long-term system b-cellprogenitors · proc. natl. acad. sci. usa vol. 92, pp. 1570-1574, february...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 92, pp. 1570-1574, February 1995Immunology
Long-term culture system for selective growth of humanB-cell progenitors
(cord blood/gene expression/interleukin 7/stem cell factor/murine stroma)
DAVID J. RAWLINGS*, SHIRLEY G. QuANt, ROBERTA M. KATO*, AND OWEN N. WiTrE**Department of Microbiology and Molecular Genetics, and tHoward Hughes Medical Institute, University of California, 5-748 MacDonald Research Laboratory,10833 Le Conte Avenue, Los Angeles, CA 90024-1662
Communicated by Max D. Cooper, University ofAlabama, Birmingham, AL, October 21, 1994
ABSTRACT We describe a simple reproducible system forenrichment and long-term culture of human B-cell progeni-tors. Enriched CD34- cord blood mononudear cells areseeded onto a murine stromal cell line to establish a biphasicculture system. These cultures are characterized by transientgrowth of myeloid cells followed by outgrowth of cells highlyenriched for early B-cell progenitors. Cultures consisting of>90%o early B-lineage cells [expressing CD10, CD19, CD38,and CD45 but lacking CD20, CD22, CD23, and surface IgM]are maintained for > 12 weeks without growth factor addition.Cells remain predominantly germ line at the immunoglobulinlocus and express only low levels of cytoplasmic ,u chain,terminal deoxynucleotidyltransferase, and recombination-activating gene 1 product. They are unresponsive to the pre-B-cell growth factors interleukin 7 or stem cell factor, or both,suggesting that growth support is provided by a cross-reactivemurine stromal cell factor. Cultured B-cell progenitors aregenerated in large numbers (>108 cells from a typical cordblood specimen) suitable for use in biochemical analysis andgene-transfer studies. This system should be useful for studyof normal and abnormal early human B-lymphopoiesis.
Antigen-independent early B-cell development (B-lymphopoi-esis) is characterized by an orderly expression of regulatorygenes, surface molecules, and gene rearrangements leading tothe generation of surface IgM+ (sIgM+), sIgD+ mature B cells.This process is controlled through cell-cell interactions andsoluble or cell-associated hematopoietic growth factors (1-3).B-lymphopoiesis occurs first in the fetal liver and bone marrowand continues after birth only in the bone marrow. The com-plexity of mammalian bone marrow precludes detailed in vivoanalysis of these developmental events and has been signifi-cantly advanced through the establishment of long term B-lymphoid culture systems (4-10). Murine lymphoid progenitorcultures maintain murine immunoglobulin-locus-unrear-ranged pro-B-cells that undergo limited differentiation. Mu-rine long-term bone marrow cultures support growth andspontaneous maturation of pre-B-cells into sIgM+ cells. How-ever, cultured cells from both systems can fully reconstituteB-cell function in immunodeficient animals (8, 9, 11).
Several culture models have been useful in phenotypic andmolecular analysis and in defining the growth factor require-ments of early human B-cell progenitors (12-19). In thesesystems, cultured B-cell progenitors survive for relatively shortperiods (<4 weeks), and cell expansion is limited (usually<10-fold). Isolation of highly purified human bone marrowstem or B-cell progenitor populations and use of growthfactors has been required to maintain optimal cell growth andexpansion. Studies of early human B-lymphopoiesis would besignificantly advanced by the development of long-term cul-ture models similar to those available using murine cells.
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
1570
Establishment and maintenance of murine long-term culturesis dependent upon an adherent stromal cell microenvironment(4-6). Cultured cells develop and proliferate in direct associationwith stromal cells. While growth factors including interleukin 7(IL-7) and stem cell factor (SCF) are important for early B-celllineage expansion, additional as-yet-unidentified stromal factorsprobably play a critical role in these processes (4, 5). HumanB-lymphopoiesis is also likely to require close association with astromal microenvironment. Human bone marrow stromal cellshave been used to support short-term growth and proliferation ofCD10+, sIgM- human B-lineage cells (12, 14-16, 19). Establish-ment of long-term B-lymphoid cultures using human stroma hasbeen limited in part by the need to repeatedly establish primarystromal cultures and problems associated with transformed lines(20, 21).
