just how many species of spirula are there ... - pascal neige · pascal neige*1 2and kerstin warnke...
TRANSCRIPT

Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. (eds.) 2010. Cephalopods - Present and PastTokai University Press, Tokyo, p. 77-84.
Introduction
Spirula, with its well-developed internal chambered shell, is one of the most unusual Recent cephalopods. Empty shells of Spirula can be found in great numbers on oceanic beaches in the Atlantic, Indian and West Pacific Oceans (Bruun 1943, Clarke 1966). As with many other epi- and mesopelagic circumtropical spe-cies, Spirula’s distribution is patchy, apparently there is no gene flow around South Africa between the Atlantic and Indo-Pacific parts of its range (Nesis, 1998). This observation challenges the classical taxonomic opinion that only one biological Spirula species exists. Cryptic species could be relatively easily detected by molecular analysis but, unfortunately, well-preserved material of whole Spirula is scarce, and most collections of Spirula were preserved in formol; and so it is quite rare to find ethanol fixed tissue on which DNA molecular analyses can be conducted. A preliminary study of four Spirula specimens caught near Fuerteventura (Canary Islands, Spain) and sequences (from the gene bank) of one Spir-ula from New Caledonia were examined for indications about the diversity of the geographically fragmented populations (Warnke, 2007). Fragments of mitochon-drial genes, 16S rRNA (16S) and cytochrome oxidase subunit III (COIII) were analysed. The 16S sequences proved to be nearly identical (99.8–100%) whereas the nucleotide sequences of COIII revealed moderate di-
vergence (0.7–1.3%) between the sequences of Spirula from Fuerteventura and New Caledonia. The translated COIII amino acid sequences displayed substantial di-vergence (3.9–4.6%), but this result was supported by single sequence from a single Spirula from New Cale-donia, so the marked sequence divergence may be an ar-tefact. In short, the analysis provided no clear answer to whether there are multiple Spirula species. The present study proposes further evidence on this issue based on shell shape and growth. These morphometric analyses are computed using intrapopulational (i.e. geographi-cal), interpopulational and ontogenetic investigations.
Taxonomic framework
Since Linnaeus (1758) first defined the species he termed Nautilus spirula, taxonomic history has re-corded a rise in the number of reported Spirula species (see Young and Sweeney, 2007 for a review) before the eventual return to just the one Spirula spirula (Linnaeus, 1758). The genus Spirula was defined by Lamarck (1799). It is now generally considered that this genus is monospecific. Some have suggested the occurrence of geographical subspecies (Bruun 1943), one in the Atlan-tic and the other in the Indo-West Pacific (Nesis, 1998), but evidence for this is still lacking.
Just how many species of Spirula are there? A morphometric approach
PASCAL NEIGE*1 AND KERSTIN WARNKE2
1Laboratoire Biogeosciences, Université de Bourgogne, CNRS, 6 Boulevard Gabriel, F-21000 Dijon, France (*Corresponding author; e-mail:[email protected])2Freie Universität Berlin, FR Paläontologie, Malteserstr. 74-100, Haus D, 12249 Berlin, Germany (e-mail: [email protected])
Received May 7, 2008; Revised manuscript accepted November 7, 2008
Abstract. We compare 110 Spirula shells from five geographical areas. Morphometry provides a criterion for determining when shell growth ends (decrease in whorl height). Characteristics of adult shells apparently vary with geographical origin: specimens from Madagascar, New Zealand and Brazil are larger than those from North-West Africa and Australia. These findings challenge the monospecific status of the genus Spirula but fall short of proving the occurrence of more than one species. Supplementary molecular investigations are called for.
Key words: Spirula, morphometry, ontogeny

Pascal Neige and Kerstin Warnke78
Figure 1. Geographical distribution of Spirula spirula after Okutani (1995) (dark grey) and Reid (2005) (light grey), and location of sampling areas (each sampling area is indicated on the map by its code and the number of specimens measured). Question marks indicate imprecise locations. Stars indicate specimens reported by Haimovici et al. (2007). Five main geographical units are defined: NWAF (North-West Africa), BRA (Brazil), MADA (Madagascar), AUS (West Australia) and NZEL (New Zealand). See Table 1 for further abbreviations.
