lsm1102_inheritance and bio genesis of organelles in the secretory pathway
TRANSCRIPT

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 1/11
Eukaryotic cells have membrane-bound organelles in orderto compartmentalize and organize cellular functions. Thisenables a diverse range of different environments to co-exist within a single cell, and has enormous implicationsin terms of the diversity of functions that can be carriedout. At the same time as proteins are being degraded inthe acidic environment of the lysosomes, new proteins arebeing synthesized in the surrounding cytoplasm and traf-ficked through the organelles of the secretory pathway. It istherefore not surprising that failure of this system can leadto a number of diseases in humans. This can take numer-ous different forms, which include congenital disorders of glycosylation, specific mutations in proteins unable to foldcorrectly and achieve their normal localization, or muta-tions in proteins required for secretory pathway func-tion1,2. The problem with possessing membrane-boundorganelles such as the endoplasmic reticulum (ER) andGolgi apparatus is that they have to be maintained. In non-dividing cells this is a simple housekeeping task — lipidand protein biosynthesis are associated with the ER, andthe required components can then be delivered to the
other parts of the system, such as the Golgi, by vesiculartrafficking. However, what happens when cells divide?Do they create additional copies of their organelles in thedaughter cells by dividing the existing copies, or build newones de novo? Here we will discuss the different aspectsof ER and Golgi inheritance, and how these involve boththe division of pre-existing copies of these organelles and de novo formation of new copies.
Models for organelle biogenesis and inheritance
In principle, there are a number of ways in whichthe problem of organelle biogenesis can be solved(FIG. 1). The most simple solution is de novo synthesis.
In its purest form, this means that, provided with theinformation encoded in the genetic material andthe machinery needed to interpret this, the cell can pro-duce the organelle with no information in the form of atemplate or copy of the organelle. For membrane-boundorganelles, the concept of de novo synthesis needs to bemodified to take into account the source of the mem-brane — both lipid and protein components. Therefore,a cell can create the organelles it needs to live.
The alternative to de novo synthesis is the inheritanceof the organelle in question, or of a template that is neces-sary for building the organelle. This problem can be sub-divided, depending on whether the organelle is present insingle or multiple copies. A single-copy organelle can beduplicated and then segregated prior to cell division, orbroken down into parts that are then shared out betweenthe two daughter cells. In theory, multi-copy organellescan simply be shared out and do not need to be dismantled.A further issue concerns the segregation or inheritanceprocess itself. If there are sufficient copies of the organelleor fragments derived from the original intact organelle,
then a stochastic partitioning mechanism can explainefficient inheritance. However, for organelles present inlow copy numbers, this mechanism fails, and an activesegregation process has to be invoked. Once cell divisionis accomplished, a growth phase then ensues. Organellesincrease in amount, a process termed biogenesis,to be ready for the next cell-division event.
For both single- and multiple-copy organelles, bio-genesis itself can occur in two ways. Either the organellecan grow and then divide once it has reached a criticalsize, or it might form a template from which a newcopy of the organelle is created alongside the original.It is important to note that there is nothing exclusive
*Faculty of Life Sciences,
University of Manchester,
The Michael Smith Building,
Oxford Road, Manchester
M13 9PT, UK.‡University of Liverpool
Cancer Studies Centre,
200 London Road,
Liverpool L3 9TA, UK.
Correspondence to F.A.B.
e-mail: [email protected]
doi:10.1038/nrm2179
Published online 16 May 2007
Inheritance and biogenesis of organelles in the secretory pathwayMartin Lowe* and Francis A. Barr ‡
Abstract | In eukaryotic cells, cellular functions are compartmentalized into membrane-
bound organelles. This has many advantages, as shown by the success of the eukaryotic
lineage, but creates many problems for cells, such as the need to build and partition these
organelles during cell growth and division. Diverse mechanisms for biogenesis of the
endoplasmic reticulum and Golgi apparatus have evolved, ranging from de novo synthesis tothe copying of a template organelle. The different mechanisms by which organelles are
inherited in yeasts, protozoa and metazoans probably reflect the differences in the structure
and copy number of these organelles.
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 429
REVIEWS

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 2/11
Single copy Multiple copy
Cell growth
Cell division
de novo nucleation/growth
Templated assembly/growth
Growth/fission
Duplication
Active segregation
Duplication
Stochastic partitioning
a
b
Centrosome
The main microtubule
organizing centre of animal
cells.
Rab GTPases
Rab proteins are Ras-like
GTPases that regulate
membrane-trafficking events in
eukaryotic cells. Different Rab
proteins are specific for
different transport pathways
and different subcellular
compartments.
Kinesin-1
A member of the kinesin family
of microtubule-based motors,
which typically move towards
microtubule plus ends.
about these models. Depending on the environmentalsituation, life cycle or developmental stage, it mightbe possible to switch between de novo synthesis andinheritance of pre-existing organelles. A good exampleof this is provided by the centrosome, which is normally copied and then segregated between the two daughtercells3. However, in the mouse embryo, the early divisionsoccur in the absence of centrosomes, and these are thenproduced de novo prior to implantation4.
The endoplasmic reticulum
In all organisms, the ER is a continuous tubular-reticularnetwork that is connected with the nuclear envelope(NE), yet some aspects of its organization differ (FIG. 2a,b).This topic has been extensively reviewed for yeast andanimal cells elsewhere5,6, and we will therefore focus onthe aspects of ER structure that are most relevant to ERinheritance in yeast and animal cells here.
In budding yeast, the perinuclear ER can be viewed as asubdomain of the NE, whereas the bulk of the ER, referredto as peripheral or cortical ER, lies under the plasmamembrane. By contrast, in most metazoan cells, includ-ing mammalian cells, the ER forms a spread hexagonalnetwork joined to the NE. Although little is known about
how the ER is shaped in any organism, recent progresssuggests that there are likely to be some common themes.Using a combination of biochemistry in a frog extractsystem and yeast genetics, integral membrane proteinsof the reticulon family and Yop1, an integral membraneprotein that interacts with the yeast reticulon Rtn1, wereidentified as factors able to target to and promote theformation of ER tubules7,8. Deletion of yeast Rtn1 incombination with Yop1 causes a change in cortical ERmorphology from predominantly tubular-reticular to amore sheet-like cisternal morphology 7,8. Reticulons arethought to adopt a hairpin structure, and it has been pro-posed that this promotes the formation of, or stabilizes,the tubular structure of the ER. How Yop1 contributes toER structure is less clear. It was originally identified as aninteraction partner of another integral membrane protein,Yip1, which belongs to a family of factors that are thoughtto be involved in the insertion of Rab GTPases into theirtarget membranes9–11. Therefore, Rab-mediated mem-brane tethering and fusion events might be important formaintaining the normal organization of the ER.
Several other observations regarding the organizationof the ER are likely to be specific for animal cells and donot appear to apply to budding yeast. It has long beenknown that the extended tubular-reticular network of the ER in animal cells is aligned with microtubules12, yetis highly dynamic13,14. This is due to a combination of direct anchoring of the ER to microtubules and exten-sion of the ER along microtubules by the kinesin-1 motorprotein. Kinesin-1 can extend the ER along microtubulesto form a tubular network in vitro15,16, and this is facili-tated through an integral membrane protein receptor onthe ER called kinectin17,18. More recent findings haveidentified cytoskeleton-associated protein 4 (CKAP4;also known as CLIMP63), another integral membraneprotein, as a microtubule attachment factor for the ER19.This combination of extension and anchoring is probably required to maintain the ER network, as microtubules arehighly dynamic structures that continuously polymerizeand depolymerize.
Mechanism of ER inheritance in yeast
Yeasts have a closed mitosis in which the NE does not breakdown and the mitotic spindle is inside the nucleus (FIG. 3).This has the consequence that the NE and perinuclear ERare partitioned together with the DNA through the actionof astral microtubules. These position the nucleus such thatit will be divided into two equal parts in the mother and
daughter cells when cell division occurs20,21. In buddingyeast, the fate of the cortical ER is radically different to thatof the perinuclear ER. Morphological studies have revealedthat the cortical pool of ER first extends into the budand then becomes anchored at the tip of the growingbud, before expanding along the cortex to fill the bud22.
The machinery for ER inheritance. Genetic approachesin budding yeast have started to elucidate the machinery required for localizing the ER to the cell cortex, and fordirecting its inheritance during cell division. Defects incomponents of the coatomer complex protein-I (COPI)pathway, required for retrograde transport from the
Figure 1 | Conceptual models for organelle biogenesis and inheritance. a | In principle,
there are a number of ways that organelle biogenesis can occur in proliferating cells.
Biogenesis can occur by de novo synthesis, which in its purest sense means that a new copy
of the organelle is generated in the absence of a template or existing copy of the organelle.
Alternatively, organelle biogenesis can occur through templated assembly and growth, or
through growth followed by fission. b | The principles that govern organelle inheritance will
depend on how many copies of the organelle are present in the cell. A single-copy
organelle can be duplicated and then segregated prior to cell division, or broken down into
parts that are then shared out between the two daughter cells. Multiple-copy organelles
can, in theory, simply be shared out and do not need to be dismantled. In this case, a
stochastic-partitioning mechanism can explain efficient inheritance of the organelle, but
for low-copy-number organelles this mechanism is not sufficient to ensure equal
partitioning, and an active segregation process needs to be invoked.
R E V I E W S
430 | JUNE 2007 | VOLUME 8 www.nature.com/reviews/molcellbio
8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 3/11
a
b
c
d
Mitotic spindle
A highly dynamic array of
microtubules that forms during
mitosis and serves to move the
duplicated chromosomes
apart.
COPI
Coat-protein complexes that
are required for vesicle
formation and trafficking
between the endoplasmic
reticulum and Golgi.
Unknown function
essential-1
(Ufe1). A SNARE protein
involved in both fusion
between endoplasmic
reticulum membranes and
vesicle fusion with the
endoplasmic reticulum.
