life in the suburbs: artificial heat source selection for
TRANSCRIPT

Web Ecol 20 161ndash172 2020httpsdoiorg105194we-20-161-2020copy Author(s) 2020 This work is distributed underthe Creative Commons Attribution 40 License
Life in the suburbs artificial heat source selection fornocturnal thermoregulation in a diurnally active
tropical lizard
NioKing Amadi1 Robert Belema1 Harrison Obodo Chukwu1 Daniele Dendi123 Amuzie Chidinma1Roger Meek4 and Luca Luiselli123
1Department of Animal and Environmental Biology Rivers State University Nkpolu-OroworukwoPO Box 5080 Port Harcourt Nigeria
2Institute for Development Ecology Conservation and Cooperation via G Tomasi di Lampedusa 3300144 Rome Italy
3Department of Zoology University of Lomeacute Lomeacute Togo4British Herpetological Society Regents Park London UK
Correspondence Roger Meek (rogermeek85aolcom)
Received 19 June 2020 ndash Revised 29 September 2020 ndash Accepted 21 October 2020 ndash Published 4 December 2020
Abstract The rapid expansion of urban environments invariably presents a novel series of pressures on wildlifedue to changes in external environmental factors In reptiles any such changes in temperature are critical sincethermoregulation is the key driver in the function of many physiological processes How reptiles adapt to suchchanges may vary from those species that are impacted negatively to others that have the behavioural flexibilityto exploit new conditions In this paper we describe retreat site selection movements and aspects of the thermalecology of the African lizard Agama agama in urban environments of West Africa In early evening lizards beganmovement from late-afternoon core activity areas and ascended the walls of houses for overnight retreats A highproportion retreated to locations in groups under or on top of warm electrical panels The thermal potential thesepanels offered was the attainment of body temperatures equal to or higher than the minimum preferred bodytemperature (PBTasymp 36 C in A agama) and hence increased physiological performance The lizards that tookadvantage of the heat sources travelled further each day to and from diurnal activity areas than individuals thatspent the night high on walls but not next to heat panels There were both potential costs (enhanced predationpressures) and benefits (impacts on thermal ecology retreat site selection) of this behaviour for lizards living inurban environments
1 Introduction
Retreat site selection is a key aspect in the study of adapta-tion sociality and predation risk in vertebrates (eg Camp-bell and Tobler 1984 Rasoloharijaona et al 2003 Rial etal 2010) The benefits from the thermal qualities of retreatsites are an important aspect for reptiles since body tem-perature levels influence among others rate of digestionsprint locomotion predation success and anti-predator tactics(Huey 1982 Downes and Shine 1998 Kearney 2002 Mor-ris and Lattanzio 2020) Additionally the effect of tempera-ture on embryo fitness and offspring survival may be depen-
dent on female reptiles selecting optimum location for eggdeposition in a thermal context (Loumlwenborg et al 2011) or inachieving optimum temperatures by diurnal basking (Lour-dais et al 2004) However the benefits involve risks Forexample gravid female aspic vipers (Vipera aspis) bask inexposed locations to enhance embryo development throughachieving optimum body temperatures but risk predation bypredators such as raptors (eg Lourdais et al 2004) andgravid female grass snakes (Natrix natrix) travel to commu-nal egg-laying sites that provide optimum temperatures forembryonic fitness (eg Meek 2017) but risk predation as aconsequence of increased movement and exposure (eg Bon-
Published by Copernicus Publications on behalf of the European Ecological Federation (EEF)
162 N Amadi et al Heat source selection of diurnally active tropical lizards
net et al 1999 Meek 2017) Therefore costs and benefitsof thermal ecology are a critical aspect of reptile life history
Selection of overnight retreats controls the microclimatesthat lizards experience when they are inactive thus enablingthem to thermoregulate by subtle movements within retreatareas (eg Porter et al 1973 Hitchcock and McBrayer2006) Indeed Porter et al (1973) showed that a hypothet-ical reptile might more easily control body temperatures inovernight retreats than during periods of diurnal activity andmay even be capable of maintaining preferred temperaturesfor longer
Although retreat site selection in reptiles has been stud-ied previously (eg Clark and Gillingham 1990 Poche etal 2005 Singhal and Johnson 2007 Meek 2008 Cabrera-Guzmaacuten and Reynoso 2010) data on overnight retreat se-lection and diurnal movement in relation to retreat site avail-ability especially in rapidly changing anthropogenic envi-ronments are limited Given that lizards possess limited dis-persal capacities they are particularly vulnerable to habi-tat changes Ecological research efforts are traditionally de-voted to natural and semi-natural habitats but studies on rep-tiles living in anthropogenic environments can provide in-sight into the degree of adaptability that has enabled somespecies for example A agama to be a widespread ecologicalsuccess both in terms of population densities and geographi-cal distribution Additionally optimal escape theory suggeststhat the risks and benefits gained from the distances involvedin travelling to retreat sites will impact on retreat site selec-tion (Schlesinger and Shine 1994 Webb et al 2004 Meek2008) and how reptiles respond to anthropogenic environ-ments is important from a conservation perspective
Agama lizards (family Agamidae) are a diverse group oftropical reptiles characterized by a great ecological plastic-ity (eg Harris 1964 Cloudsley-Thompson 1981) that hasenabled some species to survive even in areas outside theirnative African range (Enge et al 2004 Leache et al 2009Wagner et al 2009) An example of this ecological plastic-ity is the rainbow lizard (Agama agama) an agamid that hasadapted to life in suburban habitats throughout West Africa(Chapman and Chapman 1964 Akani et al 1999 2011Leacheacute et al 2017) and has been the focus of numerousstudies across African regions (eg Yeboah 1982 Anibaldiet al 1998 Adeoye and Ogunbanwo 2007) including theNiger Delta of Nigeria (eg Akani et al 2011 2013) A diur-nal species (Cloudsley-Thompson 1981 Yeboah 1982 Ani-baldi et al 1998) A agama on occasion may also be activenocturnally (Pauwels et al 2004) In the present paper weinvestigate the links between retreat site selection and move-ment ecology of A agama in urban areas of Rivers Statesouthern Nigeria We address the following key questions
1 Does A agama retreat to low-level or elevated perchesat night This question is relevant because agamidlizards have been observed to shift from terrestrial toarboreal habits by night (Cloudsley-Thompson 1981)
2 Does A agama select vertical or horizontal substrataas retreats sites This question is relevant becauselizards often show species-specific preferences for ver-tical or horizontal perches as they can maximize theirsprint performances in the event of attempted predation(Cooper 2006)
3 In retreat sites are A agama solitary or do they cluster ingroups This question is relevant because A agama hasnumerous predators (eg Luiselli et al 2002) and whenin inappropriate retreat sites may be at much higher riskof predation Selection of appropriate retreat sites mayminimize predation risk by employing group behaviourat night (Yorzinski and Platt 2012 Ellison et al 2019)
4 Is there any thermalanti-predation benefit in the associ-ation with heat panels as retreat sites This question isrelevant because thermal sensitivity in lizards impactson their ability to feed and evade predators due to in-creased muscular energy and running speeds with in-creasing body temperatures
5 Do the lizards exhibit sexual and age differences inmovement patterns between retreat sites and feeding ar-eas This question is relevant because we need to under-stand whether the benefits of increased heat availabilityare sex- or age-specific two aspects that are relativelypoorly known in tropical lizards studied at present Inthe context of question 5 we also report some observa-tions on the predation events suffered by A agama atthe study areas
2 Materials and methods
21 Study area
The field work was carried out in the Rivers State in the east-ern Niger Delta region of southern Nigeria The area has awet tropical climate with a temperature average of 25ndash28 C(up to 32ndash34 C by day) throughout the year The wet seasonoccurs from April to September and the dry season from Oc-tober to March (Amadi 2017) The area is characterized by aforestndashplantation mosaic with numerous small villages anda few large urban centres with Port Harcourt the largest cityin the state (Amadi 2017)
22 Protocol
The field study was carried out from 12 January to 22 Febru-ary 2020 around 81 houses and surrounding gardens in 61communities of Rivers State Field surveys were made everyday during this time period The geographic coordinates ofthe various sites are given in Appendix A
Data were gathered daily between 1700 and 2400 (Lagosstandard time) during which time the whole gardenprivateproperty of each sampled house was systematically searched
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 163
with the aid of flashlights from mobile devices to detect thepresence of lizards in their overnight retreats The lizardsin each microhabitat type were carefully counted and pho-tographed without disturbance The following microhabi-tatsubstratum types were classified trees and bushes ce-ment basements and walls When lizards were observedon cement basements and walls we recorded whether theywere sleepingresting within nearby electrical changeovers orswitches (ie electric panels) which were normally locatedat the front or back of houses We recorded whether the vari-ous individuals were sleepingresting (i) on a vertical or hor-izontal substratum (ii) alone or in a group and (iii) the num-ber of individuals forming each group was also noted We de-fined ldquoin a grouprdquo lizards that had at least another conspecificindividual at less than 30 cm linear distance The tempera-ture (C) of the substratum used by lizards for resting wasrecorded by Benetech Lasergrip GM320 laser electronic ther-mometer (01 C resolution and accuracy of plusmn02 C) Theheight of the lizards from the ground (in cm) was measuredwith a tape Furthermore we recorded the presence of preda-tors (cats and snakes) within each of the surveyed privateproperties and catalogued all instances of direct predationon A agama Between 1700ndash1800 we also conducted sys-tematic observations of several individuals in order to mea-sure exactly the linear distances (in m) they moved from theirldquolate-afternoon core activityrdquo area to overnight retreat sitesEighty-four lizards were monitored 20 juveniles 31 malesand 33 females by focal observations Each individual wasfollowed for at least 30 min from a distance of about 10 min such a way to avoid direct disturbance by the observer onits movements Then after each individual was observed inan overnight retreat the distance was measured with a tapefrom the ldquocorerdquo area (ie a point situated roughly in the mid-dle of the area where the lizard was seen while active) andthe overnight resting location Given that we did not monitorany individual for more than one day we could not determinewhether distance travelled by lizards could be influenced byfactors such as climate andor social interaction on a dailybasis
