kallikrein gene transfer protects against ischemic stroke...
TRANSCRIPT

Kallikrein Gene Transfer Protects Against Ischemic Strokeby Promoting Glial Cell Migration and Inhibiting Apoptosis
Chun-Fang Xia, Hang Yin, Cesar V. Borlongan, Lee Chao, Julie Chao
Abstract—Kallikrein/kinin has been shown to protect against ischemia/reperfusion-induced myocardial infarction andapoptosis. In the present study, we examined the potential neuroprotective action of kallikrein gene transfer in cerebralischemia. Adult, male Sprague-Dawley rats were subjected to a 1-hour occlusion of the middle cerebral artery followedby intracerebroventricular injection of adenovirus harboring either the human tissue kallikrein gene or the luciferasegene. Kallikrein gene transfer significantly reduced ischemia-induced locomotor deficit scores and cerebral infarctionafter cerebral ischemia injury. Expression of recombinant human tissue kallikrein was identified and localized inmonocytes/macrophages of rat ischemic brain by double immunostaining. Morphological analyses showed thatkallikrein gene transfer enhanced the survival and migration of glial cells into the ischemic penumbra and core, asidentified by immunostaining with glial fibrillary acidic protein. Cerebral ischemia markedly increased apoptotic cells,and kallikrein gene delivery reduced apoptosis to near-normal levels as seen in sham control rats. In primary culturedglial cells, kinin stimulated cell migration but inhibited hypoxia/reoxygenation-induced apoptosis in a dose-dependentmanner. The effects of kinin on both migration and apoptosis were abolished by icatibant, a bradykinin B2 receptorantagonist. Enhanced cell survival after kallikrein gene transfer occurred in conjunction with markedly increasedcerebral nitric oxide levels and phospho-Akt and Bcl-2 levels but reduced caspase-3 activation, NAD(P)H oxidaseactivity, and superoxide production. These results indicate that kallikrein gene transfer provides neuroprotection againstcerebral ischemia injury by enhancing glial cell survival and migration and inhibiting apoptosis through suppression ofoxidative stress and activation of the Akt–Bcl-2 signaling pathway. (Hypertension. 2004;43[part 2]:1-8.)
Key Words: ischemia � stroke � kinins � gene transfer � apoptosis
Stroke is the third leading cause of death and the mostcommon cause of disability in the United States. Al-
though drug therapy remains the sole choice of treatment forstroke patients, there has been no conclusive evidence oflong-lasting motor and cognitive improvement with any ofthe current drugs.1 Therefore, a search for suitable regimensto rescue the central nervous system after ischemia has beena major research endeavor.
Reperfusion injury is thought to play a critical role in thepathophysiology of cerebral ischemia. Reactive oxygen spe-cies (ROS) are implicated in reperfusion injury after cerebralischemia.2 Increased oxidative stress during reperfusion ex-acerbates cerebral ischemia–induced neuronal apoptosis.3,4
Tissue kallikrein is capable of cleaving low-molecular-weightkininogen to form vasoactive kinin peptides.5 The ability ofkinin to dilate cerebral arterial vessels is due in part to therelease of endothelium-derived relaxing factor/nitric oxide(NO).6 Other than vasodilation, recent studies have shownthat kallikrein/kinin also plays a vital role in angiogenesis andapoptosis in response to hindlimb or cardiac ischemia.7–10 NOhas been known to play a complex role in cerebral ischemia.
Excitotoxic or ischemic conditions excessively activate neu-ronal NO synthase (NOS), resulting in production of NO thatis toxic to surrounding neurons.11 Inducible NOS, which isnot normally present in healthy tissues, is induced shortlyafter ischemia and contributes to secondary late-phase dam-age.11 Conversely, NO generated from endothelial NOS(eNOS) is critical in maintaining cerebral blood flow andreducing infarct volume.11 Upregulation of eNOS resulted inneuroprotection against cerebral ischemia and reduction ofstroke damage.12 Taken together, these results suggest thatthe tissue kallikrein/kinin system, through activation of eNOSand subsequent NO production, might modulate endothelialfunction and promote cell survival and could have protectiveeffects on the brain in the setting of ischemic stroke.
Our previous study demonstrated that adenovirus-mediatedgene transfer of human tissue kallikrein ameliorated a salt-induced blood pressure rise and cerebral damage and reducedthe stroke-induced mortality rate in Dahl salt-sensitive rats.13
To determine whether kallikrein/kinin has neuroprotectiveeffects against ischemic stroke, we used a rat model of focalcerebral ischemia by 1-hour occlusion of the middle cerebral
Received September 29, 2003; first decision October 31, 2003; revision accepted November 17, 2003.From the Department of Biochemistry and Molecular Biology (C.-F.X., H.Y., L.C., J.C.), Medical University of South Carolina, Charleston, and the
Department of Neurology and Institute of Molecular Medicine and Genetics (C.V.B.), Medical College of Georgia, Augusta.Correspondence to Julie Chao, PhD, Department of Biochemistry and Molecular Biology, Medical University of South Carolina, 173 Ashley Ave,
Charleston, SC 29425. E-mail [email protected]© 2003 American Heart Association, Inc.
