local transgenic expression of granulocyte macrophage ... · local transgenic expression of...
TRANSCRIPT

of September 16, 2018.This information is current as
Initiates AutoimmunityMacrophage-Colony Stimulating Factor Local Transgenic Expression of Granulocyte
and Frank AlderuccioMark Biondo, Zeyad Nasa, Aiden Marshall, Ban Hock Toh
http://www.jimmunol.org/content/166/3/2090doi: 10.4049/jimmunol.166.3.2090
2001; 166:2090-2099; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/3/2090.full#ref-list-1
, 24 of which you can access for free at: cites 60 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Local Transgenic Expression of GranulocyteMacrophage-Colony Stimulating Factor InitiatesAutoimmunity 1
Mark Biondo, Zeyad Nasa, Aiden Marshall, Ban Hock Toh, and Frank Alderuccio2
Mechanisms leading to breakdown of immunological tolerance and initiation of autoimmunity are poorly understood. Experi-mental autoimmune gastritis is a paradigm of organ-specific autoimmunity arising from a pathogenic autoimmune response togastric H/K ATPase. The gastritis is accompanied by autoantibodies to the gastric H/K ATPase. The best characterized model ofexperimental autoimmune gastritis requires neonatal thymectomy. This procedure disrupts the immune repertoire, limiting itsusefulness in understanding how autoimmunity arises in animals with intact immune systems. Here we tested whether localproduction of GM-CSF, a pro-inflammatory cytokine, is sufficient to break tolerance and initiate autoimmunity. We generatedtransgenic mice expressing GM-CSF in the stomach. These transgenic mice spontaneously developed gastritis with an incidenceof about 80% after six backcrosses to gastritis-susceptible BALBc/CrSlc mice. The gastritis is accompanied by mucosal hyper-trophy, enlargement of draining lymph nodes and autoantibodies to gastric H/K ATPase. An infiltrate of dendritic cells andmacrophages preceded CD4 T cells into the gastric mucosa. T cells from draining lymph nodes specifically proliferated to thegastric H/K ATPase. CD4 but not CD8 T cells transferred gastritis to nude mouse recipients. CD41 CD251 T cells from the spleenretained anergic suppressive properties that were reversed by IL-2. We conclude that local expression of GM-CSF is sufficient tobreak tolerance and initiate autoimmunity mediated by CD4 T cells. This new mouse model should be useful for studies oforgan-specific autoimmunity. The Journal of Immunology,2001, 166: 2090–2099.
A utoimmune gastritis, the underlying cause of perniciousanemia in humans, is an organ-specific autoimmune dis-ease associated with loss of parietal and zymogenic cells
in the gastric mucosa (1, 2). Human gastritis is characterized bycirculating autoantibodies to thea- andb-subunits of the gastricH/K ATPase and to intrinsic factor. Experimental autoimmunegastritis (EAG)3 induced in susceptible mouse strains (3) is similarto human autoimmune gastritis, making it a relevant model tostudy organ-specific autoimmunity (1). EAG can be initiated inBALB/c mice by lymphopenia; the best characterized being thatinduced by neonatal thymectomy (4). EAG can also be initiated byimmunization with autoantigen (5) and develops spontaneously in30% of C3H/He mice (6). EAG is characterized by a chronic in-flammatory infiltrate in the gastric mucosa with loss of parietal andzymogenic cells. It is also associated with autoantibodies to thea-andb-subunits of the gastric H/K ATPase (7). Adoptive transferand in vivo depletion studies have shown that EAG is a CD41 Tcell mediated disease (8, 9) with no role for CD81 T cells (10).The early gastric lesion in EAG observed at 4 wk after neonatalthymectomy consists mainly of macrophages and CD41 T cells(11). Transgenic (tg) mice expressing the gastric H/K ATPase
b-subunit in the thymus are resistant to the development of gastricautoimmunity. These studies suggest that an immune response tothe H/K ATPaseb-subunit is necessary for disease induction (12,13) although T cell responses have been shown to both thea- andb-subunits of the H/K ATPase (14–16).
The mechanisms of disease induction in mouse models of EAGare unknown. A role has been suggested for regulatoryCD41CD251 T cells in maintaining tolerance to autoantigens suchas the gastric H/K ATPase (17–21). Sakaguchi and colleagues (18)have proposed that day 3 thymectomy prevented seeding to theperiphery of thymic-derived CD41CD251 regulatory T cells.They have shown that normal splenocytes depleted ofCD41CD251 T cells induce autoimmune gastritis when trans-ferred to nude (nu/nu) mice (17, 22). Conversely, adoptive transferof these regulatory T cells prevented autoimmunity induced byneonatal thymectomy or by adoptive transfer of pathogenic T cells(18). EAG induced by lymphopenia requires drastic manipulationof the immune system such as neonatal thymectomy. This limitstheir usefulness in studies aimed at understanding how tolerance inan intact immune system can be broken to initiate autoimmunity.
Our study was designed to address whether tolerance can bebroken and gastric autoimmunity can be initiated in mice with anintact immune system by local expression of the pro-inflammatorycytokine GM-CSF in the stomach. We selected this cytokine forlocal tissue expression because we have previously identified GM-CSF in gastric lesions of mice with EAG (11) and because GM-CSF is a key cytokine required for proliferation and differentiationof not only granulocytes and macrophages but also of dendriticcells (23). Vaccination with GM-CSF strongly augments the immuneresponse (24, 25). Too much GM-CSF can be lethal, because sys-temic tg expression of GM-CSF results in excessive accumulation andactivation of granulocytes and macrophages (26, 27). Local GM-CSFexpression has been shown to induce local inflammatory responses.For instance, mice infected with GM-CSF-expressing adenovirus
Department of Pathology and Immunology, Monash University Medical School,Prahran, Victoria, Australia
Received for publication May 4, 2000. Accepted for publication November 9, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was funded by the National Health and Medical Research Council ofAustralia and the Alfred Hospital Healthcare Group.2 Address correspondence and reprint requests to Dr. Frank Alderuccio, Department ofPathology and Immunology, Monash University Medical School, Commercial Road,Prahran, Victoria, 3181 Australia. E-mail address: [email protected] Abbreviations used in this paper: EAG, experimental autoimmune gastritis; tg, trans-genic; PC-GMCSF tg, parietal cell-GM-CSF tg;nu/nu, nude.
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

have infiltrates of granulocytes and mononuclear cells in the lung(28). Tg expression of GM-CSF in GM-CSF deficient mice (29)corrects alveolar proteinoisis associated with the deficient mice(30). Mice deficient in GM-CSF have a marked reduction in inci-dence and pathology of collagen-induced arthritis (31), while ad-ministration of GM-CSF accelerates the onset and pathology ofarthritis (32).
Here we report that local expression of GM-CSF in the stomachof gastritis-susceptible BALB/cCrSlc mice results in developmentof autoimmune gastritis associated with circulating parietal cellautoantibodies to the gastric H/K ATPase. These characteristicsare identical with those of autoimmune gastritis induced by neo-natal thymectomy, immunization, altered T cell repertoire, suble-thal irradiation, and spontaneously in C3H/He mice (5, 6, 33–35).
Materials and MethodsMice
BALB/c and (BALB/c 3 C57BL/6)F1 mice used for tg mice productionand BALB/cCrSlc were maintained at Monash University Medical Schoolanimal facilities (Victoria, Austrailia). PC-GM-CSF tg mice were back-crossed to BALB/cCrSlc at least four times.
Parietal cell GM-CSF (PC-GMCSF) transgene construction
The transgene directing GM-CSF expression to parietal cells of the stom-ach was generated as follows. The gene encoding murine GM-CSF wasisolated from pUC8 (27) byBamHI andEcoRI restriction enzyme diges-tion to release a fragment of;2.6 kbp. This included a 310-bpHaeIIIFX174 fragment that was originally cloned into theSspI site in the 39untranslated region (27). This fragment was subcloned into theSpeI site ofp11SV between 10.9 kbp of murine gastric H/K ATPaseb-subunit 59un-translated region (36) and an SV40-derived polyadenylation signal. Allfragments were blunt ended before ligation and orientation of the GM-CSFgene was determined by restriction fragment mapping. The 13.95-kbptransgene was isolated byNotI andXhoI restriction enzyme digestion andpurified on a nucleic acid chromotography system 52 column (Life Tech-nologies, Gaithersburg, MD).