Data from individuals were collected once at each houseand not repeated within the structure thus avoiding pseudo-replication Frequency of predation events by snakes wasrecorded by inducing regurgitation of ingested meals bysnakes that were encountered in the studied suburban areas(see Luiselli and Amori 2016) predation by cats was ob-served directly
In this paper we discuss some of the possible benefits ofattaining preferred body temperatures when in retreat sitesby invoking Q10 effects on physiology Q10 is a measure ofthermal sensitivity of physiological processes with increasesof 10 C This enables lizards and other ectotherms to (for ex-ample) run faster or increase digestion rates Most of theQ10effects on physiological processes in reptiles range between
2 and 3 and are determined from
Q10 = (R2R1)10C(t2t1) (1)
where Q10 the measure of rate change is a unitless quantityand derived from R the rates of change between tempera-tures t2 and t1 in C AQ10 of 1 would indicate thermal inde-pendence values greater than 1 indicated the physiologicalprocesses have increasing thermal dependence An exampleof Q10 effects is increased muscle power and subsequent in-creases in reptile running speeds with increasing body tem-perature (Bennett 1990) This in turn impacts on the abilityof a reptile to secure prey or escape predators
23 Statistical analyses
Frequency differences of lizards in retreats were evaluatedby a three-way contingency table χ2 to test the indepen-dence of substratum type group or alone and vertical ver-sus horizontal substratum Correlation between height of thewall and number of sleeping lizards was assessed by Pear-sonrsquos product moment correlation coefficient Differences inthe substratum temperature of lizard sleeping spots betweenelectric panels versus other substratum types were evaluatedby two-way ANOVA with type of substratum and daytime(Lagos standard time) as the factors For distances movedversus association or non-association with electric compo-nents data sets were first examined for normality using An-derson a2 tests These showed normal distributions in bothsexes (a2 from 021 to 032 P values from 049 to 077)and hence comparisons were made using ANOVA Tem-peratures of electric boxes where lizards were found werecompared with temperatures of retreat sites without artificialheating using simple percentages These results were thenevaluated against known minimum preferred body temper-atures (PBTs) in A agama (James and Porter 1979) usingtwo-tailed probability z scores of the differences in percent-ages A sample precision test which evaluates sample sizeindicated maximum error was plusmn89 at the 95 intervalWe used one-way ANCOVA for testing the heterogeneity ofthe slopes of the regression line of the general correlationof wall height (cm) against the number of sleeping lizardsbetween electric panels and other retreat sites Comparisonof means was by the MannndashWhitney U test and comparisonof variances by Levenersquos test Both tests are less sensitive todepartures from normality The null hypothesis for Levenersquostest is H0 σ 2
1 σ22 = 1 at α = 005 In the text means are fol-
lowed byplusmn1 standard deviation All statistical analyses wereperformed with SigmaPlot 140 software and Minitab V17with α at 005
3 Results
A total of 166 electric panels were surveyed and 43 of them(348 ) were occupied by at least one A agama lizard
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
164 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 1 Example of Agama agama resting on (a) or below (b)warm electric panels for overnight retreats
Overall 435 lizards were observed in overnight retreats96 were observed on cement walls at an average height of1018plusmn 868 cm (median= 78 cm) Only 4 of the observedindividuals were on a treebush within the properties (fre-quency differences in the use of retreat site location were sig-nificantly different χ2 test P lt 00001) On multiple occa-sions (n= 14) we observed that from around 1800 lizardsstarted heading towards retreat sites from nearby bushesfences and ascended walls and positioned themselves un-der or on top of warm electrical panels The panels regu-late electrical energy sourced from personal generators orpower from national grid and emitted heat Lizards (n= 144331 ) were observed within 5 cm distance (Fig 1a) un-der or on top of these electric panels (Fig 1b) Most wereclustered in groups (Fig 2) only 14 of the total observed(n= 435) were solitary during nocturnal inactivity (χ2 testP lt 00001) When lizards were found in groups numbersranged from 2 to 12 and averaged 36plusmn 28 individuals (me-dia n= 3 individuals) Moreover 926 of the observed in-
Figure 2 Example of overnight clustering on walls in Agamaagama from the study locality
dividuals were found on vertical substrata and only 74 onhorizontal substratum (χ2 test P lt 00001) The numbersof lizards were not correlated to the height of the wall whenthey used electric panels (P gt 005) whereas it was signifi-cantly positively correlated when they selected other retreatsites (r = 0765 P lt 0001) A heterogeneity of slopes testrevealed that the two regressions did not differ significantly(one-way ANCOVA P gt 005 Fig 3) Lizards that werelocated next to electrical panels (mean= 1355plusmn 643 cm)were found at significantly higher elevations than those thatwere not next to panels (mean= 989plusmn 824 cm the MannndashWhitney U test w = 1418 P = 00009) with the variancesof heights around the means in good agreement (LevenersquosW = 0001 P = 098)
Mean substratum temperature was significantly higher atelectric panel retreat sites than non-artificially heated areas(F = 222 P = 0007 Fig 4) and also when diurnal tem-peratures were included in the analysis (two-way ANOVAF = 1763 P = 0023) So they were different in both struc-ture (panels versus no panels) and temperatures (heated ar-eas versus non-heated areas) The frequency distributions oflizard heights on walls next to heat panels and heights oflizards not located on unheated wall areas are given in Fig 5Diurnal temperatures were in good agreement for all the dif-
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 165
Figure 3 Relationship between wall height and number of sleep-ing rainbow lizards (Agama agama) in Port Harcourt city (southernNigeria) The two regression lines respectively show lizards usingelectric panels as retreat sites and lizards using other retreat sitesFor the statistical details see the text
Figure 4 Box plots showing differences between substratum tem-perature (C) at electric panels (symbol 1 on the x axis) and at non-artificially heated locations used by lizards (symbol 2 on the x axis)Mean range standard deviation and outliers are shown
ferent substratum types (F = 069 P = 0913) Percentagecomparisons of temperatures at electric boxes versus tem-peratures at non-artificially heat areas indicated that 593 of lizards close to the electric boxes could potentially achievenocturnal PBTs equal to or higher than the known minimumPBT in A agama (asymp 36 C) compared to 75 that were atnon-artificially heated localities (z= 748 P lt 0001) Thetemperatures for both areas in relation to minimum PBT areshown in Fig 6
A greater number of adults (males 74 females 576 )spent the night close to the electrical panels than juve-niles (25 n= 25 juveniles versus males z= 38P lt00001 n= 31 juveniles versus females z= 245 P =0014 n= 33) However there were no significant differ-ences between males and females (z= 14 P = 017 all
two-tailed tests) Males (n= 31 mean= 156plusmn 101 m) trav-elled greater distances between ldquolate-afternoon core diurnalactivity areasrdquo than females (n= 33 mean= 101plusmn 67 m)and both sexes travelled further than juveniles (n= 20mean= 52plusmn 41 m F(281) = 995 P lt 00001) with Tukeypost hoc tests indicating that the differences between eachcategory were significantly different in all groups Lizardsthat retreated next to or under electric panels travelledfurther each day to ldquolate-afternoon core activity areasrdquothan individuals that did not spend the night next to elec-trical panels (males= 21plusmn 152 versus 137plusmn 874 m fe-males= 148plusmn 77 versus 85plusmn 41) but the difference wasonly significant in females (males F(129) = 276 P = 011females F(1 31) = 906 P = 0005)
Recorded predation events upon A agama at the study ar-eas (total n= 66) included domestic cats (n= 21) and thesnakes Lamprophis fuliginosus (n= 32) Lamprophis linea-tus (n= 7) Crotaphopeltis hotamboeia (n= 5) and Najamelanoleuca (n= 1)
4 Discussion
Our study has presented several new findings on the biol-ogy of A agama in the suburban areas of Port Harcourt(i) most lizards used vertical surfaces for overnight retreats(ii) most spent the night in groups rather than as solitary in-dividuals (iii) the lizards used locations on relatively highcement based walls for overnight retreats (iv) adults pref-erentially spent the night next to or on electric panels butthis was less evident in juveniles and (v) there were signifi-cant differences between males females and juveniles in thedistances travelled between late-afternoon core activity ar-eas and overnight retreats In addition cats and snakes werefound to be the main observed predators for A agama liv-ing in suburban environments However it is implicit in thisstudy that we did not measure lizard body temperatures sincewe were careful to avoid disturbing behaviour which mayimpact on subsequent behaviour Nevertheless we are con-fident that lizards resting on or next to heat panels gainedextra heat compared to those lizards that did not and werebetter capable of achieving or exceeding minimum preferredbody temperatures
There are three possible advantages that elevated retreatsites can confer (1) reduced risk of predation from terres-trial predators (2) shelter from rain (Port Harcourt has anextremely wet climate especially from April to September)and (3) passive basking close to warm electric panels po-tentially enables lizards to achieve body temperatures withinthe minimum PBT threshold Overnight clustering in groupsby A agama contrasts with territorial behaviour in diurnalhabitats (eg Yeboah 1982 Madsen and Loman 1987 Ani-baldi et al 1998) and might suggest that group behaviouris an anti-predatory strategy (Landeau and Terborgh 1986Krause et al 2002 Beauchamp 2013) or that electrical pan-
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

162 N Amadi et al Heat source selection of diurnally active tropical lizards
net et al 1999 Meek 2017) Therefore costs and benefitsof thermal ecology are a critical aspect of reptile life history
Selection of overnight retreats controls the microclimatesthat lizards experience when they are inactive thus enablingthem to thermoregulate by subtle movements within retreatareas (eg Porter et al 1973 Hitchcock and McBrayer2006) Indeed Porter et al (1973) showed that a hypothet-ical reptile might more easily control body temperatures inovernight retreats than during periods of diurnal activity andmay even be capable of maintaining preferred temperaturesfor longer