Hypertension is available at http://www.hypaha.org DOI: 10.1161/01.HYP.0000110905.29389.e5
1
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

artery (MCA) followed by reperfusion14 and immediateintracerebroventricular injection of adenovirus carrying thehuman tissue kallikrein gene. We examined the potentialeffects of kallikrein gene transfer on cerebral cell survival andangiogenesis in the brain after ischemia/reperfusion (I/R).Our results show that kallikrein/kinin protects against ische-mic stroke, characterized by a significant reduction in motordeficits, cerebral infarction, and apoptosis, as well as promo-tion of glial cell survival and migration.
MethodsAnimals and TreatmentsTwenty-four male, Sprague-Dawley rats (225 to 250 g body weight;Harlan Sprague-Dawley, Indianapolis, Ind) were used in this exper-iment. All procedures complied with standards for the care and useof animal subjects, as stated in the Guide for the Care and Use ofLaboratory Animals (Institute of Laboratory Resources, NationalAcademy of Sciences, Bethesda, Md).
Cerebral I/R surgery was performed as previously described.14 Inbrief, the rats were anesthetized intraperitoneally with ketamine/xylazine (90 mg/10 mg per kg body weight). A nylon filament(Ethicon 4-0, with the tip diameter tapered to the size of a 26-gaugeneedle) was inserted from the right external carotid artery by way ofthe internal carotid artery to the base of the right MCA to stop theblood flow. After 1 hour of ischemia, the filament was withdrawn,blood reperfusion was established, and the skin was sutured. Ade-novirus containing human tissue kallikrein cDNA (Ad.CMV-TK) orcontrol virus (Ad.CMV-Luc) (1�1011 plaque-forming units in 10�L)10 was stereotactically delivered into the right lateral cerebralventricle (1.5 mm lateral, 1.0 mm posterior to the bregma and4.5 mm deep from the skull surface) immediately after reperfusion(n�9). Body temperature of the animals was maintained at 37°Cduring surgery until they recovered from anesthesia. An additionalgroup of animals (n�6) underwent sham surgery as controls.
Behavioral TestRats were tested for neurologic deficits after cerebral I/R. Thescoring scale was as follows: 0�no observable neurologic deficits;1�failure to extend right forepaw (mild); 2�circling to the contralat-eral (moderate); and 3�loss of walking or righting reflex (severe).15
Rats with neurologic deficit scores �2 after full recovery fromanesthesia and surgery were used in the experiment.
Cerebral Infarct Staining With TTCAt day 9 after surgery, the animals were again anesthetized intra-peritoneally with ketamine/xylazine (90 mg/10 mg per kg bodyweight) and perfused transcardially with 0.9% saline, and the brainswere removed and weighed. Serial coronal brain sections (�2 mmthick) were immersed in normal saline containing 2% 2,3,5-triphe-nyltetrazolium chloride (TTC) for 30 minutes at 37°C. To minimizeartifacts produced by postischemic edema in the infarcted area, theinfarct volume was calculated with an alternate technique, aspreviously described.16 In brief, the infarcted area in the ipsilateralhemisphere was indirectly measured by subtracting the noninfarctedarea in the ipsilateral hemisphere from the total intact area of thecontralateral hemisphere.
Immunohistochemistry andDouble ImmunofluorescenceSections (5 �m) of paraffin-embedded brain were subjected toimmunohistochemistry with use of a staining kit (Universal EliteABC, Vector) and performed according to the manufacturer’sinstructions. An affinity-purified anti-human kallikrein antibody (5�g/mL)17 and an anti-glial fibrillary acidic protein (GFAP, Chemi-con) antibody were used for immunostaining. For double immuno-fluorescence analyses, sections were incubated with a mixture ofanti-human kallikrein (5 �g/mL) and anti-monocyte/macrophage
(ED1) antibodies (Chemicon), followed by a mixture of anti-mouseIgG–tetramethylrhodamine B isothiocyanate and anti-rabbit IgG–fluorescein isothiocyanate antibodies (Sigma). Cerebral apoptosiswas analyzed by TdT-mediated dUTP nick end-labeling (TUNEL)assay.18 The TUNEL-positive (apoptotic) cells were counted in 10microscopic fields of each brain section.
