lactic acid production by lactobacillus ... vol 1-no 3/lactic acid...plications are the major use of...
TRANSCRIPT

ABS
TRAC
T
J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 201138
Hot-water extracted hemicelluloses from sugar maple wood chips were hydrolyzed by dilute acid at an elevated temperature and concen-trated using a nano-filtration membrane process to obtain a fermentable sugar stream containing arabinose, galactose, glucose, mannose, rhamnose, and xylose. Lactobacillus pentosus was directly adapted to using the concentrated wood extract hydrolysate to produce lactic acid. The effect of initial sugar loading was investigated by diluting the concentrated wood hydrolysate to obtain four sugar concentrations: 54.08 g/L, 61.47 g/L, 94.10 g/L, and 129.50 g/L; the media are labelled as med1, med2, med3, and med4 respectively. After 55 hours of fermentation in a 1-L bioreactor at 37°C and pH 6.0, medium med1 had the highest lactic acid yield (Yp/s) of 0.83 g-lactic acid/g-sugar, rep-resenting approximately 97.3% of theoretical yield. Acetic acid was produced after glucose was depleted as the main by-product at up to 49% of the obtained lactic acid concentration. Adaptation of an L. pentosus strain to concentrated wood-extract hydrolysate led to a 10-h reduction in fermentation time and a 15.5% increase in lactic acid production. L. pentosus simultaneously utilized both six-carbon and five-carbon sugars; arabinose, galactose, glucose, and rhamnose were preferably utilized, whereas mannose and xylose were slowly utilized. Acetic acid was also produced after glucose had been completely consumed.
JOHN P. BUYONDO, SHIJIE LIU*
LACTIC ACID PRODUCTION BY LACTOBACILLUS PENTOSUS FROM WOOD EXTRACT HYDROLYSATES
Lactic acid is an industrially important product with a large global market. World-wide production in 2005 (exclusive of polymers) was 100,000 tons, with a 15% annual growth rate [1]. Food-related ap-plications are the major use of lactic acid in the United States, accounting for ap-proximately 85% of the commercially produced product [2]. Lactic acid is used as a buffering agent, an acidic flavoring agent, an acidulant, and a bacterial inhibi-tor in many processed foods. Lactic acid and salts are preferred to other acids used in the food industry because they do not dominate other flavours and also act as preservatives [3,4]. Other applications of lactic acid include its use in the chemical industry for de-liming and metal etching and various uses in the pharmaceutical, textile, and cosmetics industries [4]. Cur-rently, the most important application of lactic acid may be its use as a monomer in the production of a biodegradable and biocompatible polymer, polylactic acid (PLA), which is an environmentally friendly alternative to non-biodegradable
plastics derived from petrochemicals. PLA is a thermo-plastic polyester, is 100% com-postable, and has a life cycle which could potentially reduce carbon dioxide levels in the Earth’s atmosphere.
Lactic acid can be produced by ei-ther microbial fermentation or chemical synthesis. Fermentation offers several ad-vantages over chemical synthesis, includ-ing low cost of substrates, low production temperature, low energy consumption, and high product specificity, meaning that fermentation yields optically pure L-(+)- and D-(−)- lactic acid, a desired stereoisomer [1,5,6]. Optically pure lactic acid is required for the production of the highly crystalline PLA which is suitable for commercial uses. These polymers have potential markets in commodity packag-ing, fabrication of prosthetic devices, and controlled delivery of drugs in humans [1,4]. PLA can also be used to enhance the strength properties of paper sheets made from bleached or unbleached Kraft pulp produced from hot-water extracted and unextracted sugar maple wood chips [7].
INTRODUCTIONIn commercial processes, glucose
and corn starches have been widely used as substrates for biological lactic acid production. However, this is economi-cally unfavourable because pure sugars have a higher economic value than the lactic acid produced [8]. For biologi-cal production of lactic acid to be fea-sible, cheap raw materials are necessary
*Contact: [email protected]
JOHN P. BUYONDO Department of Paper and Bioprocess Engineering, SUNY College of Environmental Science and Forestry, 1 Forestry Drive, Syracuse, NY, USA , 13210
SHIJIE LIUDepartment of Paper and Bioprocess Engineering, SUNY College of Environmental Science and Forestry, 1 Forestry Drive, Syracuse, NY, USA , 13210

J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 2011 39
SPECIAL BIOREFINERY ISSUE
to meet the large demand of polymer pro-ducers and other industrial users at a rela-tively low cost. Therefore, a cellulosic feed-stock (such as wood) should be used rather than refined substrates (glucose, sucrose, etc.) [9,10]. Lignocellulosic biomass offers a favourable alternative as a feedstock for the biological production of lactic acid be-cause it is readily available, has no compet-ing food value, and is less expensive than either cornstarch or sugars. Hemicellulose is the second most abundant carbohydrate source and accounts for 25%–35% of li-gnocellulosic material. Hemicelluloses are heterogeneous polymers made of pen-toses (D-xylose, D-arabinose), hexoses (D-mannose, D-glucose, D-galactose), and sugar acids [11,12]. Hemicelluloses in hardwood primarily contain xylans, while those in softwoods primarily contain glu-comannans [13]. Treatment of the hemi-cellulosic material with dilute acid at an elevated temperature leads to liberation of fermentable sugars [14]. However, in the
bioconversion of lignocellulosic sugars to lactic acid, product yield and volumet-ric productivity are generally low because of the presence of inhibitory compounds such as furfural, 5-hydroxymethyl furfural (HMF), and weak acids, which are gen-erated during the acid hydrolysis step [10,15,16]. Weak acids inhibit cell prolif-eration by uncoupling and intracellular anion accumulation, and furfural or HMF inactivates cell replication by inhibiting specific enzymes related to glycolysis or inhibitor reduction [16]. To improve the fermentation yields of wood hydrolysates, many methods for detoxifying inhibitory compounds have been investigated, in-cluding neutralization, over-liming, evapo-ration, the use of ion-exchange resins, and activated charcoal adsorption [15].
