lipid metabolism and dynamics during phagocytosis
TRANSCRIPT

Lipid metabolism and dynamics during phagocytosisTony Yeung1,2, Barish Ozdamar1, Paul Paroutis1 andSergio Grinstein1,2
Phagocytosis, the engulfment of particles, mediates the
elimination of invading pathogens as well as the clearance of
apoptotic cells. Ingested particles reside within a vacuole or
phagosome, where they are eventually destroyed and
digested. The phagosomal lumen acquires microbicidal and
digestive properties through interaction with various
components of the endocytic pathway, a process known as
maturation. Lipids are known to have numerous roles in
phagosome formation and maturation; recent developments in
the design of lipid-specific probes and in high-resolution
imaging have revealed that lipids, notably phosphoinositides,
are involved in signaling, actin assembly and the recruitment of
molecular motors to sites of ingestion. In addition,
phosphoinositides and other lipids also regulate multiple
membrane budding, fission and fusion events required for
maturation.
Addresses1 Division of Cell Biology, The Hospital for Sick Children,
555 University Avenue, Toronto, M5G 1X8, Canada2 Institute of Medical Sciences, University of Toronto, Toronto,
M5S 1A8, Canada
T Yeung and B Ozdamar contributed equally to this work.
Corresponding author: Grinstein, Sergio ([email protected])
Current Opinion in Cell Biology 2006, 18:429–437
This review comes from a themed issue on
Membranes and organelles
Edited by Pietro de Camilli and Antonella de Matteis
Available online 14th June 2006
0955-0674/$ – see front matter
# 2006 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.ceb.2006.06.006
IntroductionPhagocytosis, the uptake of particles �0.5 mm in dia-
meter, is a central event in innate immunity and in tissue
remodeling. Professional phagocytes, mainly neutrophils
and macrophages, internalize invading microorganisms to
resolve infection and can also engulf apoptotic bodies,
contributing to the clearance of senescent or damaged
cells. Phagocytosis is triggered by surface receptors that
become cross-linked upon binding to cognate molecules
on the surface of the particles, which act as a large
multivalent ligand. Engagement of such receptors, typi-
fied by Fc and complement receptors, initiates a complex
signaling cascade that induces extensive actin and mem-
brane remodeling, driving the extension of pseudopods
and culminating with particle engulfment. Immediately
www.sciencedirect.com
after sealing, the phagosome undergoes multiple rounds
of fission and fusion with other organelles, a carefully
choreographed sequence of events collectively known as
‘maturation’ [1,2]. It is becoming increasingly apparent
that lipid-mediated signaling is intimately involved in
both phagosome formation and maturation. This review
will highlight recent advances in our understanding of
lipid metabolism and redistribution during phagocytosis.
Phagosome formationAs is the case for most mammalian cells, the plasma
membrane of phagocytes is rich in cholesterol and sphin-
golipids. Microdomains enriched in these lipids, often
called rafts, have been proposed to play a crucial role in
phagocytosis of IgG-coated particles, which is mediated
by Fcg receptors [3,4]. This class of phagocytic receptors
has been studied in most detail and will be used as a
paradigm hereafter. Fcg-receptors become activated
upon tyrosine phosphorylation of their immunoreceptor
tyrosine-based activation motif (ITAM), a process cata-
lyzed by Src-family kinases. Src-family kinases such as
Lyn, which are acylated by multiple saturated chains,
partition preferentially to cholesterol-rich microdomains
and this association is thought to be required for their
clustering and activation in the vicinity of cross-linked
receptors [5,6]. This notion is based on experiments
where depletion of cellular cholesterol using statins or
b-methyl-cyclodextrin impaired Fc receptor-mediated
signaling and phagocytosis [3,7,8]. However, statins also
affect the prenylation of Rho-family GTPases that are
essential for pseudopod extension, and depletion of cho-
lesterol interferes with processes that are not mediated by
rafts [9]. Moreover, other investigators failed to detect
inhibition of phagocytosis following cholesterol depletion
[10]. It is therefore premature to conclude that cholesterol
and sphingolipid-rich microdomains are involved in pha-
gosome formation.
The role of other lipids in phagosome formation is better
defined. Phosphoinositides have been studied in most
detail by biochemical methods and, more recently, by
fluorescence imaging using genetically encoded biosen-
sors. Chimeric constructs consisting of fluorescent pro-
teins fused to pleckstrin homology (PH) domains that
recognize the headgroups of phosphatidylinositol-4,5-
bisphosphate (PI(4,5)P2) or phosphatidylinositol-3,4,5-tri-sphosphate (PI(3,4,5)P3) have been used to monitor the
dynamics of these lipids in intact cells during particle
ingestion [11,12]. Both of these lipids undergo acute,
exquisitely localized changes during phagosome forma-
tion. PI(4,5)P2,which is constitutively present in the
Current Opinion in Cell Biology 2006, 18:429–437

430 Membranes and organelles
plasma membrane, becomes transiently and modestly
accumulated in pseudopods of forming phagosomes,
but disappears rapidly thereafter, being undetectable in
the membrane of sealed phagosomes (Figure 1a–c). Sti-
mulation of type I phosphatidylinositol-phosphate-5-
kinase (PIP5KI) is likely to be responsible for the tran-
sient accumulation of PI(4,5)P2. PIP5KI is present at the
phagosomal cup [11,13], where it may be activated by
Arf6 or Rho-family GTPases and/or by lipid mediators
like phosphatidic acid that are generated during particle
ingestion [14,15]. The causes of the subsequent disap-
pearance of PI(4,5)P2 are likely to be more complex. It is
clear that a fraction of the inositide is hydrolyzed by a
phospholipase C, most likely PLCg in Fcg-mediated
phagocytosis [11]. This is suggested by the accompanying
increase in cytosolic [Ca2+], attributed to inositol 3,4,5-
trisphosphate, and by the generation of diacylglycerol at
the cup, which coincides spatially and temporally with the
disappearance of the phosphoinositide (Figure 1d). In
addition, it is conceivable, though not yet demonstrated,
that PI(4,5)P2 may be degraded at sites of phagocytosis by
phosphoinositide 4- and/or 5-phosphatases like those
recently shown to be active during endocytosis [16].