ELISA for Human Tissue KallikreinThe levels of immunoreactive human tissue kallikrein in rat brainwere measured by ELISA.19
Primary Cultured Glial Cells and Migration AssayIn brief, brains were removed from fetal Sprague-Dawley rats onembryonic day 15 and incubated in Hanks’ solution chilled on ice.The tissues were chopped into small pieces and incubated withCa2�-free Hanks’ solution containing trypsin (0.05 mg/mL) andcollagenase (0.01 mg/mL) at 37°C for 30 minutes, followed by theaddition of soybean trypsin inhibitor (0.1 mg/mL) and DNase (0.1mg/mL). The tissue was then centrifuged for 5 minutes (1000 rpm),and the pellet was resuspended in Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 10% fetal bovine serum andD-glucose (4.5 mg/mL). Cells were plated on 60-mm tissue-culturedishes. The culture dishes were kept in a humidified incubator under5% CO2 and 95% air at 37°C for 14 to 21 days. After 14 to 21 days,glial cells were stained by GFAP antibody for confirmation.
Cell migration assays20 were performed in 24-well modifiedBoyden transwell microchambers (Costar) containing 8-�m poresize polyethylene terephthalate. The lower surface of each filter wascoated with phosphate-buffered saline containing 6.5 �g/mL ratfibronectin (30 minutes at 37°C) and then blocked with 1% bovineserum albumin in serum-free DMEM for 30 minutes at 37°C. Foreach assay, 104 cells were resuspended in 200 �L of serum-freeDMEM and plated on the upper chamber, and the lower wells werefilled with either serum-free DMEM (background migration),DMEM with bradykinin (0.1, 1, or 5 �mol/L), in the absence orpresence of icatibant (5 �mol/L), or DMEM supplemented with 10%fetal calf serum (maximal migration control) and incubated at 37°Cfor 6 hours. Cells remaining in the upper surface of the filter wereremoved with a cotton swab, and cells that had migrated (lowersurface) were fixed and stained with a Diff-Quik stain set (DadeBehring). The number of cells that had migrated through the filterwas counted under an inverted microscope and expressed as thepercentage of migrated cells measured in the presence of 10% fetalcalf serum.
Hypoxia/Reoxygenation-Induced GlialCell ApoptosisCultured glial cells were growth-arrested in serum-free medium for18 hours at 37°C before the experiments. Cells were incubated for 12hours under hypoxic conditions (95% N2 and 5% CO2), followed by24-hour reoxygenation (95% O2 and 5% CO2). Before hypoxia/reox-ygenation (H/R), glial cells were incubated with bradykinin (0.1 and1 �mol/L) with or without icatibant (10 �mol/L) for 60 minutes.Apoptotic glial cells were identified (fixed in 4% paraformaldehyde)by TUNEL staining (Roche), as previously described.13 The positivecells were determined by counting �400 to 500 glial cells in 5randomly chosen fields.
Western Blot Analyses of Bcl-2, Akt, andCleaved Caspase-3Brain extracts (80 to 120 �g) were subjected to Western blotanalyses.10 Antibodies to Bcl-2 (Santa Cruz Biotech), phospho-Akt,total Akt (Cell Signaling Technology), and cleaved caspase-3 (CellSignaling Technology) were used in the study.
Caspase-3 Activity AssayBrain extracts were incubated at 37°C for 30 minutes with caspase-3fluorogenic substrate II (Calbiochem).21 To measure nonspecifichydrolysis of caspase-3 substrate II, a preparation of an inhibitor-
2 Hypertension February 2004 Part II
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

treated brain extract was used. Caspase-3 activity was quantified ina Perkin-Elmer fluorometer with excitation at 380 nm and emissionat 460 nm.
Measurements of Nitrate and Nitrite Levels,NAD(P)H Oxidase Activity, andSuperoxide FormationNitrate and nitrite (NOx) levels in brain extracts were measured aspreviously described.22 NADH/NADPH oxidase activity was mea-sured by lucigenin-enhanced chemiluminescent detection of super-oxide in a luminometer (TD-20/20, Turner Designs).23 Superoxideproduction was measured by ferricytochrome c reduction, and thesuperoxide dismutase–inhibitable reduction of cytochrome c wasmeasured at 550 nm.24
Statistical AnalysisResults are expressed as mean�SEM. Comparisons among groupswere made by ANOVA followed by Fisher protected least significantdifference test or by unpaired Student t test. Differences wereconsidered significant at P�0.05.
ResultsKallikrein Gene Transfer Reduced NeurologicDeficit Scores and Cerebral InfarctionNeurologic deficit scoring was performed at days 1 and 9after gene transfer. At day 9, neurologic deficit scores weresignificantly decreased in the kallikrein group compared withthose in the luciferase group after cerebral I/R (0.44�0.24versus 1.67�0.49; n�9, P�0.05; Figure 1A). However, therewas no difference in neurologic deficit scores between the 2groups at 1 day after gene delivery (data not shown). Figure1B shows representative images of serial coronal brainsections stained with TTC from each group. Brain sectionsfrom the sham rat without infarction are stained red. Focalcerebral infarction in the right hemisphere in the rat from theAd.CMV-Luc group after cerebral I/R is stained white. Afterkallikrein gene delivery, brain sections appeared reddish andrelatively normal. The total infarct volume in theAd.CMV-TK group was significantly reduced compared withthat of the Ad.CMV-Luc group (11.6�0.7 versus 8.4�1.8 mm3/rat; P�0.05, n�4; Figure 1C).
