key mechanisms of systemic rna interference in the desert...
TRANSCRIPT

Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Vegetal
Key mechanisms of systemic RNA interference in the desert locust
Dulce Cordeiro dos Santos
Dissertação de Mestrado
Mestrado em Biologia Molecular e Genética
2013

Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Vegetal
Key mechanisms of systemic RNA interference in the
desert locust
Dulce Cordeiro dos Santos
Dissertação de Mestrado orientada pelo Prof. Doutor Jozef Vanden Broeck e com a orientação interna do Prof. Doutor
Jorge M.L. Marques da Silva
Mestrado em Biologia Molecular e Genética
2013

Acknowledgments In the first place, I would like to thank my promoter Professor Doctor Jozef Vanden Broeck for the opportunity to join the research group of Molecular Developmental Physiology and Signal Transduction of KU Leuven and develop this project in such an enthusiastic research environment. Secondly, I want to thank my mentor Niels Wynant for all the teachings, interesting scientific discussions and the very careful reading and suggestions to this report, as well as for the endless patience and the contagious enthusiasm concerning the research work. In addition, I would like to thank for the words of support in some difficult times. In the third place, I would like to thank Professor Doctor Jorge M. L. Marques da Silva for accepting to be my internal supervisor and for the prompt help in every situation. I would like to thank everyone in the Molecular Developmental Physiology and Signal Transduction research group for the affection and good working environment, in special to Doctor Liesbeth Badisco for guiding me in my first contact with insect research. I want to thank all my friends in Portugal, particularly Rita Mansilha, Mariana Inácio and Patrícia Silva for the constant devotion and friendship. Moreover, I address my word of thanks to my friends from Leuven, for all the support during the past academic year. I also thank my family and family friends, especially my brother Jorge Santos and my grandmother Dulce Cordeiro, for always making me feel loved and cared. I would like to address a very special word of thanks to my mother Maria Dulce Santos, for the unconditional love and constant support. I am also very thankful for the example of strength and moral values throughout my life. I want to dedicate this important moment to her. Finally, I would like to thank João Nery for all the love and dreams came true, that allowed me to face this challenging year with a big smile and a lot of joy in my heart.
To you all, Thank you very much.

Resumo
O RNA de interferência (RNAi) é um mecanismo de silenciamento genético
desencadeado por estruturas de RNA de cadeia dupla (dsRNA). Este mecanismo apresenta
uma importante função antiviral e, devido à sua elevada especificidade, capacidade de
silenciamento e potencial efeito sistémico, tem-se demonstrado uma eficaz ferramenta de
indução de perda-de-função em investigação. Além disso, este mecanismo tem sido proposto
como potencial pesticida para o controlo de pragas agrícolas.
A resposta RNAi é muito variável, quer entre espécies, quer dentro da mesma espécie,
podendo variar consoante o tecido, estado de desenvolvimento e método de entrega do
dsRNA. Contudo, os mecanismos envolvidos têm sido essencialmente estudados em
Drosophila melanogaster, que possui uma sensibilidade baixa ao RNAi (sistémico). Por outro
lado, o gafanhoto do deserto, Schistocerca gregaria, demonstra uma resposta RNAi de
elevada robustez e sensibilidade. Neste contexto, uma vez que esta espécie constitui uma
praga voraz e que o RNAi pode contribuir para estratégias seletivas de controlo de pragas
agrícolas, o gafanhoto do deserto constitui um organismo muito interessante para a
investigação dos mecanismos do RNAi (sistémico).
Em S. gregaria, a injeção de dsRNA é um método eficaz para a indução de uma
resposta sistémica de interferência mas, por outro lado, a entrega de dsRNA via oral não
resulta em silenciamento genético. Estudos realizados no nosso laboratório demonstraram a
existência de atividade de degradação de dsRNA no suco do intestino médio do gafanhoto do
deserto e identificaram, com base numa dsRNase isolada do suco digestivo de Bombyx mori,
quatro dsRNases no transcritoma de S. gregaria (denominadas dsRNase-1, -2, -3 e -4). Desta
forma, o primeiro objetivo desta tese consistiu em identificar as nucleases responsáveis pela
atividade degradadora de dsRNA no suco do intestino médio nesta espécie. Neste contexto,
um perfil de transcrição das dsRNase-1, -2, -3 e -4 foi realizado e, de acordo com os nossos
resultados, estas são maioritariamente expressas no intestino médio do gafanhoto do deserto,
o que está de acordo com a hipótese destas nucleases desempenharem um papel na
degradação do dsRNA no suco digestivo. Para testar esta hipótese, a tecnologia do RNAi foi
usada para proceder ao “knockdown” das quatro dsRNases individualmente, através da
injeção de dsRNA específico no hemocélio de gafanhotos adultos. Uma forte diminuição
transcricional de cada dsRNase foi obtida mas, devido a uma grande similaridade entre as
sequências disponíveis, foram também observados efeitos “off-target” nos gafanhotos
tratados com dsRNA específico para a dsRNases-1 e -2. Porém, os efeitos “off-target” nunca

foram tão fortes como os efeitos de silenciamento específico. Notavelmente, a atividade de
degradação de dsRNA observada no intestino médio dos gafanhotos injetados com dsRNA
para a dsRNase-2 foi altamente comprometida, o que sugere que a dsRNase-2 contribui
fortemente para a degradação do dsRNA no intestino médio de S. gregaria. Contudo, uma vez
que neste grupo também se observou uma sub-regulação significativa da dsRNase-1, é ainda
possível que uma combinação da dsRNases-1 e -2 seja necessária para a forte atividade de
degradação de dsRNA no suco do intestino médio do gafanhoto do deserto. Em contraste, as
dsRNases-3 e -4 não parecem contribuir para a degradação do dsRNA no intestino médio.
Por outro lado, investigação realizada no nosso grupo revelou a existência de atividade
de ligação ao dsRNA na hemolinfa de S. gregaria e foi demonstrado que a apolipoforina-III
(ApoLpIII), uma componente da lipoforina, esta envolvida nesta ligação. A lipoforina é um
complexo proteico com um importante papel no transporte de lípidos e é constituída pela
apolipoforina-I e -II (provenientes do mesmo percursor, ApoLpI/II) e, em determinadas
situações fisiológicas, também pela apolipoforina–III. Desta forma, o segundo objetivo desta
tese foi investigar se as lipoforinas são responsáveis pela atividade de ligação ao dsRNA na
hemolinfa e, dependendo nos resultados obtidos, testar se estes complexos proteicos podem
proteger o dsRNA da degradação na hemolinfa (terceiro objetivo). Neste contexto,
começámos por realizar um perfil de transcrição da ApoLpI/II e da ApoLpIII, o que indicou
que estas são fortemente expressas no corpo gorduroso do gafanhoto do deserto. De seguida,
procedemos a um ensaio de ligação ao dsRNA com lipoforinas isoladas da hemolinfa de S.
gregaria. A observação de uma alteração da mobilidade em gel indicou que a lipoforina
possui, de facto, atividade de ligação ao dsRNA. Seguidamente, para testar se as lipoforinas
protegem o dsRNA da degradação, suco do intestino médio e serum de gafanhoto do deserto
foram utilizados, separadamente, em ensaios de proteção ex vivo. Os resultados não revelaram
proteção do dsRNA pelas lipoforinas em nenhum dos casos.
Em quarto lugar, pretendemos avaliar o papel das componentes da lipoforina no RNAi
(sistémico). Para tal, testámos a influência da ApoLpI/II e da ApoLpIII no RNAi (sistémico)
através de uma abordagem “RNAi em RNAi” in vivo. Esta metodologia consiste em silenciar
um gene-teste através da técnica de RNAi e, seguidamente, medir o efeito deste silenciamento
na potência da resposta RNAi para um gene-marcador. O “knockdown” transcricional da
ApoLpI/II e da ApoLpIII foi bem sucedido, mas a abordagem “RNAi em RNAi” não revelou
resultados conclusivos. Quando a ApoLpI/II se encontrava transcricionalmente sub-regulada,
a resposta RNAi (sistémica) foi parcialmente comprometida. Contudo, este resultado não foi
significativo e um controlo para verificar se o “knockdown” da ApoLpI/II não interfere

diretamente com a expressão do gene-marcador não foi realizado. No caso da ApoLpIII, não
foi possível observar nenhuma diferença na potência da resposta RNAi (sistémica). Ambas as
situações se referem a um “knockdown” e, desta forma, as proteínas podem estar presentes,
apesar de em níveis reduzidos. Neste contexto, e considerando que a injeção de quantidades
muito reduzidas de dsRNA é suficiente para causar um forte “knockdown” transcricional, os
níveis de ApoLpI/II e ApoLpIII presentes podem ser suficientes para se ligar a pequenas
quantidades de dsRNA e desempenhar o seu papel. Desta forma, não foi possível concluir
acerca do papel dos componentes da lipoforina no RNAi (sistémico) do gafanhoto do deserto.
De seguida, considerando um possível mecanismo de “uptake” de dsRNA, o quinto
objetivo desta tese foi avaliar o envolvimento da endocitose dependente de clatrina no RNAi
(sistémico), após injeção de dsRNA na cavidade abdominal de S. gregaria. Neste contexto,
utilizámos uma abordagem “RNAi em RNAi” para testar dois genes envolvidos em distintos
passos da endocitose dependente de clatrina, nomeadamente clath (do inglês clathrin heavy
chain) e vha16 (do inglês vacuolar H-ATPase 16). Estes dois genes estão envolvidos no
“uptake” de dsRNA exógeno na mosca da fruta e, de forma similar ao que foi observado em
Drosophila, o silenciamento individual destes genes comprometeu a resposta RNAi
(sistémica) no gafanhoto do deserto, o que indica que a endocitose dependente de clatrina está
envolvida no processo (sistémico) de RNAi, provavelmente no mecanismo de “uptake” de
dsRNA.
Finalmente, o sexto objetivo deste trabalho foi investigar se os recetores scavenger
(RS) desempenham um papel no RNAi (sistémico) do gafanhoto do deserto. Para tal,
avaliámos o efeito da inibição farmacológica dos RSs, com conhecidos inibidores gerais desta
família de recetores, no RNAi (sistémico) in vivo. De acordo com os nossos resultados,
quando os RSs estavam inibidos, a resposta RNAi (sistémica) foi comprometida, o que indica
que estes receptores estão envolvidos no mecanismo de RNAi (sistémico) do gafanhoto do
deserto, provavelmente no “uptake” endocítico de dsRNA, de uma forma semelhante ao que
acontece em Drosophila.
Em suma, o presente estudo reporta a identificação de uma nuclease que contribui para
a degradação do dsRNA no suco do intestino médio do gafanhoto do deserto, denominada
dsRNase-2, e a identificação da lipoforina como efetora da atividade de ligação ao dsRNA na
hemolinfa. Adicionalmente, o mecanismo endocítico dependente de clatrina e mediado por
RSs é proposto como um processo de importação celular de dsRNA na mesma espécie. Estes
resultados contribuem para uma melhor compreensão do mecanismo de RNAi sistémico no
gafanhoto do deserto.

Palavras chave: RNA de interferência, gafanhoto, dsRNA, dsRNase, lipoforina, endocitose,
recetores scavenger.

Abstract
RNA interference (RNAi) is a gene silencing mechanism triggered by double-stranded RNA
(dsRNA) structures. Among other biological activities, this pathway exerts an important
antiviral function. Notably, due to its strong silencing potency, specificity and potential
systemic (sys) effect, RNAi has proven to be very effective as a loss-of-function tool in
bioresearch and, in addition, this pathway has potential to contribute to the control of insect
crop pests. However, the interference response displays a strong inter-species variation and,
even within the same organism, it can vary between different tissues, developmental stages or
even according to the dsRNA delivery method. The desert locust, Schistocerca gregaria,
constitutes an interesting organism to study these matters. This species displays a very robust
and sensitive (sys)RNAi response upon injection of dsRNA in the hemocoel but does not
respond to dsRNA delivered by feeding. In addition, the desert locust constitutes an
agricultural pest of concern, making it a potential target for RNAi-based pest control.
Previous work from our lab demonstrated dsRNase activity in the midgut juice and in the
hemolymph, and four dsRNases were identified in the transcriptome of the desert locust.
Moreover, additional dsRNA-binding activity was found in the hemolymph and
Apolipophorin-III was demonstrated to be involved. The current study reports on the
identification of a nuclease, named dsRNase-2, that contributes to the degradation of dsRNA
in the midgut juice of the desert locust and on the identification of lipophorins as effectors of
the dsRNA-binding activity in the hemolymph of S. gregaria. Furthermore, we identified
Clathrin-dependent scavenger receptors-mediated endocytosis as a mechanism for cellular
uptake of dsRNA in vivo. Taken together, the present results contribute to a better
understanding of the sysRNAi mechanism in the desert locust.
Key words: RNA interference, locust, dsRNA, dsRNase, lipophorin, endocytosis, scavenger
receptors