Recently, several murine stromal lines have allowed supportof human hematopoietic progenitor cells in long-term culturesand colony assays (22-24). Most notably, Baum et al. (24)established long-term cultures of Thy-1+, CD34+ cells onmurine stroma without additional growth factors. Those cellsremain capable of both myeloid and B-cell engraftment of in-tact human bone marrow implanted in severe combined im-munodeficient (SCID) mice. In this paper we describe adap-tation of these observations and conditions previously devel-oped for culture of murine B-cell progenitors to establish along-term culture system for selective growth of human B-cellprogenitors.
METHODSCord Blood Enrichment for CD34+ Cells, Culture Condi-
tions, and Flow Cytometry. Heparinized (10 units/ml) cordblood was obtained from umbilical and placental tissues sched-uled to be discarded and stored up to 24 hr at room temper-ature. Low-density mononuclear cells were collected by Ficoll/Hypaque separation (Pharmacia), and enrichment of CD34+cells was performed with a Celprate LC34 Biotin cell separa-tion column (CellPro, Bothell, WA) as recommended by themanufacturer. Briefly, cord mononuclear cells were stainedwith anti-CD34 biotin-conjugated antibody and passedthrough an avidin column. Collected absorbed cells were 40-80% CD34+ by flow cytometry and represented approximately0.5-1% of input cells. Sixty percent confluent S17 stromal cellmonolayers (25) were established 1-2 days prior to addition ofCD34+ cord mononuclear cells by plating 2.5 x 105 S17 cellsper 10-cm2 tissue culture dish (Falcon). CD34+-enriched cellswere plated at a density of 0.5-1 x 105 cells per ml onto theS17-coated dishes (10 ml) in RPMI 1640 medium (IrvineSciences) supplemented with 3% (vol/vol) defined fetal calfserum (HyClone) and 50 ,uM 2-mercaptoethanol and 200 mM
Abbreviations: SCF, stem cell factor; IL-2 and IL-7, interleukins 2 and7; sIgM, etc., surface IgM, etc.; RAG1, recombination-activating gene1; TdT, terminal deoxynucleotidyltransferase; C, constant region.tTo whom reprint requests should be addressed.

Proc. NatL Acad Sci USA 92 (1995) 1571
glutamine. Single-cell suspensions of cultured cells were incu-bated on ice for 30 min in saturating amounts of human-specific monoclonal antibodies (Becton Dickinson, SouthernBiotechnology Associates) in phosphate-buffered saline with2% calf serum, washed, and fixed in 1% paraformaldehyde.Samples were simultaneously analyzed by using isotype con-trols. Two-color flow cytometry was performed in a FACScanand analyzed using Lysis II software (Becton Dickinson). Thefluorokine human IL-7-conjugated biotin and SCF-conjugatedbiotin (R & D Systems) were used to detect IL-7 and SCFreceptors. Immunofluorescent terminal deoxynucleotidyl-transferase (TdT) staining (Supertechs, Bethesda, MD) wasperformed on cytospin preparations.
Nucleic Acid Analysis. High molecular weight DNA andtotal RNA were prepared and analyzed by standard methods.The human JH DNA probe (Oncogene Science) and otherprobes were labeled by random primerDNA labeling (Primelt,Stratagene) and had specific activities of 109 cpm/,ug ofDNA.
Cytokines and Growth Factor Stimulation. Recombinanthuman SCF was provided by K Zsebo and I. McNiece (Am-gen). Recombinant human IL-7 and anti-IL-7 monoclonalantibody were purchased from Collaborative Biomedical andGenzyme, respectively. For [3H]thymidine incorporation as-says, 105 cultured cells per well were grown in medium de-scribed above or in S17-conditioned medium in the presenceof growth factor for 48 hr and then were pulsed with 0.5 ,uCi(1 ,uCi = 37 kBq) of [3H]thymidine for 18 hr before scintilla-tion counting with a multiwell harvester.
