lasur les cates de llllngleterre au cours des mois ..... de septembre et octobre, de 1970 1975 (tire...
TRANSCRIPT

Excellence scientifique Protection et conservation des ressources Bandfices aux Canadiens Scientific Excellence Resource Protection & Conservation Benefits for Canadians
a, la marue franche, orhua: réalités biologiques
. Cantin, P. Lauzier, M. Naud,
a J. Munro et R. Bailey f 1 i
-4. i '
'A . . f .. 4 - i P.,
< <. ; .r<--:-- .A* - - * &- > L* .$y*. . Direction des Sciences biologiques??.; ' : , - 1
- - Ministère des Pêches et des Océany' . - -- e 4
I.
Institut. Maurice-Lamontagne C.P. 1000 Mont-Joli (Québec) G5H 324
- .,.
- , - - . -. ,..:'- .
5 '
7 -+
Novembre 1989 . ' D . , -- 4
! i
-1
. Rapport canadien à l'industrie sur .. . A : - - , .- . 7
les sciences halieutiques -.-,, : , ~ . ~ > j
-..R 7
$t aquatiques . ,
__-- - -- - - . - - . L . s
Fisheries s and Oeeans

Rapport canadien à l'industrie sur les sciences halieutiques et aquatiques
Les rapports à l'industrie contiennent les résultats des activités de recherche et de développement qui peuvent être utiles à l'industrie pour des applications immédiates ou futures. Ils sont surtout destinés aux membres des secteurs primaire et secondaire de l'industrie des pêches et de la mer. Il n'y a aucune restriction quant au sujet; de fait, la série reflète la vaste gamme des intérêts et des politiques du ministère des Pêches et des Océans, c'est-à-dire les sciences hali.eutiques et aquatiques.
Les rapports à l'industrie peuvent être cités comme des publications complètes. Le titre exact paraît au-dessus du résumé de chaque rapport. Les rapports à l'industrie sont résumés dans la revue Résutnés des sciences aquatiques et halieutiques, et ils sont classés dans l'index annuel des publications scientifiques et techniques du Ministère.
Les numéros 1 à 9 1 de cette série ont été publiés à titre de rapports sur les travaux de la Direction du développement industriel, de rapports techniques de la Direction du développement industriel, et de rapports techniques de la Direction des services aux pêcheurs. Les numéros 92 à 110 sont parus à titre de rapports à l'industrie du Service des pêches et de la mer, ministère des Pêches et de l'Environnement. Le nom actuel de la série a été établi lors de la parution du numéro 11 1 .
Les rapports à l'industrie sont produits à l'échelon régional, mais numérotés à l'échelon national. Les demandes de rapports seront satisfaites par l'établissement auteur dont le nom figure sur la couverture et la page d u titre. Les rapports épuisés seront fournis contre rétribution par des agents commerciaux.
Canadian Industry Report of Fisheries and Aquatic Sciences
Industry reports contain the results of research and development useful to industry for either immediate or future application. They are directed primarily toward individuals in the primary and secondary sectors of the fishing and marine industries. No restriction is placed on subject matter and the series reflects the broad interests and policies of the Department of Fisheries and Oceans, namely, fisheries and aquatic sciences.
Industry reports may be cited as full publications. The correct citation appears above the abstract of each report. Each report is abstracted in Aquatic Sciences and Fisheries Ahstrac,ts and indexed in the Department's annual index to scientific and technical publications.
Numbers 1-9 1 in this series were issued as Project Reports of the Industrial Devel- opment Branch, Technical Reports of the Industrial Development Branch, and Technical Reports of the Fisherman's Service Branch. Numbers 92-1 10 wereissued as Department of Fisheries and the Environment, Fisheries and Marine Service Industry Reports. The current series name was changed with report number 11 1 .
Industry reports are produced regionally but are numbered nationally. Requests for individual reports will be filled by the issuing establishment listed on the front cover and title page. Out-of-stock reports will be supplied for a fee by commercial agents.

Rapport canadien A 1' industrie sur les sciences halieutiques et aquatiques
No 200
Novembre 1989
L'ELEVAGE DE LA MORUE FRANCHE, GMU3 MORHUA: REALITES BIOLOGIQUES ET ECONOMIQUES
Direction des Sciences biologiques Hinisthre des Péches et des Océans
Institut Maurice-Lamontagne C.P. hBOO
Mont-Joli (Qukbec) G5H 324
Division de l'aquaculture, Région du Québec Divis ion des services économiques, Reg ion du Québec

Hinistdre des Approvisionnements et Services Canada 1989 No de catalogue Fs 97-14/200F ISSN 0704-3708
On devra referer comme suite A cette publication:
Dutil, Je-D., C. Cantin, P. Lauzier, H. Naud, J. Hunro et R. Bailey, 1989. L'&levage de la =rue franche, &dus arorhua: rdalitds biologiques et Bconomiques. Rapp. can. ind. scf. halieut. aquat. no 200: x + 41 p.

TABLE DES FLATIERES
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . LISTEDESTABLEAUX
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . LISTE DES FIGURES
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . RPSUMP / ABSTRACT.
P R B F A C E . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . INTRODUCTION
PARTIE 1 - CONTEXTE BCOMOMIQUE
. . . . . . . . . . . . . . . . . . . . L ' O F F R E M O N D I A L E . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . Débarquements mondiaux
Importance du marché américain pour le Québec . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . lvolution de la consommation américaine
. . . . . . . . . . . . . . . . . . . Consommation des produits de la mer Consommation des autres sources de protéines . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . Marché américain du filet de morue . . . . . . . . . . . . . . . . . . . . . . . . . . . Inventaire américain Importations américaines . . . . . . . . . . . . . . . . . . . . . . . . . P r i x . . . . . , . . . . . . . . . . . .,........
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . Marché du sale-séché
. . . . . . . . . . . . . . . . . . . . . . . . . . . . POTENTIEL AQUICOLE AU QUBBEC #
. . . . . . . . . . . . . . . . . . . . . . . Considérations micro-bconomiques

TABLE DE3 MATIERES [suite)
PARTIE II . BIOLOGIE ET BLEVAGES EXPPRIMENTAUX
REPRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Maturation sexuelle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Pecondation
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . DPVELOPPEMENT EMBRYONNAIRE
. . . . . . . . . . . . . . . . . . . . . . . . . . . . LARVE VITELLINE ET POSTLARVE
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Mertalit&
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Croissance
. . . . . . . . . . . . . . . . . . . . . . . . . . . . STADES JUVdNILE3 ET ADULTES
Distribution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Alimentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Metabolisme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Ration de base . . . . . . . . . . . . . . . . . . . . . . . . . . Ingestion et digestion
Taux de conversion . . . . . . . . . . . . . . . . . . . . . . . . . . . . Croissance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Maladies
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Parasites.,

LISTE DES TAlLEAUX
Tableau h ~ g
1 Estimation des cotîts de production des poissons marins en cages flottantes en Europe et du saumon de l'Atlantique au Canada . , . . . . . . 11
2 Résumé des cazactézistiques biologiques de cycle de vie de la morue franche . . . , , . , . , . , . . . . . . . . . . , . . . . , . . . . . , 3 6


- vii -
LISTE DES FIGURES (suite)
13 Exportations québécoises de morue légbrement salée, par pays de destination pour la période 1985 A 1987 . . . . . . . . . . . . . . . . . . . .
14 $volution des prix de la morue légérement salée, Italie et Gtatç-Unis, entre 1985 et 1988
15 $volution des débarquements québécois de morue entre 1984 et 1988 . . . . . . .
16 Pvolution des prix au débarquement pour les trois catégories de palangriers du Québec entre 1984 et 1988 . . . . . . . . . . . . . . . . . . . .
17 Saisonnalité des prix du filet de morue congeli! au marché de Boston (moyenne des prix mensuels.de 1983 à 1988) . . . . . . . . . . . . . . . . . . .
18 Potentiel de fécondation en fonction de la durée de la période de '
. . . . . . . . . . . . . . . . . . . . remisage (tiré de Davenport et al. 1981)
19 Durée de la période d'incubation en fonction de la température pour les stades larvaires lA, IB, 2, 3, 4 et 5 (dtaprbs Thompson et Riley 1984) . . .
20 Distribution des fréquences de longueur de larves de morue avec ( A droite) ou sans ( A gauche) vessie natatoire aprés 30 jours (dtapr&s Howell 1984) . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
21 Distribution des morues agées de moins d'un an en fonction de la salinite sur les cates de llllngleterre au cours des mois de septembre et octobre, . . . . . . . . . . . . . . . . . de 1970 1975 (tire de Riley et Parnell 1984)
22 Augmentation de la consommation dloxyg&ne d'une morue nourrie A satieté en fonction du temps. Les points representent la consommation moyenne sur 24 h. Les repas sont indiqués sur ltabcisse (tir4 de
. . . . . . . . . . . . . . . . . . . . . . . . . . . Soofiani et Hawkins 1982)

- vi i i -
LISTE DES FIGURES (suite)
2 3 Décroissance du poids de la nourriture dans l i e s t o m c en fonction du nombre diheures suivant le repas A 2 et lOoC (tire de Tyler 19701 . , . . . a . 2 O
24 Taux de croissance en fonction du poids de morues nourries A satiété, h 8,50C (tiré de Braaten 1984) . . . . . , . . . . . . . . . . . . . . . . . , . 22

Dutil, J.-D., C. Cantin, P. Lauzier, M. Naud, J . Hunro e t R . Bailey, 1989. L'élevage de l a morue franche, &dus mrhua: r éa l i t é s biologiques e t économiques, Rapp, can. ind. s c i . hal ieut . aquat. no 200.: x t 4 1 p.
Du s t r i c t point de vue biologique, l 'élevage de l a morue franche semble prometteur, La morue tolere de larges écarts des conditions environnementales e t la production de larves e t de juvéniles à pa r t i r de stocks de géniteurs e s t relativenient simple. Par contre, l a production de juvéniles pourrait & t r e f ac i l i t é e s i nous connaissions mieux l e s besoins nutr i t ionnels des larves, Hous disposons de peu d'informations sur l e s taux de niortalité de morue pendant l a phase d engraissement. Les mladies bactériennes, l e cannibalisirie e t l e paras i t isrne sont des facteurs qu ' il faudra contraler pour améliorer l e rendement des élevages. Au Québec, cer taines indications t e l l e s la baisse des débarqueslents e t l a ra re té de morue de for te t a i l l e suggtlrent une poss ib i l i t é de earchb pour une production aquicole de morue. Par contre, comme l e marche des produits halieutiques e s t influence par des variables ayant une portée A l 'échel le mondiale, c ' es t h ce niveau que s e dessine l a tendance des prix. Actuellement, l e jeu de l ' o f f r e e t de l a demande mondiale f a i t en sor te que les prix sont A l a baisse. L'élevage de la morue, que ce s o i t en Bcosse ou au Québec, sur l a Basse CBte-Nord, ne semble pas rentable dans l e s conditions actuel les .
Dutil, J.-D., C. ~ a n t i n ; P. Lauzier, M. Naud, J . Hunro e t R. Bailey, 1989. L'élevage de l a morue franche, &dus aorhua: r éa l i t e s biologiques e t économiques. Rapp. can. ind. s c i . hal ieut . aquat. no 200: x t 4 1 p.
Froia a biological perspective, Atlantic c d rearing would appear promising. Atlantic cod can t o l e r a t e a vide range of environsaenta1 conditions and the production of larvae and juvenile c d £rom brood stocks does not present major probleas. Nevertheless, the production of juvenile cod could be enhanced should ve have a be t te r knovledge of the nutr i t ional requireiaents of the larvae. L i t t l e inforaiation is available on the mortality r a t e of cod in on-groving f a c i l i t i e s . Bacterial diseases, cannibalisa and parasites a r e a l 1 factors that need t o be handled in order t o increase production. In Quebec, the scarc i ty of large-size cod and decreased catches of wild cod vould suggest that cod farming sould be considered a s a complewnt t o harvest f isheries . Hovever, prices a r e a l so influenced by international markets. Presently, prices tend t o decrease, Cod rearing in Scotland and in Quebec, in the Lowr North Shore region of the Gulf of S t . Lawrence, does not appear t o be profitable.

PRBFACE
LI industrie aquicole connaft un essor formidable qui pourrait h moyen terme bouleverser l e marche des produits marins. .Si nous voulons part ic iper A ce t essor industr ie l , nous devons régler cer ta ins probl&mes inhérents h l a production d'organismes en milieu froid, t e l s un courte saison de croissance e t l a présence de glaces en hiver, ce qui sous-entend un t r a v a i l de recherche axé sur de nouvelles esphces preaentant la meilleur potentiel économique. La mrue franche a déjA f a i t l P o b j e t d 'essais d'élevage A Terre-Neuve e t dans l e nord de l'Europe e t son élevage pourrait ê t r e envisagi5 par cer tains aquiculteurs de l'Est du Canada. Le personnel de l a Division des Services economiques e t de l a Division de l 'Aquiculture, Région du Québec, a prépare l e présent rapport dans l e but d'analyser les facteurs biologiques e t économiques pouvant avoir une incidence sur l e succAs d'élevage de. ce poisson marin au Québec.

INTRODUCTION
La demande mondiale de produits marins est en croissance, mais il semble peu probable que la pêche traditionnelle suif ise h la satisfaire. Dans ce contexte, l'aquaculture paraît &tre l'alternative A privilégier. La production aquicole contribue déjà pour 12% des débarquements mondiaux de produits marins, Halgré un potentiel intéressant, l'industrie aquicole qubbécoise est encore bien modeste: la production aquicole contribuait en 1988 h 2% du volume des débarquements commerciaux. Cette production, peu diversifiée, est composée exclusivement de truite arc-en-ciel, d'omble de fontaine, de moule bleue et de saumon de l'Atlantique.
Dans le but de favoriser l'essor des productions aquicoles par le biais de leur diversification, nous envisageons ici l'élevage d'une esphce' de poisson marin, la morue franche (Gados morhua). Le présent rapport vi5e quatre objectifs:
- Déterminer si le contexte économique actuel, tant national qu'international, est favorable ou dtlfavorable h cet élevage.
- Relater les résultats des essais d'élevage de la morue;
- Discuter des résultats de recherche pouvant avoir une incidence sur les pratiques d'élevage de la morue; et
- Souligner les domaines oh un isanque de connaissances de base pourrait nuire A l'expansion d'une industrie basée sur l'élevage de la morue.
La morue est une esphce indighne tolérant une gamme varide de conditions environnementales (Scott et Scott 1988) et
dont l'élevage ne date pas d'aujourd'hui. Les premiers essais connus remontent A 1865 lorsque Sars en Norvège recueillit des ovules fécondés en nature pour les incuber en laboratoire. L'annbe suivante, il réussit la fécondation A partir de gamétes prélevés de morues récoltées en nature. Suite à ces essais, la Norvège inaugura en 1882 l'écloserie de Plodevigen. D'autres écloseries virent le jour h la même &poque aux dtats-Unis et A St, Johns, Terre-Neuve, La production annuelle culmina A 200 millions de larves vésicul6es en Norvége et A 2 milliards 500 millions aux Ptats-Unis. L'eff icacittl de ces programmes n'ayant jamais été démontrée, les gouvernements amér icain et norvégien y mirent fin, du moins à cette échelle, en 1952 et en 1971, respectivement. Pour les intéressés, Shelbourne (1964) et Solemdal et al, (1984) tracent un tableau historique de l'élevage de la morue au 19e et au 20' siPcles.
Le domaine n'en est donc pas A ses premiers. balbutiements et cette revue convaincra le lecteur que plusieurs laboratoires mai tr isent dé jh les techniques de base pour la fécondation, l'incubation et la production des larves. Comme les écloser ies avaient pour but la production de larves en vue de reconstituer les stocks exploités de l'Atlantique, il y a eu peu d'essais de production de juvéniles ou de morues de taille coolmerciale à l'échelle industrielle. Par contre, il y a eu récemment quelques essais en ce sens, comme ceux tentés dans des lagunes en Norvbge (Kvenseth et Oiestad 1984; Oiestad et al. 19851, et dans des bassins extérieurs A Terre-Neuve [Villiarns et Kiceniuk 1986). Il existe aussi une entreprise commerciale d'engraissage de la morue sur une base saisonnière h Terre-Neuve. H o w l l (1984) en Angleterre a réussi un élevage de la fécondation jusqulA la production de morues de prbs d'un demi kilograrme démontrant la faisabilité de 1'8levage de la morue en

