lentiviral transduction of apoai into hematopoietic...
TRANSCRIPT

Lentiviral Transduction of ApoAI Into HematopoieticProgenitor Cells and Macrophages
Applications to Cell Therapy of Atherosclerosis
Yan Ru Su, John L. Blakemore, Youmin Zhang, MacRae F. Linton, Sergio Fazio
Objective—We used genetically engineered mouse hematopoietic progenitor cells (HPCs) to investigate the therapeuticeffects of human apoAI on atherosclerosis in apoE�/� mice.
Methods and Results—Lentiviral constructs expressing either human apoAI (LV-apoAI) or green fluorescent protein(LV-GFP) cDNA under a macrophage specific promoter (CD68) were generated and used for ex vivo transduction of mouseHPCs and macrophages. The transduction efficiency was �25% for HPCs and �70% for macrophages. ApoAI was foundin the macrophage culture media, mostly associated with the HDL fraction. Interestingly, a significant increase in mRNA andprotein levels for ATP binding cassette A1 (ABCA1) and ABCG1 were found in apoAI-expressing macrophages after acLDLloading. Expression of apoAI significantly increased cholesterol efflux in wild-type and apoE�/� macrophages. HPCstransduced with LV-apoAI ex vivo and then transplanted into apoE�/� mice caused a 50% reduction in atherosclerotic lesionarea compared to GFP controls, without influencing plasma HDL-C levels.
Conclusions—Lentiviral transduction of apoAI into HPCs reduces atherosclerosis in apoE�/� mice. Expression of apoAIin macrophages improves cholesterol trafficking in wild-type apoE-producing macrophages and causes upregulation ofABCA1 and ABCG1. These novel observations set the stage for a cell therapy approach to atherosclerosis regression,exploiting the cooperation between apoE and apoAI to maximize cholesterol exit from the plaque. (Arterioscler ThrombVasc Biol. 2008;28:1439-1446)
Key Words: macrophages � hematopoietic progenitor cells � lentiviral vector � apolipoprotein AI� cholesterol efflux � cell therapy of atherosclerosis
Monocytes and macrophages play a crucial role in all stagesof atherosclerosis.1,2 Among their many functions, regu-
lation of intracellular cholesterol homeostasis is an essentialdeterminant of foam cell transformation and downstream events,including inflammatory changes and apoptosis. In addition tointernalization of modified lipoproteins via scavenger receptors,macrophages have developed sophisticated mechanisms to en-sure prompt cholesterol exit into the HDL pathway, also knownas reverse cholesterol transport (RCT).
At least 3 pathways are involved in cholesterol export frommacrophages: the ATP binding cassette A1 (ABCA1) path-way, which exports cholesterol and phospholipids to lipid-free apoAI; the ABCG1 pathway, which mediates cholesterolefflux to larger HDL particles; and the scavenger receptorclass B type I (SRB-I) pathway, which promotes bidirectionalcholesterol exchange between cells and lipoproteins, includ-ing HDL.3,4 Recent studies have suggested that ABCA1 andABCG1 work synergistically in this process. ABCA1 carriesout the initial transfer of free cholesterol and phospholipids tolipid-poor apoAI to generate nascent pre-� HDL particles,
which are then used as acceptors of cholesterol exported byABCG1.5,6
Lipid poor apoAI is not synthesized by macrophages.Thus, activation of RCT in the artery wall depends on theavailability of apoAI that has infiltrated from the plasmacompartment. Because of the obstacles to free diffusionpresented by the growing atheroma, the concentration ofapoAI in the artery wall may be reduced below a physiolog-ical threshold required for cholesterol extraction, possiblyleading to impaired cholesterol exit and expansion of thefoam cell lesion. We previously used a retroviral approach totransduce human apoAI into bone marrow cells, and showedsignificant reduction of atherosclerotic lesions in recipientapoE�/� mice without changes in HDL levels.7–9 However,apoAI expression was very low because of the inefficiency ofretroviral transduction of quiescent bone marrow cells. Thus,the effect of apoAI on cholesterol efflux was only seen inapoE�/� cells, given the dominant position of locally pro-duced apoE in regulating cholesterol homeostasis in theatheroma. This drawback obviously reduced the therapeuticimplications of our earlier studies.
Original received November 30, 2007; final version accepted April 29, 2008.From the Department of Medicine, Division of Cardiovascular Medicine, Atherosclerosis Research Unit (Y.R.S., J.L.B., Y.Z., M.F.L., S.F.), the
Department of Pharmacology (M.F.L.), and the Department of Pathology (S.F.), Vanderbilt University Medical Center, Nashville, Tenn.Correspondence to Yan Ru Su, or MacRae F. Linton, or Sergio Fazio Vanderbilt University Medical Center, Division of Cardiovascular Medicine, 383
PRB, 2220 Pierce Avenue, Nashville TN 37232-6300. E-mail [email protected], or [email protected], or [email protected]© 2008 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.107.160093
1439
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
ay 30, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

Hematopoietic progenitor cells (HPCs) possess 2 charac-teristics that make them ideal targets for gene transfer:self-renewal capacity and pluripotentiality.10 Using HPCsinstead of total bone marrow cells as the delivery vehicleincreases transduction efficiency and specificity. The humanimmunodeficiency virus type 1 (HIV-1)–based lentiviralvectors are capable of transducing both dividing and nondi-viding cells and have become the tool of choice because oflong-term stable gene transfer both in vitro and in vivo.11
We have generated a lentiviral vector-based human apoAIcDNA expression construct (LV-apoAI) driven by a macro-phage specific promoter, CD68. With this construct, we havetransduced HPCs ex vivo for transplantation into apoE�/�
mice and demonstrated its value as a cell-mediated genetherapy for atherosclerosis. We also show that expression ofapoAI increases cholesterol efflux in macrophages also ex-pressing apoE, suggesting that cooperation between these 2apolipoproteins can be exploited for cell-based approaches toplaque regulation.
Methods
Generation of a Lentiviral Human apoAI cDNAExpression ConstructThe self-inactivating (SIN) lentiviral vector (pWPT-WRPE) was agift from Dr Trono (University of Geneva). The human CD68promoter along with the first intron12 were amplified from humangenomic DNA by polymerase chain reaction (PCR). The humanapoAI cDNA with growth hormone (GH) poly-A signal sequencewas excised from an SRA-hapoAI clone previously generated in ourlaboratory.8 The final apoAI-producing lentiviral construct wasverified via DNA sequencing to ensure accuracy.
Purification and Enrichment of HematopoieticProgenitor CellsBone marrow cells from 8- to 10-week-old apoE�/� mice wereharvested from femurs and tibias. HPCs were purified by depletionof the Lin� cells with the Lineage Depletion SpinSep kit (Stem CellTechnologies). ScaI�/cKit� cells were further selected from the Lin�
cells using EasySep Scal� and cKit� selection kit (Stem CellTechnologies) to obtain the Scal�cKit�Lin� population.
Lentiviral Production and Ex Vivo Transductionof Mouse Peritoneal Macrophages and HPCs293T cells were used for lentiviral particle production according toprotocols from Trono’s Laboratory (http://tronolab.epfl.ch).Thioglycollate-elicited mouse peritoneal macrophages were col-lected and cultured in DMEM/10% FBS. Fresh media was replacedafter 2 hours, and the lentiviral particles were added at a multiplicityof infection (MOI) of 30. HPCs were transduced in Stemspan media(Stem Cell Technologies Inc) overnight before being transplantedinto recipient mice at 50 000 HPCs per mouse. The transductionefficiency of both macrophages and HPCs were measured by theGFP expression levels using flow cytometery.
