leukotrienes are indicated as mediators of hyperoxia ... · of hyperoxia-inhibited alveolarization...
TRANSCRIPT

Leukotrienes are indicated as mediators of hyperoxia-inhibited alveolarization in newborn rats
VERONIKA BOROS,lp4 JACQUELINE S. BURGHARDT,2 CATHERINE J. MORGAN,2 AND DAVID M. OLSON1-3 Departments of lpediatrics, 2Physiology, and 30bstetrics and Gynaecology, Perinatal Research Centre, The University of Alberta, Edmonton, Alberta, Canada T6G 2S2; and 4Department of Pediatrics, Szent Janos Hospital, Budapest, Hungary H-1125
Boros, Veronika, Jacqueline S. Burghardt, Cather- ine J. Morgan, and David M. Olson. Leukotrienes are indicated as mediators of hyperoxia-inhibited alveolarization in newborn rats. Am. J. Physiol. 272 (Lung Cell. Mol. PhysioE. 16): L433-L441, 1997.-We investigated the role of leukotri- enes (LT) in hyperoxia-induced changes in lung parenchyma in neonatal rat pups. Rat pups were exposed to 21% 02 (air) or >95% O2 from days 4 to 14 after birth and were administered the 5lipoxygenase (&LO) inhibitor and LTD4 receptor antago- nist Wy-50295, 5-LO-activating protein inhibitor MK-0591, or vehicle from days 3 to 14. All measurements were done on days 12-14. There was a significant (P < 0.05) increase in peptido-LT output from lung slices of animals exposed to 02 compared with air-exposed animals. Both WY-50295 and MK-0591 significantly lowered (P < 0.05) peptido-LT output in 02-exposed animals. The 6-ketoprostaglandin F1, output was increased similarly in both vehicle- and drug-treated 02-exposed animals. O2 exposure also caused a significant increase in bronchoalveolar lavage fluid protein and extravas- cular lung water that could not be ameliorated by WY-50295 or MK-0591. Hyperoxia-induced inhibition of alveolarization, indicated by a significantly (P < 0.05) lower parenchymal tissue density, specific internal surface area, and airspace perimeter-to-area ratio, and a significantly (P < 0.05) higher mean linear intercept and airspace unit volume than air- exposed animals, was prevented by both WY-50295 and MK-0591. Although hyperoxia had no effect on septal thick- ness, WY-50295 caused significant thickening in both air- and 02-exposed pups. Our studies provide evidence that hyperoxia- induced peptido-LT may mediate Oz-induced inhibition of alveolarization and that this is not caused by an arachidonic acid shunt to cyclooxygenase.
oxygen toxicity; Wy-50295; MK-0591; bronchopulmonary dys- plasia; morphometry
HYPEROXIA IS ONE of the major pathogenic factors of neonatal lung disease, causing endothelial and epithe- lial cell damage. Histologically, the lung, in diseases such as bronchopulmonary dysplasia (BPD), is com- posed of large simplified alveoli. Margraf et al. (21) documented a severe reduction in alveolar number, an increased mean linear intercept (i.e., an estimate of alveolar diameter), and a reduced internal surface area (ISA) in morphometric studies of humans with fatal chronic BPD. Impaired lung development has also been associated with prolonged hyperoxic exposure in neona- tal rats, especially during the period of rapid alveolar development that occurs in normal rats from days 4 to 14 after birth (2,3, 12).
The mechanism by which O2 interferes with alveolar development is unknown. Although O2 radicals directly injure the neonatal lung, damage and impaired healing
may be potentiated by the release of inflammatory mediators. Lipid mediators such as leukotrienes (LT) may be involved. Elevated LT concentrations have been found in the bronchoalveolar lavage (BAL) fluid of infants with BPD (14,32). In a previous study, we found that exposure to >95% O2 from days 4 to 14 elevated peptido-LT levels in the lungs of neonatal rats (4). In the same study, we found that administration of the 5lipoxygenase (&LO) inhibitor and LTD4 receptor an- tagonist WY-50295 (Wyeth Ayerst) lowered lung pep- tido-LT levels and prevented hyperoxia-induced airway smooth muscle thickening and hyperreactivity in 4-wk- old rat pups (4).
We reasoned that if elevated LT caused by hyperoxic exposure altered airway morphometry and function at 4 wk, then hyperoxia-induced LT might be expected to alter alveolar development by 2 wk of age. Therefore, the objective of this study was to examine the effect of LT inhibition on hyperoxia-induced changes in lung parenchymal structure in neonatal rat pups. However, because WY-50295 is both a 5-LO inhibitor and an LTD4 receptor antagonist plus a platelet-activating factor (PAF) synthesis inhibitor (19), we compared its effects at 14 days to MK-0591 (Merck-Frosst, Dorval, PQ, Canada), for which the only known action is to inhibit 5-LO activating protein (1,9,25).
METHODS
Animals
Sprague-Dawley albino rat pups (Charles River Laborato- ries) of both sexes were used. They were housed in the Health Sciences Laboratory Animal Service of the University of Alberta under veterinary care. The guidelines of the Cana- dian Council of Animal Care were followed in all experimen- tal procedures. Dams were maintained on regular laboratory rodent pellets and water ad libitum and kept on a 12:12-h light-dark cycle.
The experimental design was separate for each drug. Hence, each drug had its own vehicle control groups for both air and 02 exposures. For each drug, four experimental groups were developed: 21% 02 + vehicle (RA + V), RA + drug, >95% 02 + vehicle (02 + V), and 02 + drug.
O2 Exposure
Parallel litters of randomly divided rat pups and their dams were placed into 0.14-m3 Plexiglas exposure chambers containing >95% or 21% 02 (room air) from days 4 to 14 of life. 02 concentrations were monitored daily (Ventronic oxy- gen analyzer no. 5517). 02 and air were filtered through barium hydroxide lime (Baralyme; Chemetron Medical Divi- sion, St. Louis, MO), to keep COZ under 0.5%, and through
1040-0605/97 $5.00 Copyright o 1997 the American Physiological Society L433

L434 LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION
charcoal. Temperature and humidity were maintained at 26°C and 75-80%, respectively. Chambers were opened for 15 min daily to switch dams between O2 and air environments, change dirty cages, and administer the drugs.