RESULTSEstablishment of Long-Term Human B-Cell Progenitor
Cultures. Cord blood is a rich, readily available source ofhuman hematopoietic stem cells (26-29). Cultures were es-tablished by using a three-step procedure (Fig. 1). Cord mono-nuclear cells were isolated on a Ficoll/Hypaque gradient, and
HUMAN CORD BLOOD
j 1. FICOLL/HYPAQUE
CORD MONONUCLEARCELLS
2. CD34+ BIOTIWI AVIDIN COLUMN
CD34+ ENRICHEDCELLS
3. CULTURE ONMURINE STROMA
FIG. 1. Three-step method to establish human B-cell progenitorcultures. Cord blood mononuclear cells are collected on a Ficoll/Hypaque gradient, partially enriched for CD34+-expressing cells, andtransferred to semiconfluent S17 murine stromal cell cultures in RPMI1640 medium containing 3% defined fetal calf serum. Cultures wererefreshed biweekly and harvested at various time points for analysis.
then mononuclear cells were enriched for CD34+ progenitorcells by using a CD34+ biotin avidin column. This enrichmentstep resulted in a mononuclear cell population containingbetween 50% and 80% CD34+ cells (average, =60% in fouranalyses), and further lineage depletion or CD34+ cell enrich-ment was not performed. Finally, CD34+-enriched cells weretransferred onto 12-well or 10-cm2 tissue culture dishes con-taining a subconfluent monolayer of the murine bone marrowstromal cell line S17. The S17 stromal cell line was chosenbecause of its ability to maintain long-term murine pro-B-cellgrowth (9, 10, 25, 30, 31). A seeding density of 0.1-5 x 105 cellsper ml resulted in the most consistent establishment of cul-tures. Attempts to establish cultures by using unfractionated oradherence-depleted cord mononuclear cells were unsuccess-ful. For analyses, cultured cells were harvested with the ad-herent-cell monolayer by vigorous pipetting ahd passagethrough a needle and or cell sieve (40 ,um) to eliminate cellclumps. S17 cells could be easily distinguished from long-termcultured cells.
Establishment of long-term cultures followed three rela-tively distinct phases: (i) acellular period-during weeks 1-2,cultures contained only small numbers of nonadherent cellsand significant cell death was observed; (ii) myeloid expan-sion-during weeks 2-3, there was a marked increase in cel-lularity (up to 5 x 105 cells per ml) consisting primarily ofnonadherent myeloid cells identified by Wright stain and flowcytometry; and (iii) outgrowth of B-cell progenitors-beginning at 3-4 weeks, there was a decline in nonadherent(myeloid) cells and progressive outgrowth of small round cellstightly adherent to and burrowed within the stroma. Fig. 2Ashows a Wright stain of these long-term cultured cells, dem-onstrating their monomorphic cellular appearance character-istic of small- to medium-size lymphocytes. Close associationof some cultured cells with stroma remained even after dis-ruption and manipulation of culture monolayers.The surface phenotype of long-term cultured cells was eval-
uated by flow cytometry with antibodies specific for early andmore mature stages of B-cell development (Fig. 2B). Cultureswere progressively enriched for CD10+, CD19+ cells andusually contained 80-95% CD10+, CD19+ cells after 6 weeks.More mature B-lineage markers, including CD20, CD22,CD23, and sIgM, were not expressed on cultured cells (Fig.2B). Cells stained positively (>95%) for pancellular markersCD45 and CD38 but contained <1% CD34+-expressing cells.Myeloid cells (CD13, CD15, or CD33) were present during thefirst 3-4 weeks in culture but typically represented <5% ofcultured cells at 6 weeks. Erythroid and T-lineage cells werenot detected. These findings are consistent with the outgrowthand enrichment of early B-lineage progenitors.CD34+ cells (1-2 x 106) from an average cord blood spec-
imen (20-40 ml) were used to establish four to eight 10-cm2cultures. Culture dishes contained an average of 2-5 x 107 cellsat 6-8 weeks. Thus, a typical cord blood specimen couldgenerate -1-5 x 108 B-lineage progenitors. Established cul-tures could be expanded by transfer to new S17 feeder layers.A period of limited cell growth characterized by the presenceof small adherent foci was followed by expansion of popula-tions consisting of CD10+, CD19+, CD20-, sIgM- cells(D.J.R., S.G.Q., and O.N.W., unpublished data). Culturescould be maintained for periods of greater than 12 weekswithout significant change in cell surface phenotype. Cultureshave been established in >70% of attempts generating >30independent cultures.