Western Blot Analysis of ABCA1, ABCG1, SRBI,Human ApoAI, and Murine ApoEMacrophage culture media was collected after 24 hours of incubationand concentrated with Microcon-10 (Millipore Corporation) andsubjected to apoAI and apoE Western blot analysis as previouslydescribed.7,8 Western blot analyses for ABCA1, ABCG1, and SR-BIin transduced macrophages were conducted according to a methoddescribed previously.13
Fast Protein Liquid Analysis of MacrophageCulture MediaMacrophage culture media was collected, concentrated, and sub-jected to fast protein liquid chromatography (FPLC) separation.Pools of 3 fractions were concentrated and Western blot analysis ofapoAI and apoE performed as described above.
Real-Time RT-PCR Quantitation of ApoAI, ApoE,ABCA1, ABCG1, and SR-BI mRNATotal RNA was collected using the Purescript kit (Genetra System).The cDNAs were reverse-transcribed using the High Capacity cDNAArchive kit (ABI, Applied Biosystems). Primers and probes werepurchased from ABI, and the real-time RT-PCR reactions werecarried out on a Taqman 7900 (ABI) according to a modified methoddescribed previously.14
Enzyme-Linked Immunosorbent Assay forDetection of Human ApoAIHuman apoAI protein levels from macrophage culture media as wellas serum from mice receiving apoAI transduced HPCs were deter-mined by enzyme-linked immunosorbent assay (ELISA) as previ-ously described.8,9
Immunofluorescent Staining of Human ApoAI andCD68 in the Proximal Aortic LesionsFive-micrometer frozen sections were fixed in cold acetone, blockedwith 4% BSA in PBS, then incubated with anti-human apoAIantibody (Biodesign) or anti-mouse CD68-Alexa Fluor 488 (Sero-tec). The slides were then washed with PBS and incubated withCy3-conjugated donkey antigoat Ig-G (Jackson Immuno Researchlaboratories Inc). Vectashield mounting medium with DAPI (VectorLaboratories Inc) was used to stain nuclei. Images were capturedusing an Olympus DP71 microscope and Olympus DP controllerimage software.
Cholesterol Efflux StudiesCholesterol efflux from murine peritoneal macrophages was mea-sured by a modified procedure from Mazzone15,16 and Yancey.17
Briefly, macrophages were transduced with LV-apoAI or LV-GFP,cultured in DMEM/10% FBS for 24 hours, and then incubated with100 �g/mL acLDL and 5 �Ci/mL 3H-cholesterol (NEN) in DMEM/0.1% FBS for 72 hours. Cells in controlwells (T0) were washed withDPBS, air dried, and extracted with isopropanol to provide a baselinevalue for total [3H] cholesterol content. Cells were incubated withefflux media (DMEM with or without 10 �g/mL apoAI) at 37°C for48 hours. At the end of the efflux period, the media were collected,the cells were incubated with isopropanol at room temperature for 4hours, and radioactivity in the media and cell extract was measuredby scintillation counting. Three independent experiments wereconducted.
Transplantation of HPCs into ApoE�/� Mice andQuantitation of Atherosclerotic LesionsHPCs were isolated from male apoE�/� mice (B6.129P2-Apoetm1Unc)at 12 to 13 weeks of age, and then transduced with LV-GFP orLV-apoAI at an MOI of 30 and cultured 22 to 24 hours in Stemspanmedia (Stem Cell technology). Female apoE�/� mice at 16 to 18weeks old were lethally irradiated (9 Gy) and transplanted with thetransduced HPCs. The mice were placed on regular chow diet.Plasma lipid profiles were monitored every 6 weeks posttransplan-tation. Plasma HDL-cholesterol was measured using the ACEHDL-C reagent (Diagnostic Technologies LLC). Mice were eutha-nized at 12 weeks posttransplantation. Spleen samples were collectedfrom mice that received LV-GFP transduced HPCs. Spleen cellswere incubated with fluorescently labeled antibodies for CD4, CD8a,B220, CD11b, and CD11C, and analyzed by flow cytometry. Thequantitative analysis of atherosclerosis in the proximal aorta and enface preparation was performed as described previously.18,19
1440 Arterioscler Thromb Vasc Biol August 2008
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

Statistical AnalysisStatistical analysis was performed with GraphPad PRISM software(GraphPad Software Inc). Results were expressed as the mean�SEintragroup and between-group comparisons were achieved usingeither 1-way ANOVA followed by the Bonferroni’s post test whenstatistical significance was detected, or the unpaired 2-tailed t testwhen appropriate.
ResultsHigh-Level Lentiviral-Mediated ApoAI Expressionin HPCs and Mouse Peritoneal MacrophagesWe have generated a lentiviral apoAI expressing constructand a GFP expressing construct driven by a macrophagespecific promoter, CD68. Supplemental Figure IA (availableonline at http://atvb.ahajournals.org) shows diagrams of theseconstructs. Using the conditions described above, we wereable to achieve greater than 70% and 25% transductionefficiency in primary cultured macrophages and HPCs, re-spectively, at an MOI of 50 (supplemental Figure IB).
ApoAI was efficiently secreted from macrophages, as itwas detected in the culture media and was not found in thecell lysate (Figure 1A). The distribution of apoAI in themedia was analyzed by FPLC in wild-type and apoE�/�
macrophages transduced with LV-apoAI. Endogenouslysynthesized apoAI was primarily detected in the HDLrange in both cell types, but was associated with largerparticles in apoE�/� macrophages, and relegated to smaller
particles in the presence of apoE (Figure 1B). Similar patternswere detected by ELISA in FPLC fractions (Figure 1D).
ApoAI was detected in the culture media of mouse peritonealmacrophages up to 4 weeks after transduction, indicating long-term stable expression of the apoAI transgene. The concentra-tion of apoAI in the media ranged from 440 to 510 ng/mL/106
cells in 24-hour cultures. When the apoAI-expressing HPCswere transplanted into lethally irradiated apoE�/� recipient mice,human apoAI was detected in plasma 12 weeks post-HPCtransplantation at a concentration of 8.4�1.2 �g/dL. Thisrepresents a nearly 50-fold improvement over our previousresults with retroviral apoAI–mediated bone marrow transduc-tion, which produced plasma apoAI at concentrations of 100 to200 ng/dL 12 weeks after BMT.9
The distribution of endogenous apoE in the media ofLV-apoAI–transduced wild-type macrophages was also ana-lyzed in the FPLC fractions. In complete agreement with thedata reported above for apoAI, we found that when macro-phages produce both apoE and apoAI, apoE was primarilyassociated with larger particles compared to control LV-GFPtransduced macrophages (Figure 1C).
