location ofpupillomotor and accommodation fibres ... · pressure under direct vision to various...
TRANSCRIPT

J. Neurol. Neurosurg. Psychiat., 1964, 27, 473
Location of pupillomotor and accommodation fibresin the oculomotor nerve: Experimental observations
on paralytic mydriasis1FREDERICK W. L. KERR AND 0. WEEMS HOLLOWELL
From the Section of Neurologic Surgery, Mayo Clinic and Mayo Foundation, Rochester, Minnesota, U.S.A.
Hutchinson (1867-68) in his 'Lectures on compressionof the brain' was the first to indicate that with anacutely expanding intracranial mass, the ipsilateralpupil frequently dilated and became unresponsive tolight. This he attributed to compression of the thirdnerve. Von Bergmann (1880) and subsequentlyMacewen (1887) came to similar conclusions re-garding the significance of paralytic mydriasis indetermining on which side such lesions were to befound. These authors and most subsequent writerson the subject have favoured compression ofthe thirdnerve as the most likely mechanism.A number of reports over the years, however, have
suggested another basis for this sign. Cushing (1908)expressed the view that compression of the cerebralhemisphere was responsible and that brief narrowingof the pupil preceded progressive mydriasis. Adrogueand Balado (1925) said that compression of somestructure other than the third nerve should be con-sidered, at least in some instances, since in theirexperimental and clinical studies the nerve occasion-ally showed no evidence of trauma. Schorcher(1937), on the basis of experimental and clinicaldata, stated categorically that compression of theoculomotor nerve was not a factor in the earlyphases of paralytic mydriasis and that irritation ofsympathetic pathways in the brain-stem should beregarded as the predominant mechanism. Reid andCone (1939) concluded that pressure on the thirdnerve was the cause of the dilated pupil and pointedout that because of the close association of pupillo-constrictor centres in the midbrain it was highlyimprobable that unilateral mydriasis would occurfrom pressure there. Nelson (1942) and Penfield andMcEachern (1949), however, attributed paralyticmydriasis to compression of the brain-stem.
Current opinion favours trauma to the third nerve,and among more recent studies sponsoring this view-
'This investigation was supported in part by research grant B-3296from the National Institutes of Public Health Service, and the paperwas read at the meeting of the Society of Neurological Surgeons,Rochester, Minnesota, 7-9 May 1964.
point are those of Welte (1943), Sunderland andHughes (1946), Sunderland and Bradley (1953),Lazorthes, Pigassou, and Gaubert (1955), Sunder-land (1958), Weintraub (1960), Jennett and Stem(1960), and Keefe, Rucker, and Kernohan (1960).
In a recent study of pupillomotor pathways in thespinal cord in the cat (Kerr and Brown, 1964), theease with which paralytic mydriasis could be pro-duced, occasionally lasting for a number of hours, inresponse to a single episode of trauma to the cervicalor upper thoracic cord, again raised the possibilitythat compression of the brain-stem and irritation ofthe sympathetic pupillomotor pathway might play asignificant role in this response. The fact that lateralcompression of the cord led to ipsilateral pupillo-dilatation appeared to detract from the point madeby Reid and Cone (1939) that proximity of pupillo-motor centres ruled out unilateral pupillary changesin response to pressure, since in the thoracic spinalcord of the cat the right and left ciliospinal centresare separated by a distance of less than 2 mm.For these reasons we decided to re-evaluate the
mechanisms of pupillary-dilatation secondary to in-creased pressure and to test the theories by applyingpressure under direct vision to various supraten-torial structures.As the study progressed, it became necessary also
to determine the course of the pupillomotor fibresin the third nerve, since some conflict of opinionexists in the literature on this point. The most gener-ally accepted view is that of Sunderland and Hughes(1946) who stated that small fibres are most numer-ous on the superior arc of the third nerve in its proxi-mal course near the brain-stem and concluded thatthese were the fibres that passed to the inferior divi-sion and thence to the ciliary ganglion. Lyle (1954),however, noted that the pupillomotor fibres probablyproceed down the central part of the nerve, a viewfirst proposed by St. Bernheimer (1897) and later byFuchs (1907). Lazorthes et al. (1955) stated that thesefibres emerge from the brain-stem on the medialaspect of the nerve from whence they pass laterally
473
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Frederick W. L. Kerr and 0. Weems Hollowell
to reach the inferior surface. The precise location ofthese fibres is of some practical importance in tryingto determine the mechanism of paralytic mydriasis,particularly in view of the well-known observation(Wilson, 1949; Huber, 1961) that a relatively pro-nounced degree of pupillary dilatation may resultfrom pressure, without any clinically detectable im-pairment of extraocular motility but Jennett andStern (1960) cast some doubt on this observation.The dissociation of intraocular and extraocularmotility has been variously accounted for. Someinvestigators have considered the pupillomotorfibres more vulnerable to pressure, a suggestion whichis at variance with the greater resistance of smallfibres to pressure block, while others think that thepupillomotor fibres are located in a more vulnerableposition.