Disappearance of PI(4,5)P2 from nascent phagosomes is
not, however, due solely to its hydrolysis. A fraction of the
phosphoinositide is converted to PI(3,4,5)P3 by class I
phosphatidylinositol 3-kinase (PI3K), which is recruited
and activated by Syk or by adaptor proteins like Gab2
[12,17–19]. Chimeras containing the PH domain of Akt
that preferentially bind 30-polyphosphoinositides
undergo a remarkable accumulation at sites of phagosome
formation and remain briefly associated with nascent
phagosomes (Figure 1f–g) [12,20]. As in the case of
diacylglycerol, the distribution of PI(3,4,5)P3 is confined
to the forming phagosome. It is remarkable how the lipid
species generated de novo during phagocytosis, a rela-
tively slow process lasting minutes, fail to diffuse past the
boundary of the cup. It is tempting to speculate that a
diffusional barrier is formed at the edge of advancing
pseudopods, but other mechanisms should not be
excluded. Localized generation at the base of the cup
and a combination of diffusion and hydrolysis at the edge
of the forming phagosome could equally explain the
observations. Active endocytosis near the tip of the pseu-
dopods could similarly limit the redistribution of the
lipids to the unengaged plasma membrane. Lastly, bind-
ing of lipids to immobile complexes within the cup may
restrict their lateral mobility, a notion indirectly sup-
ported by evidence obtained by measuring the behavior
of lipid-anchored fluorescent proteins after photobleach-
ing (Yeung et al., unpublished).
The accumulation of PI(3,4,5)P3 is not only highly loca-
lized, but is also very brief and hence difficult to capture
by means other than imaging. There is good evidence that
the 5-phosphatase SHIP contributes to PI(3,4,5)P3
Current Opinion in Cell Biology 2006, 18:429–437
hydrolysis: SHIP isoforms are recruited to sites of pha-
gocytosis, ostensibly by association of their SH2 domain
with phosphotyrosines [21,22], and their overexpression
curtails PI(3,4,5)P3 accumulation and phagocytic effi-
ciency [22,23]. Overexpression of the 30-phosphatase
PTEN can similarly depress phagocytosis, but whether
this enzyme is normally present and active at sites of
ingestion is less clear.
The fate of other phospholipids during phagocytosis
remains largely unexplored. Phosphatidylinositol 4-phos-
phate (PI(4)P) not only serves as a substrate for the
synthesis of PI(4,5)P2 by PIP5KI [13], but may also recruit
this kinase to the plasma membrane via its inositide-
specific activation loop [24]. Recently, the PH domain of
Osh2p (the yeast homologue of the oxysterol-binding
protein) has been developed as a probe for PI(4)P [25].
Preliminary experiments using PH-Osh2p chimeras sug-
gest that PI(4)P is metabolized during phagocytosis
(Yeung et al., unpublished). This result is consistent with
its role as a substrate for the stimulated biosynthesis of
PI(4,5)P2 and may also help to explain the dissociation of
PIP5KI from the phagosome following its internalization
[11,13].
What are the functional consequences of localized phos-
phoinositide metabolism during phagocytosis? That ino-
sitides are crucial for successful phagocytosis is best
illustrated by the effects of PI3K inhibitors like wort-
mannin, which can abrogate particle internalization
[26,27]. The picture, however, is complicated by the
size-dependence of the inhibitory effect: wortmannin
obliterates phagocytosis of large particles (e.g. >5 mm),
while particles the size of most bacteria (�1 mm) are
barely affected [26]. This suggests that not all phagocy-
tosis is the same and that generalizations should be
avoided. Nevertheless, it is clear that plasmalemmal
polyphosphoinositides play multiple, complementary
roles in orchestrating phagocytosis, signaling its initiation
and contributing to the cytoskeletal and membrane remo-
deling (Figure 2). The formation of PI(3,4,5)P3 stimulates
the hydrolysis of PI(4,5)P2 by promoting the recruitment,
phosphorylation and activation of PLCg [28]. In addition,
the headgroup of PI(3,4,5)P3 is likely to contribute to the
association of exchange factors that activate Rho-family
GTPases within the phagocytic cup, though this event is
not an absolute requirement to trigger actin polymeriza-
tion, which persists when PI3K is inhibited [1,26]. Dyna-
min II is also recruited to the phagocytic cup along with
amphiphysin II in a PI3K-dependent manner. Their
precise mode of action is unclear, but interference with
either dynamin II or amphiphysin II function was shown
to inhibit phagocytosis [29,30].