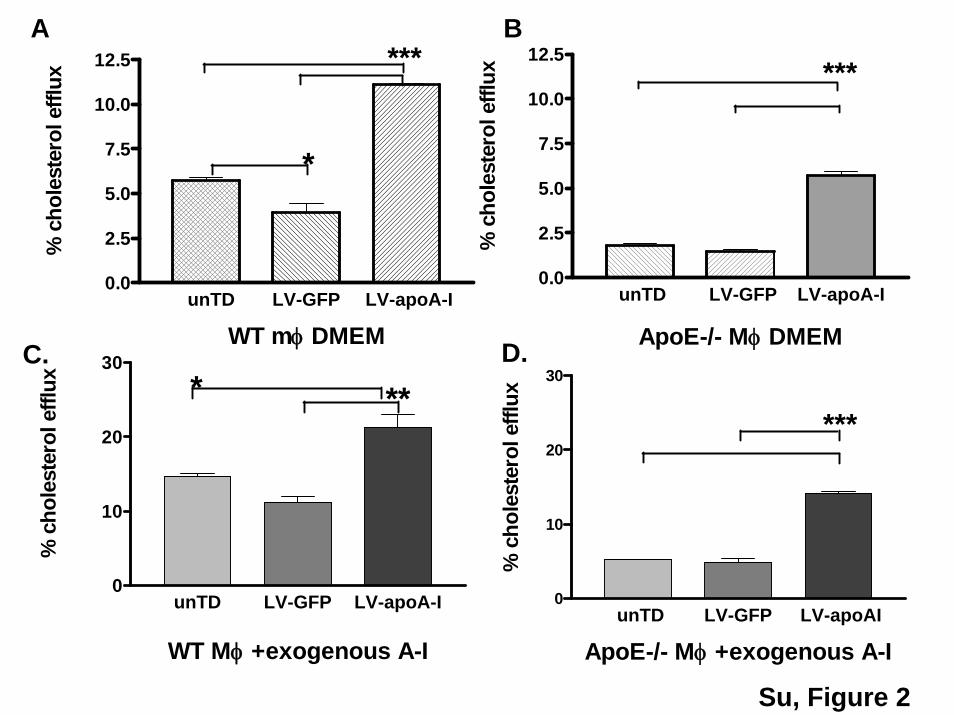
Cholesterol Efflux in Macrophages TransducedWith Lentiviral ApoAILV-apoAI transduction increased cholesterol efflux by 2.5-fold in wild-type macrophages (Figure 2A) and by 3.5-fold in
FPLC fractions
1 2 3 4 5 6wtLy wtMd E-/-Ly E-/-Md
A
apoA-I 28 kd
BH
um
A-I
14-1
7
18-2
0
21-2
3
24-2
6
27-2
9
30-3
2
33-3
5
Wt M
E-/- MapoA-I 28Kd
C
b.Wt LV-apoA-I
a. Wt LV-GFP
14-1
7
18-2
0
21-2
3
24-2
6
27-2
9
30-3
2
33-3
5
apoE 34Kd
D
15 17 19 21 23 25 27 29 31 33 35 370
25
50
75
100W T
ApoE-/-
Fraction #
Ap
oA
-I n
g/d
l
φφ
Figure 1. Detection of humanapoAI in transduced macro-phage culture media by West-ern blot and ELISA. A, Lane1:human plasma 1:1000; lane 2:LV-GFP macrophages; lanes 3and 4: LV-apoAI wt macro-phage cell lysate (wtLy) andculture media (wtMd); lanes 5and 6: LV-apoAI apoE�/� mac-rophage cell lysate (E�/�Ly)and culture media (E�/�Md).B, FPLC analysis of culturemedia. In apoE producing (wt)macrophages, apoAI is associ-ated with relatively small parti-cles, first appearing in fractions21 to 23, and still found infractions 33 to 35. In apoE�/�
macrophages, apoAI is associ-ated with relatively larger parti-cles, first appearing in fractions18 to 21 and disappearing infractions 33 to 35. C, Similarpatterns were detected byELISA in FPLC fractions. D,ApoE in FPLC fractions of theculture media. In LV-apoAI wtmacrophages, apoE was asso-ciated with large particles(fractions 14 to 32), whereas inthe LV-GFP wt macrophages,apoE was associated with rela-tively smaller particles (frac-tions 18 to 35).
Su et al Lentiviral Transduction of ApoAI Into HPCs and Macrophages 1441
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

apoE�/� macrophages (Figure 2B) compared to the LV-GFPtransduced controls. Our observation suggests that apoAIproduced in the apoE�/� macrophages is as efficient as apoEin mediating cholesterol efflux. Interestingly, the expressionof apoAI completely corrected the defective efflux caused byapoE deficiency (from 1.6 to 5.7%) and reinstated the samelevel of efflux seen in wild-type macrophages (Figure 2A).The addition of exogenous apoAI in the efflux media (10�g/mL) resulted in a further increase in cholesterol efflux inboth wild-type (from 11% to 21%) and apoE�/� (from 5.7%to 14%) macrophages compared to DMEM alone, indicatingthat endogenous and exogenous apoAI have a synergisticeffect on cholesterol efflux.
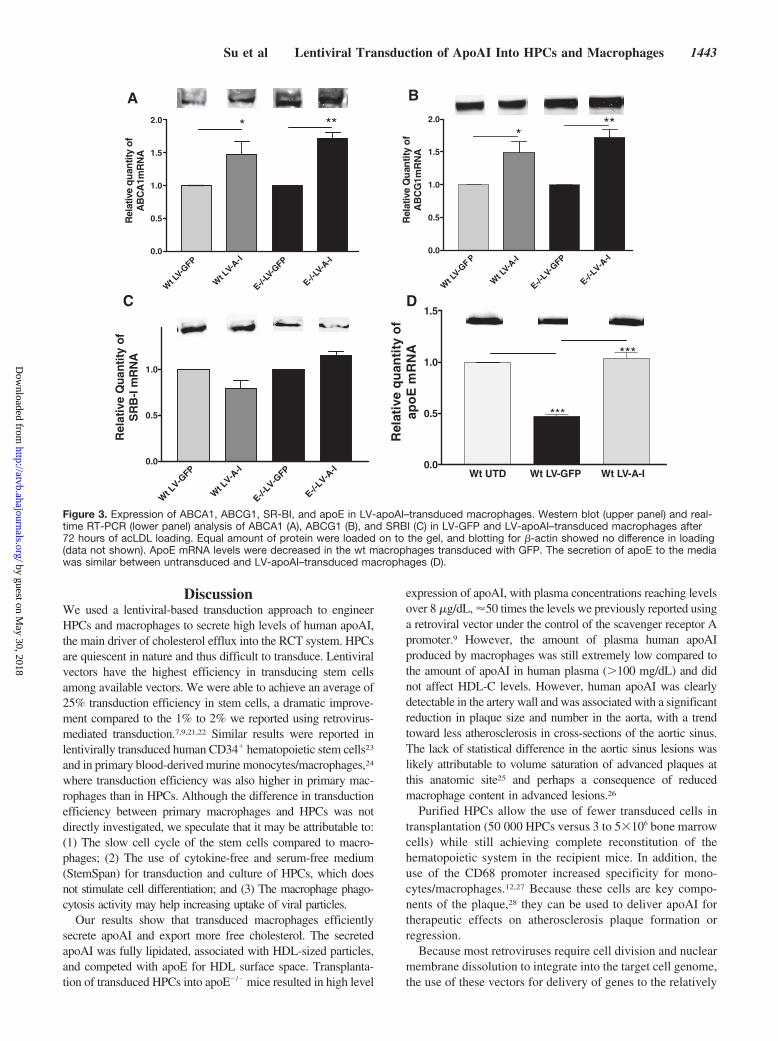
Effects of ApoAI Expression on CholesterolTransporters in MacrophagesABCA1 and ABCG1 mRNA levels were significantly in-creased in both wild-type and apoE�/� macrophages afteracLDL loading when compared to GFP-transduced macro-phages (Figure 3A and 3B, lower panel). These were accom-panied by an increase in ABCA1 and ABCG1 protein levels(Figure 3A and 3B, upper panel). Interestingly, SR-BI mRNAlevels were decreased by 20% in LV-apoAI–transducedwild-type macrophages (but not in apoE�/� macrophages)compared to GFP expressing cells (Figure 3C, lower panel)However, SR-BI protein levels were not affected (Figure 3C,upper panel). In addition, transduction of wild-type macro-
phages with LV-GFP reduced apoE mRNA levels by 50%,whereas LV-apoAI–transduced macrophages showed no sig-nificant changes in apoE secretion (Figure 3D).