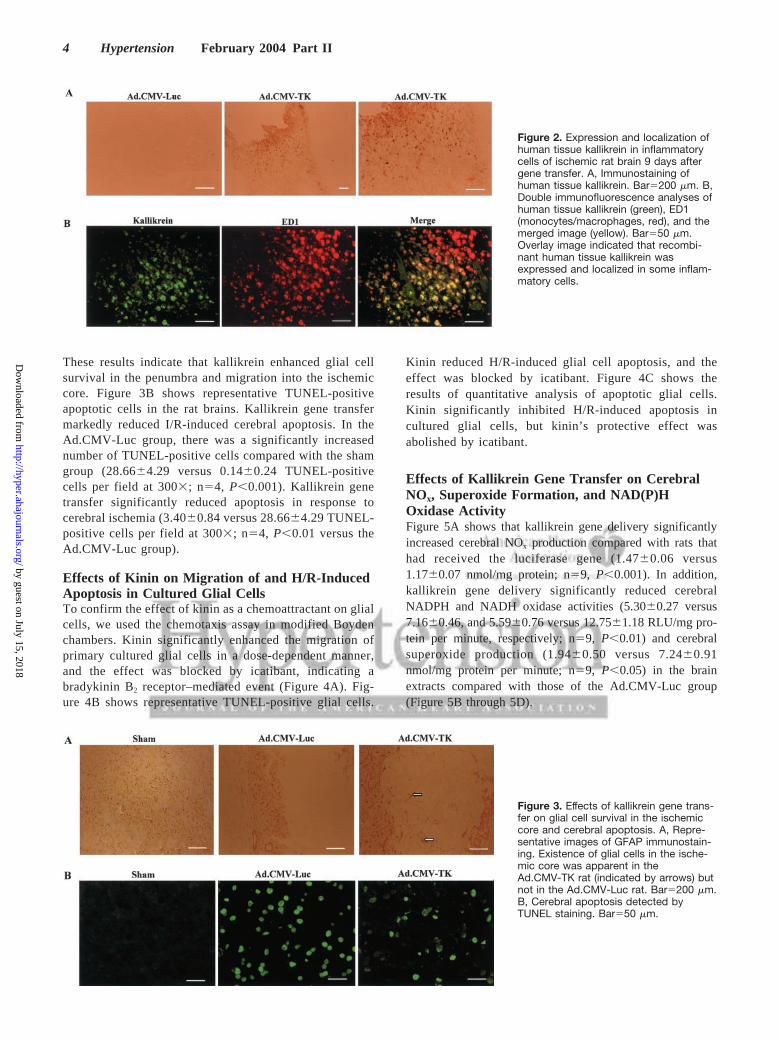
Expression and Localization of Human TissueKallikrein in Ischemic Rat Brain AfterGene TransferRecombinant human kallikrein in the rat brain after genedelivery was identified by double immunostaining andELISA. Figure 2A shows representative immunostaining ofhuman tissue kallikrein in the ischemic brains of rats that hadreceived kallikrein gene transfer. Figure 2B shows doubleimmunofluorescence of human tissue kallikrein and ED1 inthe ischemic brains of rats. Human tissue kallikrein and ED1were colocalized near areas of tissue damage after genetransfer. Human kallikrein levels in the rat brain reached alevel of 0.29�0.12 ng/mg protein (n�3), as measured byELISA 9 days after kallikrein gene delivery but were notdetected in control rats.
Effects of Kallikrein Gene Transfer on Glial CellSurvival and Cerebral ApoptosisFigure 3A shows immunostaining of GFAP, a marker forglial cells. Increased staining of GFAP surrounding the
ischemic core was observed in the Ad.CMV-Luc group,indicating activation of glial cells or reactive gliosis. In theAd.CMV-TK group, GFAP staining not only accumulated inthe penumbra but also appeared inside the ischemic core.
Figure 1. Neurologic deficit scores of rats and TTC stainingafter cerebral I/R injury. A, Scoring was performed at day 9 afterintracerebroventricular gene delivery (n�9). B, Representativeimages of serial coronal brain sections with TTC staining. C,Quantitative analysis of infarct volume (n�4).