In addition to their role in signaling, phosphoinositides are
central to the cytoskeletal rearrangements that underlie
engulfment. PI(4,5)P2 can promote actin polymerization
www.sciencedirect.com
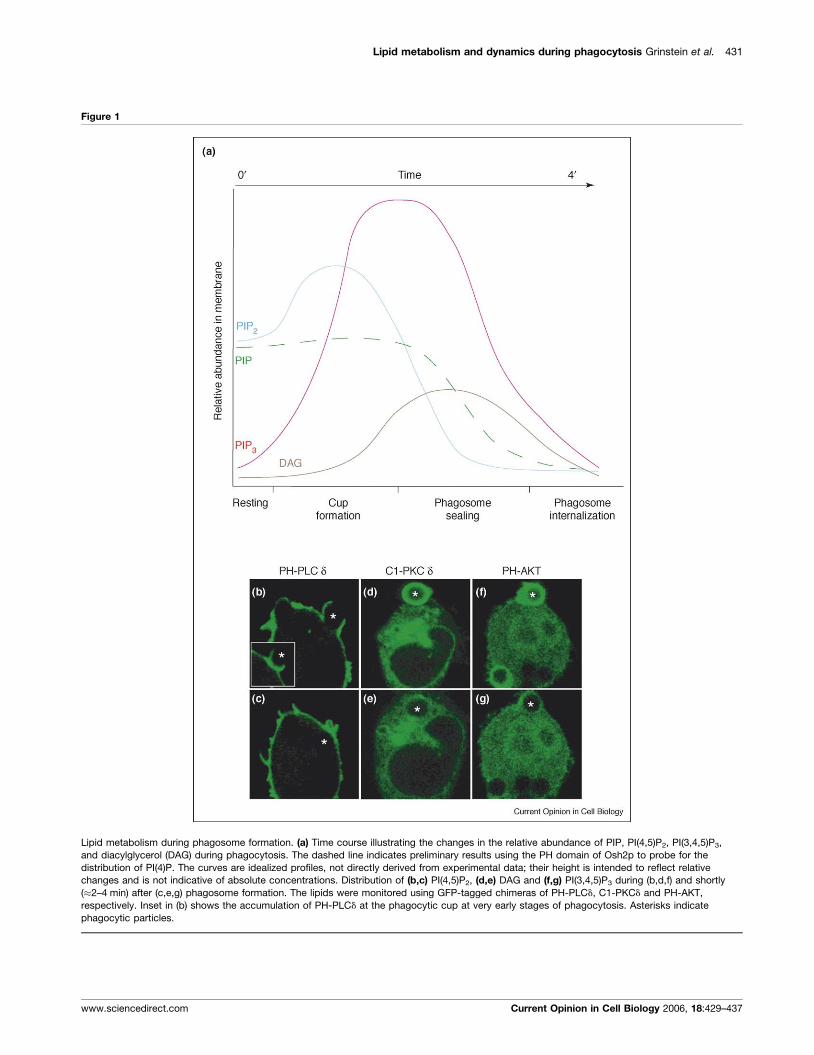
Lipid metabolism and dynamics during phagocytosis Grinstein et al. 431
Figure 1
Lipid metabolism during phagosome formation. (a) Time course illustrating the changes in the relative abundance of PIP, PI(4,5)P2, PI(3,4,5)P3,
and diacylglycerol (DAG) during phagocytosis. The dashed line indicates preliminary results using the PH domain of Osh2p to probe for the
distribution of PI(4)P. The curves are idealized profiles, not directly derived from experimental data; their height is intended to reflect relative
changes and is not indicative of absolute concentrations. Distribution of (b,c) PI(4,5)P2, (d,e) DAG and (f,g) PI(3,4,5)P3 during (b,d,f) and shortly
(�2–4 min) after (c,e,g) phagosome formation. The lipids were monitored using GFP-tagged chimeras of PH-PLCd, C1-PKCd and PH-AKT,
respectively. Inset in (b) shows the accumulation of PH-PLCd at the phagocytic cup at very early stages of phagocytosis. Asterisks indicate
phagocytic particles.
www.sciencedirect.com Current Opinion in Cell Biology 2006, 18:429–437

432 Membranes and organelles
Figure 2
Functional consequences of lipid metabolism during phagosome formation. Schematic representation of a forming phagosome induced by
activation of Fcg receptors. PI(4,5)P2 (shown in blue) plays an important role in phagocytosis by mediating actin remodeling through its interaction
with cytoskeletal proteins including the capping protein profilin, WASP family proteins and actin-severing proteins like gelsolin (not shown).
PI(3,4,5)P3 (shown in pink) promotes localized membrane remodeling by recruiting amphiphysin, dynamin and myosin X to the phagocytic cup.
It may also contribute to the activation of Rac1 by recruiting the guanine nucleotide exchange factor Vav, although PI3K-independent activation
of Rac1 has been reported.
in at least three ways: first, by inducing Arp2/3 activation by
WASP; second, by removing capping proteins like profilin
from the ends of actin filaments; and third, by stimulating
gelsolin to sever filaments, thereby generating fast-grow-
ing barbed ends. PI(3,4,5)P3 also contributes directly to the
mechanical events leading to particle intake by recruiting
WAVE [31] and myosin X to forming phagosomes [32]. Of
note, the dissociation of the cytoskeleton from the sealed
phagosome, which is believed to be required for matura-
tion to proceed, coincides with the disappearance of
PI(4,5)P2 from the phagosomal membrane, suggesting that
the inositide plays a role in the termination of actin
assembly. In support of this concept, Scott et al. found
Current Opinion in Cell Biology 2006, 18:429–437
that preventing the loss of PI(4,5)P2 by impairing PLC
caused retention of phagosomal actin [33�]. Conversely,
these authors also found that when inositide hydrolysis
proceeded normally, actin dissociated from the phago-
somes even when constitutively active Rac1 remained
associated with their membrane.
Phosphoinositides, particularly PI(3,4,5)P3, are seemingly
also involved in the membrane remodeling that enables
phagocytic cells to retain near-normal surface area while
engulfing large and/or multiple particles. Fusion of endo-
membranes at sites of phagocytosis is now believed to
mediate the conservation of surface area and to
www.sciencedirect.com
Lipid metabolism and dynamics during phagocytosis Grinstein et al. 433
simultaneously initiate the maturation sequence. Such
fusion seemingly involves products of PI3K, as it is
inhibited by wortmannin [26]. The specific lipids
involved and their precise mode of action remain to be
defined.
Phagosome maturationThe membrane of the phagosome is originally derived
from the plasmalemma and its soluble contents are a
sampling of the extracellular space. The lumen of the
nascent phagosome is therefore a rather benign environ-
ment, incapable of eliminating invading microorganisms
or of disposing apoptotic bodies. These capabilities are
acquired subsequently, as a result of maturation. In many
respects, phagosomal maturation recapitulates progres-
sion from endocytic vesicles to lysosomes. Early phago-
somes acquire Rab5 and EEA1, quintessential markers of
early endosomes. Phagosomes next adopt late endosomal
features. This transition is accompanied by divestment of
Rab5 and acquisition of Rab7 and by the appearance of
specific markers such as LAMP1 and the mannose-6-
phosphate receptor [34]. Ultimately, phagosomes merge
with lysosomes, acquiring cathepsins and other hydro-
lases. Throughout this sequence, the phagosomal lumen
becomes progressively more acidic.