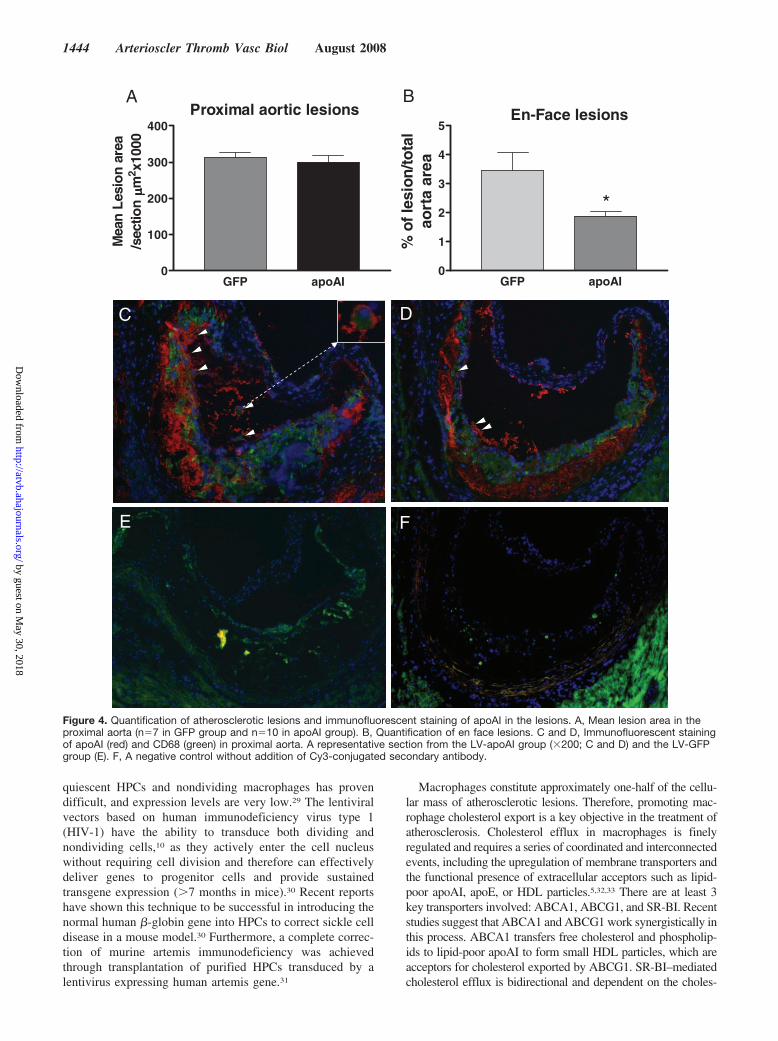
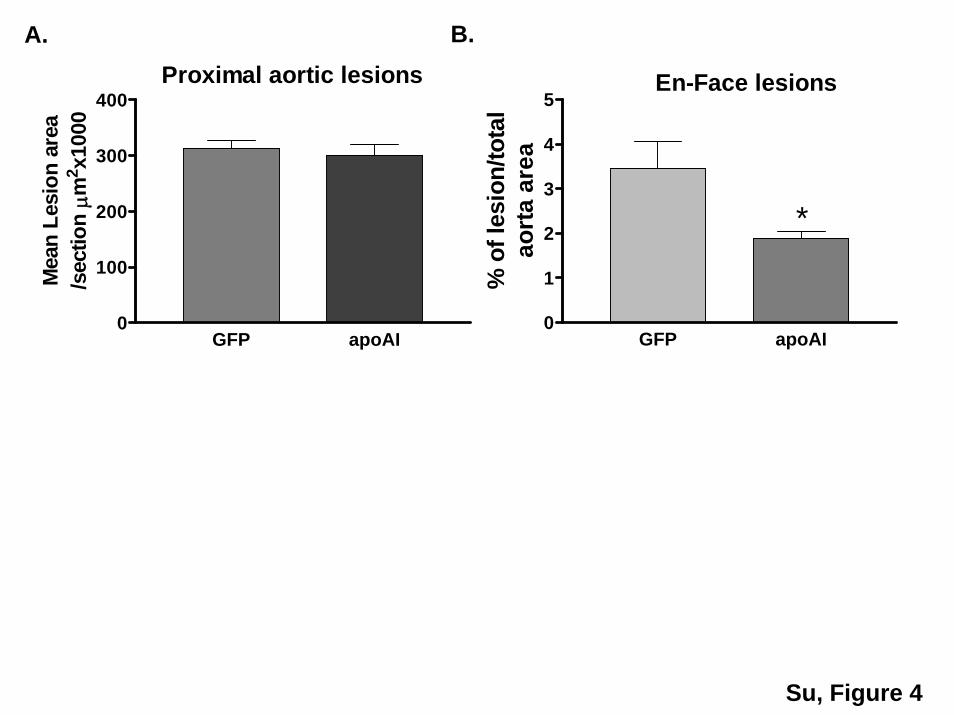
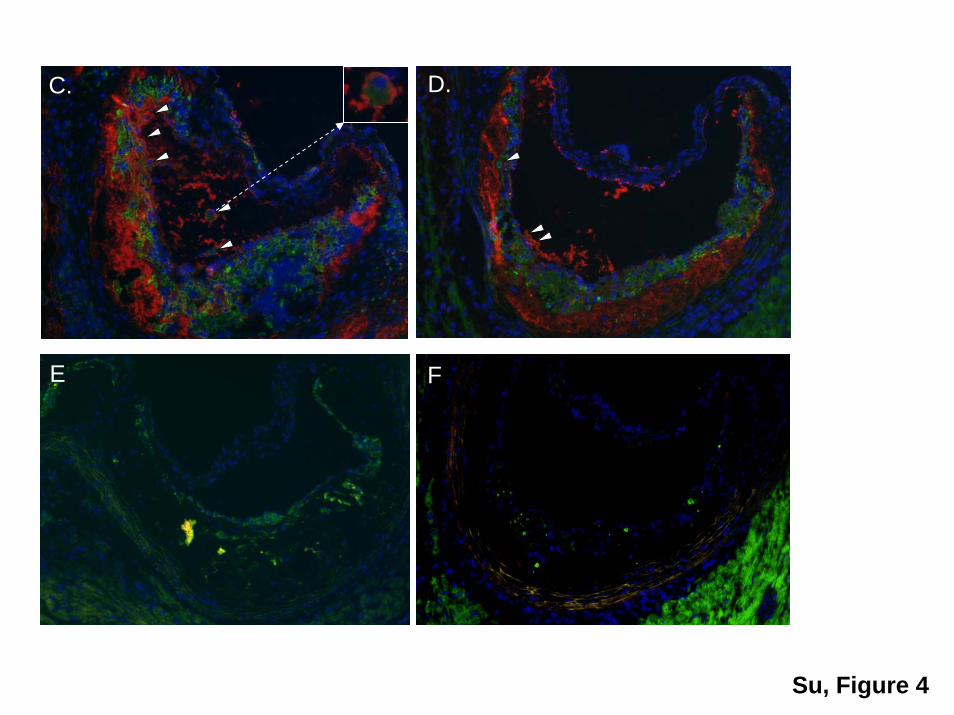
Transplantation of ApoAI-Producing HPCs intoApoE�/� Mice Reduces Atherosclerosis WithoutChanging Plasma Lipid ProfilesTwelve weeks after transplant, atherosclerotic lesions inapoAI-HPC recipient apoE�/� mice were 50% smaller than inGFP-HPC recipient controls. No significant differences wereseen in the aortic sinus (Figure 4A and 4B). Human apoAIwas identified in the proximal aortic area by immunofluores-cence staining in LV-apoAI HPC recipients (Figure 4C and4D). No differences were detected between groups in totalcholesterol, HDL-C, and lipoprotein profiles during thecourse of study (Figure 5A, 5B, and 5C).