Bioconversion of hemicellulosic sugars to lactic acid requires a strain which is capable of fermenting sugar mixtures of hexoses and pentoses. Although much research has been focused on genetically
engineering strains that can efficiently utilize both five-carbon and six-carbon sugars [17,18], the recombinant cells in-volved have a tendency to become ge-netically unstable on repeated application.The versatility of Lactobacillus pentosus makes it a potential alternative to geneti-cally modified strains. L. pentosus ferments hexoses using the EMP (Embden-Meyer-hoff-Parnas) pathway (Fig. 1) and pentoses using the PK (phosphoketolase) pathway (Fig. 2) [19,20].
In this work, wood-extract hydro-lysate from sugar maple wood chips was concentrated by a nano-filtration mem-brane process to generate a mixture of five-and six-carbon hemicellulose sugars. The bacterial strain Lactobacillus pentosus was challenged by different concentra-tions of wood-extract hydrolysate (WEH) to produce lactic acid in a batch fermenta-tion process. The effect of adaptation of Lactobacillus pentosus cells to concentrated WEH was also examined.
Fig. 1 - Biochemical steps in the homolactic fermentation of glucose (redrawn based on Bustos et al. [18]).
Fig. 2 - Biochemical steps in the heterolactic fermentation of pentoses (redrawn based on Bailey and Ollis [21]).

23 J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 201140
MATERIALS AND METHODS
Production of Hemicellulose Hy-drolysate from Sugar Maple Wood ChipsSugar maple wood chips were extracted by hot water at approximately 160°C for 2 h. The hot-water extraction was carried out in a 1840.5-L digester with a wood-to-water ratio of about 1:4. The wood extract was concentrated approximately ten-fold using a nano-filtration mem-brane. The concentrated wood extract was hydrolyzed at 135°C for 25 min with 1.5% wt sulphuric acid added. After cooling to room temperature, acid-insoluble lignin was separated from the wood-extract hy-drolysate by centrifugation (CEPA high-speed centrifuge Z81G, cylinder speed 16,000 rpm, cylinder diameter 125 mm, New Brunswick Scientific, NJ, USA). The wood-extract hydrolysate was neutralized by calcium hydroxide at room temperature, with just enough calcium hydroxide added to neutralize the sulphuric acid. The neu-tralized hydrolysate (at a pH of approxi-mately 3.5 due to the high concentration of acetic acid) was again freed of solids by centrifugation. The solids-free hydrolysate was then diluted with fresh water and frac-tionated twice to remove low-molecular-weight substances by the nano-filtration membrane process. The membrane used in this study had a molecular weight cut-off of 100 g/mol [22].
Microorganisms and Seed Culture PreparationThe bacterial strain Lactobacillus pentosus (ATCC 8041) used was obtained from the American Type Culture Collection (ATCC). The strain was maintained on MRS agar medium slant and stored at 4°C. The strain was transferred to a fresh medium every three to four weeks. The MRS medium (Difco, Maryland, U.S.A.) contained 10.0 g/L proteose peptone 3, 10.0 g/L beef extract, 5.0 g/L yeast extract, 20.0 g/L dextrose, 1.0 ml/L Tween 80, 2.0 g/L ammonium citrate, 0.1 g/L MgSO4, 0.05 g/L MnSO4, 2.0 g/L K2HPO4, and 5.0 g/L CH3COONa. The
MRS medium was supplemented by 20 g/L agar to make slant. The seed culture was prepared by picking one or two large colonies from the slant and inoculating them into 50 mL MRS (55 g/L) medium contained in 125-mL screw-capped plas-tic flasks (NALGENE, Rochester NY, U.S.A.).
The effect of adapting the strain to concentrated wood-extract hydrolysate (WEH) before fermentation was also studied. Plated MRS solid medium supple-mented with 10% WEH was inoculated with two or three large colonies picked from MRS medium slants and incubated at 37°C for 24 hours. A seed culture of the adapted strain was prepared by pick-ing one or two large colonies from an MRS-WEH medium plate and inoculating them into 50-mL MRS medium supple-mented with 20% (v/v) WEH. The seed culture was incubated at 37°C for 15–24 h on a rotary shaker (GYROMAXTM 747R, Amerex Instruments, Lafayette CA, U.S.A.), operating at 150 rpm. The media was sterilized by autoclaving (HIRAYA-MA HICLAVETM HV-110, Amerex In-struments, Lafayette CA, U.S.A.) at 121°C for 20 min.
Batch FermentationBatch fermentation experiments were conducted in a 1.0-L New Brunswick bio-reactor (BIOFLO 110; New Brunswick Scientific, New Brunswick NJ, U.S.A.) with an 800-mL working volume. The desired wood hydrolysate concentration was obtained by diluting the concentrated wood hydrolysate with the appropriate volume, V, of distilled water. The fermen-tation medium contained wood-extract hydrolysate supplemented by 10 g/L yeast extract, 2 g/L K2HPO4, 2 g/L KH2PO4, and 0.5% (v/v) Tween 80. All medium components were purchased from Fisher Scientific, Pittsburgh PA, U.S.A. Wood-ex-tract hydrolysate was sterilized by filtration through a 0.22-µm sterile filter (nitrocellu-lose membrane, Millipore) which was held on a filter holder with a receiver (1000 mL, NALGENE, Rochester NY, U.S.A.). The fermentation-medium nutrient sup-
plements were dissolved in the desired amount of distilled water in a bioreac-tor and autoclaved at 121°C for 20 min. After cooling to room temperature, an amount of filtered sterilized concentrated wood-extract hydrolysate was added to the bioreactor and the pH adjusted to 6.0 before inoculation. The bioreactor was in-oculated with 40 mL of actively growing 15–24 h-old seed culture and incubated at 37°C. Agitation speed was set to 2.5 Hz, and air flow rate was set to 25 mL/min. The pH was maintained at 6.0 by dosing with a 5 mol/L NaOH solution. Two par-allel fermentations were performed, one with native seed culture, and the other with adapted seed culture. Samples (2 mL) were taken at given fermentation times and centrifuged at 4000 rpm for 5 min. The supernatants were stored at -10°C for sugar, lactic acid, and acetic acid analyses. Experimental data were obtained in trip-licate, and the values reported represent sample means.
