two-fold increase in the number of cells reachingthe threshold of detectable Dlx1 expression in sections through thehippocampus of Dlx1as4xPA/4xPA (n¼3 brains/n¼76 sections) versus
Dlx1asþ /þ (n¼3 brains/n¼72 sections) P27–P30 brains (Fig. 3A),essentially generating a Dlx1 gain-of function phenotype as conse-quence of the functional ablation of the Dlx1 antisense NAT lncRNA.The phenotype of the Dlx1as4xPA/4xPA generated and validated here isopposing aspects of the Dlx1 and Dlx1/2 loss-of-function mutation,which reportedly showed an age dependent reduction in a subtypeof GABAeric interneurons of the cortex and hippocampus (Coboset al., 2005; Jones et al., 2011). To understand possible consequencesof the surplus of Dlx1 expressing cells, we performed section in-situ
hybridization (SISH) probing for Somatostatin (Som) and Gad67expression of adjacent sections from Dlx1as4xPA/4xPA versus
Dlx1asþ /þ P27–P30 brains (Fig. 3A). We chose to assay forGad67 expression because ISH for Gad67 mRNA allows for areliable and complete labeling of GABAeric neurons in the dentategyrus (Houser, 2007). Previously, a �30% reduction in the numberof Gad67-expressing neurons in the hippocampus was identifiedin the brains of 1-month old Dlx1� /� mice (Cobos et al., 2005;Petryniak et al., 2007). Interestingly, we have not noticed a changein the number of GABAeric neurons when assaying for Gad67expression by SISH, nor in the number of Somþ neurons in thehippocampus of Dlx1as4xPA/4xPA (n¼4) when comparing them toDlx1asþ /þ at P27–P30 (n¼6) (Fig. 3A). Gad67 has also been

Fig. 2. The absence of the Dlx1as transcript is demonstrated by SISH using the sense probe for Dlx1, detecting cells expressing the Dlx1as transcript in the hippocampus of
Dlx1asþ /þ P30 animals in an overview (A) and close ups (B,C), while the Dlx1as transcript is absent in corresponding regions of Dlx1as4xPA/4xPA age matched controls (D–F).
The successful targeting of the Dlx1as locus is further demonstrated by RT-PCR (G) assaying for the presence of the Dlx1as transcript using RNA extracted from P30 brain
samples for Dlx1asþ /þ (lanes 1 and 5) and Dlx1as4xPA/4xPA (lanes 3 and 7) in the presence (lanes 1, 3, 5 and 7) and absence (lanes 2, 4, 6 and 8) of reverse transcriptase using
the presence of the ß2-microglobulin (B2m) transcript as a control. We further demonstrate that the Dlx1as4xPA/4xPA mice are fertile (H) and of overtly normal appearance (I).
Relative expression by qPCR of Dlx1as Exon 1 (primer pair qPCRF4/R4) upstream and Dlx1as Exon 2 (primer pairs qPCRF4/R4 and E2SYBRF3/R3) downstream of the 4xPA
insertion site, shown for RNA extracted from two Dlx1as4xPA/4xPA (#270, #275) and one Dlx1asþ /þ E13.5 embryos.
P. Kraus et al. / Developmental Biology 376 (2013) 224–235228

Fig. 3. Neurochemical changes in the ganglionic eminences in the absence of Dlx1as
transcript without a detectable impact on the adult GABAeric interneuron number
is demonstrated by SISH on coronal/horizontal sections (A). Dlx1as expression is
abolished while Dlx1 expression is increased in Dlx1as4xPA/4xPA embryos at E12.5,
E13.5 and E15.5 compared to age and region matched Dlx1asþ /þ controls. Serial
sections of Dlx1, Olig2 and Mash1 expression are analyzed at E12.5 and E15.5. While
Dlx1 expression is elevated at both stages in Dlx1as4xPA/4xPA embryos compared to
the control, Olig2 and Mash1 appear unchanged at E12.5, but by E15.5 Mash1
expression appears more diffuse in the VZ and SVZ of Dlx1as4xPA/4xPA embryos. While
in the absence of functional Dlx1as RNA the number of Dlx1 expressing cells remains
about two-fold higher in the hippocampus of Dlx1as4xPA/4xPA adults compared to age
and region matched controls, the number of GABAeric neurons assayed for by Gad67
and Somatostatin expression remains unchanged. In embryos however, reduced
levels of Gad67 expression can be observed in Dlx1as4xPA/4xPA embryos compared to
Dlx1asþ /þ controls by E15.5. Abbreviations: (LV) lateral ventricle, (VZ) ventricular
zone, (SVZ) subventricular zone, (T) thalamic region, (E) eye, (MGE) medial gang-
lionic eminence, (LGE) lateral ganglionic eminence. Relative quantification of
expression levels for Dlx1, Dlx2, Olig2, Mash1 and Gad67 at E13.5 by qPCR validation
of total RNA form Dlx1as4xPA/4xPA FFPE heads (4xPA/4xPA) compared to Dlx1asþ /þ
FFPE heads (þ/þ). One-tailed Student’s test 3,** po0.005 (B). (For interpretation of
the references to color in this figure legend, the reader is referred to the web version
of this article.)
P. Kraus et al. / Developmental Biology 376 (2013) 224–235 229
identified as a trans-target of Evf2, the lncRNA transcribed fromthe Dlx5/Dlx6 locus paralogous to the Dlx1/Dlx2 locus, yet in theadult brain Gad67 expression levels are described as normal, yetreduced in the embryonic brain in the absence of Evf2 (Bond et al.,2009) as well as in the Dlx1/2 loss-of-function mutation (Petryniaket al., 2007). Analyzing Gad67 expression in the embryonic brainof Dlx1as4xPA/4xPA versus Dlx1asþ /þ by SISH at E12.5 (n¼3) andE15.5 (n¼4) Gad67 expression appeared reduced in Dlx1as4xPA/4xPA
embryos (Fig. 3) when comparing matching sections for bothgenotypes by SISH (Fig. 3A), yet no significant difference in Gad67expression levels was detected by qPCR using RNA extracted fromentire heads (n¼18, Fig. 3B). An additional interneuron markerLhx6 was analyzed by SISH and showed clearly reduced expressionby E15.5 in Dlx1as4xPA/4xPA embryos (Fig. 3A). This finding attests tosimilar mechanism for two different NAT lncRNAs, Evf2 (the NATlncRNA of the Dlx5/Dlx6 locus) and Dlx1as (the NAT lncRNA of theDlx1 locus) targeting two different Dlx family members, bothpossibly affecting early, but not late interneuron development.
Does Dlx1as RNA impact on oligodendral precursor cell fate decision?