Golgi to the ER, or the SNARE protein unknown function
essential-1 (Ufe1)23, alter normal ER structure from atubular to a sheet-like morphology, with a correspond-ing relocation of peripheral ER away from the cellcortex24. Presumably, this is because the compositionof the ER changes if recycling back from the Golgi isblocked. Consistent with this idea, mutants in the signalrecognition particle that is needed to target secretory and membrane proteins to the ER, also have altered ER
morphology 24.The cortical pool of tubular ER is inherited by an
actin-dependent process involving the myosin V family motor Myo4 and an adaptor protein, She3 (REF. 25).Disruption of actin through the use of drugs, the dele-tion of Myo4 or mutations that inactivate the ATP-binding domain in Myo4 reveal that this actin- andmotor-activity-dependent-pathway is required forthe directed extension of ER tubules into the bud 25.In addition to actin and myosin, a number of othercomponents required for cortical ER inheritance havebeen identified. Ice2 (inheritance of cortical ER-2) isa multi-spanning transmembrane protein that, when
disrupted, leads to a collapse of the cortical ER in themother cell and a failure of the cortical ER to enter thebud26. The precise function of Ice2 remains to be estab-lished, but it seems to be required for both the normaltubular morphology of the cortical ER and its transportinto the bud. Mutants in the yeast auxilin Aux1 (alsoknown as synthetic lethal with Arf1 (Swa2)) causespecific defects in the inheritance of the cortical but notthe perinuclear ER or NE27. One interesting possibility
to account for this finding comes from the identificationof a ubiquitin-associated (UBA)-type ubiquitin-bindingdomain within Aux1 (REF. 28). Ubiquitin-mediatedpathways have many functions at the ER, as part of the ER-associated degradation pathway that deals withmisfolded proteins29, and as regulatory components of specific signalling pathways30. The role of Aux1 in corti-cal ER inheritance suggests that ubiquitylation mightalso be important for this process. This idea is supportedby observations that the ubiquitin-dependent chaperoneCdc48 (cell division cycle-48; homologous to themammalian protein p97) is required for remodellingthe nucleus and ER during yeast mating31,32.
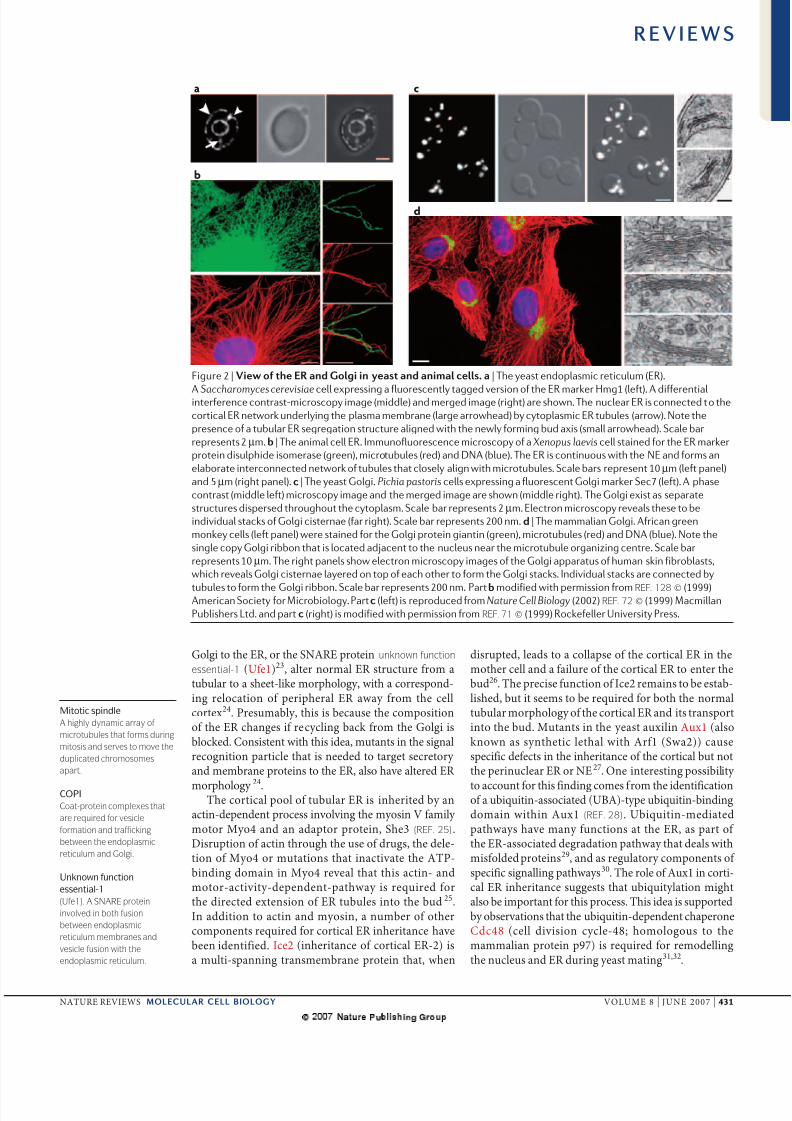
Figure 2 | View of the ER and Golgi in yeast and animal cells. a | The yeast endoplasmic reticulum (ER).
A Saccharomyces cerevisiae cell expressing a fluorescently tagged version of the ER marker Hmg1 (left). A differentialinterference contrast-microscopy image (middle) and merged image (right) are shown. The nuclear ER is connected to the
cortical ER network underlying the plasma membrane (large arrowhead) by cytoplasmic ER tubules (arrow). Note the
presence of a tubular ER segregation structure aligned with the newly forming bud axis (small arrowhead). Scale bar
represents 2μm. b | The animal cell ER. Immunofluorescence microscopy of a Xenopus laevis cell stained for the ER marker
protein disulphide isomerase (green), microtubules (red) and DNA (blue). The ER is continuous with the NE and forms an
elaborate interconnected network of tubules that closely align with microtubules. Scale bars represent 10μm (left panel)
and 5μm (right panel). c | The yeast Golgi. Pichia pastoris cells expressing a fluorescent Golgi marker Sec7 (left). A phase
contrast (middle left) microscopy image and the merged image are shown (middle right). The Golgi exist as separate
structures dispersed throughout the cytoplasm. Scale bar represents 2μm. Electron microscopy reveals these to be
individual stacks of Golgi cisternae (far right). Scale bar represents 200 nm. d | The mammalian Golgi. African green
monkey cells (left panel) were stained for the Golgi protein giantin (green), microtubules (red) and DNA (blue). Note the
single copy Golgi ribbon that is located adjacent to the nucleus near the microtubule organizing centre. Scale bar
represents 10μm. The right panels show electron microscopy images of the Golgi apparatus of human skin fibroblasts,
which reveals Golgi cisternae layered on top of each other to form the Golgi stacks. Individual stacks are connected by
tubules to form the Golgi ribbon. Scale bar represents 200 nm. Partb modified with permission from REF. 128 © (1999)American Society for Microbiology. Partc (left) is reproduced fromNature Cell Biology (2002) REF. 72 © (1999) Macmillan
Publishers Ltd. and part c (right) is modified with permission from REF. 71 © (1999) Rockefeller University Press.
R E V I E W S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 431

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 4/11
Chromosomes
Exocyst
c Cortical ER
Centrosome/spindlepole body
Mitotic spindle
NE
NE
Cortical ER
Actin
Actin
ER tubule
ER tubule
Myo4
She3
a
b Nuclear envelope
Exocyst
A multisubunit protein
complex that is important for
docking secretory vesicles with
the plasma membrane, and
anchoring the endoplasmic
reticulum to the plasma
membrane during cell division.
Sec61 transloconAn endoplasmic reticulum
(ER)-localized protein complex
that facilitates the insertion of
newly synthesized secretory
and membrane proteins into
the ER.
Cdc42
A small Rho-family GTPase that
regulates localized actin
dynamics in cells.
CDK1
The catalytic subunit of the
principal serine/threonine
kinase that regulates entry intomitosis.
B-type cyclin
The regulatory subunit of the
principal mitotic serine/
threonine kinase. In mammals,
cyclin B binds to the catalytic
subunit CDK1.
Cdc28–cyclin-2
A serine/threonine kinase that
regulates entry into S phase,
comprising a catalytic subunit
(Cdc28) and a regulatory
subunit (cyclin-2).
Landmarks for ER inheritance. Once cortical ER tubulesenter the bud, they become anchored at the bud tip, andthis process requires the function of the exocyst, a multisubunit tethering complex required for polarized trans-port of secretory vesicles into the bud33,34. A numberof exocyst subunits have been linked to ER inheritance,one of which is Sec335–37. Sec3 is not an essential componentof the exocyst, and in its absence cells are still capable of secretion35. However polarization of the bud is altered,and inheritance of the cortical ER but not the Golgi ormitochondria is defective35. Conversely, overproduc-tion of Sec3 appears to promote the capture of ER in
the bud, and it has therefore been proposed that thisis a key component of the inheritance machinery thatmight function as a spatial landmark for cortical ERinheritance35. Significantly, another exocyst subunit,Sec6, was identified as an interaction partner of Rtn1(REF. 8), which as discussed above functions in shapingthe cortical ER7. Therefore, Rtn1 might form part of areceptor on the ER that can interact with the exocystand promote anchoring of the cortical ER at the budtip. Intriguingly, the exocyst can also interact withthe Sec61 translocon, and this might form a furtherway to link the ER to the bud tip38. Although failing togive a complete picture, taken together, these findings
suggest that multiple activities associated with the ER arecrucial for its normal morphology and inheritance.