Contents List of abbreviations ................................................................................................................... 1 1. Introduction ......................................................................................................................... 2
1.1. The desert locust, Schistocerca gregaria ..................................................................... 2 1.1.1. Taxonomy .............................................................................................................. 2 1.1.2. Life cycle and swarm formation ............................................................................ 2
1.2. RNA interference ......................................................................................................... 3 1.2.1. Small interfering RNA .......................................................................................... 4 1.2.2. Systemic RNA interference (sysRNAi) ................................................................ 4 1.2.3. Persistency of the dsRNA ..................................................................................... 6 1.2.4. DsRNA uptake mechanisms .................................................................................. 7
2. Objectives .......................................................................................................................... 11 3. Methodology ..................................................................................................................... 12
3.1. Rearing of the desert locust, S. gregaria .................................................................... 12 3.2. Production of dsRNA constructs ................................................................................ 12
3.2.1. PCR amplification of the gene specific fragment ............................................... 12 3.2.2. Electrophoretic analysis of the amplified products and gel extraction ............... 13 3.2.3. Cloning, transformation and purification of plasmid DNA ................................ 13 3.2.4. Sequencing .......................................................................................................... 13 3.2.5. DsRNA synthesis ................................................................................................ 13
3.3. Intra-abdominal injections .......................................................................................... 14 3.4. Micro-dissections and collection of hemolymph and midgut juice ........................... 14 3.5. RNA extractions and cDNA synthesis ....................................................................... 15 3.6. Quantitative reverse transcription PCR (qRT-PCR) .................................................. 15 3.7. Quantification of total protein amount in the midgut juice ........................................ 15 3.8. Purification of lipophorins ......................................................................................... 16 3.9. Phenol/chloroform extraction and ethanol precipitation ............................................ 16
4. Results ............................................................................................................................... 16 4.1. Identification of dsRNases in the midgut juice .......................................................... 16 4.2. Identification of lipophorin dsRNA-binding activity in the hemolymph ................... 18 4.3. The role of lipophorin in the sysRNAi-response ....................................................... 21 4.4. Clathrin-dependent endocytosis in (sys)RNAi ........................................................... 22 4.5. Scavenger Receptors in (sys)RNAi ............................................................................ 25
5. Discussion ......................................................................................................................... 26 5.1. Future perspectives ..................................................................................................... 30
6. References ......................................................................................................................... 31 6.1. Web-references ........................................................................................................... 35
i) Annex 1 .............................................................................................................................. 36 ii) Annex 2 ............................................................................................................................. 37 iii) Annex 3 ............................................................................................................................ 38

1
List of abbreviations Ago-2 Argonaute-2 ApoLp Apolipophorin B. mori; Bm Bombyx mori BCA Bicinchoninic Acid Assay BSA Bovine Serum Albumin C. elegans; Ce; Caenorhabditis elegans cDNA Complementary DNA clath clathrin heavy chain crq croquemort CS Chondroitin Sulphate D. melanogaster Drosophila melanogaster DNA Deoxyribonucleic acid dNTP Deoxynucleotide triphosphate DS Dextran Sulphate dsRNA double-stranded RNA ef1a elongation factor 1a emp epithelial membrane protein FAO Food and Agriculture Organization gapdh glyceraldehyde 3-phosphate dehydrogenase gfp green fluorescent protein HDLp High Density Lipophorin LB Luria-Bertani LDLp Low Density Lipophorin LPS Lipopolysaccharide M. sexta Manduca sexta miRNA micro RNA mRNA messenger RNA ninaD neither inactivation nor afterpotential D PCR Polymerase Chain Reaction piRNA piwi-interacting Poly(A) Polyadenosine Poly(I) Polyinosine PRR Pattern Recognition Receptors PTGS Post-Transcriptional Gene Silencing qPCR quantitative PCR qRT-PCR quantitative Reverse Transcription PCR RdRP RNA-dependent RNA polymerase RISC RNA-induced silencing complex RNA Ribonucleic Acid RNAi RNA interference rsd RNAi spreading defective S. gregaria; Sg Schistocerca gregaria S2 Schneider 2 SDS-PAGE Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis SEM Standard Error of the Mean sid systemic RNA interference-defective siRNA small interfering RNA SR Scavenger Receptor ssRNA single-stranded RNA sysRNAi systemic RNAi T. castaneum Tribolium castaneum tubu alpha-tubulin 1a ubi ubiquitin conjugating enzyme 10 UV Ultraviolet vha16 vacuolar H-ATPase 16

2
1. Introduction
1.1. The desert locust, Schistocerca gregaria
1.1.1. Taxonomy
Schistocerca gregaria, commonly named the desert locust, is one of the species of
grasshoppers that are known as locusts (Orthoptera: Acrididae). Locusts have the ability to
develop two extensively different phenotypes, the gregarious and the solitarious phases
(Figure 1), according to changes in local population density [1]. Table 1 displays the
taxonomic classification of S. gregaria.
Figure 1: A picture of (A) an adult gregarious desert locust and (B) a gregarious (left) and solitarious (right) fifth instar larvae.
Taxonomic ranks Scientific classification Superkingdom Eukaryota Kingdom Metazoa Phylum Arthropoda Superclass Hexapoda Class Insecta Subclass Neoptera Infraclass Orthopteroidea Order Orthoptera Suborder Caelifera Superfamily Acridoidea Family Acrididae Subfamily Cyrtacanthacridinae Genus Schistocerca Species Schistocerca gregaria
1.1.2. Life cycle and swarm formation
The life cycle of the desert locust can be divided into three stages: egg (and embryo),
hopper (nymph or larvae) and adult. Following oviposition by the female adult locust, the
Table 1: Taxonomic classification of Schistocerca gregaria [National Center for Biotechnology Information].

3
eggs hatch into wingless hoppers. S. gregaria passes through five larval stages, each time
growing in size. In the final moult, the wingless fifth instar hopper develops into a winged
adult. The complete life cycle of a desert locust can last up to 7 months [2].
In the gregarious phase, desert locusts can form huge groups and migrate over large
distances, either by forming marching bands of juveniles or flying swarms of winged adults.
An adult desert locust can consume roughly its own weight in fresh food per day and a single
swarm can contain billions of insects. A swarm can travel hundreds of kilometers each day
and, occasionally, even cross oceans, which makes these animals a feared agricultural pest
[1].
1.2. RNA interference
RNA interference (RNAi) is a post-transcriptional gene silencing mechanism that is
triggered by double-stranded (ds)RNA [3]. The first evidences for the existence of such a
pathway were described in plants, where the mechanism is known as post-transcriptional gene
silencing (PTGS) [4, 5], and later on in fungi, where it is referred to as quelling [6]. However,
it was not named as RNAi until 1998, when Fire et al. described the process in which
application of dsRNA triggers the silencing of the homologous endogenous transcripts in
Caenorhabditis elegans [7].
The RNAi pathway can be described in three steps. In the first step, the trigger dsRNA
is processed into small RNA duplexes (21-28bp) by RNase III enzymes (either Dicer alone or
Drosha and Dicer). Second, in a multistep process, these duplexes are unwound and one
strand is loaded into a protein complex known as RNA-induced silencing complex (RISC). In
the third step, this complex finds the potential messenger RNA (mRNA) target by Watson-
Crick base pairing. Then, the loaded single-stranded RNA (ssRNA), called the guide strand,
directs an endonuclease (Argonaute) that is present in the RISC to degrade mRNAs that
contain sequences complementary to the loaded ssRNA. In this way, the guide strand
determines the sequence specificity of the RNAi response (Figure 2) [4, 7-10].
Since it is possible to artificially introduce dsRNA structures in cells and organisms,
RNAi has become a widely used research tool to knock down and analyze the function of
genes. In addition, due to its high specificity, RNAi may contribute to novel strategies for
selectively controlling agricultural pests, including a number of insect species. Thus, this
technique has proven to be very useful and promising in several research fields, including
(reverse) genetics, genomics and biotechnology [11, 12].

4
Figure 2: RNA interference. The long dsRNA is processed into small RNA duplexes by Dicer. These duplexes are unwound and the guide strand is loaded into RISC. Finally, the guide strand directs Argonaute, that is present in the RISC, to degrade target mRNAs (complementary to the loaded ssRNA). [Adapted from The RNAi web].
1.2.1. Small interfering RNA
Many classes of small RNAs have recently emerged, but based on their origin,
structure, associated effector proteins and biological roles, it is possible to distinguish three
main categories: short interfering RNAs (siRNAs), micro RNAs (miRNAs) and piwi-
interacting RNAs (piRNAs) [13]. For the general understanding of this work, this
introduction will focus on siRNAs.
The siRNAs are short dsRNA molecules (~21-25 nucleotides) that have their origin in
long, linear, perfectly base-paired dsRNA structures [10, 14]. These duplexes are processed
by Dicer into siRNAs that direct the silencing [4, 15]. Under natural conditions, the siRNA
based silencing mechanism can be triggered as a defense mechanism against dsRNA
structures that are either produced inside cells during the replication cycle of viruses, or
endogenously generated from repetitive elements and transposons in the cellular genome [13,
16]. For instance, in Drosophila melanogaster, a system that represses transposable elements
via siRNAs in somatic tissues was identified and the observation that Argonaute 2 (Ago-2)-
defective flies were hypersensitive to viral infection allowed to conclude that this protein
mediates antiviral immunity. In addition, flies defective in the dsRNA endocytic uptake
pathway are hypersensitive to viral infection, which suggested that the effective antiviral
immunity requires the systemic spread of the RNAi-signal [17, 18]. The systemic RNAi is
discussed below.
1.2.2. Systemic RNA interference (sysRNAi)
Cell-autonomous RNAi encompasses the interference process within individual cells

5
and is defined as the silencing process limited to the cell in which the dsRNA is introduced or
expressed [12]. However, in some organisms, when the dsRNA is artificially delivered by
feeding, soaking or injection, it is able to enter the individual cells and trigger a RNAi-
response. Moreover, in some cases, the RNAi-signal is spread throughout the entire organism
(systemic RNAi) [11]. C. elegans was the first animal in which RNAi was proven to work
systemically [7] and still the best-studied organism for the mechanisms of sysRNAi. In this
worm, RNAi can be initiated either by feeding, soaking or injection of dsRNA, or by
transgenesis. SysRNAi is a common phenomenon in many organisms. Several insects, such as
many members of Orthoptera, Dictyoptera and Coleoptera possess a robust sysRNAi
response. S. gregaria is a good example, presenting an highly effective sysRNAi-response
[19-21]. Nevertheless, in other cases, the introduction of dsRNA does not result in a
sysRNAi-effect. D. melanogaster, the best-known example of such fact, displays a very
reduced sensitivity to sysRNAi. In the larvae of the fruit fly almost all tissues, with the
exception of hemocytes, lack the ability to take up dsRNA from the surrounding environment
and, consequently, this model insect is mainly refractory to a systemic interference effect [19,
20].
The systemic phenomenon of RNAi is also well characterized in plants. In this case,
siRNAs spread to neighboring cells through the plasmodesmata and over greater distances
through the phloem [22]. In both C. elegans and plants, sysRNAi is associated with a RNA-
dependent RNA polymerase (RdRP) activity, which can convert small populations of dsRNA
fragments into an abundant pool of siRNAs. Up to date, in arthropods, RdRP encoding genes
are only described in the tick lineage and the presence of RdRP activity in insects is not clear
[12, 23-25].
Several factors can affect the efficiency of RNAi. On one hand, the cell-autonomous
RNAi machinery can present different levels of sensitivity due to different expression levels
of core components of the RNAi machinery [21, 26]. On the other hand, the systemic
character of the RNAi response can be less responsive, for instance, due to an inefficient
dsRNA uptake [27, 28]. It is also possible that the presence of non-specific nucleases leads to
degradation of the dsRNA prior to its uptake by the cells [29, 30]. In this context, the dsRNA
delivery method can also compromise the efficiency of RNAi. In C. elegans, it is known that
feeding of dsRNA is less efficient when compared to direct injection [31]. This difference
also exists in insects and, in some species, injection of dsRNA can induce RNAi, while
ingestion does not, as for instance in the plant pest Lygus lineolaris [32], in the cotton
leafworm Spodoptera litura [33] and in the desert locust [unpublished results]. These

6
situations are of particular interest and gain importance as they compromise the possibility of
pest control through feeding of dsRNA expressed either by plants or by bacteria [12, 20, 34,
35]. In addition, feeding of dsRNA is much less labor intensive when compared to the other
delivery methods [31].
1.2.3. Persistency of the dsRNA
In many insects, gene silencing can be obtained following injection or feeding of
dsRNA [12]. In order to enter the cells, the dsRNA must first persist in the hemocoel or gut
lumen, respectively [29, 30, 32], and subsequently be recognized and taken up by the cells.
1.2.3.1. Activity of dsRNA degradation
Some reports emphasize the role of dsRNase activity in the (sys)RNAi mechanism.
Firstly, in Bombyx mori, an enzyme with dsRNase activity (Bm-dsRNase) was isolated from
the digestive juice. The precursor of this enzyme (51 kDa) consists of three domains: a signal
peptide, an N-terminal propeptide and a mature Bm-dsRNase. The mature Bm-dsRNase (43
kDa) is generated by post-translational processing and is classified as a DNA/RNA non-
specific nuclease. In fact, it can degrade dsRNA, but also DNA, although in a lower degree.
Expression of this protein has been detected in the middle and posterior midgut of larvae,
whereas no expression was detected in anterior midgut, silk gland, fat body and Malpighian
tubule [36]. Secondly, in the plant bug, Lygus lineolaris, it was proven that saliva was able to
degrade dsRNA and, consequently, it was suggested that this degradation could account as
one of the reasons for the failure of gene silencing upon feeding of dsRNA in this species
[32]. Thirdly, by comparing the dsRNase activity in the hemolymph of Manduca sexta
(refractory to RNAi response) and in the one of Blattella germanica (robust RNAi response),
it was proposed that, in part, the deficient RNAi response in M. sexta is due to the rapid
degradation of the dsRNA in the hemolymph [30]. Finally, dsRNA-degradation activity was
also found in the midgut juice and in the hemolymph of S. gregaria. Based on the DNA/RNA
non-specific nuclease isolated from the midgut juice of B. mori, our group has confirmed the
identity of four dsRNase sequences found in the transcriptome of the desert locust, termed
dsRNases (-1, -2, -3 and -4) [unpublished results].