In the mouse forebrain, the first of three rounds of progenitorcells are restricted to the oligodendroglial lineage. These oligo-dendral precursor cells (OPC) can be found by E13.5 (Kessariset al., 2006; Nery et al., 2001; Parras et al., 2007; Petryniak et al.,2007; Pringle and Richardson, 1993; Tekki-Kessaris et al., 2001).While the formerly debated origin of OPCs appears to have beenestablished, the complex molecular mechanisms underlying thesubsequent neuronal–glial fate decision are still to be determined(Kessaris et al., 2006; Petryniak et al., 2007; Richardson et al.,2006). Mash1 has been demonstrated to be involved in neurogen-esis and oligodendrogenesis (Casarosa et al., 1999; Parras et al.,2007), Olig2 in oligodendrogenesis (Miyoshi et al., 2007; Zhouet al., 2001) and Dlx1 and Dlx2 play a role in facilitating thedifferentiation of GABAeric neurons (Panganiban and Rubenstein,2002). Given our observation of a two-fold increase in cellsreaching the threshold for detectable Dlx1 sense expression inthe hippocampus of adult Dlx1as4xPA/4xPA mice, reflecting essen-tially a Dlx1gain-of-function phenotype, we aimed to understanda possible role of Dlx1as RNA in this complex neural/glial fatedecision process of OPCs. We analyzed Dlx1, Olig2 and Mash1expression at embryonic time points by SISH of sections throughthe developing brain. We have noted an increase in Dlx1 expres-sion in sections of Dlx1as4xPA/4xPA embryos at E12.5, E13.5 andE15.5, when comparing them to age matched Dlx1asþ /þ controls(Fig. 3A), a significant 1.4� increase in Dlx1 expression compar-ing Dlx1as4xPA/4xPA versus Dlx1asþ /þ was validated by qPCR forRNA isolated from entire heads at E13.5 (n¼18, p-value: 0.0047),while Dlx2 expression was not significantly affected in this group(Fig. 3B). Adjacent sections were analyzed for the expression ofOlig2 and Mash1, both genes being critically involved in OPC fatedecision (Petryniak et al., 2007). At E12.5, just a day after theestimated onset of oligodendrogenesis in the mouse forebrain(Kessaris et al., 2006) we did not observe a consistent differencein Olig2 or Mash1 expression between sections of Dlx1as4xPA/4xPA
and Dlx1asþ /þ embryos (n¼3 embryos/120 sections) by SISH(Fig. 3A). Quantification of expression levels by qPCR for RNAisolated from entire heads at E13.5 (n¼18) did not indicate anydifference in expression levels for Olig2 (p-value: 0.1824), yetsimilar to Dlx1, Mash1 expression was 1.4� upregulatedDlx1as4xPA/4xPA versus Dlx1asþ /þ embryos (n¼18, p-value:0.0002) (Fig. 3B). A few days later, by E15.5, at the time whenOPCs are thought to migrate into the cerebral cortex (Kessariset al., 2006), Mash1 expression appeared more broadly expressedin the ventricular and subventricular zone in embryos with
P. Kraus et al. / Developmental Biology 376 (2013) 224–235230
functionally ablated Dlx1as RNA, when analyzing matchingsections by SISH (n¼4 embryos/80 sections) (Fig. 3A).
A dysmorphic alicochlear commissure in the absence of
functional Dlx1as RNA
While mutants of the Dlx family are probably best known for theirneurological abnormalities, the Dlx genes are also crucial players indetermining skull morphology (Depew et al., 2005) hence we furtherinvestigated whether the functional ablation of the lnc Dlx1as RNAmight also influence skull development. The skull is a complexstructure largely built from branchial arch derivatives and migratingcranial neural crest cells to encapsulate and protect the vertebratebrain from external damage. Detailed studies involving chick-quailchimeras (Couly et al., 1993; Creuzet et al., 2005) have classified threemajor compartments of the avian skull based on their origin whichhave been adapted to the mouse skull anatomy (Depew et al., 2005;
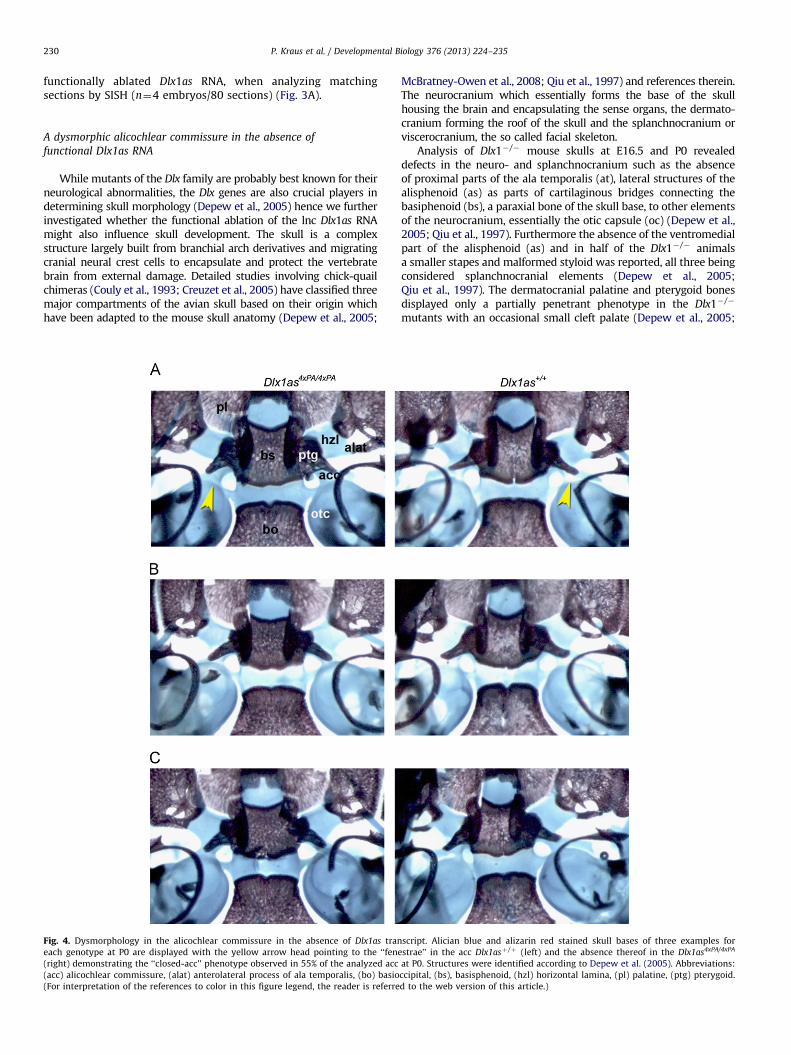
Fig. 4. Dysmorphology in the alicochlear commissure in the absence of Dlx1as tran
each genotype at P0 are displayed with the yellow arrow head pointing to the ‘‘fene
(right) demonstrating the ‘‘closed-acc’’ phenotype observed in 55% of the analyzed acc
(acc) alicochlear commissure, (alat) anterolateral process of ala temporalis, (bo) basio
(For interpretation of the references to color in this figure legend, the reader is referre
McBratney-Owen et al., 2008; Qiu et al., 1997) and references therein.The neurocranium which essentially forms the base of the skullhousing the brain and encapsulating the sense organs, the dermato-cranium forming the roof of the skull and the splanchnocranium orviscerocranium, the so called facial skeleton.
Analysis of Dlx1� /� mouse skulls at E16.5 and P0 revealeddefects in the neuro- and splanchnocranium such as the absenceof proximal parts of the ala temporalis (at), lateral structures of thealisphenoid (as) as parts of cartilaginous bridges connecting thebasiphenoid (bs), a paraxial bone of the skull base, to other elementsof the neurocranium, essentially the otic capsule (oc) (Depew et al.,2005; Qiu et al., 1997). Furthermore the absence of the ventromedialpart of the alisphenoid (as) and in half of the Dlx1� /� animalsa smaller stapes and malformed styloid was reported, all three beingconsidered splanchnocranial elements (Depew et al., 2005;Qiu et al., 1997). The dermatocranial palatine and pterygoid bonesdisplayed only a partially penetrant phenotype in the Dlx1� /�
mutants with an occasional small cleft palate (Depew et al., 2005;
script. Alician blue and alizarin red stained skull bases of three examples for
strae’’ in the acc Dlx1asþ /þ (left) and the absence thereof in the Dlx1as4xPA/4xPA
at P0. Structures were identified according to Depew et al. (2005). Abbreviations:
ccipital, (bs), basisphenoid, (hzl) horizontal lamina, (pl) palatine, (ptg) pterygoid.
d to the web version of this article.)