Cultured Human B-Cell Progenitors Retain Immunoglob-ulin Genes in Germ-Line Configuration. DNA blot analysis offive independent cord blood cultures showed that the majorityof cultured cells at 6-8 weeks maintained immunoglobulinheavy chain genes in the germ line configuration (Fig. 3). Ex-tended exposures of these autoradiographs revealed no evi-dence of outgrowth of clonally rearranged subpopulations but
Immunology: Rawlings et aL

1572 Immunology: Rawlings et al.
A *-_A
B126 200
0010 9
0 0
200 -200-
0020 0022
0 0 -
200 - 200 -
CD023 1GM
0 0
200 - 200 -
0013 0034
0 0100 102 104 100 102 104
FIG. 2. Analysis of long-term cultures at 6 weeks. (A) Wright stainof cultured cells showing small-to-medium lymphocytes and largerstromal cells. Note rosette-like appearance of lymphoid cells sur-rounding the larger stromal cell in the upper right area that haveremained adherent despite vigorous disruption of culture and needlepassage. (B) Fluorescent antibody cell profiles of long-term-culturedcells demonstrating the expression of CD10 and CD19 B-lymphoidmarkers and the absence or low-level expression of more matureB-lineage markers-CD20, CD22, CD23, sIgM, the myeloid markerCD13, and the progenitor cell marker CD34. Staining with isotypecontrol antibody is shown for comparison.
cannot completely rule out independent rearrangementswithin a small subpopulation of cells. The data summarized inTable 1 suggests that several components of the recombinationprocess may be limiting immunoglobulin heavy chain gene
rearrangement in the long-term cultured cells, including ac-cessibility of the immunoglobulin locus and low-level expres-
Cord cultures 4
1 2 3 4 5
5.4 kbGerm line
FIG. 3. Long-term-cultured B-cell progenitors retain germ lineimmunoglobulin genes. High molecular weight DNA (10 ,g) from fiveindependent (6 week) cord blood cultures (1-5), murine S17 stroma,the human cervical carcinoma cell line HELA, and the human pre-B-cell line NALM-6 was used for Southern blot analysis afterBamHI/HindIII restriction digest. Blots were probed with a human JH frag-ment, and the position of the 5.4-kb germ-line immunoglobulin heavychain fragment is indicated. Rearrangement of both NALM-6 immu-noglobulin heavy chain alleles is present.
sion of recombination-associated genes. Transcription of thelocus encoding the ,u chain correlates with accessibility of thislocus for recombination. Less than 5% of cultured cells stainedpositively for the constant region ,u chain, and very low levelsof C,, transcript were detected by mRNA blot analysis. Ter-minal deoxynucleotidyltransferase (TdT) and the recombina-tion-activating gene 1 (RAGI) product are associated withcells active in the recombination process. Cytoplasmic stainingfor TdT expression was present in <3% of long-term-culturedcells. Low levels of RAG1 transcript were detectable bymRNA blot analysis. This signal was lower by a factor of 10-50in comparison with expression levels in human thymocytes(D.J.R., R.M.K., and O.N.W., unpublished data).
Cultured Human B-Cell Progenitors Do Not Proliferate inResponse to IL-7 or SCF. IL-7 and SCF are key factors formaintenance and expansion of early B-lineage cells. The S17stromal line does not produce detectable IL-7 byRNA (North-ern) blot, PCR, or bioassay (8, 30). While this line doesproduce SCF, murine SCF has limited effect on human SCF-responsive cells. The capacity of IL-7 and SCF to supportproliferation of long-term-cultured B-cell progenitors was ex-amined in both established and newly initiated cultures. Es-tablished (4 weeks old) cultures were harvested, and equal cellnumbers were transferred to new stroma for 2 weeks. Mediumcontaining IL-7 (5-50 ng/ml), SCF (10-100 ng/ml), or bothfactors was then added and refreshed after 3 days. At 7 days,cells were enumerated and analyzed by cell staining. B-cellprogenitors remained present, and growth factor additionresulted in no significant change in cell number or phenotypecompared with cells maintained in medium without addedfactors. Control IL-7-dependent cells proliferated 10- to 15-fold in IL-7-containing medium. Addition of IL-7 (10 ng/ml)upon initiation of cultures and biweekly feeding for 6 weeksalso revealed no significant difference in cell number or sur-face phenotype (D.J.R., S.G.Q., and O.N.W., unpublisheddata). To eliminate any possible role of IL-7 produced by theS17 stroma or by the cultured mononuclear cells, an IL-7-neutralizing antibody (directed against both human and mu-rine IL-7; 2.5 ,ug/ml) was added to established cultures (one
Table 1. Expression of recombination-associated genes
Gene Cell staining mRNA expression*
C,IL1-5% (4) Undetectable to low (5)TdT 1-3% (3) NDRAGI ND Undetectable to low (4)
The number of independent cultures is shown in parentheses. ND,not done.*Expression relative to control human B-cell progenitor line or thy-mocytes, respectively.