Generation of PC-GM-CSF tg mice
Isolated transgene was resuspended in 10 mM Tris-HCl (pH 8.0) and 1 mMEDTA, at a concentration of 2–5 ng/ml, injected into pronuclei of fertilized(BALB/c 3 C57BL/6) 3 BALB/c oocytes and transferred to oviducts ofpsuedopregnant BALB/c mice according to the method of Hogan et al.(37). Tg founders were identified by PCR analysis of extracted mouse-tailDNA. Oligonucelotides 59-CCT CAC ACA GAG GAG ACT A-39and59-GTT AGA GAC GAC TTC TAC CTC TTC-39were designed to gen-erate a 350-bp fragment spanning the H/K ATPaseb-subunit promoter andthe GM-CSF gene. Control PCR to confirm DNA integrity contained oli-gonucleotides 59-CGA GCT CGA GCC TGC CTA TCT TTC AGG-39and59-CGG GAT CCT AGT TGC AGT AGT TCT CCA G-39designed togenerate a 374-bp product from the mouse insulin gene. PCR was per-formed in 25-ml reaction volumes containing amplification buffer; 10 mMTris-HCl (pH 8.3); 50 mM KCl; 1.5 mM MgCl2; 0.1% gelatin; 200mMdATP, dCTP, dGTP and dTTP; 50 pmols oligonucelotide primers; and 1.5U Taq DNA polymerase (Life Technologies, Melbourne, Australia). Re-action mixtures were incubated at 95°C for 2 min and 30 cycles of 92°C for30 s, 60°C for 30 s, and 72°C for 1 min, with a final cycle at 72°C for 5min. Fifteen-microliter samples of PCR product were separated by agarosegel electrophoresis and visualized using UV illumination. Images werecaptured by digital camera and inverted for publication.
Messenger RNA analysis
Transgene expression of GM-CSF mRNA was detected by RT-PCR es-sentially as previously described (13). Total RNA was isolated from lung,spleen, heart, liver, and stomach using the Ultraspec II RNA isolation kit(Biotecx Laboratories, Houston, TX). Briefly, 2mg of total RNA was re-verse transcribed with Moloney murine leukemia virus reverse transcrip-tase (Life Technologies, Gaithersburg, MD) using oligo(dT) primer in atotal volume of 20ml. Two microliters of the reaction mixture was sub-jected to PCR using primers designed to amplify the tg GM-CSF or actincDNA. Actin primers were 59-ATGGATGACGATATCGCTG-39and 59-ATGAGGTAGTCTGTCAGGT-39and generated a product of 568 bp. TgGM-CSF was detected following two rounds of PCR. The first-round prim-
ers were sense 59-CTATAAGCCCTAGAGGACGC-39and anti-sense 59-CCGCATAGGTGGTAACTTGTGTTTC-39to generate a product of 400bp. In the second-round PCR, 2ml of PCR product were amplified using aninternal anti-sense primer 59-GGCAGTATGTCTGGTAGTAGCTGG-39to generate a product of 275 bp. PCR was comprised of 1 cycle at 92°C for2 min and 30 cycles at 92°C for 30 s, 60°C for 30 s, and 72°C for 1 min,with a final cycle at 72°C for 5 min. PCR products were subjected toagarose gel electrophoresis and visualized by UV illumination. Imageswere captured by digital camera and inverted for publication.
ELISA, indirect immunofluorescence, and flow cytometry
Circulating anti-H/K ATPase autoantibodies were assayed by ELISA on96-well plates coated with purified pig H/K ATPase as previously de-scribed (13). Anti-parietal cell autoantibodies were detected by indirectimmunofluorescence on frozen or paraffin-embedded sections of normalmouse stomach (13). Gastric H/K ATPasea- andb-subunit reactivity wasdetected by immunofluorescence reactivity with Sf9 cells infected withbaculovirus encoding rat ATPasea- or b-subunit. Recombinant baculovi-rus was obtained from E. Shevach (National Institutes of Health, Bethesda,MD). mAbs 1H9 and 2B6, reactive with the gastric H/K ATPasea-subunitandb-subunit, respectively, were used as controls.
Immunohistochemistry was performed on frozen tissue sections withAbs reactive with CD4 T cells (FITC-anti-CD4; clone RM4-5), CD8 Tcells (FITC-anti-CD8; clone 53-6.7), dendritic cells (FITC-anti-CD11c;clone HL3), macrophages (FITC-anti-CD11b; clone M1/70), B cells (anti-B220; clone RA3.3A1) and granulocytes (FITC-anti-Gr1; clone RB6–8C5). Sections were blocked with 1% normal swine serum for 10 min atroom temperature and incubated with Ab for 60 min at room temperature.Sections were washed twice with PBS/1% Tween 20 for 5 min andmounted. To visualize parietal cells, sections were double stained togetherwith biotinylatedDolichos biflorus(Sigma, St. Louis, MO; Ref. 38) fol-lowed by streptavidin-Texas Red. Sections were viewed with a Bio-Rad(Richmond, CA) confocal microscope.
For FACS analysis, 1–23 106 cells were stained in 30-ml volumescontaining APC-anti-CD4 (clone RM4-5), PerCP-anti-CD8 (clone 53-6.7),PE-anti-B220 (clone RA3.3A1) and PE-anti-CD25 (IL-2a-chain, clonePC61) diluted in HBSS/1% FCS. Cells were analyzed on a FACScaliberusing CellQuest software (Becton Dickinson, Mountain View, CA).
Histology
Tissues were fixed in 10% formalin in PBS and embedded in paraffin.Five-micrometer stomach sections were stained with hematoxylin and eo-sin and viewed by light microscopy. Gastritis was assessed by the presenceof cellular infiltrate within the gastric mucosa. Destructive gastritis com-prised the presence of cellular infiltrate within the gastric mucosa withdestruction of the cells within gastric glands. Other tissues were also ex-amined for the presence of pathology.
Gastric and liver membranes and purified gastric H/K ATPase
Purified porcine gastric H/K ATPase was prepared by tomato-lectin chro-motography as previously described (39). Mouse gastric and liver mem-branes were prepared as follows. Tissues were homogenized in ice-coldsucrose buffer (0.25 M sucrose, 2 mM EDTA, 5 mM Tris (pH 7.5), 1 mMPMSF) with a polytron homogenizer (Kinematica, Lucerne, Switzerland).Samples were centrifuged at 3603 g for 10 min at 4°C to remove nucleiand cell debris. The supernatant was centrifuged at 55003 g for 15 min at4°C to pellet mitochondria. The supernatant was collected and centrifugedat 100,0003 g for 1 h at 2°C to pellet membranes. Membranes wereresuspended in cold HEPES buffer (50 mM HEPES (pH 7.6), 1 mMEDTA, 1 mM PMSF) and protein concentration determined using microBCA protein assay (Pierce, Rockford, IL). Samples were stored at220°C.Presence of H/K ATPase in gastric membranes was confirmed by ELISAreactivity with mAbs 1H9 and 2B6 specific for gastric H/K ATPasea- andb-subunits, respectively (40).
In vitro proliferation assay
Pooled single cell suspensions of lymphocytes from tg (n 5 5) and non-tg(n 5 6) littermates were prepared by gently grinding lymphoid tissuesbetween frosted glass slides and used as responders in in vitro proliferationassays. Non-tg splenocytes were treated with ammonium chloride solution(0.9%) to lyse RBC and irradiated (3000 rads) for use as APCs. Cells weresuspended in RPMI 1640 culture media supplemented with 10% FCS, 100U/ml penicillin, 100mg/ml streptomycin, 2 mML-glutamine, and 50mM2-ME. Proliferation assays were performed in 96-well tissue culture plates(Dynex Technologies, Chantilly, VA) in a total volume of 200ml contain-ing 2.53 105 responder cells, 2.53 105 irradiated APCs, and Ag. For in
2091The Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

vitro assay for CD41CD251 regulatory T cells, splenocytes were sortedinto CD41CD251 and CD41CD22 populations, with purity of 99% and98%, respectively. Proliferation assay was performed in a total volume of200 ml containing 23 104 responders, 53 104 irradiated APCs, and Ag.Con A was used at a final concentration of 3mg/ml and IL-2 was used at100 U/ml. Cells were incubated for 48 h at 37°C/10% CO2 followed by anadditional overnight incubation in the presence of 1mCi [3H]thymidine(NEN, Boston, MA). Cells were harvested onto glass filters (Skatron, Ster-ling, VA) suspended in scintillant and [3H]thymidine incorporation wasdetermined on a Wallac 1205 Betaplate liquid scintillation counter (Phar-macia, Uppsala, Sweden). Control wells were comprised of responder cellsalone, APCs alone or proliferation in the absence of Ag.