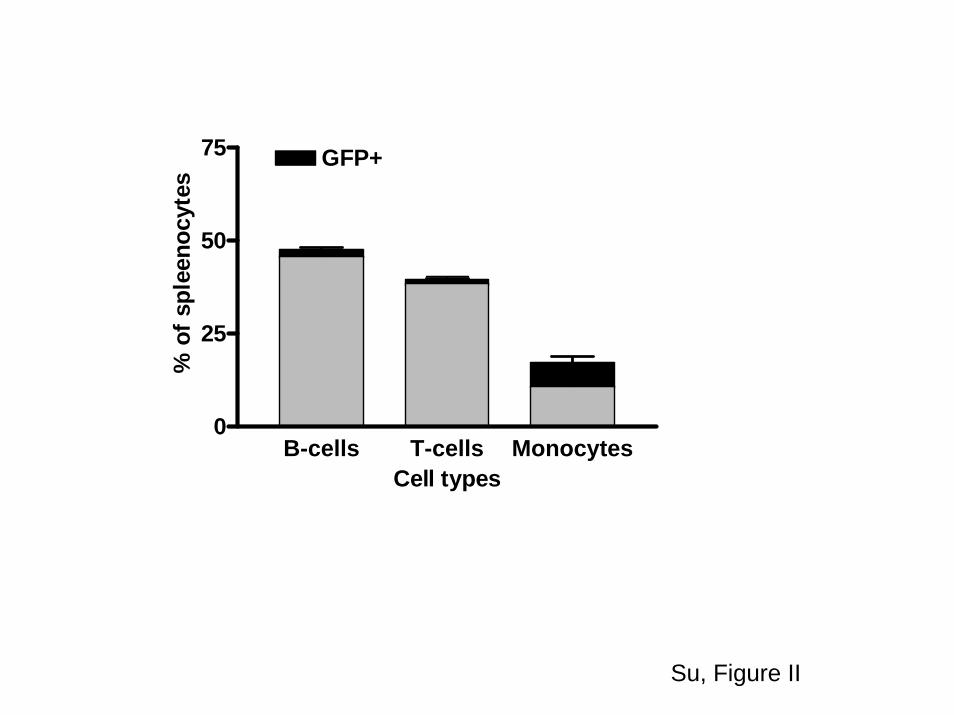
HPC transplantation into lethally irradiated apoE�/� micereconstituted the normal lineage of hematopoietic cells. Flowcytometric analysis of splenocytes collected from recipientsof LV-GFP–transduced HPCs showed similar percentages ofB-cells, T-cells, and monocytes as compared to nontrans-planted age-matched apoE�/� mice (supplemental FigureII).20 The use of the CD68 promoter resulted in a significantlyhigher percentage of GFP expression among CD11b� orCD11C� cells (macrophages) relative to B-cells (B-220�) andT-cells (CD4� and CD8�).
A B
C
unTD LV-GFP LV-apoA-I0
10
20
30
* **
WT Mφφφφ +exogenous A-I
% c
ho
lest
ero
l eff
lux
unTD LV-GFP LV-apoA-I0.0
2.5
5.0
7.5
10.0
12.5 ***
*
WT mφφφφ DMEM
% c
ho
lest
ero
l eff
lux
unTD LV-GFP LV-apoA-I0.0
2.5
5.0
7.5
10.0
12.5***
ApoE-/- Mφφφφ DMEM
% c
ho
lest
ero
l eff
lux
unTD LV-GFP LV-apoAI0
10
20
30
***
ApoE-/- Mφφφφ +exogenous A-I
% c
ho
lest
ero
l eff
lux
D
Figure 2. Cholesterol efflux in macrophages transduced with LV- apoAI. A and B, The effects of apoAI expressed from wt and apoE�/�
macrophages on cholesterol efflux to DMEM (endogenous apoA-I only). C and D, Cholesterol efflux with both endogenous and exogne-ous apoAI in the efflux media (DMEM �10 �g/mL of apoAI).
1442 Arterioscler Thromb Vasc Biol August 2008
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
DiscussionWe used a lentiviral-based transduction approach to engineerHPCs and macrophages to secrete high levels of human apoAI,the main driver of cholesterol efflux into the RCT system. HPCsare quiescent in nature and thus difficult to transduce. Lentiviralvectors have the highest efficiency in transducing stem cellsamong available vectors. We were able to achieve an average of25% transduction efficiency in stem cells, a dramatic improve-ment compared to the 1% to 2% we reported using retrovirus-mediated transduction.7,9,21,22 Similar results were reported inlentivirally transduced human CD34� hematopoietic stem cells23
and in primary blood-derived murine monocytes/macrophages,24
where transduction efficiency was also higher in primary mac-rophages than in HPCs. Although the difference in transductionefficiency between primary macrophages and HPCs was notdirectly investigated, we speculate that it may be attributable to:(1) The slow cell cycle of the stem cells compared to macro-phages; (2) The use of cytokine-free and serum-free medium(StemSpan) for transduction and culture of HPCs, which doesnot stimulate cell differentiation; and (3) The macrophage phago-cytosis activity may help increasing uptake of viral particles.
Our results show that transduced macrophages efficientlysecrete apoAI and export more free cholesterol. The secretedapoAI was fully lipidated, associated with HDL-sized particles,and competed with apoE for HDL surface space. Transplanta-tion of transduced HPCs into apoE�/� mice resulted in high level
expression of apoAI, with plasma concentrations reaching levelsover 8 �g/dL, �50 times the levels we previously reported usinga retroviral vector under the control of the scavenger receptor Apromoter.9 However, the amount of plasma human apoAIproduced by macrophages was still extremely low compared tothe amount of apoAI in human plasma (�100 mg/dL) and didnot affect HDL-C levels. However, human apoAI was clearlydetectable in the artery wall and was associated with a significantreduction in plaque size and number in the aorta, with a trendtoward less atherosclerosis in cross-sections of the aortic sinus.The lack of statistical difference in the aortic sinus lesions waslikely attributable to volume saturation of advanced plaques atthis anatomic site25 and perhaps a consequence of reducedmacrophage content in advanced lesions.26
Purified HPCs allow the use of fewer transduced cells intransplantation (50 000 HPCs versus 3 to 5�106 bone marrowcells) while still achieving complete reconstitution of thehematopoietic system in the recipient mice. In addition, theuse of the CD68 promoter increased specificity for mono-cytes/macrophages.12,27 Because these cells are key compo-nents of the plaque,28 they can be used to deliver apoAI fortherapeutic effects on atherosclerosis plaque formation orregression.
Because most retroviruses require cell division and nuclearmembrane dissolution to integrate into the target cell genome,the use of these vectors for delivery of genes to the relatively
Wt UTD Wt LV-GFP Wt LV-A-I0.0
0.5
1.0
1.5
***
***
Rel
ativ
e q
uan
tity
of
apo
E m
RN
A
Wt L
V-GFP
Wt L
V-A-I
E-/-LV
-GFP
E-/-LV
-A-I
0.0
0.5
1.0
1.5
2.0 * **R
elat
ive
qu
antit
y o
fA
BC
A1m
RN
A
Wt L
V-GF P
Wt L
V-A-I
E-/-LV-G
FP
E-/-LV-A
-I0.0
0.5
1.0
1.5
2.0
Rel
ativ
e Q
uan
tity
of
AB
CG
1mR
NA
***
Wt L
V-GFP
Wt L
V-A-I
E-/-LV-G
FP
E-/-LV-A
-I0.0
0.5
1.0
Rel
ativ
e Q
uan
tity
of
SR
B-I
mR
NA
A B
C D
Figure 3. Expression of ABCA1, ABCG1, SR-BI, and apoE in LV-apoAI–transduced macrophages. Western blot (upper panel) and real-time RT-PCR (lower panel) analysis of ABCA1 (A), ABCG1 (B), and SRBI (C) in LV-GFP and LV-apoAI–transduced macrophages after72 hours of acLDL loading. Equal amount of protein were loaded on to the gel, and blotting for �-actin showed no difference in loading(data not shown). ApoE mRNA levels were decreased in the wt macrophages transduced with GFP. The secretion of apoE to the mediawas similar between untransduced and LV-apoAI–transduced macrophages (D).