c l i m t tempesd, Soulignons que la mrue Piqure 1. Partage des debarquements de semble %tre an modele intéressant pour l e morue dans les pmincipaux pays en chercheur intéressd par les poissons mazlns 1987, Les statistiques pour le de sorte gus il y a beaucoup de renseigneaents Canada excluent le Quebec. pertinents l'aquaculture de la morue dans la littérature seientif ique.
Quebec
Danemark 2.3%
LfOFPBE MONDIALE
L'offre wndiale de produits marins par ltindustrfe des pêches est importante à Islande 28.4%
PARTIE E - CONTBXTB PCONOHIQUB
considérer dans lthventwli%é de la production aquieele d'une e s w e mr ine . beislp Pe mmctà6 du poisson est Figure 2. Débarquements de morue dans les international, lBef frre mondiale est un principaux pays de 1980 à 1988. indicateur du . niveau de eoncurrence que Les donnees de 1988 constituent devra a l f ronter la production aquicsle, des projeet ions.
6
Au cours des dernieres années, les débarquements mndiaux de morue franche ont evolué autour des 1,9 millions de tonnes dtr iques ( t a ) . Cinq pays se partagent plus de 60% des débarqueaents totaux, I l s 'agit par ordre dtinportance, en 1987, du Canada, de 1' Islande, de la lotvdqe, do Danemark e t des Etats-Unis. Des 1 325 100 tn dbbarquées p c cas pap en 1987, le Qubbec compte pour 2,3% (Pig. 1) (Lauzier, 1988).
Deguis 1983 les dIbarqueœnts de mrue de ces pays sont relativement stables, ce qui indique que l'offre de morue nta pas bté le facteur deterninant dans les fluctuations de prix des produits de cette espdce au cours des derniemes annbes (Pig. 2) .
millions 1.m. 2-
1 Danemark Etats-Unis
1 Norvege
Islande Canada
Q
k stabili té des débarquements e t % shistorique de cette exploitation peri@téant de croire A une exploitation paximale des stocks mondiaux, Bien que la FM estime que lson gent augmenter les dé%iarquei@nts par une stratégie de gestion plus efficace, il est peu probable que l'offre da morue par la pêcha trraditi~nnelle augmmte de fason significative au niveau wndial,

LA DEMANDE
Dans un contexte de stabilisation de l'offre, la demande influence les prix fixés sur un marché libre. Il est donc important de connaf tre 1 ' &volut ion de cette dernibre dans un projet d'entreprise aquicole. Cette demande sera en partie responsable de la viabilitb bconomique du projet, De plus, l'analyse de la demande permet d t identif ier les marchés les plus lucratifs et d'y déceler les exigences au niveau de la matiere première.
Importance du marché ambricain pour Pe Québec
En 1987, le Québec a exporte pour 32'2 millions $ de produits de la morue. En valeur, 77% de nos exportations ont été dirigées vers les Etah-Unis. L ' Italie constitue la deuxième destination en importance% avec près de 16% de la valeur totale, suivent le Portugal avec 3,8%, le Royaume-Uni .avec 2%, tandis que les autres destinations ne comptent que pour un peu plus de 1% (Piq. 3).
Figure 3. Exportations québécoises de morue par pays de destination en 1987.
Portugal Autres ,talie 3.8% 1
Une analyse plus dhtailltie de nos exportations nous permet de constater que l'importance du marché américain varie selon le type de produit exporté. Ainsi, les produits de morue sal&e/;&chtie qui totalisent 41% des 3 ? , 2 millions $ sont A 50% exportes aux Etats-Unis (Fiq. 4).
Figure 4. Exportations québécoises de morue par categorie de produit; en 198?,
Autres
Filets
Ctest dans ce 'type de production que se concentrent aussi nos exportations vers l'Italie et le Portugal. Far ailleurs, nos exportations de blocs de morue sont dirigées A 96% vers les Etats-Unis. Il en est de même pour les filets. Dans ces deux cas, le Royaume-Uni constitue le deuxieme débouché. Partant de ce qui précede on comprendra mieux l'impact que peuvent avoir les fluctuations de la demande américaine sur nos produits.
Pvolution de la consommation amer icaine
Les btats-Unis representent un marché potentiel de plus de 220 millions de consommateurs. Une faible variation de la consommation per capita peut produire des effets tres importants sur l'industrie des p8ches. Par exemple, une augmentation de

0 , l ki lo de la consomation annuelle de poisson per capita aux Etats-Unis cree une augmentation de 22000 tonnes métriques de la demande.
Depuis 1982, l a consommation américaine de5 produits de la mer a connu une augmentation importante, passant de 5,58 kg per capita en 1932 h 7,03 kg per capi ta en 1987 s o i t une hausse moyenne de 5% par an (Fig. 51, Cependant, i l e s t intéressant d'examiner l '&volution des derniéres années A la lumiere des projections qui s e dessinaient en 1986. Les analystes prevopien t que l a consommation américaine pourrai t a t te indre 1,71 kg per cap i ta en 1981, si l e s approvisionnenients s 'av4ralent suf f i san ts .
Figure 5. Censornt ion américaine des produits de l a mer.
kg / capita (Pr = projection) 8
Pr'86 ,, 1
Des 1986 il e s t devenu &vident que l e s espéces traditionnellenient c o n s o d e s sur l e marche américain ne pourraient combler la demande. I l s'en su iv i t une hausse importante des prix des produits de ces espéces e t une recherche de nouvelles sources d'approvisionnement pouvant combnbler c e t t e demande (Mahi-Hahi, Orange rughi, e t c ) .
Malheureusement, la hausse rapide des prix a aa len t i %a consommation. Selon les dernigres s t a t i s t i ques , l a coraacrmrnat ion américaine de produits de l a mer e s t en baisse. Elle e s t passée de 7 kg per capi ta en 1987 h 6.8 kg en 1983. Les projections les plus recentes la i ssen t c ro i re que l 'on pourrait a s s i s t e r A une lbgére baisse, ou A tout, l e moins A une s t ab i l i s a t i on de la sonsommation américaine des produits de la mer pour 1988.
Pour se convaincre de l ' inf luence de la hausse des prix des produits de l a mer sur l a consommation, i l s u f f i t de comparer la hausse de pr ix des principales sources de prot4ines. Les indices amdricains de prix montrent depuis 1984 une hausse plus rapide des prix du poisson comparativement A ceux des autres. soumces de proteines.
Figure 6. Indice américain des pr ix par '
source de psstdines de 1974 A 1987.
A n d e (P poisson; b boeuf; v volaille; p porc)
La aesistance des consommateurs A une hausse trop rapide des prix e s t d 'autant plus frappante dans l e cas des poissons, que l a consommation am0ricaine de poissons e s t loin d 'a t te indre c e l l e des autres sources de protéines (Fig. 7 ) .


Les volumes dtimportations sont 4galement un indicateur de la demnde.sur un niarché. Une demande forte i n c i t e ~ a les acheteurs A faire preuve dsagressivit4 sur les mmchés internationaux afin de stassurer un approvisionnement adéquat . Ceci aufa comme effe t dlaagmnter le niveau des importations annuelies. A 1 inverse, une baisse de la demande aura c o r n effe t la r4duetion des importations.
L n iiiipact de %a hausse des inventaires, dQ B %a baisse de la c o n s o ~ t i o n adr ica ine de poisson de fond, se répercute sur toutes l e s sources d ' iqor ta t ions adr ica ines (Fig. 9 ) . Cependant, l e Canada qui comptait pour 60% des iqor ta t ions américaines au 38 juin 1987, c o s t e p u r 63,7% A la &sip date en 1981. CPest Pe Danemark e t Pa Norvége qui voient leur part diminuer, alors que leurs exportations sur l e siarche a d r i c a i n baissent : respct iveent de 55'1 e t 27%. L'ef feé des taux de change joue sftresent e t ces pays dirigent psehblenient une pilias grande partie de Peux production vers l'Europe s b le taux de change leur e s t plus favorable.
Figure 9. Iqortationé aaéricaines de f i l e t s de morne par pays dseriqine pour la période de ~anvier A juin.
Le contexte actuel du niarche amkricain de la nome se sésume par une stabilisation de l so%fre e t une baisse de la demnde. Quel es t donc l s e f f e t de ce contexte particulier sur les prix ?
Dans un projet de production aquicole, li6volution des prix pyés h l i industrie des pêches peut donner une indication de lt4volution du prix que pourra recevoir 1 'aquiculteur pour éon produit. La tendance indiquera a i Isaquiculteur peut s'attendre A une hausse ou A une baisse de prix dans l'avenir.
A la hausse des inventaires américains de f i l e ta de auPrne aerorcée A l a Sin de 1986, correspond l a baisse et/ou Pa stabilisation des prix des différents types de f i l e t s congelés de morue. Le f i l e t canadien a a t te int un sopirot en ianviet 1987, puis a connu une baiaiae prsg~essive jusqu ' A S%.15U%/lb en octobre 1988, On ssatéend h ce que l e prix se s tabi l i se A plus ou moins $2.00 UtI/lb.
Le taux de change es t un autre facteur qui a eu de l t influence sur le prix de la m u e (Piq. 10). La devaluation du dollar adr ica in par rapport: au dollar canadien a oblige les transformpateurs canadiens en ghndral, A absorber jusqusA 8% de la baisse du prix du f i l e t . Lgapprbcàationdela - - e i o n ~ i e canadienne n'a pas induit la baisse
II Autres des prix sur l e marche a d r i c a i n m i s a Oslapide plut& a ~ % i f i e le rpouveneent. Norv@e Danemark
l
8 8 8 % 88 Année
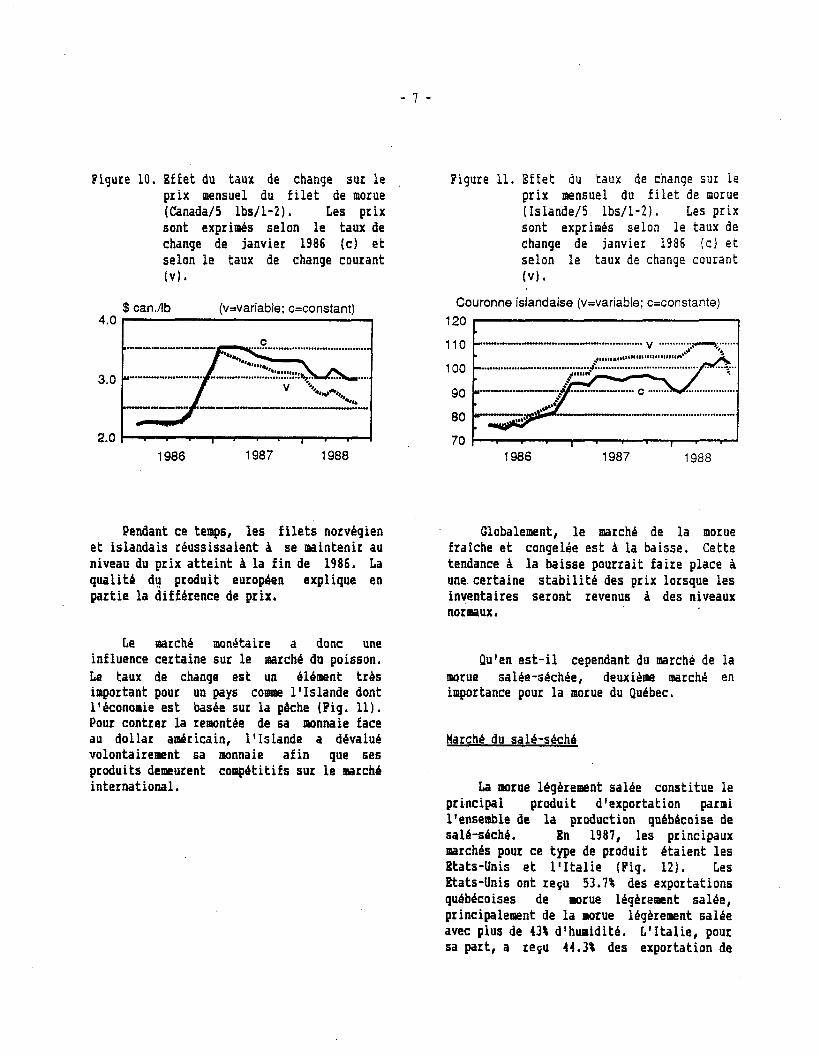
igure 10. Bffet du taux de chanqe sur l e prix mensuel du f i l e t de morue (Canadal5 lbs/l-2). Les prix sont exprimés selon l e taux de change de janvier 1986 (cl e t selon l e taux de change courant ( V I .
Pendant ce temps, l e s f i l e t s norvégien e t islandais reussissaient à se maintenir au niveau du prix a t te in t A l a f in de 1986. La qualité du produit européen explique en partie l a différence de prix.
Le riiarche mondtaire a donc une influence certaine sur l e marché du poisson. Le taux de chanqe est un élément trés important pour un pays coeiaie l 'Islande dont l'économie est basee sur la pêche (Pig. 11). Pour contrer la remontde de sa monnaie face au dollar adr ica in , l'Islande a dévalué volontairemnt sa monnaie afin que ses produits demeurent coippéti t ifs sur l e eaarchb international.
Figure 11. Ef fe t du taux de chanqe sur l e prix mensuel du f i l e t de morue (Islande/5 lbs/ l-2). Les prix sont exprimés selon le taux de change de janvier 1986 (cl e t selon l e taux de change courant ( V I .
Couronne islandaise (v=variable; c=constante) 120
Globalement, l e marché de la morue fraîche e t congelée es t I la baisse. Cette tendance à l a h i s s e pourrait faire place à une. certaine s t a b i l i t é des prix lorsque les inventaires seront revenus A des niveaux norQbaux.
Qu'en es t - i l cependant du marche de la morue salée-séchée, deuxiéipe laarche en importance pour l a morue du Québec,
La morue légkrement salbe constitue l e principal produit d'exportation parmi l'ensenble de l a production qoébecoise de salé-séché. Rn 1987, les principaux marchés pour ce type de produit etaient les Rtats-Unis e t l ' I t a l i e (Fig. 121. Les gtats-Unis ont reçu 53.7% des exportations québdcoises de wrue léqérenaent salée, principalement de l a norue légérement salbe avec plus de 43% d'humidité. L'Italie, pour sa part, a repu 44.3% des exportation de

morue 14géremnt salée principalement A 43% d'humidité ou moins.
Figure 12. Exportations qubb4coises de morue 14qhrement salée par pays de destination en 1987.
Autres 2.1 O h
Depuis 1985, on observe un ckangel~ent d ' iqjortance des marchas d'exportation pour l a morue sal4e-séchbe (Fig. 131. La valeur des expor f a t ions qu4bécoises vers 1 1 t a 1 i e aug~en te continuelleeient a lors que l a valeur: des bpor ta t ions américaines pour l e m@m produit e s t A l a baisse.
Figure 13, Bxportat ions qu4b4coises de morue 14gdrement salbe, par pays de destination pour l a pbriode 1985 (a 1987.
millions $ can. 'O - 8 6 4 I Etats-Unis 2 Italie O
1985 1986 1987 Année
Le prix de l a morue légkrement salee exportée en I t a l i e a progresse plus rapidement que sur l e mazché américain (Pig. 149. La deieande sur l e marchk i t a l i en semble s e maintenir puisque l e s prix en 1988 ont conserv4 leurs niveaux de l'anntie 1987 soratraireeaent A l a tendance génkrale des prix A la baisse sur tous l e s autres marches de l a morue canadienne.
Figure 14. dvolution des prix de la morue legerement salée, I t a l i e e t Etats-Unis, entre 1985 e t 1988.
8
6 4 Italie 2 Etats-Unis O
Re plus, l e taux de change en t re %a l i r e Italienne e t l e dol lar canadien dewtnre favorable aux exportations canadiennes.
POTEMPIEL AQUICOLB AU QUPBEC
Certaines indications du contexte g u ~ ~ o i s de l a mrue sugghrent une poss ib i l i t4 de mrch4 pour une pzoduction aquicole de morue.
Les dbbarqueaents qubbbcois de morue sent l a baisse depuis 1985 (Fig, 151. Les rbsu l ta t s p rh l ia lna i res de l a saison 1988 indiquent que l e s captures seront encore nioins importantes qu'en 1987. Au niveau de toute la region du Golfe du Saint-Laurent, cc sent %es pdcheurs u t i l i s a n t des engins