P. Kraus et al. / Developmental Biology 376 (2013) 224–235 231
Qiu et al., 1997). These same abnormalities and some exceedingthose have been described for Dlx2� /� and Dlx1� /�Dlx2� /� doublemutants (Depew et al., 2005; Qiu et al., 1997).
On the contrary, in the absence of functional Dlx1as RNA wehave never observed an obvious cleft palate nor any obviousgenotype specific changes in the ossicles when comparingDlx1as4xPA/4xPA versus Dlx1asþ /þ P0 and P1 skulls. However, wedid observe a dysmorphic alicochlear commissure (acc), whencomparing postnatal skulls of Dlx1as4xPA/4xPA and Dlx1asþ /þ agematched pups and E16.5 embryos (see Fig. 4 yellow arrowheads).The dysmorphology of the acc essentially showed itself in the lackof a gap observed where the acc bridges towards the oc. This‘‘closed-acc’’ morphology was observed in 55% of the analyzed acc(n¼60) of Dlx1as4xPA/4xPA skulls while it could only be seen in 14%of the analyzed acc (n¼94) of Dlx1asþ /þ skulls, an observationalso made at E16.5 for 36% of Dlx1as4xPA/4xPA acc (n¼14) but noneof the Dlx1asþ /þ acc (n¼20) analyzed (data not shown).
Discussion
With increasing knowledge of gene regulatory networks(GRNs), and increasing evidence pointing to the fact that GRNsare not restricted to TF target sets, it becomes evident that TFsthemselves might be fine-tuned and modulated post-transcriptionally by several classes of ncRNAs (Lipovich et al.,2010). Particularly in the CNS, ncRNAs seem to play an importantrole in directing cell fate of neural progenitor cells (NPCs) as wellas reactivity and plasticity of neural cells and their affiliated GRNsin response to external stimuli (for review see (Qureshi et al.,2010)). We are only beginning to understand possible mechan-isms of how these ncRNAs might act on their targets, for exampleby impacting on sense RNA stability via sense–antisense duplexformation, targeting the sense transcript for degradation (forreview see (Faghihi and Wahlestedt, 2009; Wang and Chang,2011)). In recent years the function of a few lncRNAs in generegulation has been demonstrated (Babajko et al., 2009; Blin-Wakkach et al., 2001; Feng et al., 2006; Lyle et al., 2000; Naganoet al., 2008; Sleutels et al., 2002).
While the actual levels of Dlx1 and or Dlx2 mRNA are likelycausal for any neuropathological or other morphological changesin Dlx1 and Dlx1/Dlx2 mutant mice, the targeting strategydescribed for these mutations would not allow for any functionalanalysis of the Dlx1as RNA, as the genomic region encoding theDlx1as RNA transcript would have also been deleted (Qiu et al.,1997). Naturally, the approach to target any NAT of a gene isdifficult, as it is extremely important not to disrupt the function ofthe gene itself so to avoid the misattribution of a phenotype asfunction of the lncRNA in question in its gene expressionmodulating function, development or pathogenesis. Severalapproaches have been taken to knockdown antisense RNA usingsiRNA technology (Janowski et al., 2007; Modarresi et al., 2012;Rapicavoli et al., 2011; Sheik Mohamed et al., 2010; Watts et al.,2010). Often though, the siRNA knockdown methodology suffersfrom off-target side effects, impacting on the interpretation of theresulting phenotype (Clark et al., 2008; Harborth et al., 2003;Scacheri et al., 2004; Sioud, 2011). Furthermore, given epigeneticand therefore nuclear functions of many lncRNAs, siRNA knock-downs are problematic because they employ the cytoplasmicRNA-Induced Silencing Complex (RISC) and hence do not com-pletely attenuate lncRNAs whose localization is partly or mostlynuclear, while commercial antisense oligonucleotide technologyfor epigenetic lncRNA targeting is not yet widely available. Henceit was our uttermost concern to avoid off-target effects and not todisrupt Dlx1, while rendering the Dlx1as RNA non-functional.Proof that we have succeeded lies in the fact that the Dlx1as
RNA can no longer be detected by either qPCR or RNA-ISH, whileDlx1 sense RNA is still present and expressed in the appropriatedomains. To date, via homologous recombination (Capecchi,1989), we have generated the first successful functional ablationof the Dlx1as RNA, a mouse NAT lncRNA, using classic gene-targeting technology.
The Dlx1as RNA is genomically encoded on the opposite strandof the Dlx1 gene. According to public cDNA and EST evidence inGenbank (e.g. AK132348, BF469576, AK139461, and BB647803),the lncRNA initiates from a transcription start region downstreamof the Dlx1 30end, and overlaps the entire extent of the Dlx1 gene,continuing well into the upstream neighbor gene of Dlx1, Metap1d
(Kawai et al., 2001). Since Metap1d encodes a mitochondrial-localized aminopeptidase, while Dlx1 has known functions in thebrain, we have made the assumption that our Dlx1as loss-of-function phenotype is due to the effect of Dlx1as on Dlx1.
Gamma-amino butyric acid (GABA) is the major neurotrans-mitter used by the highly diverse group of interneurons found inthe hippocampus (Houser, 2007), yet multiple subpopulations ofinterneurons can still be distinguished within this group ofGABAeric interneurons based on morphological and further neu-rochemical identities (Houser, 2007) with one of the largersubgroups being Somatostatin and/or Neuropeptide Y expressingneurons (Houser, 2007; Tallent, 2007) and references therein. Inadult Dlx1� /� mice a reduction in Somatostatin, Caltretinin andNeuropeptide Y producing GABAeric interneurons was reportedcausing seizures, hyperexitability and epilepsy when this muta-tion was maintained on a mixed genetic background (C57BL/6J:CD1) to allow for postnatal survival (Cobos et al., 2005; Joneset al., 2011).