LT Inhibition
Wy-50295, MK-0591, or vehicle (Hz0 plus Tween 80 in a 4:1 ratio) was administered subcutaneously to rat pups from days 3 to 14 of life. Wy-50295 was injected at a dose of 30 mg/kg daily. This dose of Wy-50295 was selected because preliminary trials proved it to effectively inhibit peptido-LT output from parenchymal lung slices in vitro when adminis- tered subcutaneously. Tissue levels of the drug were not determined, but this concentration (if evenly distributed without metabolism or excretion) would inhibit both 5-LO and PAF acetyltransferase activities (19).
MK-0591 was injected at a dose of 20 mg/kg daily. Prelimi- nary results using daily injections of 10 mg/kg resulted in only 3% inhibition of lung slice peptido-LT output in air- exposed pups and 22% inhibition in 02-exposed pups (data not shown). Administration of 20 mg/kg resulted in 71% inhibition of lung slice peptido-LT output in air-exposed pups and 75% inhibition in Oz-exposed pups (see Fig. 1).
Eicosanoid Production
Lung eicosanoid production was measured using a modi- fied short-term lung explant technique (15). Briefly, pups were killed with an intraperitoneal overdose of pentobarbital sodium (100 mg/kg Euthanyl; MIC Pharmaceuticals, Cam- bridge, ON, Canada). A catheter (Insyte-W, 24 gauge, 1.9 cm; Becton-Dickinson Vascular Access, Sandy, UT) was intro- duced into the right ventricle of the heart, and the vascula- ture was washed with 5 ml of ice-cold saline. The lungs were removed, and the left lobe was transversely sliced (tissue slicer; Stoelting, Wood Dale, IL) to a thickness of 500 urn. Three to four lung slices were placed into a tissue culture well (Costar, Cambridge, MA) containing 800 ul of Hanks’ bal- anced salt solution with 1.67 mM CaC12 and buffered with 15 mM N-2-hydroxyethylpiperazine-N’-2-ethanesulfonic acid to pH 7.36 and were incubated for 30 min at 37°C. LT released by the lung slices into the medium were assayed by enzyme- linked immunosorbent assay. Peptido-LT were assayed using a LTC,&TDd/LTE4 enzyme-linked immunoassay kit (Oxford Biomedical Research, Oxford, MI). Typical results yield 50% of the sample or standard bound/medium bound (B/Be) at 1.93 rig/ml and an 80% B/Be at 0.83 rig/ml. The specificity of the kits used were 100% for LTC4, >80% for LTD4 and LTE4, ~2% for LT&, and < 1% for LTB4. The 6-ketoprostaglan- din F1, (6-keto-PGFi,) production was quantified by radio- immunoassay. 6-Keto-[5,8,9,11,12,14,15(n)-3H]PGF1,, was ob- tained from Amersham. The 6-keto-PGFi, antibody (Cayman Chemical) was used at a 1:15,000 dilution. The total activity of 6-keto-[“H]PGF1, was 7,000-10,000 cpm, and the concen- trations of standard (Cayman Chemical) used were 3.8-1,000 pg/ml. All eicosanoid levels were normalized by the total DNA [assay modified from Downs and Wilfinger (lo)] contained in the lung tissue.
Measurement of Acute Lung Damage
Extravascular lung water (ELW) was assessed in 12-day- old rats by calculating wet-to-dry weight ratios. Animals were killed and exsanguinated. Then lungs were removed from the chest cavity and blotted minimally, and the wet weights were determined. The lungs were then allowed to dry at 37°C. The tissue was considered dry when the weight was constant for 2 consecutive days, and dry weight was then recorded.
Total protein in BAL fluid was measured on day 13. Physiological saline (1 ml) was instilled into the trachea via a polyethylene cannula [inner diameter (ID) 0.86 mm, outer diameter (OD) 1.27 mm, Intramedic; Becton-Dickinson, Par- sippany, NJ] and then recovered. This was repeated two additional times, and all recovered fluid was combined. Samples were used only if >80% instilled saline was recov- ered. The BAL fluid was then centrifuged for 5 min at 200 g. Total protein in the supernatant was measured using a Bradford (Bio-Rad Laboratories, Hercules, CA) microassay using bovine serum albumin (Sigma Chemical, St. Louis, MO) as the standard.
Lung Morphometry
Lung preparation. Lungs of 14-day-old rat pups were fixed in situ through a polyethylene tracheal cannula (0.86 mm ID X 1.27 mm OD, Intramedic; Becton-Dickinson) with 2.5% glutaraldehyde (Sigma Chemical) at a constant pressure of 20 cmH20 for 2 h. Then the trachea was ligated, and the lungs were excised and immersed in glutaraldehyde for 24 h. Lung volumes were measured by water displacement (31) before and after the 24-h fixation period to detect shrinkage. Be- cause of minimal shrinkage (O-2%), data did not have to be corrected.
After fixation, transverse sections of superior, middle, and inferior portions of the right lung and superior and inferior sections of left lung were embedded in paraffin. The entire transverse sections were cut 3 urn thick and stained with Gomori-trichromealdehyde. Slides were initially examined to eliminate sections with evidence of inadequate preparation.
Morphometric measurements. Light level morphometric assessment of lung parenchymal tissue was performed in a blinded fashion on coded slides from six 14-day-old rats from each experimental group. Five 3-day-old rats were examined. Eight to ten randomly selected fields were examined from each lung. Histological specimens observed with the micro- scope (Carl Zeiss Jenamed Variant) were put into a gray image analyzer system (IS Tech) via a video camera (MT1 S 68). The measurements and calculations were performed using Genias 25 Image Analysis software (Joyce-Loebl). All measurements were made on images magnified X 140.