fixes qui subissent le plus cette diminution.
Figure 15. Pvolution des dbbarquements québbcois de morue entre 1984 e t 1988.
'000 t.m. (1 988 = projection) 1
84 85 8 6 87 88 Année
Ceci implique une raretb encore plus prononcbe de la grosse mrue sur les rsarchbs puisque les engins fixes ( les palangriers surtout) capturent une large part de - la morue de grande tail le. Tout ceci se traduit par une difficulté croissante pour les trans f ormteurs de s fi assurer un approvisionnement de morue leur permettant de produire pendant la saison entibre.
L'offre sur le marché de la mrue salke-sbchée est restreinte par la disponibilitb de morue de grande ta i l l e nécessaire h cette production. La rareté de la grosse morue s 'es t traduite en 1988 par un prix plus blevb payb aux pkheurs qui en capturaient.
Au Quebec coam dans tout le Colfe, ce sont les phcheurs utilisant des engins fixes (les palanqr iers) qui capturent les grosses morues. Les gros palangriers ( > 50 pieds) ayant un rayon d'autonomie p lus grand que les petits sont p l u en mesure d'aller chercher la grosse morue. Les prix payés au dbbarqueraent en 1988, ref lbtaient la diffhrence de ta i l le dans les captures pour
les palangriers d u Golfe ( P i g . 16). Donc, un aquiculteur élevant de la morue pourrait avoir avantage h produire de l a grosse morue qui couunandait des prix plus élevks. Encore faut-il que. le différentiel de prix q u i prdvalait en 1988 se répbte dans l'avenir,
Figure 16. Bvolution des prix au débarquement pour les trois catégories de palangriers du Québec entre 1984 e t 1988.
11 semble exister une légbre saisonnalitb dans les prix du f i l e t de morue sur le marche de Boston (t 5 O hors saison) (Piq. 17 ) . Un producteur aquicole pourrait peut-être obtenir un meilleur prix en vendant son produit hors saison, soit durant la p4r iode dbcembre - janvier. Cependant, la moyenne des prix wnsuels au dbbrquenient pour la morue qubbecoise e t la morue de palangre de la region Scotia-Fundy ne montre aucune saisonnalité particulibre,
La mrue de l'Islande ou de Norvbge corande des prix plus blevks que la morue du Qudbec. Cette diffbrence de prix est souvent l iee A la qualitb supérieure que les acheteurs attribuent ces produits, Une production aquicole pourrait @tre en mesure d tad l iorer la qualité des produits e t ainsi obtenir un prix équivalent ou même supérieur aux produits islandais.

Figure 17. Yaisonnalité des prix du f i le t de morue congel6 au eiarché de Boston (moyenne des prix mensuels de 1983 a isea,.
considérat ions micro-écononiioues
I l existe peu d' informtion disponible A lfheuze actuelle s u les entreprises aquico%es qu i produisent de la mr%~sme. L'dlevaqe de la mrue est pratique en Nemvdga. ' &a g~duct ion extensive de juvdniles est faite dans de larges baies ferdes. Les poissons atteignent un poids de deux kilos en deux ans environ. La viabilité commerciale de cette acéivitd n'est pas encore démontrbe. Une entreprise conjointe &osse - Norvdge est A planifier la production de mrue en cages flottantes dans le nord-est de llEcosse. Les norvdgiens devraient fournir 5 000 a 10 OQO alevins servant h ddbubr les o@ratioraa en 1988 - 19819. I l existe dgaleeient A Terre- Neuve une entreprise aquicole privée qui se spdcialise dans l'engnaissersent de la petite morue de trappe possédant une faible valeur initiale.
production des poissons marins en général (Tableau 1).
Selon Hyrseth (1988) , les poissons mains don t le prix sur le wrchb est supbrieur ou kgal A 5/10 $ ean/kg pourraient être produits en aquiculture de faqon rentable en Europe. Au Nouveau-Brunswick, i l est h noter que le coQt de production pour élever du saumon en cage s'éliverait A $ %,24/kg (Piander et Good, 1988). Acéuiellement, la morue est considérée comme un poisson h faible valeur et son prix sur le mrchb e s t encore loin de 5,70 $/kg. Par contre, certaines eagdcas co rn la truite e t le hrhu de rivibre @nt des coûts de gr&uCt ion nsttemnt infbr ieurs aux co8ts prksentks ic i . La morue a peut-Otze une structure de coats de production qui s'apparente A ces espdces, mais ceci reste h prouver.
Les infsrmtrfasts dispenibbes A se sujet proviennent d'une dtude plgoduite par Jones ( 198 4 ) . Ce dernier a simulé f inawcibreiagwt l'exploitation d'une cntzapriae commerciale citélevage de morue de lsseuf A la ta i l le cerspaercia%e. Ses sonc%usisns , Btaient celles-ci: nTeshniquement, % 4levage de merue est possible, mais les projections économiques indiquent un coat de production auphrieur au prix du marché. Dans les conditions actuelles, ce type d'élevage se peut @ére rentable. Une petite entreprise ayant accbs A une alimentation geu coûteuse e t A un mrekd spécialiad pourrait Otre rentableme Bst-ce qu'i l serait rentable d'élever de la morue au Québec? Voilh la question h laquelle tentent da rbpondre Cantin e t Bourdaqes ( 1989 1 ,
L 6tat actuel des connaissaweea sur l'blevage des poissons mrins en cages flottantes periiiet de faire certaines hypothdses réalistes sur les cotits de

TABLEAU 1. Estimation des co8ts d e production des poissons marins en cages flottantes en Europe et du saumon de l'Atlantique au Canada.
Hypo t hbses Poissons marins Yaumon de 1 'Atlantique Europe Canada
1 nvestissements
Taux de conversion alimentaire
Coiits d'alimentation
Production (/employé(e )/année)
Dbpréciat ion
Alimentation
Alevins
Main d'oeuvre
Autres cofits d'op4ration
Bntretien/Haintenance
Administration
Arsort issement
Intér4ts sur investissement
Intér&ts sur marge de crhdit
Total de frais de production
115
1/12 $/kg
50 tm
109
) 5 ans
2 / 2
0,90 $/kg
12,6 tm
109
) 5 ans*
* A l'exception des moteurs et véhicules

PARTIR 11 - BIOLOGIE ET PLEVAGES EXPPRIHENTAUX
REPRODUCTION
La reproduction rev&t une double importance pour 1 'entreprise aquicole . Ainsi, 1 'entreprise visant l'engraissement doit connaître les facteurs de déclenchement de la maturat ion sexuelle, puisque celle-ci ralentira l a croissance, ce qui se traduira par une augmentation du coiit de production. De m&me, si e l l e maintient un stock de géniteurs, 1 'entreprise doit contrôler l e cycle de maturation sexuelle de manière A obtenir des gad tes de façon synchrone e t s i possible h tout moment de l'année,
géniteurs, plusieurs mois A l'avance. Halheureusement, il n'existe pas de caractères sexuels secondaires visibles, sauf peint-être Pa formation de tubercules nuptiaux chez les &les (Vladykov e t a l . 1985). Les morues en voie de maturation sexuelle, du moins en captivité, cessent de sialimenter; pendant 2 mois leur croissance est négative ce qui entraîne une perte de poids rel iée A l a fraie de 16% chez les mâles e t de 27% chez les femelles (Braaten 1984).
La fécondité est &levée. Elle peut être calculée au moyen de la formule suivante :
oh L e s t l a longueur (cm) e t P l a fécondité en million dsovules.
Maturation sexuelle
La saison de reproduction de la morue es t t rès ' etendue. Entre Terre-Neuve e t l e banc Georges, e l l e .a lieu de janvier h juin (Pahay 1983; Scott e t Scott 1988). Dans l e sud du Golfe du Saint-Laurent, e l l e commence en mai, culmine A l a fin-juin (Powles 1958) pour se terminer A l'automne (Scott e t Scott 19889. Par contre, l a t a i l l e e t conséquemrsent la v iabi l i té des oeufs diminuent au cours de l a saison de reproduction (Sars c i t e par Knutsen e t Tilseth 1985).
La t a i l l e minimale t la nuturation sexuelle varie avec les stocks, m i s se si tue généralement autour de 30 B 35 cm. La t a i l l e médiane es t autour de 40 cm e t toutes les morues de plus de 50 ca sont matures sexuellement (Beacham 1983 a, .b; Baird e t a l . 1986). Holdway e t Beamish (1985) e t Walsh e t al . (1986) ont identifie des c r i teres histologiques permettant de distinguer les morues immatures des
Donc, une morue de 40 cm produit environ 150,000 ovules e t une morue de 80 cm environ 1,600,000 ovules. Hême si l a fécondité es t fonction du' poids, le nombre dsovules par unit& de poids est constant (Bliassen e t Vahl 1982).
Fécondation
Les techniques de fécondation i n vitro, suite au prélèvement des ganrètes dsun mâle e t d'une fereglle ou de plusieurs &les e t femlles sont les ni&mes que pour d'autres poissons. Les gamètes peuvent ê t ~ e conservés plusieurs heures A sec A des températures de 50C (Kjorsvik e t Lonning 19831. Par contre, les oeufs gardes en eau de mer deviennent vite s tbr i les (Fig. 18) (Davenport e t a l . 1981). La methode sèche e t la dthode humide ont donné de bons résultats , La niéthode sèche consiste A fdconder les oeufs dans un rbcipient ne contenant pas d'eau de mer, bien que l e

rbcipient doit Btre m o u i l l b au prbalable. Dans la méthode humide, les oeufs sont mis dans l'eau de mer, generaleraent à 5 ou 6 OC, temperature & laquelle la Eraie a lieu, e t l a laitance est versee. Le mélange est laissb A reposer de quelques minutes & une heure avant d 'être rincé plusieurs fo i s avec de l'eau de mer [Shelbourne 1964; Kjorsvik e t Eonning 1983; Iversen e t Danielssen 1984).
Figure 18. Potentiel de fer t i l i sa t ion en fonction de la durée de la période de remisage ( t i r 8 de Davenport e t a l . 1981).
O 3 6 9 12 15 18 Temps (h)
Certains auteurs privildgient la technique de l a "raie naturellen dans un bassin ou un btang (Laurence e t Rogers 1976; Jones 1984). D'ailleurs, cet te technique n'est pas récente puisqu'elle a étd uti l isde t rés t d t en Norvège (Soleadal e t a l . 1984). Les oeufs fécondés sans intervention humaine sont rdcoltes en f i l t r an t l'eau, normalement A ' la surface, avec un f i l e t A fine m i l l e (Laurence e t Rogers 1976; Hove11 1984). La f lo t tabi l i té des oeufs est fonction de nombreux facteurs que nous examinons ai l leurs dans ce document. Quelques semaines avant la fraie, les morues deviennent agressives les unes envers les autres alors qulun &le vigoureux é tab l i t sa
doniinance e t cree un te r r i to i re . Les individus dominés se regroupent A l 'extbrieur de ce ter r i to i re , Toute la fraie se déroule dans le ter r i to i re de ce &le dominant, gentiraleisent la nuit près de la surface (Bravn 1 9 6 1 a, b) . Les morues ne relachent pas tous leurs oeufs au même moment, Elles le font sur une période de 2 à 3 searaines (Shelbourne 1964 ; Ho1dway e t Beamish 1985), les derniers oeufs relâchés btant plus pet i t s e t moins viables que les oeuEs relâchés les premiers (Knutsen e t Tilseth 1985 1 . La fraie naturelle produirait des oeufs de meilleure qualit4 (Shelbourne 1964).
Les oeufs de l a morue sont pélagiques. 11s ont un diamètre de 1.2 & 1.6 mm lorsquli ls ont dtb a i s en contact avec l'eau. Le chorion transparent laisse voir un vi tel lus homogbne sans globule d' huile (Fahay 1983). Leur f lo t t ab i l i t e e s t attribuable A leur contenu bleve en eau (92%) (CIaik e t Harvey 1987). Plusieurs auteurs ont décri t des méthodes d'incubation consues pour des oeufs f lot tants (Shelbourne 1964; Thoqson e t Riley 1981; Hove11 1984; Kjorsvik e t Lonning 1983; Solberg e t Tilseth 1984) .
Les stades du développement embryonnaire ont bt6 ddf inis e t décrits par plusieurs auteurs dont Laurence e t Rogers (19761, Thoqson e t Riley (19811, Fahay (19831, e t Makhotin e t al . (1984). La longueur de la période d f incubation e s t directement proportionnelle A l a température (Fiq. 1 9 ) e t ce t te relation peut Btre décrite par lf6quation suivante:
Y = 21.96 - 1.30 X (Laurence e t Rogers 1976)
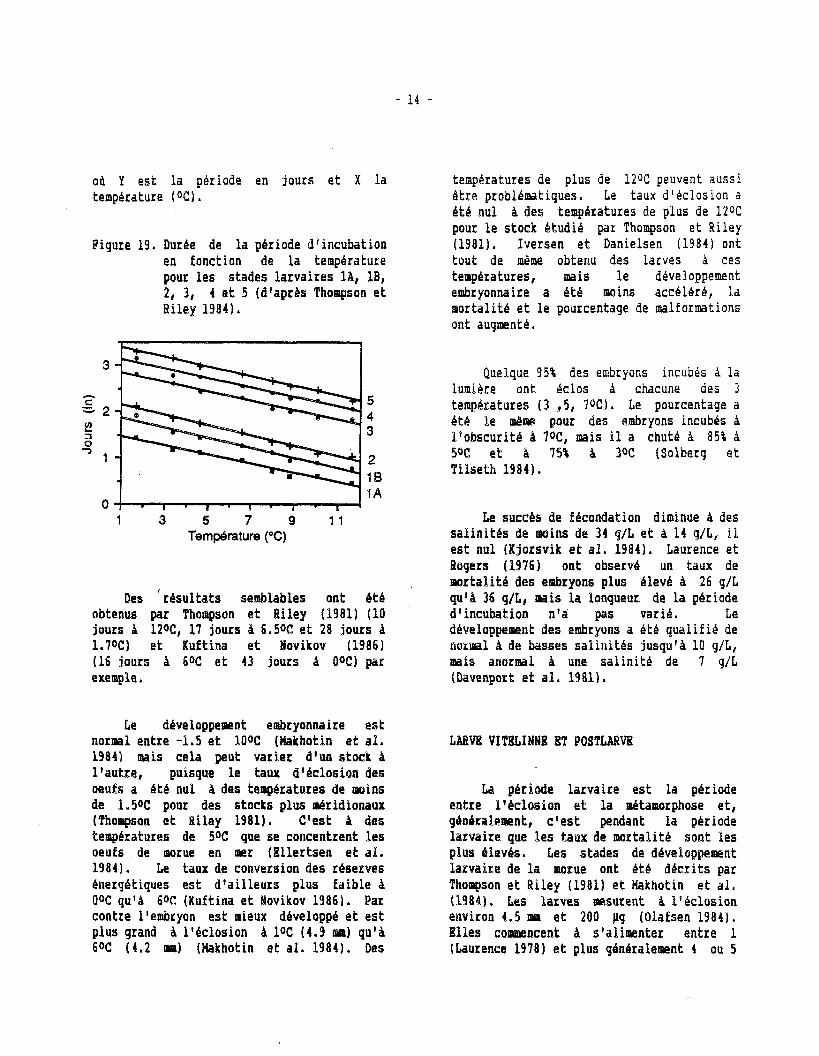
od Y e s t l a periode en jours e t X l a temperature ( O C ) .
Figure 19. Dur& de l a période dsincubatioe en fonction de l a temperature pour l e s stades la rva i res l A , l B , 2, 3, 4 e t 5 (dvapr&s T h o q ~ o n e t Riley 1984).
teapkratures de plus de 1206 peuvent aussi êtme pmobléip;akiques. Le taux d'tislosion a kéb n u l à des t e q b r a t u r e s de plus de 1206 pour l e stock Btudib p a n Thompson e t Riley (19819. Iversen e t Danielsen (1984) ont tout de mime obtenu des larves h ces tempiratures, mais l e developpement embsgonnaire a é t é m i n s accblkré, l a s lor tal i te e t l e pourcentage de malformations ont augnient4.
1 3 5 7 9 1 1 Temperature (OC)
Des ' résu l ta t s semblables ont é t é obtenus par Thompson e t Riley (1981) (10 jours à lZoC, 17 jours A 6.50C e t 28 jours A l.7oC) e t Kuftina e t lovikov (1986) (16 jours à 60C e t 43 jours à BW) par e x e q l e .
Le développement embryonnaire e s t norisal entre -1.5 e t lOoC (Ilakhotin e t a l . 1984) reais cela peut varier d'un stock A l ' au t re , puisque l e taux dl&closien des oeufs a é t é nul à des températures de moins de 1.50C p o u des stocks plus miridionaux ( ' thoqssn e t Riley 1981). C'est A des terapératures de 50C que s e concentrent %es oeufs de aiomue en mer (El ler tsen e t a%. 1984). Le taux de conversion des réserves energétiques e s t d ' a i l l eurs plus fa ib le A OoC qu'A 606 (Kuftina e t Novikov 19869. Par contre l'embryon e s t mieux dbveloppé e t e s t plus grand à lvdclosion à loC (4.9 m) qu'A 60C ( 4 . 2 nual (Hakhotin e t a l . 1984). Des
Quelque 95% des embryons incubés h la lumigre ont &clos h chacune des 3 t eqb ra tu r e s (3 ,a, 7 W ) . Le pourcentage a dtk le utdm pour des embryons incubes A %'ctbscuritB A ?OC, mais 19 a chute .A 85% A 5OC e% % 75% A 3oC (So lbe~g e t Tilseék 19841.
Le éuccLIs de fbilondation diminue à des s a l i n i t d s de sisirts da 34 g/E e t A 11 g/LI i l e s t nul (Kjorsvik e t al . 1984). Laumenee e t Rogers (1976) ont observe un taux de m r t a l i t é des embryons plus Blevé A 26 g/L qua% 36 g/L, mis l a longueur de l a période d'incubation n'a pas varie. Le ddveloppment des embrpns a &te qualif i4 de nozmP $ da basses s a l i n i t b s jusquilà 18 g/E, m i s anormal à une s a l i n i t e de 7 g/L (Davenpo~t e t a l . 1981).
ka périede la rva i re e s t l a période entre l 'éclosion e t l a dtarwrphose e t , gdnénéilaent, c i e s t pendant l a pér iode la%vaire que l e s taux de montalité sont %es plias élev4s. Les stades de dévelsgpemnt la~cvaire de l a morue ont btb dec r i t s par Thompson e t Riley il981 1 e t Hakhoti~ e t a l . (198%). Les larves mesurent % l 'éclosion e a v i ~ o n 4.5 mm e t 200 pg (Olafsen 19841. Elles comaencent A sqalioaenter en t re 1 (Lamrenee 1978) e t plus généraleisent 4 ou 5