MATERIAL AND METHODS
The animals used for the compression and stimulationstudies included 10 dogs, one Macacus rhesus, and onebaboon (Papio papio); in addition, some of the studieson dogs in which direct pressure was applied to the brain-stem and to the third nerve were repeated on three cats.For all experimental procedures deep pentobarbitalsodium anesthesia was used; supra-collicular decerebra-tion with removal of the brain rostral to this plane wascarried out in those experiments in which stimulation ofthe third nerves was to be done.
COMPRESSION STUDIES Pressure was applied to the con-vexity and to the medial and the inferior aspects of thehemisphere. A Chatillon gaugewas used in these and otherstudies of this type to estimate in grams per squarecentimetre the actual pressure applied. Pressure was ap-plied to the lateral aspect of the brain-stem at the levelof the tentorial notch by means of a foam rubber cushioncut to the approximate size and shape of the medialaspect of the temporal lobe. In other studies, pressurewas applied vertically on the quadrigeminal plate, alsoby means of a small foam rubber cushion, or trans-versely to compress the tegmentum at the level of thethird nerve nucleus without lateral displacement. Finally,pressure, measured by a Chatillon gauge, was applieddirectly to the third nerve in a similar manner. Theseobservations on the effects of pressure can be regardedonly as relatively gross approximations.
STIMULATION STUDIES The usual stimulus employed was6 v., 60/sec., 1 msec., square wave pulses delivered by aGrass stimulator via standard insulated monopolar steelelectrodes, the indifferent electrode being placed in theparavertebral musculature. When the preganglionicpathway in the third nerve was being traced, steel micro-electrodes were used to restrict the area of stimulation to aminimum, and stimuli which were just suprathreshold tomaximal were used. Thus discrete identification of thefibres was possible. Since marked miosis is present indecereblate animals in good condition, it was necessary
in most instances to produce pupillary dilatation as aprelirninary step to stimulation of the pupillomotorfibres in the third nerve. This was usually obtained bymeans of mild compression of the nerve as it left thebrain-stem and was cross-checked in experiments inwhich the nerve had not been touched; in others in whichthe nerve was completely transected at the brain-stem,the nerve was stimulated distal to the section, and in stillother experiments mydriasis was produced by sustainedstimulation of the cervical sympathetic chain, and duringthe resultant mydriasis various aspects of the third nervewere stimulated. In addition to stimulation of the thirdnerve, multiple points of the cortex, again including themedial and inferior aspects of the hemisphere, werestimulated to determine the areas from which pupillo-motor responses could be obtained before carrying outcompression experiments.The effect of sectioning various quadrants of the third
nerve was studied also. The four quadrants of the nervewere marked by introducing fine rigid steel wires throughthe nerve at right angles to each other (a procedure whichdid not influence the diameter of the pupil or the re-sponse); following this, individual nerve quadrants wererelatively accurately sectioned with fine iris scissors. Inother instances, the nerve was sectioned progressivelyfrom lateral to medial or vice versa.
Pupillary responses were photographed by means of aGrass camera triggered by a relay at 2 or 5 frames persecond; a cystoscope lamp bulb on the animal's foreheadsignalled the beginning and end of stimulation. Otherdetails of technique are as described elsewhere (Kerr andBrown, 1964).