Cell transfer studies
Single-cell suspensions were prepared from pooled spleens, paragastriclymph nodes, inguinal lymph nodes, and stomachs (41) in HBSS/1% FCS.Two groups of PC-GMCSF tg (n5 6 andn 5 3) mice with circulatingparietal cell and H/K ATPase Abs and one group of non-tg (n5 3) litter-mates were used in these experiments. Splenocytes were treated with am-monium chloride solution (0.9%) to lyse RBC. Cells to be injected werewashed and resuspended in HBSS in a total volume of 150–200ml. Cellswere transferred to BALB/cnu/numice by i.v. tail vein injection. Recipientmice received 43 107 splenocytes, 1–23 107 inguinal lymph node cells,1.3–23 107 paragastric lymph node cells, or 53 106 stomach infiltratecells. Mice were killed at 8–12 wk following cell transfer and sera wereanalyzed for H/K ATPase and parietal cell autoantibodies. Stomachs andother tissues were processed for paraffin-embedded sections and examinedby histology for gastritis.
For transfer of purified CD41 and CD81 T cells, pooled paragastriclymph node cells were isolated in two separate experiments from six andfive PC-GMCSF tg mice with circulating parietal cell and H/K ATPaseAbs. Cells were stained with anti-CD4-PE and anti-CD8-FITC and sortedusing a FACScaliber cell sorter (Becton Dickinson). Analysis of sortedpopulations revealed a purity for CD41 and CD81 T cells of 97% and98%, respectively. Each BALB/cnu/nu recipient received 1.5–23 106
cells by i.v. injection. Eight weeks following transfer, mice were killed andexamined as described above.
Statistical analysis
Cell numbers and populations were compared using a two-tailedttest. Cell transfer results were compared using a Fisher’s exact test.
ResultsGeneration of PC-GMCSF tg mice
PC-GMCSF tg mice were generated by injecting into fertilizedmouse embryos, a transgene composed of murine gastric H/KATPaseb-subunit promoter and mouse GM-CSF gene (Fig. 1A).We have previously shown that expression of OVA orb-galacto-sidase under the control of the same murine H/K ATPaseb-sub-unit promoter does not induce spontaneous autoimmune gastritis inmice (K. Scarff, unpublished observations). Tg mice were identi-fied by PCR using primers spanning the H/K ATPaseb-subunitpromoter and GM-CSF gene junction (Fig. 1,A and B). DNAintegrity was assessed by PCR amplification of mouse insulin gene(Fig. 1B). Demonstration of GM-CSF expression was attempted byimmunohistochemistry with no success (data not shown). RT-PCRwas then used to demonstrate tg expression of GM-CSF in stom-achs of PC-GMCSF tg mice. RNA isolated from tg and non-tglung, spleen, heart, liver, and stomach was subjected to RT-PCR.The quality and integrity of the cDNA generated was assessed byPCR amplification of mouseb-actin gene (Fig. 1C). Expression oftg GM-CSF was observed only in tg stomachs and not in other tgor non-tg tissues (Fig. 1C). Two rounds of PCR were required tovisualize mRNA expression suggesting low levels of transgenemRNA or as previously observed, a short biological half-life ofGM-CSF mRNA (30, 42).
PC-GMCSF tg mice develop spontaneous gastritis and H/KATPase autoantibodies
Tg and non-tg mice backcrossed four times to gastritis-susceptibleBALB/cCrSlc mice were examined at 12 wk of age for gastritisand autoantibodies to gastric parietal cells H/K ATPase. Six of 16(38%, p 5 0.02) PC-GMCSF tg mice developed gastric H/KATPase reactive Abs assessed by ELISA, compared with 0/16non-tg littermates (Fig. 2). ELISA-positive sera reacted by immu-nofluorescence with baculovirala- and b-subunits of the gastric
FIGURE 1. Generation and analysis of PC-GMCSF tg mice.A, The parietal cell GM-CSF transgene was constructed by ligating the murine GM-CSFgene to the 10.9-kbp H/K ATPaseb-subunit 59nontranslated region. The black and open boxes indicate the GM-CSF exons and introns, respectively. ThefX174 fragment previously introduced into the gene (27) is indicated by the speckled box and the SV40-derived poly(A) signal by a striped box.NotI andXhoI sites were used to excise the transgene from the cloning vector and position of oligonucleotides used for PCR screening and RT-PCR analysis areindicated.B, PCR was used to discriminate between tg and non-tg (N-tg) mice. Oligonucleotides A and A9were designed to span the H/K ATPaseb-subunitand GM-CSF gene to amplify a product of 350 bp. DNA integrity was confirmed by PCR analysis of the mouse insulin gene and inclusion of positive (1ve)and negative (-ve) DNA controls. M, DNA markers.C, RT-PCR analysis was used to confirm expression of tg GM-CSF (Tg-GM-CSF) in the stomachsof tg mice. Total RNA was isolated from lung (L), spleen (Sp), liver (Liv), heart (H), and stomach (St) of tg and non-tg mice. mRNA was reversedtranscribed and used in two rounds of PCR. Oligonucleotides B/B9 were used in the first round followed by oligonucleotides B/B0 in the second reactionto produce a product of 275 bp. PCR of actin mRNA was used to confirm integrity of RNA.
2092 AUTOIMMUNE GASTRITIS IN GM-CSF TRANSGENIC MICE
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
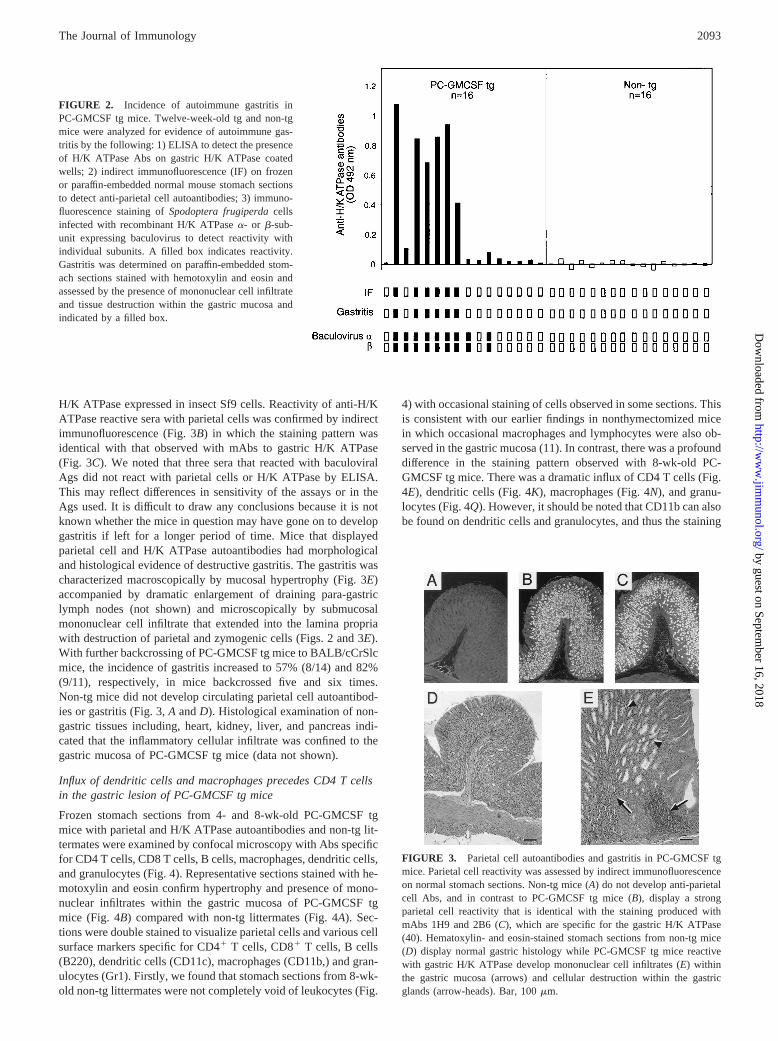
H/K ATPase expressed in insect Sf9 cells. Reactivity of anti-H/KATPase reactive sera with parietal cells was confirmed by indirectimmunofluorescence (Fig. 3B) in which the staining pattern wasidentical with that observed with mAbs to gastric H/K ATPase(Fig. 3C). We noted that three sera that reacted with baculoviralAgs did not react with parietal cells or H/K ATPase by ELISA.This may reflect differences in sensitivity of the assays or in theAgs used. It is difficult to draw any conclusions because it is notknown whether the mice in question may have gone on to developgastritis if left for a longer period of time. Mice that displayedparietal cell and H/K ATPase autoantibodies had morphologicaland histological evidence of destructive gastritis. The gastritis wascharacterized macroscopically by mucosal hypertrophy (Fig. 3E)accompanied by dramatic enlargement of draining para-gastriclymph nodes (not shown) and microscopically by submucosalmononuclear cell infiltrate that extended into the lamina propriawith destruction of parietal and zymogenic cells (Figs. 2 and 3E).With further backcrossing of PC-GMCSF tg mice to BALB/cCrSlcmice, the incidence of gastritis increased to 57% (8/14) and 82%(9/11), respectively, in mice backcrossed five and six times.Non-tg mice did not develop circulating parietal cell autoantibod-ies or gastritis (Fig. 3,A andD). Histological examination of non-gastric tissues including, heart, kidney, liver, and pancreas indi-cated that the inflammatory cellular infiltrate was confined to thegastric mucosa of PC-GMCSF tg mice (data not shown).