Upon ablation of Dlx1as RNA in a mixed C57BL/6J:129SvS2genetic background, the homozygous animals were fertile and ofnormal appearance and survival rate. Mice with an Evf2 loss offunction have been equally described as fertile and morphologi-cally normal (Bond et al., 2009). When assaying for the expressionof Dlx1, Gad67 and Somatostatin by SISH, we found a two-foldincrease in the number interneurons reaching the detectablethreshold of Dlx1 in the hippocampus of Dlx1as4xPA/4xPA adultmice, essentially reflecting a Dlx1 gain-of-function phenotype.Similar to data previously generated in vitro, where in a gain-of-function assay Dlx2 and Dlx5 but not Dlx1 expression was capableof inducing the expression of glutamic acid decarboxylases(Stuhmer et al., 2002), we did not observe significant changes inthe number of Gad67 or Somatostatin expressing cells whencomparing Dlx1as4xPA/4xPA and Dlx1asþ /þ brains, suggesting a lessdramatic effect in the case of excess Dlx1 transcript compared tothe lack thereof. Hypothetically the observation by Stuhmer et al.(2002) could be interpreted as a titration effect of the Dlx1as RNAon excess Dlx1 sense transcript yet not excess transcript of theother Dlx family members delivered by the over-expressionvector in this gain-of-function experiment. Since surplus Dlx1
transcript delivered by a potent pCAGGS expression vector (Niwaet al., 1991) does not impact on the number of GABAeric neurons,the observed two-fold increase in cells reaching detectable levelsof Dlx1 RNA after functional ablation of the Dlx1as RNA is unlikelyto affect the number of GABAeric neurons, yet it might impact onother interneurons and subpopulations or different cell types, e.g.,cholinergic neurons or glia not accounted for in our essay(Furusho et al., 2006; Miyoshi et al., 2007; Ono et al., 2008). Itis of interest though; that Gad67 expression appears unaffected inthe adult hippocampus yet possibly regionally reduced inembryos after functional ablation of Dlx1as the NAT lncRNA ofthe Dlx1/Dlx2 cluster as well Evf2 the NAT lncRNA of the Dlx5/Dlx6
cluster two paralogous genomic regions in close proximity onmouse chromosome 2. It remains to be discovered to what extentthe apparently reciprocal regulatory modality (Dlx1as down,

Fig. 5. Functional ablation of the Dlx1as RNA transcript increases Dlx1 transcript
abundance mimicking a Dlx1 gain-of function phenotype. (A) Fate determination
of OPCs by controlling transcription levels of Dlx1, Mash1 and Olig2 has been
described previously in various loss-of function experiments (Petryniak et al.,
2007). (B) We believe that the Dlx1as RNA plays a role in modulating Dlx1
transcript availability by impacting on its life span. In a wild type situation, the
hypothetical hybridization between Dlx1 sense transcripts and Dlx1as RNA might
lead to decay of the sense transcript, hence regulating levels and availability of
this transcription factor. In the absence of Dlx1as RNA, the decay could be delayed.
(C) In this gain-of function context owing to a lack or delay of Dlx1 mRNA
degradation, Mash1 and Dlx1 transcript levels were found to be similarly
increased. Excess Dlx1 protein could possibly either direct or via additional
mechanisms impact on Mash1 transcription.
P. Kraus et al. / Developmental Biology 376 (2013) 224–235232
Dlx1þMash1 up) of the Dlx1 NATlncRNA appears to be differentfrom the more complex and synergistic regulatory impact of Evf2,an lncRNA capable of both increasing and decreasing the level ofits NAT partner, Dlx6 (Bond et al., 2009).
Homeodomain (HD) containing proteins, alongside proteins ofthe basic helix-loop-helix type (bHLH), are fundamental inorchestrating neural/glial development in the spinal cord andbrain (Kessaris et al., 2001; Ono et al., 2008). While our qPCRvalidation of Olig2 expression on RNA extracted from entire headsof E13.5 embryos did not show any significant difference betweenthe two genotypes, our SISH data on matching serial sections atE15.5 hints that the increase in Dlx1 expression in Dlx1as4xPA/4xPA
embryos could possibly have a suppressive effect on Olig2
expression during the differentiation phase of the first wave ofOPCs around the time the second wave of OPCs is generated(Kessaris et al., 2006). This observation would at least be theore-tically supported by previous data suggesting a negative regula-tion of Olig2 by Dlx1and Dlx2 given the increase in Olig2
expression in E15.5 Dlx1/Dlx2 mutants (Petryniak et al., 2007).However, increased Mash1 expression was described in the Dlx1/
Dlx2 mutants (Yun et al., 2002) which would also have deleted thegenomic sequence coding for the Dlx1as transcript. Whileexpanded expression of Dlx1 and Dlx2 was described in theabsence of Mash1 (Casarosa et al., 1999; Horton et al., 1999;Poitras et al., 2007), Mash1 levels have not been addressed in theearlier Dlx family gain-of function study in vitro (Stuhmer et al.,2002). Our qPCR validation shows a statistically significant 1.4�upregulation of Mash1 expression on RNA extracted from entireheads of Dlx1as4xPA/4xPA E13.5 embryos compared to age matchedDlx1þ /þ . We also observed a slightly broader area of Mash1
expression by SISH in the ventricular and subventricular zone ofDlx1as4xPA/4xPA embryos at both E12.5 and E15.5 when comparingmatching serial sections. A potential cross-regulation betweenMash1 and Dlx1 and Dlx2 has been described recently (Parraset al., 2007; Petryniak et al., 2007; Yun et al., 2002). We have notobserved a significant change in Dlx2 expression levels at E13.5 inthe absence of functional lnc Dlx1as RNA but we have noted asignificant increase in Dlx1 sense transcript levels both by qPCRvalidation and SISH. Interestingly in this Dlx1 gain-of-functionsituation where the lnc Dlx1as RNA that normally appears toregulate Dlx1 sense transcript availability, possibly by facilitatingits decay is not functional we also find Mash1 expression in theembryonic brain upregulated at a similar level as Dlx1 expressionitself. We hypothesize that increased Dlx1 sense transcript couldpotentially stimulate Mash1 expression, which itself may regulateDlx1 sense transcript availability via a feedback loop. However,given the fact that an increased Mash1 expression was describedin the Dlx1/Dlx2 mutants (Yun et al., 2002) in which based on theconstruct design the locus encoding the lnc Dlx1as RNA was alsodeleted, a direct supressive effect of this lnc RNA on Mash1
expression cannot be excluded.Despite a clear effect of the Dlx1as RNA on Mash1 and a
potential effect on later stage Olig2 expression, likely indirectvia modulating Dlx1 transcript stability, our observations andthose of Bond et al. (2009) that the number of GABAeric neuronsin the Dlx1as4xPA/4xPA and Evf2 loss-of-function adult hippocampusappears unaffected in the absence of functional Dlx1as RNA mightbe further explained by studies by Kessaris et al. (2006).Richardson et al. (2006) showing that in the brain some of theoriginal OPC populations gradually disappear after birth and areeventually replaced in the adult. Even more intriguing is anexperiment showing replacement of deliberately killed OPC bycell populations in the vicinity (Kessaris et al., 2006). Also,GABAeric neurons, despite being the major neuronal subtypegenerated by Olig2þ cells, can develop to normal density in anOlig2 loss-of function context (Ono et al., 2008). While Dlx1as RNA
might modulate the complex HD/bHLH interplay during embryo-nic neural/glial development, pathways controlling the numbersof GABAeric neurons in the adult brain can tolerate the two-foldincrease in Dlx1 expressing cells after its functional ablation.Effects on other neuronal subtypes and/or their supportive gliaremain to be investigated.