Regulation of yeast ER inheritance
An important issue concerns the regulation and propertiming of ER inheritance during the cell cycle. After exit-ing mitosis, yeast cells grow throughout the G1 phase.This is reflected by the random organization of theactin cytoskeleton, and hence non-polarized delivery of secretory vesicles to the cell surface39. At the G1–Stransition, bud formation is initiated, and this is coupledto the polarization of the actin cytoskeleton through a
mechanism involving local activation of the Rho-family GTPase Cdc42
39–41. Cdc42 also interacts with the exocystand is required for the polarized delivery of secretory
vesicles to the site of the forming bud42. Therefore, theyeast switches from so-called isotropic growth to budding— a form of polarized growth. This process is inhibitedin mitosis and promoted during entry into S phaseby different cyclin-dependent kinase complexes.
During mitosis, the mitotic kinase Cdc28 (the bud-ding yeast homologue of CDK1), which is associated withone of a number of B-type cyclins, prevents formation of anew bud43,44. At the onset of S phase, activation of Cdc28–
cyclin-2 (Cln2) triggers the relocation of Cdc24, the
Figure 3 | Inheritance of the ER in yeast. a | In Saccharomyces cerevisiae, the nuclear envelope (NE) is connected by
tubules to the cortical endoplasmic reticulum (ER) network, which is located close to the plasma membrane.
b | Inheritance of the NE takes place during mitosis. Microtubules elongate inside the nucleus along the mother–bud axis
to form the mitotic spindle, pushing the NE into the bud. The NE is therefore segregated alongside the chromosomes
between the mother and daughter cells. c | The cortical ER uses the actin cytoskeleton for its inheritance. ER tubules
move along actin cables across the mother–bud axis using the Myo4 myosin motor protein, which is attached to the ER
membrane by its adaptor protein, She3. After reaching the bud tip, the ER tubule is anchored by the exocyst subunit
Sec3. Another exocyst subunit, Sec6, also appears to have a role in cortical ER inheritance, perhaps by binding to the ER
reticulon protein Rtn1 (not shown). Following attachment to the cortex at the bud tip, ER tubules subsequently spread
along the cell periphery to form the lattice-like cortical ER network in the daughter cell.
R E V I E W S
432 | JUNE 2007 | VOLUME 8 www.nature.com/reviews/molcellbio

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 5/11
ER networkNE
Microtubule
CentrosomeMitoticspindleChromosomes
G2
G1 Telophase Anaphase
Prophase Metaphase
Dynein
A multisubunit microtubule-
based motor, typically movingtowards microtubule minus
ends.
Nuclear lamina
A structure composed of lamin
intermediate-filament proteins
that is important for integrity of
nuclear envelope.
Golgi matrix
A biochemically defined
meshwork of proteins that
retains the characteristic
cisternal shape of Golgi
membranes.
guanine exchange factor for Cdc42, from the nucleus tothe cell cortex in the region of the presumptive bud site.Here it promotes Cdc42 activation and subsequent actinpolarization45. Because ER inheritance is actin dependent,this process will start only once the actin cytoskeletonhas polarized and bud formation has commenced at theG1–S transition.
Recent observations have indicated that a mitogen-activated protein kinase (MAPK) pathway activated by cell-wall integrity defects can control ER inheritance inyeast46. Although it is not immediately apparent howthis relates to the cell-cycle control of bud formation,there is an intriguing link between these two processes.Environmental stresses can cause perturbations in theactin cytoskeleton, and this triggers a delay in bud form-ation while the cell adapts to these altered conditions47.It has therefore been proposed that ER inheritance is linkedto sensing of osmotic stress and cell-wall integrity sensingpathways46,48. These signalling mechanisms together
with active actin–myosin-dependent transport couplethe inheritance of the cortical ER with the process of budformation in yeast.
ER inheritance in mammalian cells. In mammalian cells,the NE breaks down in mitosis and NE integral mem-brane proteins become spread throughout the ER49,50.However, although undergoing large structural changes,the ER is not actually broken down into fragments or ves-icles (FIG. 4). Measurements of long-range diffusion withinthe ER show that this is unaltered in mitosis. This findingsuggests that the reticulum remains intact, and that whatis happening is simply a topological change51,52.
Cell-biological studies have shown that microtubulesand the motor protein dynein work together to peel theNE and associated ER from the surface of the chroma-tin53,54. This requires the action of mitotic kinases suchas cyclin-dependent kinase-1 (CDK1), which results inthe disassembly of the nuclear lamina and releases theanchoring between membranes and chromatin50,55. Oncereleased from the chromatin, what happens to the ER? Asdiscussed above, the ER is attached to and moved alongmicrotubules, but this does not seem to be relevant for ERinheritance. Rather, the attachment between the ER andmicrotubules appears to be lost, in part owing to the phos-phorylation of CLIMP63 in mitosis19. Consistent with thisidea, the ER is excluded from the region of the cell that isoccupied by the mitotic spindle and chromosomes, butspreads throughout the rest of the cell volume52.
The Golgi apparatus
The Golgi apparatus is a cisternal array of membranes,either highly organized into a stack as in plants, ani-mals and protozoa, or a more dynamic arrangement
of free cisternae and stacks as seen in fungi (FIG. 2c,d).Mammalian cells organize these stacks into a single-copy array or ribbon in interphase cells, whereas plants andflies have multiple single stacks dispersed throughoutthe cytoplasm. These different types of organizationseem to be important in terms of function. Duringdevelopment in Drosophila melanogaster , it is possiblefor a single cell to use different groups of Golgi stacks,termed exocytic units, to transport a selected set of cargo proteins to a defined destination on the cell sur-face. This creates a polarized signal that is importantfor controlling the development of the anteroposteriorand dorsoventral axes of the oocyte56. Mammaliancells, with their functionally single-copy Golgi, cannotdo this and must therefore solve similar problems in adifferent way, perhaps by additional specific membrane-trafficking pathways. Some of these differences reflectthe different interactions of Golgi with the cytoskeletonin different organisms. The perinuclear Golgi ribbonseen in mammalian cells is explained by interactionswith the microtubule cytoskeleton and the activities of dynein and other microtubule-dependent motors57,58.
Yeast genetics and cell-biological approaches inmammalian cells have identified many factors that arerequired for normal Golgi organization, and this hasbeen reviewed in detail elsewhere59–61. It is striking thatmany of the components identified seem to be compo-
nents of the transport machinery, which suggests thatcontinuing vesicular transport is a key factor in deter-mining Golgi structure62. Electron microscopy andbiochemical approaches indicate that there is a struc-tural matrix (the Golgi matrix) of proteins analogous tothe nuclear lamina at the surface of the Golgi cisternae,and this has been implicated in Golgi stack and ribbonformation63–68. However, it is now clear that no singlecomponent acts alone to mediate Golgi organization;rather, this matrix is made up of many redundant com-ponents acting in different pathways. Indirect supportfor this idea is provided by the identification of a mam-malian cell line that is temperature sensitive for secretion
Figure 4 | Inheritance of the ER in mammalian cells. In interphase, the endoplasmic
reticulum (ER) extends throughout the cytoplasm as an interconnected network of
tubules (purple), organized by the microtubule cytoskeleton (orange). The ER is
continuous with the nuclear envelope (NE). During prophase, nuclear lamins become
phosphorylated, releasing the anchoring between membrane and chromatin, but the NE
remains intact at this point. NE breakdown occurs as cells enter prometaphase as a
consequence of microtubule-induced tearing, and NE membrane components become
dispersed throughout the metaphase ER network (green ER network). Connectionsbetween the ER and microtubules are lost in mitosis owing to the phosphorylation of
proteins that link these structures. ER partitioning into the newly forming daughter cells
is ensured by the spreading of the network throughout the cytoplasm of the dividing
mother cell. During telophase, the NE begins to reform around the segregated
chromosomes, ultimately becoming continuous with the extended ER network, which
itself has re-attached to the microtubule cytoskeleton.
R E V I E W S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 433

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 6/11
2:12 4:12 6:00 11:00 13:54
Existing Golgi elements (red)adjacent to ER exit sites (green)
De novo assembly of ER exit sites,closely followed by a new Golgi
Nucleus GolgiBasal
bodies
Anterior Posterior
a
bc
ER exit site
A specialized site in the
endoplasmic reticulum (ER)where vesicles are generated
that transport proteins from
the ER to the Golgi.
Basal body
The microtubule organizing
centre of protozoa, equivalent
to the centrosome of animal
cells, typically the nucleating
site for flagella and cilia.
Centrin
A calcium-binding protein that
is associated with microtubule
organizing centres.
and growth owing to the lack of Golgi matrix proteinof 130 kDa (GM130)69. GM130 has been implicated in
shaping Golgi cisternae and in the assembly of Golgistacks into a ribbon-like array 64,65,68, yet in its absencecells appear to have a functional Golgi apparatus andgrow normally at 34°C (REF. 69). Exactly what this meanscan be debated, but it does suggest that GM130, andby extension other Golgi matrix proteins, contributeto Golgi organization and oppose the disruptive effectsof thermal and perhaps other stresses. However, they might not be essential under all conditions. A parallelcould be drawn here with the role of lamins in theNE. Whereas lamin B is essential for NE integrity andmorphology, lamins A and C are dispensable for thisboth in vitro and in vivo. However, it would be wrong to
conclude that lamin A or C are not important structuralcomponents of the nucleus. Mutations or loss of laminA or C result in subtle alterations to the properties of the nucleus, and this manifests itself in the form of anumber of human diseases — termed laminopathies— that include Hutchinson–Gilford progeria, a prematureageing syndrome70.
Diverse mechanisms of Golgi biogenesis
Reflecting the different types of Golgi organization in dif-ferent organisms, there seems to be diverse mechanisms of Golgi biogenesis — from de novo synthesis to templatedassembly (FIG. 1). In yeast, Golgi biogenesis is linked tothe organization and function of the ER, since it formsde novo at ER exit sites
71,72(FIG. 5a). There is a mounting
body of evidence to support this idea, most importantly the imaging of de novo Golgi formation by the Glick andNakano laboratories73,74.