Parenchymal tissue includes alveolar septa, alveolar ducts, respiratory bronchiolar tissues, and blood vessels with a diameter (10 urn and their contents. Volume density of parenchymal tissue (V,) was calculated as [field area (FA) - airspace area (A,,)]/FA X 100 from each analyzed field. Mean septal thickness (Th,,,,) was calculated from parenchymal tissue area and length of gas exchange surface.
As an indication of mean alveolar diameter, the mean linear chord length (L,) was calculated by dividing the length of the computer-generated horizontal test line by the number of intercepts of the septal wall (23). Mean volume of airspace units (Vasp unit ) was computed as (Li x n)/3 (33). To detect the structural changes of alveolar airspaces, the perimeter-to- area ratio (P/A) of the airspace was calculated from each field.
ISA of the lung available for respiratory exchange was calculated using the formula (4 X lung volume)/L, (36). ISA data were normalized to 100 g body wt and used as specific ISA.
Statistical Analysis
Experimental data were analyzed for significant differ- ences by one-factor analysis of variance. Duncan’s New Multiple Range post hoc test was used to determine differ- ences between groups when a significant F-value was ob-

LEUKOTRIENES AND HYPEROXIA-INHIBITED AIVEOLARIZATION L435
tained. Significance was achieved at P 5 0.05. Data are expressed as means k SD.
RESULTS
Eicosanoid Production
LT production by lung slices over 30 min is shown in Fig. 1. Peptido-LT output was significantly (P < 0.05) elevated in both O2 + V groups after the lo-day O2 exposure (Fig. 1, A and B) compared with their respec- tive controls. As Fig. 1A shows, WY-50295 significantly (P < 0.05) inhibited O2 -induced LT output. Peptido-LT
3.0 A 1 1
2.5
2.0 1 1.5 1 a
AIR OXYGEN
1.81 B
1.6
1.0 -
0.8 -
0.6 -
0.4 -
b
I, C
b
a
I n=9 n=3
OXYGEN
output from air-exposed Wy-50295-treated animals de- creased; however, it was not significantly different from RA + V animals. Figure 1B shows that MK-0591 inhibited peptido-LT production from both air and 02-exposed animals; however, peptido-LT levels in the RA + MK-0591 group were not statistically signifi- cantly different from the RA + V group. Peptido-LT output from O2 + V animals was significantly (P < 0.01) higher than that of the three other groups, including O2 + MK-0591 (Fig. 1B).
The 6-keto-PGF1, output, a stable metabolite of PG12 (prostacyclin), was measured to detect a possible arachi- donic acid shunt to the cyclooxygenase pathway be- cause of 5-LO inhibition. We detected significantly (P < 0.05) increased 6-keto-PGF1, output from all four OZ- exposed groups compared with their respective air- exposed controls (Fig. 2, A and B). The 6-keto-PGF1, production was neither elevated nor diminished be- cause of WY-50295 or MK-0591 administration, indicat- ing that neither shunting of arachidonic acid to, nor inhibition of, the cyclooxygenase pathway occurred.
Fig. 1. Effect of >95% 02 exposure, WY-50295 (30 mg/kg daily, A), and MK-0591 (20 mg/kg daily, 23) on lung tissue peptido-leukotriene (LT) output of 14-day-old rat pups. Open bars, vehicle; filled bars, WY-50295 or MK-0591.02 exposure significantly (P < 0.05) increased peptido-LT production. WY-50295 and MK-0591 were able to reduce peptido-LT output significantly (P < 0.05) from Oz-exposed animals only Data are expressed as means + SD. Different letters indicate statistical differences (P < 0.05) between groups.
Subjective observations under the light microscope suggested that 02-exposed animals had fewer and enlarged alveoli and alveolar ducts compared with air-exposed controls (Fig. 5, B and C). Their architec- ture was similar to 3-day-old animals (Fig. 5A). RA + WY-50295 lungs showed increased tissue mass, and the alveolar septa appeared thick and hypercellular (Fig. SD). O2 + WY-50295 animals had lung architecture similar to air-treated animals. They had an increased amount of parenchymal tissue and more and smaller air-space units compared with O2 + V pups. The alveolar septae appeared to be markedly thicker (Fig. 5E). RA + MK-0591 were indistinguishable from RA + I
Acute Lung Injury
Body weight and lung size of the eight groups are shown in Table 1. There were no significant differences among the groups in body weight or lung volume at the end of the experimental period. Body weights and lung volumes were smaller for all four groups in the MK- 0591 experiment compared with the animals in the WY-50295 experiment. This is because of the fact that litter sizes were kept larger for a longer period of time during the MK-0591 experiment, i.e., fewer animals were removed on days 12 and 13 for wet-to-dry weight determinations or BAL, making the litters larger at day 14.
The ELW, as indicated by lung tissue wet-to-dry weight ratios, was significantly elevated in both O2 + V and OZ + drug groups (P < 0.01) compared with the air-exposed groups (Fig. 3, A and B). Neither WY-50295 nor MK-0591 diminished lung edema caused by O2 exposure. ELW was similar in the RA + V and RA + drug lungs. In a trend similar to that of ELW, BAL fluid protein levels (Fig. 4, A and B) were increased in all 02-exposed groups compared with air-exposed groups (P < 0.01). Neith er d rug diminished protein levels.
Morphometry

L436 LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION
1~ A
25 - B
20-
15-
lo-
5-
0 -
a a
a a
I s- n=4 n=4
AIR
b
T b
OXYGEN
b
T b
1T
OXYGEN
6.0
1 A
Fig. 2. Effect of >95% 02 exposure, Wy-50295 (30 mg/kg daily, A), and MK-0591(20 mg/kg daily, B) on lung tissue output of 14-day-old rat pups. Open bars, vehicle; filled bars, Wy-50295 or MK-0591. 02 exposure significantly (P < 0.05) stimulated 6-ketoprostaglandin Fi, (6-keto-PGFi,) production, but Wy-50295 or MK-0591 did not affect it. Data are expressed as means + SD. Different letters indicate statistical differences (P < 0.05) between groups.