Analytical Methods Lactic acid, acetic acid, and inhibitor com-pound concentrations were determined by 600-MHz 1H-NMR, whereas sugar con-centrations were determined by 600-MHz 13C-NMR (Bruker BioSpin Corporation, Billerica MA, U.S.A.). After centrifugation, samples were prepared by mixing 100 μL sample and 100 μL internal standard (glu-cosamine, Sigma-Aldrich, St. Louis MO, U.S.A.) with 800 μL D2O. The NMR op-erating conditions were as follows: probe type: Broadband Observe Probe; tem-perature: 30°C; 90° pulse: 11 μs; interval between pulses: 10 s; acquisition interval: 2.73 s; sweep width: 10 ppm; center of spectrum: 4.5 ppm; reference: acetone at 2.2 ppm. A calibration curve was gener-ated by plotting peak/area ratios of indi-vidual standard solutions and the internal standard. Peak/area ratios (analyte/inter-nal standards) were compared with stan-dard curves to quantify the analytes. The area of each peak was integrated using the MestReNova software, and the integral of glucosamine was set to 100. Lactic acid yield was defined as the amount of acid
J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 2011 41
SPECIAL BIOREFINERY ISSUE
produced divided by the total sugars uti-lized.
RESULTS AND DISCUSSION
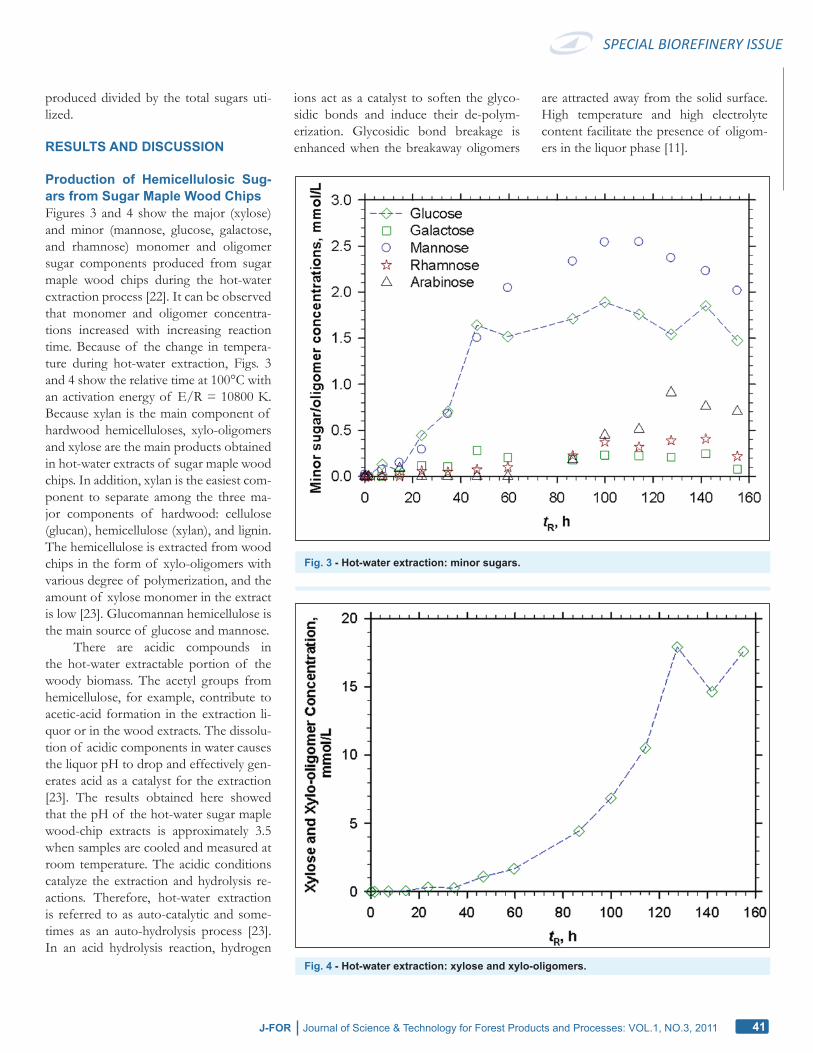
Production of Hemicellulosic Sug-ars from Sugar Maple Wood ChipsFigures 3 and 4 show the major (xylose) and minor (mannose, glucose, galactose, and rhamnose) monomer and oligomer sugar components produced from sugar maple wood chips during the hot-water extraction process [22]. It can be observed that monomer and oligomer concentra-tions increased with increasing reaction time. Because of the change in tempera-ture during hot-water extraction, Figs. 3 and 4 show the relative time at 100°C with an activation energy of E/R = 10800 K. Because xylan is the main component of hardwood hemicelluloses, xylo-oligomers and xylose are the main products obtained in hot-water extracts of sugar maple wood chips. In addition, xylan is the easiest com-ponent to separate among the three ma-jor components of hardwood: cellulose (glucan), hemicellulose (xylan), and lignin. The hemicellulose is extracted from wood chips in the form of xylo-oligomers with various degree of polymerization, and the amount of xylose monomer in the extract is low [23]. Glucomannan hemicellulose is the main source of glucose and mannose.