Main geographical areas Label n
NWAF West Morocco (coll. Warnke) Mo 7Fuerteventura (coll. Warnke) Fu 14Playa del Ingles (coll. Warnke) Ca 6
BRA Caponga Beach (coll. J. Geraldo de Aratanha) BRA-F 3Canoa Quebrada (coll. M.A. Reis Jr.) BRA-A 10
MADA Port Ostafrika (coll. Museum für Naturkunde, Berlin) PoO 8Madagascar NW (coll. S. Kiel) MaNW2 10Manakara (coll. J. Hartmann) MKSE 22Madagascar SE (coll. J. Hartmann) MaSE 9
AUS South West Australia (coll. K. Bandel) KB 3
NZEL Ninety Mile Beach (coll. F. Riedel) NMB 9New Zealand (coll. F. Riedel) NZ 9
Table 1. Location and number of specimens used in this study (see also Fig. 1). coll. = collection. n = number of measured speci-mens for a given locality.

How many species of Spirula? 79
Material and methods
A set of 110 shells was measured in this study. All were collected from beaches. The specimens measured were the best preserved ones from a larger sample. They come from various areas covering a broad span of the known geographical distribution of Spirula. Five main geographical areas are defined here based on sampling localities (see Figure 1 and Table 1): North-West Africa (NWAF), with specimens from the Canary Islands and from the coast of Morocco; Brazil (BRA), all specimens from two localities close to Fortaleza; the Madagascar area (MADA), with specimens from the east and west coasts of Madagascar and from Mozambique; South-West Australia (AUS), with specimens collected not far from Perth; and the North Island of New Zealand (NZEL). Because this analysis is based on shells col-lected from beaches, we cannot exclude the effect of post-mortem drift. This may be the case particularly
for BRA shells, located outside the known geographi-cal distribution of Spirula. The results will thus be dis-cussed (see “Discussion”) on the assumption that ana-lyzed samples might represent nearby populations.
A new measurement protocol was defined for de-ciphering shell growth as accurately as possible. The anterior boundaries of ventral and dorsal sutures (sensu Naef, 1923) were used as landmarks (Figure 2A). Their coordinates being known throughout growth (i.e., mea-sured for each septa), basic trigonometric equations were used to calculate several parameters with which to describe the shell (Figure 2B).
A particularly complicated issue is to define the cen-tre of the supposed logarithmic spiral of Spirula shell growth. Several workers have addressed this issue (e.g., Landman 1987, Neige 1997 for ammonites). Schindel (1990) attempted to locate such a starting point for growth (the apex) when studying gastropod ontogeny. He concluded that radius measurements included an oscillating error term arising from mislocation of the
Figure 2. Shell measurements. A. Location of P1 (0;0) and details (left) showing the locations of the successive landmarks. B. Cal-culated parameters (from landmark coordinates, see main text) used to study shell growth: r is radius, H is whorl height, and α is the angle between two successive septa.

Pascal Neige and Kerstin Warnke80
Figure 4. Comparative shell growth for the five main geographical areas (see Table 1 for abbreviations).
Figure 3. Example of individual growth (case of specimen NZ-02). A. radius versus whorl height. This pattern shows a decrease in whorl height at the end of growth (see main text). B. radius versus α.

How many species of Spirula? 81
apex (see also Neige 1997 for similar considerations on ammonites). Certain mathematical calculations might help in defining a theoretical initial growth point of the spiral but would necessarily assume a perfect loga-rithmic growth throughout ontogeny. Such mathemati-cal extrapolation might mask ontogenetic changes in growth of the shell. Because the present study explores ontogeny, we prefer to locate the initial growth point at an anatomical landmark: the dorsal attachment of septa number 1. By convention, P1 coordinates (dorsal attach-ment of septa 1, Figure 2A) are x = 0 and y = 0. This excludes any artificial correction of growth parameters, but produces more prominent artificial oscillations dur-ing early growth stages.
The anterior boundaries of ventral and dorsal sutures were located using a microscope with transmitted light. Measurements were made with an optic measuroscope (Nikon Measuring Microscope MM-60; precision of 1 μm). Measurement error was estimated by measuring one specimen nine times with complete repositioning and measurement. Error was very low compared with differences between specimens from a given locality.
