lecture 20: membrane transport energetics of transmembrane transport active transport passive...
Post on 21-Dec-2015
240 views
TRANSCRIPT

Lecture 20:Membrane Transport
Energetics of Transmembrane Transport
Active Transport
Passive Transport

FreeEnergy
Equilibrium
ConcentrationDifference
Progress of Reaction
Molecules Diffuse from High to Low Concentration
HighConcentration
Low Concentration
Equal
Concentrations
Equilibrium
There is an energy difference associated with a transmembrane concentrationdifference. The energy difference become zero at equilibrium, when thetwo concentrations are equal.

HighConcentration
Low Concentration
C2
C1
ConcentrationDifference
ACME
Battery
Co.
+
-
VoltageDifference
+++
+
+
+
+
Charged Molecules Create an Electrical Energy Difference
Charged molecules repel each other, increasing the energy differenceof a transmembrane concentration gradient.

Energy is Required to Create a Transmembrane Concentration Gradient
G = RT ln ( C2 / C1 ) + Z F V
The energy stored in a transmembrane concentration gradientdepends on the concentration difference and the electrical potential difference.
R 8.314 J/ ( mol K ) (Gas Constant)T Temperature in KelvinC1,C2 ConcentrationsZ Charge on moleculeF 96.5 kJ/ ( mol V ) (Faraday Constant)V Electrical potential difference in Volts
Contribution of concentration
difference
Contribution of electricalpotential
difference

Example: Glucose transportC1 = 1 mM, C2 = 66 mM, T = 310K , V = -50 mV
Example: Na+ transportC1 = 143 mM, C2 = 14 mM, T = 310K, V = -50 mV
G = RT ln ( C2 / C1 ) + Z F V
= (8.314 J/(molK))(310K) ln (66mM/1mM) + 0 = 8.314 x 310 x 4.190 J/mol = 10.8 kJ/mol
G = RT ln ( C2 / C1 ) + Z F V = (8.314 J/(molK))(310K) ln (14 mM / 143 mM) + (1)(96.5 kJ/(mol V))(-50mV) = 8.314 x 310 x (-2.324) J/mol + 96.5 x ( -50 ) J/mol = -6.0 kJ/mol + -4.8 kJ/mol = -10.8 kJ/mol (transport from high to low concentration is favorable, especially if aided by the electrical potential)
(transport from low concentration to high is unfavorable)

ABC Transporters
The ABC transporters are a large family of proteins which have thefunction of transporting substances across a membrane using the energyof ATP hydrolysis. ABC stands for ATP Binding Cassette.
An example is the multidrug resistance proteinMDR1. This protein is capable of using ATP toexpel a wide variety of small molecules fromcells. Cultured cancer cells can becomeresistant to a variety of drugs through its action.
Similar proteins are implicated in the acquisitionof drug resistance in a variety of cancers andcan contribute to the failure of chemotherapy.Drugs that specifically inhibit these proteinsare under development.

P-type ATPases as Examples of Ion PumpsAnother large family of transmembrane transporters is the P-type ATPases. Proteins in this family transport ions and other substances across membranes using the energy of ATP hydrolysis.
An example is the Ca2+ ATPase found in the membrane of the sarcoplasmic reticulum.
Muscle contraction is triggered by high levels of calcium ions in the cytosol of muscle cells. The Ca2+ ATPase enables the muscle to relax by pumping the cytosolic calcium into a specialized calcium storage compartment, the sarcoplasmic reticulum (SR).
The Ca2+ ATPase is able to pump calciumagainst a rather steep gradient. The pump maintains a concentration of Ca2+ in the cytosol of ~0.1M, and in the SR it is ~1.5 mM, a 15,000-fold increase.

Structure of the Ca2+ ATPase
The calcium ATPase is a 110 kD protein with4 domains.
The membrane-spanning domain contains 10-helices.
The three cytosolic domains (N,P,A domains)have different functions.
The N domain binds ATP. (N for Nucleotide-binding)
The P domain contains a phosphorylation site (Asp351) which accepts a phosphate from ATP during thecatalytic cycle. (P for Phosphorylation)
The A domain regulates Ca2+ binding and release.(A for Actuator)

Mechanism of the Ca2+ ATPase 1. Binding of ATP and 2 Ca2+ ions on the cytoplasmic side to the E1 state.
2. Phosphorylation of Asp 351 by ATP.
3. Transition to E2 state: opening of the Ca2+ siteto the luminal side of the membrane. (eversion)
4. Calcium release (low affinity due to phosphorylation.)
5. Phosphate hydrolysis and release.
6. Return to E1 state. (eversion)
The catalytic cycle of the pump involvesconformational changes between two states E1and E2 in which the calcium-binding sites areaccessible to different sides of the membrane.ATP hydrolysis provides the energy to powerthese changes in conformation. Two Ca2+ ionsare transported in each cycle of ATP hydrolysis.