Xia et al Kallikrein Gene Transfer in Ischemic Stroke 3
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from
These results indicate that kallikrein enhanced glial cellsurvival in the penumbra and migration into the ischemiccore. Figure 3B shows representative TUNEL-positiveapoptotic cells in the rat brains. Kallikrein gene transfermarkedly reduced I/R-induced cerebral apoptosis. In theAd.CMV-Luc group, there was a significantly increasednumber of TUNEL-positive cells compared with the shamgroup (28.66�4.29 versus 0.14�0.24 TUNEL-positivecells per field at 300�; n�4, P�0.001). Kallikrein genetransfer significantly reduced apoptosis in response tocerebral ischemia (3.40�0.84 versus 28.66�4.29 TUNEL-positive cells per field at 300�; n�4, P�0.01 versus theAd.CMV-Luc group).
Effects of Kinin on Migration of and H/R-InducedApoptosis in Cultured Glial CellsTo confirm the effect of kinin as a chemoattractant on glialcells, we used the chemotaxis assay in modified Boydenchambers. Kinin significantly enhanced the migration ofprimary cultured glial cells in a dose-dependent manner,and the effect was blocked by icatibant, indicating abradykinin B2 receptor–mediated event (Figure 4A). Fig-ure 4B shows representative TUNEL-positive glial cells.
Kinin reduced H/R-induced glial cell apoptosis, and theeffect was blocked by icatibant. Figure 4C shows theresults of quantitative analysis of apoptotic glial cells.Kinin significantly inhibited H/R-induced apoptosis incultured glial cells, but kinin’s protective effect wasabolished by icatibant.
Effects of Kallikrein Gene Transfer on CerebralNOx, Superoxide Formation, and NAD(P)HOxidase ActivityFigure 5A shows that kallikrein gene delivery significantlyincreased cerebral NOx production compared with rats thathad received the luciferase gene (1.47�0.06 versus1.17�0.07 nmol/mg protein; n�9, P�0.001). In addition,kallikrein gene delivery significantly reduced cerebralNADPH and NADH oxidase activities (5.30�0.27 versus7.16�0.46, and 5.59�0.76 versus 12.75�1.18 RLU/mg pro-tein per minute, respectively; n�9, P�0.01) and cerebralsuperoxide production (1.94�0.50 versus 7.24�0.91nmol/mg protein per minute; n�9, P�0.05) in the brainextracts compared with those of the Ad.CMV-Luc group(Figure 5B through 5D).
Figure 2. Expression and localization ofhuman tissue kallikrein in inflammatorycells of ischemic rat brain 9 days aftergene transfer. A, Immunostaining ofhuman tissue kallikrein. Bar�200 �m. B,Double immunofluorescence analyses ofhuman tissue kallikrein (green), ED1(monocytes/macrophages, red), and themerged image (yellow). Bar�50 �m.Overlay image indicated that recombi-nant human tissue kallikrein wasexpressed and localized in some inflam-matory cells.
Figure 3. Effects of kallikrein gene trans-fer on glial cell survival in the ischemiccore and cerebral apoptosis. A, Repre-sentative images of GFAP immunostain-ing. Existence of glial cells in the ische-mic core was apparent in theAd.CMV-TK rat (indicated by arrows) butnot in the Ad.CMV-Luc rat. Bar�200 �m.B, Cerebral apoptosis detected byTUNEL staining. Bar�50 �m.
4 Hypertension February 2004 Part II
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Effects of Kallikrein Gene Transfer on Akt–Bcl-2Signaling, Cleaved Caspase-3 Levels, andCaspase-3 ActivityFigure 6A through 6C shows Western blot analyses (upperpanel) and quantitative analyses (lower panel). Kallikreingene delivery in rats with cerebral I/R significantly increasedphosphorylation of Akt but not total Akt compared with thevalues in rats that had been injected with the luciferase gene(0.82�0.14 versus 0.34�0.09; n�3, P�0.05; Figure 6A).Kallikrein gene delivery also markedly increased Bcl-2 levelsin the ischemic brains of rats compared with those that hadreceived the luciferase gene (1.21�0.11 versus 0.72�0.05
arbitrary units; n�3, P�0.05; Figure 6B) but reduced cleavedcaspase-3 protein levels (433.3�56.3 versus 1041�186.7arbitrary units; n�3, P�0.001) (Figure 6C) and activity(15.9�3.7 versus 27.2�5.4 pmol/mg per minute; n�4 or 5,P�0.05) (Figure 6D) compared with those in rats injectedwith the luciferase gene.
DiscussionThis is the first study to demonstrate that tissue kallikreinprotects against neurologic deficit and cerebral infarctioninduced by ischemic stroke. The protective effects of kal-likrein gene transfer were correlated with the enhancement of
Figure 4. Effect of kinin on migration andapoptosis of primary cultured glial cells.Glial cell migration was analyzed in mod-ified Boyden chambers. A, Migrationassay. Migrating cells were counted in10 fields under 400 � magnification andexpressed as the percentage of migrat-ing cells to those in the presence of 10%serum. B, Representative TUNEL-positive glial cells. Kinin (1 �mol/L) andicatibant (10 �mol/L) were used. C,Quantitative analysis presented as a per-centage of TUNEL-positive nuclei.