There are acidic compounds in the hot-water extractable portion of the woody biomass. The acetyl groups from hemicellulose, for example, contribute to acetic-acid formation in the extraction li-quor or in the wood extracts. The dissolu-tion of acidic components in water causes the liquor pH to drop and effectively gen-erates acid as a catalyst for the extraction [23]. The results obtained here showed that the pH of the hot-water sugar maple wood-chip extracts is approximately 3.5 when samples are cooled and measured at room temperature. The acidic conditions catalyze the extraction and hydrolysis re-actions. Therefore, hot-water extraction is referred to as auto-catalytic and some-times as an auto-hydrolysis process [23]. In an acid hydrolysis reaction, hydrogen
ions act as a catalyst to soften the glyco-sidic bonds and induce their de-polym-erization. Glycosidic bond breakage is enhanced when the breakaway oligomers
are attracted away from the solid surface. High temperature and high electrolyte content facilitate the presence of oligom-ers in the liquor phase [11].
Fig. 3 - Hot-water extraction: minor sugars.
Fig. 4 - Hot-water extraction: xylose and xylo-oligomers.

23 J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 201142
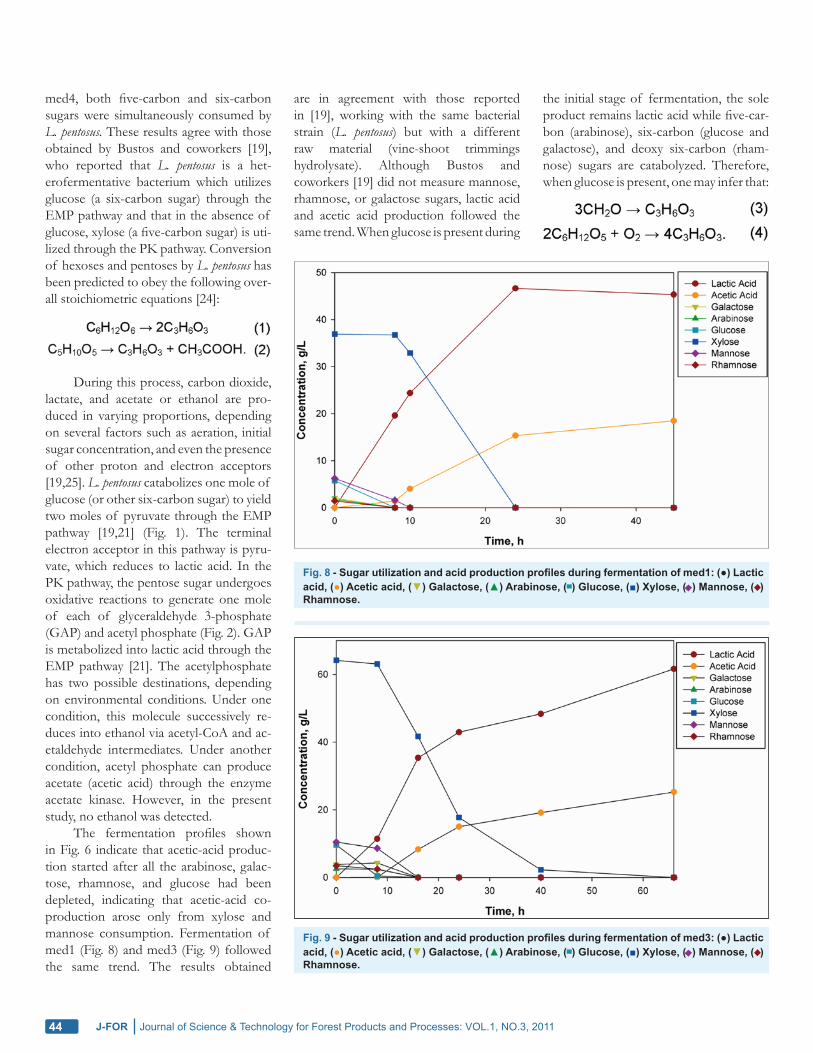
Production of Fermentable Hemicel-lulosic SugarsHot-water extract from sugar maple wood chips was hydrolyzed by 1.5% (wt.) con-centrated sulphuric acid at elevated tem-perature (135°C) to obtain fermentable hemicellulosic sugars. After cooling to room temperature, the acidic (pH 0.5) wood-extract hydrolysate was stoichio-metrically neutralized (with respect to added sulphuric acid) with calcium hy-droxide to approximately pH 3.5. The acid hydrolysis of hot-water extract from sugar maple wood chips led to simulta-neous generation of hemicellulosic sug-ars and fermentation inhibitors such as acetic acid, formic acid, furfural, and hy-droxymethyl furfural (HMF). Upon nano-filtration membrane processing, water and other low-molecular-weight compounds such as acetic acid, formic acid, HMF, and furfural were preferentially sent to the permeate stream, while sugar monomers and oligomers along with high-molecular-weight compounds were concentrated in the concentrate stream [15]. Figure 5 shows the 2D NMR spectrum for the ob-tained mixture of hemicellulosic sugars in the sugar maple wood-extract hydrolysate. The beta and alpha sugar components are located between the 5.0–5.3 ppm and 4.4–4.95 ppm chemical shifts respectively. The concentration of the respective sug-ars was obtained by determining the aver-age of the concentration of the beta and alpha sugar components.
After membrane separation, the concentrations of sugars in the wood hy-drolysate were 123 g/L xylose, 21.9 g/L mannose, 16.5 g/L glucose, 11.5 g/L ga-lactose, 5.52 g/L arabinose, and 4.13 g/L rhamnose. Here xylose, a five-carbon sugar, is the dominant component of the wood-extract hydrolysate. Arabinose is another five-carbon sugar. Mannose, glucose, and galactose are six-carbon sug-ars, while rhamnose is a deoxy six-carbon sugar. The ratio of glucose to mannose is close to one, which suggests that their main origin was glucomannan [23]. No formic acid, furfural, or HMF was de-tected in the concentrated wood-extract hydrolysate, and the concentration of
acetic acid was significantly reduced (Table 1).