Su et al Lentiviral Transduction of ApoAI Into HPCs and Macrophages 1443
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
quiescent HPCs and nondividing macrophages has provendifficult, and expression levels are very low.29 The lentiviralvectors based on human immunodeficiency virus type 1(HIV-1) have the ability to transduce both dividing andnondividing cells,10 as they actively enter the cell nucleuswithout requiring cell division and therefore can effectivelydeliver genes to progenitor cells and provide sustainedtransgene expression (�7 months in mice).30 Recent reportshave shown this technique to be successful in introducing thenormal human �-globin gene into HPCs to correct sickle celldisease in a mouse model.30 Furthermore, a complete correc-tion of murine artemis immunodeficiency was achievedthrough transplantation of purified HPCs transduced by alentivirus expressing human artemis gene.31
Macrophages constitute approximately one-half of the cellu-lar mass of atherosclerotic lesions. Therefore, promoting mac-rophage cholesterol export is a key objective in the treatment ofatherosclerosis. Cholesterol efflux in macrophages is finelyregulated and requires a series of coordinated and interconnectedevents, including the upregulation of membrane transporters andthe functional presence of extracellular acceptors such as lipid-poor apoAI, apoE, or HDL particles.5,32,33 There are at least 3key transporters involved: ABCA1, ABCG1, and SR-BI. Recentstudies suggest that ABCA1 and ABCG1 work synergistically inthis process. ABCA1 transfers free cholesterol and phospholip-ids to lipid-poor apoAI to form small HDL particles, which areacceptors for cholesterol exported by ABCG1. SR-BI–mediatedcholesterol efflux is bidirectional and dependent on the choles-
GFP apoAI0
100
200
300
400Proximal aortic lesions
Mea
n L
esio
n a
rea
/sec
tionµµ µµm
2 x10
00
GFP apoAI0
1
2
3
4
5 En-Face lesions
*
% o
f les
ion/
tota
lao
rta
area
A B
C D
E F
Figure 4. Quantification of atherosclerotic lesions and immunofluorescent staining of apoAI in the lesions. A, Mean lesion area in theproximal aorta (n�7 in GFP group and n�10 in apoAI group). B, Quantification of en face lesions. C and D, Immunofluorescent stainingof apoAI (red) and CD68 (green) in proximal aorta. A representative section from the LV-apoAI group (�200; C and D) and the LV-GFPgroup (E). F, A negative control without addition of Cy3-conjugated secondary antibody.
1444 Arterioscler Thromb Vasc Biol August 2008
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

terol gradient across the cell membrane.3 Our LV-apoAI–transduced macrophages showed an upregulation of ABCA1and ABCG1 expression after acLDL loading and, as a result, asubstantial increase in cholesterol efflux.
It is worth noting that expression of apoAI increasedcholesterol efflux in wild-type macrophages, suggesting acooperative effect of apoAI and apoE on cholesterol efflux.To our knowledge, this is the first report showing additiveeffects of endogenously produced cholesterol acceptors(apoAI and apoE) on increasing cholesterol exit from mac-rophage. This supports the contention that genetically engi-neered macrophages can serve as a useful tool for promoting
RCT in the plaque. In addition, the expression of apoAI inapoE�/� macrophages completely normalized cholesterol ef-flux to the level of wild-type macrophages, suggesting thatapoAI can compensate and rescue reduced cholesterol effluxattributable to the loss of apoE.
Crucial aspects of this work include: (1) lentiviral apoAIwas sufficient to completely normalize the impaired efflux inapoE�/� macrophages; (2) Lentiviral apoAI increased choles-terol efflux in normal macrophages, thus adding to the effectof apoE; (3) Adding exogenous apoAI caused a furtherincrease in cholesterol efflux in both wild-type and apoE�/�
macrophages, possibly because of the ability of exogenousapoAI to increase secretion of endogenous acceptors.34 Re-cent studies in our laboratory suggest that apoAI and apoEwork cooperatively in the formation of buoyant HDL bycholesterol-enriched macrophages17; (4) Lentivirally medi-ated apoAI transduction in HPCs revealed a long-term sus-tained expression of apoAI in transplanted recipient micewith reduction of aortic lesion size. One can anticipate thatthe concurrent production of both apoAI and apoE frommacrophages in the artery wall will significantly promotecholesterol exit from these cells via both ABCA1-dependentand independent pathways.
It must be emphasized that with the current approach weare finally able to demonstrate an effect of macrophage apoAIin the presence of physiological amounts of apoE, and thatapoAI and apoE produced by macrophages work coopera-tively in promoting cholesterol efflux and maintaining cho-lesterol homeostasis. This is an essential milestone notachieved in previous studies7,8 where the effect of lowerexpression levels of apoAI was only detectable in the absenceof apoE.
In wild-type macrophages, where apoAI and apoE competefor binding on the lipoprotein surface, apoE associates withlarger HDL-size particles as compared to apoAI. Whetherthese large particles were generated as a result of enhancedcholesterol efflux through the ABCA1 pathway or due to anindependent mechanism is not clear. It can be speculated thatthe larger apoE-containing particles are better acceptors forABCG1-mediated cholesterol efflux.17
The protective effects of apoAI can be attributable toanti-inflammatory and antioxidant properties not directlyrelated to plasma HDL cholesterol levels.35 This is in linewith human studies showing that injections of recombinantHDL containing a protective human apoAI variant are suffi-cient to induce plaque regression without effects on plasmaHDL.36
The recent negative outcomes of several trials of the CETPinhibitor torcetrapib37–40 are a reminder that plasma HDLmay not be the sole target for therapeutic strategies directedat removing cholesterol from the arterial plaque. Our studiessupport the notion that increasing the concentration of apoAIin the lesion is a bona fide objective of gene therapyapproaches to control the atheroma.
AcknowledgmentsWe thank Drs Patricia G. Yancey and Amy S. Major for expertadvice and discussions, James Higginbotham at the Flow CytometryCore, and Ashwath Jayagopal at Biomedical Engineering Depart-ment at Vanderbilt University for excellent technical assistance.
A
C
B
GFP apoA-I0
25
50
75
100
HD
L ch
oles
tero
l mg/
dl
0 6 120
100
200
300
400
500
600 GFPApoA-I
Weeks Post-HPCT
Tota
l ser
um
cho
lest
ero
l (m
g/d
l)
10 15 20 25 30 35 400
10
20
30
40
50
60GFPapoA-I
Fractions
Ch
ole
ster
ol m
g/dl
Figure 5. Lipid profile in apoE�/� mice transplanted with apoAIexpressing HPCs. A, Total serum cholesterol in HPCs trans-planted apoE�/� recipient mice. B, Plasma HDL-C levels inapoE�/� recipient mice 12 weeks after HPC transplantation.C, Representative FPLC analysis of plasma lipid profile of apoE�/�
recipient mice 12 weeks after HPC transplantation.
Su et al Lentiviral Transduction of ApoAI Into HPCs and Macrophages 1445
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

Sources of FundingThis work was supported by NIH grant HL65709 (to S.F.) and in partby NIH grants HL57986 (to S.F.), HL65405 (to M.F.L.), andP60DK20593 (to Y.R.S.).
DisclosuresNone.
References1. Linton MF, Fazio S. Macrophages, inflammation, and atherosclerosis. Int
J Obes Relat Metab Disord. 2003;27 Suppl 3:S35–S40.2. Maxfield FR, Tabas I. Role of cholesterol and lipid organization in
disease. Nature. 2005;438:612–621.3. Jessup W, Gelissen IC, Gaus K, Kritharides L. Roles of ATP binding
cassette transporters A1 and G1, scavenger receptor BI and membranelipid domains in cholesterol export from macrophages. Curr OpinLipidol. 2006;17:247–257.
4. Zannis VI, Chroni A, Krieger M. Role of apoA-I, ABCA1, LCAT, andSR-BI in the biogenesis of HDL. J Mol Med. 2006;84:276–294.