The Na/K Pump
Another P-type ATPase is the Na/K pump.This enzyme uses the energy of ATP hydrolysis tosimultaneously transport 3 Na+ ions out of the celland 2 K+ ions into the cell.
The concentration gradient established by theNa/K pump enables nerve impulses and musclecontraction, controls cell volume, and providesenergy for uptake of sugars and amino acids.
The importance of this protein is indicated by thefact that fully one-third of the ATP consumed by aresting animal is used to establish theseconcentration gradients.
The Na/K pump has very similar mechanism to theCa2+ ATPase.
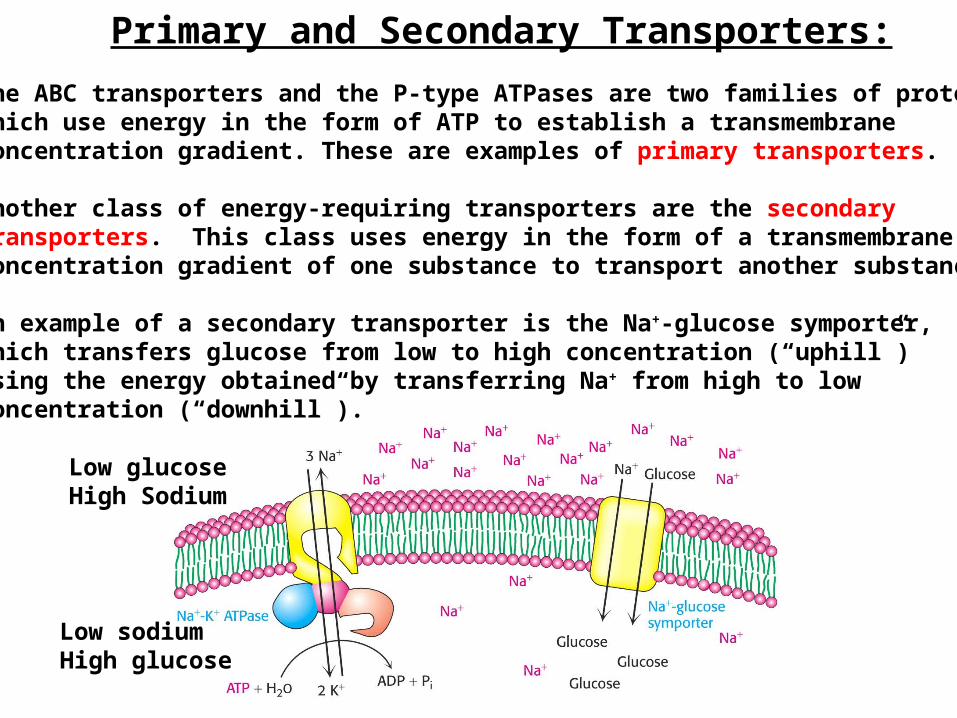
Primary and Secondary Transporters:
The ABC transporters and the P-type ATPases are two families of proteinswhich use energy in the form of ATP to establish a transmembraneconcentration gradient. These are examples of primary transporters.
Another class of energy-requiring transporters are the secondarytransporters. This class uses energy in the form of a transmembraneconcentration gradient of one substance to transport another substance.
An example of a secondary transporter is the Na+-glucose symporter,which transfers glucose from low to high concentration (“uphill”)using the energy obtained by transferring Na+ from high to lowconcentration (“downhill”).
Low glucoseHigh Sodium
Low sodiumHigh glucose

Secondary Transporters
Secondary transporters use one concentration gradient to create another.One gradient is the “fuel” molecules, whereas the other is the “cargo”molecules.
Symporters transport the “fuel” and “cargo” molecules in the samedirection.
Antiporters transport the “fuel” and “cargo” molecules in oppositedirections.

Energetics of Glucose Transport
Glucose uptake by the Na+/Glucose symport uses the Na+ gradientestablished by the Na/K pump to power glucose uptake.
The total free energy change is the sum of the energies in the twotransport processes.
Glucose transport process:C1 = 1 mM, C2 = 66 mM, T = 310K , V = -50 mV G = +10.8 kJ/mol
Na+ transport process:C1 = 143 mM, C2 = 14 mM, T = 310K, V = -50 mV G = -10.8 kJ/mol
The net free energy change is close to zero.
The Na+ gradient “pays” for the glucose transport.

Passive Transport
Primary and secondary transporters use energy to transport ions or molecules “uphill” against a concentration gradient.
Passive transport proteins facilitate the rapid “downhill” flow of ions or molecules, from higher concentration to lower concentration.
Flow ceases when equilibrium is reached, namely when theG of transport is zero.
In the absence of an electrical potential, equilibrium occurswhen the concentrations on either side of the membrane are equal.
Passive transporters only change the rate of approach to equilibriumbut not the equilibrium itself.
An example of passive transporters are ion channels.