Aside from the neurological defects described for Dlx1and Dlx2
null animals, Depew et al. (2005) identified a greatly reducedalicochlear commissure (acc) as an additional phenotype to thedeletion of the proximal ala temporalis in approximately 50% of theDlx2� /� and in the Dlx1� /�/Dlx2� /� mutants (Liu et al., 1997; Qiuet al., 1997). Hence we further examined a potential role of the lncDlx1as RNA in skull development. We found in the Dlx1as4xPA/4xPA
mice similar anatomical structures affected as in the Dlx1and Dlx2
null animals, yet in a different way. Depew et al. (2005) describe agreatly reduced alicochlear commissure (acc) as an additionalphenotype to the deletion of the proximal ala temporalis inapproximately 50% of the Dlx2� /� and in the Dlx1� /�/Dlx2� /�
mutants (Liu et al., 1997; Qiu et al., 1997). In the mouse, the originof the basisphenoid bone, the hypophyseal cartilage appears justbelow the forming pituitary gland by late E13, with the lateralplaced basitrabecular and ala temporalis cartilages being estab-lished by E15 and the murine chondrocranium being fully formedby E16 (60). We have observed the appearance of a dysmorphic or‘‘closed’’ acc at E15.5 in 36% (n¼14) of the Dlx1as4xPA/4xPA genotype(n¼14) yet none (n¼20) of the Dlx1asþ /þ genotype alicochlearcommissures. This ‘‘closed’’ acc is somewhat opposing the reduced

P. Kraus et al. / Developmental Biology 376 (2013) 224–235 233
acc phenotype described for the Dlx2� /� and in the Dlx1� /�/Dlx2� /�
mutants. Interestingly, McBratney-Owen et al. (2008) mentioned thepresence of a number of fenestrae in the developing murine chon-drocranium that are not functioning as openings for nerves or bloodvessels and disappear with ongoing maturation. The ‘‘closed acc’’dysmorphology we frequently observed for the Dlx1as4xPA/4xPA ispossibly a pre-mature closure of one of those fenestrae in the absenceof functional Dlx1as RNA without any obvious survival consequencesfor the animal itself.
In summary, we have proven that Dlx1as RNA is functional, byusing an in vivo gene-targeting approach. We have demonstrated askeletal phenotype in the skull base as well as a neurological effectafter the functional ablation of the Dlx1as RNA. Both phenotypescould possibly be attributed to our abrogation of the negativeregulation of the Dlx1 sense transcript by the Dlx1as RNA transcript.The precise mechanism remains yet to be discovered: in particular,it is not clear whether the regulation is nuclear or cytoplasmic, andwhether it is mediated by epigenetic mechanisms (e.g. repressivehistone modifications as a result of Dlx1as-mediated cis-recruitmentof Prc2 to the Dlx1 locus) or post-transcriptional events (e.g. sense–antisense hybridization of the two mature, capped, polyadenylatedRNAs, that lessens the half-life or stability of the Dlx1 mRNA),respectively. There is evidence for Evf2 utilizing both mechanisms asa NAT of Dlx6 (Bond et al., 2009). Future studies in this field shouldaddress both the subcellular localization of Dlx1as and its possibleinteractions with RNA-binding proteins. By adding a new piece tothe puzzle of oligodendral/neural fate decision we conclude thatDlx1as RNA has a function in fine-tuning the accumulation of Dlx1
transcripts and either directly or indirectly impacting on Mash1
transcript levels (Fig. 5).
Author contributions
All authors contributed to drafting the manuscript. PK, LL andTL were involved in the design of the study. LL carried outcomputational analysis and advised on the field of NATlncRNAs.PK interpreted the data, carried out most animal work andgenerated histology and qPCR data. VS and PK carried out SISH.SLL carried out the validation of the targeting event, RT–PCR andqPCR. SLL and XX assisted with genotyping. All authors have readand approved the final manuscript.
Acknowledgements
We are grateful to Yang Sun Chan, Thien Peiling, Geraldine Naiand Christopher Wong for their expertise and help with mRNAextraction from FFPE material and all members of the Lufkin Labin particular Jie Wei Goh and Song Jie as well as BRC AnStar staff,Valerie Tan, Geraldine Leong and Hsiao Yun Chan for support withthe animal work.
References
Babajko, S., Petit, S., Fernandes, I., Meary, F., LeBihan, J., Pibouin, L., Berdal, A., 2009.Msx1 expression regulation by its own antisense RNA: consequence on toothdevelopment and bone regeneration. Cells Tissues Organs 189, 115–121.
Berdal, A., Lezot, F., Pibouin, L., Hotton, D., Ghoul-Mazgar, S., Teillaud, C., Robert, B.,MacDougall, M., Blin, C., 2002. Msx1 homeogene antisense mRNA in mousedental and bone cells. Connect Tissue Res. 43, 148–152.
Blin-Wakkach, C., Lezot, F., Ghoul-Mazgar, S., Hotton, D., Monteiro, S., Teillaud, C.,Pibouin, L., Orestes-Cardoso, S., Papagerakis, P., Macdougall, M., Robert, B.,Berdal, A., 2001. Endogenous Msx1 antisense transcript: in vivo and in vitroevidences, structure, and potential involvement in skeleton development inmammals. Proc. Nat. Acad. Sci. U.S.A. 98, 7336–7341.
Bond, A.M., Vangompel, M.J., Sametsky, E.A., Clark, M.F., Savage, J.C., Disterhoft, J.F.,Kohtz, J.D., 2009. Balanced gene regulation by an embryonic brain ncRNA iscritical for adult hippocampal GABA circuitry. Nat. Neurosci 12, 1020–1027.
Brockdorff, N., Ashworth, A., Kay, G.F., Cooper, P., Smith, S., McCabe, V.M.,Norris, D.P., Penny, G.D., Patel, D., Rastan, S., 1991. Conservation of positionand exclusive expression of mouse Xist from the inactive X chromosome.Nature 351, 329–331.
Capecchi, M.R., 1989. The new mouse genetics: altering the genome by genetargeting. Trends. Genet 5, 70–76.
Casarosa, S., Fode, C., Guillemot, F., 1999. Mash1 regulates neurogenesis in theventral telencephalon. Development 126, 525–534.
Chen, L.L., Carmichael, G.G., 2010. Decoding the function of nuclear long non-coding RNAs. Curr. Opin. Cell. Biol 22, 357–364.
Chooniedass-Kothari, S., Vincett, D., Yan, Y., Cooper, C., Hamedani, M.K., Myal, Y.,Leygue, E., 2010. The protein encoded by the functional steroid receptor RNAactivator is a new modulator of ER alpha transcriptional activity. FEBS. Lett.584, 1174–1180.
Clark, P.R., Pober, J.S., Kluger, M.S., 2008. Knockdown of TNFR1 by the sense strandof an ICAM-1 siRNA: dissection of an off-target effect. Nucleic Acids. Res. 36,1081–1097.
Clemson, C.M., McNeil, J.A., Willard, H.F., Lawrence, J.B., 1996. XIST RNA paints theinactive X chromosome at interphase: evidence for a novel RNA involved innuclear/chromosome structure. J. Cell. Biol. 132, 259–275.
Cobos, I., Calcagnotto, M.E., Vilaythong, A.J., Thwin, M.T., Noebels, J.L., Baraban, S.C.,Rubenstein, J.L., 2005. Mice lacking Dlx1 show subtype-specific loss ofinterneurons, reduced inhibition and epilepsy. Nat. Neurosci 8, 1059–1068.
Costa, F.F., 2010. Non-coding RNAs: meet thy masters. Bioessays 32, 599–608.Couly, G.F., Coltey, P.M., Le Douarin, N.M., 1993. The triple origin of skull in higher
vertebrates: a study in quail-chick chimeras. Development 117, 409–429.Creuzet, S., Couly, G., Le Douarin, N.M., 2005. Patterning the neural crest
derivatives during development of the vertebrate head: insights from avianstudies. J. Anat. 207, 447–459.
Depew, M.J., Simpson, C.A., Morasso, M., Rubenstein, J.L., 2005. Reassessing the Dlxcode: the genetic regulation of branchial arch skeletal pattern and develop-ment. J. Anat. 207, 501–561.