Aside from a general assessment of the total lipid com-
position at specific stages in the maturation process,
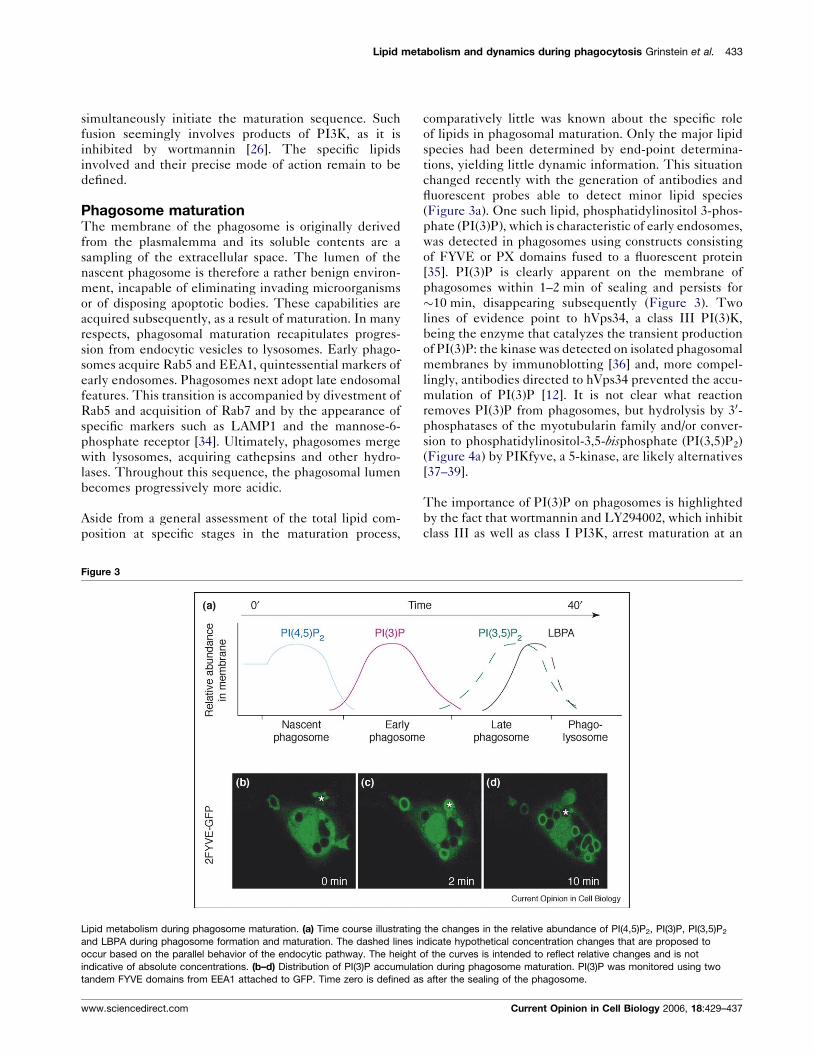
Figure 3
Lipid metabolism during phagosome maturation. (a) Time course illustrating
and LBPA during phagosome formation and maturation. The dashed lines in
occur based on the parallel behavior of the endocytic pathway. The height
indicative of absolute concentrations. (b–d) Distribution of PI(3)P accumulat
tandem FYVE domains from EEA1 attached to GFP. Time zero is defined as
www.sciencedirect.com
comparatively little was known about the specific role
of lipids in phagosomal maturation. Only the major lipid
species had been determined by end-point determina-
tions, yielding little dynamic information. This situation
changed recently with the generation of antibodies and
fluorescent probes able to detect minor lipid species
(Figure 3a). One such lipid, phosphatidylinositol 3-phos-
phate (PI(3)P), which is characteristic of early endosomes,
was detected in phagosomes using constructs consisting
of FYVE or PX domains fused to a fluorescent protein
[35]. PI(3)P is clearly apparent on the membrane of
phagosomes within 1–2 min of sealing and persists for
�10 min, disappearing subsequently (Figure 3). Two
lines of evidence point to hVps34, a class III PI(3)K,
being the enzyme that catalyzes the transient production
of PI(3)P: the kinase was detected on isolated phagosomal
membranes by immunoblotting [36] and, more compel-
lingly, antibodies directed to hVps34 prevented the accu-
mulation of PI(3)P [12]. It is not clear what reaction
removes PI(3)P from phagosomes, but hydrolysis by 30-phosphatases of the myotubularin family and/or conver-
sion to phosphatidylinositol-3,5-bisphosphate (PI(3,5)P2)
(Figure 4a) by PIKfyve, a 5-kinase, are likely alternatives
[37–39].
The importance of PI(3)P on phagosomes is highlighted
by the fact that wortmannin and LY294002, which inhibit
class III as well as class I PI3K, arrest maturation at an
the changes in the relative abundance of PI(4,5)P2, PI(3)P, PI(3,5)P2
dicate hypothetical concentration changes that are proposed to
of the curves is intended to reflect relative changes and is not
ion during phagosome maturation. PI(3)P was monitored using two
after the sealing of the phagosome.