A Kruskal-Wallis test was used to test for differences in the adult-stage parameters: radius, whorl height, number of septa and rotational angle. The Kruskal-Wal-lis test can be applied as a single factor non-parametric ANOVA case. It is useful for situations where the usual ANOVA normality assumptions may not apply, which is the case here because of small sample sizes for Austra-lia (n = 3) and Brazil (n = 7). This test indicates whether observed differences between means of a given param-eter and for several populations are statistically signifi-cantly different. H0 is that observed differences are due to a sampling effect. H1 is that at least two populations have statistically different means.
Results
Two complementary analyses were conducted: one focusing on individual growth; the other comparing the various growth parameters for the five main geographi-cal areas defined here.
GrowthThe growth parameters of each of the 110 specimens
were investigated. Because the anterior boundaries of suture attachments were measured for all the septa of each specimen, growth could be described very pre-cisely, particularly in terms of whorl height versus shell radius.
One pattern is largely dominant (68 specimens; i.e., 64.5% of the complete sample). This pattern involves
a characteristic decrease in whorl height at the end of growth (Figure 3A). Only in three specimens (i.e., 2.7% of the complete sample) is this decrease followed by a fresh increase in shell height, and even then it is a tiny increase limited to only one or two septa. 39 specimens display no decrease in whorl height during growth. These are interpreted as juvenile or broken shells. The decrease in whorl height may be considered as an indi-cation of the imminent end of growth in Spirula. It is considered here as a homologous feature, indicative of a homologous stage of growth. For convenience, this stage (onset of the end of growth) is termed the ‘adult stage’ in what follows.
Angle variations between two successive septa are similar between specimens. Specimen NZ-02 (Figure 3B) is used as an example. A first phase displays high values (nearly 55°). These large initial angles can be explained by the bulbous shape of the first chamber. Thereafter the angle decreases in the course of growth to about 20°. A further small decrease in septal angle occurs at the very end of growth.
Population approachA population analysis is made on adult shells only
(i.e., those meeting the criterion for the end of growth defined above) to explore geographical differences in shell characteristics. The five main geographical areas are used for that purpose (see Figure 1).
Shells from the same areas display similar patterns of growth in terms of whorl height increase (Figure 4). Moreover, the growth of specimens is conservative at the complete sample scale. The main difference between specimens from the five main geographical areas occurs at the end of growth. On average (see Figure 4), shells from Madagascar attain the greatest whorl heights at the end of growth. Shells from New Zealand and from Brazil display intermediate whorl heights; those from
Area Mean Min Max n
AUS 27 25 29 3
BRA 28.7 25 31 7
MADA 30.5 27 34 28
NWAF 27 24 30 17
NZEL 29 26 34 13
Table 2. Mean, minimum and maximum of septa at adult stage (see main text for end of growth criterion) for the five main geographical areas (NWAF (North-West Africa), BRA (Brazil), MADA (Madagas-car), AUS (West Australia) and NZEL (New Zealand). n = number of adult shells.