7
1.2.3.2. Activity of dsRNA-binding
Activity of dsRNA-binding has been reported in the hemolymph of B. mori. Lipophorin,
that is an important lipoprotein synthetized in the fat body and present in the insects’
hemolymph, was identified as the molecular effector of this activity [37, 38]. The lipophorin
complex, that is particularly important in the lipid transport mechanism, contains three
different apoproteins: two non-exchangeable structural components processed from the same
precursor - apolipophorin-I (ApoLpI) and apolipophorin-II (ApoLpII) – and one component
that can associate to the lipophorin or be present as a free form in the hemolymph -
apolipophorin-III (ApoLpIII) [39]. Under most physiological conditions, lipophorin exists in
the hemolymph as an high-density lipophorin (HDLp), which contains one molecule of
ApoLpI and one of ApoLpII. Under certain conditions, for instance during lipid transport,
several ApoLpIII can bind to the HDLp, generating the low-density lipophorin (LDLp) [38,
39]. Interestingly, lipophorin has lately been suggested to be involved in the innate immune
response in insects [40-45].
Research performed in our lab identified dsRNA binding activity in the hemolymph of
S. gregaria, but not in the midgut juice. Through an electrophoretic mobility shift assay
followed by protein purification and determination of the amino acid sequence by Edman
degradation, the protein was revealed to be a desert locust homolog of ApoLpIII. In addition,
transcript sequences of the precursor of ApoLpI and ApoLpII (ApoLpI/II) and of ApoLpIII
were found in the transcriptome of S. gregaria [unpublished results].
1.2.4. DsRNA uptake mechanisms
Two mechanisms of cellular uptake of dsRNA have been described in animals: a
transmembrane channel-mediated uptake mechanism and an endocytosis-mediated uptake
mechanism [12].
1.2.4.1. Transmembrane channel-mediated uptake mechanism
The best-studied mechanism of cellular uptake of dsRNA in animals is the
transmembrane channel-mediated uptake mechanism by SID-1 in C. elegans. The systemic
RNA interference-deficient 1 (sid-1) mutant was identified in a screen conducted by Winston
et al. (2002) and SID-1 is a transmembrane protein necessary for the passive uptake of
dsRNA silencing signals into C. elegans cells [46, 47]. It has been demonstrated that the
expression of SID-1 in Drosophila cells was sufficient to enhance the uptake of dsRNA from

8
the medium and that this factor acted as a passive gated-channel selective for dsRNA [48, 49].
Furthermore, it was reported that SID-1 is required for the import but not for the export of
RNAi triggers [50].
Several genes related to sid-1 were identified in insects (Table 2) but their clear
involvement in the dsRNA uptake remains uncertain [12]. In Apis mellifera, the involvement
of sid-1 in the RNAi process was suggested due to the observed up-regulation of Am-sid-1
right before the knockdown of a target gene [51]. On the other hand, silencing of the three
Tribolium castaneum sid-1 genes, either individually or all together, did not influence the
RNAi response [11]. Furthermore, it was recently suggested that SID-1 is not required for the
sysRNAi in Locusta migratoria [52].
Curiously, sequence analysis has revealed that, in some insects, SID-1 shows more
similarity to C. elegans TAG-130 than to Ce-SID-1 [11, 12]. Ce-TAG-130, nowadays also
denominated as Cholesterol Uptake Protein-1 (ChUP-1), is a membrane-associated protein
involved in the uptake of dietary cholesterol [53]. In addition, it has been observed that
ChUP-1 does not affect the sysRNAi in C. elegans, which may indicate that insect sid-1-like
genes may have a different function rather than dsRNA uptake [53]. Nevertheless, in other
arthropods, SID-1 seems to be involved in the RNAi mechanism, as in the case of the Pacific
white leg shrimp, Litopenaeus vannamei [54]. Furthermore, fish and mammalian sid-1
homologs also mediate dsRNA uptake [55, 56]. In this context, further research is required in
order to elucidate the role of the insect sid-1 related sequences in the RNAi response.
Table 2: Overview on the presence of sid-1 related genes in different insect orders and species. # sid-1 rel. genes: number of sid-1 related genes. Exp.: expression. Ref.: reference. +: expressed. /: not tested. Np: not published. Adapted from [12].
Species # sid-1 rel. genes
Exp. Ref. Species # sid-1 rel. genes
Exp. Ref.
Coleoptera Hymenoptera Tribolium castaneum 3 + [11] Apis mellifera 1 + [51] Diptera Nasonia vitripennis 1 / [11] Aedes aegypti 0 / [11] Lepidoptera Anopheles gambiae 0 / [11] Bombyx mori 3 / [11] Culex quinquefasciatus 0 / [12] Spodoptera exigua 1 / [12] Drosophila melanogaster 0 / [47] Orthoptera Hemiptera Schistocerca americana 1 + [57] Acyrthosiphon pisum 1 / [12] Locusta migratoria 1 + [52] Aphys gossypii 1 / [58] Schistocerca gregaria 1 + Np Rhodnius prolixus 0 / [12] Phtiraptera Rhopalosiphum padi 1 / [12] Pediculus humanus corporis 1 / [12] Sitobion avenae 1 / [58]

9
1.2.4.2. Endocytosis-mediated uptake mechanism
The best-known model insect D. melanogaster, which has no robust sysRNAi, lacks
sid-1 related genes. However, S2 cells are still able to develop an RNAi response when
soaked in medium with dsRNA, which suggested the existence of an alternative dsRNA
uptake mechanism. Two independent functional screens, Saleh et. al (2006) [59] and Ulvila et
al. (2006) [60], verified that several components of the Clathrin-dependent endocytosis were
needed for effective dsRNA-uptake and processing. Furthermore, it was demonstrated that
blocking the Clathrin-dependent endocytic pathway in S2 cells impaired the RNAi response
and that the dsRNA was associated with vesicles upon entrance in the cell. These results
indicated that dsRNA fragments were internalized in S2 cells by Clathrin-dependent
endocytosis [59, 60].
The scavenger receptors (SRs), a major class of pattern recognition receptors (PRRs),
were demonstrated to mediate the Clathrin-dependent endocytosis of dsRNA [59, 60]. They
constitute a group of structurally unrelated cell surface proteins that are able to recognize and
mediate the endocytosis of negatively charged macromolecules and (modified) lipoproteins.
More recently, SRs have been shown to perform an important role in the innate immunity by
functioning as PRRs. They participate in the recognition and removal of pathogen agents, in
particular of bacteria [61]. SR-based bacterial recognition seems to be conserved in insects
and humans and may represent one of the most primitive forms of microbial recognition [62].
In mammals, this family of receptors is categorized into, at least, six classes according to their
structural characteristics (classes A-F) [63]. SRs, mainly belonging to classes B and C, have
been characterized in Drosophila (Table 3).
Curiously, Drosophila SR-CI functionally resembles the mammalian class A SRs. It
plays a relevant role in the phagocytosis of Gram-positive and -negative bacteria in S2 cells
[62]. In addition, another Drosophila SR, named Eater, has been described to mediate
phagocytosis of both Gram-positive and -negative bacteria in Drosophila [67, 68].
Class Symbol Name Reference C SR-CI Scavenger receptor class C, type I [62] C SR-CII Scavenger receptor class C, type II [62] C SR-CIII Scavenger receptor class C, type III [62] C SR-IV Scavenger receptor class C, type IV [62] B crq croquemort [64] B emp epithelial membrane protein [65] B ninaD neither inactivation nor afterpotential D [66] / eater eater [67]
Table 3: Overview of the SRs characterized in Drosophila melanogaster. /: not classified yet. Adapted from [59].

10
Interestingly, the eater homologs in Sarcophaga peregrina and C. elegans are implicated in
the removal of apoptotic cells [69, 70]. However, it is noteworthy that it has recently been
suggested that the role of the mammalian SR A in bacterial phagocytosis is functionally
distinct from the SR A mediated clearance of the lipopolysaccharide (LPS), which is
consistent with a the idea that the bacterial phagocytic ligands and the LPS (endocytic ligand)
may bind to distinct sites of the SR A [71-73]. Thus, in the context of receptor-mediated
endocytosis of dsRNA and through the use of general SR inhibitors, Saleh et al. were able to
demonstrate that a combination of SRs participates in dsRNA uptake in S2 cells [59].
Moreover, using a RNAi targeting approach, Ulvila et al. investigated the role of specific SRs
in the process. The silencing of SR-CI and of eater led to a significant decrease in the
internalization of dsRNA fragments, while silencing the class B scavenger receptors crq, emp,
and ninaD had no detectable effect on the uptake of dsRNA. In addition, the stable
transfection of mammalian cells with SR-CI was enough to markedly increase the dsRNA
internalization in these cells [60]. Taken together, these results made clear that, in Drosophila
S2 cells, dsRNA is internalized by SR-mediated endocytosis. Research work performed in our
lab identified five class B SRs and one class C SR in the transcriptome of the desert locust
[unpublished results].
Notably, it has been suggested that, in C. elegans, the dsRNA is imported from the gut
lumen via endocytosis, in a SID-2 dependent manner [74]. SID-2 is a transmembrane protein
expressed in all gut cells and is mainly localized in the apical/luminal membrane. The sid-2
mutants are specifically defective in developing a sysRNAi effect upon ingestion of dsRNA.
The expression of Ce-SID-2 in S2-cells significantly enhanced the uptake of dsRNA but,
nevertheless, sid-1 mutants fed with dsRNA failed to show silencing in gut cells and in all
other cells, which means that both SID-1 and -2 are required for the delivery of dsRNA from
the lumen into gut cells in order to cause a (sys)RNAi after ingestion of dsRNA. Thus, it is
possible that, upon being imported to the gut cells via the endocytic uptake mediated by SID-
2, the dsRNA is released from internalized vesicles in a step mediated by the dsRNA channel
SID-1 [46, 74, 75]. Recently, another C. elegans sysRNAi defective mutant was described,
namely sid-3. It was proven that cells lacking SID-3 cells could properly perform RNAi
silencing but poorly import dsRNA. In addition, upon overexpression of SID-3, cells were
able to import dsRNA more efficiently and this import required an intact SID-3 kinase
domain. Together with the fact that SID-3 is widely expressed in most tissues, the results led
the authors to suggest that SID-3 functions to enhance the endocytosis-mediated dsRNA
import in the recipient cell, playing a regulatory role [76]. Another interesting protein

11
identified in the (sys)RNAi process is the endosome-associated protein SID-5. It was
described as a regulator of the cellular export of RNAi silencing signals and required for
efficient sysRNAi in C. elegans, in response to both ingested and expressed dsRNA [77]. In
addition, the identification of C. elegans RNAi Spreading Defective 3 (rsd-3) mutant and
characterization of RSD-3, also suggested that endocytosis is involved in the spreading of
dsRNA in this worm species. RSD-3 is highly expressed in endocytic cells and was suggested
to possess a regulatory role in the vesicle-trafficking pathway specific for sysRNAi [78, 79].
2. Objectives
The sysRNAi response in insects is highly species-dependent. However, the underlying
mechanisms have been mostly investigated in D. melanogaster, which is well known to
possess poor sensitivity towards sysRNAi. On the other hand, the desert locust, S. gregaria,
demonstrates a highly robust and sensitive sysRNAi response [21]. Since this species is a
voracious pest insect and the RNAi response has the potential to contribute to strategies for
the selective control of agricultural pests, the desert locust constitutes an amenable insect to
investigate the mechanisms of sysRNAi. In S. gregaria, injection of dsRNA is an effective
method to induce sysRNAi but, in contrast, oral delivery of dsRNA does not result in gene
silencing [unpublished results]. Previous research in our lab revealed dsRNase activity in the
midgut juice of the desert locust. Therefore, the first objective of this thesis was to identify
the nucleases responsible for the dsRNA degradation in the midgut juice. Moreover, ex-vivo
experiments from our group identified additional dsRNA-binding activity in the hemolymph
of S. gregaria and ApoLpIII was demonstrated to be involved [unpublished results].
Therefore, the second objective of this thesis was to investigate whether lipophorins are the
effectors of this activity and, depending on the obtained results, to test if lipophorins can
protect dsRNA from degradation in the hemolymph (third objective). Next, in the fourth
place, we intended to assess the role of lipophorin components in the (sys)RNAi. Then,
regarding the dsRNA uptake mechanism, the fifth objective was to evaluate the involvement
of Clathrin-dependent endocytosis in the (sys)RNAi following the injection of dsRNA into
the abdominal body cavity of S. gregaria and, finally, to investigate whether SRs are playing
a role in this pathway.

12
3. Methodology
3.1. Rearing of the desert locust, S. gregaria
Gregarious S. gregaria were reared under crowded conditions with controlled temperature
(32 ± 1 °C), light (13h photoperiod) and relative humidity (40-60%). They were fed daily
with fresh cabbage and dried oat flakes. Fifth larval stage or adult locusts were
developmentally synchronized by transferring them to a different cage directly after the fourth
or final moult, respectively.
3.2. Production of dsRNA constructs
Gene specific dsRNA constructs were synthesized. Firstly, for each construct (except for
gfp), a gene specific fragment flanked by two T7 sequences was amplified by polymerase
chain reaction (PCR). Secondly, the PCR product was analyzed by electrophoresis and the
expected band was excised for sequencing. Thirdly, after the sequences were confirmed,
sense and antisense RNA strands were synthetized in the same reaction by a T7 enzyme. In
the case of gfp, a single T7 promoter-containing vector, cloned in the sense and antisense
direction, was directly used as template. Therefore, the RNA sense and antisense strands were
synthetized in different reactions and posteriorly annealed. The DNA and ssRNA were
removed by nuclease digestion and the proteins and mono/oligonucleotides by solid phase
adsorption. These steps are explained in more detail below. Transcript sequence information
of clathrin heavy chain (clath), vacuolar H-ATPase 16 (vha16), glyceraldehyde 3-phosphate
dehydrogenese (gapdh), alpha-tubulin 1a (tubu), dsRNase-1, -2, -3, -4, ApoLpI/II and
ApoLpIII was available in our laboratory, as well as the green fluorescent protein (gfp)
sequence-containing vector [[21] and unpublished results].
3.2.1. PCR amplification of the gene specific fragment
A DNA template flanked by two T7 promotor sequences was produced for each
transcript: dsRNase-1 (576 bp), -2 (346 bp), -3 (496 bp), -4 (646 bp), ApoLpI/II (631 bp),
ApoLpIII (277 bp), vha16 (453 bp), clath (561 bp), gapdh (447 bp) and tubu (545 bp). The
primers used in this step (Sigma–Aldrich co.) are displayed in table 4 (Annex 1). REDTaq®
ReadyMixTM (Sigma–Aldrich co.) was used as a source of DNA Taq polymerase, dNTPs and
PCR buffer. S. gregaria midgut tissue cDNA was used as template for the amplification of
clath, vha16, gapdh, tubu, dsRNase-1, -2, -3 and -4. Total fourth larval stage S. gregaria

