lecture 10 m230 feigon - ucla-doe...
TRANSCRIPT

Lecture 10 M230 Feigon• Sequential resonance assignments in
DNA (and RNA): homonuclear method• 2º structure determination
Reading resourcesEvans Chap 9

Why do NMR of nucleic acids?
More than a simple double helix!!
triplexesUnusual DNA structures quadruplexes
bent DNADrug-DNA complexesChemically modified DNA, e.g. cis-Pt (anti-cancer drug)Cation localizationProtein-DNA complexes
plus, the wonderful world of RNA!RibozymesPseudoknotsviral RNAs, e.g. HIV RNAsnRNAssnoRNAs & scaRNAsAptamersRiboswitchesmiRNAs, siRNAsTelomerase RNAlncRNAs and lincRNAsRNA-protein complexesEtc!
Kang, et al PNAS 2014

Nucleic acids different problem than proteins
Ø Only four bases, one sugar Simpler!Ø No through-bond connectivity between base and its
sugar (ribose or deoxyribose)Ø No through-bond connectivity from nucleotide to
nucleotide (except through P). You will see next week how through-bond connectivities in (13C, 15N labeled) protein backbone simplify protein assignments using triple-resonance experiments.
Ø Lower proton densitynucleotide is 3X MW of aa
Ø For helices, no long range constraints
More difficult!

1H NMR of Nucleic Acids: Assignments
1) Proton resonancesNon-exchangeable
DNA Base AH8, GH8AH2TH6, CH6CH5, TMe
Deoxyribose 1’, 2’, 2’’, 3’, 4’, 5’, 5’’RNA same, except UH5, UH6 (like CH5, CH6)
ribose, no 2’’ (2’ only)
Exchangeable (only seen in H2O)iminosamino
2) Proton spin systems- D2O only; no coupled labile spin systems (except aminos)
CH5-CH6, UH5-UH6 ~7 HzTMe-TH6 ⇐ 4 bond coupling ~1.5 Hz
deoxyriboseribose
aromatic
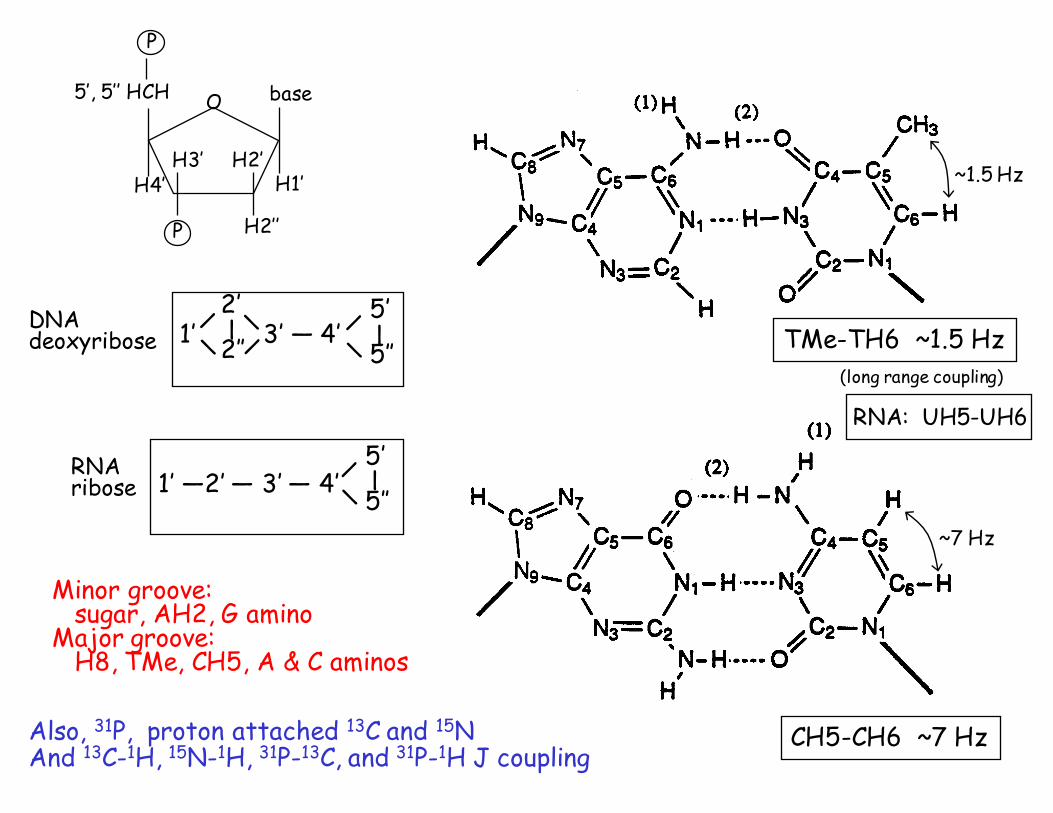
CH5-CH6 ~7 Hz
~7 Hz
~1.5 Hz
Minor groove:sugar, AH2, G amino
Major groove:H8, TMe, CH5, A & C aminos
TMe-TH6 ~1.5 Hz1’ 3’ — 4’ 2’
2’’
— 5’
5’’
—
5’
5’’1’ —2’ — 3’ — 4’
—
DNAdeoxyribose
RNAribose
O
H3’ H2’
base
H1’
H2’’
H4’
5’, 5’’ HCH
P
P
(long range coupling)
Also, 31P, proton attached 13C and 15NAnd 13C-1H, 15N-1H, 31P-13C, and 31P-1H J coupling
RNA: UH5-UH6