V lungs (Fig. 6, A and C). O2 + MK-0591 animals had a markedly improved structure compared with O2 + V animals without any changes in septal thickness (Fig. 6,BandD).
Table 1. Rat body weights and lung volumes at 14 days of age
WY-50295 MK-0591
Body Lung Body Lung n weight, g volume, ml n weight, g volume, ml
Air+Vehicle 6 29.6k3.0 1.562088 7 26.223.4 1.38kO.13 Air + Drug 6 25.7k1.9 1.5520.09 7 26.320.9 1.36kO.11 Oz+Vehicle 6 28924.5 1.5220.12 7 27.323.8 1.4320.16 02 + Drug 6 28.422.9 1.5320.96 8 26.121.9 1.3OkO.18
Values are means + SD. There are no significant differences among groups within each experiment.
a a
CONTROL
b b
HYPEROXIC
b b
a
a
CONTROL HY PEROXIC
Fig. 3. Effect of >95% 02 exposure, Wy-50295 (30 mg/kg daily, A), and MK-0591 (20 mg/kg daily, B) on wet-to-dry lung weight ratios of 14-day-old rat pups. Open bars, vehicle; filled bars, Wy-50295 or MK-0591. The extravascular lung water content was significantly (P < 0.01) in creased in all Oa-exposed groups. Neither Wy-50295 nor MK-0591 altered wet-to-dry lung ratios. Data are expressed as means + SD. Different letters indicate statistical differences (P < 0.05) between groups.
The comparative lung morphometric findings for Wy-50295treated and control rat pups before O2 ex- posure and after 10 days of hyperoxia are shown in Table 2. The VP of O2 + V pups was significantly lower compared with both air-treated groups and the Oz + Wy-50295-treated group. Furthermore, the parenchy- ma1 tissue volume density showed a significant eleva- tion in RA + WY-50295 pups compared with RA + V pups. No difference was noted in Th,,,t among 02- and air-exposed pups. In contrast, both WY-50295 groups had increased septal wall thickness compared with vehicle-treated controls. The L, was increased in the O2 + V group at day 14. The alveolar diameter of the O2 + V pups was similar to the day 3 values. Mean- while, the O2 + WY-50295 animals had an alveolar diameter close to the air-exposed animals. The Vaspunit of O2 + V animals was similar to that of 3-day-old

LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION L437
a a
CONTROL HYPEROXIC
a a
b
b
T b
T
CONTROL HYPEROXIC
Fig. 4. Effect of >95% 02 exposure, WY-50295 (30 mg/kg daily, A), and MK-0591(20 mg/kg daily, B) on protein content of bronchoalveo- lar lavage (BAL) fluid of 14-day-old rat pups. Open bars, vehicle; filled bars, WY-50295 or MK-0591.02-exposed pups had significantly (P < 0.01) hi g h er protein levels. Neither WY-50295 nor MK-0591 altered the protein content. Data are expressed as means 2 SD. Different letters indicate statistical differences (P < 0.05) between groups.
animals, whereas the airspace volume of RA + V and RA + WY-50295 was significantly smaller. WY-50295 had no effect on airspace volume in air exposed animals compared with its relative vehicle controls; however, the airspace volume was significantly lower in the O2 + WY-50295 group compared with the O2 + V group. The airspace perimeter-to-area ratio (P/A,,,) followed a similar pattern to that of Vaspunit. The ratio was signifi- cantly smaller in the O2 + V group, much like the values of the 3-day-old animals. The O2 + WY-50295 ratio was larger than the OZ + V group and similar to both air groups. A decrease was observed in specific ISA development of O2 + V animals, which was not ob- served in the O2 + WY-50295 group. There was no difference in alveolar diameter or specific ISA between the 14-day-old air-treated groups.
Table 3 shows the morphometric measurements for MK-0591 and vehicle-treated rats pups. O2 exposure in this experiment caused a significantly (P < 0.05) lower V, in O2 + V pups compared with RA + V, RA + MK-0591, and 02 + MK-0591. MK-0591 caused no effects on tissue density in air-exposed animals. As with Wy-50295, no difference was noted in Thsept among 02- and air-exposed pups. Unlike Wy-50295, MK-0591 caused no increases in Thsept. L, and Vaspunit were increased in the O2 + V group, whereas the values in the O2 + MK-0591 group were similar to air-exposed groups. PlA,,, followed a similar pattern, with O2 exposure causing a significant decrease that was prevented by MK-0591. The specific ISA was significantly lower in the O2 + V group. Again, MK-0591 treatment was able to prevent this decrease.
DISCUSSION
Hyperoxic exposure upsets the normal cellular oxi- dant-antioxidant defense equilibrium by producing marked increases in O2 free radical production (38). Prolonged exposure of newborn and adult rats to high concentrations of OZ causes lung damage characterized by interstitial and intra-alveolar edema, followed by infiltration of protein, entry of cells, and finally hemor- rhage into the alveolar space (4, 6). Inhibition of the process of alveolarization is also a hallmark of hyper- oxic exposure during the newborn period (3,22,37).
Hyperoxia is one of several potential mechanisms that play a role in the O2 effect on lung growth and maturation. It slows lung maturation and causes a reduction in alveolar number and surface area by permanently inhibiting the process of septation (22). Saccule septation is permanently diminished and the parenchymal airspace is enlarged with irregular dilata- tion of alveoli and alveolar ducts (35). This exists until young adulthood of rats (30). Decreased cell prolifera- tion is a well-known concomitant of hyperoxic exposure in the newborn lung. Because the process of septation undoubtedly involves a very coordinated proliferation of specific lung cells, an inhibitor of DNA synthesis such as hyperoxia can easily disrupt this developmental process of the lung (13). Exposing rat pups to 100% O2 from days 4 to 13 also affects the connective tissue of the lung, altering elastin fiber structure and concentra- tion (2), suggesting another potential mechanism by which hyperoxia may interfere with alveolarization.