13
cDNA was used as template for the amplification of ApoLpI/II and ApoLpIII. The PCR
reaction was prepared according to the REDTaq® ReadyMixTM protocol.
3.2.2. Electrophoretic analysis of the amplified products and gel extraction
The amplified products were analyzed by 1% agarose gel electrophoresis followed by
fluorescent visualization with UV-light. The agarose gel was prepared with agarose (Sigma–
Aldrich co.), 10x Tris-Acetate EDTA (TAE) buffer and GelRed™ (Biotium). For every case,
1Kb ladder (Fermentas) was utilized. The REDTaq® ReadyMixTM was also the source for
loading dye. The bands with the expected length were excised and the DNA template was
extracted for future cloning, transformation and sequencing. GenElute™ Gel extraction Kit
(Sigma–Aldrich co.) was used according to the manufacturer’s instructions.
3.2.3. Cloning, transformation and purification of plasmid DNA
Cloning of the previously extracted fragments was performed using the TOPO TA
Cloning® Kit for Sequencing (Life Technologies Co.) and TOP10 Chemically Competent E.
coli cells were used for transformation, according to the manufacturer’s instructions. The
transformation mix was spread on Luria-Bertani (LB) agar plates containing 50 µg/mL
ampicillin and the plates were incubated overnight at 37 ºC. Six colonies were picked and
incubated individually in LB broth medium (50 µg/mL ampicillin), overnight, at 37 ºC in a
shaking incubator. The plasmid isolation was performed with GenElute TM HP plasmid
Miniprep kit (Sigma–Aldrich co.), according to the manufacturer’s instructions.
3.2.4. Sequencing
The sequences of the inserted DNA fragments were determined using ABI PRISM
BigDye Terminator Ready Reaction Cycle Sequencing Kit (Applied Biosystems), according
to the protocol provided by the manufacturer. The obtained sequences were compared with
the transcript sequence information previously identified by our research group [unpublished
results].
3.2.5. DsRNA synthesis
DsRNA constructs for dsRNase-1, -2, -3, -4, ApoLpI/II, ApoLpIII, vha16, clath, gapdh
and tubu were synthesised using the MEGAscript® RNAi kit (Ambion). PCR products for
dsRNase-1, -2, -3, -4, ApoLpI/II, ApoLpIII, vha16, clath, gapdh and tubu were directly used
for dsRNA production according to manufacturers protocol. Synthesis of gfp dsRNA (589 bp)
was performed using a TOPO 4.1 sequencing vector (Life Technologies Co.) containing a gfp

14
sequence as DNA template. Since only one T7 promoter site is present in this vector, the
fragment was cloned both in the sense and antisense direction. RNA was then synthesised by
the T7 Enzyme Mix of the MEGAscript RNAi kit (Ambion). Both gfp RNA strands were first
synthesised independently before being mixed to anneal, while transcripts made from a single
template with opposing T7 promoters (all remaining cases) were hybridized during the
transcription reaction. After the production of dsRNA, the remaining DNA and ssRNA were
removed by nuclease treatment, and proteins and mono/oligonucleotides were removed by
solid phase adsorption purification, according to the manufacturers' specifications (Ambion).
The dsRNA-concentration was determined by means of a Nanodrop spectrophotometer
(Thermo Fisher Scientific, Inc.), and the integrity of the dsRNA was assessed by gel
electrophoresis, prepared as described in 3.2.2. The dsRNA was stored at −20 °C until further
use.
3.3. Intra-abdominal injections
S. gregaria Ringer solution (1L: 150 mM NaCl; 1.7 mM CaCl2; 10 mM KCl; 4.3 mM
MgCl2; 4 mM NaHCO3; 90 mM sucrose; 5 mM trehalose; pH 7.2 ) was used to dilute the
dsRNA to the desired concentration. Fifth larval stage locusts were each intra-abdominally
injected with a volume of 6 µl of dsRNA-solution and adult locusts with a volume of 10 µl. In
the cases of gapdh, tubu and gfp, each locust was injected with 150 ng of dsRNA. In the cases
of dsRNase-1, -2, -3, -4, ApoLpI/II, ApoLpIII, clath and vha16, an amount of 300 ng per
locust was used.
Adult locusts were each injected with a solution of 0.1 mg poly(I) (Sigma-Aldrich co.),
poly(A) (Sigma-Aldrich co.), dextran sulphate (Sigma-Aldrich co.) or chondroitin sulphate
(Sigma-Aldrich co.), diluted in 10 µl of Sg-Ringer solution.
3.4. Micro-dissections and collection of hemolymph and midgut juice
The micro-dissections were performed in Sg-Ringer solution under a binocular
microscope. Equal volumes of hemolymph were collected per locust, with the use of capillary
tubes. S. gregaria serum was prepared by pelleting the hemocytes from the hemolymph by
centrifugation. Upon midgut dissection, similar volumes of midgut juice were collected per
locust. The midgut juice samples of three individual locusts were pooled and subsequently
diluted in 150 µl Sg-Ringer solution. All samples were immediately transferred to liquid
nitrogen to prevent RNA and protein degradation, and stored at -80 ºC until further usage.

15
3.5. RNA extractions and cDNA synthesis
RNA extractions were performed with the Lipid tissue extraction kit (Qiagen), as
described in the corresponding protocol. A Nanodrop spectrophotometer was used to assess
quality and concentration of the extracted RNA. In order to produce cDNA, equal amounts of
RNA were used. The cDNA synthesis was performed with the PrimeScript™ First strand
cDNA Synthesis Kit (TaKaRa), according to manufacturers’ specifications. The cDNA-
solution was diluted 15 times in MilliQ water and analyzed immediately or stored at -20ºC
until further usage.
3.6. Quantitative reverse transcription PCR (qRT-PCR)
Quantitative real time RT-PCR (qRT-PCR) was used to measure the relative quantities of
transcripts. For this, qRT-PCR primers were design with primer express software (Applied
Biosystems) (Table 5, Annex 1). In order to validate the primers, a standard curve based on a
serial dilution of cDNA was made to determine the primer annealing efficiency, the presence
of primer dimers and the production of a single PCR product. For all transcripts, only a single
melting peak was found. Based on the study of Van Hiel et al. [80] and with the use of the
geNorm program [81], our lab established the two most stably expressed genes for these
experiments, namely ubiquitin conjugating enzyme 10 (ubi) and elongation factor 1a (ef1a).
In this context, in order to correct for sample-to-sample variations, the relative expression
levels were normalized against these two reference genes. In addition, the data were further
normalized against a calibrator cDNA sample to account for possible variations in the PCR-
efficiency in different PCR runs. Each qRT-PCR reaction contained 10 µl SYBR green
solution (Life Technology Co.), 0.75 µl of each 10 mM forward and reverse primers (Sigma
Aldrich), 3.5 µl milliQ water and 5 µl cDNA. In every experiment, a no-template control was
included to check for possible contaminations and all reactions were performed in duplicate.
The qRT-PCR reaction was performed and analyzed in a 96 well plate and by using the
StepOne System (ABI Prism, Applied Biosystems). Since the efficiency of the different
primers was the same, the relative transcript quantity was calculated according to the delta-
delta Ct method.
3.7. Quantification of total protein amount in the midgut juice
The total protein concentration in the midgut juice samples was determined according to
the bicinchoninic acid (BCA) assay. For this purpose, two reagents named A and B were
mixed in a 1/49 ratio. Reagent A was prepared by adding sodium bicinchoninate (0.1 g),

16
Na2CO3.H2O (2 g), sodium tartrate (dihydrate) (0.16 g), NaOH (0.4 g) and NaHCO3 (0.95 g)
in 100 mL of distilled H2O and adjusting the pH to 11.25. Reagent B consisted of 0.4%
CuSO4.5H2O. Bovine serum albumin (BSA) standard solutions were used as reference and
the measurements were performed by means of a Nanodrop spectrophotometer (Thermo
Fisher Scientific, Inc.). Afterwards, Sg-Ringer solution was used to dilute the samples to the
same concentration.
3.8. Purification of lipophorins
Lipophorins were purified from the hemolymph of desert locusts by ultracentrifugation in
a potassium bromide density gradient, according to the protocol provided in the
supplementary data (Annex 2). Next, the K+ and Br- ions were removed from the solution
through dialysis in Sg-Ringer solution. The size of the purified proteins was confirmed by
sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE), prepared with
NuPAGE Bis-Tris Mini Gels (Life technologies Co.). The staining was performed with
SimplyBlue™ Safe stain (Life technologies Co.). The used ladder was SeeBlue Plus2 Pre-
Stained Standard (Life technologies Co.).
3.9. Phenol/chloroform extraction and ethanol precipitation
The phenol/chloroform extraction and the ethanol precipitation were performed according
to the protocol provided in the supplementary data (Annex 2).
4. Results
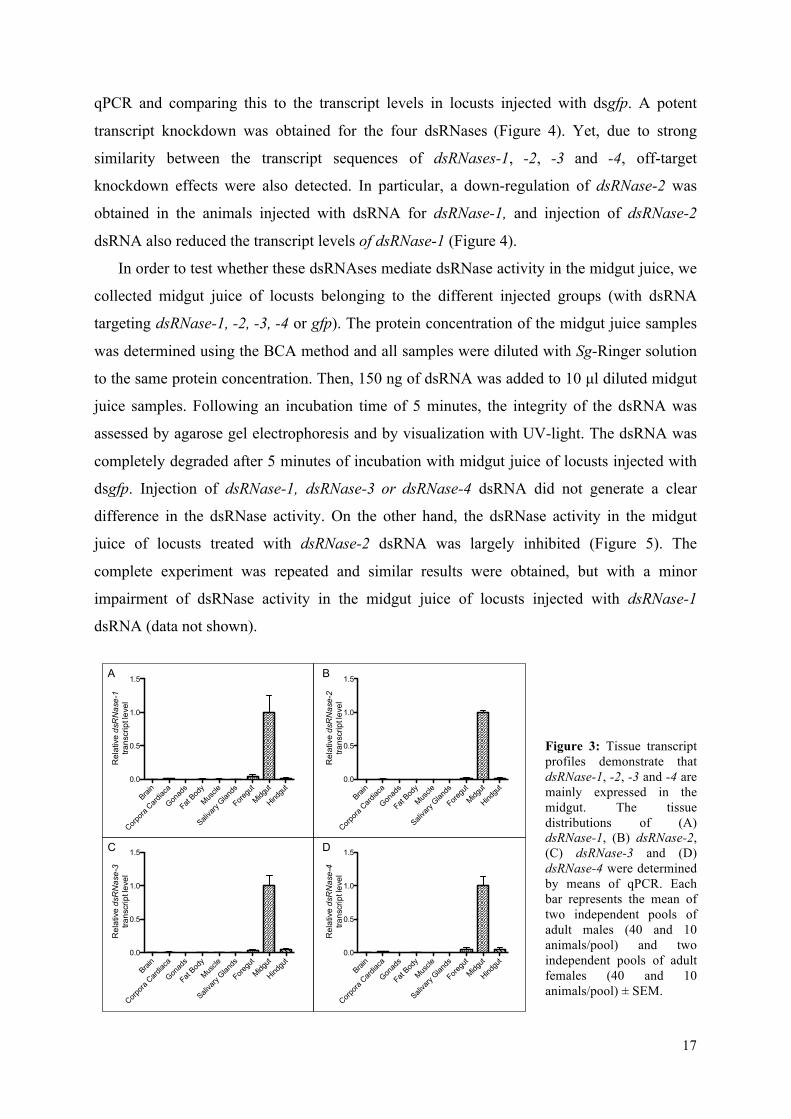
4.1. Identification of dsRNases in the midgut juice
In the available S. gregaria transcriptome database, four transcripts with high sequence
similarity to the DNA/RNA non-specific nuclease of B. mori [36] were identified and termed
dsRNase-1, -2, -3 and -4 [unpublished results]. By means of qPCR, the tissue transcript
profile of these dsRNases was determined for seven-days old adult desert locusts. Our data
demonstrate that they are mainly expressed by the midgut tissue of the desert locust (Figure
3). In order to investigate the role of these nucleases in the degradation of dsRNA in the
midgut juice, transcript-specific dsRNA constructs were generated for the four dsRNases. The
dsRNA fragments were injected intra-abdominally to induce gene silencing. The knockdown
was assessed six days later by measuring the transcript levels in the midgut tissue by means of
17
qPCR and comparing this to the transcript levels in locusts injected with dsgfp. A potent
transcript knockdown was obtained for the four dsRNases (Figure 4). Yet, due to strong
similarity between the transcript sequences of dsRNases-1, -2, -3 and -4, off-target
knockdown effects were also detected. In particular, a down-regulation of dsRNase-2 was
obtained in the animals injected with dsRNA for dsRNase-1, and injection of dsRNase-2
dsRNA also reduced the transcript levels of dsRNase-1 (Figure 4).
In order to test whether these dsRNAses mediate dsRNase activity in the midgut juice, we
collected midgut juice of locusts belonging to the different injected groups (with dsRNA
targeting dsRNase-1, -2, -3, -4 or gfp). The protein concentration of the midgut juice samples
was determined using the BCA method and all samples were diluted with Sg-Ringer solution
to the same protein concentration. Then, 150 ng of dsRNA was added to 10 µl diluted midgut
juice samples. Following an incubation time of 5 minutes, the integrity of the dsRNA was
assessed by agarose gel electrophoresis and by visualization with UV-light. The dsRNA was
completely degraded after 5 minutes of incubation with midgut juice of locusts injected with
dsgfp. Injection of dsRNase-1, dsRNase-3 or dsRNase-4 dsRNA did not generate a clear
difference in the dsRNase activity. On the other hand, the dsRNase activity in the midgut
juice of locusts treated with dsRNase-2 dsRNA was largely inhibited (Figure 5). The
complete experiment was repeated and similar results were obtained, but with a minor
impairment of dsRNase activity in the midgut juice of locusts injected with dsRNase-1
dsRNA (data not shown).
Figure 3: Tissue transcript profiles demonstrate that dsRNase-1, -2, -3 and -4 are mainly expressed in the midgut. The tissue distributions of (A) dsRNase-1, (B) dsRNase-2, (C) dsRNase-3 and (D) dsRNase-4 were determined by means of qPCR. Each bar represents the mean of two independent pools of adult males (40 and 10 animals/pool) and two independent pools of adult females (40 and 10 animals/pool) ± SEM.
Brain
Corpora
Card
iaca
Gonad
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-1
trans
crip
t lev
el
Brain
Corpora
Card
iaca
Gonad
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-3
trans
crip
t lev
el
Brain
Corpora
Card
iaca
Gonad
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-2
trans
crip
t lev
el
Brain
Corpora
Card
iaca
Gonad
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-4
trans
crip
t lev
el
A B
C D