Dinger, M.E., Amaral, P.P., Mercer, T.R., Pang, K.C., Bruce, S.J., Gardiner, B.B.,Askarian-Amiri, M.E., Ru, K., Solda, G., Simons, C., Sunkin, S.M., Crowe, M.L.,Grimmond, S.M., Perkins, A.C., Mattick, J.S., 2008. Long noncoding RNAs inmouse embryonic stem cell pluripotency and differentiation. Genome. Res. 18,1433–1445.
Eberhart, J.K., He, X., Swartz, M.E., Yan, Y.L., Song, H., Boling, T.C., Kunerth, A.K.,Walker, M.B., Kimmel, C.B., Postlethwait, J.H., 2008. MicroRNA Mirn140modulates Pdgf signaling during palatogenesis. Nat. Genet. 40, 290–298.
Faghihi, M.A., Wahlestedt, C., 2009. Regulatory roles of natural antisense tran-scripts. Nat. Rev. Mol. Cell. Biol. 10, 637–643.
Feng, J., Bi, C., Clark, B.S., Mady, R., Shah, P., Kohtz, J.D., 2006. The Evf-2 noncodingRNA is transcribed from the Dlx-5/6 ultraconserved region and functions as aDlx-2 transcriptional coactivator. Genes. Dev. 20, 1470–1484.
Furusho, M., Ono, K., Takebayashi, H., Masahira, N., Kagawa, T., Ikeda, K., Ikenaka, K.,2006. Involvement of the Olig2 transcription factor in cholinergic neurondevelopment of the basal forebrain. Dev. Biol. 293, 348–357.
Ghanem, N., Jarinova, O., Amores, A., Long, Q., Hatch, G., Park, B.K., Rubenstein, J.L.,Ekker, M., 2003. Regulatory roles of conserved intergenic domains in verte-brate Dlx bigene clusters. Genome. Res. 13, 533–543.
Ginger, M.R., Shore, A.N., Contreras, A., Rijnkels, M., Miller, J., Gonzalez-Rimbau, M.F.,Rosen, J.M., 2006. A noncoding RNA is a potential marker of cell fate duringmammary gland development. Proc. Nat. Acad. Sci. U.S.A. 103, 5781–5786.
Gupta, R.A., Shah, N., Wang, K.C., Kim, J., Horlings, H.M., Wong, D.J., Tsai, M.C.,Hung, T., Argani, P., Rinn, J.L., Wang, Y., Brzoska, P., Kong, B., Li, R., West, R.B.,van de Vijver, M.J., Sukumar, S., Chang, H.Y., 2010. Long non-coding RNAHOTAIR reprograms chromatin state to promote cancer metastasis. Nature464, 1071–1076.
Harborth, J., Elbashir, S.M., Vandenburgh, K., Manninga, H., Scaringe, S.A., Weber, K.,Tuschl, T., 2003. Sequence, chemical, and structural variation of small inter-fering RNAs and short hairpin RNAs and the effect on mammalian genesilencing. Antisense Nucleic Acid Drug Dev 13, 83–105.
Horton, S., Meredith, A., Richardson, J.A., Johnson, J.E., 1999. Correct coordinationof neuronal differentiation events in ventral forebrain requires the bHLH factorMASH1. Mol. Cell. Neurosci 14, 355–369.
Houser, C.R., 2007. Interneurons of the dentate gyrus: an overview of cell types,terminal fields and neurochemical identity. Prog. Brain. Res. 163, 217–232.
Hung, T., Chang, H.Y., 2010. Long noncoding RNA in genome regulation: prospectsand mechanisms. RNA. Biol. 7, 582–585.
Hung, T., Wang, Y., Lin, M.F., Koegel, A.K., Kotake, Y., Grant, G.D., Horlings, H.M.,Shah, N., Umbricht, C., Wang, P., Kong, B., Langerod, A., Borresen-Dale, A.L.,Kim, S.K., Van de Vijver, M., Sukumar, S., Whitfield, M.L., Kellis, M., Xiong, Y.,Wong, D.J., Chang, H.Y., 2011. Extensive and coordinated transcription ofnoncoding RNAs within cell-cycle promoters. Nat. Genet 43, 621–629.
Janowski, B.A., Younger, S.T., Hardy, D.B., Ram, R., Huffman, K.E., Corey, D.R., 2007.Activating gene expression in mammalian cells with promoter-targetedduplex RNAs. Nat. Chem. Biol. 3, 166–173.
Jeong, J., Li, X., McEvilly, R.J., Rosenfeld, M.G., Lufkin, T., Rubenstein, J.L., 2008. Dlxgenes pattern mammalian jaw primordium by regulating both lower jaw-specificand upper jaw-specific genetic programs. Development 135, 2905–2916.
Jones, D.L., Howard, M.A., Stanco, A., Rubenstein, J.L., Baraban, S.C., 2011. Deletionof Dlx1 results in reduced glutamatergic input to hippocampal interneurons.J. Neurophysiol 105, 1984–1991.
Katayama, S., Tomaru, Y., Kasukawa, T., Waki, K., Nakanishi, M., Nakamura, M., Nishida,H., Yap, C.C., Suzuki, M., Kawai, J., Suzuki, H., Carninci, P., Hayashizaki, Y., Wells, C.,

P. Kraus et al. / Developmental Biology 376 (2013) 224–235234
Frith, S., Ravasi, T., Pang, K.C., Hallinan, J., Mattick, J., Hume, D.A., Lipovich, L.,Batalov, S., Engstrom, P.G., Mizuno, Y., Faghihi, M.A., Sandelin, A., Chalk, A.M.,Mottagui-Tabar, S., Liang, Z., Lenhard, B., Wahlestedt, C., 2005. Antisense transcrip-tion in the mammalian transcriptome. Science 309, 1564–1566.
Kawai, J., Shinagawa, A., Shibata, K., Yoshino, M., Itoh, M., Ishii, Y., Arakawa, T.,Hara, A., Fukunishi, Y., Konno, H., Adachi, J., Fukuda, S., Aizawa, K., Izawa, M.,Nishi, K., Kiyosawa, H., Kondo, S., Yamanaka, I., Saito, T., Okazaki, Y.,Gojobori, T., Bono, H., Kasukawa, T., Saito, R., Kadota, K., Matsuda, H.,Ashburner, M., Batalov, S., Casavant, T., Fleischmann, W., Gaasterland, T.,Gissi, C., King, B., Kochiwa, H., Kuehl, P., Lewis, S., Matsuo, Y., Nikaido, I., Pesole, G.,Quackenbush, J., Schriml, L.M., Staubli, F., Suzuki, R., Tomita, M., Wagner, L.,Washio, T., Sakai, K., Okido, T., Furuno, M., Aono, H., Baldarelli, R., Barsh, G., Blake,J., Boffelli, D., Bojunga, N., Carninci, P., De Bonaldo, M.F., Brownstein, M.J., Bult, C.,Fletcher, C., Fujita, M., Gariboldi, M., Gustincich, S., Hill, D., Hofmann, M., Hume,D.A., Kamiya, M., Lee, N.H., Lyons, P., Marchionni, L., Mashima, J., Mazzarelli, J.,Mombaerts, P., Nordone, P., Ring, B., Ringwald, M., Rodriguez, I., Sakamoto, N.,Sasaki, H., Sato, K., Schonbach, C., Seya, T., Shibata, Y., Storch, K.F., Suzuki, H., Toyo-oka, K., Wang, K.H., Weitz, C., Whittaker, C., Wilming, L., Wynshaw-Boris, A.,Yoshida, K., Hasegawa, Y., Kawaji, H., Kohtsuki, S., Hayashizaki, Y., 2001. Functionalannotation of a full-length mouse cDNA collection. Nature 409, 685–690.