Proc. Natl. Acad ScL USA 92 (1995)
Proc. Natl. Acad. Sci. USA 92 (1995) 1573
A 150 - 150 -
0 0
100 102 104 100 102 104
B 1 2 3
IL-2R~
.. .......... ..........~ ~~~~... ...
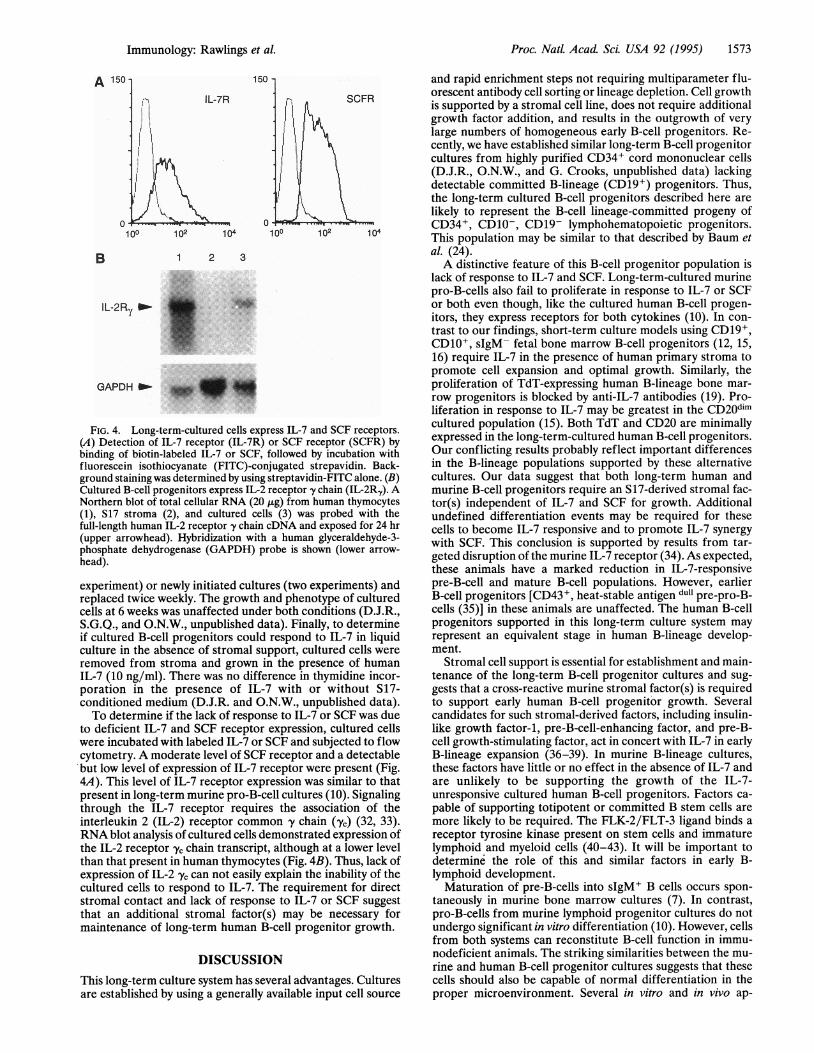
FIG. 4. Long-term-cultured cells express IL-7 and SCF receptors.
(A) Detection of IL-7 receptor (IL-7R) or SCF receptor (SCFR) by
binding of biotin-labeled IL-7 or SCF, followed by incubation with
fluorescein isothiocyanate (FITC)-conjugated strepavidin. Back-
ground staining was determined by using streptavidin-FITC alone. (B)Cultured B-cell progenitors express IL-2 receptor y chain (IL-2Ry). ANorthern blot of total cellular RNA (20 jitg) from human thymocytes
(1), S17 stroma (2), and cultured cells (3) was probed with the
full-length human IL-2 receptor y chain cDNA and exposed for 24 hr
(upper arrowhead). Hybridization with a human glyceraldehyde-3-
phosphate dehydrogenase (GAPDH) probe is shown (lower arrow-
head).