18
Figure 5: The dsRNase-2 contributes strongly to dsRNA degradation in the midgut juice. The midgut juice of each pool of animals was diluted with Sg-Ringer solution to the same protein concentration and incubated with 150 ng of dsRNA (≈500bp) for 5 minutes. The analysis was performed by means of agarose gel (1%) electrophoresis. L: ladder; Control dsRNA: dsRNA incubated with Sg-Ringer solution; dsdsR1, dsdsR2, dsdsR3, dsdsR4 and dsgfp: dsRNA incubated with midgut juice of pools of locusts injected, respectively, with transcript-specific dsRNA of dsRNase-1, -2, -3, -4 and gfp. Each well represents one pool, with n≥2.
4.2. Identification of lipophorin dsRNA-binding activity in the hemolymph
Previous work from our lab demonstrated that the desert locust homolog of ApoLpIII is
involved in the ex-vivo dsRNA-binding activity in the hemolymph and it was suggested that
dsgfp dsdsR1 dsdsR2 dsdsR3 dsdsR40.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-1
trans
crip
t lev
el
*****
dsgfp dsdsR1 dsdsR2 dsdsR3 dsdsR40.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-3
trans
crip
t lev
el
**
dsgfp dsdsR1 dsdsR2 dsdsR3 dsdsR40.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-2
trans
crip
t lev
el
**
**
dsgfp dsdsR1 dsdsR2 dsdsR3 dsdsR40.0
0.5
1.0
1.5
Rel
ativ
e dsRNase-4
trans
crip
t lev
el
***
A B
C D
Figure 4: A strong transcript knockdown was obtained for the four dsRNases. (A) dsRNase-1, (B) -2, (C) -3 and (D) -4 transcript levels were determined 6 days after the injection of dsRNA for gfp (dsgfp), dsRNase-1 (dsdsR1), dsRNase-2 (dsdsR2), dsRNase3 (dsdsR3) and dsRNase4 (dsdsR4) (**: p≤0.01; ***: p≤0.001; n≥4; each bar represents the mean ± SEM).

19
the entire lipophorin might bind to the dsRNA [unpublished results]. In this context, we
initially assessed the transcript profiles of the precursors of ApoLpI and ApoLpII, termed
ApoLpI/II, and of ApoLpIII. Our data demonstrated that ApoLpI/II and ApoLpIII have a
pronounced expression in the fat body (Figure 6). In order to test whether lipophorins are the
effectors of the dsRNA-binding activity in the hemocoel, hemolymph from a group of adult
locusts was collected and lipophorins were isolated by means of potassium bromide density
gradient ultracentrifugation. The K+ and Br- ions were (partially) removed from the lipophorin
solution through dialysis. Next, the identity of the purified lipophorins was analyzed with
SDS-PAGE. The predicted sizes of ApoLpII and ApoLpIII are 17 KDa and 76KDa,
respectively, and by means of SDS-PAGE it was indeed possible to detect two clear bands
corresponding to proteins with a mass of ≈20 KDa and ≈70 KDa. In addition, it was possible
to detect protein presence in the wells, likely corresponding to ApoLpI (predicted mass of 294
KDa) (Figure 14, Annex 3). Next, the lipophorin-extract was incubated with dsRNA for 5
minutes. By means of agarose gel electrophoresis, the binding of the lipophorin to the dsRNA
was confirmed by demonstrating a shift in the electrophoretic mobility of the dsRNA
incubated with lipophorins, in comparison to the dsRNA incubated in Sg-Ringer solution (that
functioned as a control) (Figure 7A). Next, we analyzed if lipophorins could protect the
dsRNA from the enzymatic degradation in the midgut juice. Following the 5 minutes
incubation of 150 ng of dsRNA with 10 µl lipophorin solution, 5 µl of midgut juice was
added to digest dsRNA for 5 minutes. As a control, the dsRNA was first incubated with 10 µl
Sg-Ringer solution for 5 minutes and subsequently 5 µl of midgut juice was added to digest
the dsRNA for 5 minutes. In addition, a second control was also taken into account that
consisted of dsRNA incubated with Sg-Ringer solution for 10 minutes. A phenol-chloroform
extraction followed by an ethanol precipitation was performed in order to dissociate the
dsRNA from all dsRNA-binding proteins. The integrity of the dsRNA was analyzed by
comparing the intensity of the bands after electrophoresis. Yet, there were no observable
differences when lipophorins were bound to the dsRNA (Figure 7B). With the aim of testing
whether lipophorins are able to protect dsRNA from the dsRNase activity in the hemolymph,
a similar test was performed with the desert locust serum. Once again, we were unable to
observe clear differences between the degradation condition and the one previously incubated
with the lipophorin extract (Figure 7C).

20
Figure 6: ApoLpI/II and ApoLpIII have a pronounced expression in the fat body. (A) ApoLpI/II and (B) ApoLpIII tissue distributions were measured by means of qPCR. Each bar represents the mean of two independent pools of adult males (40 and 10 animals/pool) and two independent pools of adult females (40 and 10 animals/pool) ± SEM. For the male and female reproductive systems only males or females were used, respectively.
Figure 7: Lipophorins bind to dsRNA but do not perform dsRNA-protection. (A) Lipophorins perform dsRNA-binding activity: a gel mobility shift band was obtained when dsRNA was incubated with lipophorins. The analysis was performed by means of 1% agarose gel electrophoresis. L: ladder. Cont: dsRNA incubated with Sg-Ringer solution for 5 minutes. Lp+dsRNA: dsRNA incubated with S. gregaria lipophorin extract (diluted in Sg-Ringer) for 5 minutes. (B) Lipophorins do not protect the dsRNA against degradation activity in the midgut juice. The midgut juice equally degraded the dsRNA that was previously incubated with lipophorins or Sg-Ringer solution. The integrity of the dsRNA was analyzed through 1% agarose electrophoresis. L: ladder. Cont: dsRNA incubated with Sg-Ringer solution for 10 minutes. MGj+dsRNA: dsRNA incubated with midgut juice for 5 minutes. MGj+Lp+dsRNA: dsRNA incubated first with S. gregaria lipophorin extract for 5 minutes and, second, with midgut juice for 5 minutes. (C) Lipophorins do not perform dsRNA-protection from the dsRNA digestion in the serum. The serum equally degraded the dsRNA that was previously incubated with lipophorins or Sg-Ringer solution. The integrity of the dsRNA was analyzed through 1% agarose electrophoresis..L: ladder. Cont: dsRNA incubated with Sg-Ringer solution for 10 minutes. S+dsRNA: dsRNA incubated with serum for 5 minutes. S+Lp+dsRNA: dsRNA incubated first with S. gregaria lipophorin extract for 5 minutes and, second, with serum for 5 minutes. In every case, 150 ng of dsRNA (≈500bp) were added.
Brain
Corpora
Card
iaca
Corpora
Allata
Thorac
ic Gan
glia
Suboe
soph
agea
l Gan
glion
Testes
Ovarie
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5R
elat
ive ApoLpI/II
trans
crip
t lev
el
Brain
Corpora
Card
iaca
Corpora
Allata
Thorac
ic Gan
glia
Suboe
soph
agea
l Gan
glion
Testes
Ovarie
s
Fat Bod
y
Muscle
Saliva
ry Glan
ds
Foregu
t
Midgut
Hindgu
t0.0
0.5
1.0
1.5
Rel
ativ
e ApoLpIII
trans
crip
t lev
el
A B

21
4.3. The role of lipophorin in the sysRNAi-response
In order to test whether lipophorins play a role in the (sys)RNAi-response, we used the
‘RNAi on RNAi’ approach. This method consists of silencing a test gene with RNAi and
subsequently measuring the effect on the RNAi-potency of a marker gene by means of qPCR.
Therefore, we proceeded to the production of transcript-specific dsRNA fragments for
ApoLpI/II and ApoLpIII (dsApoLpI/II and dsApoLpIII). Next, to obtain a transcript
knockdown, 150 ng of dsRNA of ApoLpI/II or ApoLpIII was injected into the abdominal body
cavity of the adult locusts. Yet, the resulting knockdown was rather moderate (data not
shown). Thus, to improve the RNAi-efficiency, fifth larval stage locusts were intra-
abdominally injected with 300 ng of dsRNA and, immediately after their final moult, locusts
were again injected with 300 ng of dsRNA. The relative transcript levels were measured six
days later in the fat body tissue, by means of qPCR, and compared with these of locusts
injected with dsgfp. Potent knockdowns were obtained for both ApoLpI/II and ApoLpIII
(Figure 8). Afterwards, the locusts received an intra-abdominal injection with transcript-
specific dsRNA for glyceraldehyde 3-phosphate dehydrogenese (gapdh) (dsgapdh). We
selected gapdh as a marker gene since it is one of the most stably expressed genes in S.
gregaria [80]. At the same time, a maximum-knockdown (maximum-KD) control group was
also assessed, in which the delivery of dsApoLpI/II was replaced by the delivery of dsgfp. The
transcript levels of gapdh in the midgut were measured 16 hours later, by qPCR, and
statistically compared between the test group and the respective maximum-KD group.
Previous results of the lab indicated that 16 hours post injection is a time point that allows for
an accurate detection of differences in RNAi-efficiency [21]. Moreover, it was previously
demonstrated that the midgut tissue displayed high sensitivity towards injected dsRNA [21].
In order to assess the physiological expression levels of gapdh, a no-knockdown (no-KD)
control group was also assessed, in which the deliveries of dsApoLpI/II and dsgapdh were
replaced by the delivery of dsgfp. The gapdh levels in the no-KD control group represent the
physiological levels, but were not used for statistical analyses. The results revealed a small
difference in the gapdh expression levels between the group with reduced ApoLpI/II levels
and the maximum-KD control. Yet, the difference was not significant (Figure 9A). The same
experiment was performed for ApoLpIII and, in this case, no differences were observed
between the knockdown levels of gapdh in the test and the control group (Figure 9B).

22
Figure 8: A knockdown of ApoLpI/II and ApoLpIII was obtained. (A) A strong down-regulation of ApoLpI/II was observed. The relative ApoLpI/II transcript level was compared, in the fat body, between a group of locusts injected with dsgfp and a group injected with dsApoLpI/II, 6 days after injection. (B) A strong down-regulation of ApoLpIII was observed. The relative ApoLpIII transcript level in the fat body was compared between a group of locusts injected with dsgfp and a group injected with dsApoLpIII, 6 days after injection. (**: p≤0.01; n≥5; each bar represents the mean ± SEM). The relative transcript levels were measured by means of qPCR.
Figure 9: ‘RNAi on RNAi’ approach on ApoLpI/II and ApoLpIII. (A) Locusts were intra-abdominally injected with dsgfp or dsApoLpI/II and, 6 days later, with dsgfp or dsgapdh, as indicated by dsgfp-dsgfp, dsgfp-dsgapdh and dsApoLpI/II-dsgapdh. 16 hours later, qPCR was performed to determine the gapdh transcript level in the midgut. Statistical analysis was performed between dsgfp-dsgapdh and dsApoLpI/II-dsgapdh (n≥6). (B) Locusts were intra-abdominally injected with dsgfp or dsApoLpIII and, 6 days later, with dsgfp or dsgapdh, as indicated by dsgfp-dsgfp, dsgfp-dsgapdh and dsApoLpIII-dsgapdh. 16 hours later, qPCR was performed to determine the gapdh transcript level in the midgut. Statistical analysis was performed between dsgfp-dsgapdh and dsApoLpIII-dsgapdh (n≥10; each bar represents the mean ± SEM).
4.4. Clathrin-dependent endocytosis in (sys)RNAi
In order to test whether the Clathrin-dependent endocytosis plays a role in the (sys)RNAi
mechanism of the desert locust, the genes of two distinct endocytic components were silenced
in vivo by means of RNAi. The selected test genes, namely the clathrin heavy chain (clath)
and the vacuolar H-ATPase 16 (vha16), are involved in different steps of the clathrin-
mediated endocytosis, specifically in the formation of coated vesicles and in the acidification
of the lysosomes, respectively. In addition, down-regulation of these genes significantly
impaired the uptake of dsRNA in S2 cells [59]. The silencing of clath and vha16 was obtained
dsgfp dsApoLpI/II0.0
0.5
1.0
1.5
**
Rel
ativ
e ApoLpI/II
trans
crip
t lev
el
dsgfp dsApoLpIII0.0
0.5
1.0
1.5
**
Rel
ativ
e ApoLpIII
trans
crip
t lev
el
A B
dsgfp - dsgfp - dsApoLpI/II -0.0
0.5
1.0
1.5
dsgfp dsgapdh dsgapdh
Rel
ativ
e gapdh
trans
crip
t lev
el
dsgfp - dsgfp - dsApoLpIII -0.0
0.5
1.0
1.5
dsgfp dsgapdh dsgapdh
Rel
ativ
e gapdh
trans
crip
t lev
el
A B

23
through intra-abdominal injection of transcript-specific dsRNA fragments (dsclath and
dsvha16) in adult locusts. A period of 6 days after injection was originally chosen in order to
obtain a potent transcript knockdown. However, after the injection of dsclath, mortality was
observed starting from 7 days. Thus, for clath, the knockdown was measured 4 days post-
injection, instead of 6 days, as was the case for vha16. The knockdown levels were assessed
in the midgut tissue by measuring the transcript levels with qPCR and by comparing these
levels with the ones of locusts injected with dsgfp. Potent knockdown levels were obtained for
both test genes (Figure 10). Afterwards, an ‘RNAi on RNAi’ approach was used. Therefore,
locusts were injected with dsclath and, 4 days later, with dsgapdh. At the same time, a
maximum-KD control group was performed. Midgut dissections were performed 16 hours
after the injection of dsgapdh. The transcript levels of gapdh in the midgut were measured by
using qPCR and subsequently statistically compared between the test group and the respective
maximum-KD group. Moreover, in order to assess the physiological expression levels of
gapdh, a no-KD control group was also accounted for, in which the locusts were twice
injected with dsgfp. No statistical analysis was performed with the data of the no-KD group
(Figure 11A). An identical experiment was performed for vha16. However, in this case, the
selected marker gene was alpha-tubulin 1a (tubu), also one of the most stably expressed
genes in S. gregaria [80]. A different marker gene was chosen to ensure that the results were
not specific for gapdh. In addition, the second injections occurred 6 days after the first ones,
instead of 4 days (Figure 11B). According to our data, the knockdown of gapdh and tubu was
significantly less potent when clath and vha16 were down-regulated, respectively (Figure 11).
However, it was still possible that the delivery of dsRNA directed against the test genes
interfered directly with the expression levels of the marker genes. Thus, in order to ensure that
this was not the case, the expression levels of gapdh and tubu of a group of animals treated
individually with dsclath and dsvha16 were compared with these of a group treated with
dsgfp, 4 and 6 days after the injection, respectively (Figure 12). The expression levels of
gapdh and tubu were identical between the analyzed groups.