Current Opinion in Cell Biology 2006, 18:429–437
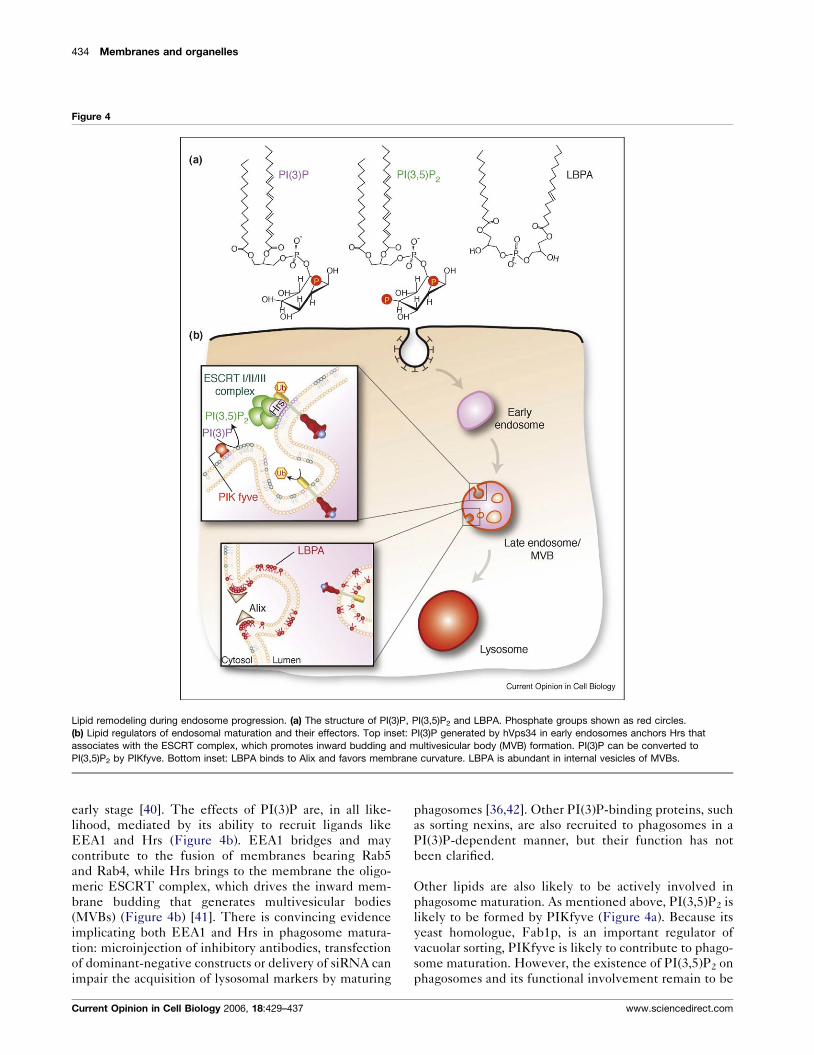
434 Membranes and organelles
Figure 4
Lipid remodeling during endosome progression. (a) The structure of PI(3)P, PI(3,5)P2 and LBPA. Phosphate groups shown as red circles.
(b) Lipid regulators of endosomal maturation and their effectors. Top inset: PI(3)P generated by hVps34 in early endosomes anchors Hrs that
associates with the ESCRT complex, which promotes inward budding and multivesicular body (MVB) formation. PI(3)P can be converted to
PI(3,5)P2 by PIKfyve. Bottom inset: LBPA binds to Alix and favors membrane curvature. LBPA is abundant in internal vesicles of MVBs.
early stage [40]. The effects of PI(3)P are, in all like-
lihood, mediated by its ability to recruit ligands like
EEA1 and Hrs (Figure 4b). EEA1 bridges and may
contribute to the fusion of membranes bearing Rab5
and Rab4, while Hrs brings to the membrane the oligo-
meric ESCRT complex, which drives the inward mem-
brane budding that generates multivesicular bodies
(MVBs) (Figure 4b) [41]. There is convincing evidence
implicating both EEA1 and Hrs in phagosome matura-
tion: microinjection of inhibitory antibodies, transfection
of dominant-negative constructs or delivery of siRNA can
impair the acquisition of lysosomal markers by maturing
Current Opinion in Cell Biology 2006, 18:429–437
phagosomes [36,42]. Other PI(3)P-binding proteins, such
as sorting nexins, are also recruited to phagosomes in a
PI(3)P-dependent manner, but their function has not
been clarified.
Other lipids are also likely to be actively involved in
phagosome maturation. As mentioned above, PI(3,5)P2 is
likely to be formed by PIKfyve (Figure 4a). Because its
yeast homologue, Fab1p, is an important regulator of
vacuolar sorting, PIKfyve is likely to contribute to phago-
some maturation. However, the existence of PI(3,5)P2 on
phagosomes and its functional involvement remain to be
www.sciencedirect.com

Lipid metabolism and dynamics during phagocytosis Grinstein et al. 435
established. By contrast, there is convincing evidence that
another important lipid mediator accumulates in phago-
somes. Lysobisphosphatidic acid (LBPA) (also known as
bismonoacylphosphatidic acid), which is generated by
degradation of phosphatidylglycerol and cardiolipin
[43], is normally abundant in the internal membranes
of MVBs and late endosomes [44] (Figure 4). It has also
been identified in maturing endosomes, though it is
unclear whether it is formed in situ or is imported by
fusion with late endosomes. LBPA has a unique cone-
shaped structure that may induce membrane curvature,
facilitating the budding that generates MVBs [45] (Figure
4b). Recently, Gruenberg and colleagues demonstrated
that the addition of LBPA to pure lipid liposomes causes
them to adopt an MVB-like multivesicular morphology,
but only if the pH is acidic, as is the case in the phago-
somal lumen [46�]. However, LBPA is unlikely to be
solely responsible for budding. Indeed, Gruenberg’s team
identified ALIX, an E class Vps protein, as a regulator of
MVB formation. ALIX is an LBPA-binding protein and
its depletion by means of siRNA reduces the number of
acidic compartments in the cells [46�]. The presence of
ALIX in phagosomes has not been reported and much
remains to be learned about its possible role and that of
LBPA in maturation.
The internal vesicles of MVBs also contain significant
amounts of cholesterol. Typically implicated in the stabi-
lization of microdomains at the plasma membrane, cho-
lesterol is also found in the endocytic pathway. Nearly 80%
of the cholesterol content in the endocytic pathway is
confined to the recycling compartments of the early and
late endosomes [47]. Intriguingly, cholesterol is nearly
absent from lysosomal membranes. Insights into to the
role of cholesterol in endomembrane traffic have been
gleaned from studies in cells derived from Niemann-Pick
type C (NPC) patients. NPC is an inherited cholesterol-
storage disorder that typically results in a marked accu-
mulation of cholesterol in late endosomal structures [48].