experiment) or n'ewly initiated cultures (two experiments) and
replaced twice weekly. The growth and phenotype of cultured
cells at 6 weeks was unaffected under both conditions (D.J.R.,S.G.Q., and O.N.W., unpublished data). Finally, to determine
if cultured B-cell progenitors could respond to IL-7 in liquidculture in the absence of stromal support, cultured cells were
removed from stroma and grown in the presence of human
IL-7 (10 ng/ml). There was no difference in thymidine incor-
poration in the presence of IL-7 with or without S17-
conditioned medium (D.J.R. and O.N.W., unpublished data).To determine if the lack of response to IL-7 or SCF was due
to deficient IL-7 and SCF receptor expression, cultured cells
were incubated with labeled IL-7 or SCF and subjected to flow
cytometry. A moderate level of SCF receptor and a detectable
'but low level of expression of IL-7 receptor were present (Fig.
4A). This level of IL-7 receptor expression was similar to that
present in long-term murine pro-B-cell cultures (10). Signaling
through the IL-7 receptor requires the association of the
interleukin 2 (IL-2) receptor common y' chain ('yc) (32, 33).RNA blot analysis of cultured cells demonstrated expression of
the IL-2 receptor yc chain transcript, although at a lower level
than that present in human thymocytes (Fig. 4B). Thus, lack of
expression of IL-2 yc can not easily explain the inability of the
cultured cells to respond to IL-7. The requirement for direct
stromal contact and lack of response to IL-7 or SCF suggest
that an additional stromal factor(s) may be necessary for
maintenance of long-term human B-cell progenitor growth.
ON
This long-term culture system has several advantages. Culturesare established by using a generally available input cell source
and rapid enrichment steps not requiring multiparameter flu-orescent antibody cell sorting or lineage depletion. Cell growthis supported by a stromal cell line, does not require additionalgrowth factor addition, and results in the outgrowth of verylarge numbers of homogeneous early B-cell progenitors. Re-cently, we have established similar long-term B-cell progenitorcultures from highly purified CD34+ cord mononuclear cells(D.J.R., O.N.W., and G. Crooks, unpublished data) lackingdetectable committed B-lineage (CD19+) progenitors. Thus,the long-term cultured B-cell progenitors described here arelikely to represent the B-cell lineage-committed progeny ofCD34+, CD10-, CD19- lymphohematopoietic progenitors.This population may be similar to that described by Baum etal. (24).A distinctive feature of this B-cell progenitor population is
lack of response to IL-7 and SCF. Long-term-cultured murinepro-B-cells also fail to proliferate in response to IL-7 or SCFor both even though, like the cultured human B-cell progen-itors, they express receptors for both cytokines (10). In con-trast to our findings, short-term culture models using CD19+,CD10+, sIgM- fetal bone marrow B-cell progenitors (12, 15,16) require IL-7 in the presence of human primary stroma topromote cell expansion and optimal growth. Similarly, theproliferation of TdT-expressing human B-lineage bone mar-row progenitors is blocked by anti-IL-7 antibodies (19). Pro-liferation in response to IL-7 may be greatest in the CD20dimcultured population (15). Both TdT and CD20 are minimallyexpressed in the long-term-cultured human B-cell progenitors.Our conflicting results probably reflect important differencesin the B-lineage populations supported by these alternativecultures. Our data suggest that both long-term human andmurine B-cell progenitors require an S17-derived stromal fac-tor(s) independent of IL-7 and SCF for growth. Additionalundefined differentiation events may be required for thesecells to become IL-7 responsive and to promote IL-7 synergywith SCF. This conclusion is supported by results from tar-geted disruption of the murine IL-7 receptor (34). As expected,these animals have a marked reduction in IL-7-responsivepre-B-cell and mature B-cell populations. However, earlierB-cell progenitors [CD43+, heat-stable antigen dull pre-pro-B-cells (35)] in these animals are unaffected. The human B-cellprogenitors supported in this long-term culture system mayrepresent an equivalent stage in human B-lineage develop-ment.