3) 1D spectra of a DNA dodecamer with N6A mod.A. Non-exchangeable
1) intensities Me 3x aromatics2) chemical shifts
-- see regions on spectrum3) coupling constants
-- CH5-CH6-- all sugar protons are coupled
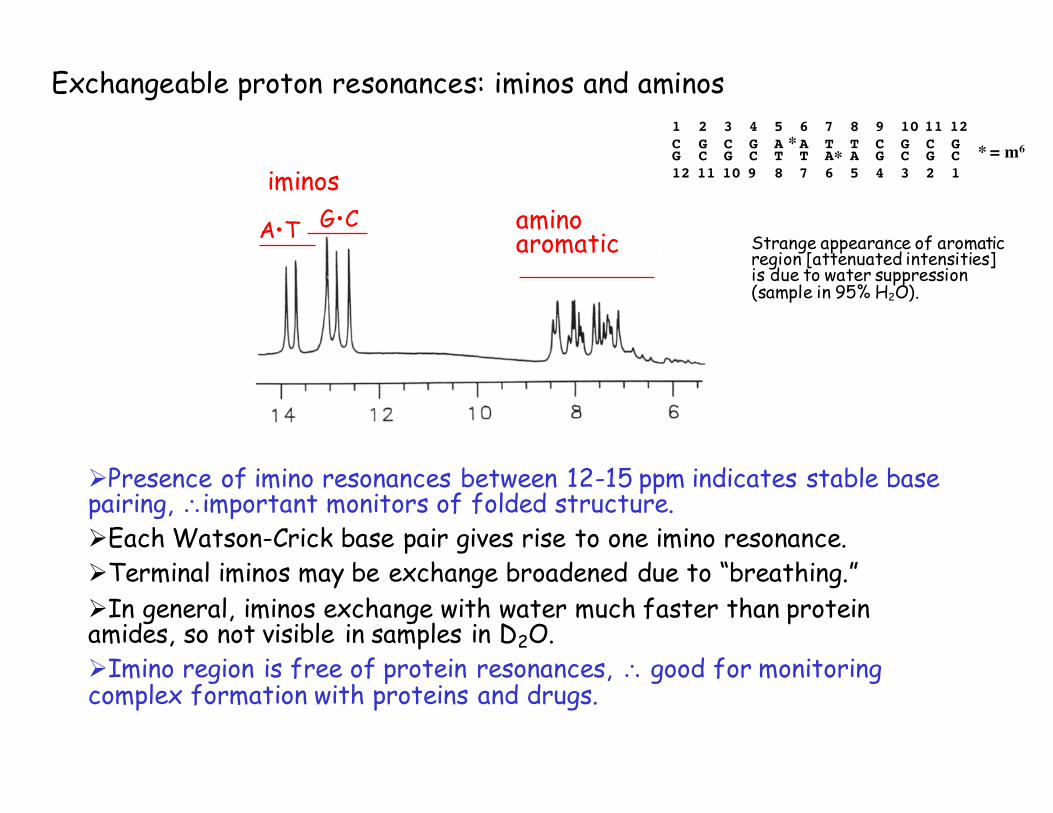
1 2 3 4 5 6 7 8 9 10 11 12 C G C G A A T T C G C GG C G C T T A A G C G C12 11 10 9 8 7 6 5 4 3 2 1
** * = m6
m6AHDO
aromaticH1’,H5
H3’
H4’,H5’H5”
H2’,H2”
TMe
H8H2
H6
* * **
1 2 3 4 5 6 7 8 9 10 11 12 C G C G A A T T C G C GG C G C T T A A G C G C12 11 10 9 8 7 6 5 4 3 2 1
** * = m6
iminos
A•T G•C aminoaromatic
Exchangeable proton resonances: iminos and aminos
ØPresence of imino resonances between 12-15 ppm indicates stable base pairing, ∴important monitors of folded structure.ØEach Watson-Crick base pair gives rise to one imino resonance.ØTerminal iminos may be exchange broadened due to “breathing.”ØIn general, iminos exchange with water much faster than protein amides, so not visible in samples in D2O.ØImino region is free of protein resonances, ∴ good for monitoring complex formation with proteins and drugs.
Strange appearance of aromatic region [attenuated intensities] is due to water suppression (sample in 95% H2O).