HISTOLOGICAL STUDY Ten oculomotor nerves from man,two from macaques, and four from dogs were studied.The lateral and superior aspects of all the nerves weremarked with a narrow stripe of red and of green plasticpaint, respectively; this was allowed to dry beforeplacing the specimen in 10%. formalin. These identifica-tion marks were applied with the nerve and brain in situsince, on sectioning the nerve just distal to the pointwhere it enters the dura, considerable external rotationtended to occur. Three segments were cut from each nerve,one at the emergence from the brain-stem, one at themidpoint in the subarachnoid space, and one at the levelwhere the nerve pierces the dura.The plastic marking resisted subsequent processing of
the nerve until sections were attached to the slides; de-paraffinizing frequently removed the markers. Therefore,their position was identified on the slide with a diamondpoint before deparaffinizing.
Sections were cut at 8 microns and stained by theBodian technique.
RESULTS
COMPRESSION STUDIES Pressure applied to any orall points of the cerebral hemispheres (including thebasal aspects) for 30 to 60 seconds or more had noeffect on the size of the pupils provided the directionin which the pressure was applied did not compress
474
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from
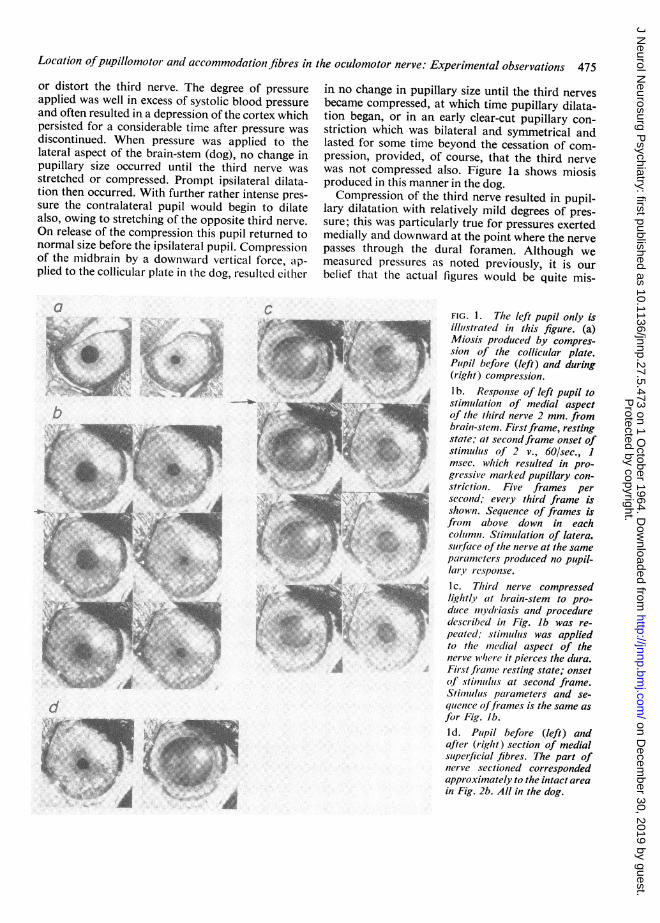
Location ofpupillomotor and accommodationi fibres in the oculomotor nerve: Experimental observations 475or distort the third nerve. The degree of pressureapplied was well in excess of systolic blood pressureand often resulted in a depression of the cortex whichpersisted for a considerable time after pressure wasdiscontinued. When pressure was applied to thelateral aspect of the brain-stem (dog), no change inpupillary size occurred until the third nerve wasstretched or compressed. Prompt ipsilateral dilata-tion then occurred. With further rather intense pres-sure the contralateral pupil would begin to dilatealso, owing to stretching of the opposite third nerve.On release of the compression this pupil returned tonormal size before the ipsilateral pupil. Compressionof the midbrain by a downward vertical force, ap-plied to the collicular plate in the dog, resulted either
a
CSot' 'O
I~~.t4 ...I;Ie
in no change in pupillary size until the third nervesbecame compressed, at which time pupillary dilata-tion began, or in an early clear-cut pupillary con-striction which was bilateral and symmetrical andlasted for some time beyond the cessation of com-pression, provided, of course, that the third nervewas not compressed also. Figure la shows miosisproduced in this manner in the dog.Compression of the third nerve resulted in pupil-
lary dilatation with relatively mild degrees of pres-sure; this was particularly true for pressures exertedmedially and downward at the point where the nervepasses through the dural foramen. Although wemeasured pressures as noted previously, it is ourbelief that the actual figures would be quite mis-
FIG. 1. The left pupil only isillutstrated in this figure. (a)Miosis produced by compres-sion of the collicular plate.Pupil before (left) and during(right) compression.lb. Response of left pupil tostimulation of medial aspectof the third nerve 2 mm. frombrain-stem. Firstframe, restingstate; at secondframe onset ofstimulus of 2 v., 60/sec., 1msec. which resulted in pro-gressive marked pupillary con-striction. Five frames persecond; every third frame isshown. Sequence of frames isfrom above down in eachcolumn. Stimutlation of latera.surface ofthe nerve at the sameparaimeters produced no pupil-lary response.Ic. Third nerve compressedlightly at brain-stem to pro-duce mydriasis and proceduredescribed in Fig. lb was re-