Xia et al Kallikrein Gene Transfer in Ischemic Stroke 5
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

glial cell survival and migration and attenuation of I/R-induced cerebral apoptosis and oxidative stress. The protec-tive effects were not mediated by the blood pressure–lowering effect of tissue kallikrein, because the bloodpressures of these rats after gene transfer remained normal(data not shown). A previous study showed that neuronaldamage induced to the MCA becomes apparent at day 1 andreached maximal levels from 2 to 7 days after reperfusion.25
After intracerebroventricular kallikrein delivery, expressionand localization of recombinant human tissue kallikrein couldbe detected in the ischemic rat brain 9 days after genetransfer. In addition, our previous results showed expressionof human tissue kallikrein in various rat brain regions at 1 to7 days after intracerebroventricular delivery of the kallikreingene.26 These observations taken together suggest that ex-pression of recombinant kallikrein in the ischemic brain couldrescue the ischemic penumbra and subsequently reduce I/R-induced infarction after gene transfer. Most important, weobserved that after occlusion of the MCA, I/R-inducedlocomotor deficit scores were significantly ameliorated in ratsthat had received the kallikrein gene transfer. It has beendocumented that spontaneous partial recovery of motorand/or cognitive dysfunction in stroke patients often occurs,but the factors that affect such functional improvement havenot been well elucidated.27 In this study, we have shown thatkallikrein gene transfer promotes a near-total recovery fromstroke-induced locomotor disability.
It is well documented that ROS are generated and play adetrimental role in cerebral I/R injury.2 Induction of oxidative
stress during reperfusion exacerbates cerebral ischemia–in-duced neuronal apoptosis.3,4 Our results show that expressionof kallikrein in the ischemic brain resulted in markedlyincreased cerebral NOx levels in conjunction with reducedNAD(P)H oxidase activity and superoxide production. NO isknown to be a potent antioxidant, because it inhibits NADPHoxidase activity and thus, ROS formation in neutrophils.28
Moreover, a recent study showed that NO plays a vital role inneuroprotection after MCA occlusion,12 indicating that up-regulation of eNOS could protect against cerebral ischemia.Production of NO in the ischemic brain was capable ofreducing generation of ROS, such as superoxide, by inhibit-ing NAD(P)H oxidase activity, and it subsequently facilitatedthe protective effects on ischemia-induced cerebral apoptosis.Furthermore, increased ROS production during reperfusionmight also contribute to the induction of caspase-8, therebyexacerbating apoptosis after focal cerebral ischemia.3 There-fore, these results indicate that kallikrein/kinin, through NOformation, suppresses oxidative stress in ischemic stroke.
Our present study shows that kallikrein gene transfersignificantly reduced I/R-induced cerebral apoptosis, whichwas accompanied by increased phospho-Akt and Bcl-2 levelsbut reduced caspase-3 activation. Akt, a serine/threoninekinase, is believed to promote cell viability in various celltypes. Akt phosphorylation was upregulated in the early stageof MCA occlusion in mice,29 suggesting that prosurvival cellsignaling is initiated in an active fashion before cell deathpathways are activated in the ischemic penumbra. In addition,Bcl-2 was downregulated in neurons after cerebral ische-
Figure 5. Effects of kallikrein gene transfer on (A) cerebral NOx levels, (B) NADPH oxidase activity, (C) NADH oxidase activity, and (D)superoxide formation in ischemic rat brain after I/R injury.
6 Hypertension February 2004 Part II
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

mia.30 Therefore, the antiapoptotic effect of kallikrein genetransfer on cerebral ischemia is most likely mediated byactivation of Akt and increased Bcl-2 levels, which subse-quently resulted in reduced activation of caspase-3 activity.NO has also been shown to inhibit apoptosis by inhibitingcaspase-3 activation.31 Taken together, these findings indicatethat kallikrein gene transfer protected against ischemia-induced stroke mainly through activation of the Akt–Bcl-2signaling pathway, which could lead to increased eNOSactivity and thus, cerebral NO formation and neuroprotection.
We observed that kallikrein gene transfer resulted in theappearance of glial cells in the ischemic core. This might beattributed to migration of glial cells from the ischemicpenumbra. In in vitro studies, we showed that kinin was ableto enhance the migration of primary cultured glial cells and toinhibit glial cell apoptosis induced by H/R in a dose-
dependent manner. These effects of kinin on cell migrationand apoptosis were blocked by icatibant, indicating a brady-kinin B2 receptor–mediated event. These findings reinforcethe notion that enhancement of glial cell survival and migra-tion in the ischemic penumbra and core through activation ofbradykinin B2 receptors is crucial in neuroprotection againstcerebral ischemia.14 However, a recent study showed thatearly treatment with a bradykinin B2 receptor antagonist (0.25and 6.25 hours after the onset of ischemia) attenuated thedamage of transient cerebral ischemia by inhibiting edemaformation.32 This result indicated that kinin might have adetrimental effect in the early stage of ischemic onset. Incontrast, we have shown that kallikrein gene transfer pro-tected against neurologic deficits and cerebral infarction afterfocal cerebral injury. One explanation is that the expressionof recombinant tissue kallikrein was delayed until 3 to 5 days
Figure 6. Effects of kallikrein gene transfer on Akt–Bcl-2 cell survival signaling pathway. Upper panels, Western blots. Lower panels,quantitative analysis. A, Phospho-Akt and total Akt. B, Bcl-2. C, Cleaved caspase-3 levels in ischemic brain. D, Caspase-3 activity inrat brain after ischemic stroke.