Assignment of spin systemsCOSY -- can identify all
CH5-CH6TMe-TH6H1’-H2’, H2’’, etc.
Theoretically, can isolate eachdeoxyribose spin system
but 5’, 5’’ usually difficult orimpossible due to overlap with 4’
1 2 3 4 5 6 7 8 9 10 11 12 [d( C G C G A A T T C G C G )]2*
1’ 3’ 4’2’ 5’
2’’ 5’’
TOCSY -- connects whole spin system,so can resolve more xpeaks,e.g. 2’, 2’’ - 5’, 5’’ ; 1’-3’
CH5
CH6
TMe
TH6
H1’
H2’, 2’’
H2’, 2’’
H3’
1’
3’
H4’,5’,5’’

aromatic H1’, CH5
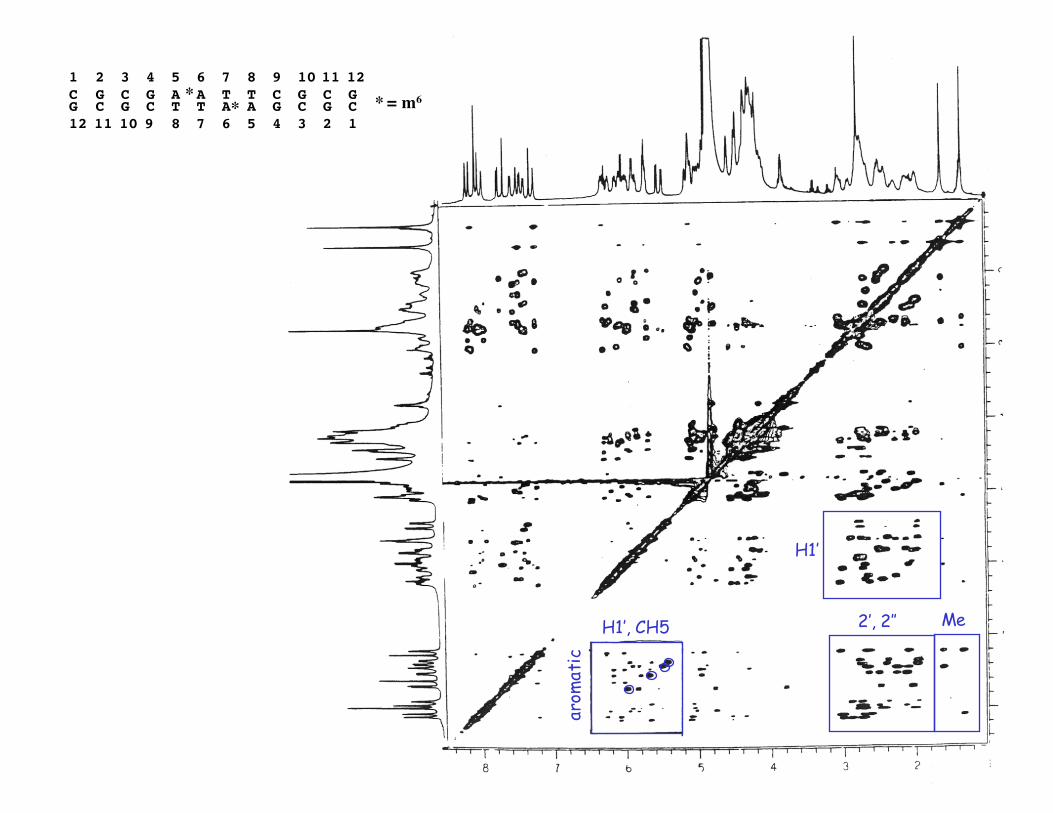
1 2 3 4 5 6 7 8 9 10 11 12 C G C G A A T T C G C GG C G C T T A A G C G C12 11 10 9 8 7 6 5 4 3 2 1
** * = m6
arom
atic
H1’, CH5 2’, 2’’ Me
H1’

⇒ Primarily relies on NOESY spectra(vs NOESY and COSY for 1H sequential assign. of proteins)
[For DNA, most of COSY information is also in NOESY sincecoupled protons generally also show NOEs]
Intensities of xpeaks will depend on sugar conformationbase H1’, 2’, 2’’
base H1’, 2’, 2’’
base H1’, 2’, 2’’
3’
5’B-DNA with anti glycosidic conformationclosest to intra 2’
inter 2’’but also directly (and indirectly) to others.
⇒ next page
∴ Can assign DNA sequentially by xpeaks from base to sugar to base...
NOEs expected base - H1’base - H2’, H2’’ → 2 eachH1’ - H2’, H2’’ → like COSY xpeaks
(next page)
Sequential Assignment of A- or B-DNA Helices
1) Base-sugar interactionsThe H6 (pyr) & H8 (pur) protons are close (within 4Å) to 1’, 2’, 2’’ protons on own base
(intranucleotide NOE) and to 1’, 2’, 2’’, protons on 5’ neighboring base (internucleotide NOE)