Influx of dendritic cells and macrophages precedes CD4 T cellsin the gastric lesion of PC-GMCSF tg mice
Frozen stomach sections from 4- and 8-wk-old PC-GMCSF tgmice with parietal and H/K ATPase autoantibodies and non-tg lit-termates were examined by confocal microscopy with Abs specificfor CD4 T cells, CD8 T cells, B cells, macrophages, dendritic cells,and granulocytes (Fig. 4). Representative sections stained with he-motoxylin and eosin confirm hypertrophy and presence of mono-nuclear infiltrates within the gastric mucosa of PC-GMCSF tgmice (Fig. 4B) compared with non-tg littermates (Fig. 4A). Sec-tions were double stained to visualize parietal cells and various cellsurface markers specific for CD41 T cells, CD81 T cells, B cells(B220), dendritic cells (CD11c), macrophages (CD11b,) and gran-ulocytes (Gr1). Firstly, we found that stomach sections from 8-wk-old non-tg littermates were not completely void of leukocytes (Fig.
4) with occasional staining of cells observed in some sections. Thisis consistent with our earlier findings in nonthymectomized micein which occasional macrophages and lymphocytes were also ob-served in the gastric mucosa (11). In contrast, there was a profounddifference in the staining pattern observed with 8-wk-old PC-GMCSF tg mice. There was a dramatic influx of CD4 T cells (Fig.4E), dendritic cells (Fig. 4K), macrophages (Fig. 4N), and granu-locytes (Fig. 4Q). However, it should be noted that CD11b can alsobe found on dendritic cells and granulocytes, and thus the staining
FIGURE 2. Incidence of autoimmune gastritis inPC-GMCSF tg mice. Twelve-week-old tg and non-tgmice were analyzed for evidence of autoimmune gas-tritis by the following: 1) ELISA to detect the presenceof H/K ATPase Abs on gastric H/K ATPase coatedwells; 2) indirect immunofluorescence (IF) on frozenor paraffin-embedded normal mouse stomach sectionsto detect anti-parietal cell autoantibodies; 3) immuno-fluorescence staining ofSpodoptera frugiperdacellsinfected with recombinant H/K ATPasea- or b-sub-unit expressing baculovirus to detect reactivity withindividual subunits. A filled box indicates reactivity.Gastritis was determined on paraffin-embedded stom-ach sections stained with hemotoxylin and eosin andassessed by the presence of mononuclear cell infiltrateand tissue destruction within the gastric mucosa andindicated by a filled box.
FIGURE 3. Parietal cell autoantibodies and gastritis in PC-GMCSF tgmice. Parietal cell reactivity was assessed by indirect immunofluorescenceon normal stomach sections. Non-tg mice (A) do not develop anti-parietalcell Abs, and in contrast to PC-GMCSF tg mice (B), display a strongparietal cell reactivity that is identical with the staining produced withmAbs 1H9 and 2B6 (C), which are specific for the gastric H/K ATPase(40). Hematoxylin- and eosin-stained stomach sections from non-tg mice(D) display normal gastric histology while PC-GMCSF tg mice reactivewith gastric H/K ATPase develop mononuclear cell infiltrates (E) withinthe gastric mucosa (arrows) and cellular destruction within the gastricglands (arrow-heads). Bar, 100mm.
2093The Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
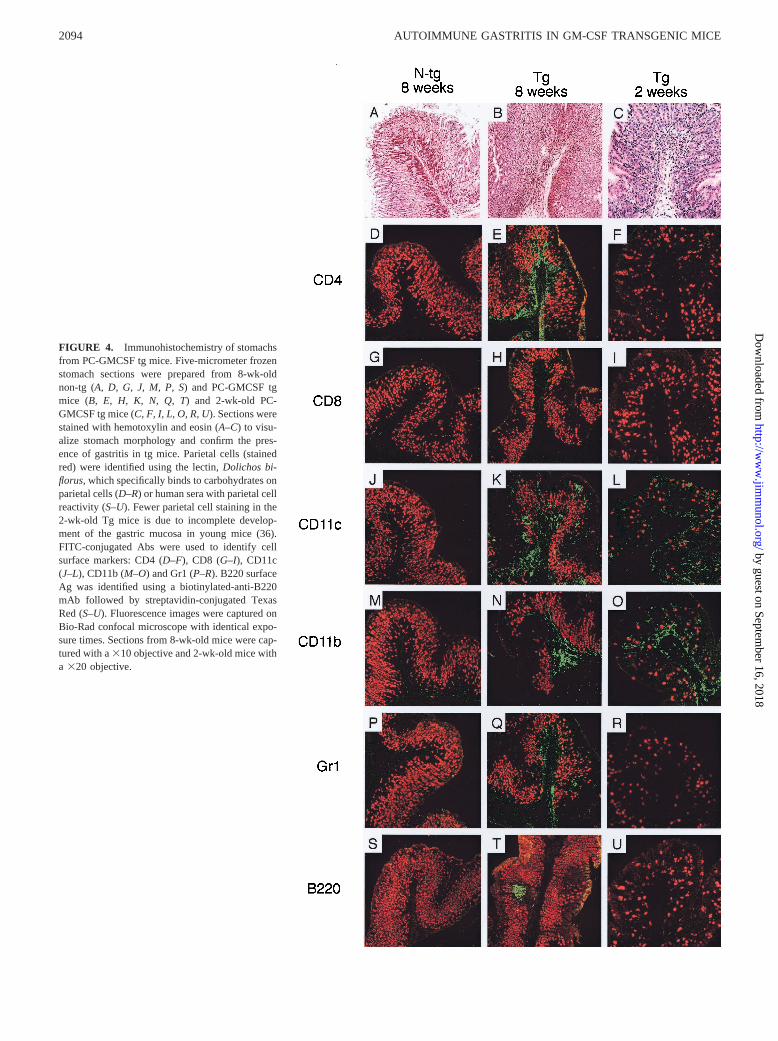
FIGURE 4. Immunohistochemistry of stomachsfrom PC-GMCSF tg mice. Five-micrometer frozenstomach sections were prepared from 8-wk-oldnon-tg (A, D, G, J, M, P, S) and PC-GMCSF tgmice (B, E, H, K, N, Q, T) and 2-wk-old PC-GMCSF tg mice (C, F, I, L, O, R, U). Sections werestained with hemotoxylin and eosin (A–C) to visu-alize stomach morphology and confirm the pres-ence of gastritis in tg mice. Parietal cells (stainedred) were identified using the lectin,Dolichos bi-florus, which specifically binds to carbohydrates onparietal cells (D–R) or human sera with parietal cellreactivity (S–U). Fewer parietal cell staining in the2-wk-old Tg mice is due to incomplete develop-ment of the gastric mucosa in young mice (36).FITC-conjugated Abs were used to identify cellsurface markers: CD4 (D–F), CD8 (G–I), CD11c(J–L), CD11b (M–O) and Gr1 (P–R). B220 surfaceAg was identified using a biotinylated-anti-B220mAb followed by streptavidin-conjugated TexasRed (S–U). Fluorescence images were captured onBio-Rad confocal microscope with identical expo-sure times. Sections from 8-wk-old mice were cap-tured with a310 objective and 2-wk-old mice witha 320 objective.
2094 AUTOIMMUNE GASTRITIS IN GM-CSF TRANSGENIC MICE
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

observed in Fig. 4Nmay not be entirely associated with macro-phages. In some sections, B cells were observed in follicle-likeaggregates (Fig. 4T) similar to that observed in the neonatalthymectomy model of EAG. CD8 T cells were not present (Fig.4H), which is also similar to the neonatal thymectomy model (11,41) in which CD8 T cells do not appear to be implicated in thepathogenesis of EAG (10). To identify early cellular events asso-ciated with the initiation of autoimmunity, we examined stomachsof 4- (with H/K ATPase reactivity) and 2-wk-old tg mice. Immu-nohistochemical analysis of the stomachs of 4-wk-old PC-GMCSFtg mice showed the presence of a heterogeneous cellular infiltratesimilar to that observed in the 8-wk-old group except that B cellfollicles were not observed (data not shown). This indicates thatautoimmune gastritis in this model is well established by 4 wk ofage and, therefore, accelerated compared with the neonatalthymectomy model in which only a minority of mice display ev-idence of disease at 4 wk of age (11). However, examination of2-wk-old tg mice did reveal a difference in the composition of thecellular infiltrate. Of the leukocyte markers examined, staining wasobserved only with CD11c and CD11b (Fig. 4,L andO) markers.No staining was observed for CD4 or CD8 T cells, B cells orgranulocytes (Fig. 4,F, I, U, andR). Taken together, these obser-vations support our hypothesis that in PC-GMCSF tg mice the firststage of autoimmune gastritis is associated with proliferation andactivation of APCs, which then leads to activation and migration ofpathogenic CD4 T cells into the gastric mucosa.