Fermentation of Wood-Extract Hydrolysate:Preparation of Fermen-tation Medium To investigate the possible effect of wood-extract hydrolysate (WEH) con-centration on the production of lactic acid, concentrated WEH was diluted with distilled water. Fermentation media were prepared by adding different amounts of distilled water to a known volume of concentrated WEH (mL) to obtain different sugar concentrations (Table
2). The main sugar component of thefermentation media was xylose, contribut-ing about 68% of the total sugar concen-tration. This agrees with the results pre-sented in the literature by Liu et al. [15] and Amidon et al. [23] that xylose is the main component of sugar maple wood-extract hydrolysate.
Lactic Acid Fermentation The seed culture (40 mL) was inoculated into the prepared fermentation medium, and the batch fermentation reaction was set to run for 55 hours. Unlike many other Lactobacillus strains, L. pentosus is
Fig. 5 - 2D NMR of standard mixture of monosaccharides.
TABLE 1 Concentrations of different inhibitor compounds in WEH before and after membrane fi ltration.
Concentration, g/LInhibitor compound
38.86Acetic acidFormic acidHMFFurfural
Before After2.17
N/A N/A0.27 N/A3.12 N/A

J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 2011 43
SPECIAL BIOREFINERY ISSUE
SO2 present in the evaporator condensate might reduce the efficiency of the bio-methane production process. To address this problem, the authors have recently used HFC technology to remove SO2 from sulphite mill evaporator condensates. Pilot trial results on the use of this technol-ogy indicated that 50%–93% of the SO2 can be removed, depending on operating
capable of metabolizing both five-carbon and six-carbon sugars. Therefore, all sug-ars in the fermentation media were po-tential carbon sources for microorganism growth and lactic acid production. Acetic acid was produced as the main by-product. As shown in Fig. 6, during fermentation of med2, arabinose, glucose, rhamnose, and galactose were utilized within the first eight hours, while utilization of mannose and xylose started after eight hours. Man-nose was consumed in the next 12 hours, and xylose was utilized only slowly.
In the fermentation of med4 (Fig. 7), arabinose, glucose, rhamnose, and galac-tose were utilized within the first 24 hours, while utilization of mannose and xylose started after 10 hours. In this case, man-nose was consumed in the next 14 hours, and xylose was utilized only slowly. After 55 hours of fermentation, all sugars con-tained in med2 were completely consumed, while in the case of med4, 118.11 g/L to-tal sugars were consumed, and 10.39 g/L xylose remained unconsumed.
The highest lactic acid concentration obtained was 43.66 g/L after 55 hours’ fermentation of med2, with a productivity of 0.99 g/(L.h) and a product yield (YP/S) of 0.72 g lactic acid /g total sugar con-sumed. Acetic acid (21.60 g/L) was pro-duced as the main by-product. Figures 6 to 9 show that the production of acetic acid became pronounced after glucose had been exhausted. In the fermentation of med4 (Fig.7), 59.62 g/L lactic acid was produced after 55 hours of fermentation, with a productivity of 1.36 g/(L.h) and a product yield (YP/S) of 0.50 g-lactic acid /g- total sugar consumed. The concentra-tion of the acetic acid produced was 21.13 g/L.
During fermentation of med2 and
Fig. 6 - Sugar utilization and acid production profiles during fermentation of med2: (●) Lactic acid, (●) Acetic acid, (▼) Galactose, (▲) Arabinose, (▄) Glucose, (■) Xylose, ( ) Mannose, ( ) Rhamnose.
TABLE 2 Hemicellulose sugar composition of prepared fermentation media.
med1med2med3med4
Medium WEH,mL
V,mL
Xylose,g/L
Mannose,g/L
Glucose,g/L
Galactose,g/L
Arabinose,g/L
Total sugars,g/L
Rhamnose,g/L
300400500600
500400300200
36.9041.2264.1586.34
6.226.91
10.4715.54
5.756.339.62
12.53
2.043.243.836.98
1.731.862.544.14
1.441.913.483.98
54.0861.4794.10
129.50
Fig. 7 - Sugar utilization and acid production profiles during fermentation of med4: (●) Lactic acid, (●) Acetic acid, (▼) Galactose, (▲) Arabinose, (▄) Glucose, (■) Xylose, ( ) Mannose, ( ) Rhamnose.
23 J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 201144
med4, both five-carbon and six-carbon sugars were simultaneously consumed by L. pentosus. These results agree with those obtained by Bustos and coworkers [19], who reported that L. pentosus is a het-erofermentative bacterium which utilizes glucose (a six-carbon sugar) through the EMP pathway and that in the absence of glucose, xylose (a five-carbon sugar) is uti-lized through the PK pathway. Conversion of hexoses and pentoses by L. pentosus has been predicted to obey the following over-all stoichiometric equations [24]:
During this process, carbon dioxide, lactate, and acetate or ethanol are pro-duced in varying proportions, depending on several factors such as aeration, initial sugar concentration, and even the presence of other proton and electron acceptors [19,25]. L. pentosus catabolizes one mole of glucose (or other six-carbon sugar) to yield two moles of pyruvate through the EMP pathway [19,21] (Fig. 1). The terminal electron acceptor in this pathway is pyru-vate, which reduces to lactic acid. In the PK pathway, the pentose sugar undergoes oxidative reactions to generate one mole of each of glyceraldehyde 3-phosphate (GAP) and acetyl phosphate (Fig. 2). GAP is metabolized into lactic acid through the EMP pathway [21]. The acetylphosphate has two possible destinations, depending on environmental conditions. Under one condition, this molecule successively re-duces into ethanol via acetyl-CoA and ac-etaldehyde intermediates. Under another condition, acetyl phosphate can produce acetate (acetic acid) through the enzyme acetate kinase. However, in the present study, no ethanol was detected.