5. Gelissen IC, Harris M, Rye KA, Quinn C, Brown AJ, Kockx M, CartlandS, Packianathan M, Kritharides L, Jessup W. ABCA1 and ABCG1 syn-ergize to mediate cholesterol export to apoA-I. Arterioscler Thromb VascBiol. 2006;26:534–540.
6. Vaughan AM, Oram JF. ABCA1 and ABCG1 or ABCG4 act sequentiallyto remove cellular cholesterol and generate cholesterol-rich HDL. J LipidRes. 2006;47:2433–2443.
7. Ishiguro H, Yoshida H, Major AS, Zhu T, Babaev VR, Linton MF, FazioS. Retrovirus-mediated expression of apolipoprotein A-I in the macro-phage protects against atherosclerosis in vivo. J Biol Chem. 2001;276:36742–36748.
8. Major AS, Dove DE, Ishiguro H, Su YR, Brown AM, Liu L, Carter KJ,Linton MF, Fazio S. Increased cholesterol efflux in apolipoprotein AI(ApoAI)-producing macrophages as a mechanism for reduced atherosclerosisin ApoAI(�/�) mice. Arterioscler Thromb Vasc Biol. 2001;21:1790–1795.
9. Su YR, Ishiguro H, Major AS, Dove DE, Zhang W, Hasty AH, BabaevVR, Linton MF, Fazio S. Macrophage apolipoprotein A-I expressionprotects against atherosclerosis in ApoE-deficient mice and up-regulatesABC transporters. Mol Ther. 2003;8:576–583.
10. Kondo M, Wagers AJ, Manz MG, Prohaska SS, Scherer DC, BeilhackGF, Shizuru JA, Weissman IL. Biology of hematopoietic stem cells andprogenitors: implications for clinical application. Annu Rev Immunol.2003;21:759–806.
11. Woods NB, Mikkola H, Nilsson E, Olsson K, Trono D, Karlsson S.Lentiviral-mediated gene transfer into haematopoietic stem cells. J InternMed. 2001;249:339–343.
12. Gough PJ, Raines EW. Gene therapy of apolipoprotein E-deficient miceusing a novel macrophage-specific retroviral vector. Blood. 2003;101:485–491.
13. Zhang W, Yancey PG, Su YR, Babaev VR, Zhang Y, Fazio S, Linton MF.Inactivation of macrophage scavenger receptor class B type I promotesatherosclerotic lesion development in apolipoprotein E-deficient mice.Circulation. 2003;108:2258–2263.
14. Su YR, Linton MF, Fazio S. Rapid quantification of murine ABC mRNAsby real time reverse transcriptase-polymerase chain reaction. J Lipid Res.2002;43:2180–2187.
15. Lin CY, Duan H, Mazzone T. Apolipoprotein E-dependent cholesterol effluxfrom macrophages: kinetic study and divergent mechanisms for endogenousversus exogenous apolipoprotein E. J Lipid Res. 1999;40:1618–1627.
16. Mazzone T, Reardon C. Expression of heterologous human apoli-poprotein E by J774 macrophages enhances cholesterol efflux to HDL3.J Lipid Res. 1994;35:1345–1353.
17. Yancey PG, Yu H, Linton MF, Fazio S. A pathway-dependent on apoE,ApoAI, and ABCA1 determines formation of buoyant high-densitylipoprotein by macrophage foam cells. Arterioscler Thromb Vasc Biol.2007;27:1123–1131.
18. Linton MF, Atkinson JB, Fazio S. Prevention of atherosclerosis in apo-lipoprotein E-deficient mice by bone marrow transplantation. Science.1995;267:1034–1037.
19. Paigen B, Morrow A, Holmes PA, Mitchell D, Williams RA. Quantitativeassessment of atherosclerotic lesions in mice. Atherosclerosis. 1987;68:231–240.
20. Major AS, Fazio S, Linton MF. B-lymphocyte deficiency increases ath-erosclerosis in LDL receptor-null mice. Arterioscler Thromb Vasc Biol.2002;22:1892–1898.
21. Dunbar CE, Cottler-Fox M, O’Shaughnessy JA, Doren S, Carter C,Berenson R, Brown S, Moen RC, Greenblatt J, Stewart FM. Retrovirallymarked CD34-enriched peripheral blood and bone marrow cells con-tribute to long-term engraftment after autologous transplantation. Blood.1995;85:3048–3057.
22. Miller DG, Adam MA, Miller AD. Gene transfer by retrovirus vectorsoccurs only in cells that are actively replicating at the time of infection.Mol Cell Biol. 1990;10:4239–4242.
23. Di Nunzio F, Piovani B, Cosset FL, Mavilio F, Stornaiuolo A. Trans-duction of human hematopoietic stem cells by lentiviral vectorspseudotyped with the RD114-TR chimeric envelope glycoprotein. HumGene Ther. 2007;18:811–820.
24. Zeng L, Yang S, Wu C, Ye L, Lu Y. Effective transduction of primary mouseblood- and bone marrow-derived monocytes/macrophages by HIV-baseddefective lentiviral vectors. J Virol Methods. 2006;134:66–73.
25. Hasty AH, Linton MF, Brandt SJ, Babaev VR, Gleaves LA, Fazio S.Retroviral gene therapy in ApoE-deficient mice: ApoE expression in theartery wall reduces early foam cell lesion formation. Circulation. 1999;99:2571–2576.
26. Nakashima Y, Plump AS, Raines EW, Breslow JL, Ross R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughoutthe arterial tree. Arterioscler Thromb. 1994;14:133–140.
27. Gough PJ, Raines EW. Advances in retroviral transduction of hemato-poietic stem cells for the gene therapy of atherosclerosis. Curr OpinLipidol. 2003;14:491–497.
28. Hansson GK. Immune mechanisms in atherosclerosis. ArteriosclerThromb Vasc Biol. 2001;21:1876–1890.
29. Challita PM, Kohn DB. Lack of expression from a retroviral vector aftertransduction of murine hematopoietic stem cells is associated with meth-ylation in vivo. Proc Natl Acad Sci USA. 1994;91:2567–2571.
30. Levasseur DN, Ryan TM, Pawlik KM, Townes TM. Correction of amouse model of sickle cell disease: lentiviral/antisickling beta-globingene transduction of unmobilized, purified hematopoietic stem cells.Blood. 2003;102:4312–4319.
31. Mostoslavsky G, Fabian AJ, Rooney S, Alt FW, Mulligan RC.Complete correction of murine Artemis immunodeficiency bylentiviral vector-mediated gene transfer. Proc Natl Acad Sci USA. 2006;103:16406–16411.
32. Duong M, Collins HL, Jin W, Zanotti I, Favari E, Rothblat GH. Relativecontributions of ABCA1 and SR-BI to cholesterol efflux to serum fromfibroblasts and macrophages. Arterioscler Thromb Vasc Biol. 2006;26:541–547.
33. Feng B, Tabas I. ABCA1-mediated cholesterol efflux is defective in freecholesterol-loaded macrophages. Mechanism involves enhanced ABCA1degradation in a process requiring full NPC1 activity. J Biol Chem.2002;277:43271–43280.
34. Kockx M, Rye KA, Gaus K, Quinn CM, Wright J, Sloane T, Sviridov D,Fu Y, Sullivan D, Burnett JR, Rust S, Assmann G, Anantharamaiah GM,Palgunachari MN, Katz SL, Phillips MC, Dean RT, Jessup W, KritharidesL. Apolipoprotein A-I-stimulated apolipoprotein E secretion from humanmacrophages is independent of cholesterol efflux. J Biol Chem. 2004;279:25966–25977.
35. Kontush A, Chapman MJ. Functionally defective high-densitylipoprotein: a new therapeutic target at the crossroads of dyslipidemia,inflammation, and atherosclerosis. Pharmacol Rev. 2006;58:342–374.