24
Figure 10: A potent knockdown of clath and vha16 was obtained. (A) A strong down-regulation of clath was observed. The relative clath transcript level was measured by qPCR in the midgut and compared between a group of locusts injected with dsgfp and a group injected with dsclath, 4 days after injection. (B) A strong down-regulation of vha16 was observed. The relative vha16 transcript level was measured in the midgut by qPCR and compared between a group of locusts injected with dsgfp and a group injected with dsvha16, 6 days after injection. (**:p≤0.01; n≥6; each bar represents the mean ± SEM).
Figure 11: Clath and vha16 in the (sys)RNAi mechanism. (A) Locusts were intra-abdominally injected with dsgfp or dsclath and, 4 days later, with dsgfp or dsgapdh, as indicated by dsgfp-dsgfp, dsgfp-dsgapdh and dsclath-dsgapdh. 16 hours later, qPCR was performed to determine gapdh transcript level in the midgut. Statistical analysis was performed between dsgfp-dsgapdh and dsclath-dsgapdh. (B) Locusts were intra-abdominally injected with dsgfp or dsvha16 and, 6 days later, with dsgfp of dstubu, as indicated by dsgfp-dsgfp, dsgfp-dstubu and dsvha16-dstubu. 16 hours later, tubu transcript level in the midgut was assessed by qPCR was performed to determine. Statistical analysis was performed between dsgfp-dstubu and dsvha16-dstubu (*: p≤0.05; **: p≤0.01; n≥8; each bar represents the mean ± SEM).
Figure 12: Injection of dsRNA targeting the test genes did not interfere directly with the marker genes transcript levels. (A) gapdh transcript level was determined 4 days after the injection of dsgfp or dsclath, by qPCR (n≥6); (B) tubu transcript level was determined 6 days after injection of dsgfp or dsvha16, by qPCR (n≥6; each bar represents the mean ± SEM).

25
4.5. Scavenger Receptors in (sys)RNAi
In order to evaluate if SRs play a role in the (sys)RNAi pathway in the desert locust,
we took into account that these receptors constitute a very large family and, instead of using a
RNAi on RNAi technique, we choose to use an approach based on the use of general SRs
inhibitors. This method proved to be efficient in demonstrating that the SRs play a role in the
RNAi response in Drosophila S2 cells [59]. The SRs were blocked by intra-abdominal
injection of well-known competitive inhibitors of the SR-family, namely polyinosine (poly(I))
and dextran sulphate (DS) [59, 82-84]. In the control groups, locusts were treated with
chemically related substances that interact with other receptors but that are not able to bind to
SRs, namely polyadenosine (poly(A)) and chondroitin sulphate (CS) [83]. The structure of
these macromolecules is displayed in Figure 15 (Annex 3). One hour later, the locusts were
injected with dstubu or dsgfp. Sixteen hours later, the knockdown of tubu was determined in
the midgut by means of qPCR. In order to assess the physiological expression level of tubu, a
group of animals was injected with dsgfp and the tubu transcript level in the midgut was
determined sixteen hours later by means of qPCR. Our data show that tubu was significantly
less down-regulated in the groups treated with the SRs inhibitors than in the groups treated
with the corresponding control substances (Figure 13A). To ensure that blocking SRs with the
chosen SRs inhibitors did not interfere with the physiological tubu expression levels directly,
tubu expression levels of groups treated with the SRs inhibitors and control substances were
determined. One hour after the correspondent injections, the measured tubu transcript levels
in the test and in the control groups were indistinguishable (Figure 13B).

26
Figure 13: SRs in the (sys)RNAi mechanism. (A) The locusts were first injected with poly(A), poly(I), CS or DS and, one hour later, with dstubu. The different groups are indicated as poly(A)-dstubu, Poly(I)-dstubu, CS-dstubu and DS-dstubu. A control group indicated as (-)-dsgfp was included but no statistical analysis was performed with the respective data. The transcript levels of tubu were assessed 16 hours latter by means of qPCR. Statistical analysis was performed between poly(A)-dstubu and poly(I)-dstubu, and between CS-dstubu and DS-dstubu groups (**: p≤0.01; ***: p≤0.001; n≥16; each bar represents the mean ± SEM). (B) Similar tubu expression levels, measured by qPCR, were observed between the groups injected with the inhibitors poly(I) and DS, and with the respective control substances Poly(a) and CS. The relative tubu transcript level was measured 1 hour upon the injection (n ≥ 6; each bar represents the mean ± SEM).
5. Discussion
The desert locust is an agricultural pest of serious concern (which is constantly
monitored by FAO - Food and Agriculture Organization) and displays a very robust
(sys)RNAi response upon injection of dsRNA in the hemolymph, which makes it a good
target for RNAi-based pest management [21]. However, this species does not respond to
dsRNA delivered by feeding [unpublished results], compromising the potential pest control
by RNAi. Therefore, understanding the contributing factors for the susceptibility towards
RNAi is an issue of major interest. Research performed in our group has revealed that the
midgut juice of S. gregaria contains dsRNA degrading activity and, based on Bm-dsRNase
[36], four dsRNases were identified in the transcriptome of S. gregaria [unpublished results].
The first objective of this study consisted of testing whether these Sg-dsRNases contribute to
the dsRNase activity in the midgut juice of the desert locust. Similar as reported for the Bm-
dsRNase [36], the expression profiles of Sg-dsRNase-1, -2, -3 and -4 illustrate that these
nucleases are mainly expressed in the midgut (Figure 3). In this context, Sg-dsRNase-1, -2, -3
and -4 constituted good candidates for the dsRNA-degradation role in the midgut juice. In
order to test this, we proceeded to the knockdown of the four dsRNases individually and
succeeded to obtain a strong down-regulation of each Sg-dsRNase (Figure 4) but, due to a

27
marked similarity between the available sequences, off-target effects were also reported in the
midgut of locusts treated with dsRNase-1 and dsRNase-2 dsRNA. However, the off-target
effects were never as strong as the specific down-regulation levels (Figure 4). Notably, the
dsRNase activity observed in the midgut juice of locusts injected with dsRNase-2 dsRNA was
highly impaired (Figure 5), which suggests that dsRNase-2 strongly contributed to the dsRNA
degradation in the midgut juice of S. gregaria. However, since in this group it was also
possible to observe a significant down-regulation of dsRNase-1, it remains possible that a
combination of both dsRNase-1 and -2 is necessary for the strong dsRNase activity in the
midgut juice of the desert locust. In contrast, dsRNase-3 and -4 did not seem to contribute to
the dsRNase activity (Figure 5). Interestingly, the transfection of Bm5 and Hi5 cells with Bm-
dsRNase resulted in a small decrease of the RNAi-response. In addition, there are no reports
of successful RNAi experiments by feeding in B. mori [29]. These facts reinforce the
suggestion that Bm-dsRNase activity in the midgut contributes to the deficiency of the RNAi
response upon dsRNA feeding in B. mori and that, in a similar way, Sg-dsRNase-2, or a
combination of Sg-dsRNase-1 and -2, mainly contributes to the inability to induce RNAi
upon feeding dsRNA to the desert locust.
In L. lineolaris, saliva was demonstrated to rapidly degrade dsRNA, which was
proposed as a major reason for the contrast between the failure of RNAi by dsRNA feeding
and the success of RNAi by injection [32]. Thus, it is important to consider that the dsRNase
activity may take place in different stages of the digestive process, which would contribute
strongly to the deficiency of RNAi through feeding of dsRNA. Curiously, it was suggested
that the effector of the dsRNA digestion activity in L. lineolaris saliva was specific for
dsRNA, which contrasts with the DNA degradation ability of Bm-dsRNase [32, 85]. This
different substrate specificity might indicate that the effectors of the dsRNA degradation in
the digestive cavities belong to different families of nucleases, likely with distinct roles in the
digestive and immune system. However, this is not clear yet and further research is needed.
Interestingly, it has been described that an isoform of Bm-dsRNase can be active and is
involved in the metabolism of nucleic acids in the cytoplasm [29]. In addition, the infection
with a cypovirus was found to induce the expression of Bm-dsRNase [86]. Considering that
viral genomes are protected by the protein capsid in the extracellular medium, the intracellular
dsRNase activity would be important for the degradation of viral genomes and replicative
intermediates in the cytoplasm. Therefore, it is possible to speculate about a potential role of
these dsRNases in the antiviral-immunity.

28
Research from our group identified dsRNA-degradation activity in S. gregaria
hemolymph [unpublished results]. However, in contrast to the situation in the midgut juice,
additional dsRNA-binding activity by ApoLpIII was also detected [unpublished results]. B.
mori hemolymph was likewise demonstrated to contain dsRNA-binding activity and
lipophorins were identified as the effectors of this activity [37]. In this context, the second
objective of this work was to test whether lipophorins might contribute to the dsRNA-binding
activity in the hemolymph of the desert locust. According to our data, ApoLpI/II and ApoLpIII
are mainly expressed in the fat body of the desert locust (Figure 6), which is according to the
fact that lipophorins have a very well established role in the lipid transport process. A
dsRNA-binding assay was performed with lipophorins isolated from the hemolymph of the
desert locust, and the observation of a gel mobility shift (Figure 7A) indicated that lipophorin
indeed possesses dsRNA-binding activity. These data are in line with the dsRNA-binding
activity of lipophorins in the hemolymph of B. mori [37]. Interestingly, by comparing
between the rate of dsRNA degradation in the hemolymph of two insect species with
discrepant susceptibilities to RNAi by dsRNA injection in the hemocoel, it was proposed that
the persistence of dsRNA in the hemolymph could be a determinant factor for the sensitivity
of insects towards RNAi [30]. In this context, it was plausible to consider that lipophorin
could perform dsRNA-protection from the degradation activity in the hemolymph. This
potential role of lipophorin would be in accordance with the fact that the desert locust
displays a robust (sys)RNAi response upon dsRNA-injection into the hemocoel, but is totally
refractory to a RNAi response upon feeding of dsRNA [[21] and unpublished results]. Thus,
the third objective of this work was to test whether lipophorins could protect dsRNA from the
degradation in the hemolymph. Protection assays with midgut juice and serum of the desert
locust were performed but the results did not reveal detectable dsRNA protection by the
lipophorin in any of the two conditions (Figures 7B and 7C). In the midgut juice situation, it
is possible that the lipophorin degradation also occurs, preventing the potential dsRNA
protection, but that is not the case for the hemolymph. Thus, it is possible that lipophorins
play a different role rather than dsRNA-protection.
Notably, vertebrate lipoproteins play an important role in the innate immune system
by recognizing and interacting with pathogen agents, as bacteria, bacterial endotoxins, viruses
and parasites [87]. In insects, lipophorin was also reported to be involved in the immunity
against bacteria, fungi and parasites [41, 42, 44, 45]. Therefore, since dsRNA-structures are
typically associated with the processes of viral infection, it is noteworthy that the lipophorin
dsRNA-binding activity may also perform an anti-viral immunity role. In addition,

29
interestingly, it was proven that the antiviral immune response in Drosophila requires the
systemic RNAi spread [18]. In this context, the fourth objective of this work was to
understand if the dsRNA-binding activity of the lipophorins contributes to the (sys)RNAi
pathway. Therefore we tested the influence of the ApoLpI/II and ApoLpIII in the (sys)RNAi
through an in vivo ‘RNAi on RNAi’ approach. The transcript knockdown for ApoLpI/II and
ApoLpIII was successful (Figure 8), but the ‘RNAi on RNAi’ approach data did not reveal
conclusive results (Figure 9). When ApoLpI/II was down-regulated, the (sys)RNAi response
seemed partially impaired (Figure 9A). However, this result was not significant and, in
addition, a control to ensure that the knockdown of ApoLpI/II did not interfere directly with
the marker (gapdh) gene expression was not performed. In the case of ApoLpIII, it was not
possible to observe any difference in the potency of the (sys)RNAi-response (Figure 9B).
Both cases refer to a situation of knockdown and, therefore, although in reduced levels,
ApoI/II and ApoIII proteins are still present. In this context, considering that the injection of
very small amounts of dsRNA, as 30 ng, is sufficient to cause a strong down-regulation effect
[21], the remaining levels of ApoI/II and ApoIII might still be sufficient to bind (very) small
quantities of dsRNA and still play a role. Thus, it was not possible to infer about the role of
the lipophorin components in the (sys)RNAi of the desert locust.
On the other hand, insect lipophorins and vertebrate lipoproteins can be internalized
by Clathrin-dependent receptor-mediated endocytosis [88-90] and, notably, this endocytic
pathway was proven to mediate dsRNA uptake in D. melanogaster [59, 60]. In this context, a
potential role for lipophorins in the cellular dsRNA-uptake mechanism was considered. This
hypothesis would explain an uptake of dsRNA from the hemocoel but not from the midgut
lumen [since dsRNA binding activity was not detected in the midgut juice [unpublished
results]], which could explain the absence of an interference effect when the dsRNA is
delivered by feeding. Thus, the fifth objective of this work was to investigate if Clathrin-
dependent endocytosis could be involved in dsRNA uptake in the desert locust. In order to do
that, and based on the work reported for D. melanogaster [59, 60], we used an ‘RNAi on
RNAi’ approach. Two genes involved in exogenous dsRNA uptake in the fruit fly were
tested, namely clath and vha16. Similarly to what was reported in Drosophila, the individual
silencing of these genes partially impaired the (sys)RNAi response (Figure 11) [59, 60].
These results indicate that the Clathrin-dependent endocytosis is involved in the (sys)RNAi
mechanism of the desert locust, likely in the dsRNA uptake mechanism.
In Drosophila, pharmacological inhibition tests and RNAi-based silencing of SRs
demonstrated that the dsRNA endocytic uptake is mediated by SRs [59, 60]. Interestingly, in