The fermentation profiles shown in Fig. 6 indicate that acetic-acid produc-tion started after all the arabinose, galac-tose, rhamnose, and glucose had been depleted, indicating that acetic-acid co-production arose only from xylose and mannose consumption. Fermentation of med1 (Fig. 8) and med3 (Fig. 9) followed the same trend. The results obtained
are in agreement with those reported in [19], working with the same bacterial strain (L. pentosus) but with a different raw material (vine-shoot trimmings hydrolysate). Although Bustos and coworkers [19] did not measure mannose, rhamnose, or galactose sugars, lactic acid and acetic acid production followed the same trend. When glucose is present during
the initial stage of fermentation, the sole product remains lactic acid while five-car-bon (arabinose), six-carbon (glucose and galactose), and deoxy six-carbon (rham-nose) sugars are catabolyzed. Therefore, when glucose is present, one may infer that:
Fig. 8 - Sugar utilization and acid production profiles during fermentation of med1: (●) Lactic acid, (●) Acetic acid, (▼) Galactose, (▲) Arabinose, (▄) Glucose, (■) Xylose, ( ) Mannose, ( ) Rhamnose.
Fig. 9 - Sugar utilization and acid production profiles during fermentation of med3: (●) Lactic acid, (●) Acetic acid, (▼) Galactose, (▲) Arabinose, (▄) Glucose, (■) Xylose, ( ) Mannose, ( ) Rhamnose.

J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 2011 45
SPECIAL BIOREFINERY ISSUE
Because of apparent acetic-acid pro-duction and the absence of ethanol pro-duction from the mixture of mannose and xylose at the end stage of fermentation, one may infer that the six-carbon sugars follow the EMP pathway while the five-carbon sugars follow PK pathways when glucose is not present.
In both med2 and med4, L. pentosus preferred to utilize arabinose (five-car-bon), glucose (six-carbon), rhamnose (six-carbon deoxy), and galactose (six-carbon) sugars first, before mannose (six-carbon) and xylose (five-carbon). In the case of med2, mannose and xylose utilization started after complete utilization of other sugars, while in med4, mannose and xy-lose utilization started after 10 hours of fermentation. Fermentation of med4 with higher initial glucose concentration (12.53 g/L) yielded a higher average rate of sugar utilization (2.68 g.L-1.h-1) than fermenta-tion of med2 with a lower initial glucose concentration (6.33 g/L), which yielded a sugar utilization rate of 1.37 g.L-1.h-1. Com-pared to glucose, the conversion of xylose requires additional enzymatic steps. Some enzymes are inducible; therefore, there is a lag time before the enzymes required for assimilation appear when cells are ex-posed to xylose [26]. It has also been pro-posed that an increase in xylose concen-tration will result in increased intracellular concentrations of intermediates such as fructose 1,6-diphosphate, which inhibits enzyme activity during xylose metabolism [27]. Nevertheless, the ability of L. pento-sus to utilize five-carbon and six-carbon sugar mixtures simultaneously makes it a potential microorganism for use in indus-trial fermentation of wood-extract hydro-lysate.
Effect of Sugar Loading on Lactic Acid ProductionThe effect of sugar loading on the pro-duction of lactic acid was studied (Fig. 10). Fermentation media containing 54.08 g/L sugars (med1), 61.47 g/L sug-ars (med2), 94.10 g/L sugars (med3), and 129.50 g/L sugars (med4) were used. As shown in Fig. 7, the final lactic
acid concentration increased with an in-crease in sugar concentration in the fer-mentation medium. The maximum lactic acid concentration (65 g/L) was obtained after 106 hours of fermentation of med4. However, the yield was higher with fer-mentation media containing lower con-centrations of WEH.
The med1 medium had the highest product yield, 0.83 g-lactic acid/g-sugar, followed by med2 (0.72 g/g), med3 (0.65 g/g), and med4 (0.50 g/g). In addition, the time required for complete fermentation (or complete substrate utilization) gener-ally increased with an increase in hydro-lysate loading. Fermentation media (med1 and med2) which contained lower WEH concentrations required a shorter fermen-tation time to attain maximum lactic acid production than media with higher con-centrations (med3 and med4). The low lactic-acid yields at higher WEH loading could be attributed largely to product in-hibition because known microbial inhibi-tor compounds (furfural, HMF, and acetic acid) had been removed from WEH dur-ing the nano-filtration membrane process.
This result is in good agreement withreports in the literature [19,28] that prod- uct inhibition leads to decreases in fermen-tation rate and in microbial growth rate.
In all fermentations, acetic acid was produced as the main by-product. Acetic-acid production followed the same trend as lactic-acid production. Although there was no significant difference in the acetic-acid production for media containing dif-ferent levels of WEH, generally acetic acid concentration increased with an increase in sugar concentration in the fermentation medium. The med4 fermentation medium produced the highest acetic-acid con-centration, 23.90 g/L, followed by med3 (22.61 g/L), med2 (21.61 g/L), and med1 (18.47 g/L). Acetic acid was produced mainly when mannose and xylose were the only remaining fermentable sugars. This observation indicates that in the presence of glucose, L. pentosus metabolizes both five-and six-carbon sugars to produce lac-tic acid exclusively, whereas in the absence of glucose, it metabolizes five-carbon sug-ars through the PK pathway to produce both lactic acid and acetic acid.
Fig. 10 - Effect of initial sugar concentrations in wood hydrolysate-based medium on lactic acid production by L. pentosus.

23 J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 201146
Adaptation of L. pentosus to WEH for Lactic Acid ProductionThe effect of adaptation of seed-culture cells to concentrated WEH was studied. Generally, fermentation of the adapted L. pentosus strain on WEH-containing me-dia showed higher lactic-acid yields and higher productivity than WEH fermenta-tions performed with a native L. pentosus strain (Table 3). The adapted L. pentosus strain also required less time to achieve maximum lactic-acid production than the native strain.