Sequential assignments of DNA (& RNA)
A
A
T

Sequential assignments of DNA (& RNA)
A
A
T

Sequential assignments of DNA (& RNA)
A
A
T

H1’
H6,
H8
*Note: This is two peaks.
T7AH2
C3C11T8
C9
AH2
G4G10G2, G12
A6A5
C1
↓
C1 →
1 2 3 4 5 6 7 8 9 101112C G C G A A T T C G C G

Ø Can also do same thing via base - H2’, H2’’ (as in schematic) but now have 4 xpeaks base → more complicated. Useful to double-check assignments (and exclude AH2s).Ø Assigned H1’ can be connected to rest of sugar spin system via COSY, TOCSY, and/or NOESY.∴ Have now assigned all H8, H6, H5, TMe, and sugar resonances (except 5’, 5’’ usually).
Note: NOEs to H1’ usually only show up clearly at long τm and are partially spindiffusion, so after assignments are made may want to use shorter τm forstructure determination.
2) The AH2 protons are usually far from any other protons and have few or no NOEs in D2O.
Often (usu. weak) NOEs to H1’ or AH2-AH2: these give information on local structure (minor groove width, propellering).
Assignment to specific base is done via NOE from iminos in H2O.
Also, 1H-13C HSQC

3) Base-base interactionsMost interactions in DNA are along strand, very
few cross strand NOEs
- pyr (CH5, TMe)⇓
- pur (H8) and pyr (H6)
Directional - goes 3’ → 5’ only
in N6MeA ‘Dickerson’dodecamer
See NOESY spectrumstrong weaker
1 2 3 4 5 6 7 8 9 10 11 12 C G C G A A T T C G C G
T8Me-T7Me T8Me-T7H6 C3H5 -G2H8T7Me-A6H8 C11H5-G10H8
C9H5 -T8H6in base-H1’
region
Note that if both strands were not symmetrical, would have to assign along each strand.
5’

1 2 3 4 5 6 7 8 9 10 11 12 C G C G A A T T C G C GG C G C T T A A G C G C12 11 10 9 8 7 6 5 4 3 2 1
** * = m6
arom
atic
H1’, CH5
⇑“fingerprint”
2’, 2’’ Me
H1’
T7T8

DNA conformation
1) a) Appearance of sequential base-sugar NOEs and base-base interactions indicates a right handed strand conformation.
b) Strong base-H2’, H2’’ and weaker base-H1’ NOEs indicates anticonformation of bases.
c) Intensity of base-base and base-sugar NOEs indicative of local conformation s.a. base tilt, twist, sugar pucker, groove width, etc.
2) Since there are few xstrand NOEs, double strand relies on:a) appearance of H-bonded imino resonances in H2Ob) melting studiesc) direct detection of H-bonds via scalar coupling (requires labeled
sample)
15N H 15N
~ -88 Hz ~2.5 Hz
~5-11 Hz
Dingley, et al Nature Protocols 3, 242 (2008)

DNA conformation3) Sugar conformation
a) Measure coupling constants via DQF-COSYb) Deduce sugar conformation from NOE intensities
B-DNA short distanceintra nucleotide H6,H8 → H2’inter nucleotide H6,H8 → H2’’
A-DNA (or RNA)intra nucleotide H6,H8-H3’; H4’-H2”inter nucleotide H6,H8-H2’; H6,H8-H3’
If use short enough τm, can distinguish.
C2’ endoJ1’, 2’ ~ 10 Hz
C3’ endoJ1’, 2’ ~ 1.5-3.3 Hz
2’
2’
1’
1’
2’’3’
P
4’
5’, 5’’ HCH
P
2’’
3’
P
4’
5’, 5’’ HCH
P
(S-type)
(N-type)

Why is RNA harder to study than DNA?
aromatic H1’, UH5CH5
H2’, H3’,H4’, H5’, H5’’ !
ØOverlap of ribose H2’, H3’, H4’, H5’, H5”!ØC and U base protons not distinguishable by J-coupling or chemical shift (since no Me on U)

Butcher, Allain, Feigon, Nature Struct. Biol. 6, 212-216 (1999)
U39
Hairpin Ribozyme Loop B: An irregular underwound helix with 7 non-Watson Crick base pairs
A40
A23
This structure only solvable with 13C, 15N labeled RNA