In our newborn rat model, 10 days of exposure to >95% O2 caused marked interstitial and intra-alveolar edema and proteinosis. Altered alveolarization oc- curred in hyperoxic animals, which corresponds to the results of investigators cited above. Thus we have a good model of hyperoxic lung damage that has allowed us to identify potential mediators involved.
Several investigators have suggested that metabo- lites of arachidonic acid, produced by inflammatory cells or by cells lining the alveoli, may be involved in hyperoxic lung damage. LTB4 is a well-known chemoat- tractant for neutrophils, and the activated inflamma- tory cells can release many other radicals and enzymes that may damage normal tissue as well as kill invading

LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION
Fig. 5. Photomicrographs of lung parenchyma. A: 3-day-old rats. B: 14-day-old rats, 21% 0s (RA) + vehicle. C: 14-day-old rats, 0s + vehicle. D: 14-day-old rats, RA + Wy-50295. E: 14-day-old rats, 0s + WY-50295 The parenchymal architecture of Os-exposed pups is simpler (fewer and enlarged air-space units; C). The alveolar septae of RA + Wy-50295treated animals are bulkier; note the apparent hypercellularity (D). Pulmonary structure of 0s + WY-50295 pups contains more and smaller alveoli compared with the 02 vehicle lung. Note again the increased septal thickness (El. Bar, 100 pm
microorganisms (34). In addition, it is known that LTC4 that was inhibited with lipoxygenase inhibitors. Simi- and LTD4 increase vascular permeability, and it has larly, increased LTD4 and LTB4 synthesis was shown in been shown that LT can provoke pressor responses and isolated rabbit lung infused with tertiary butyl perox- edema in perfused rat lungs (7, 18). Several investiga- ide. The accompanied increased vascular permeability tors have detected increased LT levels due to oxidative was ameliorated by the LT antagonist FPL-55712 (16). stress. Burghuber et al. (5) demonstrated increased Taniguchi et al. (34) found increased LTB4 levels in the 5hydroxyeicosatetraenoic acid production and edema BAL fluid of adult rats exposed to 85% O2 for 60 h. of perfused isolated rat lung caused by glucose oxidase Lipoxygenase inhibited animals had reduced mortality.
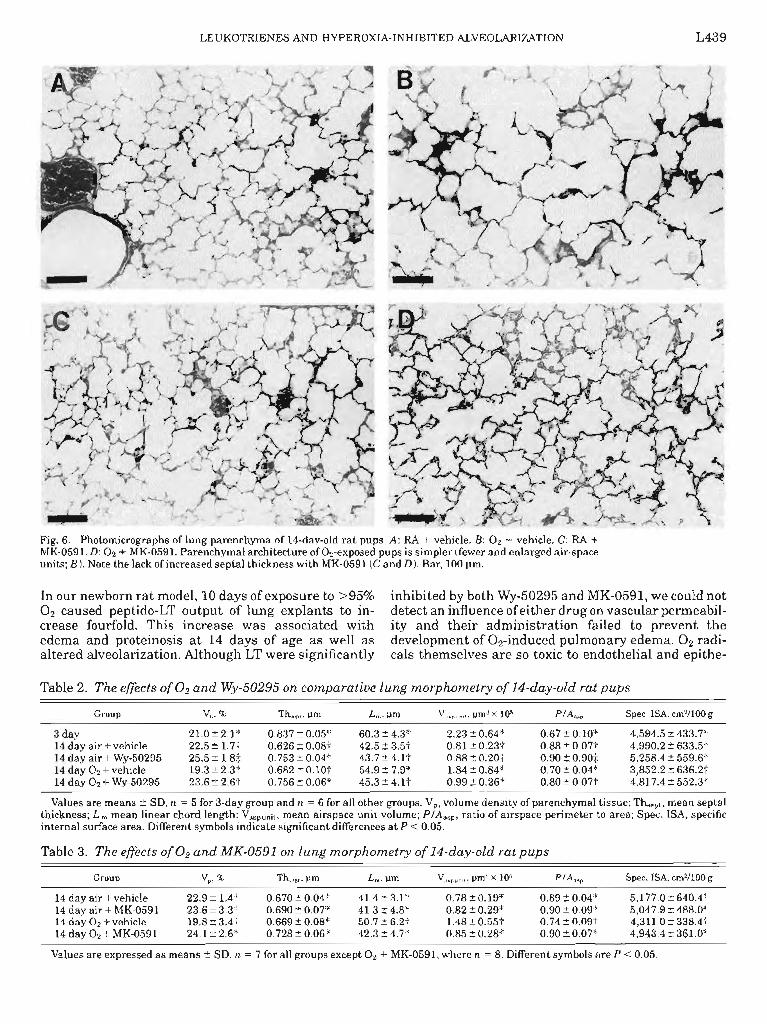
LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION L439
Fig. 6. Photomicrographs of lung parenchyma of 14-day-old rat pups. A: RA + vehicle. B: 02 + vehicle. C: RA + MK-0591. D: 02 + MK-0591. Parenchymal architecture of Oz-exposed pups is simpler (fewer and enlarged air-space units; B). Note the lack of increased septal thickness with MK-0591 (C and D). Bar, 100 pm.