Defects in the cholesterol-binding proteins NPC1 and
NPC2 account for nearly all cases of NPC. Most interest-
ingly, the NPC phenotype is also associated with defects in
traffic between late endosomes and lysosomes [49]. Under
normal conditions, the progressive decrease in cholesterol
levels along the endocytic pathway is accompanied by an
increase in the levels of ceramide [50]. Ceramide, pro-
duced from the catabolism of sphingomyelin [51], is also
capable of stabilizing lipid microdomains [52] and thus
may function in an analogous manner to cholesterol.
Though the precise contribution of the cholesterol-to-
ceramide switch during endocytic progression has not
yet been adequately explored, it is tempting to speculate
that the ratio of these two lipids may function as a vital cue
in the late-endosome-to-lysosome transition.
Despite its importance in endocytic traffic, remarkably
little is known about cholesterol in phagosomes.
www.sciencedirect.com
Evidence of its presence is sketchy and precise quantita-
tion of cholesterol concentration at the individual stages
of the maturation sequence is lacking. Cholesterol has
been invoked in regulating the fate of PI(3)P in phago-
somes, their ability to bind flotillin and even in their
microbicidal competence. However, most of this informa-
tion is preliminary and much remains to be done.
ConclusionsWhile formerly regarded as a solvent matrix for membrane
proteins or as an insulator separating cellular compart-
ments, it is now strikingly clear that lipids play an active
and very dynamic role in virtually all aspects of cell func-
tion, with phagocytosis being no exception. Lipids are
critical in transducing phagocytic signals, in remodeling
the actin cytoskeleton and in attracting molecular motors
during phagosome formation. In phagosome maturation,
lipids dictate membrane fusion and fission. Importantly,
lipid metabolism is also a critical event in host–pathogen
interactions. This is best illustrated by Salmonella, which
co-opts the host cell phosphoinositide metabolism during
invasion. The bacterium injects effectors that induce loca-
lized hydrolysis of PI(4,5)P2 to gain entrance into the cell
[53], and promote the formation of PI(3)P to establish a
favorable replicative niche [54�]. Conversely, Mycobacter-ium promotes the exclusion of PI(3)P from the phagosomal
membrane in order to arrest phagosomal maturation, pre-
venting fusion with potentially microbicidal lysosomes
[55�]. We predict that lipids will be similarly found to
control other aspects of the microbicidal response, such as
the generation of reactive oxygen intermediates. More
sophisticated lipid detection methods and the develop-
ment of additional biosensors for in situ determination will
be needed to propel this field forward.
AcknowledgmentsWork in the authors’ laboratory is supported by the Heart and StrokeFoundation of Canada and the Canadian Institutes for Health Research.T. Y is supported by a CIHR MD/PhD studentship. B. O. is supported bya ResTraComp fellowship from the Hospital for Sick Children. S.G. is thecurrent holder of the Pitblado Chair in Cell Biology.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest
�� of outstanding interest
1. Niedergang F, Chavrier P: Signaling and membrane dynamicsduring phagocytosis: many roads lead to the phagos(R)ome.Curr Opin Cell Biol 2004, 16:422-428.
2. Greenberg S, Grinstein S: Phagocytosis and innate immunity.Curr Opin Immunol 2002, 14:136-145.
3. Kwiatkowska K, Frey J, Sobota A: Phosphorylation of FcgRIIAis required for the receptor-induced actin rearrangementand capping: the role of membrane rafts. J Cell Sci 2003,116:537-550.
4. Kono H, Suzuki T, Yamamoto K, Okada M, Yamamoto T, Honda Z:Spatial raft coalescence represents an initial step in FcgRsignaling. J Immunol 2002, 169:193-203.
Current Opinion in Cell Biology 2006, 18:429–437

436 Membranes and organelles
5. Sheets ED, Holowka D, Baird B: Critical role for cholesterol inLyn-mediated tyrosine phosphorylation of FcepsilonRI andtheir association with detergent-resistant membranes.J Cell Biol 1999, 145:877-887.
6. Holowka D, Gosse JA, Hammond AT, Han X, Sengupta P,Smith NL, Wagenknecht-Wiesner A, Wu M, Young RM, Baird B:Lipid segregation and IgE receptor signaling: a decade ofprogress. Biochim Biophys Acta 2005, 1746:252-259.
7. Loike JD, Shabtai DY, Neuhut R, Malitzky S, Lu E, Husemann J,Goldberg IJ, Silverstein SC: Statin inhibition of Fc receptor-mediated phagocytosis by macrophages is modulated by cellactivation and cholesterol. Arterioscler Thromb Vasc Biol 2004,24:2051-2056.
8. Hillyard DZ, Jardine AG, McDonald KJ, Cameron AJ: Fluvastatininhibits raft dependent Fcgamma receptor signalling in humanmonocytes. Atherosclerosis 2004, 172:219-228.
9. Kwik J, Boyle S, Fooksman D, Margolis L, Sheetz MP, Edidin M:Membrane cholesterol, lateral mobility, and thephosphatidylinositol 4,5-bisphosphate-dependentorganization of cell actin. Proc Natl Acad Sci USA 2003,100:13964-13969.
10. Gatfield J, Pieters J: Essential role for cholesterol in entryof mycobacteria into macrophages. Science 2000,288:1647-1650.
11. Botelho RJ, Teruel M, Dierckman R, Anderson R, Wells A,York JD, Meyer T, Grinstein S: Localized biphasic changesin phosphatidylinositol-4,5-bisphosphate at sites ofphagocytosis. J Cell Biol 2000, 151:1353-1368.
12. Vieira OV, Botelho RJ, Rameh L, Brachmann SM, Matsuo T,Davidson HW, Schreiber A, Backer JM, Cantley LC, Grinstein S:Distinct roles of class I and class III phosphatidylinositol3-kinases in phagosome formation and maturation. J Cell Biol2001, 155:19-25.