A further example of de novo Golgi formation canbe found in the protozoan Trypanosoma brucei, whichhas a single-copy Golgi apparatus that lies adjacent
to the basal body75 (FIG. 5b). T. brucei build a new Golgide novo adjacent to the basal body by a combinationof de novo synthesis and transfer of material from theold copy 75–77. Simultaneously, the single ER exit siteis also duplicated, which suggests these two eventsare co-ordinately regulated, or reflect a single process.A caveat is that it is difficult to discriminate between truede novo assembly and templated growth that requiresthe original copy of the Golgi apparatus. Although littleis known about the molecular details of these processes,recent work suggests an intriguing link to the centrin fam-ily of calcium-binding proteins; centrins are associatedwith microtubule organizing centres in many organisms,including trypanosomes. T. brucei has two centrins. Thefirst of these is exclusively localized to the basal body andrequired for its duplication, whereas the other, centrin-2,is associated with a novel structure adjacent to the Golgiapparatus and is required for Golgi duplication (FIG. 5c)
78.As discussed elsewhere, de novo synthesis is not the
only way to build a new Golgi apparatus (FIG. 1), andother protozoa use a radically different mechanism.Toxoplasma gondii is an obligate intracellular eukaryoticparasite of medical significance that also serves asa good model system for the study of membrane-trafficking processes and organelle function79. In thisorganism, the Golgi apparatus is a single-copy organellethat grows by a lateral extension process, and then
undergoes medial fission during cell division, sothat each daughter cell obtains a functional Golgi80.A similar mechanism has also been seen in otherprotozoa such as Trichomonas81.
In mammals it is different again. Throughout S phase,when cells grow, new material is continuously deliveredto the pre-existing Golgi. Recent findings indicate thatthis is a size control mechanism that is tightly coupledto the synthesis and delivery of new Golgi enzymes fromthe ER62. As in yeast, the delivery of material from the ERis therefore crucial for the establishment and maintenanceof the Golgi, and it is not surprising that Golgi in animalcells also have an intimate relationship with ER exit sites.
Figure 5 | Golgi biogenesis and inheritance in yeast and protozoa. a | Top panels show
images captured from a movie of living Pichia pastoris yeast cells expressing fluorescently
labelled markers for endoplasmic reticulum (ER) exit sites (Sec13–GFP, in green) and the
Golgi apparatus (Sec7–DsRed, in red). Note the de novo appearance of two ER exit sites
closely followed by new Golgi structures in close proximity to them (arrowheads). The
schematic view underneath illustrates the de novo formation of ER exit sites (green) and
Golgi structures (red) in the mother cell and in the newly forming bud that will ultimately
form the daughter cell. Golgi inheritance occurs by de novo synthesis in the bud. Late
Golgi elements are transported into the bud separately from other Golgi components (not
shown).b | Protozoa such as Trypanosoma brucei typically have a single-copy Golgi thatlies adjacent to the basal body (left). Trypanosoma brucei carries out de novo Golgi
assembly adjacent to the existing Golgi apparatus. Quite how this is brought about is
currently unclear, but recent work suggests that the centrin-2 protein may be required to
guide new Golgi synthesis. c | Centrin-2 (green) is located in pools at the Golgi (indicated
by red fluorescent Golgi marker, closed arrow heads) and basal body (arrow). At an early
time point, a pool of centrin-2 is present next to the existing Golgi (left), and this marks the
site at which the new Golgi forms (right). Centrins may therefore form a template structure
that is capable of initiating de novo biogenesis of the Golgi apparatus. Part a reproduced
with permission from Nature Cell Biology REF. 72© (2002) Macmillan Publishers Ltd.
R E V I E W S
434 | JUNE 2007 | VOLUME 8 www.nature.com/reviews/molcellbio

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 7/11
G1 Telophase Anaphase
G2 Prophase Metaphase
Golgiinheritance
Mitotic Golgifragments
Nucleus
Golgi ribbon
Golgi
biogenesis ER
The difference is whether or not a pre-existing Golgi isneeded to template Golgi formation as in protozoa, or if it can occur de novo as in yeast.
Strategies for Golgi inheritance
The mechanisms of Golgi inheritance reflect the differentmodes of biogenesis discussed above. As yeast producea functional Golgi de novo it is not strictly essential toinherit old Golgi. However, this highlights the impor-tance of ER inheritance, as without a properly functionalcomplement of ER the cell will be compromised in itsability to produce Golgi de novo82. Budding yeast arealso capable of targeting Golgi elements to the bud usingan actin–myosin-dependent partitioning mechanism83,so in reality a combination of inheritance and de novo synthesis might be the most efficient option. Some pro-tozoa split their duplicated Golgi down the middle, by a binary fission mechanism that reflects their mode of cell division80,81.
In mammalian cells, it is clear that the Golgi breaksdown into many small vesicles (FIG. 6). However, there aretwo possibilities to explain what happens next, and the
debate continues84. The initial events as cells enter mitosisare not disputed. This includes the release of many periph-eral membrane proteins into the cytoplasm and break-down of the ribbon structure, followed by the unstackingand fragmentation of the individual cisternae into small
vesicles. These Golgi vesicles might then either remain asdispersed independent entities throughout mitosis thatpartition according to a stochastic mechanism52,85–89, ordeliver their content back to the ER90–92. Once deliveredback to the ER, Golgi proteins would be trapped andpartition together with the ER, as ER to Golgi traffickingis blocked at an early stage for the duration of mitosis93,94.There is evidence for both ideas, and it might depend on
the content of the vesicle, as structural components of theGolgi do not seem to recycle to the ER, whereas Golgienzymes and cargo can to some extent94–96.
So, despite the controversy, is the mechanism of inheritance in mammalian cells so different from theother organisms discussed? Ribbon splitting is actually analogous to the binary fission mechanism used by pro-tozoa, and might therefore involve the same underlyingmachinery. Rebuilding a Golgi during mitotic exit hasmany similarities with de novo Golgi biogenesis in yeast.It is clearly important to re-establish transport from theER to form a new Golgi, which suggests that the delivery of ER-derived material is necessary. However, the largepool of vesicles in the cytoplasm derived from the oldGolgi needs to assemble together with this to build newfunctional Golgi in the two daughter cells. The questionis do these vesicles reflect a template for Golgi assembly,or are they simply a supply of components directed by theER-derived material to incorporate into a new Golgi? Inboth cases, Golgi proteins must have self-assembly prop-erties and form structures specifying cisternal shape and
size, and stack organization.
Regulation of Golgi inheritance
The transitions between the different phases of the cellcycle are controlled by protein phosphorylation and ubiquit-ylation. It is therefore not surprising that the inheritanceof the Golgi apparatus is controlled by these samemechanisms. Although protein phosphorylation has his-torically been the focus of attention, more recent evidencepoints to the importance of reversible ubiquitylationin controlling the assembly state of the Golgi.
Phosphorylation. Most information about phosphor-ylation is known for mammalian cells, in which it hasbeen shown that, similar to the NE, CDK1 drives thebreakdown of the Golgi apparatus97–99. However, CDK1does not function alone and a number of other proteinkinases of the polo and MAPK family have also beenimplicated (TABLE I). These kinases phosphorylate multi-ple proteins that are required for trafficking and Golgistructure (TABLE I), and as a consequence the Golgi thendisassembles. This disassembly is in part mediated by the action of the COPI vesicle formation pathway, whichconsumes Golgi cisternae into vesicles97, 98.
As mentioned above, ER to Golgi trafficking is blockedin mitosis, which is likely to have an important role in thereorganization of the Golgi, as under these conditions
the Golgi will no longer be supplied with new components.How this transport block is achieved is still not completely understood. Multiple components of the ER to Golgitransport machinery, including the GTPase RAB1 andGM130, become phosphorylated in mitosis66,99–101, butno single crucial component has been identified. GM130phosphorylation alone is unlikely to be the critical event,as when it is depleted from interphase cells the efficiency of ER to Golgi transport is only slightly reduced102. Ratherthan a single key substrate, this supports a model in whichphosphorylation of multiple components reduces the effi-ciency of ER to Golgi transport. Other aspects of Golgifragmentation, such as the breakdown of the Golgi ribbon,
Figure 6 | Inheritance of the Golgi in mammalian cells. The interphase Golgi ribbon
is composed of stacked cisternae that are linked by lateral tubular connections to form
the Golgi ribbon, which is often located in the perinuclear region of the cell. During
prophase, the lateral connections are lost as the Golgi ribbon is converted into individual
stacks that remain close to the nucleus. From prophase through to metaphase, stacking
of Golgi cisternae is lost, and the cisternae are converted into small ~50–70 nm vesicles,
and larger vesicular and tubular elements. These mitotic Golgi fragments function as the
units of partitioning, and then become dispersed throughout the cytoplasm. An
alternative view is that the Golgi fragments fuse with the endoplasmic reticulum (ER),which merges these two compartments and Golgi proteins are partitioned with the ER.
In telophase, the Golgi fragments fuse with each other to initiate the reformation of new
Golgi stacks that ultimately connect to form a Golgi ribbon in each daughter cell.
R E V I E W S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 435

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 8/11
Golgin-84
A member of a diverse family
of coiled-coil proteins that have
been implicated in vesicle
trafficking and shaping the
Golgi.
GRASP
(Golgi reassembly stacking
protein). A family of proteins
identified in an in vitro
biochemical screen for Golgi
stack formation.
Polo-like kinase 1
A serine or threonine kinase
that is required for centrosome
and spindle function in mitosis
and cytokinesis.