Pascal Neige and Kerstin Warnke82
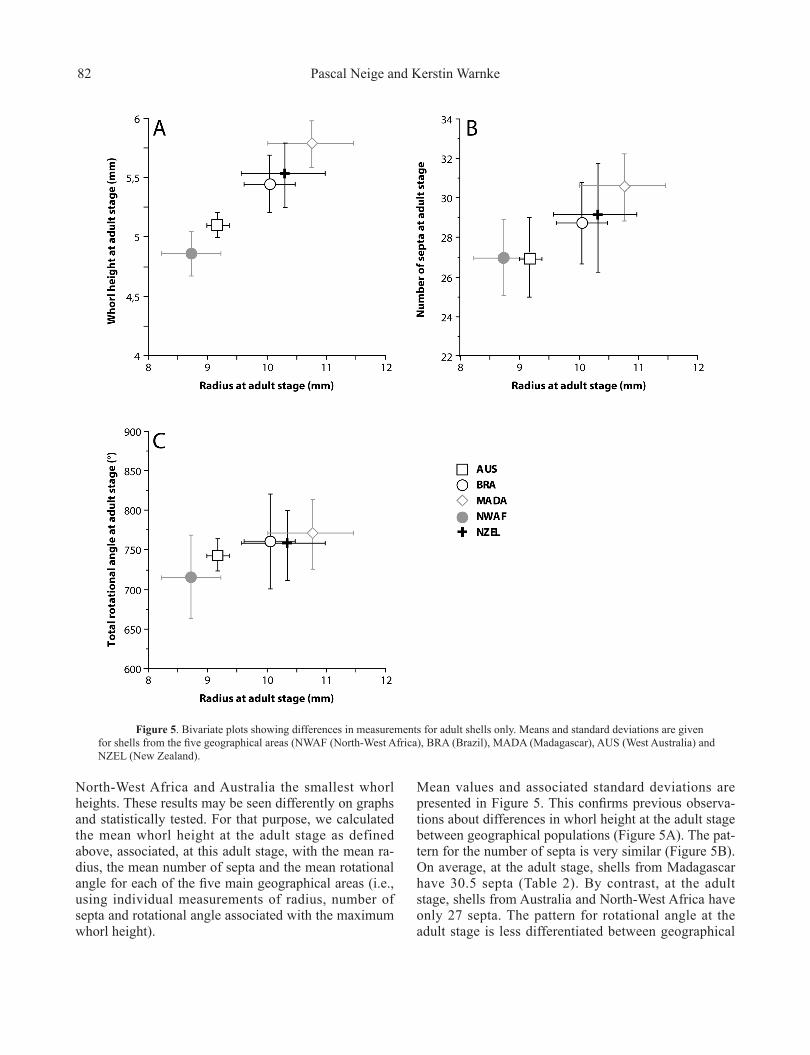
North-West Africa and Australia the smallest whorl heights. These results may be seen differently on graphs and statistically tested. For that purpose, we calculated the mean whorl height at the adult stage as defined above, associated, at this adult stage, with the mean ra-dius, the mean number of septa and the mean rotational angle for each of the five main geographical areas (i.e., using individual measurements of radius, number of septa and rotational angle associated with the maximum whorl height).
Mean values and associated standard deviations are presented in Figure 5. This confirms previous observa-tions about differences in whorl height at the adult stage between geographical populations (Figure 5A). The pat-tern for the number of septa is very similar (Figure 5B). On average, at the adult stage, shells from Madagascar have 30.5 septa (Table 2). By contrast, at the adult stage, shells from Australia and North-West Africa have only 27 septa. The pattern for rotational angle at the adult stage is less differentiated between geographical
Figure 5. Bivariate plots showing differences in measurements for adult shells only. Means and standard deviations are given for shells from the five geographical areas (NWAF (North-West Africa), BRA (Brazil), MADA (Madagascar), AUS (West Australia) and NZEL (New Zealand).

How many species of Spirula? 83
areas (Figure 5C). Table 3 shows the Kruskal-Wallis test results for these observations. All comparisons reveal statistical differences among populations, although the difference for rotational angle is less marked (p = 0.023).
Discussion
This study uses a morphometric exploration of shell growth to propose pointers for investigating species structuring within Spirula. This should be seen as a first step towards a comparative analysis using both morpho-metric and molecular approaches. However, a number of findings are worth discussing:
1. We propose a criterion for detecting the end of shell growth based on the recognition of a decrease in whorl height. This ontogenetic pattern is common in the populations under study (64.5%). It would be interesting to compare this criterion with others based on the soft parts of the animal. This criterion has made it possible to compare shells from different geographical popula-tions at an equivalent (adult) growth stage.
2. The comparisons revealed differences in measure-ments between populations at the adult stage. We sta-tistically tested differences in values of radius, whorl height, number of septa and rotational angle. Differenc-es between populations may be relatively large: for ex-ample the mean number of septa at the adult stage is 27 for North-West African specimens versus 30.5 for shells from Madagascar. Similarly, whorl height is nearly 5.78 mm for shells from Madagascar versus nearly 4.86 mm for shells from North-West Africa, a difference of nearly 20%. Schematically, two separate clusters can be recog-nized: specimens from Madagascar, New Zealand and Brazil, with larger dimensions at the adult stage, and specimens from North-West Africa and Australia. It is difficult to explain this clustering in terms of geography.