13. Coppolino MG, Dierckman R, Loijens J, Collins RF, Pouladi M,Jongstra-Bilen J, Schreiber AD, Trimble WS, Anderson R,Grinstein S: Inhibition of phosphatidylinositol-4-phosphate5-kinase Ia impairs localized actin remodeling andsuppresses phagocytosis. J Biol Chem 2002,277:43849-43857.
14. Tolias KF, Hartwig JH, Ishihara H, Shibasaki Y, Cantley LC,Carpenter CL: Type Ia phosphatidylinositol-4-phosphate5-kinase mediates Rac-dependent actin assembly.Curr Biol 2000, 10:153-156.
15. Wong KW, Isberg RR: Arf6 and phosphoinositol-4-phosphate-5-kinase activities permit bypass of the Rac1 requirement forb1 integrin-mediated bacterial uptake. J Exp Med 2003,198:603-614.
16. Shin HW, Hayashi M, Christoforidis S, Lacas-Gervais S,Hoepfner S, Wenk MR, Modregger J, Uttenweiler-Joseph S,Wilm M, Nystuen A et al.: An enzymatic cascade of Rab5effectors regulates phosphoinositide turnover in theendocytic pathway. J Cell Biol 2005, 170:607-618.
17. Gu H, Botelho RJ, Yu M, Grinstein S, Neel BG: Critical rolefor scaffolding adapter Gab2 in Fc gamma R-mediatedphagocytosis. J Cell Biol 2003, 161:1151-1161.
18. Moon KD, Post CB, Durden DL, Zhou Q, De P, Harrison ML,Geahlen RL: Molecular basis for a direct interaction betweenthe Syk protein-tyrosine kinase and phosphoinositide3-kinase. J Biol Chem 2005, 280:1543-1551.
19. Tridandapani S, Lyden TW, Smith JL, Carter JE, Coggeshall KM,Anderson CL: The adapter protein LAT enhances fcg receptor-mediated signal transduction in myeloid cells. J Biol Chem2000, 275:20480-20487.
20. Marshall JG, Booth JW, Stambolic V, Mak T, Balla T, Schreiber AD,Meyer T, Grinstein S: Restricted accumulation ofphosphatidylinositol 3-kinase products in a plasmalemmalsubdomain during Fcg receptor-mediated phagocytosis.J Cell Biol 2001, 153:1369-1380.
21. Maresco DL, Osborne JM, Cooney D, Coggeshall KM,Anderson CL: The SH2-containing 50-inositol phosphatase
Current Opinion in Cell Biology 2006, 18:429–437
(SHIP) is tyrosine phosphorylated after Fc gamma receptorclustering in monocytes. J Immunol 1999, 162:6458-6465.
22. Nakamura K, Malykhin A, Coggeshall KM: The Src homology 2domain-containing inositol 5-phosphatase negativelyregulates Fcgamma receptor-mediated phagocytosis throughimmunoreceptor tyrosine-based activation motif-bearingphagocytic receptors. Blood 2002, 100:3374-3382.
23. Cox D, Dale BM, Kashiwada M, Helgason CD, Greenberg S:A regulatory role for Src homology 2 domain-containinginositol 50-phosphatase (SHIP) in phagocytosis mediated byFcg receptors and complement receptor 3aMb2; CD11b/CD18). J Exp Med 2001, 193:61-71.
24. Kunz J, Wilson MP, Kisseleva M, Hurley JH, Majerus PW,Anderson RA: The activation loop of phosphatidylinositolphosphate kinases determines signaling specificity.Mol Cell 2000, 5:1-11.
25. Roy A, Levine TP: Multiple pools of phosphatidylinositol4-phosphate detected using the pleckstrin homology domainof Osh2p. J Biol Chem 2004, 279:44683-44689.
26. Cox D, Tseng CC, Bjekic G, Greenberg S: A requirement forphosphatidylinositol 3-kinase in pseudopod extension.J Biol Chem 1999, 274:1240-1247.
27. Araki N, Hatae T, Furukawa A, Swanson JA: Phosphoinositide-3-kinase-independent contractile activities associated withFcg-receptor-mediated phagocytosis and macropinocytosisin macrophages. J Cell Sci 2003, 116:247-257.
28. Falasca M, Logan SK, Lehto VP, Baccante G, Lemmon MA,Schlessinger J: Activation of phospholipase C gamma byPI 3-kinase-induced PH domain-mediated membranetargeting. Embo J 1998, 17:414-422.
29. Gold ES, Morrissette NS, Underhill DM, Guo J, Bassetti M,Aderem A: Amphiphysin IIm, a novel amphiphysin II isoform,is required for macrophage phagocytosis. Immunity 2000,12:285-292.
30. Gold ES, Underhill DM, Morrissette NS, Guo J, McNiven MA,Aderem A: Dynamin 2 is required for phagocytosis inmacrophages. J Exp Med 1999, 190:1849-1856.
31. Oikawa T, Yamaguchi H, Itoh T, Kato M, Ijuin T, Yamazaki D,Suetsugu S, Takenawa T: PtdIns(3,4,5)P3 binding is necessaryfor WAVE2-induced formation of lamellipodia. Nat Cell Biol2004, 6:420-426.
32. Cox D, Berg JS, Cammer M, Chinegwundoh JO, Dale BM,Cheney RE, Greenberg S: Myosin X is a downstreameffector of PI(3)K during phagocytosis. Nat Cell Biol 2002,4:469-477.
33.�
Scott CC, Dobson W, Botelho RJ, Coady-Osberg N,Chavrier P, Knecht DA, Heath C, Stahl P, Grinstein S:Phosphatidylinositol-4,5-bisphosphate hydrolysis directsactin remodeling during phagocytosis. J Cell Biol 2005,169:139-149.
This study explores the mechanisms leading to actin disassembly duringphagocytosis and suggests that PI(4,5)P2 hydrolysis rather than Rac1inactivation is mainly responsible for actin depolymerization during com-pletion of phagocytosis.
34. Pitt A, Mayorga LS, Stahl PD, Schwartz AL: Alterations in theprotein composition of maturing phagosomes. J Clin Invest1992, 90:1978-1983.