Kessaris, N., Fogarty, M., Iannarelli, P., Grist, M., Wegner, M., Richardson, W.D.,2006. Competing waves of oligodendrocytes in the forebrain and postnatalelimination of an embryonic lineage. Nat. Neurosci 9, 173–179.
Kessaris, N., Pringle, N., Richardson, W.D., 2001. Ventral neurogenesis and theneuron-glial switch. Neuron 31, 677–680.
Kino, T., Hurt, D.E., Ichijo, T., Nader, N., Chrousos, G.P., 2010. Noncoding RNA gas5is a growth arrest- and starvation-associated repressor of the glucocorticoidreceptor. Sci. Signal. 3, ra8.
Kraus, P., Fraidenraich, D., Loomis, C.A., 2001. Some distal limb structures developin mice lacking Sonic hedgehog signaling. Mech. Dev 100, 45–58.
Kraus, P., Leong, G., Tan, V., Xing, X., Goh, J.W., Yap, S.P., Lufkin, T., 2010. A morecost effective and rapid high percentage germ-line transmitting chimericmouse generation procedure via microinjection of 2-cell, 4-cell, and 8-cellembryos with ES and iPS cells. Genesis 48, 394–399.
Kraus, P., Lufkin, T., 1999. Mammalian Dlx homeobox gene control of craniofacialand inner ear morphogenesis. J. Cell. Biochem. Suppl. 32�33, 133–140.
Kraus, P., Xing, X., Lim, S.L., Fun, M.E., Sivakamasundari, V., Yap, S.P., Lee, H.,Karuturi, R.K., Lufkin, T., 2012. Mouse strain specific gene expression differencesfor Illumina microarray expression profiling in embryos. BMC. Res. Notes. 5,232.
Lengacher, S., Magistretti, P.J., Pellerin, L., 2004. Quantitative rt-PCR analysis ofuncoupling protein isoforms in mouse brain cortex: methodological optimiza-tion and comparison of expression with brown adipose tissue and skeletalmuscle. J. Cereb. Blood. Flow. Metab. 24, 780–788.
Lewandoski, M., Wassarman, K.M., Martin, G.R., 1997. Zp3-cre, a transgenic mouseline for the activation or inactivation of loxP-flanked target genes specificallyin the female germ line. Curr. Biol. 7, 148–151.
Lipovich, L., Johnson, R., Lin, C.Y., 2010. MacroRNA underdogs in a microRNAworld: evolutionary, regulatory, and biomedical significance of mammalianlong non-protein-coding RNA. Biochim. Biophys. Acta 1799, 597–615.
Liu, J.K., Ghattas, I., Liu, S., Chen, S., Rubenstein, J.L., 1997. Dlx genes encode DNA-binding proteins that are expressed in an overlapping and sequential patternduring basal ganglia differentiation. Dev. Dyn. 210, 498–512.
Lyle, R., Watanabe, D., te Vruchte, D., Lerchner, W., Smrzka, O.W., Wutz, A., Schage-man, J., Hahner, L., Davies, C., Barlow, D.P., 2000. The imprinted antisense RNA atthe Igf2r locus overlaps but does not imprint Mas1. Nat. Genet 25, 19–21.
Mao, R., Page, D.T., Merzlyak, I., Kim, C., Tecott, L.H., Janak, P.H., Rubenstein, J.L.,Sur, M., 2009. Reduced conditioned fear response in mice that lack Dlx1 andshow subtype-specific loss of interneurons. J. Neurodev. Disord 1, 224–236.
Martianov, I., Ramadass, A., Serra Barros, A., Chow, N., Akoulitchev, A., 2007.Repression of the human dihydrofolate reductase gene by a non-codinginterfering transcript. Nature 445, 666–670.
McBratney-Owen, B., Iseki, S., Bamforth, S.D., Olsen, B.R., Morriss-Kay, G.M., 2008.Development and tissue origins of the mammalian cranial base. Dev. Biol. 322,121–132.
McGuinness, T., Porteus, M.H., Smiga, S., Bulfone, A., Kingsley, C., Qiu, M., Liu, J.K.,Long, J.E., Xu, D., Rubenstein, J.L., 1996. Sequence, organization, and transcrip-tion of the Dlx-1 and Dlx-2 locus. Genomics 35, 473–485.
Meza-Sosa, K.F., Valle-Garcia, D., Pedraza-Alva, G., Perez-Martinez, L., 2012. Role ofmicroRNAs in central nervous system development and pathology. J. Neurosci. Res.90, 1–12.
Miyoshi, G., Butt, S.J., Takebayashi, H., Fishell, G., 2007. Physiologically distinct temporalcohorts of cortical interneurons arise from telencephalic Olig2-expressingprecursors. J. Neurosci 27, 7786–7798.
Modarresi, F., Faghihi, M.A., Lopez-Toledano, M.A., Fatemi, R.P., Magistri, M.,Brothers, S.P., van der Brug, M.P., Wahlestedt, C., 2012. Inhibition of naturalantisense transcripts in vivo results in gene-specific transcriptional upregula-tion. Nat. Biotechnol 30, 453–459.
Mu, X., Beremand, P.D., Zhao, S., Pershad, R., Sun, H., Scarpa, A., Liang, S., Thomas, T.L.,Klein, W.H., 2004. Discrete gene sets depend on POU domain transcription factorBrn3b/Brn-3.2/POU4f2 for their expression in the mouse embryonic retina.Development 131, 1197–1210.
Nagano, T., Mitchell, J.A., Sanz, L.A., Pauler, F.M., Ferguson-Smith, A.C., Feil, R.,Fraser, P., 2008. The Air noncoding RNA epigenetically silences transcriptionby targeting G9a to chromatin. Science 322, 1717–1720.
Nery, S., Wichterle, H., Fishell, G., 2001. Sonic hedgehog contributes tooligodendrocyte specification in the mammalian forebrain. Development 128,527–540.
Niwa, H., Yamamura, K., Miyazaki, J., 1991. Efficient selection for high-expressiontransfectants with a novel eukaryotic vector. Gene 108, 193–199.
Ono, K., Takebayashi, H., Ikeda, K., Furusho, M., Nishizawa, T., Watanabe, K.,Ikenaka, K., 2008. Regional- and temporal-dependent changes in the differ-entiation of Olig2 progenitors in the forebrain, and the impact on astrocytedevelopment in the dorsal pallium. Dev. Biol. 320, 456–468.
Panganiban, G., Rubenstein, J.L., 2002. Developmental functions of the Distal-less/Dlx homeobox genes. Development 129, 4371–4386.
Parras, C.M., Hunt, C., Sugimori, M., Nakafuku, M., Rowitch, D., Guillemot, F., 2007.The proneural gene Mash1 specifies an early population of telencephalicoligodendrocytes. J. Neurosci 27, 4233–4242.
Petryniak, M.A., Potter, G.B., Rowitch, D.H., Rubenstein, J.L., 2007. Dlx1 and Dlx2control neuronal versus oligodendroglial cell fate acquisition in the developingforebrain. Neuron 55, 417–433.