With the adapted L. pentosus strain, batch fermentation of med2 (containing 50% WEH (v/v)) led to a 15.5% increase in lactic-acid production, compared to a 2.0% increase in lactic-acid production in med4 (containing 75% WEH (v/v)). These results show that even with a WEH-adapt-ed L. pentosus strain, fermentation of lactic acid from a medium containing high levels of WEH is affected by product inhibition, which is in agreement with the observa-tions made earlier in this paper.
Although wood-extract hydrolysate contains high concentrations of mixed sugars including xylose, mannose, glucose, arabinose, and rhamnose, little research has been reported in which woody bio-mass has been used as a carbon source for lactic-acid production. In the present study, batch fermentation of different WEH-containing media led to high lactic-acid production levels ranging between 43.2 g/L and 65.02 g/L and representing 0.83 g/g and 0.54 g/g product yield respective-ly. The lactic-acid production and yield val-ues obtained are higher than most values that have been reported in the literature. For example, Walton and co-workers [29] used the bacterial strain Bacillus coagulans MXL-9 to obtain 33 g/L lactic acid and a yield of 0.73 g/g from wood hemicel-lulose extracts; Bustos and coworkers [19] reported 46.0 g/L lactic-acid concentra-tion and yield of 0.78 g/g from vine-trim-ming hydrolysate using a L. pentosus strain. Other research results that have been reported include lactic-acid production with a yield of 0.82 g/g from acid-treated softwood hydrolysates using L. casei subsp. Rhamnous strain [28] and lactic-acid prod-
uction from wheat-straw hemicellulose hydrolysates at a concentration of 9 g/L (representing a yield of 0.90 g/g) using L. pentosus and 10 g/L (representing a yield of 0.61 g/g ) using L. brevis [30].
CONCLUSIONSThis study has demonstrated that batch fermentation of sugar maple wood-extract hydrolysate-based media using L. pentosus yields high productivity of lactic acid. The strain simultaneously utilized hexose and pentose sugars in the fermentation media. In the first 12 hours of fermentation, the strain preferably utilized arabinose, glucose, rhamnose, and galactose, while utilization of mannose and xylose started after 12 h, with mannose being consumed in the next 12 h. However, L. pentosus growth rate and product yield were inhibited by high concentrations of lactic acid produced during fermentation. Although use of an L. pentosus strain adapted to concentrated WEH led to significant increases in lactic-acid productivity and yield for fermentation media containing lower concentrations of WEH, there was no significant effect on product yields with fermentation media containing high WEH concentrations. The low lactic-acid yield could be attributed largely to product inhibition because known microbial inhibitors contained in wood hydrolysate had been removed by a nano-filtration process. The main by-product, acetic acid, is produced after glucose has been depleted. Acetic-acid production is minimal when glucose is present in the sugar mixture.
ACKNOWLEDGEMENTSThe authors are grateful for research sup-port from the Biorefinery Research Insti-tute and the Department of Paper and Bioprocess Engineering. Thanks also to Dr. Thomas E. Amidon, Mr. Dave Kiem-le, and Mr. Christopher D. Wood for their help with these studies.
REFERENCES
Jin, B., Yin, P., Ma, Y., and Zhao, L., “Production of Lactic Acid and Fun-gal Biomass by Rhizopus Fungi from Food Processing Waste Streams”, Journal of Industrial Microbiology and Biotechnology 32(11-12):678-686 (2005). Garlotta, D., “A Literature Review of Poly (Lactic Acid)”. Journal of Poly-mers and the Environment 9(2): 63-84 (2001). Monteagudo, J.M., Rodríguez, L., Rincón, J., and Fuertes, J., “Kinetics of Lactic Acid Fermentation by Lac-tobacillus Delbrueckii Grown on Beet Molasses”. Journal of Chemical Tech-nology and Biotechnology 68(3):271-276 (1997). Schmidt, S. and Padukone, N., “Pro-duction of Lactic Acid from Wastepa-per as a Cellulosic Feedstock”. Journal of Industrial Microbiology and Bio-technology 18(1):10-14 (1997). John, R.P., Nampoothiri, K.M., and Pandey, A., “Fermentative Produc-tion of Lactic Acid from Biomass: An Overview of Process Developments and Future Perspectives”. Applied Microbiology and Biotechnology
1.
2.
3.
4.
5.
TABLE 3 Fermentation of concentrated WEH using native and adapted Lactobacillus pentosus strains.