Although retreat site selection in reptiles has been stud-ied previously (eg Clark and Gillingham 1990 Poche etal 2005 Singhal and Johnson 2007 Meek 2008 Cabrera-Guzmaacuten and Reynoso 2010) data on overnight retreat se-lection and diurnal movement in relation to retreat site avail-ability especially in rapidly changing anthropogenic envi-ronments are limited Given that lizards possess limited dis-persal capacities they are particularly vulnerable to habi-tat changes Ecological research efforts are traditionally de-voted to natural and semi-natural habitats but studies on rep-tiles living in anthropogenic environments can provide in-sight into the degree of adaptability that has enabled somespecies for example A agama to be a widespread ecologicalsuccess both in terms of population densities and geographi-cal distribution Additionally optimal escape theory suggeststhat the risks and benefits gained from the distances involvedin travelling to retreat sites will impact on retreat site selec-tion (Schlesinger and Shine 1994 Webb et al 2004 Meek2008) and how reptiles respond to anthropogenic environ-ments is important from a conservation perspective
Agama lizards (family Agamidae) are a diverse group oftropical reptiles characterized by a great ecological plastic-ity (eg Harris 1964 Cloudsley-Thompson 1981) that hasenabled some species to survive even in areas outside theirnative African range (Enge et al 2004 Leache et al 2009Wagner et al 2009) An example of this ecological plastic-ity is the rainbow lizard (Agama agama) an agamid that hasadapted to life in suburban habitats throughout West Africa(Chapman and Chapman 1964 Akani et al 1999 2011Leacheacute et al 2017) and has been the focus of numerousstudies across African regions (eg Yeboah 1982 Anibaldiet al 1998 Adeoye and Ogunbanwo 2007) including theNiger Delta of Nigeria (eg Akani et al 2011 2013) A diur-nal species (Cloudsley-Thompson 1981 Yeboah 1982 Ani-baldi et al 1998) A agama on occasion may also be activenocturnally (Pauwels et al 2004) In the present paper weinvestigate the links between retreat site selection and move-ment ecology of A agama in urban areas of Rivers Statesouthern Nigeria We address the following key questions
1 Does A agama retreat to low-level or elevated perchesat night This question is relevant because agamidlizards have been observed to shift from terrestrial toarboreal habits by night (Cloudsley-Thompson 1981)
2 Does A agama select vertical or horizontal substrataas retreats sites This question is relevant becauselizards often show species-specific preferences for ver-tical or horizontal perches as they can maximize theirsprint performances in the event of attempted predation(Cooper 2006)
3 In retreat sites are A agama solitary or do they cluster ingroups This question is relevant because A agama hasnumerous predators (eg Luiselli et al 2002) and whenin inappropriate retreat sites may be at much higher riskof predation Selection of appropriate retreat sites mayminimize predation risk by employing group behaviourat night (Yorzinski and Platt 2012 Ellison et al 2019)
4 Is there any thermalanti-predation benefit in the associ-ation with heat panels as retreat sites This question isrelevant because thermal sensitivity in lizards impactson their ability to feed and evade predators due to in-creased muscular energy and running speeds with in-creasing body temperatures
5 Do the lizards exhibit sexual and age differences inmovement patterns between retreat sites and feeding ar-eas This question is relevant because we need to under-stand whether the benefits of increased heat availabilityare sex- or age-specific two aspects that are relativelypoorly known in tropical lizards studied at present Inthe context of question 5 we also report some observa-tions on the predation events suffered by A agama atthe study areas
2 Materials and methods
21 Study area
The field work was carried out in the Rivers State in the east-ern Niger Delta region of southern Nigeria The area has awet tropical climate with a temperature average of 25ndash28 C(up to 32ndash34 C by day) throughout the year The wet seasonoccurs from April to September and the dry season from Oc-tober to March (Amadi 2017) The area is characterized by aforestndashplantation mosaic with numerous small villages anda few large urban centres with Port Harcourt the largest cityin the state (Amadi 2017)
22 Protocol
The field study was carried out from 12 January to 22 Febru-ary 2020 around 81 houses and surrounding gardens in 61communities of Rivers State Field surveys were made everyday during this time period The geographic coordinates ofthe various sites are given in Appendix A
Data were gathered daily between 1700 and 2400 (Lagosstandard time) during which time the whole gardenprivateproperty of each sampled house was systematically searched
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 163
with the aid of flashlights from mobile devices to detect thepresence of lizards in their overnight retreats The lizardsin each microhabitat type were carefully counted and pho-tographed without disturbance The following microhabi-tatsubstratum types were classified trees and bushes ce-ment basements and walls When lizards were observedon cement basements and walls we recorded whether theywere sleepingresting within nearby electrical changeovers orswitches (ie electric panels) which were normally locatedat the front or back of houses We recorded whether the vari-ous individuals were sleepingresting (i) on a vertical or hor-izontal substratum (ii) alone or in a group and (iii) the num-ber of individuals forming each group was also noted We de-fined ldquoin a grouprdquo lizards that had at least another conspecificindividual at less than 30 cm linear distance The tempera-ture (C) of the substratum used by lizards for resting wasrecorded by Benetech Lasergrip GM320 laser electronic ther-mometer (01 C resolution and accuracy of plusmn02 C) Theheight of the lizards from the ground (in cm) was measuredwith a tape Furthermore we recorded the presence of preda-tors (cats and snakes) within each of the surveyed privateproperties and catalogued all instances of direct predationon A agama Between 1700ndash1800 we also conducted sys-tematic observations of several individuals in order to mea-sure exactly the linear distances (in m) they moved from theirldquolate-afternoon core activityrdquo area to overnight retreat sitesEighty-four lizards were monitored 20 juveniles 31 malesand 33 females by focal observations Each individual wasfollowed for at least 30 min from a distance of about 10 min such a way to avoid direct disturbance by the observer onits movements Then after each individual was observed inan overnight retreat the distance was measured with a tapefrom the ldquocorerdquo area (ie a point situated roughly in the mid-dle of the area where the lizard was seen while active) andthe overnight resting location Given that we did not monitorany individual for more than one day we could not determinewhether distance travelled by lizards could be influenced byfactors such as climate andor social interaction on a dailybasis
Data from individuals were collected once at each houseand not repeated within the structure thus avoiding pseudo-replication Frequency of predation events by snakes wasrecorded by inducing regurgitation of ingested meals bysnakes that were encountered in the studied suburban areas(see Luiselli and Amori 2016) predation by cats was ob-served directly
In this paper we discuss some of the possible benefits ofattaining preferred body temperatures when in retreat sitesby invoking Q10 effects on physiology Q10 is a measure ofthermal sensitivity of physiological processes with increasesof 10 C This enables lizards and other ectotherms to (for ex-ample) run faster or increase digestion rates Most of theQ10effects on physiological processes in reptiles range between
2 and 3 and are determined from
Q10 = (R2R1)10C(t2t1) (1)
where Q10 the measure of rate change is a unitless quantityand derived from R the rates of change between tempera-tures t2 and t1 in C AQ10 of 1 would indicate thermal inde-pendence values greater than 1 indicated the physiologicalprocesses have increasing thermal dependence An exampleof Q10 effects is increased muscle power and subsequent in-creases in reptile running speeds with increasing body tem-perature (Bennett 1990) This in turn impacts on the abilityof a reptile to secure prey or escape predators
23 Statistical analyses
Frequency differences of lizards in retreats were evaluatedby a three-way contingency table χ2 to test the indepen-dence of substratum type group or alone and vertical ver-sus horizontal substratum Correlation between height of thewall and number of sleeping lizards was assessed by Pear-sonrsquos product moment correlation coefficient Differences inthe substratum temperature of lizard sleeping spots betweenelectric panels versus other substratum types were evaluatedby two-way ANOVA with type of substratum and daytime(Lagos standard time) as the factors For distances movedversus association or non-association with electric compo-nents data sets were first examined for normality using An-derson a2 tests These showed normal distributions in bothsexes (a2 from 021 to 032 P values from 049 to 077)and hence comparisons were made using ANOVA Tem-peratures of electric boxes where lizards were found werecompared with temperatures of retreat sites without artificialheating using simple percentages These results were thenevaluated against known minimum preferred body temper-atures (PBTs) in A agama (James and Porter 1979) usingtwo-tailed probability z scores of the differences in percent-ages A sample precision test which evaluates sample sizeindicated maximum error was plusmn89 at the 95 intervalWe used one-way ANCOVA for testing the heterogeneity ofthe slopes of the regression line of the general correlationof wall height (cm) against the number of sleeping lizardsbetween electric panels and other retreat sites Comparisonof means was by the MannndashWhitney U test and comparisonof variances by Levenersquos test Both tests are less sensitive todepartures from normality The null hypothesis for Levenersquostest is H0 σ 2
1 σ22 = 1 at α = 005 In the text means are fol-
lowed byplusmn1 standard deviation All statistical analyses wereperformed with SigmaPlot 140 software and Minitab V17with α at 005
3 Results
A total of 166 electric panels were surveyed and 43 of them(348 ) were occupied by at least one A agama lizard
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
164 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 1 Example of Agama agama resting on (a) or below (b)warm electric panels for overnight retreats