Anaphase promoting
complex/cyclosome
A ubiquitin ligase that is
required for progression
through mitosis.
might require additional factors. For example, what splitsmammalian-cell Golgi ribbons into single stacks or aprotozoan Golgi down the middle, and is there an activity to promote this? Alternatively, this stack splitting might bea passive process explained by phosphorylation of proteinssuch as GM130 and golgin-84, which have been implicatedin maintaining the ribbon of Golgi stacks in interphasecells68,103. Similarly, loss of the cisternal architecture of theGolgi is thought to involve phosphorylation of compo-nents linking Golgi cisternae together, and factors suchas Golgi reassembly stacking proteins (GRASPs), which aretargets for CDK1, have been proposed to contribute tothis process66,104,105. Interestingly, the processes of Golgiribbon and stack formation might be linked, as GRASP65is an interaction partner of GM130 (REF. 106). On the basisof a number of observations, it has been suggested thatGolgi-stack formation is a non-productive form of vesicletethering that does not lead to membrane fusion, butrather holds Golgi cisternae together66,107.
The roles of polo-like kinase-1 (PLK1) and MEK1(MAPK and ERK kinase-1) in Golgi fragmentation areunclear, and they do not appear to be essential for Golgibreakdown. However, both have been implicated together
with GRASP55 and GRASP65 in an organelle-breakdowncheckpoint105,108–110. Just as unattached chromosomes sig-nal to prevent cell-cycle progression, so too might the Golgithrough GRASPs, MEK1 and PLK1105,108–111. Some evi-dence also exists for this in yeast, in which Grh1 (the yeasthomologue of GRASP65) was found in a screen for mitoticcheckpoint deficiency 112. However, this is contro versial,and other evidence shows that Grh1 is a Golgi protein that,as in mammals, has a function in ER to Golgi transport113.Intriguingly, recent evidence indicates there might be a linkbetween the GRASP checkpoint and centrin-2 function inmitosis in animal cells111, which suggests that centrins may have a general function in Golgi inheritance.
Ubiquitylation. In addition to phosphorylation, othertypes of regulatory modification are important for con-trolling the passage through mitosis, and perhaps themost important of these is ubiquitylation114. In recentyears, a number of lines of evidence have pointed to therole of ubiquitylation in remodelling the ER and Golgiduring mitosis. This has mainly emerged from studiesof the ubiquitin-dependent chaperone p97 (also knownas valosin-containing protein (VCP)) and its interactingproteins p37, p47 and VCIP135 (VCP/p47 complexinteracting protein-1)115–121. Interestingly, p47 is a CDK1substrate, and this regulation appears to couple these twosystems for controlling the Golgi, as mutant forms of p47that can no longer be phosphorylated by CDK1 preventcomplete fragmentation of the Golgi during mitosis117.
Although a major function of ubiquitylation is to targetproteins for degradation by the proteasome, it is nowappreciated that it also has regulatory functions, and evi-dence is emerging that this regulatory role is importantfor Golgi inheritance. The most direct evidence for thiscomes from a study showing that the p97–p47 cofactorVCIP135 is a deubiquitylating enzyme that is required forreformation of the Golgi complex in an in vitro assay. This
suggests that ubiquitylation — through cycles of addingand removing ubiquitin to substrates — is necessary for Golgi reassembly, rather than ubiquitin-dependentproteolysis120. A number of outstanding questionsremain about the role of ubiquitylation: the first concernsthe identity of the ubiquitin ligase that is responsiblefor carrying out ubiquitin conjugation in organelleinheritance, and the second concerns the identity of the proteins targeted by this system. An obvious answerto the first question would be the anaphase promoting
complex/cyclosome, as this is a key mitotic regulatorthat is capable of modifying specific sets of substratesrequired for entry into and exit from mitosis114,122.
Table 1 | Kinases and substrates implicated in Golgi inheritance in mammals
Kinases Interactionpartners
Substrates Proposed function of substrate Refs
CDK1 Cyclin B GM130 Golgi membrane and vesicle tethering 99,101
GRASP65 Membrane tethering and cisternal stacking 105,129–131
RAB1 Golgi membrane and vesicle tethering 100
p47 Membrane fusion 117
NIR2 Phospholipid transfer 126
PLK1 GRASP65 GRASP65 Membrane tethering and cisternal stacking 105,109,129–131
RAB1 RAB1 Golgi membrane and vesicle tethering 105
PLK3 GiantinMEK1ERK2
Unknown Not applicable 132,133
MEK1 PLK3 Unknown Not applicable 133–136
ERK1/2 PLK3 GRASP55 Cisternal stacking 133,137
ERK1c Unknown Unknown Not applicable 138
Unknown Unknown Golgin-84 Golgi-membrane and vesicle tethering 103
CDK1, cyclin-dependent kinase-1; ERK1/1c/2, extracellular regulated kinase-1/1c/2; GM130, Golgi matrix protein of 130 kDa;
GRASP55, Golgi reassembly and stacking protein of 55 kDa; GRASP65, Golgi reassembly and stacking protein of 65kDa;MAP kinase, mitogen-activated protein kinase; MEK1, MAP kinase/ERK kinase kinase-1; NIR2, PYK2 amino-terminal domaininteracting receptor-2; PLK1/3, polo-like kinase-1/3.
R E V I E W S
436 | JUNE 2007 | VOLUME 8 www.nature.com/reviews/molcellbio

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 9/11
Cytokinesis
The process of cytoplasmic
division in animal cells.
Answers to both these questions will be important fordetermining how direct the role of ubiquitin modifica-tion is, and how exactly Golgi inheritance is regulatedby ubiquitylation.
Outlook and future directions
Despite recent advances, many aspects of the mechanismsneeded to build an organelle remain mysterious. Howand why do Golgi membranes adopt their characteristiccisternal shape whereas most other cellular membranesadopt vesicular or tubular morphologies? The recentidentification of factors promoting ER tubulation showsthis is surely protein mediated, but we still do not under-stand what these factors are or why this morphology isimportant.
A second issue concerns the difference between self-assembly and de novo assembly. In general, membraneorganelles self-assemble to form unique structures, thatis, the protein and lipid components alone define the finalform of the organelle123. Some yeasts provide an exampleof de novo Golgi formation, in which components self-
assemble without a template to form Golgi cisternae.However, in mammalian cells and some protozoa,although these components might also self-assemble, it isunclear whether they can do so without the assistance of a template structure. This might reflect the different scaleof the Golgi in yeast and mammals. In mammals, someform of templating might be needed to promote assembly,which would otherwise be too slow or inefficient duringthe 15–30 minute time window that is available as mam-malian cells exit mitosis. Questions remain over the fateof Golgi proteins in mitosis, and this will require furtherinvestigation of multiple classes of Golgi proteins to buildup a full picture of what happens to them. A related ques-tion is why does secretion stop in some organisms duringmitosis but not in others? It is obvious that unicellularorganisms such as yeasts might have a selective advantage
if they can continue to secrete and grow throughout thecell cycle; however, it is not clear why mammalian cellsstop secreting during cell division.
In recent years it has become apparent that organellefunction during mitosis is also important for the cell-division process itself. Asymmetrical cell divisions arerequired for determining cell fates during development,and it is now known that this is underpinned in part by asymmetry in the behaviour of organelles required forcell function (reviewed in REF. 124). It has been foundrecently that asymmetry in the activity of RAB11 associ-ated with recycling endosomes reflects altered traffickingof molecules that specify asymmetrical cell fate125.However, nothing is known about other organelles in thiscontext, and there might also be differences in moleculesthat are associated with the ER and Golgi. Signalling mole-cules involved in determining cell fate have to be syn-thesized and delivered to the cell surface, and one mightexpect that this is also a point of regulation involving theER and Golgi. Another potential mechanism of regula-tion is the release from organelles of components that
function in the division process itself. During mitosis,the lipid transfer protein NIR2 is phosphorylated by CDK1 and released from Golgi membranes, and thentranslocates to the cleavage furrow, where it contributesto the regulation of cytokinesis
126,127.Last, without a full list of the molecular components
needed to build an organelle and a better understand-ing of the rules governing organelle assembly, many of the ideas discussed above remain speculative. In recentyears, much progress has been made in achieving thesegoals, yet much remains to be done and there might bemany surprises to come. The organelles of the secre-tory pathway have been known for over 100 years, butexplaining how they are constructed and function willremain a central problem in cell biology for some timeto come.
1. Howell, G. J., Holloway, Z. G., Cobbold, C.,
Monaco, A. P. & Ponnambalam, S. Cell biology of
membrane trafficking in human disease. Int. Rev.
Cytol. 252, 1–69 (2006).
2. Olkkonen, V. M. & Ikonen, E. When intracellular
logistics fails — genetic defects in membrane
trafficking. J. Cell Sci. 119, 5031–5045 (2006).3. Delattre, M. & Gonczy, P. The arithmetic of centrosome
biogenesis. J. Cell Sci. 117, 1619–1630 (2004).
4. Szollosi, D., Calarco, P. & Donahue, R. P. Absence of
centrioles in the first and second meiotic spindles of
mouse oocytes. J. Cell Sci. 11, 521–541 (1972).
5. Du, Y., Ferro-Novick, S. & Novick, P. Dynamics and
inheritance of the endoplasmic reticulum. J. Cell Sci.117, 2871–2878 (2004).
6. Vedrenne, C. & Hauri, H. P. Morphogenesis of the
endoplasmic reticulum: beyond active membrane
expansion. Traffic 7, 639–646 (2006).
7. Voeltz, G. K., Prinz, W. A., Shibata, Y., Rist, J. M. &
Rapoport, T. A. A class of membrane proteins shaping
the tubular endoplasmic reticulum. Cell 124,
573–586 (2006).
8. De Craene, J. O. et al. Rtn1p is involved in structuring
the cortical endoplasmic reticulum. Mol. Biol. Cell 17,
3009–3020 (2006).
References 7 and 8 are key studies that identify
the reticulon family of proteins as factors that
shape the ER in yeast and animal cells.9. Calero, M., Whittaker, G. R. & Collins, R. N. Yop1p,
the yeast homolog of the polyposis locus protein 1,
interacts with Yip1p and negatively regulates cell
growth. J. Biol. Chem. 276, 12100–12112 (2001).