30
vertebrates, it is known that SRs are able to mediate the uptake of (modified) lipoproteins [91,
92]. Furthermore, in D. melanogaster, it has been proposed that lipophorin can interact with
ninaD, a class B SR, in order to mediate the cellular uptake of carotenoids [66]. This suggests
that, similar to what happens in vertebrates, SRs are mediating the uptake of insect
lipophorins. Moreover, SRs are a major class of pattern recognition receptors that recognize
polyanionic ligands, which makes them good candidates to recognize dsRNA [59]. In this
context, the sixth objective of this work was to test whether the SRs play a role in the
(sys)RNAi response of the desert locust. According to our results, pharmacological inhibition
of the SRs partially impaired the RNAi-pathway (Figure 13A). This data indicates that SRs
are involved in the (sys)RNAi mechanism of the desert locust, likely in the endocytic cellular
uptake of dsRNA, in a similar way to what is observed in Drosophila. Therefore, we suggest
that SR-mediated endocytosis contributes to the dsRNA uptake mechanism of S. gregaria.
Moreover, further research will be needed to understand whether lipophorins play a role in
this process.
In summary, this work reports on (i) the identification of an important effector of the
dsRNA degradation in the midgut, namely the dsRNAse-2, (2) the identification of the
lipophorins as effectors of the additional dsRNA-binding activity in the hemolymph and (3)
the contribution of SR-mediated Clathrin-dependent endocytosis of dsRNA to the (sys)RNAi
mechanism of the desert locust. These results brought important new insights to the
understanding of two key steps in S. gregaria (sys)RNAi response, namely the persistence of
the dsRNA in the body cavities and the cellular uptake of dsRNA. Furthermore, we discussed
the potential role of dsRNases and lipophorins in insect antiviral immunity and the fact that
lipophorin might intervene in the SR-mediated endocytosis of dsRNA in the desert locust.
5.1. Future perspectives
In order to further decipher the influence of the dsRNases and lipophorins on the RNAi-
susceptibility in the desert locust, several experiments can be proposed. For instance, it would
be relevant to test whether impairing the dsRNase-activity in the midgut juice by using RNAi
could confer the locusts susceptibility to orally delivered dsRNA. Secondly, improved
understanding of the role of the dsRNA-binding activity of lipophorins could be achieved by
assessing the involvement of the lipophorin receptor in the RNAi-pathway; therefore, a
‘RNAi on RNAi’ approach could be used. In addition, pharmacological inhibition of the
internalization of lipophorins could also generate more information about the possible role of

31
this protein complex in the (sys)RNAi of the desert locust. Finally, it would still be interesting
to investigate the possible role of Sg-SID-1 in the (sys)RNAi response. An in vivo ‘RNAi on
RNAi’ experiment would be a good approach, as well as the expression of this
transmembrane protein in S2 cells, for instance, followed by the assessment of the imported
dsRNA amount.
6. References 1. Simpson SJ, Sword GA: Locusts. Curr Biol 2008, 18(9):R364-R366. 2. Symmons PM, Cressman K: Biology and behaviour. In: Desert locust guidelines. Food and
Agriculture Organization of the United Nations (FAO) 2001:1-43. 3. Hammond SM, Caudy AA, Hannon GJ: Post-transcriptional gene silencing by double-
stranded RNA. Nature reviews Genetics 2001, 2(2):110-119. 4. Meister G, Tuschl T: Mechanisms of gene silencing by double-stranded RNA. Nature
2004, 431(7006):343-349. 5. Montgomery MK, Fire A: Double-stranded RNA as a mediator in sequence-specific
genetic silencing and co-suppression. Trends in genetics : TIG 1998, 14(7):255-258. 6. Romano N, Macino G: Quelling: transient inactivation of gene expression in Neurospora
crassa by transformation with homologous sequences. Molecular microbiology 1992, 6(22):3343-3353.
7. Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC: Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391(6669):806-811.
8. Montgomery MK, Xu S, Fire A: RNA as a target of double-stranded RNA-mediated genetic interference in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America 1998, 95(26):15502-15507.
9. Hammond SM: Dicing and slicing: the core machinery of the RNA interference pathway. FEBS letters 2005, 579(26):5822-5829.
10. Siomi H, Siomi MC: On the road to reading the RNA-interference code. Nature 2009, 457(7228):396-404.
11. Tomoyasu Y, Miller SC, Tomita S, Schoppmeier M, Grossmann D, Bucher G: Exploring systemic RNA interference in insects: a genome-wide survey for RNAi genes in Tribolium. Genome biology 2008, 9(1):R10.
12. Huvenne H, Smagghe G: Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: a review. Journal of insect physiology 2010, 56(3):227-235.
13. Carthew RW, Sontheimer EJ: Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136(4):642-655.
14. Mello CC, Conte D, Jr.: Revealing the world of RNA interference. Nature 2004, 431(7006):338-342.
15. Tomari Y, Zamore PD: Perspective: machines for RNAi. Genes & development 2005, 19(5):517-529.
16. Siomi MC, Saito K, Siomi H: How selfish retrotransposons are silenced in Drosophila germline and somatic cells. FEBS letters 2008, 582(17):2473-2478.
17. van Rij RP, Saleh MC, Berry B, Foo C, Houk A, Antoniewski C, Andino R: The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes & development 2006, 20(21):2985-2995.

32
18. Saleh MC, Tassetto M, van Rij RP, Goic B, Gausson V, Berry B, Jacquier C, Antoniewski C, Andino R: Antiviral immunity in Drosophila requires systemic RNA interference spread. Nature 2009, 458(7236):346-350.
19. Miller SC, Brown SJ, Tomoyasu Y: Larval RNAi in Drosophila? Development genes and evolution 2008, 218(9):505-510.
20. Belles X: Beyond Drosophila: RNAi In Vivo and Functional Genomics in Insects. Annu Rev Entomol 2010, 55:111-128.
21. Wynant N, Verlinden H, Breugelmans B, Simonet G, Vanden Broeck J: Tissue-dependence and sensitivity of the systemic RNA interference response in the desert locust, Schistocerca gregaria. Insect biochemistry and molecular biology 2012, 42(12):911-917.
22. Uddin MN, Kim JY: Intercellular and systemic spread of RNA and RNAi in plants. Wires Rna 2013, 4(3):279-293.
23. Smardon A, Spoerke JM, Stacey SC, Klein ME, Mackin N, Maine EM: EGO-1 is related to RNA-directed RNA polymerase and functions in germ-line development and RNA interference in C. elegans (vol 10, pg 169, 2000). Curr Biol 2000, 10(10):R393-R394.
24. Sijen T, Fleenor J, Simmer F, Thijssen KL, Parrish S, Timmons L, Plasterk RH, Fire A: On the role of RNA amplification in dsRNA-triggered gene silencing. Cell 2001, 107(4):465-476.
25. Kurscheid S, Lew-Tabor AE, Valle MR, Bruyeres AG, Doogan VJ, Munderloh UG, Guerrero FD, Barrero RA, Bellgard MI: Evidence of a tick RNAi pathway by comparative genomics and reverse genetics screen of targets with known loss-of-function phenotypes in Drosophila. Bmc Mol Biol 2009, 10.
26. Boisson B, Jacques JC, Choumet V, Martin E, Xu JN, Vernick K, Bourgouin C: Gene silencing in mosquito salivary glands by RNAi. FEBS letters 2006, 580(8):1988-1992.
27. Pei Y, Tuschl T: On the art of identifying effective and specific siRNAs. Nat Methods 2006, 3(9):670-676.
28. Cullen BR: Enhancing and confirming the specificity of RNAi experiments. Nat Methods 2006, 3(9):677-681.
29. Liu J, Swevers L, Iatrou K, Huvenne H, Smagghe G: Bombyx mori DNA/RNA non-specific nuclease: expression of isoforms in insect culture cells, subcellular localization and functional assays. Journal of insect physiology 2012, 58(8):1166-1176.
30. Garbutt JS, Belles X, Richards EH, Reynolds SE: Persistence of double-stranded RNA in insect hemolymph as a potential determiner of RNA interference success: Evidence from Manduca sexta and Blattella germanica. Journal of insect physiology 2013, 59(2):171-178.
31. Kamath RS, Martinez-Campos M, Zipperlen P, Fraser AG, Ahringer J: Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans. Genome biology 2001, 2(1).
32. Allen ML, Walker WB, 3rd: Saliva of Lygus lineolaris digests double stranded ribonucleic acids. Journal of insect physiology 2012, 58(3):391-396.
33. Rajagopal R, Sivakumar S, Agrawal N, Malhotra P, Bhatnagar RK: Silencing of midgut aminopeptidase N of Spodoptera litura by double-stranded RNA establishes its role as bacillus thuringiensis toxin receptor. Journal of Biological Chemistry 2002, 277(49):46849-46851.
34. Tian H, Peng H, Yao Q, Chen H, Xie Q, Tang B, Zhang W: Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. Plos One 2009, 4(7):e6225.
35. Tenllado F, Martinez-Garcia B, Vargas M, Diaz-Ruiz JR: Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC biotechnology 2003, 3.
36. Arimatsu Y, Kotani E, Sugimura Y, Furusawa T: Molecular characterization of a cDNA encoding extracellular dsRNase and its expression in the silkworm, Bombyx mori. Insect biochemistry and molecular biology 2007, 37(2):176-183.
37. Sakashita K, Tatsuke T, Masaki Y, Lee JM, Kawaguch Y, Kusakabe T: dsRNA Binding Activity of Silkworm Larval Hemolymph is Mediated by Lipophorin Complex. J Fac Agr Kyushu U 2009, 54(2):401-406.

33
38. Van der Horst DJ, Roosendaal SD, Rodenburg KW: Circulatory lipid transport: lipoprotein assembly and function from an evolutionary perspective. Mol Cell Biochem 2009, 326(1-2):105-119.
39. Canavoso LE, Jouni ZE, Karnas KJ, Pennington JE, Wells MA: Fat metabolism in insects. Annual review of nutrition 2001, 21:23-46.
40. Mandato CA, DiehlJones WL, Downer RGH: Insect hemocyte adhesion in vitro: Inhibition by apoliphorin I and an artificial substrate. Journal of insect physiology 1996, 42(2):143-148.
41. Niere M, Meisslitzer C, Dettloff M, Weise C, Ziegler M, Wiesner A: Insect immune activation by recombinant Galleria mellonella apolipophorin III. Bba-Protein Struct M 1999, 1433(1-2):16-26.
42. Dettloff M, Wittwer D, Weise C, Wiesner A: Lipophorin of lower density is formed during immune responses in the lepidopteran insect Galleria mellonella. Cell Tissue Res 2001, 306(3):449-458.
43. Whitten MM, Tew IF, Lee BL, Ratcliffe NA: A novel role for an insect apolipoprotein (apolipophorin III) in beta-1,3-glucan pattern recognition and cellular encapsulation reactions. Journal of immunology 2004, 172(4):2177-2185.
44. Cheon HM, Shin SW, Bian GW, Park JH, Raikhel AS: Regulation of lipid metabolism genes, lipid carrier protein lipophorin, and its receptor during immune challenge in the mosquito Aedes aegypti. Journal of Biological Chemistry 2006, 281(13):8426-8435.
45. Ma G, Hay D, Li DM, Asgari S, Schmidt O: Recognition and inactivation of LPS by lipophorin particles. Developmental and comparative immunology 2006, 30(7):619-626.
46. Zhuang JJ, Hunter CP: RNA interference in Caenorhabditis elegans: Uptake, mechanism, and regulation. Parasitology 2012, 139(5):560-573.
47. Winston WM, Molodowitch C, Hunter CP: Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 2002, 295(5564):2456-2459.
48. Feinberg EH, Hunter CP: Transport of dsRNA into cells by the transmembrane protein SID-1. Science 2003, 301(5639):1545-1547.
49. Shih JD, Hunter CP: SID-1 is a dsRNA-selective dsRNA-gated channel. Rna-a Publication of the Rna Society 2011, 17(6):1057-1065.
50. Jose AM, Smith JJ, Hunter CP: Export of RNA silencing from C. elegans tissues does not require the RNA channel SID-1. Proceedings of the National Academy of Sciences of the United States of America 2009, 106(7):2283-2288.
51. Aronstein K, Pankiw T, Saldivar E: SID-I is implicated in systemic gene silencing in the honey bee. J Apicult Res 2006, 45(1):20-24.
52. Luo Y, Wang XH, Yu D, Kang L: The SID-1 double-stranded RNA transporter is not required for systemic RNAi in the migratory locust. Rna Biol 2012, 9(5):663-671.
53. Valdes VJ, Athie A, Salinas LS, Navarro RE, Vaca L: CUP-1 Is a Novel Protein Involved in Dietary Cholesterol Uptake in Caenorhabditis elegans. Plos One 2012, 7(3).
54. Labreuche Y, Veloso A, de la Vega E, Gross PS, Chapman RW, Browdy CL, Warr GW: Non-specific activation of antiviral immunity and induction of RNA interference may engage the same pathway in the Pacific white leg shrimp Litopenaeus vannamei. Developmental and comparative immunology 2010, 34(11):1209-1218.
55. Ren R, Xu X, Lin T, Weng S, Liang H, Huang M, Dong C, Luo Y, He J: Cloning, characterization, and biological function analysis of the SidT2 gene from Siniperca chuatsi. Developmental and comparative immunology 2011, 35(6):692-701.
56. Duxbury MS, Ashley SW, Whang EE: RNA interference: a mammalian SID-1 homologue enhances siRNA uptake and gene silencing efficacy in human cells. Biochem Biophys Res Commun 2005, 331(2):459-463.
57. Dong Y, Friedrich M: Nymphal RNAi: systemic RNAi mediated gene knockdown in juvenile grasshopper. BMC biotechnology 2005, 5:25.
58. Xu WN, Han ZJ: Cloning and phylogenetic analysis of sid-1-like genes from aphids. J Insect Sci 2008, 8.