In our newborn rat model, 10 days of exposure to >95% inhibited by both WY-50295 and MK-0591, we could not O2 caused peptido-LT output of lung explants to in- detect an influence of either drug on vascular permeabil- crease fourfold. This increase was associated with ity and their administration failed to prevent the edema and proteinosis at 14 days of age as well as development of 02-induced pulmonary edema. O2 radi- altered alveolarization. Although LT were significantly cals themselves are so toxic to endothelial and epithe-
Table 2. The effects of O2 and WY-50295 on comparative lung morphometry of 14-day-old rat pups
Group V", 5% Lw ?Jm V usounit. Pm3 x 10” Pf&, Spec. ISA, cd/100 g
3 day 14 day air + vehicle 14 day air + WY-50295 14 day 02 + vehicle 14 day 02 + WY-50295
21.0 t 2.1” 0.837*0.05* 60.3 24.3" 2.23~h0.64~ 0.67 t O.lO* 4,594.5?433.7" 22.5-fl.7: 0.626-CO.08-i 42.523.5.t 0.81 -c 0.23: 0.88+0.07f 4,990.2?633.5? 25.5?1.8$ 0.75310.04* 43.724.lf 0.88+0.20t 0.90 2 0.90; 5,258.4%559.6* 19.3 f 2.3” 0.682t O.lOt 54.9 -t 7.9” 1.84 i 0.84” 0.7010.04* 3,852.2?636.2? 23.612.6t 0.756t0.06" 45.3t4.11 0.99+0.26* 0.80t0.07t 4,817.4?552.3"
Values are means -C SD, n = 5 for 3-day group and n = 6 for all other groups. V,, volume density of parenchymal tissue; Th,,rt, mean septal thickness; L ,,, mean linear chord length; Vaspunit, mean airspace unit volume; P/A,,,,, ratio of airspace perimeter to area; Spec. ISA, specific internal surface area. Different symbols indicate significant differences at P < 0.05.
Table 3. The effects of O2 and MK-0591 on lung morphometry of 14-day-old rat pups
Group v,, YfY Thsc,,, pm L Pm v,,,,,;,, pm” x 105 PJ&, Spec. ISA, cm*/100 g
14 day air + vehicle 22.9? 1.4”; 0.670t0.04" 41.423.1" 0.78 t 0.19* 0.89 t 0.04” 5J77.0t640.4" 14 day air + MK-0591 23.6k3.3" 0.690?0.07* 41.3+-4.8* 0.82-cO.29" 0.90 t 0.09" 5,047.9%-488.0* 14 day 02 + vehicle 19.8 i- 3.4t 0.669 2 0.08” 50.7k6.2.t 1.48?0.55+ 0.74Ir0.09t 4,311.0-+338.4+ 14 day 02 + MK-0591 24.1 t 2.6” 0.728t0.06" 42.3t4.7": 0.85?0.28* 0.90t0.07" 4,943.4-c361.0"
Values are expressed as means + SD. n = 7 for all groups except O2 + MK-0591, where n = 8. Different symbols are P < 0.05.

L440 LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION
lial barriers that cellular mediators may play little or no role. It is also possible that an LT effect would have been evident only for the first few days of O2 exposure.
Up to this time, no one has examined whether LTs play a role in altered lung development because of hyperoxic exposure during the critical period of alveolar- ization. We used two LT synthesis inhibitors that had separate modes of action to study the possible LT effects. In our study comparing the O2 + WY-50295 group with the OZ + V group, it was evident that the WY-50295 group had a more advanced septation pro- cess, indicating that LT inhibition has a beneficial effect. WY-50295 also caused a significant increase in septum thickness in both the O2 + WY-50295 and RA + WY-50295 groups. The bulkiest septae were detected in the O2 + WY-50295 group as a summarized effect of O2 exposure and drug administration. The same trend was evident with parenchymal tissue density. The effect of WY-50295 on air-exposed animals, despite nonsignifi- cant inhibition of peptido-LT, suggests that WY-50295 had an action on the lungs that was not related to 5-LO inhibition. This effect may account for the prevention of damage in 02-exposed animals rather than the actual inhibition of LT during the exposure.
WY-50295 also inhibits PAF production by rat perito- neal mast cells (19), and PAF has many actions that mimic those of peptido-LT (including edema, mucus production, bronchoconstriction, and hyperresponsive- ness). Whether hyperoxia increases PAF levels is uncer- tain, because prolonged exposure of adult guinea pig lungs to hyperbaric O2 causes an increase in lyso-PAF, an inactive metabolite of PAF, while not affecting the levels of PAF itself (28). Furthermore, WY-50295 is 12-fold more effective as an LT inhibitor than a PAF synthesis inhibitor (19, 26), and LT output from lung tissue correlates with the changes we have seen in lung development. However, because WY-50295 might have been inhibiting PAF production or have other unknown effects on the lung, further experiments using a more specific LT synthesis inhibitor were necessary.
In our study, MK-0591 was able to prevent the damage to alveolarization caused by O2 exposure. Essen- tially, the Og + MK-0591 group was indistinguishable from both RA groups, including RA + MK-0591. MK- 0591 caused no detectable changes in the RA group, despite its ability to inhibit the production of pep- tido-LT in these animals. The evidence suggests that any protection offered by MK-0591 to the 02-exposed group is most likely because of the inhibition of pepti- do-LT. We are unaware of any nonspecific actions of MK-0591, and none have been published to our knowl- edge.
Little is known about the role of mediators in the process of 02-induced altered alveolarization or even what controls normal alveolar development. The effect of WY-50295 in air-exposed rats might suggest a nonspe- cific effect of this drug on normal lung development. However, this effect is most likely not due to the inhibition of LT because of the lack of effect of MK-0591 in air animals. Recently, Massaro and Massaro (24) provided evidence that the treatment of newborn rats
with retinoic acid influences postnatal alveolarization. Retinoic acid administration increases the number of alveoli in newborn animals and prevents the low alveo- lar number and low specific ISA produced by dexametha- sone. Data from different in vitro studies are controver- sial; both 12-LO and 5-LO pathway metabolites can stimulate and inhibit cell proliferation (8, 20, 29). Clinical studies have suggested that the frequency of the occurrence of BPD increases after indomethacin is given during pregnancy (11). Recently, Nagai et al. (27) concluded that normal alveolar formation in rats from days 4 to 14 depends on the presence of prostaglandins (PG) since animals administered indomethacin had altered alveolar development. Also, animals receiving PG and indomethacin at the same time showed fewer abnormalities. However, they did not measure the LT production of the indomethacin-administered group; therefore the possibility of an arachidonic acid shunt to the 5-LO pathway was not excluded. We did not detect an elevation of 6-keto-PGF1, production at day 14 in the WY-50295 or MK-0591-treated animals compared with vehicle animals. We have detected high 6-keto- PGFi, levels in all hyperoxic groups due to O2 exposure. Hageman et al. (17) found a similar effect in mature rabbits after OZ exposure. Hence, it is unlikely that the effects observed in the OZ- or drug-treated groups on alveolar development could be caused by altered PG levels.