peated, stimululs was appliedto the mnedial aspect of thenerve whler it pierces the dura.First franme resting state; onsetof stimiulus at second frame.Stimulus parameters and se-quence offrames is the same asfor Fig. Jb.I d. Pupil before (left) andafter (right) section of medialsuperficial fibres. The part ofnerve sectioned correspondedapproximately to the intact areain Fig. 2b. All in the dog.
*q,.fk SA:|: f s l 8ZZes .t ............. xso :.
#
.~*_i;iftA.
d
.I..
"Y; S S'
.4.;
-1 -5.
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Frederick W. L. Kerr and 0. Weems Hollowell
leading, since minimal changes in the direction ofapplied pressure resulted in significant changes insize of the pupil, but marked changes in pressure ina given direction were often without effect.The conclusion that may be drawn, however, on
the basis of repeating the compression experimentsmany times, both in the same and other animals, isthat the third nerve is by far the most sensitivestructure to pressure in so far as the production ofmydriasis is concerned; pressures of equal or greaterintensity applied to the cerebral cortex, the lateralor dorsal aspect of the brain-stem at the level of thetentorial notch, being either without effect or, as inthe case of the quadrigeminal plate, result in miosis.
STIMULATION OF THE THIRD NERVE Using gradedelectrical stimuli as described under Methods andwith both third nerves exposed from the brain-stemto the dural penetration, the nerve was stimulated atsuccessive points around its circumference; thiswas done at the point of emergence of the nerve
from the midbrain, at the point where the dura was
penetrated, and midway between these two points.Some variation in the response occurred from animalto animal, and the description which follows cor-
responds to the most typical experiments. Stimula-tion of the superior surface of the nerve at its emerg-ence from the midbrain resulted in maximal pupillaryconstriction, particularly on the medial half of thesuperior surface. Stimulation of the lateral and in-ferior quadrants of the nerve at this level had no
effect on the pupil, while stimulation of the medialsurface resulted in progressively less response as thestimulus was applied farther from the dorsomedialaspect.At the midpoint of its subarachnoid course, the
maximal response was obtained from the medialaspect of the nerve, either exactly on the horizontalmeridian (Fig. lb) or slightly above or below thispoint. Again stimulation on the dorsal, lateral, or
ventral aspects of the nerve produced either minimalor no pupillary changes. From the area of maximalresponse, the pupillary constriction decreased pro-
gressively as stimulus was applied more dorsally or
ventrally.When the nerve was stimulated at the dural
foramen, the maximal response was obtained againfrom stimulation of the medial aspect (Fig. Ic), thatis, from stimulation ofthe fibres occupying a positionsimilar to or identical with those that gave the maxi-mal response in the mid portion of the nerve. How-ever, at the dural foramen the fibres appeared to bemore spread out and a fairly pronounced response
was usually obtained from the ventrolateral aspectof the nerve also.