Overall 435 lizards were observed in overnight retreats96 were observed on cement walls at an average height of1018plusmn 868 cm (median= 78 cm) Only 4 of the observedindividuals were on a treebush within the properties (fre-quency differences in the use of retreat site location were sig-nificantly different χ2 test P lt 00001) On multiple occa-sions (n= 14) we observed that from around 1800 lizardsstarted heading towards retreat sites from nearby bushesfences and ascended walls and positioned themselves un-der or on top of warm electrical panels The panels regu-late electrical energy sourced from personal generators orpower from national grid and emitted heat Lizards (n= 144331 ) were observed within 5 cm distance (Fig 1a) un-der or on top of these electric panels (Fig 1b) Most wereclustered in groups (Fig 2) only 14 of the total observed(n= 435) were solitary during nocturnal inactivity (χ2 testP lt 00001) When lizards were found in groups numbersranged from 2 to 12 and averaged 36plusmn 28 individuals (me-dia n= 3 individuals) Moreover 926 of the observed in-
Figure 2 Example of overnight clustering on walls in Agamaagama from the study locality
dividuals were found on vertical substrata and only 74 onhorizontal substratum (χ2 test P lt 00001) The numbersof lizards were not correlated to the height of the wall whenthey used electric panels (P gt 005) whereas it was signifi-cantly positively correlated when they selected other retreatsites (r = 0765 P lt 0001) A heterogeneity of slopes testrevealed that the two regressions did not differ significantly(one-way ANCOVA P gt 005 Fig 3) Lizards that werelocated next to electrical panels (mean= 1355plusmn 643 cm)were found at significantly higher elevations than those thatwere not next to panels (mean= 989plusmn 824 cm the MannndashWhitney U test w = 1418 P = 00009) with the variancesof heights around the means in good agreement (LevenersquosW = 0001 P = 098)
Mean substratum temperature was significantly higher atelectric panel retreat sites than non-artificially heated areas(F = 222 P = 0007 Fig 4) and also when diurnal tem-peratures were included in the analysis (two-way ANOVAF = 1763 P = 0023) So they were different in both struc-ture (panels versus no panels) and temperatures (heated ar-eas versus non-heated areas) The frequency distributions oflizard heights on walls next to heat panels and heights oflizards not located on unheated wall areas are given in Fig 5Diurnal temperatures were in good agreement for all the dif-
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 165
Figure 3 Relationship between wall height and number of sleep-ing rainbow lizards (Agama agama) in Port Harcourt city (southernNigeria) The two regression lines respectively show lizards usingelectric panels as retreat sites and lizards using other retreat sitesFor the statistical details see the text
Figure 4 Box plots showing differences between substratum tem-perature (C) at electric panels (symbol 1 on the x axis) and at non-artificially heated locations used by lizards (symbol 2 on the x axis)Mean range standard deviation and outliers are shown
ferent substratum types (F = 069 P = 0913) Percentagecomparisons of temperatures at electric boxes versus tem-peratures at non-artificially heat areas indicated that 593 of lizards close to the electric boxes could potentially achievenocturnal PBTs equal to or higher than the known minimumPBT in A agama (asymp 36 C) compared to 75 that were atnon-artificially heated localities (z= 748 P lt 0001) Thetemperatures for both areas in relation to minimum PBT areshown in Fig 6
A greater number of adults (males 74 females 576 )spent the night close to the electrical panels than juve-niles (25 n= 25 juveniles versus males z= 38P lt00001 n= 31 juveniles versus females z= 245 P =0014 n= 33) However there were no significant differ-ences between males and females (z= 14 P = 017 all
two-tailed tests) Males (n= 31 mean= 156plusmn 101 m) trav-elled greater distances between ldquolate-afternoon core diurnalactivity areasrdquo than females (n= 33 mean= 101plusmn 67 m)and both sexes travelled further than juveniles (n= 20mean= 52plusmn 41 m F(281) = 995 P lt 00001) with Tukeypost hoc tests indicating that the differences between eachcategory were significantly different in all groups Lizardsthat retreated next to or under electric panels travelledfurther each day to ldquolate-afternoon core activity areasrdquothan individuals that did not spend the night next to elec-trical panels (males= 21plusmn 152 versus 137plusmn 874 m fe-males= 148plusmn 77 versus 85plusmn 41) but the difference wasonly significant in females (males F(129) = 276 P = 011females F(1 31) = 906 P = 0005)
Recorded predation events upon A agama at the study ar-eas (total n= 66) included domestic cats (n= 21) and thesnakes Lamprophis fuliginosus (n= 32) Lamprophis linea-tus (n= 7) Crotaphopeltis hotamboeia (n= 5) and Najamelanoleuca (n= 1)
4 Discussion
Our study has presented several new findings on the biol-ogy of A agama in the suburban areas of Port Harcourt(i) most lizards used vertical surfaces for overnight retreats(ii) most spent the night in groups rather than as solitary in-dividuals (iii) the lizards used locations on relatively highcement based walls for overnight retreats (iv) adults pref-erentially spent the night next to or on electric panels butthis was less evident in juveniles and (v) there were signifi-cant differences between males females and juveniles in thedistances travelled between late-afternoon core activity ar-eas and overnight retreats In addition cats and snakes werefound to be the main observed predators for A agama liv-ing in suburban environments However it is implicit in thisstudy that we did not measure lizard body temperatures sincewe were careful to avoid disturbing behaviour which mayimpact on subsequent behaviour Nevertheless we are con-fident that lizards resting on or next to heat panels gainedextra heat compared to those lizards that did not and werebetter capable of achieving or exceeding minimum preferredbody temperatures
There are three possible advantages that elevated retreatsites can confer (1) reduced risk of predation from terres-trial predators (2) shelter from rain (Port Harcourt has anextremely wet climate especially from April to September)and (3) passive basking close to warm electric panels po-tentially enables lizards to achieve body temperatures withinthe minimum PBT threshold Overnight clustering in groupsby A agama contrasts with territorial behaviour in diurnalhabitats (eg Yeboah 1982 Madsen and Loman 1987 Ani-baldi et al 1998) and might suggest that group behaviouris an anti-predatory strategy (Landeau and Terborgh 1986Krause et al 2002 Beauchamp 2013) or that electrical pan-
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

N Amadi et al Heat source selection of diurnally active tropical lizards 163
with the aid of flashlights from mobile devices to detect thepresence of lizards in their overnight retreats The lizardsin each microhabitat type were carefully counted and pho-tographed without disturbance The following microhabi-tatsubstratum types were classified trees and bushes ce-ment basements and walls When lizards were observedon cement basements and walls we recorded whether theywere sleepingresting within nearby electrical changeovers orswitches (ie electric panels) which were normally locatedat the front or back of houses We recorded whether the vari-ous individuals were sleepingresting (i) on a vertical or hor-izontal substratum (ii) alone or in a group and (iii) the num-ber of individuals forming each group was also noted We de-fined ldquoin a grouprdquo lizards that had at least another conspecificindividual at less than 30 cm linear distance The tempera-ture (C) of the substratum used by lizards for resting wasrecorded by Benetech Lasergrip GM320 laser electronic ther-mometer (01 C resolution and accuracy of plusmn02 C) Theheight of the lizards from the ground (in cm) was measuredwith a tape Furthermore we recorded the presence of preda-tors (cats and snakes) within each of the surveyed privateproperties and catalogued all instances of direct predationon A agama Between 1700ndash1800 we also conducted sys-tematic observations of several individuals in order to mea-sure exactly the linear distances (in m) they moved from theirldquolate-afternoon core activityrdquo area to overnight retreat sitesEighty-four lizards were monitored 20 juveniles 31 malesand 33 females by focal observations Each individual wasfollowed for at least 30 min from a distance of about 10 min such a way to avoid direct disturbance by the observer onits movements Then after each individual was observed inan overnight retreat the distance was measured with a tapefrom the ldquocorerdquo area (ie a point situated roughly in the mid-dle of the area where the lizard was seen while active) andthe overnight resting location Given that we did not monitorany individual for more than one day we could not determinewhether distance travelled by lizards could be influenced byfactors such as climate andor social interaction on a dailybasis
Data from individuals were collected once at each houseand not repeated within the structure thus avoiding pseudo-replication Frequency of predation events by snakes wasrecorded by inducing regurgitation of ingested meals bysnakes that were encountered in the studied suburban areas(see Luiselli and Amori 2016) predation by cats was ob-served directly
In this paper we discuss some of the possible benefits ofattaining preferred body temperatures when in retreat sitesby invoking Q10 effects on physiology Q10 is a measure ofthermal sensitivity of physiological processes with increasesof 10 C This enables lizards and other ectotherms to (for ex-ample) run faster or increase digestion rates Most of theQ10effects on physiological processes in reptiles range between
2 and 3 and are determined from
Q10 = (R2R1)10C(t2t1) (1)
where Q10 the measure of rate change is a unitless quantityand derived from R the rates of change between tempera-tures t2 and t1 in C AQ10 of 1 would indicate thermal inde-pendence values greater than 1 indicated the physiologicalprocesses have increasing thermal dependence An exampleof Q10 effects is increased muscle power and subsequent in-creases in reptile running speeds with increasing body tem-perature (Bennett 1990) This in turn impacts on the abilityof a reptile to secure prey or escape predators
23 Statistical analyses
Frequency differences of lizards in retreats were evaluatedby a three-way contingency table χ2 to test the indepen-dence of substratum type group or alone and vertical ver-sus horizontal substratum Correlation between height of thewall and number of sleeping lizards was assessed by Pear-sonrsquos product moment correlation coefficient Differences inthe substratum temperature of lizard sleeping spots betweenelectric panels versus other substratum types were evaluatedby two-way ANOVA with type of substratum and daytime(Lagos standard time) as the factors For distances movedversus association or non-association with electric compo-nents data sets were first examined for normality using An-derson a2 tests These showed normal distributions in bothsexes (a2 from 021 to 032 P values from 049 to 077)and hence comparisons were made using ANOVA Tem-peratures of electric boxes where lizards were found werecompared with temperatures of retreat sites without artificialheating using simple percentages These results were thenevaluated against known minimum preferred body temper-atures (PBTs) in A agama (James and Porter 1979) usingtwo-tailed probability z scores of the differences in percent-ages A sample precision test which evaluates sample sizeindicated maximum error was plusmn89 at the 95 intervalWe used one-way ANCOVA for testing the heterogeneity ofthe slopes of the regression line of the general correlationof wall height (cm) against the number of sleeping lizardsbetween electric panels and other retreat sites Comparisonof means was by the MannndashWhitney U test and comparisonof variances by Levenersquos test Both tests are less sensitive todepartures from normality The null hypothesis for Levenersquostest is H0 σ 2