34
59. Saleh MC, van Rij RP, Hekele A, Gillis A, Foley E, O'Farrell PH, Andino R: The endocytic pathway mediates cell entry of dsRNA to induce RNAi silencing. Nature cell biology 2006, 8(8):793-802.
60. Ulvila J, Parikka M, Kleino A, Sormunen R, Ezekowitz RA, Kocks C, Ramet M: Double-stranded RNA is internalized by scavenger receptor-mediated endocytosis in Drosophila S2 cells. The Journal of biological chemistry 2006, 281(20):14370-14375.
61. Areschoug T, Gordon S: Scavenger receptors: role in innate immunity and microbial pathogenesis. Cell Microbiol 2009, 11(8):1160-1169.
62. Ramet M, Pearson A, Manfruelli P, Li XH, Koziel H, Gobel V, Chung E, Krieger M, Ezekowitz RAB: Drosophila scavenger receptor Cl is a pattern recognition receptor for bacteria. Immunity 2001, 15(6):1027-1038.
63. Gough PJ, Gordon S: The role of scavenger receptors in the innate immune system. Microbes Infect 2000, 2(3):305-311.
64. Franc NC, Dimarcq JL, Lagueux M, Hoffmann J, Ezekowitz RAB: Croquemort, a novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity 1996, 4(5):431-443.
65. Hart K, Wilcox M: A Drosophila gene encoding an epithelial membrane protein with homology to CD36/LIMP II. Journal of molecular biology 1993, 234(1):249-253.
66. Kiefer C, Sumser E, Wernet MF, von Lintig J: A class B scavenger receptor mediates the cellular uptake of carotenoids in Drosophila. Proceedings of the National Academy of Sciences of the United States of America 2002, 99(16):10581-10586.
67. Kocks C, Cho JH, Nehme N, Ulvila J, Pearson AM, Meister M, Strom C, Conto SL, Hetru C, Stuart LM et al: Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell 2005, 123(2):335-346.
68. Erturk-Hasdemir D, Silverman N: Eater: a big bite into phagocytosis. Cell 2005, 123(2):190-192.
69. Hori S, Kobayashi A, Natori S: A novel hemocyte-specific membrane protein of Sarcophaga (flesh fly). Eur J Biochem 2000, 267(17):5397-5403.
70. Zhou Z, Hartwieg E, Horvitz HR: CED-1 is a transmembrane receptor that mediates cell corpse engulfment in C-elegans. Cell 2001, 104(1):43-56.
71. Peiser L, De Winther MP, Makepeace K, Hollinshead M, Coull P, Plested J, Kodama T, Moxon ER, Gordon S: The class A macrophage scavenger receptor is a major pattern recognition receptor for Neisseria meningitidis which is independent of lipopolysaccharide and not required for secretory responses. Infect Immun 2002, 70(10):5346-5354.
72. Peiser L, Makepeace K, Pluddemann A, Savino S, Wright JC, Pizza M, Rappuoli R, Moxon ER, Gordon S: Identification of Neisseria meningitidis nonlipopolysaccharide ligands for class A macrophage scavenger receptor by using a novel assay. Infect Immun 2006, 74(9):5191-5199.
73. Amiel E, Acker JL, Collins RM, Berwin B: Uncoupling Scavenger Receptor A-Mediated Phagocytosis of Bacteria from Endotoxic Shock Resistance. Infect Immun 2009, 77(10):4567-4573.
74. McEwan DL, Weisman AS, Hunter CP: Uptake of extracellular double-stranded RNA by SID-2. Molecular cell 2012, 47(5):746-754.
75. Winston WM, Sutherlin M, Wright AJ, Feinberg EH, Hunter CP: Caenorhabditis elegans SID-2 is required for environmental RNA interference. Proceedings of the National Academy of Sciences of the United States of America 2007, 104(25):10565-10570.
76. Jose AM, Kim YA, Leal-Ekman S, Hunter CP: Conserved tyrosine kinase promotes the import of silencing RNA into Caenorhabditis elegans cells. Proceedings of the National Academy of Sciences of the United States of America 2012, 109(36):14520-14525.
77. Hinas A, Wright AJ, Hunter CP: SID-5 Is an Endosome-Associated Protein Required for Efficient Systemic RNAi in C. elegans. Curr Biol 2012, 22(20):1938-1943.
78. Tijsterman M, May RC, Simmer F, Okihara KL, Plasterk RH: Genes required for systemic RNA interference in Caenorhabditis elegans. Curr Biol 2004, 14(2):111-116.

35
79. Grishok A: RNAi mechanisms in Caenorhabditis elegans. FEBS letters 2005, 579(26):5932-5939.
80. Van Hiel MB, Van Wielendaele P, Temmerman L, Van Soest S, Vuerinckx K, Huybrechts R, Broeck JV, Simonet G: Identification and validation of housekeeping genes in brains of the desert locust Schistocerca gregaria under different developmental conditions. Bmc Mol Biol 2009, 10.
81. Pattyn F, Speleman F, De Paepe A, Vandesompele J: RTPrimerDB: the real-time PCR primer and probe database. Nucleic acids research 2003, 31(1):122-123.
82. Whitman SC, Daugherty A, Post SR: Regulation of acetylated low density lipoprotein uptake in macrophages by pertussis toxin-sensitive G proteins. Journal of lipid research 2000, 41(5):807-813.
83. Pearson AM, Rich A, Krieger M: Polynucleotide binding to macrophage scavenger receptors depends on the formation of base-quartet-stabilized four-stranded helices. The Journal of biological chemistry 1993, 268(5):3546-3554.
84. Krieger M, Herz J: Structures and functions of multiligand lipoprotein receptors: macrophage scavenger receptors and LDL receptor-related protein (LRP). Annual review of biochemistry 1994, 63:601-637.
85. Arimatsu YF, T.; Sugimura,Y.; Togoh, M.; Ishihara, R.; Tokizane, M.; Kotani, E.; Hayashi, Y.; Furusawa, T.: Purification and Properties of Double-stranded RNA-degrading Nuclease, dsRNase, from the Digestive Juice of the Silkworm, Bombyx mori. Journal of Insect Biotechnology and Sericology 2007, 76:57-62
86. Wu P, Li MW, Wang X, Zhao P, Wang XY, Liu T, Qin GX, Guo XJ: Differentially expressed genes in the midgut of Silkworm infected with cytoplasmic polyhedrosis virus. Afr J Biotechnol 2009, 8(16):3711-3720.
87. Khovidhunkit W, Kim MS, Memon RA, Shigenaga JK, Moser AH, Feingold KR, Grunfeld C: Effects of infection and inflammation on lipid and lipoprotein metabolism: mechanisms and consequences to the host. Journal of lipid research 2004, 45(7):1169-1196.
88. Dantuma NP, Potters M, De Winther MPJ, Tensen CP, Kooiman FP, Bogerd J, Van der Horst DJ: An insect homolog of the vertebrate very low density lipoprotein receptor mediates endocytosis of lipophorins. Journal of lipid research 1999, 40(5):973-978.
89. Willnow TE, Nykjaer A, Herz J: Lipoprotein receptors: new roles for ancient proteins. Nature cell biology 1999, 1(6):E157-E162.
90. Mousavi SA, Malerod L, Berg T, Kjeken R: Clathrin-dependent endocytosis. Biochem J 2004, 377:1-16.
91. Dhaliwal BS, Steinbrecher UP: Scavenger receptors and oxidized low density lipoproteins. Clin Chim Acta 1999, 286(1-2):191-205.
92. Xu SZ, Laccotripe M, Huang XW, Rigotti A, Zannis VI, Krieger M: Apolipoproteins of HDL can directly mediate binding to the scavenger receptor SR-BI, an HDL receptor that mediates selective lipid uptake. Journal of lipid research 1997, 38(7):1289-1298.
6.1. Web-references • http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?mode=Info&id=7010&lvl=3&lin=f
&keep=1&srchmode=1&unlock • http://www.rnaiweb.com/RNAi/What_is_RNAi/ • http://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?sid=415248 • http://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?sid=586312 • http://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?sid=135023145&loc=es_rss • http://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?sid=134989364&loc=es_rss

36
i) Annex 1
Fw-primer Rv-primer dsRNase-1 TAATACGACTCACTATAGGG
CCTTTCCAAAGGACACCTCA TAATACGACTCACTATAGGG CCTGCGGGTTTTTGAAGATA
dsRNase-2 TAATACGACTCACTATAGGG GGACACCTCGCTGCTAAGTC
TAATACGACTCACTATAGGG CGATGTACGGATTGTTGACG
dsRNase-3 TAATACGACTCACTATAGGG GCTAGCGGTAGCAGGAACAC
TAATACGACTCACTATAGGG ACTGCCCACCACAGTACCTC
dsRNase-4 TAATACGACTCACTATAGGG TCGTCTACGAGCCGAAATCT
TAATACGACTCACTATAGGG TCGTGGATCACTTGCTAACG
ApoLpI/II TAATACGACTCACTATAGGG TTTGAGATTGCTGCACGAAC
TAATACGACTCACTATAGGG CATCAACAAGCAGCTTTCCA
ApoLpIII TAATACGACTCACTATAGGG CAGGTGAACATCGCAGAGAC
TAATACGACTCACTATAGGG GGTGCCCTGGTGCTTCTC
gapdh TAATACGACTCACTATAGGG CCGTTGCTGTCGGTTCGTAGGAA
TAATACGACTCACTATAGGG TTGGGGCATCTGCACTTGGA
clath TAATACGACTCACTATAGGG AAATCTTGATCGGGCATACG
TAATACGACTCACTATAGGG CAATGTGAAGGCCACA
vha16 TAATACGACTCACTATAGGG TGCGACCAGAGCTGATTATG
TAATACGACTCACTATAGGG GAGGTGGCTGCTGTAGGAAG
tubu TAATACGACTCACTATAGGG ATTTTTTAGCGAAACTGGTGCTGG
G
TAATACGACTCACTATAGGG TGGTGTAAGTCGGGCGTTCAATGT
Fw-primer Rv-Primer
ubi GACTTTGAGGTGTGGCGTAG GGATCACAAACACAGAACGA
ef1a GATGCTCCAGGCCACAGAGA TGCACAGTCGGCCTGTGAT
dsRNase-1 GCTCATCGACGCCTGCTT CGGCGACCATCGTGAAGT
dsRNase-2 CCGAGACCAACGACTACTACTTCCT CGCGCCGAGCATGAA
dsRNase-3 CGCCCCAGCGACAGACT CGAGTTGACGCCGAAGAAG
dsRNase-4 CCACAGTGGTACGCCTTCAA AAGTCGCGCACGTCCAA
ApoLpI/II CAGTGCAAAGGCAGCATTGA CAGTGCCGCTTGTCGCTAAT
ApoLpIII GCGCGCCCAGATGCT TTGACGATCGTGTGGTTCAGTT
gapdh GTCTGATGACAACAGTGCAT GTCCATCACGCCACAACTTTC
clath AAGGACGCAATGGAGTATGCA TTTCCAGAAACCACGCAAGTAA
vha16 CTGCATACGGAACTGCAAGT CATAATCAGCTCTGGTCGATT
tubu CAGCCCGATGGGCAGAT CGCAAAGATGCTGTGATTGA
Table 4: Sequences of the primers used for the amplification of the DNA template for synthesis of dsRNA of dsRNase-1, -2, -3 and -4, ApoLpI/II, ApoLpIII, gapdh, clath, vha16 and tubu. The bold sequences correspond to the T7-promoter site.
Table 5: Sequences of the qRT-PCR primers of ubi, ef1a, dsRNase-1, -2, -3 and -4, ApoLpI/II,ApoLpIII, gapdh, clath, vha16 and tubu.

37
ii) Annex 2
Ultracentrifugation in a potassium bromide density gradient for the purification of lipophorins
• Dilute the hemolymph to a volume of 5mL, in a dilution solution consisting in 150
mM NaCl and 5 mM EDTA in 50 mM phosphate buffer, PH=6.8;
• Add 2.2 g KBr the solution;
• Add 5.5 mL of 0.9% NaCl to the top;
• Centrifuge for 10 hours, 4°C, 50,000 rpm;
• Collect the yellow band with a syringe.
Phenol/Chloroform extraction and Ethanol precipitation
• Prepare following reaction mix:
• 1 volume dsRNA (50 µl);
• 1/2 volume phenol (25 µl);
• 1/2 volume chloroform (25 µl);
• Pipette the reaction mix up and down and incubate 2-3 min at room temperature;
• Centrifuge 15 min at 13000 rpm at 4 °C, the upper layer contains the dsRNA;
• Prepare following reaction mix (ethanol precipitation):
• 1 volume dsRNA solution (upper layer);
• 3 volumes ethanol 100 %;
• 1/10 volume sodium acetate 3M;
• 2 µl glycogen;
• Incubate 1.5 h at -20 °C;
• Centrifuge 30 min at 13000 rpm at 4 °C and discard the supernatant;
• Add 1 mL ethanol 70 %;
• Centrifuge 10 min 13000 rpm at 4 °C and discard the supernatant;
• Resuspend the pellet in the desired volume of Milli-Q water.

38
iii) Annex 3
Figure 14: Bands with the size approximately corresponding to the predicted S. gregaria homologs of ApoII (≈70 KDa) and Apo III (≈20 KDa) are observable in the SDS-PAGE analysis of the lipophorin extract.
Figure 15: Structures of the scavenger receptors inhibitors (A and B) and control substances (C and D). (A) Polyinosine. (B) Dextran Sulphate. (C) Polyadenosine. (D) Chondroitin Sulphate. [National Center for Biotechnology Information].
17 KDa -
28 KDa -
62 KDa -
98 KDa -