STIMULATION OF THE CORTEX Cortical stimulation
was done to identify areas from which a pupillaryresponse to pressure might be expected and not asa study of electrically evoked pupillary responsesper se.The areas of the cerebral cortex which were
stimulated in the dog included the whole of the con-vexity, the medial aspect, including the cingulategyrus and the ventral aspect of the temporal lobe,and the hippocampal gyrus. In general, pupillaryresponses to a stimulus of 6 v., 60/sec., 1 msec. werenegligible or minimal amounting to less than 1 mm.of change in diameter from all points tested on theconvexity with the exception of the most lateralaspect of the cruciate gyrus at which point a sym-metrical dilatation of approximately 2 mm. occurred.The most pronounced dilatation resulted from
stimulation of the ventral surface of the hippocampalgyrus at about its midpoint; in these instancesmydriasis from a diameter of approximately 1 mm.up to approximately 5 to 6 mm. occurred in thecontralateral pupil while the ipsilateral pupil showeda less marked response. This pupillary responsewas obtained repeatedly with the same characteris-tics. The contralateral cervical sympathetic chainbelow the superior ganglion was then sectioned;when the same cortical area was stimulated, clear-cut but less marked pupillary dilatation occurredon the side of the sympathectomy, the response onthe side of the intact sympathetic chain being un-changed.
TRANSECTIONS OF THE THIRD NERVE With the datafrom stimulation, the third nerve was progressivelysectioned in a series of experiments to cross-checkthe validity of the former results.The sections of the nerve were carried out at the
midpoint of its course in the subarachnoid space orat the point where the nerve passed into the dura andstimulation was applied proximal to the severedportion of the nerve (Fig. 2). The lateral half of thenerve could be transected at these levels without anyeffect on pupillary response to stimulation of thenerve proximal to the lesion (Fig. 2a). Further sec-tioning of the nerve to include the central or axialportion likewise produced no change. When all thatremained of the nerve was a thin, almost translucentband of the most medial fibres, the pupillary responseremained unchanged (Fig. 2b). Progressive section-ing of this medial husk of the nerve then resultedin gradual deterioration of the pupillary response,which disappeared as the last fibres were transected.In the experiment on the baboon on which Fig. 2is based, the pupil was initially mydriatic.
Section of the third nerve in the reverse direction,that is, from medial to lateral, resulted in initialdilatation of the pupil, but full dilatation did not
476
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Location ofpupillomotor and accommodation fibres in the oculomotor nerve: Experimental observations 477
a
L
R
b c d
M
/to
2"
I1 ...LI0 5mm.
FIG. 2. Effect ofprogressive sectioning of the third nerve (baboon) at dural penetration point on electrically evokedpupillary response. Stimulus of 10 v., 60/sec., 5 msec. was applied to medial aspect of nerve 2 mm. from brain-stem.2a. Lateral (L) halfsectioned. 2b. Lateralfour-fifths sectioned. 2c. Small ventromedial bundle remaining. 2d. Responsefrom intact nerve for comparison.
Upper row. Pupil at rest. Middle and lower rows. Response at I and 2 seconds, respectively, from onset ofstimulation.Scale for pupil diameter in millimetres. Discussion in text.
occur in some instances until the whole of the nervehad been transected, and a mild response could beobtained to stimulation proximal to the section whenthe medial half of the nerve had been transected.This indicates that, despite negative results of stimu-lation, some pupillomotor fibres run in the lateralhalf of the nerve and this small number of pupillo-motor fibres is sufficient to maintain a degree ofpupillary tone. However, in some instances sectionof the most medially placed fibres at the midpointin the subarachnoid course caused immediate andpronounced paralytic mydriasis (Fig. Id). Nochanges in pupillary contour (discoria) occurred as aresult of these partial transections, the dilatationbeing regular and progressive. Section of the nerve inquadrants also confirmed these observations; insome instances the cut surface of the distal stumpof the nerve was stimulated. No pupillary response
was obtained from stimulation of the central partof the nerve, whereas the maximal response waselicited from the medial and superficial aspects.
OBSERVATIONS ON ACCOMMODATION During thecourse of these observations on pupillary constric-tion in response to localized stimulation of the thirdnerve, changes in accommodation were looked forand found. Simple inspection revealed a markedforward displacement of the iris in response tostimulation of the nerve at specific points; this dis-placement is due to the increased curvature of theanterior surface of the lens which occurs on ac-commodation (Adler, 1959). That accommodationoccurred was also clearly demonstrated by retino-scopy.