Stromal cell support is essential for establishment and main-tenance of the long-term B-cell progenitor cultures and sug-gests that a cross-reactive murine stromal factor(s) is requiredto support early human B-cell progenitor growth. Severalcandidates for such stromal-derived factors, including insulin-like growth factor-1, pre-B-cell-enhancing factor, and pre-B-cell growth-stimulating factor, act in concert with IL-7 in earlyB-lineage expansion (36-39). In murine B-lineage cultures,these factors have little or no effect in the absence of IL-7 andare unlikely to be supporting the growth of the IL-7-unresponsive cultured human B-cell progenitors. Factors ca-pable of supporting totipotent or committed B stem cells aremore likely to be required. The FLK-2/FLT-3 ligand binds areceptor tyrosine kinase present on stem cells and immaturelymphoid and myeloid cells (40-43). It will be important todetermine the role of this and similar factors in early B-lymphoid development.
Maturation of pre-B-cells into sIgM+ B cells occurs spon-taneously in murine bone marrow cultures (7). In contrast,pro-B-cells from murine lymphoid progenitor cultures do notundergo significant in vitro differentiation (10). However, cellsfrom both systems can reconstitute B-cell function in immu-nodeficient animals. The striking similarities between the mu-rine and human B-cell progenitor cultures suggests that thesecells should also be capable of normal differentiation in theproper microenvironment. Several in vitro and in vivo ap-
Immunology: Rawlings et al.

1574 Immunology: Rawlings et al.
proaches, including transfer to severe combined immunodefi-cient mice, are currently being employed to promote differ-entiation of these cells.
We are grateful to Richard Stiehm and Bonnie Ank for providingcord blood, David Schatz for human RAGI DNA, Jonathan Braun forhuman C., Warren Leonard for human IL-2 receptor y subunit cDNA,Christel Uittenbogaart for human thymocytes, Julia Shimaoka forassistance in manuscript preparation, and Doug Saffran for criticalreading of the manuscript. O.N.W. is an Investigator of the HowardHughes Medical Institute. This work was supported by grantsAR36834 and AR01912 to D.J.R. and CA12800 to O.N.W.
1. Uckun, F. M. (1990) Blood 76, 1908-1923.2. Rolink, A. & Melchers, F. (1991) Cell 66, 1081-1094.3. Cooper, M. D. (1987) N. Engl. J. Med. 317, 1452-1456.4. Kincade, P. W., Lee, G., Pietrangeli, C. E., Hayashi, S.-I. &
Gimble, J. M. (1989) Annu. Rev. Immunol. 7, 111-143.5. Kincade, P. W. (1994) Proc. Natl. Acad. Sci. USA 91, 2888-2889.6. Dorshkind, K. (1990) Annu. Rev. Immunol. 8, 111-137.7. Whitlock, C. A. & Witte, 0. N. (1982) Proc. Natl. Acad. Sci. USA
79, 3608-3612.8. Scherle, P. A., Dorshkind, K. & Witte, 0. N. (1990) Proc. Natl.
Acad. Sci. USA 87, 1908-1912.9. Saffran, D. C., Faust, E. A. & Witte, 0. N. (1992) Curr. Top.
Microbiol. Immunol. 182, 34-44.10. Faust, E. A., Saffran, D. C., Toksoz, D., Williams, D. A. & Witte,
0. N. (1993) J. Exp. Med. 177, 915-923.11. Dorshkind, K., Denis, K A. & Witte, 0. N. (1986) J. Immunol.
137, 3457-3463.12. Wolf, M. L., Buckley, J. A., Goldfarb, A., Law, C.-L. & LeBien,
T. W. (1991) J. Immunol. 147, 3324-3330.13. McGinnes, K, Letarte, M. & Paige, C. J. (1991) Blood 77, 961-
970.14. McGinnes, K, Quesniaux, V., Hitzler, J. & Paige, C. (1991) Exp.
Hematol. 19, 294-303.15. Moreau, I., Duvert, V., Banchereau, J. & Saeland, S. (1993) Blood
81, 1170-1178.16. Moreau, I., Duvert, V., Caux, C., Galmiche, M.-C., Charbord, P.,
Banchereau, J. & Saeland, S. (1993) Blood 82, 2396-2405.17. Saeland, S., Duvert, V., Pandrau, D., Caux, C., Durand, I.,
Wrighton, N., Wideman, J., Lee, F. & Banchereau, J. (1991)Blood 78, 2229-2238.
18. Villablanca, J. G., Anderson, J. M., Moseley, M., Law, C.-L.,Elstrom, R. L. & LeBien, T. W. (1990)J. Exp. Med. 172,325-334.