1 σ22 = 1 at α = 005 In the text means are fol-
lowed byplusmn1 standard deviation All statistical analyses wereperformed with SigmaPlot 140 software and Minitab V17with α at 005
3 Results
A total of 166 electric panels were surveyed and 43 of them(348 ) were occupied by at least one A agama lizard
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
164 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 1 Example of Agama agama resting on (a) or below (b)warm electric panels for overnight retreats
Overall 435 lizards were observed in overnight retreats96 were observed on cement walls at an average height of1018plusmn 868 cm (median= 78 cm) Only 4 of the observedindividuals were on a treebush within the properties (fre-quency differences in the use of retreat site location were sig-nificantly different χ2 test P lt 00001) On multiple occa-sions (n= 14) we observed that from around 1800 lizardsstarted heading towards retreat sites from nearby bushesfences and ascended walls and positioned themselves un-der or on top of warm electrical panels The panels regu-late electrical energy sourced from personal generators orpower from national grid and emitted heat Lizards (n= 144331 ) were observed within 5 cm distance (Fig 1a) un-der or on top of these electric panels (Fig 1b) Most wereclustered in groups (Fig 2) only 14 of the total observed(n= 435) were solitary during nocturnal inactivity (χ2 testP lt 00001) When lizards were found in groups numbersranged from 2 to 12 and averaged 36plusmn 28 individuals (me-dia n= 3 individuals) Moreover 926 of the observed in-
Figure 2 Example of overnight clustering on walls in Agamaagama from the study locality
dividuals were found on vertical substrata and only 74 onhorizontal substratum (χ2 test P lt 00001) The numbersof lizards were not correlated to the height of the wall whenthey used electric panels (P gt 005) whereas it was signifi-cantly positively correlated when they selected other retreatsites (r = 0765 P lt 0001) A heterogeneity of slopes testrevealed that the two regressions did not differ significantly(one-way ANCOVA P gt 005 Fig 3) Lizards that werelocated next to electrical panels (mean= 1355plusmn 643 cm)were found at significantly higher elevations than those thatwere not next to panels (mean= 989plusmn 824 cm the MannndashWhitney U test w = 1418 P = 00009) with the variancesof heights around the means in good agreement (LevenersquosW = 0001 P = 098)
Mean substratum temperature was significantly higher atelectric panel retreat sites than non-artificially heated areas(F = 222 P = 0007 Fig 4) and also when diurnal tem-peratures were included in the analysis (two-way ANOVAF = 1763 P = 0023) So they were different in both struc-ture (panels versus no panels) and temperatures (heated ar-eas versus non-heated areas) The frequency distributions oflizard heights on walls next to heat panels and heights oflizards not located on unheated wall areas are given in Fig 5Diurnal temperatures were in good agreement for all the dif-
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 165
Figure 3 Relationship between wall height and number of sleep-ing rainbow lizards (Agama agama) in Port Harcourt city (southernNigeria) The two regression lines respectively show lizards usingelectric panels as retreat sites and lizards using other retreat sitesFor the statistical details see the text
Figure 4 Box plots showing differences between substratum tem-perature (C) at electric panels (symbol 1 on the x axis) and at non-artificially heated locations used by lizards (symbol 2 on the x axis)Mean range standard deviation and outliers are shown
ferent substratum types (F = 069 P = 0913) Percentagecomparisons of temperatures at electric boxes versus tem-peratures at non-artificially heat areas indicated that 593 of lizards close to the electric boxes could potentially achievenocturnal PBTs equal to or higher than the known minimumPBT in A agama (asymp 36 C) compared to 75 that were atnon-artificially heated localities (z= 748 P lt 0001) Thetemperatures for both areas in relation to minimum PBT areshown in Fig 6
A greater number of adults (males 74 females 576 )spent the night close to the electrical panels than juve-niles (25 n= 25 juveniles versus males z= 38P lt00001 n= 31 juveniles versus females z= 245 P =0014 n= 33) However there were no significant differ-ences between males and females (z= 14 P = 017 all
two-tailed tests) Males (n= 31 mean= 156plusmn 101 m) trav-elled greater distances between ldquolate-afternoon core diurnalactivity areasrdquo than females (n= 33 mean= 101plusmn 67 m)and both sexes travelled further than juveniles (n= 20mean= 52plusmn 41 m F(281) = 995 P lt 00001) with Tukeypost hoc tests indicating that the differences between eachcategory were significantly different in all groups Lizardsthat retreated next to or under electric panels travelledfurther each day to ldquolate-afternoon core activity areasrdquothan individuals that did not spend the night next to elec-trical panels (males= 21plusmn 152 versus 137plusmn 874 m fe-males= 148plusmn 77 versus 85plusmn 41) but the difference wasonly significant in females (males F(129) = 276 P = 011females F(1 31) = 906 P = 0005)
Recorded predation events upon A agama at the study ar-eas (total n= 66) included domestic cats (n= 21) and thesnakes Lamprophis fuliginosus (n= 32) Lamprophis linea-tus (n= 7) Crotaphopeltis hotamboeia (n= 5) and Najamelanoleuca (n= 1)
4 Discussion
Our study has presented several new findings on the biol-ogy of A agama in the suburban areas of Port Harcourt(i) most lizards used vertical surfaces for overnight retreats(ii) most spent the night in groups rather than as solitary in-dividuals (iii) the lizards used locations on relatively highcement based walls for overnight retreats (iv) adults pref-erentially spent the night next to or on electric panels butthis was less evident in juveniles and (v) there were signifi-cant differences between males females and juveniles in thedistances travelled between late-afternoon core activity ar-eas and overnight retreats In addition cats and snakes werefound to be the main observed predators for A agama liv-ing in suburban environments However it is implicit in thisstudy that we did not measure lizard body temperatures sincewe were careful to avoid disturbing behaviour which mayimpact on subsequent behaviour Nevertheless we are con-fident that lizards resting on or next to heat panels gainedextra heat compared to those lizards that did not and werebetter capable of achieving or exceeding minimum preferredbody temperatures
There are three possible advantages that elevated retreatsites can confer (1) reduced risk of predation from terres-trial predators (2) shelter from rain (Port Harcourt has anextremely wet climate especially from April to September)and (3) passive basking close to warm electric panels po-tentially enables lizards to achieve body temperatures withinthe minimum PBT threshold Overnight clustering in groupsby A agama contrasts with territorial behaviour in diurnalhabitats (eg Yeboah 1982 Madsen and Loman 1987 Ani-baldi et al 1998) and might suggest that group behaviouris an anti-predatory strategy (Landeau and Terborgh 1986Krause et al 2002 Beauchamp 2013) or that electrical pan-
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-
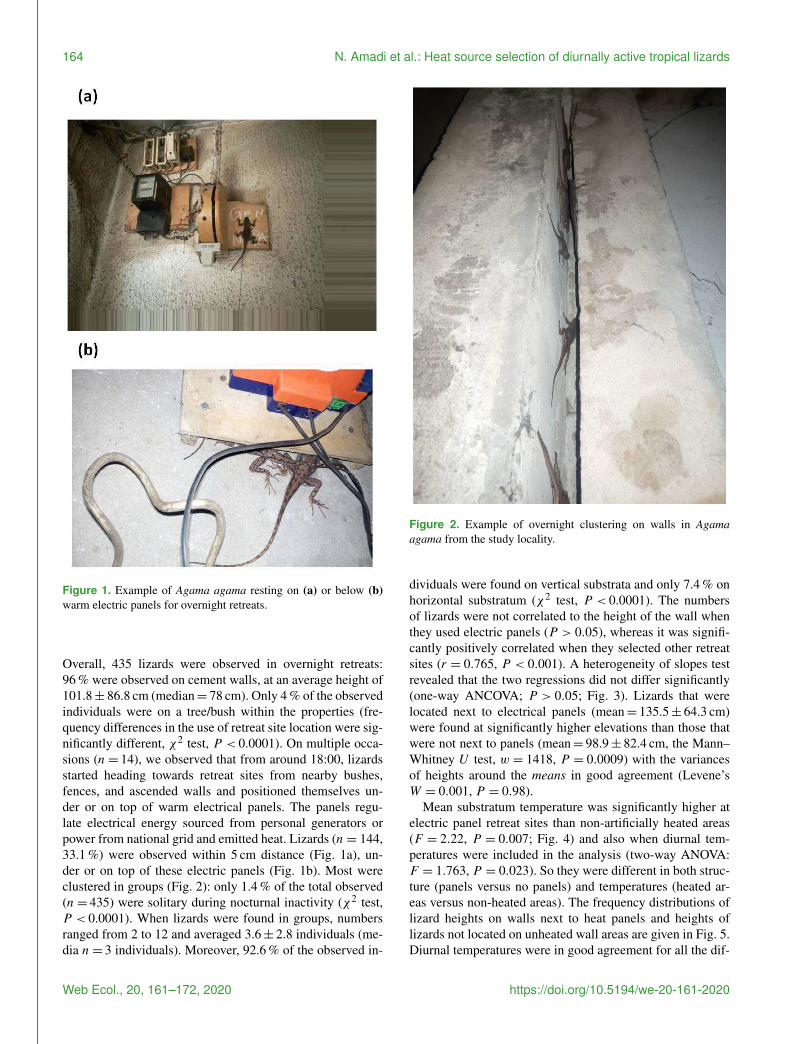
164 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 1 Example of Agama agama resting on (a) or below (b)warm electric panels for overnight retreats
Overall 435 lizards were observed in overnight retreats96 were observed on cement walls at an average height of1018plusmn 868 cm (median= 78 cm) Only 4 of the observedindividuals were on a treebush within the properties (fre-quency differences in the use of retreat site location were sig-nificantly different χ2 test P lt 00001) On multiple occa-sions (n= 14) we observed that from around 1800 lizardsstarted heading towards retreat sites from nearby bushesfences and ascended walls and positioned themselves un-der or on top of warm electrical panels The panels regu-late electrical energy sourced from personal generators orpower from national grid and emitted heat Lizards (n= 144331 ) were observed within 5 cm distance (Fig 1a) un-der or on top of these electric panels (Fig 1b) Most wereclustered in groups (Fig 2) only 14 of the total observed(n= 435) were solitary during nocturnal inactivity (χ2 testP lt 00001) When lizards were found in groups numbersranged from 2 to 12 and averaged 36plusmn 28 individuals (me-dia n= 3 individuals) Moreover 926 of the observed in-
Figure 2 Example of overnight clustering on walls in Agamaagama from the study locality
dividuals were found on vertical substrata and only 74 onhorizontal substratum (χ2 test P lt 00001) The numbersof lizards were not correlated to the height of the wall whenthey used electric panels (P gt 005) whereas it was signifi-cantly positively correlated when they selected other retreatsites (r = 0765 P lt 0001) A heterogeneity of slopes testrevealed that the two regressions did not differ significantly(one-way ANCOVA P gt 005 Fig 3) Lizards that werelocated next to electrical panels (mean= 1355plusmn 643 cm)were found at significantly higher elevations than those thatwere not next to panels (mean= 989plusmn 824 cm the MannndashWhitney U test w = 1418 P = 00009) with the variancesof heights around the means in good agreement (LevenersquosW = 0001 P = 098)
Mean substratum temperature was significantly higher atelectric panel retreat sites than non-artificially heated areas(F = 222 P = 0007 Fig 4) and also when diurnal tem-peratures were included in the analysis (two-way ANOVAF = 1763 P = 0023) So they were different in both struc-ture (panels versus no panels) and temperatures (heated ar-eas versus non-heated areas) The frequency distributions oflizard heights on walls next to heat panels and heights oflizards not located on unheated wall areas are given in Fig 5Diurnal temperatures were in good agreement for all the dif-
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 165
Figure 3 Relationship between wall height and number of sleep-ing rainbow lizards (Agama agama) in Port Harcourt city (southernNigeria) The two regression lines respectively show lizards usingelectric panels as retreat sites and lizards using other retreat sitesFor the statistical details see the text
Figure 4 Box plots showing differences between substratum tem-perature (C) at electric panels (symbol 1 on the x axis) and at non-artificially heated locations used by lizards (symbol 2 on the x axis)Mean range standard deviation and outliers are shown