It should be remembered that the analyzed popula-tions contain shells that might have been subjected to post-mortem drift. Post-mortem drift is a common phenomenon within recent shelled cephalopods and has lead to misinterpretation of geographical ranges of some of them. For example, cuttlefishes have been er-
roneously supposed to occur in the Western Atlantic Ocean because of cuttlebone post-mortem drift (see Voss, 1974). Analyzes of shell preservation and epizoan occurrences on Spirula shells collected from the beach by Dauphin (1979a, b) were inconclusive regarding the exact geographical origin of the living population, and thus the possibility of intense geographical drift. As a result, she favoured a distant origin because of the lack of any living specimen caught in the area. Similarly, shells used here for BRA locality are not included in the known geographical distribution of Spirula spirula (see Figure 1). However, Haimovici et al. (2007) recently reported S. spirula along Brazilian coasts (Bahia state) southward of the beaches studied here. Only few com-plete specimens have been found (e.g., not only shells but nor a living population). Thus, the Brazilian shells used in the present study may have been drifted, but for a short distance. In addition, the relatively homogenous distributions of morphological features within popula-tions compared to statistical differences of morphologi-cal features among populations suggest that shell popu-lations reflect natural living populations. More work is needed to clearly determine whether Spirula popula-tions live along the Brazilian coasts and thus to identify the exact extent of geographical drift.
The results obtained here clearly challenge the tradi-tional monospecific status of Spirula. However, they are only preliminary findings and do not demonstrate the existence of two or more species. A molecular analy-sis is now needed. It would be interesting also to use a similar morphometric approach but based on sexed specimens to control for any sexual dimorphism. An alternative hypothesis to the occurrence of two or more species to account for the observed morphometric dif-ferences is ecophenotypy. This could also be tested by a molecular approach. In practice, this will prove difficult because complete Spirula specimens are seldom caught. Although limited in geographical scope compared to the present study, an analogous analysis based on Nautilus may help interpreting the present results (Tanabe et al. 1990). In spite of their non-overlapping distributions (live-caught specimens were coming from the Philip-
H p-valuesRadius at adult stage, Geographical areas 43.6 <0.0001 ***Whorl height at adult stage, Geographical areas 49.7 <0.0001 ***Number of septa at adult stage, Geographical areas 25.9 <0.0001 ***Rotational angle at adult stage, Geographical areas 11.3 0.023 *
Table 3. Non parametric Kruskal-Wallis test for various data sets (see main text). H is the Kruskal-Wallis test statistic. Results are given for ex-aequo correction. *: test significant at 95% confidence level; ***: test significant at 99.9% confidence level.

Pascal Neige and Kerstin Warnke84
pines, Fiji and Palau), Nautilus from the three popula-tions shared quite similar overall shell morphology but could be distinguished by adult features such as the dimensions of the shell and total number of septa. Using such results and some genetic data (from Woodruff et al. 1987), the authors concluded that the three populations belong to the same wide-ranging species. Based on ge-netic and morphological exploration of Nautilus coming from different geographic areas, Wray et al. (1995) con-cluded similarly that some of the morphological features used to define Nautilus species may represent variation within one widespread species. Applied to the present analysis, this would mean that observed morphological differences between populations could represent varia-tion within a single species. However, it is worth noting that the geographic variation described here for Spirula is much more extensive than that for Nautilus. Finally, new genetic and morphological data are needed before to determine Spirula species number.
Acknowledgments
We thank M. Aeppler, J. Geraldo de Aratanha, K. Bandel, J. Hartmann, S. Kiel, F. Riedel, M.A. Reis Jr, Ignacio Santana and the Museum für Naturkunde (Berlin) for kindly providing Spirula shells. Thanks to Emmanuelle Pucéat for her help in computing trigono-metric equations to transform landmark coordinates into distances and angle measurements. We are grateful to participants at the 7th International Symposium, Cepha-lopods – Present and Past for their discussions during the meeting. We thank Kazushige Tanabe for his helpful comments, and Sylvain Gerber, Melanie J. Hopkins and Michael Labarbera for checking our English. This work was supported by the DFG WA 1454/2-1 grant from the Deutsche Forschungsgemeinschaft (DFG) to Kerstin Warnke. This paper is a contribution to the FED (Forme, Evolution, Diversité) team of UMR CNRS 5561 Bio-géosciences, and to GDR MEF (Morphométrie et Evo-lution des Formes).