Maximum lactic-acid concentration, g/L
Parameter Strain
Native 43.66med4med2
Fermentation medium
AdaptedNative
AdaptedNative
AdaptedNative
Adapted
Time required to attain maximum lactic-acid production, h
Lactic-acid yield, g/g
Lactic-acid productivity, g/(L×h)
50.455545
0.720.780.991.15
62.4563.96
7552
0.500.541.041.53

J-FOR Journal of Science & Technology for Forest Products and Processes: VOL.1, NO.3, 2011 47
SPECIAL BIOREFINERY ISSUE
74(3):524-534 (2007). Rao, C.S., Prakasham, R.S., Rao, A.B., and Yadav, J.S., “Production of L (+) Lactic Acid by Lactobacillus Delbrueckii Immobilized in Functionalized Al-ginate Matrices”. World Journal of Microbiology and Biotechnology 24(8):1411-1415 (2008). Hasan, A., Bujanovic, B., and Amidon, T., “Strength Properties of Kraft Pulp Produced from Hot-Water Extracted Woodchips Within the Biorefinery”. Journal of Biobased Materials and Bioenergy 4(1):46-52 (2010). Hofvendahl, K. and Hahn-Hägerdal, B., “Factors Affecting the Fermen-tative Lactic Acid Production from Renewable Resources”. Enzyme and Microbial Technology 26(2-4):87-107 (2000). Schmidt, S. and Padukone, N., “Pro-duction of Lactic Acid from Wastepa-per as a Cellulosic Feedstock”. Journal of Industrial Microbiology and Bio-technology 18(1):10-14 (1997). Sreenath, H.K., Moldes, A.B., Koe-gel, R.G., and Straub, R.J., “Lactic Acid Production from Agriculture Residues”. Biotechnology Letters 23(3):179-184 (2001). Liu, S.J., “Woody Biomass: Niche Po-sition as a Source of Sustainable Re-newable Chemicals and Energy and Kinetics of Hot-Water Extraction/Hydrolysis”. Biotechnology Advances 28(5):563-582 (2010). Kumar, R., Singh, S., and Singh, O.V., “Bioconversion of Lignocellulosic Biomass: Biochemical and Molecu-lar Perspectives”. Journal of Indus-trial Microbiology and Biotechnology 35(5):377-391 (2008). Saha, B.C., “Hemicellulose Biocon-version”. Journal of Industrial Micro-biology and Biotechnology 30(5):279-291 (2003). Hu, R., Lin, L., Liu, T.J., and Liu, S.J., “Dilute Sulfuric Acid Hydrolysis of Sugar Maple Wood Extract at Atmo-spheric Pressure”. Bioresource Tech-nology 101(10):3586-3594 (2010). Liu, T.J., Lin, L., Sun, Z., Hu, R., and Liu, S.J., “Bioethanol Fermenta-
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
tion by Recombinant E. Coli FBR5 and its Robust Mutant FBHW Using Hot-Water Wood Extract Hydrolyzate as Substrate”. Biotechnology Advanc-es 28(5):602-608 (2010). Palmqvist, E. and Hahn-Hägerdal, B., “Fermentation of Lignocellulosic Hydrolysates. I: Inhibition and De-toxification”. Bioresource Technol-ogy 74(1):17-24 (2000). Dien, B.S., Nichols, N.N., and Both-ast, R.J., “Fermentation of Sugar Mix-tures using Escherichia Coli Catabolite Repression Mutants Engineered for Production of L-Lactic Acid”. Jour-nal of Industrial Microbiology and Biotechnology 29(5):221-227 (2002). Picataggio, S.K., Zhang, M., Franden, M.A., McMillan, J.D., and Finkelstein, M., “Recombinant Lactobacillus for Fermentation of Xylose to Lactic Acid and Lactate”. U.S. Patent 5,798,237 (issued August 25, 1998). Bustos, G., Moldes, A.B., Cruz, J.M., and Domínguez, J.M., “Influence of the Metabolism Pathway on Lactic Acid Production from Hemicellulosic Trimming Vine Shoots Hydrolyz-ates Using Lactobacillus Pentosus”. Bio-technology Progress 21(3):793-798 (2005). Krueger, K.K. and Peterson, W.H., “Metabolism of Biotin and Oxybiotin by Lactobacillus pentosus 124-2”. Jour-nal of Bacteriology 55(5): 693-703 (1948). Bailey, E.J. and Ollis, F.D., Biochemi-cal Engineering Fundamentals. 2nd ed., McGraw-Hill, New York (1986). Liu, S.J., Lu, H., Hu, R., Shupe, A., Lin, L., and Liang, B., “A Sustainable Woody Biomass Biorefinery”. Bio-technology Advances. In press, DOI http://dx.doi.org/10.1016/j.bio-techadv.2012.01.013 (2012).Amidon, T.E., Wood, C.D., Shupe, A.M., Wang, Y., Graves, M., and Liu, S.J., “Biorefinery: Conversion of Woody Biomass to Chemicals, Energy, and Materials”. Journal of Biobased Materials and Bioenergy 2(2):100-120 (2008).
16.
17.
18.
19.
20.
21.
22.
23.
Lampen, J.O. and Peterjohn, H.R., “Studies on the Specificity of the Fer-mentation of Pentoses by Lactobacil-lus Pentosus”. Journal of Bacteriology 62(3): 281-292 (1951). Martínez-Anaya, M.A., Llin, M.L., Macías, M.P., and Collar, C., “Regu-lation of Acetic Acid Production by Homo- and Heterofermentative Lacto-bacilli in Whole-Wheat Sour-Doughs”. Zeitschrift für Lebensmittel-Untersu-chung und -Forschung, 199(3):186-190 (1994). Tanaka, K., Komiyama, A., Sonomo-to, K., Ishizaki, A., Hall, S., and Stan-bury, P., “Two Different Pathways for D-Xylose Metabolism and the Effect of Xylose Concentration on the Yield Coefficient of L-Lactate in Mixed-Acid Fermentation by the Lactic Acid Bacterium Lactococcus lactis IO-1”. Ap-plied Microbiology and Biotechnol-ogy 60(1-2):160-167 (2003). Wang, L., Zhao, B., Liu, B., Yu, B., Ma, C., Su, F., Hua, D., Li, Q., Ma, Y., and Xu, P., “Efficient Production of l-Lactic Acid from Corncob Molasses, a Waste By-Product in Xylitol Pro-duction, by a Newly Isolated Xylose Utilizing Bacillus Sp. Strain”. Biore-source Technology 101(20):7908-7915 (2010). Iyer, P.V., Thomas, S., and Lee, Y.Y., “High-Yield Fermentation of Pen-toses into Lactic Acid”. Applied Bio-chemistry and Biotechnology - Part A: Enzyme Engineering and Biotech-nology 84-86, 665-677 (2000).Walton, S.L., Bischoff, K.M., van Hei-ningen, A.R., and van Walsum, G.P., “Production of Lactic Acid from Hemicellulose Extracts by Bacillus coagulans MXL-9”. Journal of Indus-trial Microbiology and Biotechnology 37(8):823-830 (2010). Garde, A., Jonsson, G., Schmidt, A.S., and Ahring, B.K., “Lactic Acid Pro-duction from Wheat Straw Hemicel-lulose Hydrolysate by Lactobacillus pentosus and Lactobacillus brevis”. Biore-source Technology 81(3):217-223 (2002).
24.
25.
26.
27.
28.
29.
30.