35. Ellson CD, Anderson KE, Morgan G, Chilvers ER, Lipp P,Stephens LR, Hawkins PT: Phosphatidylinositol 3-phosphateis generated in phagosomal membranes. Curr Biol 2001,11:1631-1635.
36. Fratti RA, Backer JM, Gruenberg J, Corvera S, Deretic V: Role ofphosphatidylinositol 3-kinase and Rab5 effectors inphagosomal biogenesis and mycobacterial phagosomematuration arrest. J Cell Biol 2001, 154:631-644.
37. Kelley VA, Schorey JS: Modulation of cellularphosphatidylinositol 3-phosphate levels in primarymacrophages affects heat-killed but not viableMycobacterium avium’s transport through the phagosomematuration process. Cell Microbiol 2004, 6:973-985.
www.sciencedirect.com

Lipid metabolism and dynamics during phagocytosis Grinstein et al. 437
38. Sbrissa D, Ikonomov OC, Shisheva A: Phosphatidylinositol3-phosphate-interacting domains in PIKfyve. Bindingspecificity and role in PIKfyve. Endomenbrane localization.J Biol Chem 2002, 277:6073-6079.
39. Ikonomov OC, Sbrissa D, Mlak K, Kanzaki M, Pessin J, Shisheva A:Functional dissection of lipid and protein kinase signals ofPIKfyve reveals the role of PtdIns 3,5-P2 production forendomembrane integrity. J Biol Chem 2002, 277:9206-9211.
40. Araki N, Johnson MT, Swanson JA: A role for phosphoinositide3-kinase in the completion of macropinocytosis andphagocytosis by macrophages. J Cell Biol 1996, 135:1249-1260.
41. Raiborg C, Bremnes B, Mehlum A, Gillooly DJ, D’Arrigo A, Stang E,Stenmark H: FYVE and coiled-coil domains determine thespecific localisation of Hrs to early endosomes. J Cell Sci 2001,114:2255-2263.
42. Vieira OV, Harrison RE, Scott CC, Stenmark H, Alexander D, Liu J,Gruenberg J, Schreiber AD, Grinstein S: Acquisition of Hrs, anessential component of phagosomal maturation, is impairedby mycobacteria. Mol Cell Biol 2004, 24:4593-4604.
43. Amidon B, Brown A, Waite M: Transacylase and phospholipasesin the synthesis of bis(monoacylglycero)phosphate.Biochemistry 1996, 35:13995-14002.
44. Kobayashi T, Startchev K, Whitney AJ, Gruenberg J: Localizationof lysobisphosphatidic acid-rich membrane domains in lateendosomes. Biol Chem 2001, 382:483-485.
45. Kobayashi T, Hirabayashi Y: Lipid membrane domains in cellsurface and vacuolar systems. Glycoconj J 2000, 17:163-171.
46.�
Matsuo H, Chevallier J, Mayran N, Le Blanc I, Ferguson C, Faure J,Blanc NS, Matile S, Dubochet J, Sadoul R et al.: Role of LBPA andAlix in multivesicular liposome formation and endosomeorganization. Science 2004, 303:531-534.
This study reports the ability of the unique lipid LBPA to induce theformation of multivesicular structures in a pH-dependent manner.
47. Mobius W, van Donselaar E, Ohno-Iwashita Y, Shimada Y,Heijnen HF, Slot JW, Geuze HJ: Recycling compartments andthe internal vesicles of multivesicular bodies harbor most ofthe cholesterol found in the endocytic pathway. Traffic 2003,4:222-231.
www.sciencedirect.com
48. Chang TY, Reid PC, Sugii S, Ohgami N, Cruz JC, Chang CC:Niemann-Pick type C disease and intracellular cholesteroltrafficking. J Biol Chem 2005, 280:20917-20920.
49. Kobayashi T, Beuchat MH, Lindsay M, Frias S, Palmiter RD,Sakuraba H, Parton RG, Gruenberg J: Late endosomalmembranes rich in lysobisphosphatidic acid regulatecholesterol transport. Nat Cell Biol 1999, 1:113-118.
50. Megha, London E: Ceramide selectively displaces cholesterolfrom ordered lipid domains (rafts): implications for lipid raftstructure and function. J Biol Chem 2004, 279:9997-10004.
51. Osiecki-Newman K, Legler G, Grace M, Dinur T, Gatt S,Desnick RJ, Grabowski GA: Human acid beta-glucosidase:inhibition studies using glucose analogues and pH variation tocharacterize the normal and Gaucher disease glycon bindingsites. Enzyme 1988, 40:173-188.
52. Xu X, Bittman R, Duportail G, Heissler D, Vilcheze C,London E: Effect of the structure of natural sterols andsphingolipids on the formation of ordered sphingolipid/steroldomains (rafts). Comparison of cholesterol to plant, fungal,and disease-associated sterols and comparison ofsphingomyelin, cerebrosides, and ceramide. J Biol Chem 2001,276:33540-33546.
53. Terebiznik MR, Vieira OV, Marcus SL, Slade A, Yip CM,Trimble WS, Meyer T, Finlay BB, Grinstein S: Elimination ofhost cell PtdIns(4,5)P(2) by bacterial SigD promotesmembrane fission during invasion by Salmonella.Nat Cell Biol 2002, 4:766-773.
54.�
Hernandez LD, Hueffer K, Wenk MR, Galan JE: Salmonellamodulates vesicular traffic by altering phosphoinositidemetabolism. Science 2004, 304:1805-1807.
Reports the ability of the bacterial phosphatase SopB to catalyze thegeneration of PI(3)P, which modulated membrane traffic.
55.�
Vergne I, Chua J, Lee HH, Lucas M, Belisle J, Deretic V:Mechanism of phagolysosome biogenesis block by viableMycobacterium tuberculosis. Proc Natl Acad Sci USA 2005,102:4033-4038.
A mycobacterial phosphatase is found to reduce the phagosomal contentof PI(3)P, contributing to maturation arrest.
Current Opinion in Cell Biology 2006, 18:429–437