10. Sivars, U., Aivazian, D. & Pfeffer, S. R. Yip3 catalyses
the dissociation of endosomal Rab–GDI complexes.
Nature 425, 856–859 (2003).
11. Martincic, I., Peralta, M. E. & Ngsee, J. K. Isolation
and characterization of a dual prenylated Rab and
VAMP2 receptor. J. Biol. Chem. 272, 26991–26998
(1997).12. Terasaki, M., Chen, L. B. & Fujiwara, K. Microtubules
and the endoplasmic reticulum are highly
interdependent structures. J. Cell Biol. 103,
1557–1568 (1986).
13. Lee, C. & Chen, L. B. Dynamic behavior of
endoplasmic reticulum in living cells. Cell 54, 37–46
(1988).14. Dabora, S. L. & Sheetz, M. P. The microtubule-
dependent formation of a tubulovesicular network
with characteristics of the ER from cultured cell
extracts. Cell 54, 27–35 (1988).
15. Vale, R. D. & Hotani, H. Formation of membrane
networks in vitro by kinesin-driven microtubule
movement. J. Cell Biol. 107, 2233–2241
(1988).
16. Allan, V. & Vale, R. Movement of membrane tubules
along microtubules in vitro: evidence for specialised
sites of motor attachment. J. Cell Sci. 107,
1885–1897 (1994).
17. Toyoshima, I., Yu, H., Steuer, E. R. & Sheetz, M. P.
Kinectin, a major kinesin-binding protein on ER. J. Cell
Biol. 118, 1121–1131 (1992).
18. Kumar, J., Yu, H. & Sheetz, M. P. Kinectin, an essential
anchor for kinesin-driven vesicle motility. Science 267,
1834–1837 (1995).
19. Vedrenne, C., Klopfenstein, D. R. & Hauri, H. P.
Phosphorylation controls CLIMP-63-mediated
anchoring of the endoplasmic reticulum to
microtubules. Mol. Biol. Cell 16, 1928–1937
(2005).
20. Huffaker, T. C., Thomas, J. H. & Botstein, D. Diverse
effects of β-tubulin mutations on microtubule
formation and function. J. Cell Biol. 106, 1997–2010
(1988).
21. Jacobs, C. W., Adams, A. E., Szaniszlo, P. J. & Pringle,
J. R. Functions of microtubules in the Saccharomyces
cerevisiae cell cycle. J. Cell Biol. 107, 1409–1426
(1988).
22. Preuss, D. et al. Structure of the yeast endoplasmicreticulum: localization of ER proteins using
immunofluorescence and immunoelectron microscopy.
Yeast 7, 891–911 (1991).23. Patel, S. K., Indig, F. E., Olivieri, N., Levine, N. D. &
Latterich, M. Organelle membrane fusion: a novel
function for the syntaxin homolog Ufe1p in ER
membrane fusion. Cell 92, 611–620 (1998).
24. Prinz, W. A. et al. Mutants affecting the structure of
the cortical endoplasmic reticulum in
Saccharomyces cerevisiae. J. Cell Biol. 150,
461–474 (2000).
25. Estrada, P. et al. Myo4p and She3p are required for
cortical ER inheritance in Saccharomyces cerevisiae.
J. Cell Biol. 163, 1255–1266 (2003).
Identification of myosin as the motor driving the
active inheritance of cortical ER in yeast. See
reference 83 for related evidence for the role of
myosin in Golgi inheritance.
R E V I E W S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 437

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 10/11
26. Estrada de Martin, P., Du, Y., Novick, P. &
Ferro-Novick, S. Ice2p is important for the distribution
and structure of the cortical ER network in
Saccharomyces cerevisiae. J. Cell Sci. 118, 65–77
(2005).
27. Du, Y., Pypaert, M., Novick, P. & Ferro-Novick, S.
Aux1p/Swa2p is required for cortical endoplasmic
reticulum inheritance in Saccharomyces cerevisiae.
Mol. Biol. Cell 12, 2614–2628 (2001).28. Xiao, J., Kim, L. S. & Graham, T. R. Dissection of
Swa2p/auxilin domain requirements for
cochaperoning Hsp70 clathrin-uncoating activityin vivo. Mol. Biol. Cell 17, 3281–3290 (2006).
29. Denic, V., Quan, E. M. & Weissman, J. S. A luminal
surveillance complex that selects misfolded
glycoproteins for ER-associated degradation. Cell 126,
349–359 (2006).
30. Rape, M. et al. Mobilization of processed, membrane-
tethered SPT23 transcription factor by CDC48(UFD1/
NPL4), a ubiquitin-selective chaperone. Cell 107,
667–677 (2001).
31. Latterich, M. & Schekman, R. The karyogamy gene
KAR2 and novel proteins are required for
ER-membrane fusion. Cell 78, 87–98 (1994).
32. Latterich, M., Frohlich, K. U. & Schekman, R.
Membrane fusion and the cell cycle: Cdc48p
participates in the fusion of ER membranes. Cell 82,
885–893 (1995).
33. TerBush, D. R., Maurice, T., Roth, D. & Novick, P. The
exocyst is a multiprotein complex required for
exocytosis in Saccharomyces cerevisiae. EMBO J. 15,
6483–6494 (1996).
34. Guo, W., Roth, D., Walch-Solimena, C. & Novick, P.
The exocyst is an effector for Sec4p, targeting
secretory vesicles to sites of exocytosis. EMBO J. 18,
1071–1080 (1999).
35. Wiederkehr, A., Du, Y., Pypaert, M., Ferro-Novick, S. &
Novick, P. Sec3p is needed for the spatial regulation of
secretion and for the inheritance of the cortical
endoplasmic reticulum. Mol. Biol. Cell 14,
4770–4782 (2003).
References 35 and 36 are important papers
demonstrating that the exocyst is a spatial
landmark for secretion and ER inheritance.36. Finger, F. P., Hughes, T. E. & Novick, P. Sec3p is a
spatial landmark for polarized secretion in budding
yeast. Cell 92, 559–571 (1998).37. Finger, F. P. & Novick, P. Sec3p is involved in secretion
and morphogenesis in Saccharomyces cerevisiae.
Mol. Biol. Cell 8, 647–662 (1997).38. Toikkanen, J. H., Miller, K. J., Soderlund, H., Jantti, J.
& Keranen, S. The β subunit of the Sec61p
endoplasmic reticulum translocon interacts with theexocyst complex in Saccharomyces cerevisiae. J. Biol.
Chem. 278, 20946–20953 (2003).
39. Pruyne, D. & Bretscher, A. Polarization of cell growth
in yeast. I. Establishment and maintenance of polarity
states. J. Cell Sci. 113, 365–375 (2000).
40. Pruyne, D. & Bretscher, A. Polarization of cell growth
in yeast. J. Cell Sci. 113, 571–585 (2000).
41. Wedlich-Soldner, R., Wai, S. C., Schmidt, T. & Li, R.
Robust cell polarity is a dynamic state established by
coupling transport and GTPase signaling. J. Cell Biol.
166, 889–900 (2004).42. Zhang, X. et al. Cdc42 interacts with the exocyst and
regulates polarized secretion. J. Biol. Chem. 276,
46745–46750 (2001).
43. Padmashree, C. G. & Surana, U. Cdc28–Clb mitotic
kinase negatively regulates bud site assembly in the
budding yeast. J. Cell Sci. 114, 207–218 (2001).
44. Yeong, F. M., Lim, H. H., Padmashree, C. G. &
Surana, U. Exit from mitosis in budding yeast:
biphasic inactivation of the Cdc28–Clb2 mitotickinase and the role of Cdc20. Mol. Cell 5, 501–511
(2000).
45. Gulli, M. P. et al. Phosphorylation of the Cdc42
exchange factor Cdc24 by the PAK-like kinase Cla4
may regulate polarized growth in yeast. Mol. Cell 6,
1155–1167 (2000).
46. Du, Y., Walker, L., Novick, P. & Ferro-Novick, S. Ptc1p
regulates cortical ER inheritance via Slt2p. EMBO J.
25, 4413–4422 (2006).
47. Harrison, J. C., Bardes, E. S., Ohya, Y. & Lew, D. J.
A role for the Pkc1p/Mpk1p kinase cascade in the
morphogenesis checkpoint. Nature Cell Biol. 3,
417–420 (2001).
48. Levin, D. E. Cell wall integrity signaling in
Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev.
69, 262–291 (2005).
49. Yang, L., Guan, T. & Gerace, L. Integral membrane
proteins of the nuclear envelope are dispersed
throughout the endoplasmic reticulum during mitosis.
J. Cell Biol. 137, 1199–1210 (1997).50. Burke, B. & Ellenberg, J. Remodelling the walls of the
nucleus. Nature Rev. Mol. Cell Biol. 3, 487–497
(2002).
51. Ellenberg, J. et al. Nuclear membrane dynamics and
reassembly in living cells: targeting of an inner nuclear
membrane protein in interphase and mitosis. J. Cell
Biol. 138, 1193–1206 (1997).52. Axelsson, M. A. & Warren, G. Rapid, endoplasmic
reticulum-independent diffusion of the mitotic Golgi
haze. Mol. Biol. Cell 15, 1843–1852 (2004).53. Salina, D. et al. Cytoplasmic dynein as a facilitator of
nuclear envelope breakdown. Cell 108, 97–107
(2002).
54. Beaudouin, J., Gerlich, D., Daigle, N., Eils, R. &
Ellenberg, J. Nuclear envelope breakdown proceeds
by microtubule-induced tearing of the lamina. Cell
108, 83–96 (2002).
References 53 and 54 report a major finding that
microtubules actively promote the peeling of the
NE and ER away from chromatin during mitotic
entry.55. Peter, M., Nakagawa, J., Doree, M., Labbe, J. C. &
Nigg, E. A. In vitro disassembly of the nuclear lamina
and M phase-specific phosphorylation of lamins by
Cdc2 kinase. Cell 61, 591–602 (1990).