jours plus tard (Gamble e t Houde 1984) Pendant ces quelques jours e l l e s t i r e n t leurs besoins nutrit ionnels de leur sac v i t e l l i n , jusqu'h ce que l e s yeux, l a bouche e t l ' i n t e s t i n deviennent fonctionnels (Yin e t Blaxter 1987a). La résorption du sac v i t e l l i n e s t terminée vers l e 7e jour suivant l 'éclosion. C'est h ce moment que l ' a c t i v i t h aliieentaire e s t h son maximum. Les larves doivent entre l e 5e e t l e l l e jour réussir à t i r e r leur nourriture exclusivement du milieu extkrieur. Si e l l e s n'ont pas rencontré leurs besoins énergétiques au bout du l l e jour, e l l e s sont condamnées, & une température ambiante de 7oC, h mourir dans les 5 jours qui suivent ( Y i n e t Blaxter 1987a).
ans une s i tua t ion d'élevage, l a t a i l l e des proies e s t un Eacteur cr i t ique, sur tout dans l e s quelques jours suivant 1 'éclosion. Lorsqu'elles commencent h slaliieenter, l e s larves ne peuvent ingérer l e s nauplii d ' Artemia sp., un organisme zooplanctoniq'ue couramment u t i l i s e pour 1 'élevage des larves, ' De plus, une d ié te constitube exclusivenent de ro t i fé res ou de nauplii dlArtemia sp. a des carences e t ne permet pas aux larves de survivre jusqu'h l a miitamorphose (Howell 19841. Howell suggére de nourrir l e s larves avec des ro t i fé res , puis de passer des rot i fbres aux métanauplii dtArtemia sp. préalablement nourris d'un mélange de deux algues, I l n 'exis te A l 'heure actuel le aucune nourriture a r t iE i c i e l l e pour des larves de ce t t e t a i l l e (Hjelmeland e t a l , 1984; Oiestad e t a l . 1985). Les techniques de production d'algues, de ro t i fé res e t d'llrtemia sp. de m6me que les densites de ces organismes e t autres organismes zooplanctoniques fournis aux larves sont decr i t s entre au t res par Thompson e t Riley I1981), Howell (1973, 19841, Solberg e t Tilseth (19841, e t Ti lseth e t El ler tsen (19841.
Peu de renseignements sont disponibles sur l es facteurs in£ luençant l a capture des proies. Les larves de morues sont des predateurs visuels . El les s 'alimentent sporadiquement e t requibrent une intensi té lumineuse de l ' o rd re de 0 . 1 l u x pour détecter l es proies (El ler tsen e t a l . 1984; Tilseth e t El ler tsen 1984). En nature, l es larves s e tiennent prbs de la surface où e l l e s s e nourrissent de nauplii de copkpodes dont l a t a i l l e e s t fonction de la t a i l l e des larves, l e s plus pe t i tes larves se nourrissant de plus p e t i t s crustacks (Lough 1984). Graduellement, e l l e s d i la i ssen t l e s nauplii pour des s tades plus avancbs de copépodes calanoides jusqu'h l a métamorphose (Kvenseth e t Oiestad 1984). En laboratoire, si un choix leur e s t donné, l es larves sé lec t ionnent l e s naupl i i de copépodes (El ler tsen e t a l . 1984; Ti lseth e t El ler tsen 19841, El les consomment un nauplii h l 'heure jusquth un aiaximum de 3 ou 4 nauplii , Le temps de digestion d'un nauplii h 5oC e s t de 30 minutes (Ti lseth e t Ellertsen 19841. I l ex is te plusieurs méthodes visuel les (Oiestad 1984; Ti lseth e t El ler tsen 1984; Yin e t Blaxter 19861 e t biochimiques (Buckley 1979, 1981, 1984; Raae e t a l . 19881 permettant de dktecter l es troubles d'ordre nutr i t ionnel des larves.
Mortalité
Howell (1984) dans une s e r i e d ' essa i s a obtenu un taux de survie des larves de 10% jusqu'h l a metamorphose e t de 5 A 7% au bout dsune phriode de 72 jours h une t i i l l e f ina le de 3.5 cm. Parmi l e s facteurs de mortalith, mentionnons des d ié tes inadéquates e t l e cannibalisme. Howell (1984) a observe l e 30e jour une dis t r ibut ion bimodale des longueurs dû h un retard dans l a croissance des larves qui n'avaient pas reussi h remplir leur vessie natatoire (Pig. 201, Lorsque la période c r i t ique e s t passée, l e taux de mortalité reste constant (Raae e t a l . 1988). Le

transfert de. larves Sgkes de 5 jousa A des lagunes en No~vhge a donné l e s deux premihrea annbes un taux de survie de 29 jusqu à Pa mbtamorphose . Le pourcentage es t passé A 30 e t dnne 509 par la sui te lorsque des mesures ont été prises pour eliminer l e probllsrpe de prédation par les niéduses sur les larves (Kvenseth e t Oiestad 1984; Oiestad e t a l . 19851.
Figure 20. Distribution des fr&quences de longueur de larves de morue avec (à droite) ou sans ( à gauche) vessie natatoire aprBs 30 jouas idgapz8s Hove11 1984).
6 7 8 9 .IO 11 Longueur (mm)
La température e t la sa l in i t0 le ta les (LCso, 2 4 h l pour des larves d jeun sont respectivement de 16 h 18 O C ( A 32 9/11 e t de 2 à 3 g/P (A 6oC) (Yin e t BPaxter f987b).
Croissance
La métanorphose se produit a une t a i l l e de 10 iua e t 1000 pg (poids sec) au bout dsune période de 4 4 jours d lOoC e t de 52 jours A ?OC (Laurence 1978) A 1 2 sip e t 18130 pg (poids sec) au bout d'une période de 35-40 jours (Kvenseth e t Oiestad 1984, Oiestad e t a l . 1985). La croissance en longueur à 7oC a Bt4 de 0.083 W l o u r entre l'éclosion e t l e 32e jour de vie larvaire:
Longueur = 4.502t0.081 Age (jours) (Yin e t Rlaxter 1986)
Elle a étb de 1,8%/jour entre le 23e e t le 62e jour suivant l'éclosion:
Ln (Longueur) = 1.50t0.018 Age ours) (Ruckley 1981)
Des courbes de croissance en masse des larves jusqul& la métamorphose ont é t é pm&sent&es par Kvenseth e t Oiestad (1984) e t Lau~enee ( 1978, 1979 .
STADES JUWILBS ET ADULTES
Les morues féquentent les fonds de 30 à 400 a A des températures de O A 130C e t des sa l in i tés de 31 d 34 g/l, mis on l e s trouve qenéralement A des profondeurs de 40 il 100 m e t à des temp4ratur.e~ de 3-4oC (au nord) à 7-IQC (au sud) (Scott 191%). Dans le Golfe du Saint-Laurent, Pa distribution verticale des morues change avec %a ta i l le , les morues de moins de 20 cm se tenant plus prés de la ebte dans la couche intermédiaire (Jean 1964 1 , C'est également l e cas dans l 'Atlantique-kt (Pig, 21) 04 les juvéniles friiquentené les eaux moins profondes, les individus de moins dsun an rementant mêw les estuaires jusquqA des eaux dont la sa l in i te n'est que de 10 A 20 q/l dont les mfuea plus 8g6es sont absentes (RiPey e t Parnell 1984). Les morues de moins de 20 cm ne aeiablent pas migrer hors du golfe du Saint-Laurent en hiver, alors que les autres =rues font des migzations saisonnieres vems les eaux profondes e t l'Atlantique. Ces g l ~ ~ u e s sont de retour prbs de la côte e t pirds de la surface vers la ni-juin (Jean 1964 1.

Figure 21. Distribution des norues Aqées de moins d'un an en fonction de l a sa l in i té sur les côtes de l'Angleterre au cours des mois de septembre e t octobre, de 1970 A 1975 ( t i r é de Riley e t Parnell 19841.
Alimentation
Les mgrues juveniles e t adultes, tout cowœ les larves, sont des prédateurs visuels. Blles peuvent repbrer e t sa i s i r des particules aussi fines que 2 m. Le passage h la vision scotopique a l ieu h des intensités luaineuses de 1 'ordre de à 1 lux A l i 1 9 7 1 . Les morues sont aussi capables de détecter l a nourriture cachde dans l e substrat ou les particules fines sur le fond à l 'aide des papilles gustatives sur le barbillon e t les nageoires pelviennes (8rani 19691. 11 semble que les acides aminés glycine e t alanine soient de bons déclencheurs de l a séquence d'alimentation (Bllingsen e t Doving 1986). Des qu'elles mesurent 2.5 4 3 .0 Ca, les morues se nourrissent volontiers de moulée e t A part ir de ce t te t a i l l e la survie e s t la &me que la diete so i t constituee de moulbe ou dlAztemia sp. (Bromley e t 3ykes 19851. Des wrues sauvages transportées A nos installations s'alimentaient de moulée ou de poissons quelques jours seulement suivant leur arrivée.
Dans leur milieu naturel, les morues s'alimentent de plusieurs espèces de crustacés e t de poissons. Halheureusement, l'incidence du cannibalisme n'est pas négligeable e t pourrait être significative en production, puisque les souches non-domes tiquées de poisson sont caracterisées par de fortes variations individuelles du taux de croissance. Or, l'incidence du cannibalisme augmente avec la t a i l l e en nature: e l l e es t nulle chez les morues de moins de 50 cm e t augmente progressivereent jusqulA 7% de la dihte chez les morues de plus de 81 cm (Waiwood e t Hajkowski 19841. De même dans le Saguenay, 10% des poissons dans les estomacs de morue sont des tnoru'es (Lalancette 19841, Les morues dont l a t a i l l e se situe entre 1 0 e t 25 cm semblent faire les f ra is de ce phénomène (Templeman 19761 ce qui pourrait expliquer la segréqat ion des t a i l l e s en milieu naturel (Jean 1964, Riley e t Parnell 1984). Dans les Olevaqes, le cannibalisme se manifeste A toute t a i l l e . Broaley e t Sykes (1985) ont note que plusieurs cas de rnoztalité étaient dus au cannibalisme chez des morues de 2.5 à 3.0 cm. Bien que le cannibalisae a i t généralement lieu entre individus de t a i l l e s différentes, les attaques peuvent se produire entre individus de &me t a i l l e (Bravn 1961a; Bromley e t Sykes 1985). L'agressivité entre les morues de même t a i l l e es t associhe A l'établissement d'une hiérarchie entre ces individus. Dans les cas extr2mes, c'est-A-dire peur les laorues au bas de l a hierarchie, les Bienaces e t ieursures répétbes conduisent A des blessures e t h un jeQne force pouvant entralner la moeort. Ce comportement a Btb observe au sein de pe t i t s groupes de mrues de 3 classes de longueur entre 5 e t 90 cm (Brawn 1961a). Les cas de cannibaliseie sont p lu s fréquents chez les morues recevant une ration moindre (Oiestad e t a l . 19851 e t , à ration constante, pourraient augmenter 'avec une élévation de la temperature, comme ce Eut l e cas dans nos bassins l e printemps dernier. Vraisemblablement, de te l les interactions

ont moins de chances de se produire dans les bassins h t rks fortes densites.
Le nrétabolisrne des poissons es t fonction de plusieurs facteurs, en particulier leur poids e t la tempbeature de leur envlronaemnt . Le &tabo%isnie se mesure habituellement par la consonmat ion d'oxygine puisque les facteurs influengant le métabo%isme influencent aussi la coaisoslei;itiow d oxygéna, La consommation dqoixyg&ne en fonction du poids du poisson e s t dbemite par lvdquation:
Qot (mq Op/heure) = 0.0204 P(g)aa8z* (Braaten 1984)
A une temperature de 8,4QC, et:
1
$ une temperature de 1206. Ainsi, des morues de 500 g consoament environ 25 sag Bs/heure A 3 et 46 q Ba/hpu~e A I50C contre 1'98 e t 354 mg O~/haure b 3 e t l5QC respectivement gour des morues de 6 kg (Saunders 1963). L9augmentat ion post-pandriale (suite d 1 "angestion de nourriture 1 de la eonsom t ion d oxygkne chez la morue es t trks naarqube. Elle es t fonction de la ration, du neabre de repas e t de la tesegdrature (Piql. 221. k consosirsation reto&e au niveau de base environ 24 h suivant l e repas ou aprés plusieurs jours lorsque la morue a consomd plinsieurs repas sur une courte pbriode (Soef iani et Havk inis 1982 1 . Ces considbrations sont d~termimntes puisque la morue ne toldre pas des eoncentraticsw d'oxygéne sous les 3 ag/L A 10oC (Saundems 1963 1 .
Figure 22. Augmentation de la consoinmation dsoxygéne d'une morue nourrie a sa t ib té en fonction du temps. Les points représentent la consoinraat~on moyenne sur 2 4 h. Les repas sont indiqués sur lsabcisse ( t i r 4 de Soofiani e t Hawkins 19821.
La teqbrature aura aussi une incidence sur la q u n t i t d de nourriture ingéree, la proportion de l'dneacjie cswsommtie requise poux assumer les besoins fondamentaux de lssngcinisaag, e t la proportion de l'tsnergie s e n s o d e qui %ma A Pa eaeisàaance. Be nidme, la composition de la diète ama une incidence sur la consommation e t la croissance. Comme l a nourriture représente un fort pourcentage du cott dtexploi tation d'une pisciculture, ces variables peuvent ddcider du succes ou de lt&chec, ou A tout le moins de l a marge de profi t de la pisciculture. loué n8exaainons ci-dessous que lsinformtion spbcifique h la isoria~, les principes gbwbraiax du mitabol isw des ectothermes, c'est-A-dire les organisrees dont la tanipdrature interne su i t la éeiaperatune @xterne, ktant valables aussi pour: la morue.