ferent substratum types (F = 069 P = 0913) Percentagecomparisons of temperatures at electric boxes versus tem-peratures at non-artificially heat areas indicated that 593 of lizards close to the electric boxes could potentially achievenocturnal PBTs equal to or higher than the known minimumPBT in A agama (asymp 36 C) compared to 75 that were atnon-artificially heated localities (z= 748 P lt 0001) Thetemperatures for both areas in relation to minimum PBT areshown in Fig 6
A greater number of adults (males 74 females 576 )spent the night close to the electrical panels than juve-niles (25 n= 25 juveniles versus males z= 38P lt00001 n= 31 juveniles versus females z= 245 P =0014 n= 33) However there were no significant differ-ences between males and females (z= 14 P = 017 all
two-tailed tests) Males (n= 31 mean= 156plusmn 101 m) trav-elled greater distances between ldquolate-afternoon core diurnalactivity areasrdquo than females (n= 33 mean= 101plusmn 67 m)and both sexes travelled further than juveniles (n= 20mean= 52plusmn 41 m F(281) = 995 P lt 00001) with Tukeypost hoc tests indicating that the differences between eachcategory were significantly different in all groups Lizardsthat retreated next to or under electric panels travelledfurther each day to ldquolate-afternoon core activity areasrdquothan individuals that did not spend the night next to elec-trical panels (males= 21plusmn 152 versus 137plusmn 874 m fe-males= 148plusmn 77 versus 85plusmn 41) but the difference wasonly significant in females (males F(129) = 276 P = 011females F(1 31) = 906 P = 0005)
Recorded predation events upon A agama at the study ar-eas (total n= 66) included domestic cats (n= 21) and thesnakes Lamprophis fuliginosus (n= 32) Lamprophis linea-tus (n= 7) Crotaphopeltis hotamboeia (n= 5) and Najamelanoleuca (n= 1)
4 Discussion
Our study has presented several new findings on the biol-ogy of A agama in the suburban areas of Port Harcourt(i) most lizards used vertical surfaces for overnight retreats(ii) most spent the night in groups rather than as solitary in-dividuals (iii) the lizards used locations on relatively highcement based walls for overnight retreats (iv) adults pref-erentially spent the night next to or on electric panels butthis was less evident in juveniles and (v) there were signifi-cant differences between males females and juveniles in thedistances travelled between late-afternoon core activity ar-eas and overnight retreats In addition cats and snakes werefound to be the main observed predators for A agama liv-ing in suburban environments However it is implicit in thisstudy that we did not measure lizard body temperatures sincewe were careful to avoid disturbing behaviour which mayimpact on subsequent behaviour Nevertheless we are con-fident that lizards resting on or next to heat panels gainedextra heat compared to those lizards that did not and werebetter capable of achieving or exceeding minimum preferredbody temperatures
There are three possible advantages that elevated retreatsites can confer (1) reduced risk of predation from terres-trial predators (2) shelter from rain (Port Harcourt has anextremely wet climate especially from April to September)and (3) passive basking close to warm electric panels po-tentially enables lizards to achieve body temperatures withinthe minimum PBT threshold Overnight clustering in groupsby A agama contrasts with territorial behaviour in diurnalhabitats (eg Yeboah 1982 Madsen and Loman 1987 Ani-baldi et al 1998) and might suggest that group behaviouris an anti-predatory strategy (Landeau and Terborgh 1986Krause et al 2002 Beauchamp 2013) or that electrical pan-
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-
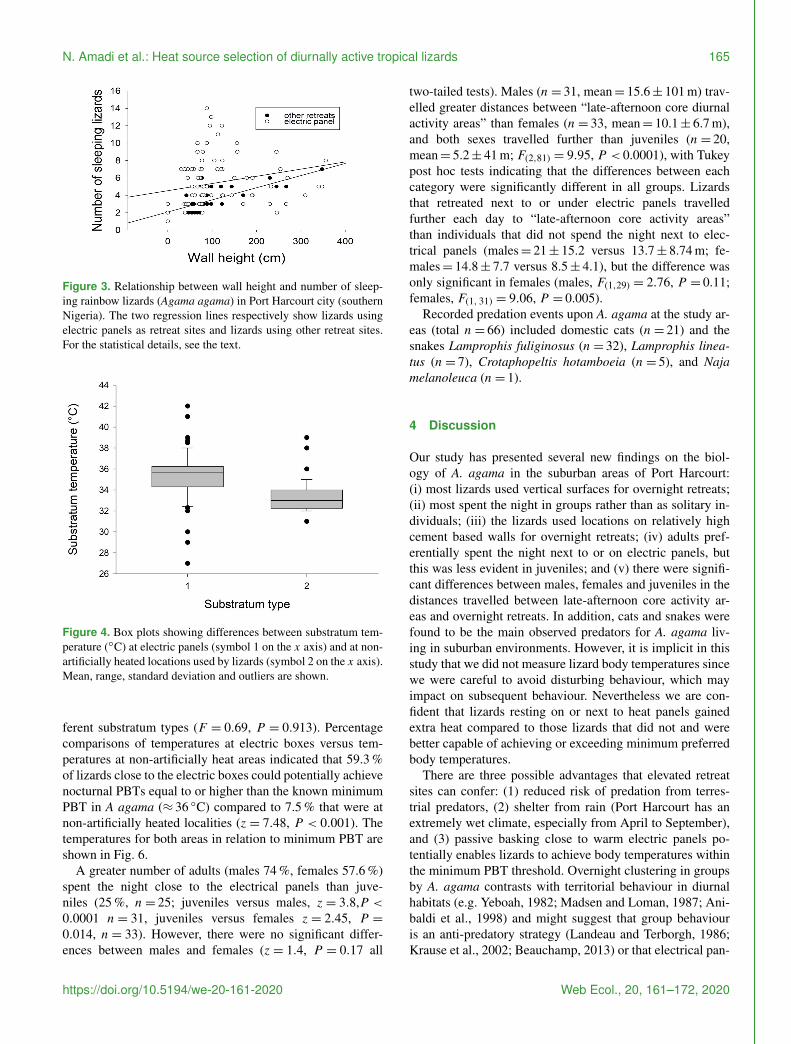
N Amadi et al Heat source selection of diurnally active tropical lizards 165
Figure 3 Relationship between wall height and number of sleep-ing rainbow lizards (Agama agama) in Port Harcourt city (southernNigeria) The two regression lines respectively show lizards usingelectric panels as retreat sites and lizards using other retreat sitesFor the statistical details see the text
Figure 4 Box plots showing differences between substratum tem-perature (C) at electric panels (symbol 1 on the x axis) and at non-artificially heated locations used by lizards (symbol 2 on the x axis)Mean range standard deviation and outliers are shown
ferent substratum types (F = 069 P = 0913) Percentagecomparisons of temperatures at electric boxes versus tem-peratures at non-artificially heat areas indicated that 593 of lizards close to the electric boxes could potentially achievenocturnal PBTs equal to or higher than the known minimumPBT in A agama (asymp 36 C) compared to 75 that were atnon-artificially heated localities (z= 748 P lt 0001) Thetemperatures for both areas in relation to minimum PBT areshown in Fig 6
A greater number of adults (males 74 females 576 )spent the night close to the electrical panels than juve-niles (25 n= 25 juveniles versus males z= 38P lt00001 n= 31 juveniles versus females z= 245 P =0014 n= 33) However there were no significant differ-ences between males and females (z= 14 P = 017 all
two-tailed tests) Males (n= 31 mean= 156plusmn 101 m) trav-elled greater distances between ldquolate-afternoon core diurnalactivity areasrdquo than females (n= 33 mean= 101plusmn 67 m)and both sexes travelled further than juveniles (n= 20mean= 52plusmn 41 m F(281) = 995 P lt 00001) with Tukeypost hoc tests indicating that the differences between eachcategory were significantly different in all groups Lizardsthat retreated next to or under electric panels travelledfurther each day to ldquolate-afternoon core activity areasrdquothan individuals that did not spend the night next to elec-trical panels (males= 21plusmn 152 versus 137plusmn 874 m fe-males= 148plusmn 77 versus 85plusmn 41) but the difference wasonly significant in females (males F(129) = 276 P = 011females F(1 31) = 906 P = 0005)
Recorded predation events upon A agama at the study ar-eas (total n= 66) included domestic cats (n= 21) and thesnakes Lamprophis fuliginosus (n= 32) Lamprophis linea-tus (n= 7) Crotaphopeltis hotamboeia (n= 5) and Najamelanoleuca (n= 1)
4 Discussion
Our study has presented several new findings on the biol-ogy of A agama in the suburban areas of Port Harcourt(i) most lizards used vertical surfaces for overnight retreats(ii) most spent the night in groups rather than as solitary in-dividuals (iii) the lizards used locations on relatively highcement based walls for overnight retreats (iv) adults pref-erentially spent the night next to or on electric panels butthis was less evident in juveniles and (v) there were signifi-cant differences between males females and juveniles in thedistances travelled between late-afternoon core activity ar-eas and overnight retreats In addition cats and snakes werefound to be the main observed predators for A agama liv-ing in suburban environments However it is implicit in thisstudy that we did not measure lizard body temperatures sincewe were careful to avoid disturbing behaviour which mayimpact on subsequent behaviour Nevertheless we are con-fident that lizards resting on or next to heat panels gainedextra heat compared to those lizards that did not and werebetter capable of achieving or exceeding minimum preferredbody temperatures
There are three possible advantages that elevated retreatsites can confer (1) reduced risk of predation from terres-trial predators (2) shelter from rain (Port Harcourt has anextremely wet climate especially from April to September)and (3) passive basking close to warm electric panels po-tentially enables lizards to achieve body temperatures withinthe minimum PBT threshold Overnight clustering in groupsby A agama contrasts with territorial behaviour in diurnalhabitats (eg Yeboah 1982 Madsen and Loman 1987 Ani-baldi et al 1998) and might suggest that group behaviouris an anti-predatory strategy (Landeau and Terborgh 1986Krause et al 2002 Beauchamp 2013) or that electrical pan-
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-
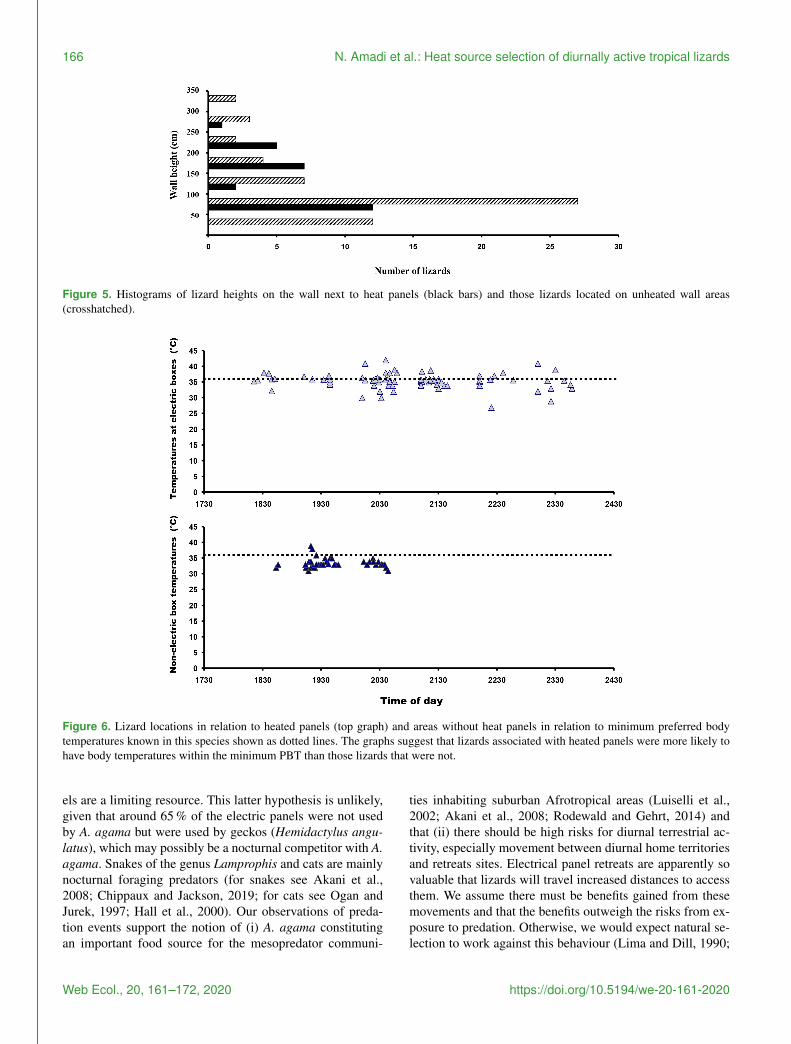
166 N Amadi et al Heat source selection of diurnally active tropical lizards
Figure 5 Histograms of lizard heights on the wall next to heat panels (black bars) and those lizards located on unheated wall areas(crosshatched)
Figure 6 Lizard locations in relation to heated panels (top graph) and areas without heat panels in relation to minimum preferred bodytemperatures known in this species shown as dotted lines The graphs suggest that lizards associated with heated panels were more likely tohave body temperatures within the minimum PBT than those lizards that were not
els are a limiting resource This latter hypothesis is unlikelygiven that around 65 of the electric panels were not usedby A agama but were used by geckos (Hemidactylus angu-latus) which may possibly be a nocturnal competitor with Aagama Snakes of the genus Lamprophis and cats are mainlynocturnal foraging predators (for snakes see Akani et al2008 Chippaux and Jackson 2019 for cats see Ogan andJurek 1997 Hall et al 2000) Our observations of preda-tion events support the notion of (i) A agama constitutingan important food source for the mesopredator communi-
ties inhabiting suburban Afrotropical areas (Luiselli et al2002 Akani et al 2008 Rodewald and Gehrt 2014) andthat (ii) there should be high risks for diurnal terrestrial ac-tivity especially movement between diurnal home territoriesand retreats sites Electrical panel retreats are apparently sovaluable that lizards will travel increased distances to accessthem We assume there must be benefits gained from thesemovements and that the benefits outweigh the risks from ex-posure to predation Otherwise we would expect natural se-lection to work against this behaviour (Lima and Dill 1990
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

N Amadi et al Heat source selection of diurnally active tropical lizards 167
Fahrig 2007) which it clearly does not However we do notknow when the lizards adopted this behaviour and whetherthere is a long-term selective advantage to retreat site se-lection Nevertheless gaining heat from warm overnight re-treat sites to elevate body temperatures must increase thecapacity for fast movement due to Q10 effects on bodytemperature (Bennett 1990) and even reduce early-morningbasking costs Increased movements and greater variation inmale movement may involve territoriality even in respect ofovernight retreat sites For example males of A agama arehighly territorial and males with higher body temperatureswould have an advantage in territory disputes due to Q10 ef-fects on muscular energy Distances travelled by males werehighly variable but significantly greater than travelled by fe-males and subadults This likely also involves territory de-fence since this necessitates movement (patrols) to detect thepresence of rival males and is well known in other species oflizards including agamids (eg Meek and Avery 2008)