In every instance accommodation was associatedwith pupillary constriction, whereas no accommoda-
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

f:P
W...kLbs jS
'ti, t ,
v Y ^#4*st i
:% :>w fi
Q t.;1
FIG. 3a
.* Wt.§t
.ttu.,½4/<
:i, 4, */
479~~~~~~~~~-
,-A N4~~~~~~~~~~~~~~~~~~~ ~
Atft" S.
no.3b
* ,. ' v A
r *# #w w &'
* ~
tiPt.0
4 A:7
* ~~ ~....:**v
>ee\1t *- -ih ~**;0s
FIG. 3c FnG. 3dFIG. 3. Pupillomotorfibres in the third nerve in man, monkey, and dog. 3a. At brain-stem in man thesefibres are containedin afewfine strands in the superomedial angle ( x 500). 3b. At the midpoint in the subarachnoid space ( x 500). 3c. At thebrain-stem in the monkey (M. rhesus) they form a conspicuous layer beneath the epineurium ( x 500). 3d. In the dog theyare also conspicuous and can be seen superficially as well as spreading more deeply between the larger fibres ( x 425)(shown here at the mid-point of the nerve in the subarachnoid space).
A
.N.
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Location ofpupillomotor and accommodationi fibres in the oculomotor nerve: Experimental observations 479
tion occurred when there was no pupillary responseto stimulation of the third nerve. It is thus clear thatthe two responses are mediated by fibres intimatelyassociated in the nerve. Furthermore, when weakpupillary responses were elicited, correspondinglyweak accommodation occurred.
HISTOLOGICAL OBSERVATIONS As noted, sections ofthe third nerve at the brain-stem, at the point wherethe nerve pierces the dura, and midway between thesetwo levels were studied.
Fibres fell into two main groups. One of thesecomprised the large fibres ranging between 10 and16 p. The other was represented by small fibres of adiameter between about 5 and 1-0 p, which wererather distinctive in the dog and monkey (Fig. 3),since the myelin sheath tended to stain pink topurple and clusters of such fibres were readilyidentified. They were located readily in both specieson the dorsomedial aspect of the nerve at the level ofexit from the brain-stem; subsequently, at the middle
a
Dog
Monkey _
and more distal parts of the nerve they were foundalong the most medial aspect of the nerve. Thesefibres were arranged in small bundles, many of thembeing placed superficially immediately beneath theepineurium; some of the bundles retained thisposition while others extended inward for variablebut usually small distances between the larger fibres,from which they were quite distinct.The superficial lateral and ventral aspects of the
nerve were devoid of these clusters of small fibres,although isolated groups of small fibres were notuncommon throughout the deeper layers ofthe nerve.In man the small fibres at or near the surface of thenerve are difficult to demonstrate for reasons whichare not clear. Possibly immediate fixation is neces-sary for good results. We were able to demonstratethem satisfactorily in only two of the 10 humannerves studied. Even in these two they were far lessobvious than in the dog and monkey, and it shouldbe noted that in the sections in Fig. 4 an interroga-tion mark indicates that the fibres seen at the level
b
L M
Q,
C
Man <> +DVI
Mo
FIG. 4. Location ofppupillomotor and accommodationfibres in the third nerve ofdog, monkey, and man. 4a. (first column).At emergencefrom brain-stem. 4b. Midpoint in subarachnoid space. 4c. At point where nerve pierces dura. All sections arearranged in identical manner, the superior surface being upward; L corresponds to lateral andM to medial aspect, for alldiagrams.
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Frederick W. L. Kerr and 0. Weems Hollowell
of penetration of the dura, in man, although small,did not show the characteristic clustered arrange-ment of the fibres we have considered to be pupillo-motor in the dog and monkey. Therefore, the loca-tion of pupillomotor fibres at the dural foramen inman has not been unequivocally demonstrated, buton a comparative anatomical basis and from the moreproximal location ofthese fibres in the nerve and theirultimate destination to the inferior division, it maybe concluded that they lie medially or ventromedially.