NOUS entendons par rat ion de base la rat ion requise pour quf un organisme maintienne constant son poids. La rat ion de base e s t fonction du poids:
RD = 0.023 P O 1 a O h 13oC (Jones e t Hislop 1978)
Ra = 0.017 P O S ' ' A l O O C (Jobling 1982)
RD = 0.025 P A 8°C (Braaten 19841
oh Ra e s t l a rat ion de base (kcal / jour) e t P le poids ( g ) .
La re la t ion entre l a rat ion de base e t l e poids du poisson e s t influencée par l a température puisque l a rat ion de base augmente avec l a temperature de fason exponentielle. Par exemple, pour des morues de moins dk 40 cm, à des températures de 7 A 18OC:
Log Ra = 0.065 T - 1.018 (Hawkins e t a l . 1985)
oh RD e s t l a ration de base ( % de Pl e t T l a température ( O C ) . Ainsi A 10oC, l a rat ion de base correspond à 0.4% du poids de la morue/jour, contre 0.9% A 15oC (Hawkins e t a l . 19851. Par contre curieusement, Jobling (1982) n'a trouve aucune re la t ion entre la rat ion de base e t la température, Kohler (1964) estinte que des morues de 510 e t 750 g maintenues h des températures ambiantes variant entre 2.5 e t 150C requidrent individuellement en un an 1176 e t 1375 g de hareng pour min t en i r leur poids cons tant .
La quant i té de nourriture que l a morue inghre pendant un repas augmente avec son poids (Jobling 19831 e t diminue avec la température, mais i l e s t h noter que la morue continue h consommer la nourriture of fe r te A des temperatures aussi basses que O ou loC (Saunders 1963; dans nos élevages, données non-pub1 ikes 1 . Dans un essai d'engraissement, Williams e t Kiceniuk 11986) ont observe une consommation moyenne de 130 g/kg/semaine chez des morues de 40 h 6 0 cm nourries à s a t i é t é t r o i s fo i s par semaine h des temperatures variant de 7 h 150C. Notons que la consommation de nourriture e s t 1.5 fo i s plus élevée lorsque les morues sont nourries à sa t i t - t e deux fo i s par jour au l i eu d'une fo i s A 4.50C (Braaten e t Gokstad 19809, ce qui r e f l é t e vraisemblablement l e ryth~te d'évacuation de l a nourriture de l'estomac h l ' i n t e s t i n (Tyler 1970). Celui-ci var ie avec l a température (Fig. 231: l'estomac se vide en 2.5 jours h 5%' e t en 24 h h 10oC. Le taux semble ê t r e maximum h 15% e t diminuer h des températures plus hautes, ce qui indiquerait que la temperature optimale de croissance se s i t ue autour de 150C (Tyler 1970). Grâce A ces données, il e s t possible de calculer l e nombre de repas h donner par semaine e t l a ration h donner par poisson pour fournir un maximum d'bnergie à l a morue selon sa t a i l l e e t l a température de 1 'environnement,
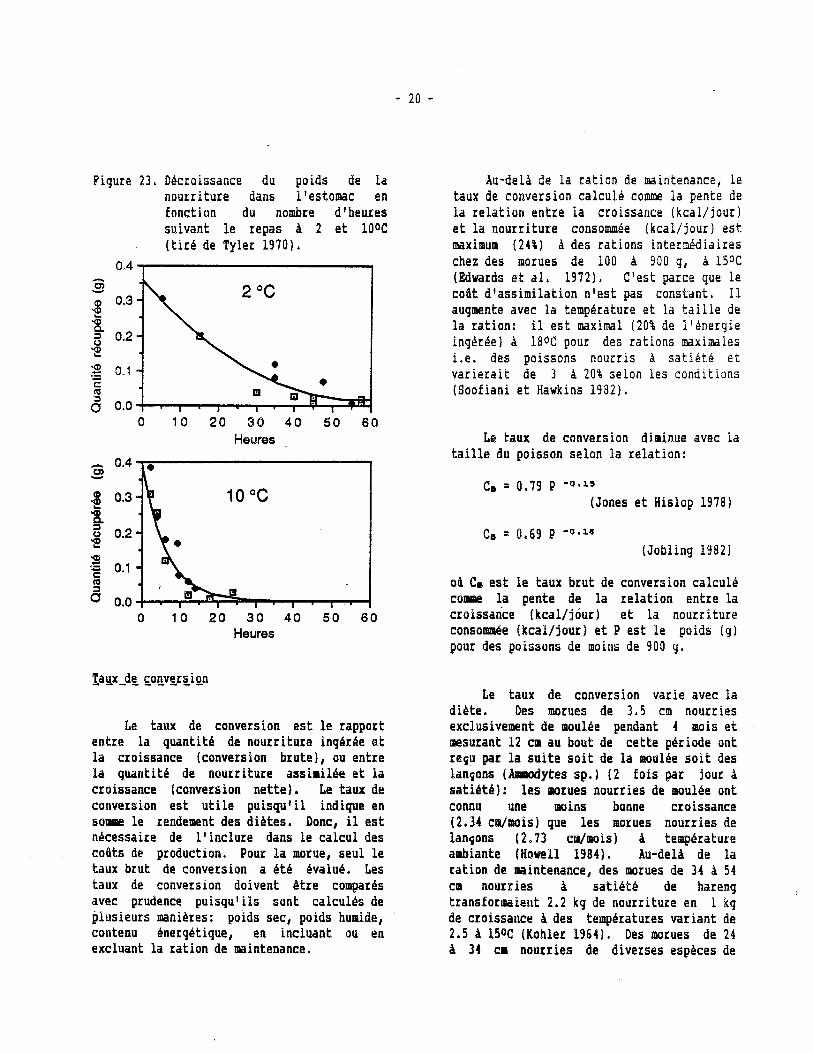
Figure 2 3 , Décroissance du. poids de la nourriture dans l'estomac en fonction du nombre d'heures suivant l e repas h 2 e t POOC ( t i r é de Tyler 19701
O I O 20 30 40 50 60 Heures .
O 10 20 30 40 5 8 68 Heures
Le taux de conversion es t l e rapport entre la quantite de nourriture ingerbe e t l a croissance (conversion brute), ou entre Ica quantith de nourriture assimilbe e t la croissance (conver~;ion nette). Le taux de conversion es t u t i le puisqu9 i l indique en soerpe le rendement des dietes. Donc, il es t nkessaire de l'inclure dans le calcul des cslits de production. Pour %a morue, seul Pe taux brut de conversion a été 4val.uk. Les taux de conversion doivent @tre coaiparbs avec prudence puisqu ' i l s sont calculks de plusieurs nunieres: poids sec, poids humide, contenu énerghtique, en incluant ou en excluant l a ration de maintenance.
Au-delA de la ration de maintenance, l e taux de c~wversion calculé c o r n la pente de la relation entre la croissance (keai/jour) e t la nourriture consomm6e (kcal/ jour ) es t maximum (249) A des rations i n t e r m i a i r e s ckezdes morues de 1 0 0 A 9 0 0 g, à 150C (Edvards e t a l . 1 9 7 2 ) . C'est parce que le codt d'assimilation n'est pas constant. I l augmnte avec la température e t la t a i l l e de la ration: i l es t maximal ( 2 0 % de lt4nergie ingdréel A l8oC pour des rations maximales i.e. des poissons nourris A satibté e t varierait de 3 A 20% selon les candltians (300f iani e t Hawkins 1982).
Le taux de conversion diainue avec la t a i l l e du poisson selon la relation:
Cm = 0.79 P '011' (Jones e t Hislop 1978)
oil Cm es t l e taux brut de conversion calculé csmrpe l a pente de l a relation entre l a croissance (kcal/jQur 1 e t la nouzsiture c o n s o ~ e (kcaP/jour 1 e t P est le poids (g$ pour des poissons de moins de 900 q,
Le taux de conversion varie avec l a diete. Des morues de 3.5 cm nourries exclusivement de mou%&e pendant 4 mois e t ,
mesurant 1 2 cm au bout de cette phriode ont rep! par l a sui te so i t de la moulée so i t des lanpons (-@es sg. 1 ( 2 fois par jour A, satikth): les morues nourries de mulhe ont connu une moins bonne croissance (2.34 cdmois) que les morues nourries de lanpona (2,73 eWmois1 h teqbratume arabiante (HowP1 19841. Au-delA de l a ration de iaaintenance, des morues de 34 h 54 cm nourries h satibté de hareng transfermient 2.2 kg de nourri tu~e en E kq de croissance h des tempbratures variant de 2.5 h P50C (Kohler 19641. Des morues de 2 4 à 34 c i nourries de diverses esphces de

poissons requéraient 2.3 kg de nourriture par kg de croissance h des températures de 6 A 180C (Howell 1984). Williams e t Kiceniuk ont mesure des taux de onv version moins élevés chez des morues de 40 A 60 cm nourries de capelan h s a t i é t é 3 fois/semaine A des températures variant de 7 A 150C.
Croissance
Le taux de croissance des morues e s t rapide, Braaten (19841 produisit en deux ans e t s i x mois suivant l a fécondation des morues de 4 kg. Dans des lagunes, Oiestad e t a l . (19851 ont. produit h par t i r de larves métamorphosées, des morues de 18 cm (60 g) dans une période de moins de 6 mois. Howell (1984) a produit des morues mesurant 12 cm (18 g1 au bout de 18 semaines e t 34 cm (475 g l a u bout de 60 semaines, Wil l iamset Kiceniuk (19861 ont obtenu un taux moyen de croissance en poids de 66 e t 79% en 12 e t 1 4 semaines lors de deux essais réa l i s& sur des morues de 40 A 60 cm nourries s a t i é t é 3 fois/se&ine h des températures de 7 h 15oC.
La croissance s e r a i t maximale h des températures de 13 A 150C (Jobling 19831, L'étude de Tyler (19701 basée sur l e taux d ' évacuation gastrique suggére l a même conclusion. Ces températures sont l e s températures sélectionnées par l a morue placée dans un gradient thermique (Bohle 1974), mais ne coïncident pas avec l e s températures auxquelles on retrouve l a morue en nature (Jean 1964, Scott 19821. Howell (1984) sugg4re plutôt des températures de 10 à 130C.
Selon Hawkins e t a l . (19851, l a croissance specif ique e s t fonction de la rat ion A toutes les températures e t , à ration constante, e l l e e s t maximale aux basses températures puisque l a ra t ion de
base- augmente avec l a température. Far contre, méme s i l e taux de conversion diminue avec une augmentation de température, l a consommation de nourriture augmente e t l e taux de croissance des morues nourries h s a t i é t é e s t aiaxiaial A des temperatures plus élevées (Hawkins e t a l . 19851, Les équations suivantes décr ivent la relat ion entre l e taux de croissance spécifique e t l a rat ion alimentaire:
Ca = -0.0520 i 0.1894 R a (Hawkins e t a l , 1 9 8 5 )
pour un poisson de 100 g A 10oC,
Cs = -0,1939 t 0.2428 Ra (Hawkins e t a l . 1985)
pour un poisson de 400 g A 10oC,
Ca = -0.58 t 1.13 Rs (Houlihan e t a l . 1988)
pour des poissons de 300 g gardés 3 mois h IOOC. Ca e s t l e taux de croissance spécif ique ( 8 du poids f r a i s par jour) e t R l a rat ion alimentaire ( 8 du poids f r a i s par jour). Braaten (19841 fournit l 'équation:
Log C = -1.16 t 0.52 Log R (Braaten 1984)
où C e s t l a croissance (kcal/jour) e t R l a ration alimentaire (kcal/jour) h 8.40C pour des poissons pesant entre 200 e t 300 g. Cette relat ion entre croissance e t ra t ion a aussi é t é demontrée pour des morues de moins de 900 g (Edwards e t a l . 1972, Jones e t Hislop 1978).
Plusieurs esphces de poissons emmagasinent l 'énergie consommée en e x c h des besoins de base du métabolisme l e long de 1' in tes t in e t sous l e derme SOUS forme de lipides, mais l a morue canalise ce t t e Qnergie vers la croissance hépatique e t l a croissance du t i s s u musculaire (Edwards

e t a l . 1 9 7 2 ) . Ainsi, l e rapport poids du foie/poida total augmente avec la ration de faeon linhaire. Le poids du foie varie de fason saisonnibre e t reflete %es variations du contenu en lipides de mèiae que l e taux de croissance de la morue (Holdway e t Beanish 198%). Holdway e t Beamish (19841 fournissent des courbes permettant de déterminer l e contenu en lipides, en protéines, en eau e t en energie h par t i r de l a mesure d'une seule des ces variables.
Le taux de croissance des morues diminue avec leur t a i l l a :
Log, C = 2.930 - 0.441 Log, P (Jobling 1983)
oh C es t l a croissance spécif ique e t P l e poids (9) pour des morues de 100 % 1000 g A des t e q k a t u r e a da 10 à 11oC, e t
C = 9.68 P (Braaten 1984)
od C es t ' 1k croissance spkcifique e t P l e poids (gl pouf: des morues de 4 kg e t moins A - des températures de 6 B 90C0 Ainsi, Braaten (1984) a déterminé que la croissance spécifique en poids des Ilterues é t a i t de 1 , 4 A 2.0% p a ~ jour entre 3,6 e t 90 g, 0,8 A 1.7% par jour entre 90 e t 800 g e t 0.2 % 0.39 par jour au-delà (Pig. 2 4 ) . Hovell (1984) a =sure un taux de croissance de 27 &mis pour des morues de mins de PZ cm e t de 19 W ~ o i s Qour des niorues de 12 B 34 cm h des teqkratures variant entre 6 e t 160C. Des mrues de 34 cm (450 g) e t 42 cm (760 g) nourries A sa t ié té pendant un an A des teqéra tures variant entre 2.5 e t 15eC se sont accrues de 15 cm e t 11 cm respectivement peur un accroissement en poids de 989 pour les asmues de 42 cm contre 157% pour les morues de 34 cm (Kehler 19641.
Figure 2 4 . Taux de croissance en fonction du poids de morues noussies h satiété, à 8.50C ( t i r é de Braaten 19841.
Les conditions de promiscuité decoulant des for tes densités d'élevage requihrent une benne connaissance des myens de prbsvention e t de traiteisent des rsraladies bacter iennes', virales au fangiques. Ha lheu~euse~n t , %es &thodes aises au peint pour le traitement des =ladies bactériennes des salmonides par exemple peuvent s'avérer totalement inutiles pour les poissons marins. Quoiqui il en soi t , l'dlevaqe requiert en partant une benne connaissance des maladies les plus communes. Or, peu de teavaux ont 4 t h publies sur les maladies de la aorue, que ce soi t sur les stocks sauvages ou SUE les mrtpes gardées en captivité, vraisemblableaient parce qu ' i l y a eu peu de travaux sur l a question. Seuleslent deux mladies ont étk rapportbes juaqulh maintenant. La plus doeimageable es t %a vibriese causbe par l a basthrie Vibrio anquillarun. Les sanaptbmes les p l u s communs sont des lesions cutanbes, des Euroncles e t des hkmorragies, l 'érosion de la nageoire caudale e t l'exophtalmie. Plusieurs souches semblent être en cause e t les traiteieents