Ion Channels
Ion channels are proteins that enable rapid flows of ions through membranes,often at rates close to free diffusion in solution. Examples include Na+ channels,K+ channels, and Cl- channels.
Ion passage is regulated.Ligand-gated channels allow ion passage in response to binding of signalmolecules. Voltage-gated channels allow ion passage in response to changes in thetransmembrane electrical potential.
Ion channels are very selective about ion type.

Ion Channels Enable Nerve Cells to Transmit Electrical Impulses
Nerve Cell
In the resting state, nerve cells have an electrically polarized membrane(~-60 mV), with Na+ and K+ gradients established by the Na/K pump.
A nerve impulse is a propagating electrical disturbance of the membranepotential caused by the flow of ions across the membrane.
The membrane becomes temporarily permeable to ions when ionchannels open.
High K+ Low Na+-60 mV
High Na+ Low K+
Resting Firing Recovering
K+
Na+ (channels) (Na/K pump)
Na+
K+
Up to +30 mV(propagates)
Back to -60 mV

The Acetylcholine Receptor: A Ligand-Gated Channel
Nerve impulses are traverse synapses by causing fusion of vesicles containing neurotransmitters (such as acetylcholine) with the plasma membrane, releasing them into the synaptic cleft.
Acetylcholine diffuses across the synapse to the postsynaptic membrane, and binds to the acetylcholine receptor. The acetylcholine receptor is an ion channel which allows passage of Na+ and K+ across the membrane.
Binding of acetylcholine to theacetylcholine receptor opensthe channel and allows Na+ toflow into the cell and K+ to flowout. This depolarization triggersan action potential whichpropagates down the nerve cell.

Model for Structural Basis of Acetylcholine Receptor Action
The acetylcholine receptor has the quaternary structure 2 and the5 chains are arranged in a ring with approximate 5-fold symmetry,forming a pore at the center. Acetylcholine binds at the - and- interfaces and induces an allosteric shift between the closedand opened conformations. Transmembrane helices lining the poreare believed to rotate upon acetylcholine binding.
In the closed state, hydrophobic side-chains from these helicesocclude the pore.In the open state, the rotation of the helices shifts the hydrophobicside chains out of the way and causes the walls of the pore to be linedwith hydrophilic side-chains, facilitating ion transport.

Sodium and Potassium Channels: Voltage-Gated Channels
Sodium and Potassium channels in nerve cell membranes are sensitive tothe membrane potential. They open when the membrane is depolarized.The ions that flow through open channels further depolarize the membrane,propagating the disturbance down the nerve cell.
Na+ channels open first, initially causing the membrane potential to becomeless negative. These close spontaneously as the K+ channels open- thepassage of K+ ions in the opposite direction allows the membrane potentialto return to near its starting value.
Only a miniscule fraction of the Na+ and K+ ions in the cell actually cross themembrane. Small ion fluxes can change the transmembrane voltagevery effectively.

Relationships of Voltage-Gated Channels
Eukaryotic sodium channels consistof 4 pseudo-repeats, each of whichprobably contains 6 transmembranehelices.
Sequence similarities between sodium channels and potassium channelssuggest a similar structure. Bacterial potassium channels are simpler,containing only the pore-forming transmembrane helices 5 and 6. Thesesimpler molecules have proved suitable for structural studies.

Structure of the Bacterial Potassium Channel
The bacterial potassium channel forms a tetramer of subunits each containing2 transmembrane helices- the pore is formed at the center of the tetramer.A water-filled channel in the lower parts of the molecule can allow hydrated K+to enter. But a constriction at the top is too narrow for hydrated potassium to enter. K+ ions are desolvated (very unfavorable) during passage through theconstriction in the channel, but polar groups in the constriction regionprovide interactions which stabilize the desolvated ions.

SelectivityThe K+ channel is 100-fold more permeable to K+ ions than Na+ ions.The selectivity can be understood as a difference in stabilization ofdesolvated K+ and Na+ ions when passing through the constriction.
K+ ions are larger and the protein is capable of forming sufficient interactions with K+ that offset the energetic cost of breaking bonds to water, enabling K+ to pass through.
Na+ is smaller and the protein is not capable of forming sufficient interactions with Na+ to offset the larger energetic cost of desolvatingNa+, making it more difficult for Na+ to pass through.

Summary: Transmembrane concentration differences are not at equilibrium andtherefore are a source of energy which can be used for other purposes.
Two categories of active transporters are pumps, which use the energyof ATP hydrolysis to transport molecules across membranes, and secondary transporters, which use the energy stored in a transmembraneconcentration gradient.
Passive transporters facilitate diffusion of molecules through membranesand allow equilibrium to be more rapidly established. Ion channelsAre examples of passive transporters and play important roles innerve cell function.Key Concepts:Thermodynamics of transport
Active TransportPrimary transporters (pumps)Secondary transporters (antiporters, symporters)
Passive transportIon channelsLigand-gated and Voltage-gated ion channels