An early study showing that Cdk1 controls the
assembly status of the nuclear lamina, and
therefore the structure of a membrane organelle.56. Herpers, B. & Rabouille, C. mRNA localization and ER-
based protein sorting mechanisms dictate the use of
transitional endoplasmic reticulum–golgi units
involved in gurken transport in Drosophila oocytes.
Mol. Biol. Cell 15, 5306–5317 (2004).
57. Allan, V. J., Thompson, H. M. & McNiven, M. A.
Motoring around the Golgi. Nature Cell Biol. 4,
E236–E242 (2002).
58. Barr, F. A. & Egerer, J. Golgi positioning: are we
looking at the right MAP? J. Cell Biol. 168, 993–998
(2005).
59. Shorter, J. & Warren, G. Golgi architecture and
inheritance. Annu. Rev. Cell Dev. Biol. 18, 379–420
(2002).
60. Gillingham, A. K. & Munro, S. Long coiled-coil proteins
and membrane traffic. Biochim. Biophys. Acta. 1641,
71–85 (2003).61. Short, B., Haas, A. & Barr, F. A. Golgins and GTPases,
giving identity and structure to the Golgi apparatus.
Biochim. Biophys. Acta 1744, 383–395 (2005).
62. Guo, Y. & Linstedt, A. D. COPII–Golgi protein
interactions regulate COPII coat assembly and Golgi
size. J. Cell Biol. 174, 53–63 (2006).63. Cluett, E. B. & Brown, W. J. Adhesion of Golgi
cisternae by proteinaceous interactions: intercisternal
bridges as putative adhesive structures. J. Cell Sci.
103, 773–784 (1992).64. Slusarewicz, P., Nilsson, T., Hui, N., Watson, R. &
Warren, G. Isolation of a matrix that binds medial
Golgi enzymes. J. Cell Biol. 124, 405–413 (1994).
65. Nakamura, N. et al. Characterization of a cis-Golgi
matrix protein, GM130. J. Cell Biol. 131, 1715–1726
(1995).
66. Barr, F. A., Puype, M., Vandekerckhove, J. & Warren, G.
GRASP65, a protein involved in the stacking of Golgi
cisternae. Cell 91, 253–262 (1997).
67. Seemann, J., Jokitalo, E., Pypaert, M. & Warren, G.
Matrix proteins can generate the higher order
architecture of the Golgi apparatus. Nature 407,
1022–1026 (2000).
68. Puthenveedu, M. A., Bachert, C., Puri, S., Lanni, F. &
Linstedt, A. D. GM130 and GRASP65-dependent
lateral cisternal fusion allows uniform Golgi-enzymedistribution. Nature Cell Biol. 8, 238–248 (2006).
69. Vasile, E., Perez, T., Nakamura, N. & Krieger, M.
Structural integrity of the Golgi is temperature
sensitive in conditional-lethal mutants with no
detectable GM130. Traffic 4, 254–272 (2003).70. Scaffidi, P. & Misteli, T. Lamin A dependent nuclear
defects in human aging. Science 312, 1059–1063
(2006).
71. Rossanese, O. W. et al. Golgi structure correlates with
transitional endoplasmic reticulum organization in
Pichia pastoris and Saccharomyces cerevisiae. J. Cell
Biol. 145, 69–81 (1999).72. Bevis, B. J., Hammond, A. T., Reinke, C. A. & Glick, B. S.
De novo formation of transitional ER sites and Golgi
structures in Pichia pastoris. Nature Cell Biol. 4,
750–756 (2002).
73. Losev, E. et al. Golgi maturation visualized in living
yeast. Nature 441, 1002–1006 (2006).
74. Matsuura-Tokita, K., Takeuchi, M., Ichihara, A.,
Mikuriya, K. & Nakano, A. Live imaging of yeast Golgi
cisternal maturation. Nature 441, 1007–1010
(2006).
References 71–74 are a series of landmark papers
that identify a role for ER exit sites in directing the
formation and inheritance of new Golgi stacks in
yeast. This work paved the way for the
re-evaluation of the importance of the ER in Golgi
biogenesis in other organisms.75. Hartmann, J. et al. Golgi and centrosome cycles in
Toxoplasma gondii . Mol. Biochem. Parasitol. 145,125–127 (2006).
76. He, C. Y. et al. Golgi duplication in Trypanosoma
brucei . J. Cell Biol. 165, 313–321 (2004).
77. Ho, H. H., He, C. Y., de Graffenried, C. L., Murrells, L. J.
& Warren, G. Ordered assembly of the duplicating
Golgi in Trypanosoma brucei . Proc. Natl Acad. Sci.
USA 103, 7676–7681 (2006).
Hints at underlying mechanistic similarities
between centrosome duplication and Golgi
biogenesis in these organisms.
78. He, C. Y., Pypaert, M. & Warren, G. Golgi duplication
in Trypanosoma brucei requires Centrin2. Science
310, 1196–1198 (2005).
Identification of centrin-2 as a template for Golgi
biogenesis in trypanosomes.79. Joiner, K. A. & Roos, D. S. Secretory traffic in the
eukaryotic parasite Toxoplasma gondii : less is more.
J. Cell Biol. 157, 557–563 (2002).80. Pelletier, L. et al. Golgi biogenesis in Toxoplasma
gondii . Nature 418, 548–552 (2002).
81. Benchimol, M., Ribeiro, K. C., Mariante, R. M. &
Alderete, J. F. Structure and division of the Golgi
complex in Trichomonas vaginalis and
Tritrichomonas foetus. Eur. J. Cell Biol. 80, 593–607
(2001).
A key study showing that, in some protozoa, the
Golgi undergoes lateral growth and then divides
into two by medial fission for equal partitioning
into the daughter cells.82. Reinke, C. A., Kozik, P. & Glick, B. S. Golgi inheritance
in small buds of Saccharomyces cerevisiae is linked to
endoplasmic reticulum inheritance. Proc. Natl Acad.
Sci. USA 101, 18018–18023 (2004).
83. Rossanese, O. W. et al. A role for actin, Cdc1p, and
Myo2p in the inheritance of late Golgi elements in
Saccharomyces cerevisiae. J. Cell Biol. 153, 47–-62
(2001).
84. Barr, F. A. Golgi inheritance: shaken but not stirred.
J. Cell Biol. 164, 955–958 (2004).85. Lucocq, J. M. & Warren, G. Fragmentation and
partitioning of the Golgi apparatus during mitosis inHeLa cells. EMBO J. 6, 3239–3246 (1987).
86. Jesch, S. A. & Linstedt, A. D. The Golgi and
endoplasmic reticulum remain independent during
mitosis in HeLa cells. Mol. Biol. Cell 9, 623–635
(1998).
87. Terasaki, M. Dynamics of the endoplasmic reticulum
and golgi apparatus during early sea urchin
development. Mol. Biol. Cell 11, 897–914 (2000).88. Jokitalo, E., Cabrera-Poch, N., Warren, G. &
Shima, D. T. Golgi clusters and vesicles mediate
mitotic inheritance independently of the endoplasmic
reticulum. J. Cell Biol. 154, 317–330 (2001).
89. Jesch, S. A., Mehta, A. J., Velliste, M., Murphy, R. F. &
Linstedt, A. D. Mitotic Golgi is in a dynamic
equilibrium between clustered and free vesicles
independent of the ER. Traffic 2, 873–884 (2001).
90. Thyberg, J. & Moskalewski, S. Reorganization of the
Golgi complex in association with mitosis:
redistribution of mannosidase II to the endoplasmic
reticulum and effects of brefeldin A. J. Submicrosc.Cytol. Pathol. 24, 495–508 (1992).
91. Zaal, K. J. et al. Golgi membranes are absorbed into
and reemerge from the ER during mitosis. Cell 99,
589–601 (1999).
92. Altan-Bonnet, N. et al. Golgi inheritance in
mammalian cells is mediated through endoplasmic
reticulum export activities. Mol. Biol. Cell 17,
990–1005 (2006).
References 91 and 92 provide evidence for the
recycling of Golgi proteins to the ER in mitosis.
93. Featherstone, C., Griffiths, G. & Warren, G. Newly
synthesized G protein of vesicular stomatitis virus is
not transported to the Golgi complex in mitotic cells.
J. Cell Biol. 101, 2036–2046 (1985).
94. Prescott, A. R. et al. Evidence for prebudding arrest of
ER export in animal cell mitosis and its role in
generating Golgi partitioning intermediates. Traffic 2,
321–335 (2001).
R E V I E W S
438 | JUNE 2007 | VOLUME 8 www.nature.com/reviews/molcellbio

8/6/2019 LSM1102_Inheritance and Bio Genesis of Organelles in the Secretory Pathway
http://slidepdf.com/reader/full/lsm1102inheritance-and-bio-genesis-of-organelles-in-the-secretory-pathway 11/11
95. Pecot, M. Y. & Malhotra, V. Golgi membranes remain
segregated from the endoplasmic reticulum during
mitosis in mammalian cells. Cell 116, 99–107
(2004).
A study providing evidence for discrete nature of
the ER and Golgi in mitosis. Also refer to references
85–89.
96. Seemann, J., Pypaert, M., Taguchi, T., Malsam, J. &
Warren, G. Partitioning of the matrix fraction of the
Golgi apparatus during mitosis in animal cells. Science
295, 848–851 (2002).
97. Misteli, T. & Warren, G. COP-coated vesicles areinvolved in the mitotic fragmentation of Golgi stacks in
a cell-free system. J. Cell Biol. 125, 269–282 (1994).
98. Misteli, T. & Warren, G. Mitotic disassembly of the
Golgi apparatus in vivo. J. Cell Sci. 108, 2715–2727
(1995).
99. Lowe, M. et al. Cdc2 kinase directly phosphorylates
the cis-Golgi matrix protein GM130 and is required
for Golgi fragmentation in mitosis. Cell 94, 783–793
(1998).