t en tés A ce jour sont restés sans succés, mais les Norvégiens mettent présentement au point un vaccin (Egidius e t Andersen 1984). L'autre maladie rapportée e s t de nature v i ra le : l a nécrose érythrocytaire v i r a l e (Viral Erythrocytic Necrosis) e s t due h un virus de l a famille des Iridoviridae (ICDV). I l n'y a pas de mortalité associée A l a présence de ce virus, sauf peut-être chez l e s Salmonidae (Reno e t Nicholson 1981). Des morues inoculées avec ce virus ont développ4 la maladie en un mois, sont restées malades 3 mois, puis ont retrouvé leur santb, Le virus cause l'anémie e t perturbe l e métabolisr~e des érythrocytes sans causer de mortalites chez l a morue (Reno e t a l , 1986).
Mentionnons aussi que l a morue a tendance spontanément, SOUS l'impulsion d'un s t ress , A développer un probléme d'exophtalmie, par l'accumulation de gaz dans l ' o e i l . Comme c ' e s t le cas pour tous l e s poissons, l'exophtalmie peut & t r e causée par l a sursaturation en gaz de l 'eau ou par une brusque remontée vers l a surface au moment de l a p&che, mais chez la morue une perturbation des conditions d'élevage ou l e s t r e s s sous diverses formes peuvent entraf ner un déreglement dans l e fonctionnement de l 'axe pseudobranchie - anhydrase carbonique- glande choroide, ce qui cause une production
accrue de gaz dans l ' o e i l m4me (Dehadrai 1966). La blessure qui en résul te cause l a perte de l a vue e t pourrait constituer un s i t e d'entrée pour 1' infection bactérienne.
Parasites
Les parasites de l a morue sont indésirables dans l e contexte aquicole pour deux raisons. Parfois, ils peuvent entraîner la mort directement ou encore indirectement en rendant l es poissons plus vulnérables aux maladies. I l s peuvent aussi diminuer la valeur du produit de deux
maniéres, parce que leur présence répugne aux consoramateurs e t parce que l e coUt d'extraction e s t klevé. Comme c ' e s t souvent l e cas pour les organismes en milieu naturel, l a morue héberge une variéth de parasites (Margolis e t Arthur 1979; Margolis e t Kabata 1988).
Parmi ces parasites, w l g r é leur t a i l l e microscopique, l e s protozoaires ne sont pas les moins néfastes. Trichodina, u n c i l i e de la famille des Urceolariidai! (40-,100 Fm) a ét4 observe sur les branchies e t sur des lésions cutanées, causant l a mort de 20% d'un stock de géniteurs en cage (Egidius e t Andersen 1984). Le f l age l l é Trypanosoma murmanensis, un parasite du sang, a cause des troubles pers i s ten ts au niveau circulatoire: une augmentation de l'abondance des f lage l lés dans l e sang su i t e A une infestat ion expérimentale a entraf né une baisse de l 'hématocrite e t de l'hbmoglobine. Le parasi te e s t transmis A l a morue par l a sangsue marine Johanssonia a rc t ica , Des concentrations de l 'o rdre de 105 organismes par mL ont é t é observées (Khan 1976, 1971). Ce parasite e s t moins fréquent (prévalence 4 % ) dans l e Golfe du Saint-Laurent que dans 1 'Atlantique (Khan e t a l . 1980). Le microsporozoaire Lama morhua a également é t é signalé sous Eorme de kis tes sur l e s branchies, mais aussi sur l e s organes internes (Morrison 1983), mais on ne s a i t pas s ' i l cause des troubles pathologiques. C'est aussi un protozoaire, non-identifié, qui semble causer l e s lésions par des cel lules-x (Egidius e t a l , 1981; Vaterman e t Dethlefsen 1982). Les cas d ' infect ion sont plus fréquents chez les individus de moins de 40 cm e t près de l a cate ( 2 % dans l a Baie des Chaleurs, Horrison e t a l . 1982). Le parasi te provoque des -lésions semblables à des tumeurs, généralement au niveau des pseudobranchies. I l pourrait Btre la cause d'un ralentissement de l a maturation sexuelle e t d'un taux plus élevé de mortalité associée h

des infections bactériennes chez des morues gardbes en cap t iv i té (Morrison e t a l . 19821
Les parasites l e s mieux connus de l a morue sont l es nématodes de la famille des Anisakidae. Le ver de morue, Phocanema decipiens, commence son cycle dans l e phoque, principalement l e phoque g r i s : l a femelle l ibére quelques 400 mille oeufs (Odense 19781 qui éclosent, A des températures de 2 h 240C, en 5 A 20 jours. Les oeufs coulent de sorte que l e s larves qui emerqent se Eixent au substrat od e l l e s peuvent survivre jusqu'ëi 1 4 0 jours A une température de 5% Ces larves sont avalées par des copépodes benthiques (Harpacticoida, Cyclopoidal, jamais par des organismes planctoniques t e l s l es Calanoidea. La tzansmission ne s e f a i t pas directement au poisson (HcClelland 1982) . Ces copépodes sont A leur tour mangés par la morue ou d 'autre poissons t e l s l 'éperlan, Osmerus mordax, dont l a morue se nourrit . La larve pénètre ë i t ravers l ' e s t omc jusque dans l e muscle od e l l e reste enroulée (3-4 cm). Lorsque 1h morue e s t consomm4e par l e phoque, l a température interne élevbe du phoque déclenche l a mue e t l a maturation (Odense 1978). Le ver de morue n ' e s t pas dangereux pour l'homme; il meurt l o r s de l a congélation (24 h h -200C) ou de l a cuisson (7OoC pendant 7 minutes). I l e s t plus fréquent dans le sud du Golfe Saint-Laurent (Odense 1978; McClelland 1982). HcClelland e t a l . (19831 on% dénombré de O h 20 parasi tes par morue pour des morues de 20 A 80 cm, le nombre augmentant proportionnel- lement h l a t a i l l e du poisson. Seulement 20% des morues de 20 cm étaient infestées a lors que ce pourcentage e s t de 100% pour des morues de 80 cm. P. decipiens s e loge principalement dans les f i l e t s (86%), mais auss i dans les muscles recouvrant l'abdomen (11%) e t dans l a cavité coélomique ( 3 % ) . Anisakis sp., un autre nématode de la m6me famille, e s t moins fréquent e t se loge dans les mbsenthhres hépatiques e t viscéraux,
rarement dans l e s f i l e t s (3% de; cas) (McClelland e t a l . 1983).
Le crustacé Lernaeocera branchialis, comme son nom l'indique, s'accroche aux branchies. Les stades intermédiaires de ce parasite s e trouvent chez la poule de mer, Cyclopterus lumpus, h proximité des chtes, ce qui peut expliquer que ce sont surtout l es morues de moins de 50 cm qui sont parasitées (parfois 15% des individus). Le nombre de parasi tes par poisson e s t rarement plus que deux. Malqré cela, i l semble que Pa présence du. parasi te s o i t corrélée ii une perturbation du cycle de maturation sexuelle (Templeman e t a l . 1976). Voir Kabata (19881 pour plus de dé t a i l s sur ce parasi te .
DISCUSSION
L'élevage de l a morue franche présente de façon générale beaucoup moins d'inconnues que celui de nombreux autres poissons mar ins (Tableau 2, page 36). Csest dû en par t ie au f a i t que les premiers essa i s de psoduction de post-larves pour l e repeuplement remontent h au-delh de 100 ans. En Pcosse, où les conditions seraient idéales pour la production de morue, on estime que l'aquaculture intensive de l a morue présente peu de problPmes e t qu ' i l e s t possible de produire A pa r t i r dsinne morue de 5 g un poisson de 2 kg au bout de 18 mois. Soulignons l ' e s s a i r éa l i s é en Norvége O & une solution a étb envisagée par laquelle les larves sont élevbes en milieu lagunaire de fason plus économique vraisemblablement (Kvenseth e t Oiestad 1984; Oiestad e t a l . 1985). L'élevage complet ne semblant pas rentable, l e s producteurs se dir igent pàutdt vers l sengraissement de juvbni les prélevés dans l e milieu naturel. C'est l e cas dans l'Est du Canada (Williams e t Rlceniuk 1986, Fisher 1988).

Les mécanismes de déclenchement de la maturation sexuelle chez la morue ne di£ fèrent vraisemblablement pas de ceux d'autres poissons, mais lorsque l'on vise l'engraissement de morues juvéniles pour la commercialisation, il est nécessaire de connaître avec précision le moment et les conditions propices b ce déclenchement. L'inconvénient de la maturation sexuelle dans un élevage tient h ce que la maturation sexuelle entraîne une croissance négative et consequemment, une baisse de la qualité et du volume de production. Le probléme est documente (Braaten 1984), mais il reste h trouver des solutions si l'on veut commercialiser des morues de plus de 30 h 35 cm. Jusqu'A maintenant, on ne contrôle pas le cycle de maturation sexuelle de la morue, contrairement A ce qui est le cas pour le turbot européen par exemple chez lequel la maturation sexuelle peut, être provoquée en tout temps de l'année en manipulant la photopériode ou en injectant des hormones. Lorsqu'un poisson a une longue saison de reproduction, il est possible d'envisager se procurer en nature les génitkurs requis pour la fécondation artificielle et ce, sans risquer de rater une saison. C'est le cas de la Htorue qui présente un autre avantage, celui de frayer naturellement en élevage. Néanmois, une production intensive devra tat ou tard produire ses propres géniteurs et ceci requiert un bon contrale de la reproduction.
Bien que l'on connaisse en partie les besoins nutritionnels et environnementaux des larves de morue, des renseignements additionnels sont requis. La morue produit un grand nombre de petits oeufs pélagiques donnant de minuscules larves planctoniques. Elle est en cela comparable & la sole ou au turbot en Europe (Jones 1984). La période larvaire est critique car c'est au cours de cette période que se produisent le plus de mortalitds. Parmi les problèmes à résoudre, nous en mentionnerons trois principaux. Premièrement, le taux de mortalité n'est pas
le même d'un lot b un autre. Ceci est dû A une certaine variabilité de la qualit6 des gamètes, mais il n'existe pas de méthodes pour déterminer la qualité des oeuEs. Deuxiémement, pour les régions où l'eau de mer est diluée par des apports d'eau douce, l'incubation est problématique: l'embryon coule ce qui multiplie le risque que les maladies se propagent. Peut-être est-il possible de manipuler les geniteurs de maniére h ajuster la densité de l'oeuf, et donc sa tlottabilit&, sans modifier le taux de mortalité des embryons. La survie des embryons et des larves en eau saumatre n'a été évaluée que partiellement et les résultats obtenus sont contradictoires (Laurence et Rogers 1976; Davenport et al. 1981; Kjorsvik et Lonning 19831. Finalement, il faudrait trouver une diète de remplacement pour les larves et les morues juvéniles de moins de 3.5 cm. Les tentatives de remplacer la nourriture vivante, coûteuse en main d'oeuvre, par une moulée n'ont pas réussi jusqu'h present (Hjelnteland et al. 1984; Oiestad et al. 19851.
L'élevage des morues aux stades juvéniles et adultes ne présente pas de problèmes particuliers. Ces morues tolerent de larges gammes de conditions environnementales, ce qui est un avantage non négligeable si on en envisage l'élevage, particulièrement dans les eaux de surface. Les morues se nourrissent volontiers de poisson (hareng, capelan, etc.) et les taux de conversion observés sont excellents puisqu'ils varient de 2:l A 3:l.
Malheureusement, il n'existe que trtis peu de données sur les taux de mortalité observés a u cours des esvais d'engraissement. Le principal problème à. résoudre est sans doute celui du cannibalisme. Dans un bassin, il s'établit des hiérarchies de dominance entre les individus, les individus soumis ayant un

taux de croissance faible ou même nul. Les individus dont la croissance est nulle peuvent éventuellement être dévorés par les plus gros individus. Habituellement, ce problème est solutionné en séparant régul ibrement les poissons par classe de taille, ce qui requiert une manipulation des poissons. Des solutions alternatives pourraient être testées: trouver une cédule de nourr issage qui satisfasse les besoins des individus dominants puis ceux des individus soumis dans la dme journée, ou encore déterminer s1 il existe une densité au-delh de laquelle ce problème ne se pose plus,
L'augmentation de la densité ne va pas sans augmenter les risques de propagation des maladies. Or, il existe trés peu d'informations sur les maladies de Pa morue dans la litterature scientifique. Présentement, nous poursuivons A notre laboratoire, en collaboration avec l'Institut Armand-Frappier et 1 'école de MCdeelne vétérinaire de St-Hyacinthe, un programme ' visant A identifier les virus et bactCr ies pathogènes pouvant causer des problémes lorsque les poisson5 sauvages sont mis en stabulation.
L'engraissement de morues sauvages A partir de poissons tels le capelan ne permet pas de produire des morues exemptes de parasites. Dans un élevage intensif, la production de morues exemptes de parasites permettrait de réduire le coltt de la transformation et d'offrir un produit di£ fbrent au consommateur. Pour réduire 1 ' incidence du parasitisme dans un élevage de morues sauvages, on ne peut que sélectionne^ des tailles ou des stocks qui en sont relativement: exempts et les nourrir de poissons ou moulées dont les parasites ont été tués, par exemple par congélation.
L'élevage de la morue en milieu cdtier dans le Golfe du Saint-Laurent se heurtera au problème physique des glaces en hiver, et dans les cas où un engraissement sur une base saisonnière est envisagé, au fait que dans les milieux protégés, tels les estuaires des rivières ou par exemple la Baie des Chaleurs, la salinité des eaux de surface varie et est plutôt faible. La résistance de la morue aux basses temperatures de son environnement est bien connue. La morue sécrete une glycoprotéine plasmatique qui lui permet de survivre à des températures bien inférieures A OoC. Par contre, on ne connaît rien de sa capacité de tolérer des salinités infbrieures A la salinité de l'eau de mer. Les résultats préliminaires de tests effectués dans nos laboratoires, conjointement avec l'Université du Québec à Rimouski et l'Institut National de la Recherche Scient if igue, Océanologie, sont h cet Cgard prometteurs a Des norues tzansf &rées A divers temps de l'année h des eaux de salinités intermédiaires (7, 14 et 21 g/L) survivent, alors que des salinités plus faibles causent des mortalités, Sous rbaerve de tests en cours h des températures plus élevées et de comparaisons du taux de croissance entre ces traitements, il semble possible de conluxe que les milieux estuarien et cdtier sont propices A l'élevage de la morue sous ce strict point de vue. Plus généralement, il est permis de penser que bien que les poissons marins vivent dans des eaux dont la salinite se situe près de 32 g/L, ils pourraient très bien vivre A des eaux dont la salinité est inférieure, ce qui permet d'envisager divers sc4narios pour l'utilisation dseaux plus chaudes (eau de mer diluée avec des eaux douces plus chaudes) ou le choix de sites d'&levage en milieu cbtier.
Le Québec h 1' image du Canada est un exportateur de poisson. Cette caractéristique le soumet aux forces du marchb international. Toute fluctuation sur