Our studies provide evidence that chronic LT inhibi- tion prevents 02-induced inhibition of the formation of alveoli. These results suggest that chronically elevated LT play a role as mediators of 02-induced changes in lung morphometry in newborn rats, and therefore may play a role in the pathogenesis of diseases such as BPD. Whether LT act alone or in concert with other media- tors such as growth factors remains to be discovered.
We acknowledge the technical assistance of Damyanti Bhardwaj and Anna Radomski. We also thank Wyeth Ayerst for providing WY-50295 and Merck-Frosst for providing MK-0591.
Salary and/or project support came from the Alberta Heritage Foundation for Medical Research, the Medical Research Council of Canada, the Natural Sciences and Engineering Research Council of Canada, the Molly Towel1 Perinatal Research Foundation, and the Alberta Lung Association.
Address for reprint requests: J. S. Burghardt, The Perinatal Research Centre, 220 Heritage Medical Research Centre, The Univ. of Alberta, Edmonton, Alberta, Canada T6G 2S2.
Received 2 May 1996; accepted in final form 30 October 1996.
REFERENCES
Briedeau, C., C. Chan, S. Charleson, D. Denis, J. F. Evans, A. W. Ford-Hutchinson, J. W. Gillard, J. Guay, and D. Guevermont. Pharmacology of MK-0591 (3- [ 1-(4-chloroben- zyl)-3-(t-butylthiol-5-(quinolin-2yl-methoxy)-indol-2-yl]-2,2-di- methylpropanoic acid) a potent, orally active leukotriene biosyn- thesis inhibitor. Can. J. Physiol. Pharmacol. 70: 799-807, 1992. Bruce, M. C., R. Pavlowski, and CJ. F. Tomashefski. Changes in lung elastic fiber structure and concentration associated with hyperoxia in the developing rat lung. Am. Reu. Respir. Dis. 140: 1067-1074,1989. Bucher, J. R., and R. J. Roberts. The development of the newborn rat lung in hyperoxia: a dose-response study of lung growth, maturation and changes in antioxidant enzyme activ- ites. Pediatr. Res. 15: 999-1008,198l.

LEUKOTRIENES AND HYPEROXIA-INHIBITED ALVEOLARIZATION L441
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
Burghardt, J. S., V. Boros, D. F. Biggs, and D. M. Olson. Lipid mediators in oxygen-induced airway remodeling and hyper- responsiveness in newborn rats. Am. J. Respir. Crit. Care Med. 154: 837-842,1996. Burghuber, 0. C., R. J. Strife, J. Zirolli, P. M. Henson, M. M. Mathias, J. T. Reevers, R. C. Murphy, and N. F. Voelkel. Leukotriene inhibitors attenuate rat lung injury induced by hydrogen peroxide. Am. Rev. Respir. Dis. 131: 778-785,1985. Clark, J. M., and C. J. Lambersten. Pulmonary oxygen toxicity Pharmacol. Rev. 23: 37-133, 1971. Dahlen, S. E., J. Bjork, P. Hedqvist, K. E. Arforst, S. Hammarstrom, J. A. Lindgren, and B. Samuelsson. Leukotrienes promote plasma leakage and leukocyte adhesion in post capillary venules: in vivo effects with relevance to the acute inflammatory response. Proc. NatZ. Acad. Sci. USA 78: 3883- 3891,198l. Dethlefsen, S. M., D. Shepro, and P. A. D’Amore. Arachidonic acid metabolites in bFGF-, PDGF- and serum-stimulated vascu- lar cell growth. Exp. CeZZ Res. 212: 263-273,1994. Diamant, Z., M. C. Timmers, H. VanderVeen, B. S. Fried- man, M. DeSmet, M. Dupre, D. Hilliard, E. Bel, and P. J. Stark. The effect of MK-0591, a novel 5-lipoxygenase activating protein inhibitor, on leukotriene biosynthesis and allergen- induced airway responses in asthmatic subjects in vivo. J. Allergy Clin. Immunol. 95: 42-51, 1995. Downs, T. R., and W. W. Wilfinger. Fluorometric quantification of DNA in cells and tissue. Anal. Biochem. 131: 538-547,1983. Eronen, M., E. Pesonen, T. Kurki, K. Teramo, OYlikorkala, and M. Hallman. Increased incidence of bronchopulmonary dysplasia after antenatal administration of indomethacin to prevent preterm labor. J. Pediatr. 124: 782-788,1994. Frank, L. Oxygen toxicity in neonatal rats: the effect of endo- toxin treatment on survival during and post-02 exposure. Pedi- atr. Res. 21: 109-115,1987. Frank, L., and G. E. McLaughlin. Protection against acute and chronic hyperoxic inhibition of neonatal rat lung develop- ment with the 21-aminosteroid drug, U74389F. Pediatr. Res. 33: 623-638,1993. Groneck, P., B. Goetze-Speer, M. Opperman, H. Effert, and C. P. Speer. Association of pulmonary inflammation and in- creased microvascular permeability during the development of bronchopulmonary dysplasia: a sequential analysis of inflamma- tory mediators in respiratory fluids of high-risk preterm neo- nates. Pediatrics 93: 712-718, 1994. Gross, L., G. Walker, W. Smith, M. Maniscalo, M. Czajka, C. Wilson, and S. Rooney. An organ culture model for studying biochemical development of fetal rat lung. J. AppZ. Physiol. 