36. Nissen SE, Tsunoda T, Tuzcu EM, Schoenhagen P, Cooper CJ, Yasin M,Eaton GM, Lauer MA, Sheldon WS, Grines CL, Halpern S, Crowe T,Blankenship JC, Kerensky R. Effect of recombinant ApoA-I Milano oncoronary atherosclerosis in patients with acute coronary syndromes: arandomized controlled trial. JAMA. 2003;290:2292–2300.
37. Kastelein JJ, van Leuven SI, Burgess L, Evans GW, Kuivenhoven JA,Barter PJ, Revkin JH, Grobbee DE, Riley WA, Shear CL, Duggan WT,Bots ML. Effect of torcetrapib on carotid atherosclerosis in familialhypercholesterolemia. N Engl J Med. 2007;356:1620–1630.
38. Nissen SE, Tardif JC, Nicholls SJ, Revkin JH, Shear CL, Duggan WT,Ruzyllo W, Bachinsky WB, Lasala GP, Tuzcu EM. Effect of torcetrapib onthe progression of coronary atherosclerosis. N Engl J Med. 2007;356:1304–1316.
39. Tall AR. CETP inhibitors to increase HDL cholesterol levels. N EnglJ Med. 2007;356:1364–1366.
40. Barter PJ, Caulfield M, Eriksson M, Grundy SM, Kastelein JJ, Komajda M,Lopez-Sendon J, Mosca L, Tardif JC, Waters DD, Shear CL, Revkin JH,Buhr KA, Fisher MR, Tall AR, Brewer B. Effects of torcetrapib in patientsat high risk for coronary events. N Engl J Med. 2007;357:2109–2122.
1446 Arterioscler Thromb Vasc Biol August 2008
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from

Yan Ru Su, John L. Blakemore, Youmin Zhang, MacRae F. Linton and Sergio FazioApplications to Cell Therapy of Atherosclerosis
Lentiviral Transduction of ApoAI Into Hematopoietic Progenitor Cells and Macrophages:
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2008 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.107.1600932008;28:1439-1446; originally published online May 22, 2008;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/28/8/1439World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2008/05/23/ATVBAHA.107.160093.DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 30, 2018
http://atvb.ahajournals.org/D
ownloaded from
A B
C.
Su, Figure 2
unTD LV-GFP LV-apoA-I0
10
20
30* **
WT Mφ +exogenous A-I
% c
hole
ster
ol e
fflux
unTD LV-GFP LV-apoA-I0.0
2.5
5.0
7.5
10.0
12.5 ***
*
WT mφ DMEM
% c
hole
ster
ol e
fflux
unTD LV-GFP LV-apoA-I0.0
2.5
5.0
7.5
10.0
12.5***
ApoE-/- Mφ DMEM
% c
hole
ster
ol e
fflux
unTD LV-GFP LV-apoAI0
10
20
30
***
ApoE-/- Mφ +exogenous A-I
% c
hole
ster
ol e
fflux
D.

B.A.
Wt LV-G
FP
Wt LV-A
-I
E-/-LV-G
FP
E-/-LV-A
-I
0.0
0.5
1.0
1.5
2.0
Rel
ativ
e Q
uant
ity o
fA
BC
G1m
RN
A
***
Wt LV-G
FP
Wt LV-A
-I
E-/-LV-G
FP
E-/-LV-A
-I
0.0
0.5
1.0
1.5
2.0 * **
Rel
ativ
e qu
antit
y of
AB
CA
1mR
NA
C.
Wt UTD Wt LV-GFP Wt LV-A-I0.0
0.5
1.0
1.5
***
***R
elat
ive
quan
tity
ofap
oE m
RN
A
D.
Wt LV-G
FP
Wt LV-A
-I
E-/-LV-G
FP
E-/-LV-A
-I
0.0
0.5
1.0
Rel
ativ
e Q
uant
ity o
fSR
B-I
mR
NA
Su, Figure 3
GFP apoAI0
100
200
300
400Proximal aortic lesions
Mea
n Le
sion
are
a/s
ectio
nµm
2 x10
00
GFP apoAI0
1
2
3
4
5 En-Face lesions
*
% o
f les
ion/
tota
lao
rta
area
A. B.
Su, Figure 4
Su, Figure 4
C. D.
E F

Su, Figure 5
A.
C.
B.
GFP apoA-I0
25
50
75
100
HD
L ch
oles
tero
l mg/
dl
0 6 120
100
200
300
400
500
600 GFPApoA-I
Weeks Post-HPCT
Tota
l ser
um c
hole
ster
ol (m
g/dl
)
10 15 20 25 30 35 400
10
20
30
40
50
60GFPapoA-I
Fractions
Cho
lest
erol
mg/
dl

ApoA-I
SD SA
5’ LTR cPPT 3’ LTR SIN
WPREψ
apoA-ICD68 Prmt
SD SA
5’ LTR cPPT 3’ LTR SIN
WPREψ
GFPCD68 PrmtGFP
Su, Figure I
A. The constructs

B. Expression of GFP with varying multiplicities of infection (MOI).
0 25 50 75 100 125 1500
25
50
75
100
MOI
% G
FP p
ositi
ve c
ells
c.
Su, Figure. I
Phase GFP+ Mφ
d.
Phase GFP+ HPC
a. Macrophages b.HPCs
0 10 20 30 40 500
10
20
30
MOI
% o
f GFP
pos
itive
Cel
ls
B-cells T-cells Monocytes0
25
50
75 GFP+
Cell types
% o
f spl
eeno
cyte
s
Su, Figure II

Figure legends
FigureI. Lentiviral Expression constructs and LV-GFP expression in HPCs
and macrophages. A. Schematic illustration of LV-GFP and LV-apoAI cDNA
expression constructs. The human apoAI cDNA and Green Fluorescent Protein
(GFP) are driven by a human macrophage specific promoter, CD68.13 The HIV-1
3’ long terminal repeat (LTR) is shown with a self inactivating (SIN) deletion. Ψ:
Packaging signal. SD, SA: splicing donor and acceptor site. CPPT: central
polypurine tract or DNA flap/central DNA termination sequence. HGH: human
growth hormone poly-A signal sequence. WPRE: woodchuck hepatitis virus post-
transcriptional regulatory element. B. Expression of GFP with varying
multiplicities of infection (MOI). a. LV-GFP transduced macrophages. At an MOI
of 50, more than 70% of the transduced macrophages express GFP three days
after transduction. b. LV-GFP transduced HPCs. At an MOI of 50, more than
25% of the transduced HPCs expressed GFP three days after transduction.
Microscopic images of lentiviral-CD68-GFP transduced macrophages (c) and
HPCs (d).
Figure II: Flow cytometeric analysis of splenocytes from apoE-/-recipient
mice transplanted with LV-GFP. About 50% of the splenocytes were positive
for B-220 and among these cells less than 2% were GFP positive. There were
less than 1% GFP positive cells that were CD4+ or CD8a+. Notably, 16% of the
splenocytes were monocytes, positive for (CD11b+, or CD11C+) and among
those 7% were positive for GFP.

FPLC fractions
1 2 3 4 5 6wtLy wtMd E-/-Ly E-/-Md
A.
apoA-I 28 kd
B.
Hum
A-I
14-1
7
18-2
0
21-2
3
24-2
6
27-2
9
30-3
2
33-3
5
Wt MΦE-/- MΦ
apoA-I 28Kd
Su, Figure 1
C.
b.Wt LV-apoA-Ia. Wt LV-GFP
14-1
7
18-2
0
21-2
3
24-2
6
27-2
9
30-3
2
33-3
5
apoE 34Kd
D.
15 17 19 21 23 25 27 29 31 33 35 370
25
50
75
100W T
ApoE-/-
Fraction #
Apo
A-I
ng/d
l