l es marchés mondiaux, que ce s o i t au niveau de l ' o f f r e ou de l a demande de produits marins, a un impact d i rec t sur l es produits de l a pêche québécoise. Comme l 'aquaculture pourrait en quelque sorte compléter l a production du Québec, e l l e sera aussi soumise aux mêmes tensions du marché.
Dans l e cas de l a morue, l e contexte qui prévaut actuellement e s t ca rac té r i sé d'une part par une s t a b i l i t é des débarquements mondiaux e t d 'autre par t par une baisse de la demande. Cette dernihre étant induite par l a résis tance des consommateurs face A une hausse t rop rapide des prix de ces derniCres années. Cette combinaison de variables a t 8 t f a i t de f a i r e baisser l es prix de la morue sur l e marché. Cette tendance A l a baisse pourrait f a i r e place ci une certaine s t a b i l i t é des prix lorsque l e s inventaires seront revenus A des niveaux normaux.
Au Québec, certaines indications t e l l e s la baisse' des débarquements, l a ra re té de morue de for te t a i l l e e t l a baisse du taux de croissance des morues de nos stocks suggèrent une poss ib i l i t é de marché pour une production aquicole de morue franche.
Du c6té micro-économique, i l ex is te peu d'informations sur la r en t ab i l i t é de l 'élevage de l a morue. Jones dans une étude produite en 1984 r e l a t a i t tout de m&me qu'ci ce t te époque i l n ' é ta i t pas rentable d'élever de la morue en Bcosse. Malgré cela, ci Terre-Neuve, des entrepreneurs ont lancé en 1986 une entreprise d'engraissement de morue en milieu cBtier (Fisher 1988). Des morues capturées ci l a f in de la période de f ra ie ont & té transportées jusqulA des cages pour des essais d'engraissement entre j u i l l e t e t décembre, c'est-A-dire entre l e moment où les températures relativement chaudes de l ' é t é favorisent une croissance rapide e t l e moment où la croissance r a l en t i t
a lors que l 'énergie e s t h nouveau dir igée vers l a production des gonades. Le taux de mortalité au cours du transport a varié selon la méthode de capture e t selon l e temps requis pour parcourir la distance entre l e l i eu de pêche e t l e s i t e d'engraissement. La mortalité s2 produisait principalement dans l e s deux semaines suivant l a capture. Les morues recevaient environ 6% de leur poids par repas e t é taient nourries tous les 2 jours environ, généralement de capelan ou autres poissons. Les résu l ta t s de ces essa i s sont d i f f i c i l e s A évaluer puisque plusieurs facteurs n 'é taient pas sous contrale: la densi té n ' é ta i t pas l a m@me d'une cage A l ' au t r e , les t a i l l e s individuelles n 'htaient pas homoghnes d'od des problimes de cannibalisme, l es morues ont pu se nourrir de crustacés e t p e t i t s poissons que l e s mailles des parois des cages la i ssa ien t pénétrer, e tc . Les morues qui mesuraient 47.3 cm 11.07 kg) en j u i l l e t , en mesuraient 53.2 cm (1.89 kg1 en septembre e t 56.8 cm (2.02 kg) en décembre (Fisher 1988). Ce type de projet pourrait connaftre un cer ta in essor dans l ' aveni r . Cantin e t Bourdages (19891 ont abordé ce t te question dans l e contexte québécois.
Ali, H.A., 1971. Les réponses rétinomo- t r ices: caractères e t mécanismes. Vision Res. 11: 1225-1288.
Baird, J.W., C.A. Bishop e t R. Wells, 1986. Estimates of age and length a t maturity for cod in NAFO Divisions 2Jt3KLNO. HAFO SCR Dac, 86/1?2. Serial No N 1251: 3 p.

Beacharn, T.D., 1983a. Grovth and maturity of At lant ic cod (Gadus morhua) i n the southern GulE of S t . Lawrence. Can. Tech. Rep. Fish. Aquat. Sci . 1142: i v t 31 p.
Beachaa, T.D., 1983b. Var iab i l i ty in rsedian s i z e and age a t sexual maturity of At lant ic cod, &dus rmrhua, on the Scotian shelf i n the Northvest At lan t i c Ocean. Fish, B u l l , 81: 303-321.
Bohle, B., 1974. Temperaturpreferanse hos tork (Gadus morhua) . Fisken Hav. 3er. B 20: 1-28.
Braaten, B., 1984. Grovth of cod i n r e l a t i o n t o f i s h s i z e and r a t i o n ievel , p. 677-712. In: B. Dahl, D.3. Danielssen, E. Hoksness e t P. S o l e d a l (Ris), The propagation of cod a d u s morhua. Flodevigen rappor t s e r . 1.
Braaten, B., e t 3.L. Gokstad, 1980. Appetite feeding experiments with cod-preliminary r e s u l t s . 1 .C.E.S. C.H. 1980/F: 20.
Braun, V.H., 1961a. Aggressive behaviour in the cod (Gadus c a l l a r i a s ) . Behaviour 18: 107-147.
Braun, V.H., 1961b. Reproductive behaviour of the cod (Gadus c a l l a r i a s ) . Behaviour 18: 177-198.
Bravn, V.H., 1969. Peeding behavior of ced (Gadus; morhua). J. Fish. Res. Board Can. 26: 583-596.
Broaley, P.J., e t P.A. Sykes, 1985, üeaning d i e t s f o r tu rbo t (Ssopththalmus mximus), s o l e (Solea so lea ) and cod (Gaàus m r h u a ) , p. 191-211. In: C.B. Covey, A.H. Mackie, and J.G. Bell (Eds.), Nutr i t ion and feeding in f i sh . Academic Press, New York. 489 p.
Buckley, L.J., 1979. Relationships betwen RNA-DNA r a t i o , prey density, and qrowth r a t e in At lan t i c cod (Cadus mrhua) larvae. J. Fish. Res. Board Can. 36: 1497-1502.
Buckley, L. J., 1981. Biochernical changes during ontogenesis of cod (Gaduç rwrhua) and v in te r f lounder (Pseudopleuronectes amer icanusl larvae . Rapp. P.-v. Rbun. Cons. i n t . Explor. #er 178: 547-552.
Buckley, L.J., 1984. RNA-DNA r a t i o : an index of l a r v a l Eish growth in the sea. Har. Biol. 8Q: 291-298.
Cantin, C. e t J. Bourdages, 1989. F a i s a b i l i t b bconomique d'un blevage-type de morue sur l a basse Ciite-Nord. Rapport d 'analyse 4conomique e t co l s~~erc ia le no. 27. ISBN 0-662-95599-4: 61 pages.
Craik, C A e t S.M. Harvey, 1984. Biocheaical changes occurring during f i n a l maturation of eggs of soeie marine and Ereshvater t e l eos t s . J. Pish Biol. 24: 599-610.
Craik, J.C.A., e t S.H. Harvey, 1987. The causes of buoyancy in eqgs of marine t e l e o s t s . J. m r . b iol . Ass. U.K. 67: 169-182.

Dahl, E. , D.S. Danie l ssen , E. Hoksness e t P. So lemfa l ( E d s ) , The propaqa t ion of cod Gadus morhua. Flodevigen r a p p o r t s e r , 1, 895 p.
Davenport, J., S. Lonninq e t E, Kjorsv ik , 1981. Osmotic and s t r u c t u r a l changes d u r i n g e a r l y development of eggs and l a r v a e of t h e cod, Gadus morhua. J . F i s h B i o l , 19: 317-331.
Dehadrai, P.V., 1966. Mechanisus of qaseous exophthalmia i n t h e A t l a n t i c cod, Gadus morhua. J . F i sh . Res, Bd. Canada 23: 909-914
D i v i s i o n de l a s t a t i s t i q u e e t d e l ' i n f o r m a t i q u e , 1988. Revue s t a t i s t i q u e a n n u e l l e 1984-1987. H i n i s t e r e d e s P@ches e t d e s Océans, Canada. ISBN 0-662-95010-0.
Edwards, ~ . R . c . , D.H. Finlayson, e t J.H. S t e e l e , 1972. An exper imenta l s t u d y of t h e oxygen consumption, growth, and metabol ism of t h e cod (Gadus morhua) . J. exp. mar. Bio l . Ecol. 8: 299-309.
Eqid ius , E., e t K . Andersen, 1984. Disease problems i n cod r e a r i n q , p . 761-772. In: E. Dahl, D.S. Danielssen, E. Moksness e t P. Solemdal ( E d s ) , The propaga t ion of cod &dus morhua. F lodeviqen r a p p o r t s e r . 1.
Eqid ius , E.C., J .V. Johannessen, e t E. Lange, 1981. Pseudobranchial tumours i n A t l a n t i c cod, Gaduç morhua, from t h e Baren ts Sea. 3 . F i s h Dis. 4: 527-532.
p r o t e i n and water c o n t e n t of cod, Gadus morhua, muscle from Northern Norway, J . P i s h B i o l . 20: 527-533.
E l l e r t s e n , B . , P. Possum, P. Solemia l , S. Sundby, e t S. T i l u e t h , 1984. A c a s e s t u d y on t h e d i s t r i b u t i o n of cod l a r v a e and a v a i l a b i l i t y of p r e y o r g a n i s u s i n r e l a t i o n t o p h y s i c a l p r o c e s s e s i n Lofoten, p. 453-478, In: E. Dahl, D.S. Danie l ssen , E. Moksness e t P . Solemdal ( E d s ) , The propaqa t ion of cod, Gadus morhua, F lodevigen r a p p o r t u e r .
E l l i n q s e n , O.F., e t K.B. Dovinq, 1986. Chernical. f r a c t i o n a t i o n of shr imp e x t r a c t s ' induc inq bot tom food s e a r c h behaviour i n cod (Gadus morhua 1. J . Chem. Ecol . 12: 155-168.
Fahay, H.P., 1983. Guide t o t h e e a r l y s t a g e s of marine f i s h e s o c c u r r i n g i n t h e Western North A t l a n t i c Ocean, Cape H a t t e r a s t o t h e Southern S c o t i a n S h e l f . J , Northw. A t l . F i s h . S c i . 4: 423 p.
F iander - Good A s s o c i a t e s Ltd, 1988. Economic assessment of salmonid cage c u l t u r e i n d u s t r y i n s o u t h westhern New Brunswick : 104 pages.
F i s h e r , R . 1988. Assessments and o b s e r v a t i o n s of a cod farminq o p e r a t i o n i n Newfoundland. Can. Ind. Rep. F i sh . Aquat. S c i . 194: v i i t 8 1 p.
E l i a s s e n . J.-E., e t O. Vahl, 1982. Seasona l v a r i a t i o n s i n t h e gonad s i z e and t h e

Gamble, J.C., e t E.D. Houde, 1984. Growth, mor t a l i t y and feeding of cod (Gadus morhua) l a rvae i n enclosed water c o l u m s and In labomatory tanks , p. 123-144. In : B. Dahl, D.S. Danielssen, E. Hoksness e t P. Solemdal (Eds) , The propagation of cod, Gadus morhua. Flodevigen rappor tser . 1.
Hawkins, A.D., N.H. Soof i an i , e t G.W. Smith, 1985. Growth and feeding of j uven i l e cod {Gadus morhua). J, Cons, i n t . Explor. Mer 42: 11-32.
Haelmeland, K,, E , Huse, T. Jorgensen, G. Molvik, e t J . Raa, 1984. Tfypsin and trypsinogen a s indices of growth and s u r v i v a l p o t e n t i a l of csd (Gadus morhua) la rvae , p. 189-202. In: E. Dahl, D.S. Danielssen, E. Hoksness e t P . Solemdal {Eds), The propagation of cod, Gadus morhua. Flodevigen r appor t se r , 1.
Holdvay, D.A., et, F.W.H. Bearaish, 1984. - f p e c i f i c gaowth r a t e an6 proximate body composition of A t l an t i c cod (Gadus momhua]. J . Exp. Mar. BioP. Ecol. 81: 147-170.
Holdvay, D.A., e t F,W,H. Beamish, 1985. The e f f e c t of growth r a t e , s i z e and season on oocyte development and matur i ty of A t l a n t i c cod (Gadus mor-hua). J. Exp. Hat. Biol. Ecol. 85: 3-19,
Houlihan, D.F., 3 . J , Hall, C. G~ay, e t B.S. Noble, 1988. Growth r a t e s and p r o t e i a turnover in At l an t i c cod, Gadus morhua. Can. J . Fish. Aquat. Sc i . 45: 951-964.
Hovcll, B , R . , 1973. Marine f i s h c u l t u r e i n E r i t a i n VIII , A marine r o t i f e r , Baachionus p l i c a t i l i s , and the l a rvae of the mimssel, Hy t i lu s edu l i s , a s foods fo r l a r v a l f l a t f i s h . J . Cons. i n t . Explor. Mer 35: 1-6.
Howell, B.R., 1984. The in t ens ive r ea r ing of juveni le cod, Gadus morhua, p . 657-676. In: E. Dahl, D.S. Danielssen, E. Moksness e t P. Solemdal (Eds), The propagat ion of cod Gadus rnorhua, Flodevigen rappor b e r . 1.
Iversen, $ , A , , e t B,S. Banielssen, 1984, DevePspaient and m o r t a l l t y of cod fGadus morhua) egqs and l a rvae in d i f f e r e n t temperatures, p. 49-66. In: E. Dahl, D.S. Danielssen, E. Moksness e t P. Solendal (Edsf , The propagation of cod &dus msrhua. Flodevigen rappor tser . 1.
Jean, Y., 1964. Seasonal d i s t r i b u t i o n of cod (Qàus morhuaf along the canadïan At l an t i c r o a s t i n r e l a t i o n t o va t e r temperature, 9. Pish , Res, Bd. Canada 21: 429-460.
Jobling, H., 1982. Food and gaowth r e l a t ionsh ips of t he cod, &dus morhua, with s p e c i a l r e fe rence t o Balsf jorden, nor th Norway. J . Fish Biol . 21: 357-371.
Jobling, ii, 1983, Growth s t u d i e s with %ish-overcominq the problems of s i z e v a r i a t i o n , J . F i s h Eliol. 22: 153-157.

Jones , A., 1984. Does cod farminq have a commercial f u t u r e ? A t e c h n i c a l and economic assessment , p. 773-786. I n : E. Dahl, D.S. Danielssen, E. Moksness e t P. Solemdal (Eds) , The p r o p a g a t i o n of cod, Gadus morhua. F lodevigen r a p p o r t s e r , 1.
Jones, R . , e t J.R.G. Hiulop, 1978. P u r t h e r o b s e r v a t i o n s on t h e r e l a t i o n between food i n t a k e and growth of qadoids i n c a p t i v i t y . J. Cons. i n t , Explor . Mer. 38: 244-251,
Kabata, 2., 1988. Copepoda and Branchiura , p. 3-127. In: L. Margol is , e t 2. Kabata ( E d s ) , Guide t o t h e p a r a s i t e s of f i s h e s of Canada. P a r t II - C r u s t a c e a . Can. i p e c . Publ . P i sh . Aquat. i c i , 101: 184 p.
Khan, R.A., 1976. The I i f e . c y c l e of Trypanosoma murmanensis. Can, J. 2001. 54: '1840-1849.
Khan, R.A. , 1977. Blood changes i n A t l a n t i c cod (Gadus morhua) i n f e c t e d w i t h Trypanosoma murmanensis. J . F i s h . Res . Board Can. 34: 2193-2196.
Khan, R.A., J . Murphy, e t D. Taylor , 1980. Preva lence of trypanosoœe i n A t l a n t i c cod ( h d u s morhua) e s p e c i a l l y i n r e l a t i o n t o s t o c k s i n t h e Newfoundland a r e a . Can. J. F i s h . A q u a t . S c i , 37: 1467-1475.
Kjorsv ik , E., A. S tene , e t S. Lonninq, 1984. Morpholoqical, p h y s i o l o g i c a l and q e n e t i c a l tud dies of eqq q u a l i t y i n cod (Gadus morhua), p. 67-86. In: E. Dahl, D.S. Danielssen, E. Moksness e t P. Solemdal (Eds) , The propaga t ion of
cod, Cadua murhua. Plodeviqen r a p p o r t s e r . 1.
Kjorsv ik , E., e t S. Lonninq, 1983. E f f e c t s of egg q u a l i t y on normal f e r t i l i z a t i o n and e a r l y development of t h e cod, &dus morhua. J . P i s h B i o l . 23: 1-12.
Knutsen, G.M., e t 5. T i l s e t h , 1985. Growth, development, and f e e d i n q s u c c e s s of A t l a n t i c cod l a r v a ~ Gadus morhua r e l a t e d t o eqq s i z e . T r a n s , Am. P i s h , Soc. 114: 507-511.
Kohler, A.C., 1964. V a r i a t i o n s i n t h e qrowth of A t l a n t i c cod (Gadus morhual . J. F i s h . Res. Bd, Canada 21: 57-100.
Kuf t ina , N.D., e t G.G. Novikov, 1986. Embryonic qrowth and u t i l i z a t i o n of yolk s t o r a g e p r o t e i n s d u r i n q e a r l y ontogeny of cod, Gadus morhua, a t d i f f e r e n t t empera tures . J . I c h t h y o l . 26: 76-88.
Kvenseth, P.G., e t V. Oies tad , 1984. Large s c a l e r e a r i n g of cod f r y on t h e n a t u r a l food p r o d u c t i o n i n a n e n c l o s e d pond, p. 645-666. In: E. Dahl, D.3, Danie l ssen , E. Moksness e t P. Solemdal (Eds) , The propaga t ion of cod, Qdus morhua . Plodevigen r a p p o r t s e r . 1.
L a l a n c e t t e , L.-M., 1984. Cro issance , r e p r o d u c t i o n e t régime a l i m e n t a i r e de l a morue, Gadus morhua, v i v a n t d a n s l e F j o r d du Sagnenay a u Québec. Canadian F i e l d - N a t u r a l i s t 98: 305-314.