Poitras, L., Ghanem, N., Hatch, G., Ekker, M., 2007. The proneural determinantMASH1 regulates forebrain Dlx1/2 expression through the I12b intergenicenhancer. Development 134, 1755–1765.
Pringle, N.P., Richardson, W.D., 1993. A singularity of PDGF alpha-receptorexpression in the dorsoventral axis of the neural tube may define the originof the oligodendrocyte lineage. Development 117, 525–533.
Qiu, M., Bulfone, A., Ghattas, I., Meneses, J.J., Christensen, L., Sharpe, P.T.,Presley, R., Pedersen, R.A., Rubenstein, J.L., 1997. Role of the Dlx homeoboxgenes in proximodistal patterning of the branchial arches: mutations of Dlx-1,Dlx-2, and Dlx-1 and -2 alter morphogenesis of proximal skeletal and softtissue structures derived from the first and second arches. Dev. Biol. 185,165–184.
Qureshi, I.A., Mattick, J.S., Mehler, M.F., 2010. Long non-coding RNAs in nervoussystem function and disease. Brain Res 1338, 20–35.
Rapicavoli, N.A., Poth, E.M., Zhu, H., Blackshaw, S., 2011. The long noncoding RNASix3OS acts in trans to regulate retinal development by modulating Six3activity. Neural. Dev. 6, 32.
Redon, S., Reichenbach, P., Lingner, J., 2010. The non-coding RNA TERRA is anatural ligand and direct inhibitor of human telomerase. Nucleic Acids Res. 38,5797–5806.
Richardson, W.D., Kessaris, N., Pringle, N., 2006. Oligodendrocyte wars. Nat. Rev.Neurosci 7, 11–18.
Rinn, J.L., Kertesz, M., Wang, J.K., Squazzo, S.L., Xu, X., Brugmann, S.A., Goodnough, L.H.,Helms, J.A., Farnham, P.J., Segal, E., Chang, H.Y., 2007. Functional demarcation ofactive and silent chromatin domains in human HOX loci by noncoding RNAs. Cell129, 1311–1323.
Robledo, R.F., Rajan, L., Li, X., Lufkin, T., 2002. The Dlx5 and Dlx6 homeobox genesare essential for craniofacial, axial, and appendicular skeletal development.Genes. Dev. 16, 1089–1101.
Saino-Saito, S., Berlin, R., Baker, H., 2003. Dlx-1 and Dlx-2 expression in the adultmouse brain: relationship to dopaminergic phenotypic regulation. J. Comp.Neurol. 461, 18–30.
Sana, J., Faltejskova, P., Svoboda, M., Slaby, O., 2012. Novel classes of non-codingRNAs and cancer. J. Transl. Med 10, 103.
Scacheri, P.C., Rozenblatt-Rosen, O., Caplen, N.J., Wolfsberg, T.G., Umayam, L.,Lee, J.C., Hughes, C.M., Shanmugam, K.S., Bhattacharjee, A., Meyerson, M.,Collins, F.S., 2004. Short interfering RNAs can induce unexpected and diver-gent changes in the levels of untargeted proteins in mammalian cells. Proc.Nat. Acad. Sci. U.S.A. 101, 1892–1897.
Sheik Mohamed, J., Gaughwin, P.M., Lim, B., Robson, P., Lipovich, L., 2010.Conserved long noncoding RNAs transcriptionally regulated by Oct4 andNanog modulate pluripotency in mouse embryonic stem cells. RNA 16,324–337.
Sinclair, M.S., Perea-Martinez, I., Dvoryanchikov, G., Yoshida, M., Nishimori, K.,Roper, S.D., Chaudhari, N., 2010. Oxytocin signaling in mouse taste buds. PLoS.One 5, e11980.
Sioud, M., 2011. Promises and challenges in developing RNAi as a research tool andtherapy. Methods Mol. Biol. 703, 173–187.
Sleutels, F., Zwart, R., Barlow, D.P., 2002. The non-coding Air RNA is required forsilencing autosomal imprinted genes. Nature 415, 810–813.
St Laurent 3rd, G., Faghihi, M.A., Wahlestedt, C., 2009. Non-coding RNA transcripts:sensors of neuronal stress, modulators of synaptic plasticity, and agents ofchange in the onset of Alzheimer’s disease. Neurosci. Lett. 466, 81–88.
Stecca, B., Ruiz i Altaba, A., 2009. A GLI1-p53 inhibitory loop controls neural stemcell and tumour cell numbers. EMBO J 28, 663–676.
Stuhmer, T., Anderson, S.A., Ekker, M., Rubenstein, J.L., 2002. Ectopic expression ofthe Dlx genes induces glutamic acid decarboxylase and Dlx expression.Development 129, 245–252.
Tallent, M.K., 2007. Somatostatin in the dentate gyrus. Prog. Brain. Res. 163,265–284.
Tekki-Kessaris, N., Woodruff, R., Hall, A.C., Gaffield, W., Kimura, S., Stiles, C.D.,Rowitch, D.H., Richardson, W.D., 2001. Hedgehog-dependent oligodendrocytelineage specification in the telencephalon. Development 128, 2545–2554.
Thakur, N., Tiwari, V.K., Thomassin, H., Pandey, R.R., Kanduri, M., Gondor, A.,Grange, T., Ohlsson, R., Kanduri, C., 2004. An antisense RNA regulates thebidirectional silencing property of the Kcnq1 imprinting control region. Mol.Cell. Biol. 24, 7855–7862.
Vanhee-Brossollet, C., Vaquero, C., 1998. Do natural antisense transcripts makesense in eukaryotes? Gene 211, 1–9.

P. Kraus et al. / Developmental Biology 376 (2013) 224–235 235
Wang, K.C., Chang, H.Y., 2011. Molecular mechanisms of long noncoding RNAs.Mol. Cell 43, 904–914.
Wang, K.C., Yang, Y.W., Liu, B., Sanyal, A., Corces-Zimmerman, R., Chen, Y., Lajoie, B.R.,Protacio, A., Flynn, R.A., Gupta, R.A., Wysocka, J., Lei, M., Dekker, J., Helms, J.A.,Chang, H.Y., 2011. A long noncoding RNA maintains active chromatin tocoordinate homeotic gene expression. Nature 472, 120–124.
Watts, J.K., Yu, D., Charisse, K., Montaillier, C., Potier, P., Manoharan, M., Corey, D.R.,2010. Effect of chemical modifications on modulation of gene expression byduplex antigene RNAs that are complementary to non-coding transcripts atgene promoters. Nucleic Acids Res. 38, 5242–5259.
Werner, A., 2005. Natural antisense transcripts. RNA. Biol. 2, 53–62.
Wonders, C., Anderson, S., 2005. Beyond migration: Dlx1 regulates interneurondifferentiation. Nat. Neurosci 8, 979–981.
Young, T.L., Matsuda, T., Cepko, C.L., 2005. The noncoding RNA taurine upregulatedgene 1 is required for differentiation of the murine retina. Curr. Biol. 15, 501–512.
Yun, K., Fischman, S., Johnson, J., Hrabe de Angelis, M., Weinmaster, G., Rubenstein, J.L,2002. Modulation of the notch signaling by Mash1 and Dlx1/2 regulatessequential specification and differentiation of progenitor cell types in thesubcortical telencephalon. Development 129, 5029–5040.
Zhou, Q., Choi, G., Anderson, D.J., 2001. The bHLH transcription factor Olig2 promotesoligodendrocyte differentiation in collaboration with Nkx2.2. Neuron 31,791–807.