T cells from PC-GMCSF tg mice proliferate to gastricmembranes and H/K ATPase
Cell suspensions from paragastric lymph nodes were pooled fromfive H/K ATPase-reactive tg mice and from six nonreactive non-tglittermates. Cells were challenged in vitro with preparations ofgastric or liver membranes or purified gastric H/K ATPase. T cellsfrom tg mice responded specifically to gastric membranes and toH/K ATPase and not to liver membranes (Fig. 5A). Non-tg T cellsdid not respond to gastric or liver membranes or to gastric H/KATPase (Fig. 5A). These observations suggest that the responseto gastric membranes is likely to be directed to the gastric H/KATPase. We next determined whether the T cell response wasconfined to cells isolated from the paragastric lymph node, orwhether T cells isolated from other lymphoid organs can respondto gastric membranes. Cells were isolated from the spleen andfrom the paragastric, mesenteric, and inguinal lymph nodes. Aproliferative response was observed only with T cells isolated fromthe paragastric lymph node (Fig. 5B). None of the cell preparationsresponded to liver membranes, included as controls (Fig. 5B).From these results it appears that PC-GMCSF T cells proliferatespecifically to the H/K ATPase of gastric membranes and that theresponse is confined to cells from the paragastric lymph node.
CD41 T cells from PC-GMCSF tg mice transfer gastritis
To determine whether PC-GMCSF tg mice can adoptively transfergastritis, cells isolated from pooled tg and non-tg stomach, spleen,or paragastric or inguinal lymph nodes were transferred to BALB/cnu/numice in two separate experiments. Eight to 12 wk followingtransfer, recipient mice were killed, sera were assessed for H/KATPase, and parietal cell autoantibodies and stomachs were ex-amined for gastritis (Fig. 6). A destructive gastritis was observedin mice that received cells from tg spleen (1/2) (Fig. 6A), inguinallymph nodes (3/5), paragastric lymph nodes (2/5), and stomach(1/1); the gastritis was associated with circulating autoantibodiesto parietal cells demonstrated by indirect immunofluorescence(Fig. 6B). These observations indicate that pathogenic lympho-
cytes are not confined to the paragastric lymph node or stomach,but were present in all lymphoid sources examined. Cells fromnon-tg spleen (n5 1), inguinal lymph node (n5 1), and paragas-tric lymph node (n5 1) did not induce gastritis innu/numice (datanot shown). This is consistent with previous observations that cellsfrom normal mice do not transfer gastritis (9, 20, 21). To determinewhether the pathogenic cells were CD41 or CD81 T cells, CD41
and CD81 cells from pooled paragastric lymph nodes were sortedby flow cytometry and transferred to syngeneicnu/numice. Pooleddata from two experiments indicate that mice transferred withCD41 T cells develop gastritis with circulating autoantibodies toparietal cells (5/6) while mice transferred with CD81 T cells re-mained disease free (0/4,p 5 0.048) (data not shown). The findingthat CD41 T cells transferred autoimmune gastritis is similar tothat observed in EAG induced by neonatal thymectomy (10), inwhich CD41 T cells have been shown to be the pathogenic T cellsresponsible for disease.
FIGURE 5. In vitro proliferation of PC-GMCSF T cells to gastric mem-branes and H/K ATPase.A, Paragastric lymph node cells were pooled fromfour PC-GMCSF tg and from six non-tg mice and proliferation was deter-mined in response to gastric or liver membranes or purified H/K ATPase.In each well, 2.53 105 responder cells were incubated with 2.53 105
irradiated normal splenocytes as APCs and Ag at various concentrations.Cells were incubated for 48 h followed by overnight incubation with 1mCiof [3H]thymidine. Each point represents the mean of duplicate wells. Con-trol wells included responder cells alone (1400 cpm), APCs alone 141cpm), and responders plus APCs in the absence of Ag (1269 cpm), withmean cpm indicated.B, Single-cell suspensions from spleen, paragastric,inguinal, and mesenteric lymph nodes were assessed for in vitro prolifer-ation to gastric and liver membranes. Assay conditions were as describedabove. Controls included responder cells alone (600–966 cpm), APCsalone (369 cpm), and responders plus APCs in the absence of Ag (500–800 cpm) with mean cpm indicated.
2095The Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Lymphocyte and CD41CD251 regulatory T cell populations arenot perturbed in PC-GMCSF tg mice
The majority of mouse models of EAG result from an inducedstate of lymphopenia; be it neonatal thymectomy (43), repeatedlymphoid irradiation (35), or tg skewing of the T cell repertoire(34). Therefore, total cell numbers and lymphocyte populationswere analyzed in 6- to 8-wk-old PC-GMCSF tg to determinewhether there were any differences in these parameters comparedwith non-tg mice. We found no difference in total number of cellsrecovered from the thymus (tg,n 5 4, 1.486 0.5 3 108; non-tg,n 5 4, 1.626 1.833 108; p 5 0.89), spleen (tg, 1.796 0.43 108;non-tg, 1.856 0.313 108; p 5 0.81), and inguinal lymph nodes(tg, 3.906 2.743 106; non-tg, 2.866 1.593 106; p 5 0.54). Notunexpectedly, given the dramatic enlargement of the draining para-gastric lymph node of tg mice, we observed a corresponding 10-fold increase in the total number of cells recovered from this site(tg, 5.536 2.083 106; non-tg, 0.586 0.193 106; p 5 0.017).FACS analysis of thymocyte and splenocyte populations usingCD4, CD8, and B220 markers revealed no difference in these pop-ulations (data not shown). These data indicate that the induction ofautoimmune gastritis was not associated with alteration or skewingof T or B lymphocyte populations.
Thymic-derived CD41CD251 T cells have been implicated inthe regulation of pathogenic CD4 T cells in mouse models of au-toimmune gastritis (18–21). Therefore, we examined for the pres-ence of this regulatory population of cells in our tg mice. We foundno difference in the proportions of thymic or peripheral popula-tions of CD41CD251 cells of tg and non-tg mice (data notshown). To address whether there was any functional alteration inthis population, we tested the in vitro proliferative and regulatory
property of these cells (20, 44). CD41CD251 and CD41CD252
cells were purified from pooled spleens of three gastritic PC-GMCSF tg mice. Although CD41CD252 cells readily proliferatedto Con A, CD41CD251 regulatory cells were resistant to Con Astimulation (Fig. 7). Furthermore, CD41CD251 regulatory cellsinhibited proliferation of CD41CD252 cells to Con A stimulationand this inhibition was abrogated by IL-2 (Fig. 7). These charac-teristics are similar to those previously described for this regula-tory population (20, 44) and indicate that autoimmune gastritis inPC-GMCSF tg mice is not due to global perturbation of theCD41CD251 regulatory population.
DiscussionEAG in mice is an animal model of human autoimmune gastritis,sharing the same target autoantigens, gastric pathology, and cir-culating autoantibodies to thea- andb-subunits of the gastric H/KATPase (1). The best characterized EAG models are those inducedby lymphopenia (4). These models are of limited use for studiesdirected toward addressing how tolerance can be broken and au-toimmunity initiated in animals with an intact immune system.
In this study, we generated tg mice that locally expressed GM-CSF in the stomach under the control of the gastric H/K ATPaseb-subunit promoter (36). PC-GMCSF tg mice spontaneously de-veloped characteristics of autoimmune gastritis with an incidenceincreasing from 40% in mice backcrossed four times to;80% inmice backcrossed six times to gastritis-susceptible BALB/c/CrSlcmice. Circulating autoantibodies to gastric parietal cells were gen-erated that reacted with thea- andb-subunits of the gastric H/KATPase. Stomachs from these PC-GMCSF tg mice with parietalcell Abs displayed an inflammatory infiltrate in the gastric mucosa.The infiltrates extended into the lamina propria with accompany-ing destruction of mucosal parietal and zymogenic cells. The gas-tric hypertropy observed in PC-GMCSF tg mice is similar to that
FIGURE 6. Transfer of autoimmune gastritis from PC-GMCSF tg miceto BALB/c nu/numice. Splenocytes from PC-GMCSF tg mice with pari-etal cell reactive autoantibodies were transferred i.v. to recipient BALB/cnu/numice. Eight-wk following transfer, mice were killed and assessed forparietal cell autoantibodies and histological evidence of gastritis.Nu/numice transferred with splenocytes from tg mice developed gastritis (A) withcharacteristic mononuclear cell infiltrate within gastric mucosa (arrow-heads) and cellular destruction within gastric glands (arrows) and autoan-tibodies (B) reactive with parietal cells by immunofluorescence and to H/KATPase by ELISA (not shown).