Laurence, G.C., 1978. Comparative growth, r e s p i r a t i o n and de layed Eeedinq a b i l i t i e s of l a r v a l cod (Gadus morhua) and haddock (Melanogrammus a e g l e f i n u s ) a s i n £ luenced by tempera ture d u r i n g l a b o r a t o r y s t u d i e s . Mar. B i o i . 50: 1-7.
Laurence, G.C., 1979, Larva l l eng th-weiqh t r e l a t i o n s f o r seven s p e c i e s of Northwest A t l a n t i c E ishes r e a r e d i n t h e l a b o r a t o r y . P i s h . Bul l . 76: 890-895.
Laurence, 6.C. e t C.A. Rogers, 1976. E f f e c t s of t empera ture and s a l i n i t y on compara t ive embr yo deve lopment and m o r t a l i t y of A t l a n t i c cod (Gadus morhua) and haddock (Nelanogrammus a e g l e f i n u s ) . J. Cons. i n t . Explor . Uer 36: 220-228.
Lauz ie r , P., 1988. Analyse &cononrique e t commerciale du po isson d e fond a u Québek. U i n i s t h r e d e s Péches e t d e s Océans. D i v i s i o n d e s S c i e n c e s économiques, Canada. ISBN 0-662-95176-x: 25 pages.
Lough, R.G., 1984. Larva l f i s h trophodynamic s t u d i e s on Georges Bank: samplinq s t r a t e g y and i n i t i a l r e s u l t s , p. 395-4134. I n : E. Dahl, D.Y. Danielssen, E. Noksness e t P. S o l e e d a l (Eds 1, The propaga t ion of cod, Gadus morhua. Flodevigen r a p p o r t s e r . 1.
Hakhotin, V.V., G.G. Novikov, S.G. Soin, e t V.N. Tirrreiko, 1984, The p e c u l i a r i t y of t h e development of White Sea cod, p. 105-122. In : E. Dahl, 5.3. Danielssen, E. Uoksness e t P. S o l e e d a l ( E d s ) , The propaga t ion of cod Gadus morhua . Flodevigen r a p p o r t s e r . 1.
Marqolis , L . e t J.R. Ar thur , 1979. Synops i s of t h e p a r a s i t e s of f i s h e s oE clanada, B u l l . F i s h . Res. Board Can. 199: 269 p.
Margolis , L., e t Z . Kabata , 1988. Guide t o t h e p a r a s i t e s of f i s h e s of Canada. P a r t I I - C r u s t a c e a . Can. Spec. Publ . F i s h . Aquat. Sci. 101: 184 p.
May, A.W., 1967. P e c u n d i t y of A t l a n t i c cod. J . P ish . Res. Bd. Canada 24: 1531- 1551,
HcClel land, G a , 1982, Phocanema d e c i p i e n s f Nematoda : Anisakinae 1 : exper imenta l i n f e c t i o n s i n m r i n e copepodes, Can, J. Zool. 60: 502-509.
McClelland, G., R.K. Misra , e t D.9. Marcoqliese, 1983. V a r i a t i o n s i n abundance of l a r v a l a n i s a k i n e s , sea lvorm (Phosanema d e c i p i e n s ) and r e l a t e d s p e s i e a i n S c o t i a n She l f (4Vs and 4V) cod and f l a t f i s h . Can. Tech. Rep. P i s h . Aquat. S c i . 1202: i x t. 27 p.
Uor r i s son , C.M., 1983. The d i s t r i b u t i o n of t h e m i c r o s p o r i d i a n Loma morhua i n t i s s u e s of t h e sod Gadus morhua. Can. J . Zeol. 61: 2155-2161.
Morrisson, C.H., G. Shum, R.G. Appy, P. Odense, e t C. Arinand, 1982, His to logy and p r e v a l e n e e of X-cel l l e s i o n s i n A t l a n t i c cod tGadus morhua). Can. J , F i s h . Aquat. S e i . 39: 1519-1530.

Hyrseth, B . , 1988, Congress Proceed ings . Aquacul ture I n t e r n a t i o n a l Congress and Expos i t ion , Vancouver, 6-9 September 1988.
Odense, P.H., 1978. Some a s p e c t s of t h e codworm problem. F i sh . Mar. i e r v . Ind. Rep. 106: 20 p.
O i e s t a d , Va, 1984. C r i t e r i a f o r c o n d i t o n evolved from e n c l o s u r e exper iments w i t h cod l a r v a popula t ions , p. 213-230. In: E. Dahl, D.S. Danie l ssen , E. Moksness e t P. io lemdal (Eds) , The p r o p a g a t i o n of cod, Gadus morhua. F lodevigen r a p p o r t s e r , 1.
Oies tad , V., P.G. Kvenseth, e t A. Folkvord, 1985. Hass p roduc t ion of A t l a n t i c cod j u v e n i l e s Gadus morhua i n a Norwegian s a l t v a t e r pond. Trans. Am. F i s h . Soc. 119: 590-595.
Olafsen , J.A., 1984. I n g e s t i o n of b a c t e r i a by cod (Gadus morhua) l a r v a e , p . 627-644. In: E. Dahl, D.3. Danie l ssen , E. Moksness e t P. Solemdal (Eds l , The propaga t ion of cod, Gadus morhua . Flodevigen r a p p o r t s e r . 1.
Powles, P .M., 1958. S t u d i e s of r e p r o d u c t i o n and f e e d i n g of A t l a n t i c cod (Gadus c a l l a r i a s ) i n t h e Southves te rn Gulf oE S t . Lawrence. J. F i s h . Res. Bd. Canada 15: 1383-1402.
Raae, A.J., 1. Opstad, P. Kvenseth, e t B.T. Walther, 1988. RNA, DNA and p r o t e i n d u r i n g e a r l y development i n f e e d i n g and s t a r v e d cod (Gadus morhua) l a r v a e . Aquaculture 73: 247-259.
Reno, P.W., K . K l e f t i s , $.Y, S h e r b u r n ~ , e t B.L. Nicholson, 19 86. Experimental i n f e c t i o n and p a t h o g e n e s i s of v i r a l e r y t h r o c y t i c n e c r o s i s (VEN) i n A t l a n t i c cod, &dus morhua, Can, J . P i s h . Aquat. S c i . 43: 945-951.
Reno, P.W., e t B.L. Nicholson, 1981. U l t r a s t r u c t u r e and preva lence of v i r a l e y t h r o c y t i c n e c r o s i s (VEN) v i r u s i n A t l a n t i c cod, Gadus morhua, £rom t h e n o r t h e r n A t l a n t i c Ocean. J . F i s h Di;. 4: 361-370.
Riis - Veste rgaard , J., 1984. Water ba lance i n cod eqgs, p . 87-104. I n : E, Dahl, D.S. Danie l ssen , E. Moksness e t P. Solemdal ( E d s ) , The propaga t ion of cod, Gadus morhua. Plodevigen r a p p o r t s e r . 1.
Riley, J . D . , e t W.G. P a r n e l l , 1984. The d i s t r i b u t i o n of young cod, p. 563-580. In: E. Dahl, D.S. Danie l ssen , B. Moksness e t P. Solemdal (Eds) , The propaga t ion of cod, Gadus morhua. Plodeviqen r a p p o r t s e r . 1.
Saunders, R.L., 1963. R e s p i r a t i o n of A t l a n t i c c o d . J. F i s h . Res. Bd. Can. 20: 373-386.
S c o t t , J .S. , 1982. Depth, t empera ture and s a l i n i t y p s e f e r e n c e s of common f i s h e s of t h e S c o t i a n s h e l f . J . Northw. Atl. F i s h . S c i . 3: 29-39.
S c o t t , W.B., e t H.G. S c o t t , 1988. A t l a n t i c f i s h e s of Canada. Can. Bull . P i s h . Aquat. i c i . 219: 731 p.


W i l l i a m , P . , e t J,W. Kiceniuk, 1986. #@eding and growth rates i n A t l a n t i c cod (Gadus morhua) he ld in capt i v i t y d u r i n g t h e summer-autumn months. Can. Tech. Rep. F i s h . Aquat, S c i , 1466: i v t 10 p.
Yin, M.C., e t J.H.$. Blax te r , 1986. Morphological changes d u r i n g growth and s t a r v a t i o n of l a r v a l cod (Gadus morhua) and f l o u n d e r ( P l a t i c h t h y s f l e s u s ) . J . Exp. Mar. B i o l . Ecol . 104: 215-228.
Yin, H.C., e t J.H.S. Blax te r , 1987a. Peeding a b i l i t y and s u r v i v a l d u r i n g s t a r v a t i o n of marine f i s h l a r v a e r e a r e d i n t h e l a b o r a t o r y , J . Exp. Mar. B i o l . Ecol . 105: 73-83,
Yin, M.C., e t J.H.$, Blax te r , 1987b. Temperature, s a l i n i t y t o l e r a n c e , and buoyancy d u r ing e a r l y development and s t a r v a t i o n of Clyde and North Sea h e r r i n g , cod, and f lounder l a r v a e . J . Exp. Mar. B i o l . Ecol . 107: 279-290.

Tableau 2 , ROsumC des caractéristiques biologiques du cycle de vie de la morue franche.
A. L'embryon
Taille de l'oeuf: 1.2 - 1.6 m.
Incubation: POlagique . (Shelbourne 1964, Thompson et Riley 1981, Kjorsvik et Lonning 1983, Howell 1984, Solberg et Tilseth 1984).
Durée fonction de la température. (Laurence et Rogers 1976, Tkompson et Riley 1981, Kjsrsvik et Novikov 1986).
Dkveloppement : Norml entre I et PZOC. (Thcampson et Riley 1981, Iversen et Danielssen 1984, Hakhotin et al. 1984, Kuftina et Novikov 1986).
Varie avec 1 ' éclairage. ,
(Solberg et Tilseth 1984).
Tolére l'eau saumAtre. (Laurence et Rogers 1976, Davenport et al. 1981, Kjorsvik et al. 1984).

B. Larve vitelline et ~ostlarve
Taille A ltéclosion: 4.5. mm / 200 pq. (Olafsen 1984).
Développement : Hétamorphose en 44 jours A lOoC et en 52 jours A 70C. (Laurence 1978).
Nourriture: Sac vitellin (7 jours); alimentation exogene dPs le 4e ou 5e jour. (Laurence 1978, Gamble et Houde 1984, Yin et Eaxter 1987a)
RotifPres, nauplii et métanaulii dtArtemia, (Howell 1973, 1984, Thompaon et Riley 1981, Solberg et Tilseth 1984, Tilseth et Ellertsen 1984).
Taille des proies fonction de la taille de la larve. (Lough 1984).
Consommation de 1 nauplii & l'heure jusquf h concurrence de 3 ou 4. (Tilseth et Ellertsen 1984 1 .
Taux de survie: 10% jusqulA la metamorphose et 5 A 7% en 72 jours, jusqufA 508. (Howell 1984, Kverseth et Oiestad 1984, Oiestad et al 19851,
Facteurs: Diete inadhquate, cannibalisrse. (Howell 1984).
Température de plus de 16 A 180C et salinites de moins de 2-3 g/L. (Yin et Blaxter 1987b).

C. Morues juveniles et adultes
Taille A la mbtamorphose: 1i)-12 mm et 1000 h 1400 pq (poids sec), (Laurence 1978, Kvenseth et Oiestad 9985, Oiestad et al. 1985).
Mourr i ture:
Cannibaf isme:
Moulee accept4e h partir de 2.5-3.0 cm. (Bromley et Sykes 1985).
Glycine et alanine déclencheurs du comportement d'alimentation. (Ellingsen et Doving 19861,
Ponction de la taille. (Waiwood et Hajkowski 1984, Bromley et Sykes 1985).
Fonction de la ration. (aiestad e t al. 1985).
Consommation dloxygéne: Mesurbe en fonction du poids et de %a tempbrature. (Saunders 1963, Braaten 19841,
Mesurée en fonction de l'alimentation, (Soofiani et Hawkins 19821,
Ration de base:
Ingestion:
Déterminée en fonction du poids du poisson. (Jones et Hislop 1978, Jobling 1982, Braaten 1984).
Déterminée en fonction. de la température. (Hawkins e t al, 1985).
Fonction du poids du poisson. (Jobling 1983).
Fonction de la tempbrature . ( Saunders 1963).

C, Morues juv4niles et adultes (suite)
Ingestion (suite): Fonction de la cédule d'alimentation. (Braaten et Gokstad 1980).
Digestion: Rythme fonction de la température. (Tyler 1970).
Taux de conversion: Maximum h des rations intermédiaires. (Eiwards et al. 1972 1.
Diminue avec la taille du poisson. (Jones et Hislop 1978, Joblinq 19821.
Fonction de la diilte, (Hove11 1984).
Coût d'assimilation varie de 3 A 20% de l'ingestion. (Soofiani et Hawkins 1982).
Croissance :
Haladies:
Rapide (taille de 34 cm en 60 semaines). (Braaten 1984, Howell 1984, Oiestad et al. 1985, Williams et Kiceniuk 1986).
Maximale h 13-150C. (Tyler 1970, Jobling 1983).
Augmente en fonction de la ration. (Edwards et al. 1972, Jones et Hislop 1978, Braaten 1984, Hawkins et al. 1985, Houlihan et al. 1988).
Diminue avec la taille (2%/jour, { 90 g; O.Z%/jour, ) 800 g). (Kohler 1964, Jobllinq 1983, Braaten 1984, Howell 19841.
Vibriose (mortelle). (Eqidiuü et Andersen 1984).

C. Morues juvéniles et adultes (suite)
Maladies (suite): Necrose erythrocytaire virale. (Reno et Nicholson 1981, Reno et al, 19861.
Exophthalmie . (Dehadrai 1966).
Parasites:
Comportement:
Trichodina sp. (mortel j. (Egidius et Andersen 1984 1.
Trypanosoma murmnens is . (Khan 1976, 1977).
Lom norhua. (Morrlsson 1983).
Phocanenta deci piens (ver de morue 1 . (Odense 1978, Mc Clelland 1982, Hc Clelland et al. 1983).
Lernaeocera branchial is. (Templeman et al. 1976, Kabata 1988).
Hiérarchies e t agressivité. (Braun l961ai.

il, Maturation sexuelle
Taille h la maturité sexuelle: { 10% h 30 cm, 100% h 50 cm. (Beacharn 1983a,b; Baird et al. 19861.
Croissance : NBgat ive. (Braaten 1984).
Saison de reproduction: De janvier A juin dans l'Atlantique, de mai A l'automne dans le Golfe. (Powles 1958, Fahay 1983, Scott et Scott 1988).
Fécond i t4 : Trbs élev4e. (May 1967).
Pbcondation artificielle: Techniques habituelles. (Shelbourne 1964, Iversen et Danielssen 1984, Kjorsvik et Lonning 1984).
Fécondation naturelle:
Comportement :
Spontanée en bassin. (Laurence e t Roqers 1976, Jones 19841.
Gametes libéres sur une période de 2 A 3 semaines, (Shelbourne 1964, Holdway et Beamish 1985).
Hibrarchies et agressivité. (Brawn 1961a,b).