References 97–99 describe the key role of Cdk1 in
controlling the assembly state of the Golgi during
mitosis.
100. Bailly, E. et al. Phosphorylation of two small GTP-
binding proteins of the Rab family by p34cdc2. Nature
350, 715–718 (1991).
101. Lowe, M., Gonatas, N. K. & Warren, G. The mitotic
phosphorylation cycle of the cis-Golgi matrix protein
GM130. J. Cell Biol. 149, 341–356 (2000).102. Seemann, J., Jokitalo, E. J. & Warren, G. The role of
the tethering proteins p115 and GM130 in transport
through the Golgi apparatus in vivo. Mol. Biol. Cell 11,
635–645 (2000).
103. Diao, A., Rahman, D., Pappin, D. J., Lucocq, J. &
Lowe, M. The coiled-coil membrane protein golgin-84
is a novel rab effector required for Golgi ribbon
formation. J. Cell Biol. 160, 201–212 (2003).
104. Shorter, J. et al. GRASP55, a second mammalian
GRASP protein involved in the stacking of Golgi
cisternae in a cell-free system. EMBO J. 18,
4949–4960 (1999).
105. Preisinger, C. et al. Plk1 docking to GRASP65
phosphorylated by Cdk1 suggests a mechanism for
Golgi checkpoint signalling. EMBO J. 24, 753–765
(2005).
106. Barr, F. A., Nakamura, N. & Warren, G. Mapping the
interaction between GRASP65 and GM130,
components of a protein complex involved in the
stacking of Golgi cisternae. EMBO J. 17, 3258–3268
(1998).
107. Shorter, J. & Warren, G. A role for the vesicle tethering
protein, p115, in the post-mitotic stacking of reassembling Golgi cisternae in a cell-free system.
J. Cell Biol. 146, 57–70 (1999).
108. Sütterlin, C., Hsu, P., Mallabiabarrena, A. &
Malhotra, V. Fragmentation and dispersal of the
pericentriolar Golgi complex is required for entry into
mitosis in mammalian cells. Cell 109, 359–369
(2002).
109. Sütterlin, C. et al. Polo-like kinase is required for the
fragmentation of pericentriolar Golgi stacks during
mitosis. Proc. Natl Acad. Sci. USA 98, 9128–9132
(2001).
References 108–109 identified a link between
GRASP proteins and cell-cycle regulation in both
yeast and mammalian cells. This might take the
form of a checkpoint reading out the status of the
Golgi. Also refer to references 105, 110 and 111.
110. Feinstein, T. N. & Linstedt, A. D. Mitogen-activated
protein kinase-dependent Golgi unlinking occurs in G2
phase and promotes the G2/M cell cycle transition.
Mol. Biol. Cell 18, 594–604 (2007).
111. Sutterlin, C., Polishchuk, R., Pecot, M. & Malhotra, V.
The Golgi-associated protein GRASP65 regulates
spindle dynamics and is essential for cell division. Mol.
Biol. Cell 16, 3211–3222 (2005).
112. Norman, T. C. et al. Genetic selection of peptide
inhibitors of biological pathways. Science 285,
591–595 (1999).
113. Behnia, R., Barr, F. A., Flanagan, J. J., Barlowe, C. & Munro, S. The yeast orthologue of GRASP65 forms a
complex with a coiled-coil protein that contributes to
membrane traffic. J. Cell Biol. 176, 255–261 (2007).
114. Peters, J. M. The anaphase promoting complex/
cyclosome: a machine designed to destroy. Nature
Rev. Mol. Cell Biol. 7, 644–656 (2006).
115. Rabouille, C., Levine, T. P., Peters, J. M. & Warren, G.
An NSF--like ATPase, p97, and NSF mediate cisternal
regrowth from mitotic Golgi fragments. Cell 82,
905–914 (1995).
116. Kondo, H. et al. p47 is a cofactor for p97-mediated
membrane fusion. Nature 388, 75–78 (1997).
117. Uchiyama, K. et al. The localization and
phosphorylation of p47 are important for Golgi
disassembly–assembly during the cell cycle. J. Cell
Biol. 161, 1067–1079 (2003).
118. Uchiyama, K. et al. VCIP135, a novel essential factor
for p97/p47-mediated membrane fusion, is required
for Golgi and ER assembly in vivo. J. Cell Biol. 159,
855–866 (2002).
119. Kano, F. et al. NSF/SNAPs and p97/p47/VCIP135 are
sequentially required for cell cycle-dependent
reformation of the ER network. Genes Cells 10,
989–999 (2005).
120. Wang, Y., Satoh, A., Warren, G. & Meyer, H. H.
VCIP135 acts as a deubiquitinating enzyme during
p97–p47-mediated reassembly of mitotic Golgi
fragments. J. Cell Biol. 164, 973–978 (2004).121. Uchiyama, K. et al. p37 is a p97 adaptor required for
Golgi and ER biogenesis in interphase and at the end
of mitosis. Dev. Cell 11, 803–816 (2006).
References115–121 are an important series of
studies that suggest ubiquitylation and
deubiquitylation might have an important role in
controlling the assembly state of the Golgi during
mitosis in mammalian cells.
122. Fry, A. M. & Yamano, H. APC/C-mediated degradation in
early mitosis: how to avoid spindle assembly checkpoint
inhibition.Cell Cycle 5, 1487–1491 (2006).
123. Misteli, T. The concept of self-organization in cellular
architecture. J. Cell Biol. 155, 181–185 (2001).124. Betschinger, J. & Knoblich, J. A. Dare to be different:
asymmetric cell division in Drosophila, C. elegans and
vertebrates. Curr. Biol. 14, R674–R685 (2004).
125. Emery, G. et al. Asymmetric Rab 11 endosomes regulate
delta recycling and specify cell fate in the Drosophila
nervous system. Cell 122, 763–773 (2005).
A groundbreaking study showing that organelle
function in mitosis has a key role in determining the
outcome of cell division. In this case, whether cells
will adopt similar (symmetrical) or different
(asymmetrical) fates. See reference 124 for
background details on this topic.126. Litvak, V. et al. Mitotic phosphorylation of the
peripheral Golgi protein Nir2 by Cdk1 provides a
docking mechanism for Plk1 and affects cytokinesis
completion. Mol. Cell 14, 319–330 (2004).
127. Barr, F. A., Sillje, H. H. & Nigg, E. A. Polo-like kinases
and the orchestration of cell division. Nature Rev. Mol.
Cell Biol. 5, 429–440 (2004).
128. Lane, J. D. & Allan, V. J. Microtubule-based
endoplasmic reticulum motility in Xenopus laevis:
activation of membrane-associated kinesin during
development. Mol. Biol. Cell 10, 1909–1922
(1999).
129. Wang, Y., Seemann, J., Pypaert, M., Shorter, J. &
Warren, G. A direct role for GRASP65 as a mitotically
regulated Golgi stacking factor. EMBO J. 22, 3279–
3290 (2003).130. Lin, C. Y. et al. Peripheral Golgi protein GRASP65 is
a target of mitotic polo-like kinase (Plk) and Cdc2.
Proc. Natl Acad. Sci. USA 97, 12589–12594
(2000).131. Wang, Y., Satoh, A. & Warren, G. Mapping the
functional domains of the Golgi stacking factor
GRASP65. J. Biol. Chem. 280, 4921–4928 (2005).132. Ruan, Q. et al. Polo-like kinase 3 is Golgi localized and
involved in regulating Golgi fragmentation during the
cell cycle. Exp. Cell Res. 294, 51–59 (2004).
133. Xie, S. et al. MEK1-induced Golgi dynamics during cell
cycle progression is partly mediated by Polo-like
kinase-3. Oncogene 23, 3822–3829 (2004).
134. Acharya, U., Mallabiabarrena, A., Acharya, J. K. &
Malhotra, V. Signaling via mitogen-activated protein
kinase kinase (MEK1) is required for Golgi
fragmentation during mitosis. Cell 92, 183–192
(1998).
The first evidence that a kinase other than Cdk1
might contribute to the control of the Golgi during
mitosis.135. Colanzi, A., Deerinck, T. J., Ellisman, M. H. &
Malhotra, V. A specific activation of the mitogen-
activated protein kinase kinase 1 (MEK1) is required
for Golgi fragmentation during mitosis. J. Cell Biol.
149, 331–339 (2000).
136. Colanzi, A., Sutterlin, C. & Malhotra, V. RAF1-activated
MEK1 is found on the Golgi apparatus in late prophase
and is required for Golgi complex fragmentation in
mitosis. J. Cell Biol. 161, 27–32 (2003).
137. Jesch, S. A., Lewis, T. S., Ahn, N. G. & Linstedt, A. D.
Mitotic phosphorylation of Golgi reassembly stacking
protein 55 by mitogen-activated protein kinase ERK2.
Mol. Biol. Cell 12, 1811–1817 (2001).
138. Shaul, Y. D. & Seger, R. ERK1c regulates Golgi
fragmentation during mitosis. J. Cell Biol. 172,
885–897 (2006).
AcknowledgementsWe would like to thank G. Warren and C. He for providing the
images of dividing Toxoplasma gondii , V. Allan for images of the ER in animal cells, B. Glick for images of the Golgi in Pichia
pastorisand Y. Du and S.Ferro-Novick for images of the bud-
ding yeast ER.
Competing interests statementThe authors declare no competing financial interests.
DATABASESThe following terms in this article are linked online to:
Entrez Genome Project: http://www.ncbi.nlm.nih.gov/
entrez/query.fcgi?db=genomeprj
Toxoplasma gondii | Trypanosoma brucei
UniProtKB: http://ca.expasy.org/sprot
Aux1 | Cdc28 | Cdc48 | CKAP4 | Ice2 | PLK1 | Ufe1 | Yip1 | Yop1
Access to this links box is available online.
R E V I E W S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | JUNE 2007 | 439