The high percentage of female lizards associated with ar-tificial heat sources could relate to the importance of highand constant body temperatures in increasing offspring fit-ness and survivorship in reptiles (Shine and Harlow 1993Angilletta et al 2000 Lourdais et al 2004 Loumlwenborg etal 2011 Moor et al 2020) Laboratory research has shownthat elevated levels of basking opportunity and higher nighttemperatures during late summer in female gecko lizardsincreased the proportion of births occurring before winter(Moor et al 2020) Attaining nocturnally high body tem-peratures in natural environments especially when noctur-nal temperatures are lower than diurnal temperatures maybe impossible and hence A agama is exploiting one of theoutcomes of anthropogenic environmental change
Overnight retreats at elevated locations for example treebranches are not exceptional components in the habitats ofAfrican agamids and are known to reduce predation risk(Reaney and Whiting 2003) In central Nigeria A agamaspend the night in trees and shrubs and on buildings contrast-ing with terrestrial diurnal activity (Cloudsley-Thompson1981) Nocturnal selection for higher perches has also beenobserved in other tropical lizards including the chameleonBrookesia decaryi normally a diurnal ground dweller thatselects higher areas such as small plants shrubs dead fallenbranches or lianas as nocturnal retreat sites (Razafimahatra-tra et al 2008)
The selection of electric panels as overnight retreat sitesdemonstrates a degree of behavioural flexibility in A agamaincluding exploitation of the almost permanent ldquodeficiency inpublic electricity supplyrdquo in Port Harcourt city and in Nige-ria generally where there is widespread reliance on privategenerators instead of public electric power (Ado and Josiah2015 Nkoro et al 2019) Despite only a small proportion oflizards being found on trees or bushes to accurately measurethis would require an estimate of how many trees and busheswere available compared to wall area availability but for lo-gistic reasons this was impossible to measure in the fieldAlso it would be necessary to have an estimate of the like-lihood to detect a lizard in a tree as opposed to a wall inorder to make firm conclusions on the issue of resting sitepreferences Thus given the importance of understanding be-haviour and ecology of a highly adaptable species in new andchanging environments a more experimental approach willbe applied in a forthcoming phase of our research For ex-ample increased insight into the heat available to A agamain its natural environment compared to urban areas will en-able understanding of the comparative thermal constraints ineach habitat and their impact on the lives of the lizards andhow they adapt to urban areas Such knowledge will be a firststep in understanding the conservation measures required toprotect A agama should they be required at a future date
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-
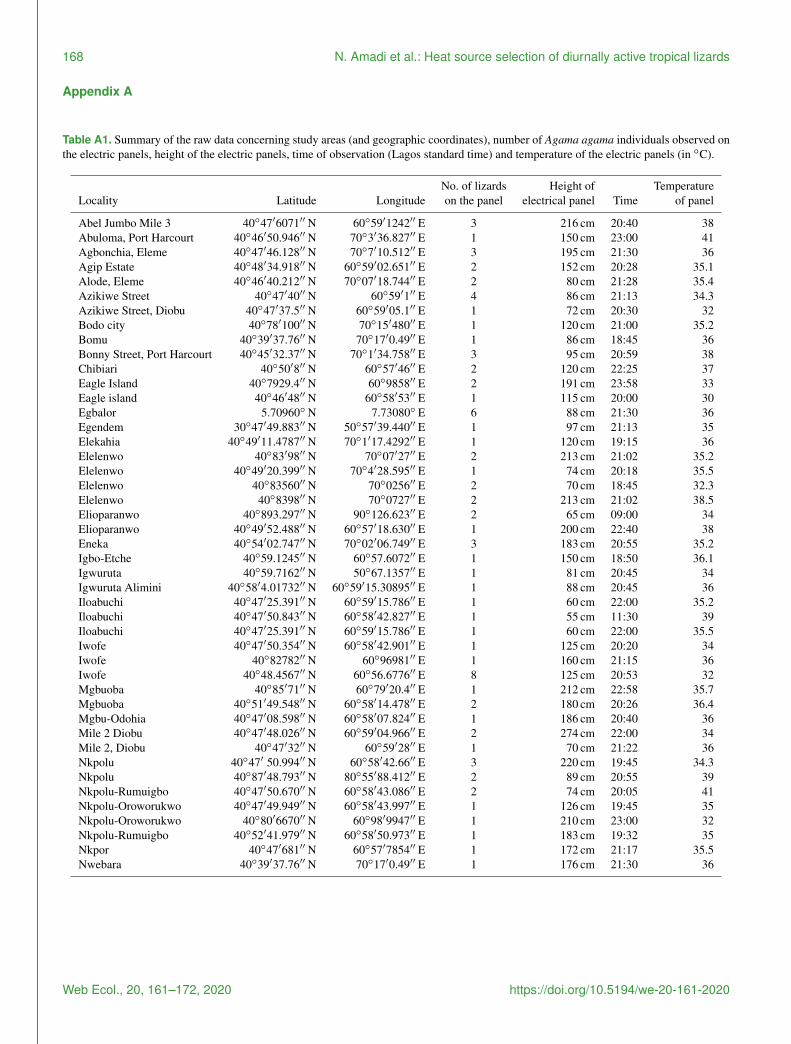
168 N Amadi et al Heat source selection of diurnally active tropical lizards
Appendix A
Table A1 Summary of the raw data concerning study areas (and geographic coordinates) number of Agama agama individuals observed onthe electric panels height of the electric panels time of observation (Lagos standard time) and temperature of the electric panels (in C)
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Abel Jumbo Mile 3 4047prime6071primeprime N 6059prime1242primeprime E 3 216 cm 2040 38Abuloma Port Harcourt 4046prime50946primeprime N 703prime36827primeprime E 1 150 cm 2300 41Agbonchia Eleme 4047prime46128primeprime N 707prime10512primeprime E 3 195 cm 2130 36Agip Estate 4048prime34918primeprime N 6059prime02651primeprime E 2 152 cm 2028 351Alode Eleme 4046prime40212primeprime N 7007prime18744primeprime E 2 80 cm 2128 354Azikiwe Street 4047prime40primeprime N 6059prime1primeprime E 4 86 cm 2113 343Azikiwe Street Diobu 4047prime375primeprime N 6059prime051primeprime E 1 72 cm 2030 32Bodo city 4078prime100primeprime N 7015prime480primeprime E 1 120 cm 2100 352Bomu 4039prime3776primeprime N 7017prime049primeprime E 1 86 cm 1845 36Bonny Street Port Harcourt 4045prime3237primeprime N 701prime34758primeprime E 3 95 cm 2059 38Chibiari 4050prime8primeprime N 6057prime46primeprime E 2 120 cm 2225 37Eagle Island 4079294primeprime N 609858primeprime E 2 191 cm 2358 33Eagle island 4046prime48primeprime N 6058prime53primeprime E 1 115 cm 2000 30Egbalor 570960 N 773080 E 6 88 cm 2130 36Egendem 3047prime49883primeprime N 5057prime39440primeprime E 1 97 cm 2113 35Elekahia 4049prime114787primeprime N 701prime174292primeprime E 1 120 cm 1915 36Elelenwo 4083prime98primeprime N 7007prime27primeprime E 2 213 cm 2102 352Elelenwo 4049prime20399primeprime N 704prime28595primeprime E 1 74 cm 2018 355Elelenwo 4083560primeprime N 700256primeprime E 2 70 cm 1845 323Elelenwo 408398primeprime N 700727primeprime E 2 213 cm 2102 385Elioparanwo 40893297primeprime N 90126623primeprime E 2 65 cm 0900 34Elioparanwo 4049prime52488primeprime N 6057prime18630primeprime E 1 200 cm 2240 38Eneka 4054prime02747primeprime N 7002prime06749primeprime E 3 183 cm 2055 352Igbo-Etche 40591245primeprime N 60576072primeprime E 1 150 cm 1850 361Igwuruta 40597162primeprime N 50671357primeprime E 1 81 cm 2045 34Igwuruta Alimini 4058prime401732primeprime N 6059prime1530895primeprime E 1 88 cm 2045 36Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 352Iloabuchi 4047prime50843primeprime N 6058prime42827primeprime E 1 55 cm 1130 39Iloabuchi 4047prime25391primeprime N 6059prime15786primeprime E 1 60 cm 2200 355Iwofe 4047prime50354primeprime N 6058prime42901primeprime E 1 125 cm 2020 34Iwofe 4082782primeprime N 6096981primeprime E 1 160 cm 2115 36Iwofe 40484567primeprime N 60566776primeprime E 8 125 cm 2053 32Mgbuoba 4085prime71primeprime N 6079prime204primeprime E 1 212 cm 2258 357Mgbuoba 4051prime49548primeprime N 6058prime14478primeprime E 2 180 cm 2026 364Mgbu-Odohia 4047prime08598primeprime N 6058prime07824primeprime E 1 186 cm 2040 36Mile 2 Diobu 4047prime48026primeprime N 6059prime04966primeprime E 2 274 cm 2200 34Mile 2 Diobu 4047prime32primeprime N 6059prime28primeprime E 1 70 cm 2122 36Nkpolu 4047prime 50994primeprime N 6058prime4266primeprime E 3 220 cm 1945 343Nkpolu 4087prime48793primeprime N 8055prime88412primeprime E 2 89 cm 2055 39Nkpolu-Rumuigbo 4047prime50670primeprime N 6058prime43086primeprime E 2 74 cm 2005 41Nkpolu-Oroworukwo 4047prime49949primeprime N 6058prime43997primeprime E 1 126 cm 1945 35Nkpolu-Oroworukwo 4080prime6670primeprime N 6098prime9947primeprime E 1 210 cm 2300 32Nkpolu-Rumuigbo 4052prime41979primeprime N 6058prime50973primeprime E 1 183 cm 1932 35Nkpor 4047prime681primeprime N 6057prime7854primeprime E 1 172 cm 2117 355Nwebara 4039prime3776primeprime N 7017prime049primeprime E 1 176 cm 2130 36
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-
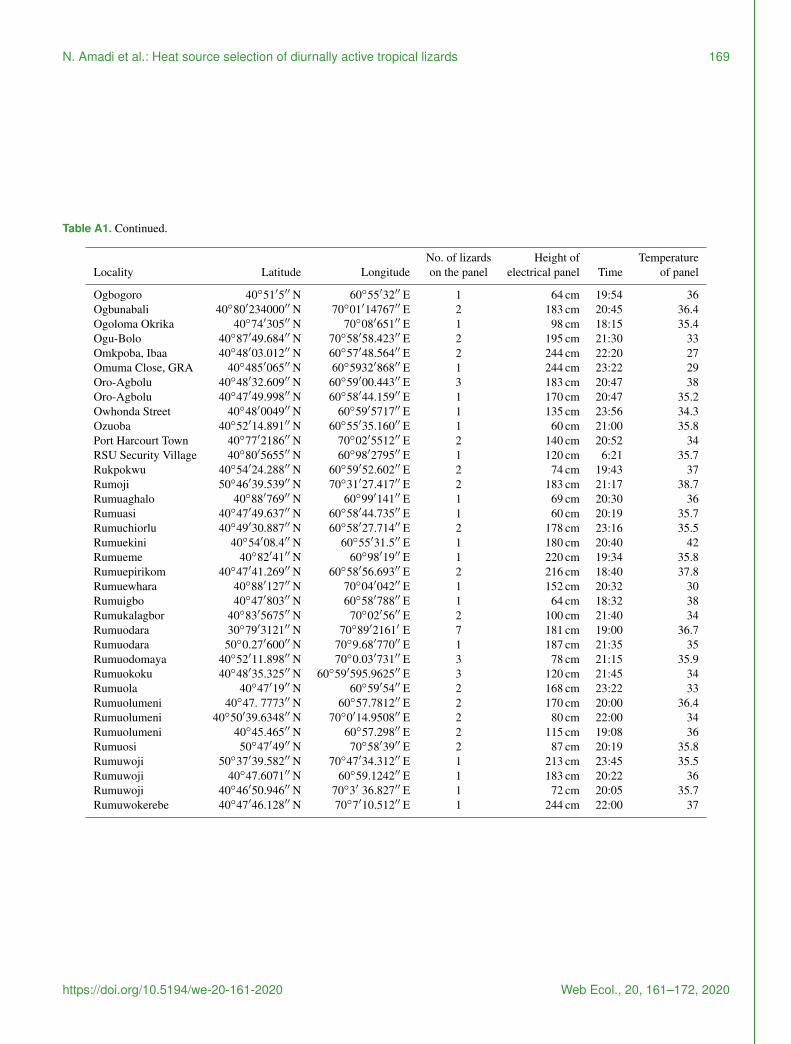
N Amadi et al Heat source selection of diurnally active tropical lizards 169
Table A1 Continued
No of lizards Height of TemperatureLocality Latitude Longitude on the panel electrical panel Time of panel
Ogbogoro 4051prime5primeprime N 6055prime32primeprime E 1 64 cm 1954 36Ogbunabali 4080prime234000primeprime N 7001prime14767primeprime E 2 183 cm 2045 364Ogoloma Okrika 4074prime305primeprime N 7008prime651primeprime E 1 98 cm 1815 354Ogu-Bolo 4087prime49684primeprime N 7058prime58423primeprime E 2 195 cm 2130 33Omkpoba Ibaa 4048prime03012primeprime N 6057prime48564primeprime E 2 244 cm 2220 27Omuma Close GRA 40485prime065primeprime N 605932prime868primeprime E 1 244 cm 2322 29Oro-Agbolu 4048prime32609primeprime N 6059prime00443primeprime E 3 183 cm 2047 38Oro-Agbolu 4047prime49998primeprime N 6058prime44159primeprime E 1 170 cm 2047 352Owhonda Street 4048prime0049primeprime N 6059prime5717primeprime E 1 135 cm 2356 343Ozuoba 4052prime14891primeprime N 6055prime35160primeprime E 1 60 cm 2100 358Port Harcourt Town 4077prime2186primeprime N 7002prime5512primeprime E 2 140 cm 2052 34RSU Security Village 4080prime5655primeprime N 6098prime2795primeprime E 1 120 cm 621 357Rukpokwu 4054prime24288primeprime N 6059prime52602primeprime E 2 74 cm 1943 37Rumoji 5046prime39539primeprime N 7031prime27417primeprime E 2 183 cm 2117 387Rumuaghalo 4088prime769primeprime N 6099prime141primeprime E 1 69 cm 2030 36Rumuasi 4047prime49637primeprime N 6058prime44735primeprime E 1 60 cm 2019 357Rumuchiorlu 4049prime30887primeprime N 6058prime27714primeprime E 2 178 cm 2316 355Rumuekini 4054prime084primeprime N 6055prime315primeprime E 1 180 cm 2040 42Rumueme 4082prime41primeprime N 6098prime19primeprime E 1 220 cm 1934 358Rumuepirikom 4047prime41269primeprime N 6058prime56693primeprime E 2 216 cm 1840 378Rumuewhara 4088prime127primeprime N 7004prime042primeprime E 1 152 cm 2032 30Rumuigbo 4047prime803primeprime N 6058prime788primeprime E 1 64 cm 1832 38Rumukalagbor 4083prime5675primeprime N 7002prime56primeprime E 2 100 cm 2140 34Rumuodara 3079prime3121primeprime N 7089prime2161prime E 7 181 cm 1900 367Rumuodara 50027prime600primeprime N 70968prime770primeprime E 1 187 cm 2135 35Rumuodomaya 4052prime11898primeprime N 70003prime731primeprime E 3 78 cm 2115 359Rumuokoku 4048prime35325primeprime N 6059prime5959625primeprime E 3 120 cm 2145 34Rumuola 4047prime19primeprime N 6059prime54primeprime E 2 168 cm 2322 33Rumuolumeni 4047 7773primeprime N 60577812primeprime E 2 170 cm 2000 364Rumuolumeni 4050prime396348primeprime N 700prime149508primeprime E 2 80 cm 2200 34Rumuolumeni 4045465primeprime N 6057298primeprime E 2 115 cm 1908 36Rumuosi 5047prime49primeprime N 7058prime39primeprime E 2 87 cm 2019 358Rumuwoji 5037prime39582primeprime N 7047prime34312primeprime E 1 213 cm 2345 355Rumuwoji 40476071primeprime N 60591242primeprime E 1 183 cm 2022 36Rumuwoji 4046prime50946primeprime N 703prime 36827primeprime E 1 72 cm 2005 357Rumuwokerebe 4047prime46128primeprime N 707prime10512primeprime E 1 244 cm 2200 37
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

170 N Amadi et al Heat source selection of diurnally active tropical lizards
Data availability Data will be provided upon request from the au-thors
Author contributions NA and LL conceived the study RM andLL carried out data analysis and wrote the manuscript includingpreparing figures and tables NA RB HOC DD and AC collectedthe data All authors read and agreed on the final version of themanuscript
Competing interests The authors declare that they have no con-flict of interest
Acknowledgements Our gratitude goes to the second-year gen-eral ecology students of the Departments of Animal and Environ-mental Biology Plant ScienceBiotechnology and Microbiology(Rivers State University of Science and Technology Port Harcourt)who were instrumental to the collection of some data while study-ing the habitat ecology of Agama lizards in Rivers State We thankthe two anonymous referees and the subject editor Sonja Knappfor helpful comments on the submitted drafts
Review statement This paper was edited by Sonja Knapp andreviewed by two anonymous referees
References
Adeoye G O and Ogunbanwo O O Helminth parasites of theAfrican lizard Agama agama (Squamata Agamidae) in LagosNigeria Rev Biol Trop 55 417ndash425 2007
Ado A and Josiah M M Impact of deficient electricity supply onthe operations of small scale businesses in North East Nigeria IJ BED 3 1ndash12 2015
Akani G Ebere N Peacuterez-Mellado V and Luiselli LStomach flushing affects survivalemigration in wildlizards a study case with rainbow lizards (Agamaagama) in Nigeria Amphibia-Reptilia 32 253ndash260httpsdoiorg101163017353711X565493 2011