FIGURE 7. In vitro CD41CD251 regulatory T cell assay.CD41CD251 and CD41CD22 cells from pooled splenocytes of three gas-tric PC-GMCSF tg mice were sorted by flow cytometry to 99% and 98%purity respectively (not shown). Each population was assayed in vitro forproliferative response to 3mg/ml Con A in the absence or presence of 100U/ml IL-2. The ability of CD41CD251 cells to prevent proliferation ofCD41CD22 cells to Con A stimulation was assessed in mixed cultureassays with reducing proportion of CD41CD251 cells. Abrogation ofCD41CD251-induced regulation was shown by addition of IL-2. Cellswere incubated for 48 h followed by overnight incubation with 1mCi of[3H]thymidine. Each point represents the mean of duplicate wells.
2096 AUTOIMMUNE GASTRITIS IN GM-CSF TRANSGENIC MICE
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

described in other models of EAG (8, 35, 43) and has been attrib-uted to the replacement of parietal and zymogenic cells with pro-liferating epithelial stem cells (7). Lymphoid cells recovered fromthe draining paragastric lymph nodes of PC-GMCSF tg mice spe-cifically proliferated in response to stimulation with gastric Agsand purified H/K ATPase. CD4, but not CD8, T cells transferredgastritis to syngeneicnu/numice, These observations confirm theimmunological nature of the gastritis. These characteristics areidentical with the autoimmune gastritis observed in mice followingneonatal thymectomy (13, 33, 43, 45), suggesting that the immu-nopathology in both models are similar.
GM-CSF is a pleiotrophic cytokine that stimulates proliferationand maturation of macrophages and dendritic cells (23, 46, 47). Inprevious studies we have shown that GM-CSF is expressed in thegastric lesion of mice with EAG induced by neonatal thymectomy(11). A role for GM-CSF in autoimmune pathology has previouslybeen suggested in collagen-induced arthritis in mice (31, 32). Al-though the exact mechanism by which GM-CSF expression in-duces autoimmune gastritis is not known, we suggest that localexpression of GM-CSF transgene in the gastric mucosa has initi-ated an autoimmune response through activation of local APCs,most likely dendritic cells. In normal mice, immature APCs areimplicated in the removal of apoptotic cells resulting from cellularturnover in the stomach (7); and in itself, this process would not beexpected to result in activation of these APCs (48). In the presenceof a pro-inflammatory cytokine such as GM-CSF, gastric APCsmay be activated directly (49) or through the action of other in-duced cytokines such as TNF-a (50) and migrate to the local drain-ing lymph node where they activate naive CD41 T cells specificfor the gastric H/K ATPase. The proposed effect of GM-CSF inenhancing dendritic cell activation and presentation has previouslybeen exploited in designing vaccines against pathogens and tumors(24, 51, 52). The earlier influx of CD11c and CD11b reactive cellswithin the gastric mucosa of PC-GMCSF tg mice observed at 2 wkafter birth preceding the influx of CD4 T cells observed at 4 wksupports a role for these APCs in the initiation of gastric autoim-munity. It is not known whether these cells are recruited to thegastric mucosa, arise from local proliferation or both. Similar find-ings have been observed following intramuscular injections withplasmids encoding GM-CSF (53). In these studies, expression ofGM-CSF in muscle resulted in local accumulation of macro-phages, dendritic cells and granulocytes but not CD4 T cells. Thelack of CD4 T cells in the DNA vaccination study compared withour findings may reflect differences in the time span of the twoexperiments. The finding that an in vitro T cell proliferative re-sponse was only observed with T cells isolated from the localdraining paragastric lymph node and not from other lymphoid or-gans supports the suggestion that activated APCs migrate to thedraining lymph node to activate naive T cells homing to the lymphnode. As with other models of autoimmune gastritis (8–10), thetransfer and histochemical studies in this report suggest that CD41
T cells are the pathogenic cells in EAG. Ag presentation and ac-tivation of T cells may also occur in the gastric mucosa, as hasbeen suggested in animal models of diabetes (54). Certainly, wehave observed organized lymphoid structures similar to those de-scribed by Ludewig and colleagues (54) in the gastric mucosa ofmice with thymectomy-induced EAG (11) and also in the PC-GMCSF mice in the present study.
Local tg expression of other pro-inflammatory cytokines such asTNF-a, IFN-g, and IL-2 have previously been attempted to induceother models of organ-specific autoimmunity. For instance, ex-pression of IL-2 or TNF-a in pancreatic islets of BL/6 mice re-sulted in insulitis without diabetes (55, 56), while in the NODmouse, islet expression of TNF-a accelerated diabetes onset (50).
These studies indicate that TNF-a expression in islets can promotelocal inflammation or aggravate diabetes in diabetes-prone mice.However, whether TNF-a can promote by itself a destructive au-toimmune lesion is not clear. Although IFN-g expression in pan-creatic islets resulted in diabetes, it is unclear whether this is aconsequence of autoimmunity or the result of local destructioninduced by IFN-g (57). In the present study, we show for the firsttime that expression of the pro-inflammatory cytokine, GM-CSF,in the gastric environment of gastritis-susceptible mice has in-duced autoimmune gastritis. This was confirmed by production ofautoantibodies to the gastric H/K ATPase, a specific T cell re-sponse to the ATPase and transfer of disease by CD4 T lympho-cytes from gastritic tonu/numice. These observations suggest thatexpression of GM-CSF in the local environment of the stomach issufficient to break tolerance and initiate autoimmunity.
A defining feature of our study is that tolerance to gastric H/KATPase has been broken without a major perturbation of the im-mune system, a feature associated with lymphopenic models ofEAG (4, 58). EAG is not observed in normal BALB/c or BALB/cCrSlc mice. Experimental evidence suggests a role for regulatoryCD41CD251 T cells in maintaining tolerance and that removal ofthis population from the normal repertoire renders the remaininglymphocytes pathogenic (17, 18). This is not the case in PC-GMCSF tg mice because we found CD41CD251 cells in the thy-mus and in the periphery of these mice. In vitro, we found thatCD41CD251 T cells from the spleens of tg mice were anergic,could prohibit proliferation of CD41CD252 cells and their anergycould be reversed by exogenous IL-2. These properties are iden-tical with those previously described for CD41CD251 regulatorycells (20, 44). Therefore, in PC-GMCSF tg mice, it appears thatlocal production of GM-CSF in the stomach has initiated a patho-genic autoimmune response, and overcome suppressor activity ofCD41CD251 regulatory T cells. Our observation that cells fromthe draining paragastric lymph node can be specifically stimulatedto proliferate in the presence of gastric Ags supports this sugges-tion. The observation that autoimmune gastritis can be transferredfrom lymphoid sites other than the draining lymph node seems tocontradict this. This apparent discrepancy may reflect the ability ofcirculating activated autoreactive T cells to expand followingtransfer to an “empty” periphery innu/nu mice. In vitro,CD41CD251 regulatory cells are naturally anergic and do notproliferate in response to stimulation through the TCR. However,the anergy and suppressive activity of CD41CD251 T cells can beabrogated by IL-2 coupled with TCR stimulation (20, 44). There-fore, it is possible that in PC-GMCSF tg mice, local IL-2 generatedby activated self-reactive CD4 T cells in the paragastric lymphnode may have rendered CD41CD251 regulatory cells nonfunc-tional. The presence of this regulatory population may also explainwhy not all PC-GMCSF tg mice develop autoimmunity. However,with the incidence of gastritis approaching 100% with increasingnumbers of backcrosses to gastritis-susceptible BALB/c/CrSlcmice, local expression of GM-CSF alone may be sufficient to breaktolerance and initiate autoimmunity in this mouse strain.