19. Ryan, D. H., Nuccie, B. L., Ritterman, I., Liesveld, J. L. & Ab-boud, C. N. (1994) J. Immunol. 152, 5250-5258.
20. Cicuttini, F. M., Martin, M., Salvaris, E., Ashman, L., Begley,C. G., Novotny, J., Maher, D. & Boyd, A. W. (1992) Blood 80,102-112.
21. Thalmeier, K, Meissner, P., Reisbach, G., Falk, M., Brechtel, A.& Dormer, P. (1994) Blood 83, 1799-1807.
22. Sutherland, H. J., Eaves, C. J., Lansdorp, P. M., Thacker, J. D. &Hogge, D. E. (1991) Blood 78, 666-672.
23. Issaad, C., Croisille, L., Katz, A., Vainchenker, W. & Coulombel,L. (1993) Blood 81, 2916-2924.
24. Baum, C. M., Weissman, I. L., Tsukamoto, A. S., Buckle,Anne-M. & Peault, B. (1992) Proc. Natl. Acad. Sci. USA 89,2804-2808.
25. Collins, L. S. & Dorshkind, K. (1987) J. Immunol. 4, 1082-1087.26. Broxmeyer, H. E., Douglas, G. W., Hangoc, G., Cooper, S., Bard,
J., English, D., Amy, M., Thomas, L. & Boyse, E. A. (1989) Proc.Natl. Acad. Sci. USA 86, 3828-3832.
27. Hows, J. M., Bradley, B. A., Marsh, J. C. W., Luft, T., Coutinho,L., Testa, N. G. & Dexter, T. M. (1992) Lancet 340, 73-76.
28. Lu, L., Xiao, M., Shen, R.-N., Grigsby, S. & Broxmeyer, H. E.(1993) Blood 81, 41-48.
29. Moritz, T., Keller, D. C. & Williams, D. A. (1993) J. Etp. Med.178, 529-536.
30. Billips, L. G., Petitte, D., Dorshkind, K, Narayanan, R., Chiu,C.-P. & Landreth, K S. (1992) Blood 79, 1185-1192.
31. Henderson, A. J., Johnson, A. & Dorshkind, K (1990) J. Immu-nol. 145, 423-428.
32. Noguchi, M., Nakamura, Y., Russell, S. M., Ziegler, S. F., Tsang,M., Cao, X. & Leonard, W. J. (1993) Science 262, 1877-1880.
33. Kondo, M., Takeshita, T., Higuchi, M., Nakamura, M., Sudo, T.,Nishikawa, S.-I. & Sugamura, K (1994) Science 263, 1453-1454.
34. Peschon, J. J., Morrissey, P. J., Grabstein, K H., Ramsdell, F. J.,Maraskovsky, E., Gliniak, B. C., Park, L. S., Ziegler, S. F., Wil-liams, D. E., Ware, C. B., Meyer, J. D. & Davison, B. L. (1994)J. Exp. Med. 180, 1955-1960.
35. Hardy, R. R., Carmack, C. E., Shinton, S. A., Kemp, J. D. &Hayakawa, K (1991) J. Exp. Med. 173, 1213-1225.
36. Gibson, L. F., Piktel, D. & Landreth, K S. (1993) Blood 82,3005-3011.
37. Landreth,K S., Narayanan, R. & Dorshkind, K. (1992) Blood 80,1207-1212.
38. Samal, B., Sun, Y., Steams, G., Xie, C., Suggs, S. & McNiece, I.(1994) Mol. Cell. Biol. 14, 1431-1437.
39. Nagasawa, T., Kikutani, H. & Kishimoto, T. (1994) Proc. Natl.Acad. Sci. USA 91, 2305-2309.
40. Hannum, C., Culpepper, J., Campbell, D., McClanahan, T.,Zurawski, S., et al. (1994) Nature (London) 368, 643-648.
41. Matthews, W., Jordan, C. T., Wiegand, G. W., Pardoll, D. &Lemischka, I. R. (1991) Cell 65, 1143-1152.
42. Rosnet, O., Marchetto, S., deLapeyriere, 0. & Birnbaum, D.(1991) Oncogene 6, 1641-1650.
43. Small, D., Levenstein, M., Kim, E., Carow, C., Amin, S., Rock-well, P., Witte, L., Burrow, C., Ratajczak, M. Z., Gewirtz, A. M.& Civin, C. I. (1994) Proc. Natl. Acad. Sci. USA 91, 459-463.
Proc. Natl Acad Sci USA 92 (1995)