Akani G C Luiselli L Eniang E A and RugieroL Life in the tropical suburbs Food type partition-ing among sympatric African house snakes of the genusLamprophis (Colubridae) Ital J Zool 75 395ndash399httpsdoiorg10108011250000801888037 2008
Akani G C Petrozzi F Rugiero L Segniagbeto G H andLuiselli L Effects of rainfall and geography on the compar-ative diets of eight rainbow lizard populations across TogoBenin and Nigeria (West Africa) Amphibia-Reptilia 34 185-192 httpsdoiorg10116315685381-00002881 2013
Akani G C Luiselli L U C A and Politano E Ecological andconservation considerations on the reptile fauna of the easternNiger Delta (Nigeria) Herpetozoa 11 141ndash153 1999
Amadi N A Herpetofauna Survey of Rivers State (Southern Nige-ria) Rivers State University Press Port Harcourt Rivers StateNigeria 63 pp 2017
Angilletta Jr M J Winters R S and Dunham AE Thermal effects on the energetics of lizard em-bryos implications for hatchling phenotypes Ecol-ogy 81 2957ndash2968 httpsdoiorg1018900012-9658(2000)081[2957TEOTEO]20CO2 2000
Anibaldi C Luiselli L and Angelici F Notes on the ecologyof a suburban population of rainbow lizards in coastal KenyaAfr J Ecol 36 199ndash206 httpsdoiorg101046j1365-2028199800119x 1998
Beauchamp G Social predation How Group Living BenefitsPredators and Prey Elsevier Paris 336 pp 2013
Bennett A F Thermal dependence of locomo-tor capacity Am J Physiol 250 253ndash258httpsdoiorg101152ajpregu19902592R253 1990
Bonnet X Naulleau G and Shine R The dangers of leavinghome dispersal and mortality in snakes Biol Conserv 89 39ndash50 httpsdoiorg101016S0006-3207(98)00140-2 1999
Cabrera-Guzmaacuten E and Reynoso V H Use of sleeping perchesby the lizard Anolis uniformis (Squamata Polychrotidae) in thefragmented tropical rainforest at Los Tuxtlas Mexico Rev MexBiodivers 81 921ndash924 2010
Campbell S S and Tobler I Animal sleep a review of sleep dura-tion across phylogeny Neurosci Biobehav R 8 269ndash300 1984httpsdoiorg1010160149-7634(84)90054-X
Chapman B M and Chapman R F Observations on the biologyof the lizard Agama agama in Ghana Proc Zool Soc Lond143 121ndash132 1964
Chippaux J-P and Jackson K Snakes of Central and WesternAfrica John Hopkins University Press Baltimore USA 448 pp2019
Clark D L and Gillingham J C Sleep-site fidelity intwo Puerto Rican lizards Anim Behav 39 1138ndash1148httpsdoiorg101016S0003-3472(05)80786-9 1990
Cloudsley-Thompson J L Bionomics of the rainbow lizardAgama agama (L) in eastern Nigeria during the dry seasonJ Arid Environ 4 235ndash245 httpsdoiorg101016S0140-1963(18)31565-9 1981
Cooper W E Risk factors affecting escape behaviour byPuerto Rican Anolis lizards Can J Zool 84 495ndash504httpsdoiorg101139z06-018 2006
Downes S and Shine R Heat safety or solitude Using habitat se-lection experiments to identify a lizards priorities Anim Behav55 1387ndash1396 httpsdoiorg101006anbe19970705 1998
Ellison G Wolfenden A Kahana L Kisingo A JamiesonJ Jones M and Bettridge C M Sleeping site selection inthe nocturnal northern lesser galago (Galago senegalensis) sup-ports antipredator and thermoregulatory hypotheses Int J Pri-matol 40 276ndash296 httpsdoiorg101007s10764-019-00085-y 2019
Enge K M Krysko K L and Talley B L Distribution andecology of the introduced African rainbow lizard Agama agamaafricana (Sauria Agamidae) Florida Sci 67 303ndash310 2004
Fahrig L Non-optimal animal movement in humanaltered landscapes Funct Ecol 21 1003ndash1015httpsdoiorg101111j1365-2435200701326x 2007
Hall L S Kasparian M A Van Vuren D and Kelt DA Spatial organization and habitat use of feral cats (Feliscatus L) in Mediterranean California Mammalia 64 19ndash28httpsdoiorg101515mamm200064119 2000
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

N Amadi et al Heat source selection of diurnally active tropical lizards 171
Harris V A The Life of the Rainbow Lizard Hutchinson TropicalMonographs Hutchinson and Company London 174 pp 1964
Hitchcock M A and McBrayer L D Thermoregulation in noc-turnal ecthotherms Seasonal and intraspecific variation in theMediterranean Gecko (Hemidactylus turcicus) J Herp 40 185ndash195 httpsdoiorg101670233-04A1 2006
Huey R B Temperature physiology and the ecology of reptilesin Biology of the Reptilia 12 Physiology C Physiological Ecol-ogy edited by Gans C and Pough F H Academic Press Lon-don 1982
James F C and Porter W C Behaviour- microclimatic relation-ships in the African rainbow lizard Agama agama Copeia 1979585ndash593 httpsdoiorg1023071443867 1979
Kearney M Hot rocks and much-too-hot rocks seasonal patternsof retreat site selection by a nocturnal ectotherm J Therm Biol27 205ndash218 httpsdoiorg101016S0306-4565(01)00085-72002
Krause J Ruxton G D Ruxton G D and Ruxton I G Livingin Groups Oxford University Press Oxford UK 228 pp 2002
Landeau L and Terborgh J Oddity and the ldquoconfu-sion effectrdquo in predation Anim Behav 34 1372ndash1380httpsdoiorg101016S0003-3472(86)80208-1 1986
Leache A D Chong R A Papenfuss T J Wagner P BohmeW Schmitz A Rodel M O LeBreton M Ineich I ChirioL Bauer A Eniang E A and Din S B E Phylogeny of thegenus Agama based on mitochondrial DNA sequence data BonnZool Beit 56 273ndash278 2009
Leacheacute A D Grummer J A Miller M Krishnan S Fu-jita M K Boumlhme W Schmitz A Lebreton M Ine-ich I Chirio L Ofori-boateng C Eniang E A Green-baum E Roumldel M-O and Wagner P Bayesian inferenceof species diffusion in the West African Agama agama speciesgroup (Reptilia Agamidae) System Biodivers 15 192ndash203httpsdoiorg1010801477200020161238018 2017
Lima S L and Dill L M Behavioural decisions made under therisk of predation a review and prospectus Can J Zool 68 619ndash640 httpsdoiorg1010801477200020161238018 1990
Lourdais O Shine R Bonnet X Guillon M and Naul-leau G Climate affects embryonic development in aviviparous snake Vipera aspis Oikos 104 551ndash560httpsdoiorg101111j0030-1299200412961x 2004
Loumlwenborg K Shine R and Hagman M Fitness disadvantagesto disrupted embryogenesis impose selection against suboptimalnest-site choice by female grass snakes Natrix natrix (Colubri-dae) J Evol Biol 24 177ndash183 httpsdoiorg101111j1420-9101201002153x 2011
Luiselli L and Amori G Diet chap 8 in ReptileEcology and Conservation A Handbook of Tech-niques Oxford University Press Oxford 97ndash109httpsdoiorg101093acprofoso978019872613500300082016
Luiselli L Angelici F M and Akani G C Comparative feed-ing strategies and dietary plasticity of the sympatric cobras Najamelanoleuca and Naja nigricollis in three diverging Afrotropicalhabitats Can J Zool 80 55ndash63 httpsdoiorg101139z01-178 2002
Madsen T and Loman J On the role of colour dis-play in the social and spatial organization of male rain-
bow lizards (Agama agama) Amphibia-Reptilia 8 365ndash372httpsdoiorg101163156853887X00135 1987
Meek R Retreat site characteristics and body temperatures ofSaurodactylus brosseti Bons and Pasteur 1957 (Sauria Sphaero-dactylidae) Morocc Bull Soc Herp Fr 128 41ndash48 2008
Meek R Repeated use of roadside tunnels of the European mole(Talpa europea) as a communal nesting area by grass snakes Na-trix natrix are there thermal benefits Herp Bull 139 16ndash192017
Meek R and Avery R A Basking in the Australian wa-ter dragon Physignathus lesueurii why do alpha malesnot respond to operative temperatures in the same wayas adults and sub-adults Amphibia-Reptilia 29 257ndash262httpsdoiorg101163156853808784125009 2008
Moor G Penniket S and Cree A Greater basking opportu-nity and warmer nights during late pregnancy advance modalbirth season in a live-bearing gecko lowering the risk of re-duced embryonic condition Biol J Linn Soc 130 128ndash141httpsdoiorg101093biolinneanblaa017 2020
Morris C L and Lattanzio M S Intraspecific variation in treelizard escape behaviour in relation to habitat and temperatureBehaviour 157 185ndash204 httpsdoiorg1011631568539X-00003586 2020
Nkoro E Ikue-John N Okeke W Amabuike I L and Ajaba JA Power supply and the performances of small and mediumscale enterprises (SMSEs) in Rivers State Int J BusinessEcosyst Strat 1 55ndash62 2019
Ogan C V and Jurek R M Biology and ecology of feral free-roaming and stray cats in Mesocarnivores of northern Cali-fornia biology management and survey techniques workshopmanual edited by Harris J E and Ogan C V The WildlifeSociety Humboldt State University 87ndash92 1997
Pauwels O S G Gramentz D and Toham A K Agama agama(Rock-headed Agama) nocturnal activity Herp Rev 35 164ndash165 2004
Poche Jr A J Powell R and Henderson R W Sleep-site selec-tion and fidelity in Grenadian Anoles Sleep 30 31ndash39 2005
Porter W P Mitchell J W Beckman W A and De-Witt C B Behavioral implications of mechanisticecology Thermal and behavioral modeling of desert ec-totherms and their microenvironment Oecologia 13 1ndash54httpsdoiorg101007BF00379617 1973
Rasoloharijaona S Rakotosamimanana B Randrianambinina Band Zimmermann E Pair-specific usage of sleeping sites andtheir implications for social organization in a nocturnal Mala-gasy primate the Milne Edwardsrsquo sportive lemur (Lepilemur ed-wardsi) Am J Phys Anthropol 122 251ndash258 2003
Razafimahatratra B Mori A and Hasegawa M Sleep-ing site pattern and sleeping behavior of Brookesia decaryi(Chamaeleonidae) in Ampijoroa dry forest northwestern Mada-gascar Current Herp 27 93ndash99 httpsdoiorg1031051881-1019-27293 2008
Reaney L T and Whiting M J Picking a tree habitat use by thetree agama Acanthocercus atricollis atricollis in South AfricaAfric Zool 38 273ndash278 2003
Rial R V Akaacircrir M Gamundiacute A Nicolau C Garau CAparicio S and Coenen A M Evolution of wakefulness sleepand hibernation from reptiles to mammals Neurosci BiobehavR 34 1144ndash1160 2010
httpsdoiorg105194we-20-161-2020 Web Ecol 20 161ndash172 2020
172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-

172 N Amadi et al Heat source selection of diurnally active tropical lizards
Rodewald A D and Gehrt S D Wildlife population dynamicsin urban landscapes in Urban wildlife conservation SpringerBoston MA 117ndash147 2014
Shine R and Harlow P Maternal thermoregulation influences off-spring viability in a viviparous lizard Oecologia 96 122ndash1271993
Singhal S and Johnson M The behavioral ecology of sleep natu-ral sleeping site choice in three Anolis lizard species Behaviour144 1033ndash1052 2007
Schlesinger C A and Shine R Selection of diurnal retreat sites bythe nocturnal gekkonid lizard Oedura lesueurii Herpetologica50 156ndash163 1994
Wagner P Glaw F Glaw K and Boumlhme W Studies on AfricanAgama IV First record of Agama agama (Sauria Agamidae)from Madagascar and identity of the alien population on GrandeComore Island Herp Notes 2 73ndash77 2009
Webb J K Pringle R M and Shine R How do nocturnal snakesselect diurnal retreat sites Copeia 4 919ndash925 2004
Yeboah S Observations on territory of the rainbowlizard Agama agama Afric J Ecol 20 187ndash192httpsdoiorg101111j1365-20281982tb00289x 1982
Yorzinski J L and Platt M L The difference between nightand day antipredator behavior in birds J Ethol 30 211ndash218httpsdoiorg101007s10164-011-0318-5 2012
Web Ecol 20 161ndash172 2020 httpsdoiorg105194we-20-161-2020
- Abstract
- Introduction
- Materials and methods
-
- Study area
- Protocol
- Statistical analyses
-
- Results
- Discussion
- Appendix A
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- Review statement
- References
-