The PC-GMCSF tg mice generated in this study will permit usto address several important questions pertaining to the develop-ment of organ-specific autoimmunity. We have shown for the firsttime that local expression of a proinflammatory cytokine in thestomach of genetically susceptible mice can induce a damagingautoimmune response. Our observations support the recent find-ings of Sarvetnick and colleges (59) that bystander damage result-ing from responses to “danger” signals (60, 61) may be sufficientto initiate autoimmunity. Taken together, these observations sup-port our proposed genesis of the gastric lesion in autoimmune gas-tritis initiated by the gastric H/K ATPase (62). The induction of
2097The Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

damaging autoimmunity in the presence of an intact immune sys-tem will make this model useful for understanding mechanismsassociated with the breakdown of immune regulation in the locallymph nodes leading to autoimmunity. Understanding this processcan be expected to lead to strategies designed to restore toleranceand/or arrest progression of damaging autoimmunity.
AcknowledgmentsWe thank Dr. R. Lang for providing pUC8 plasmid encoding the mouseGM-CSF gene and Dr. E. Randle-Barrett and D. Pellicci for cell sorting.
References1. Toh, B. H., I. R. van Driel, and P. A. Gleeson. 1997. Pernicious anaemia.N. Engl.
J. Med. 337:1441.2. Toh, B. H., P. A. Gleeson, S. Whittingham, and I. R. van Driel. 1998. Autoim-
mune gastritis and pernicious anemia. InThe Autoimmune Diseases, 3rd Ed. N.R.Rose and I.R. Mackay, eds. Academic, Sydney, p. 459.
3. Kojima, A., and R. T. Prehn. 1981. Genetic susceptibility to post-thymectomyautoimmune diseases in mice.Immunogenetics 14:15.
4. Gleeson, P. A., B. H. Toh, and I. R. van Driel. 1996. Organ-specific autoimmu-nity induced by lymphopenia.Immunol. Rev. 149:97.
5. Scarff, K. J., J. M. Pettitt, I. R. van Driel, P. A. Gleeson, and B. H. Toh. 1997.Immunization with gastric H1/K1-ATPase induces a reversible autoimmune gas-tritis. Immunology 92:91.
6. Alderuccio, F., and B. H. Toh. 1998. Spontaneous autoimmune gastritis inC3H/He mice: a new mouse model for gastric autoimmunity.Am. J. Pathol.153:1311.
7. Judd, L. M., P. A. Gleeson, B. H. Toh, and I. R. van Driel. 1999. Autoimmunegastritis results in disruption of gastric epithelial cell development.Am. J. Physiol. 277:G209.
8. Sakaguchi, S., K. Fukuma, K. Kuribayashi, and T. Masuda. 1985. Organ-specificautoimmune diseases induced in mice by elimination of T cell subset.J. Exp.Med. 161:72.
9. Smith, H., Y. H. Lou, P. Lacy, and K. S. K. Tung. 1992. Tolerance mechanismsin experimental ovarian and gastric autoimmune diseases.J. Immunol. 149:2212.
10. de Silva, H. D., I. R. Van Driel, N. La Gruta, B. H. Toh, and P. A. Gleeson. 1998.CD41 T cells, but not CD81 T cells, are required for the development of ex-perimental autoimmune gastritis.Immunology 93:405.
11. Martinelli, T. M., I. R. van Driel, F. Alderuccio, P. A. Gleeson, and B. H. Toh.1996. Analysis of mononuclear cell infiltrate and cytokine production in murineautoimmune gastritis.Gastroenterology 110:1791.
12. Alderuccio, F., P. A. Gleeson, S. P. Berzins, M. Martin, I. R. van Driel, andB. H. Toh. 1997. Expression of the gastric H/K ATPasea-subunit in the thymusmay explain the dominant role of theb-subunit in the pathogenesis of autoim-mune gastritis.Autoimmunity 25:167.
13. Alderuccio, F., B. H. Toh, S. S. Tan, P. A. Gleeson, and I. R. van Driel. 1993. Anautoimmune disease with multiple molecular targets abrogated by the transgenicexpression of a single autoantigen in the thymus.J. Exp. Med. 178:419.
14. de Silva, H. D., P. A. Gleeson, B. H. Toh, I. R. van Driel, and F. R. Carbone.1999. Identification of a gastritogenic epitope of the H/K ATPaseb-subunit.Immunology 96:145.
15. Suri-Payer, E., A. Z. Amar, R. Mchugh, K. Natarajan, D. H. Margulies, andE. M. Shevach. 1999. Post-thymectomy auoimmune gastritis: fine specificity andpathogenicity of anti-H/K ATPase-reactive T cells.Eur. J. Immunol. 29:669.
16. Katakai, T., Y. Agata, A. Shimizu, C. Ohshima, A. Nishio, M. Inaba,S. Kasakura, K. J. Mori, and T. Masuda. 1997. Structure of the TCR expressedon a gastritogenic T cell clone, II-6, and frequent appearance of similar clono-types in mice bearing autoimmune gastritis.Int. Immunol. 9:1849.
17. Sakaguchi, S., M. Sakaguchi, M. Asano, M. Itoh, and M. Toda. 1995. Immuno-logical self-tolerance maintained by activated T cells expressing IL-2 receptora-chains (CD25): breakdown of a single mechanism of self-tolerance causesvarious autoimmune diseases.J. Immunol. 155:1151.
18. Asano, M., M. Toda, N. Sakaguchi, and S. Sakaguchi. 1996. Autoimmune diseaseas a consequence of developmental abnormality of a T cell subpopulation.J. Exp.Med. 184:387.
19. Takahashi, T., Y. Kuniyasu, M. Toda, N. Sakaguchi, M. Itoh, M. Iwata,J. Shimizu, and S. Sakaguchi. 1998. Immunologic self-tolerance maintained byCD251CD41 naturally anergic and supressive T cells: induction of autoimmunedisease by breaking their anergic/suppressive state.Int. Immunol. 10:1969.
20. Itoh, M., T. Takahashi, N. Sakaguchi, Y. Kuniyasu, J. Shimizu, F. Otsuka, andS. Sakaguchi. 1999. Thymus and autoimmunity: production of CD251CD41
naturally anergic and suppressive T cells as a key function of the thymus inmaintaining immunological self-tolerance.J. Immunol. 162:5317.
21. Suri-Payer, E., A. Z. Amar, A. M. Thornton, and E. M. Shevach. 1998.CD41CD251 T cells inhibit both the induction and effector function of autore-active T cells and represent a unique lineage of immunoregulatory cells.J. Im-munol. 160:1212.
22. Sakaguchi, S., M. Toda, M. Asano, M. Itoh, S. S. Morse, and N. Sakaguchi. 1996.T cell-mediated maintenance of natural self-tolerance: its breakdown as a possi-ble cause of various autoimmune diseases.J. Autoimmun. 9:211.
23. Metcalf, D. 1985. The granulocyte-macrophage colony-stimulating factors.Sci-ence 229:16.
24. Weiss, W. R., K. J. Ishii, R. C. Hedstrom, M. Sedegah, M. Ichino, K. Barnhart,D. M. Klinman, and S. L. Hoffman. 1998. A plasmid encoding murine granulo-
cyte-macrophage colony-stimulating factor increases protection conferred by amalaria DNA vaccine.J. Immunol. 161:2325.
25. Dranoff, G., E. Jaffee, A. Lazenby, P. Golumbek, H. Levitsky, K. Brose,V. Jackson, H. Hamada, D. Pardoll, and R. C. Mulligan. 1993. Vaccination withirradiated tumor cells engineered to secrete murine granulocyte-macrophage col-ony-stimulating factor stimulates potent, specific, and long-lasting anti-tumor im-munity. Proc. Natl. Acad. Sci. USA 90:3539.
26. Johnson, G. R., T. J. Gonda, D. Metcalf, I. K. Hariharan, and S. Cory. 1989. Alethal myeloproliferative syndrome in mice transplanted with bone marrow cellsinfected with a retrovirus expressing granulocyte-macrophage colony stimulatingfactor.EMBO J. 8:441.
27. Lang, R. A., D. Metcalf, R. A. Cuthbertson, I. Lyons, E. Stanley, A. Kelso,G. Kannourakis, D. J. Williamson, G. K. Klintworth, T. J. Gonda, andA. R. Dunn. 1987. Transgenic mice expressing a hemopoietic growth factor gene(GM-CSF) develop accumulations of macrophages, blindness, and a fatal syn-drome of tissue damage.Cell 51:675.
28. Stampfli, M. R., R. E. Wiley, G. S. Neigh, B. U. Gajewska, X. F. Lei,D. P. Snider, Z. Xing, and M. Jordana. 1998. GM-CSF transgene expression inthe airway allows aerosolized ovalbumin to induce allergic sensitization in mice.J. Clin. Invest. 102:1704.