75: 355-360,1978. Gurtner, G. H., I. S. Farrukh, N. F. Adkinson, A. M. Sciuto, J. M. Jacobson, and J. R. Michale. The role of arachidonate mediators in peroxide-induced lung injury. Am. Rev. Respir. Dis. 136: 480-483,1987. Hageman, J. R., J. Zemaitis, R. B. Holtzman, S. E. Smith, and C. E. Hunt. Failure of non-selective inhibition of arachi- donic acid metabolism to ameliorate hyperoxic lung injury. Prost. Leuko. Essent. Fatty Acids 32: 145-153,1988. Hedqvist, P., S. E. Dahlen, L. Gustafsen, S. Hammarstrom, and B. Samuelsson. Biological profile of leukotrienes Cd and Dd. Acta Physiol. Stand. 110: 331-333, 1980. Hogaboam, C. M., D. Donigi-Gale, T. S. Shoupe, E. Y. Bissonnette, A. D. Befus, and J. L. Wallace. Platelet- activating factor synthesis by peritoneal mast cells and its inhibition by two quinoline-based compounds. Br. J. Pharmacol. 105: 87-92,1992. Leikauff, G. D., H. E. Claesson, C.A. Doupnik, S. Hybinette, and R. C. Grafstrom. Cvsteinvl leukotrienes enhance growth of
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
human airway epithelial cells. Am. J. Physiol. 259 (Lung CeZZ. MOL. Biol. 3): L255-L261, 1990. Margraf, L. R., J. F. Tomashefski, Jr., M. C. Bruce, and B. B. Dahms. Morphometric analysis of the lung in bronchopulmo- nary dysplasia.Am. Rev. Respir. Dis. 143: 391-400,199l. Massaro, D., N. Teich, S. Maxwell, G. Massaro, and P. Whitney. Postnatal development of alveoli: regulation and evidence for a critical period in rats. J. CZin. Invest. 76: 1297- 1305,1985. Massaro, G. D., 0. Joyce, D. Colleen, and D. Massaro. Postnatal development of lung alveoli: suppression by 13% 02 and a critical period. Am. J. Physiol. 258 (Lung CelZ. Mol. PhysioZ. 2): L321-L327, 1990. Massaro, G. D., and D. Massaro. Postnatal treatment with retinoic acid increases the number of pulmonary alveoli in rats. Am. J. Physiol. 270 (Lung CelZ. Mol. Physiol. 14): L305-L310, 1996. Menard, L., M. Laviolette, and P. Borgeat. Studies of the inhibitory activity of MK-0591 (3-[1-(4-chlorobenzyl)-3-(t-butyl- thiol-5-(quinolin-2yl-methoxy)-indol-2-yl] -2,2-dimethylpropa- noic acid) on arachidonic acid metabolism in human phagocytes. Can. J. Physiol. Pharmacol. 70: 808-813, 1992. Musser, J. H., and A. F. Krefi. 5-lipoxygenase: properties, pharmacology, and the quinolinyl (bridged) aryl class of inhibi- tors. J. Med. Chem. 35: 2501-2524,1992. Nagai, A., M. Katayama, W. M. Thurlbeck, R. Matsui, S. Yasui, and K. Konno. Effect of indomethacin on lung development in postnatal rats: possible role of prostaglandins in alveolar formation. Am. J. Physiol. 268 (Lung Cell. Mol. Physiol. 12): L56-L62,1995. Parente, L., M. Fitzgerald, G. DeNucci, and S. Moncada. The release of lyso-PAF from guinea-pig lungs. Eur. J. Pharma- col. 112: 281-284,1985. Porreca, E., C. Di Febbo, M. Reale, R. Barbacane, A. Mezzetti, F. Cuccrullo, and P. Conti. Modulation of rat vascular smooth muscle cell (VSMC) proliferation by cysteinyl leukotrienes Dd: a role for mediation of interleukin 1. Atheroscle- rosis 113: 11-18, 1995. Randell, S. H., R. R. Mercer, and S. L. Young. Neonatal hyperoxia alters the pulmonary alveolar and capillary structure of 40 day old rats. Am. J. PathoZ. 136: 1259-1266,199O. Scherle, W. F. A simple method for volumetry of organs in quantitative stereology. Mikroscopie 26: 57-60,197O. Stenmark, K. R., M. Eyzaguire, J.Y. Westcott, P. M. Henson, and R. C. Murphy. Potential role of eicosanoids and PAF in the pathophysiology of bronchopulmonary dysplasia. Am. Rev. Respir. Dis. 136: 770-772,1987. Suen, H. C., K. D. Bloch, and P. K. Donahoe. Antenatal glucocorticoid corrects pulmonary immaturity in experimentally- induced congenitally diaphragmatic hernia in rats. Pediatr. Res. 35: 523-529,1994. Taniguchi, H., F. Taki, K. Takachi, T. Satake, S. Sugiyama, and T. Ozawa. The role of LTBA in the genesis of oxygen toxicity in the lung. Am. Rev. Respir. Dis. 133: 805-808,1986. Thibeault, D. W., S. Mabry, and M. Rezaiekhaligh. Neonatal pulmonary oxygen toxicity in the rat lung changes with aging. Pediatr. Pulmonol. 9: 96-108,199O. Weibel, E. R. Morphometry of the Human Lung. Berlin: Springer, 1963, p. 151. Yam, J., L. Frank, and R. J. Roberts. Oxygen toxicity: comparison of lung biochemical responses in neonatal and adult rats. Pediatr. Res. 12: 115-119, 1978. Zweir, J. L. Electron paramagnetic response evidence that cellular toxicity is caused by the generation of superoxide and hydroxyl free radical. FEBS Lett. 252: 12-16, 1989.