Xia et al Kallikrein Gene Transfer in Ischemic Stroke 7
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

after local gene transfer to reach a high level. Moreover, theobserved protective effect of kallikrein on cerebral infarctionand apoptosis was examined 9 days after the onset ofischemia. Taken together, these results suggest that kal-likrein/kinin is beneficial during the late stage of ischemicstroke by promoting glial cell survival and migration andinhibiting apoptosis.
PerspectivesThe present study demonstrates that kallikrein/kinin protectsagainst ischemia-induced neurologic dysfunction and cere-bral infarction by enhancing glial cell survival and migrationand inhibiting apoptosis. These effects occurred in conjunc-tion with suppression of oxidative stress and activation of theAkt–Bcl-2 signaling pathway. A continuous supply of kal-likrein/kinin by gene transfer or protein infusion might offera therapeutic target for the treatment of ischemic stroke.
AcknowledgmentsThis work was supported by National Institutes of Healthgrant HL29397.
References1. Lee JM, Zipfel GJ, Choi DW. The changing landscape of ischemic brain
injury mechanisms. Nature (London). 1999;399:A7–A14.2. Love S. Oxidative stress in brain ischemia. Brain Pathol. 1999;9:
119–131.3. Morita-Fujimura Y, Fujimura M, Yoshimoto T, Chan PH. Superoxide
during reperfusion contributes to caspase-8 expression and apoptosis aftertransient focal stroke. Stroke. 2001;32:2356–2361.
4. Linnik MD, Zobrist RH, Hatfield MD. Evidence supporting a role forprogrammed cell death in focal cerebral ischemia in rats. Stroke. 1993;24:2002–2009.
5. Clements JA. The glandular kallikrein family of enzymes: tissue-specificexpression and hormonal regulation. Endocr Rev. 1989;10:393–419.
6. Erdos B, Miller AW, Busija DW. Impaired endothelium-mediatedrelaxation in isolated cerebral arteries from insulin-resistant rats. Am JPhysiol Heart Circ Physiol. 2002;282:H2060–H2065.
7. Emanueli C, Minasi A, Zacheo A, Chao J, Chao L, Salis MB, Straino S,Tozzi MG, Smith R, Gaspa L, Bianchini G, Stillo F, Capogrossi MC,Madeddu P. Local delivery of human tissue kallikrein gene acceleratesspontaneous angiogenesis in mouse model of hindlimb ischemia. Circu-lation. 2001;103:125–132.
8. Emanueli C, Salis MB, Stacca T, Gaspa L, Chao J, Chao L, Piana A,Madeddu P. Rescue of impaired angiogenesis in spontaneously hyper-tensive rats by intramuscular human tissue kallikrein gene transfer.Hypertension. 2001;38:136–141.
9. Yoshida H, Zhang JJ, Chao L, Chao J. Kallikrein gene delivery attenuatesmyocardial infarction and apoptosis after myocardial ischemia and reper-fusion. Hypertension. 2000;35:25–31.
10. Agata J, Chao L, Chao J. Kallikrein gene delivery improves cardiacreserve and attenuates remodeling after myocardial infarction. Hyper-tension. 2002;40:653–659.
11. Samdani AF, Dawson TM, Dawson VL. Nitric oxide synthase in modelsof focal ischemia. Stroke. 1997;28:1283–1288.
12. Amin-Hanjani S, Stagliano NE, Yamada M, Huang PL, Liao JK,Moskowitz MA. Mevastatin, an HMG-CoA reductase inhibitor, reducesstroke damage and upregulates endothelial nitric oxide synthase in mice.Stroke. 2001;32:980–986.
13. Zhang JJ, Chao L, Chao J. Adenovirus-mediated kallikrein gene deliveryreduces aortic thickening and stroke-induced death rate in Dahl salt-sensitive rats. Stroke. 1999;30:1925–1931.
14. Borlongan CV, Yamamoto M, Takei N, Kumazaki M, Ungsuparkorn C,Hida H, Sanberg PR, Nishino H. Glial cell survival is enhanced duringmelatonin-induced neuroprotection against cerebral ischemia. FASEB J.2000;14:1307–1317.