29. Stanley, E., G. J. Lieschke, D. Grail, D. Metcalf, G. Hodgson, J. A. Gall,D. W. Maher, J. Cebon, V. Sinickas, and A. R. Dunn. 1994. Granulocyte/mac-rophage colony-stimulating factor-deficient mice show no major perturbation ofhematopoiesis but develop a characteristic pulmonary pathology.Proc. Natl.Acad. Sci. USA 91:5592.
30. Huffman, J. A., W. M. Hull, G. Dranoff, R. C. Mulligan, and J. A. Whitsett. 1996.Pulmonary epithelial cell expression of GM-CSF corrects the alveolar proteinosisin GM-CSF-deficient mice.J. Clin. Invest.97:649.
31. Campbell, I. K., M. J. Rich, R. J. Bischof, A. R. Dunn, D. Grail, andJ. A. Hamilton. 1998. Protection from collagen-induced arthritis in granulocyte-macrophage colony-stimulating factor-deficient mice.J. Immunol. 161:3639.
32. Campbell, I. K., A. Bendele, D. A. Smith, and J. A. Hamilton. 1997. Granulocyte-macrophage colony stimulating factor exacerbates collagen induced arthritis inmice.Ann. Rheum. Dis. 56:364.
33. Fukuma, K., S. Sakaguchi, K. Kuribayashi, W.-L. Chen, R. Morishita, K. Sekita,H. Uchino, and T. Masuda. 1988. Immunologic and clinical studies in murineexperimental autoimmune gastritis induced by neonatal thymectomy.Gastroen-terology 94:274.
34. Sakaguchi, S., T. H. Ermak, M. Toda, L. J. Berg, W. Ho, B. Fazekas de St. Groth,P. A. Peterson, N. Sakaguchi, and M. M. Davis. 1994. Induction of autoimmunedisease in mice by germline alteration of the T cell receptor gene expression.J. Immunol. 152:1471.
35. Sakaguchi, N., K. Miyai, and S. Sakaguchi. 1994. Ionizing radiation and auto-immunity: induction of autoimmune disease in mice by high dose fractionatedtotal lymphoid irradiation and its prevention by inoculating normal T cells.J. Im-munol. 152:2586.
36. Morley, G. P., J. M. Callaghan, J. B. Rose, B. H. Toh, P. A. Gleeson, andI. R. van Driel. 1992. The mouse gastric H,K-ATPaseb subunit.J. Biol. Chem.267:1165.
37. Hogan, B., F. Costantini, and E. Lacy. 1986.Manipulating the mouse embryo.Cold Spring Harbor Lab Press, New York, p. 153.
38. Falk, P., K. A. Roth, and J. I. Gordon. 1994. Lectins are sensitive tools fordefining the differentiation programs of mouse gut epithelial cell lineages.Am. J. Physiol. 266:G987.
39. Callaghan, J. M., B. H. Toh, R. J. Simpson, G. S. Baldwin, and P. A. Gleeson.1992. Rapid purification of the gastric H1/K1-ATPase complex by tomato-lectinaffinity chromotography.Biochem. J. 283:63.
40. Jones, C. M., J. M. Callaghan, P. A. Gleeson, Y. Mori, T. Masuda, and B. H. Toh.1991. The parietal cell autoantibodies recognised in neonatal thymectomy-in-duced murine gastritis are thea and b subunits of the gastric proton pump.Gastroenterology 101:287.
41. Alderuccio, F., B. H. Toh, P. A. Gleeson, and I. R. van Driel. 1995. A novelmethod for isolating mononuclear cell from the stomachs of mice with experi-mental autoimmune gastritis.Autoimmunity 21:215.
42. Gasson, J. C. 1991. Molecular physiology of granulocyte-macrophage colony-stimulating factor.Blood 77:1131.
43. Tung, K. S. K., S. Smith, C. Teuscher, C. Cook, and R. E. Anderson. 1987.Murine autoimmune oophoritis, epididymoorchitis, and gastritis induced by day3 thymectomy: immunopathology.Am. J. Pathol. 126:293.
44. Thornton, A. M., and E. M. Shevach. 1998. CD41CD251 immunoregulatory Tcells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2production.J. Exp. Med. 188:287.
45. Tung, K. S. K., S. Smith, P. Matzner, K. Kasai, J. Oliver, F. Feuchter, andR. E. Anderson. 1987. Murine autoimmune oophoritis, epididymoorchitis, andgastritis induced by day 3 thymectomy: autoantibodies.Am. J. Pathol. 126:303.
46. Inaba, K., M. Inaba, N. Romani, H. Aya, M. Deguchi, S. Ikehara, S. Muramatsu,and R. M. Steinman. 1992. Generation of large numbers of dendritic cells frommouse bone marrow cultures supplemented with granulocyte/macrophage colo-ny-stimulating factor.J. Exp. Med. 176:1693.
47. Pulendran, B., J. L. Smith, G. Caspary, K. Brasel, D. Pettit, E. Maraskovsky, andC. R. Maliszewski. 1999. Distinct dendritic cell subsets differentially regulate theclass of immune response in vivo.Proc. Natl. Acad. Sci. USA 96:1036.
48. Gallucci, S., M. Lolkema, and P. Matzinger. 1999. Natural adjuvants: endoge-nous activators of dendritic cells.Nat. Med. 5:1249.
2098 AUTOIMMUNE GASTRITIS IN GM-CSF TRANSGENIC MICE
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

49. Galati, G., P. Rovere, G. Citterio, A. Bondanza, U. Scagliette, E. Bucci, S. Heltai,U. Fascio, C. Rugarli, and A. A. Manfredi. 2000. In vivo administration of GM-CSF promotes the clearance of apoptotic cells: effects on monocytes and poly-morphonuclear leukocytes.J. Leukocyte Biol. 67:174.
50. Green, E. A., E. E. Eynon, and R. A. Flavell. 1998. Local expression of TNFa inneonatal NOD mice promotes diabetes by enhancing presentation of islet anti-gens.Immunity 9:733.
51. Mahvi, D. M., M. J. Sheehy, and N. S. Yang. 1997. DNA cancer vaccines: a genegun approach.Immunol. Cell Biol. 75:456.
52. Cao, X., W. Zhang, J. Wang, M. Zhang, X. Huang, H. Hamada, and W. Chen.1999. Therapy of established tumour with a hybrid cellular vaccine generated byusing granulocyte-macrophage colony-stimulating factor genetically modifieddendritic cells.Immunology 97:616.
53. Haddad, D., J. Ramprakash, M. Sedegah, Y. Charoenvit, R. Baumgartner,S. Kumar, S. L. Hoffman, and W. R. Weiss. 2000. Plasmid vaccine expressinggranulocyte-macrophage colony-stimulating factor attracts infiltrates includingimmature dendritic cells into injected muscles.J. Immunol. 165:3772.
54. Ludewig, B., B. Odermatt, S. Landmann, H. Hengartner, and R. M. Zinkernagel.1998. Dendritic cells induce autoimmune diabetes and maintain disease via denovo formation of local lymphoid tissue.J. Exp. Med. 188:1493.
55. Heath, W. R., J. Allison, M. W. Hoffmann, G. Schonrich, G. J. Hammerling,B. Arnold, and J. F. A. P. Miller. 1992. Autoimmune diabetes as consequence oflocally produced interleukin-2.Nature 359:547.
56. Picarella, D. E., A. Kratz, C. B. Li, N. H. Ruddle, and R. A. Flavell. 1993.Transgenic TNF-a production in pancreatic islets leads to insulitis, not diabetes:distinct patterns of inflammation in TNF-a and TNF-b transgenic mice.J. Im-munol. 150:4136.
57. Sarvetnick, N., D. Liggitt, S. L. Pitts, S. E. Hansen, and T. A. Stewart. 1988.Insulin-dependent diabetes mellitus induced in transgenic mice by ectopic ex-pression of class II MHC and interferon-g. Cell 52:773.
58. Alderuccio, F., and B. Toh. 2000. Immunopathology of autoimmune gastritis:lessons from mouse models.Histol. Histopathol. 15:869.
59. Horwitz, M. S., L. M. Bradley, J. Harbertson, T. Krahl, J. Lee, and N. Sarvetnick.1998. Diabetes induced by coxsackie virus: initiation by bystander damage andnot molecular mimicry.Nat. Med. 4:781.
60. Matzinger, P. 1998. An innate sense of danger.Semin. Immunol. 10:399.61. Matzinger, P. 1994. Tolerance, danger, and the extended family.Annu. Rev.
Immunol. 12:991.62. Toh, B. H., J. W. Sentry, and F. Alderuccio. 2000. The causative H/K ATPase
antigen in the pathogenesis of autoimmune gastritis.Immunol. Today 21:348.
2099The Journal of Immunology
by guest on September 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from