DISCUSSION
From the experiments carried out, many of themgross but nevertheless mimicking a gross pheno-menon such as an acute localized rise in intracranialpressure, it appears that compression of supra-tentorial structures other than the third nerve doesnot produce mydriasis; when mydriasis does occurit is almost certainly due to transmitted pressure ordistortion of the nerve.Some authors (Cushing, 1908; Dott, 1939;
Fischer-Bruigge, 1951, among others) have stated thata brief period of ipsilateral miosis may precede theonset of paralytic mydriasis. Cushing consideredboth phenomena due to compression of the hemi-sphere while Fischer-Bruigge attributed them topressure on the third nerve. Although a transientslight constriction of the pupil was noted in responseto direct compression of the third nerve on one oc-casion in our experiments, this was so unusual thatlittle significance can be attached to it. Certainly,from the clinical point of view a response as brief asthis (lasting for a few seconds) would almost certainlypass unperceived. Compression of the collicularplate resulted in marked miosis in the cat and oc-casionally in the dog; it may be assumed that this isdue to stimulation of the underlying parasympatheticcentre. Unilateral miosis was not observed, however.Even rather intense pressure leading to persistentdeformation of the tegmentum did not result inmydriasis, and it is interesting to note in this regardthat compression with corresponding ischaemicanoxia either of the parasympathetic pupillocon-strictor outflow as described herein or of thesympathetic pupillo-dilator outflow (Budge's cilio-spinal centre, Kerr and Brown, 1964) gives rise tosigns of irritation or activation and not to paralysis.It is probable that if compression were sustainedparalysis would ensue but we did not prolongcompression much beyond 30 to 60 seconds in thesestudies. These observations may be of some signi-ficance in explaining the miosis that precedes para-lytic mydriasis. Since compression of the third nerveresults in only fleeting miosis, and rarely at that,
tegmental compression would appear to be a morelikely mechanism. Failure to observe strictly ipsi-lateral miosis in our experiments tends to weakenthis hypothesis, however.With regard to pupillary dilatation in response to
localized pressure changes, the evidence appears tobe reasonably clear-cut. A cortical origin for pressuremydriasis can be ruled out since pupillary changeswere not produced by pressure at any point on theconvexity, the medial or the ventral surface of thebrain. When pupillary changes of any significancewere produced by electrical stimulation of the cortex,the response was either bilateral and symmetrical or,as in the case of the hippocampal gyrus, the contra-lateral pupil became larger than the ipsilateral one.Thus an irritative cortical lesion would not produceipsilateral dilatation.The evidence from this study of the effect of pres-
sure applied under direct vision to supratentorialstructures indicates that mydriasis is produced mostreadily by compression of the third nerve. Thismydriasis is reversible, but with repeated compres-sion becomes fixed. The direction of applied pressurewas more significant than the degree of pressurewithin certain limits. Thus, compression of the nerveagainst medially placed structures near the point ofexit from the subarachnoid space produced mydriasiswith relatively mild pressures, whereas direct verticalpressure of the same degree on the nerve at this levelwas much less effective. This correlates with theobservation that marked generalized increase inintracranial pressure may develop (Browder andMeyers, 1939) without significant changes in pupil-lary function, whereas less pressure with displacementof the intracranial contents frequently results inparalytic mydriasis.The determination of the position of the pupillo-
motor fibres in the third nerve is based on bothanatomical and functional studies. Since the resultsof these two methods are in close agreement, it maybe stated that, between the brain-stem and the exitof the nerve from the subarachnoid space, thesefibres lie along the dorsomedial and medial aspectsof the nerve and have a gradually descending courseas they run forward. Furthermore, they are locatedvery superficially, most of them being immediatelybeneath the epineurium.One discrepancy in our experiments should be
noted, however; thus, while a fairly brisk contractionof the pupil could usually be obtained from theventrolateral aspect of the nerve near the point ofdural penetration, no definite aggregation of finefibres could be demonstrated histologically at thispoint. It is possible that some of the scattered finefibres seen throughout the nervemay mediate pupillo-motor functions and thus account for the discrepancy
480
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from

Location ofpupilloniotor and accommodation fibres in the oculomotor nerve: Experimental observations 481
between stimulation and structural findings in thisinstance.The location of the pupillomotor fibres as de-
scribed here confirms the observations of Sunderlandand Hughes (1946) and supports, from the structuralpoint of view, the mechanism of compression of thesuperior surface of the third nerve by the posteriorcerebral artery as described by Fischer-Brugge (1951),Sunderland and Bradley (1953), Keefe et al. (1960),and others. On the other hand, the medial positionof these fibres at the level of the posterior clinoidand the dural penetration points also supports thesuggestion of various authors (Reid and Cone,1939; Fischer-Briigge, 1951; Lazorthes et al., 1955)that pressure against medially placed structures atthis level is responsible.