15. Hara H, Fink K, Endres M, Friedlander RM, Gagliardini V, Yuan J,Moskowitz MA. Attenuation of transient focal cerebral ischemic injury intransgenic mice expressing a mutant ICE inhibitory protein. J CerebBlood Flow Metab. 1997;17:370–375.
16. Lin TN, He YY, Wu G, Khan M, Hsu CY. Effect of brain edema oninfarct volume in a focal cerebral ischemia model in rats. Stroke. 1993;24:117–121.
17. Chao J, Chao L, Tillman DM, Woodley CM, Margolius HS. Character-ization of monoclonal and polyclonal antibodies to human tissue kal-likrein. Hypertension. 1985;7:931–937.
18. Phanithi PB, Yoshida Y, Santana A, Su M, Kawamura S, Yasui N. Mildhypothermia mitigates post-ischemic neuronal death following focalcerebral ischemia in rat brain: immunohistochemical study of Fas,caspase-3 and TUNEL. Neuropathology. 2000;20:273–282.
19. Chao J, Jin L, Chen LM, Chen V, Chao L. Systemic and portal veindelivery of human kallikrein gene reduces blood pressure in hypertensiverats. Hum Gene Ther. 1996;7:901–911.
20. Martin S, Vincent J-P, Mazella J. Involvement of the neurotensinreceptor-3 in the neurotensin-induced migration of human microglia.J Neurosci. 2003;23:1198–1205.
21. Khurana P, Ashraf QM, Mishra OP, Delivoria-Papadopoulos M. Effect ofhypoxia on caspase-3, -8, and -9 activity and expression in the cerebralcortex of newborn piglets. Neurochem Res. 2002;27:931–938.
22. Misko TP, Schilling RJ, Salvemini D, Moore WM, Currie MG. A flu-orometric assay for the measurement of nitrite in biological samples. AnalBiochem. 1993;214:11–16.
23. Griending KK, Minieri CA, Ollerenshaw JD, Wayne Alexander R. An-giotensin stimulates NADH and NADPH activity in cultured vascularsmooth muscle cells. Circ Res. 1994;74:1141–1148.
24. Cavanagh EMV, Fraga CG, Ferder L, Felipe I. Enalapril and captoprilenhance antioxidant defenses in mouse tissues. Am J Physiol. 1997;272:R514–R518.
25. Garcia JH, Wagner S, Liu K-F, Hu X-J. Neurological deficit and extentof neuronal necrosis attributable to middle cerebral artery occlusion inrats. Stroke. 1995;26:627–635.
26. Wang C, Chao C, Madeddu P, Chao L, Chao J. Central delivery of humantissue kallikrein gene reduces blood pressure in hypertensive rats.Biochem Biophys Res Commun. 1998;244:449–454.
27. Borlongan CV. Motor activity-mediated partial recovery in ischemic rats.NeuroReport. 2000;11:4063–4067.
28. Clancy RM, Leszczynska-Piziak J, Abramson SB. Nitric oxide, an endo-thelial cell relaxation factor, inhibits neutrophil superoxide anion produc-tion via a direct action on the NADPH oxidase. J Clin Invest. 1992;90:1116–1121.
29. Shibata M, Yamawaki T, Sasaki T, Hattori H, Hamada J, Fukuuchi Y,Okano H, Miura M. Upregulation of Akt phosphorylation at the earlystage of middle cerebral artery occlusion in mice. Brain Res. 2002;942:1–10.
30. Krajewski S, Mai JK, Krajewska M, Sikorska M, Mossakowski MJ, ReedJC. Upregulation of bax protein levels in neurons following cerebralischemia. J Neurosci. 1995;15:6364–6376.
31. Weiland U, Haendeler J, Ihling C, Albus U, Scholz W, Ruetten H, ZeiherAM, Dimmeler S. Inhibition of endogenous nitric oxide synthase poten-tiates ischemia-reperfusion-induced myocardial apoptosis via a caspase-3dependent pathway. Cardiovasc Res. 2000;45:671–678.
32. Ding-Zhou L, Margaill I, Palmier B, Pruneau D, Plotkine M, Marchand-Verrecchia C. LF 16–0687 Ms, a bradykinin B2 receptor antagonist,reduces ischemic brain injury in a murine model of transient focalcerebral ischemia. Br J Pharmacol. 2003;139:1539–1547.
8 Hypertension February 2004 Part II
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Chun-Fang Xia, Hang Yin, Cesar V. Borlongan, Lee Chao and Julie ChaoMigration and Inhibiting Apoptosis
Kallikrein Gene Transfer Protects Against Ischemic Stroke by Promoting Glial Cell
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 2003 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension published online December 29, 2003;Hypertension.
http://hyper.ahajournals.org/content/early/2003/12/29/01.HYP.0000110905.29389.e5.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertensionin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 15, 2018http://hyper.ahajournals.org/
Dow
nloaded from