ACCOMMODATION There has been up to the presenttime no information regarding the course of fibresmediating accommodation, and although the ob-servations made during the course of this study coveronly a very short part of this pathway, they indicatethat, at least in the third nerve, the fibres for ac-commodation are intimately associated with thepupillomotor fibres.
SUMMARY
Study of the effect of compression and stimulationof the third nerve of several species of animals andhistological study of the nerves yielded the followingfindings.The third nerve is the only supratentorial structure
which on being compressed gives rise to mydriasis.Compression of the brain-stem tegmentum at the
level of the oculomotor nucleus results in miosis,not mydriasis.
The position of the pupillomotor fibres in thethird nerve has been determined by stimulation,partial neurotomies, and histological study, resultsfrom these methods being in close agreement.The fibres mediating accommodation run with the
pupillomotor fibres in the third nerve.The authors wish to express their appreciation toDrs. W. D. Backer and P. L. Archambeau for perform-ing retinoscopic examinations.
REFERENCES
Adler, F. H. (1959). Physiology of the Eye: Clinical Application,3rd ed. Mosby, St. Louis.
Adrogue, E., and Balado, M. (1925). Rev. Asoc. med. argent., 38, 252.Bergmann, E. von (1880). Die Lehre von den Kopfverletzungen. In
Billroth and Luecke. Deutsche Chirurgie, pt 30, p. 398.Enke, Stuttgart.
Browder, J., and Meyers, R. (1939). Ann Surg., 110, 357.Cushing, Harvey (1908). Surgery of the head. In Keen's Surgery, vol. 3,
p. 204. Saunders, Philadelphia.Dott, N. (1939). In Thomson and Miles' Manual of Surgery. Oxford
University Press, London.Fischer-Brugge, E. (1951). Acta neurochir. (Wien), 2, 36.Fuchs (1907). Arb. neurol. Inst. Univ. Wien, 16(2), 245.Huber, A. (1961). Eye Symptoms in Brain Tumours. Mosby, St. Louis.Hutchinson, J. (1867-1868). Clin. Lect. Rep. Lond. Hosp., 4, 10.Jennett, W. B., and Stern, W. E. (1960). J. Neurosurg. 17, 598.Keefe, W. P., Rucker, C. W., and Kernohan, J. W. (1960). Arch.
Ophthal., 63, 585.Kerr, F. W. L., and Brown, J. A. (1964). Arch. Neurol. (Chic.). 10, 262.Lazorthes, G., Pigassou, R., and Gaubert, J. (1955). Bull. Soc. belg.
Ophthal., 109, 16.Lyle, D. J. (1954). Neuro-ophthalmology, 2nd ed. Thomas, Springfield,
Illinois.Macewen, William (1887). Amer. J. med. Sci., 94, 123.Nelson, J. (1942). J. Amer. med. Ass., 119, 864.Penfield, W., and McEachern, D. (1949). In The Oxford Medicine,
vol. 6, pp. 137-216.Reid, W. L., and Cone, W. V. (1939). J. Amer. med. Ass., 112, 2030.St. Bernheimer (1897). Albrecht v. Graefes Arch. Ophthal., 44, 526.Schorcher, F. (1937). Dtsch. Z. Chir., 248, 420.Sunderland, S. (1958). Brit. J. Surg., 45, 422.
and Bradley, K. C. (1953). J. Neurol. Neurosurg. Psychiat.,16, 35.and Hughes, E. S. R. (1946). Brain, 69, 301.
Weintraub, C. M. (1960). Brit. J. Surg., 48, 62.Welte, E. (1943). Zbl. Neurochir., 8, 217.Wilson, W. C. (1949). Arch. Neurol. Psychiat. (Chic.), 61, 385.
Protected by copyright.
on Decem
ber 30, 2019 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.27.5.473 on 1 O
ctober 1964. Dow
nloaded from