References
Bruun, A. F., 1943: The biology of Spirula spirula (L.). Dana Reports, vol. 24, p. 1-44.
Clarke, M. R., 1966: A review of the systematics and ecology of oce-anic squids. Advances in marine Biology, vol. 4, p. 91-300.
Dauphin, Y., 1979a: Contribution à l’étude de la formation des gise-ments de cephalopods. I. Les coquilles de spirules (Dibranchiata) de Nouvelle Calédonie. Cahiers de l’Indo-pacifique, vol. 1, p. 165-194.
Dauphin, Y., 1979b: Contribution à l’étude de la formation des gise-ments de cephalopods. I. Les coquilles de spirules (Dibranchiata,
Cephalopoda) de l’île Maurice (océan indien). Cahiers de l’Indo-pacifique, vol. 1, p. 335-351.
Haimovici, M., Costa, p. A. S., Santos, R. A., Martins, A. S., and Ola-vo, G., 2007: Composição de espécies, distribuição e abundância de cefalópodes do talude da região central do Brasil. In, Costa, p. A. S.; Olavo, G.; Martins, A. S. eds., Biodiversidade da Fauna Marinha Profunda na Costa Central Brasileira. Rio de Janeiro: Museu Nacional (Série Livros n.24), p. 109-132.
Lamarck, J. B. p. A., 1799: Prodrome d’une nouvelle classification des coquilles, comprenant une rédaction appropriée des caractères génériques, et l’établissement d’un grand nombre de genres nou-veaux. Mémoires de la Société d’Histoire Naturelle de Paris, vol. 1, p. 63-91.
Landman, N. H. 1987: Ontogeny of Upper Cretaceous (Turonian-San-tonian) scaphitid ammonites from the western interior of North America: systematics, developmental patterns, and life history. Bulletin of the American Museum of Natural History, vol. 185, p. 117-241.
Linnæus, C. 1758: Systema Naturæ per regna tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differen-tiis, Synonymis, Locis. Tomus I. 824 p. Editio decima, reformata, Holmiæ.
Naef, A. 1923: Die Cephalopoden. Fauna und Flora des Golfes von Neapel. vol. 35, p. 1-863.
Neige, p. 1997: Ontogeny of the Oxfordian ammonite Creniceras renggeri from the Jura of France. Eclogae Geologicae Helvetiae, vol. 90, p. 605-616.
Nesis, K. N., 1998: Biodiversity and systematics in cephalopods: Un-resolved problems require an integrated approach. South African Journal of marine Science, vol. 20, p. 165-173.
Okutani, T. 1995: Cuttlefish and Squids of the World in Color, 185p. National Cooperative Association of Squid Processors, Tokyo.
Reid, A. 2005: Family Spirulidae Owen, 1936. In, Jereb, p. and Roper, C. F. E. eds., Cephalopods of the World. An Annotated and Il-lustrated Catalogue of Species known to Date, vol. 1, p. 211-212. FAO, Rome.
Schindel, D. E. 1990: Unoccupied morphospace and the coiled geom-etry of gastropods: Architectural constraint or geometric covaria-tion? In, Ross, R. A. and Allmon, W. D. eds., Causes of Evolu-tion, p. 270–304. University of Chicago Press, Chicago, IL.
Tanabe, K., Tsukahara, J. and Hayasaka, S., 1990: Comparative mor-phology of living Nautilus (Cephalopoda) from the Philippines, Fiji and Palau. Malacologia, vol. 31, p. 297-312.
Voss, G.L., 1974: On the absence of cuttlefish in the western Atlantic. The Veliger, vol. 16, p. 367-369.
Warnke, K., 2007: On the species status of Spirula spirula (Linné 1758) (Cephalopoda): a new approach based on divergence of amino acid sequences between the Canaries and New Caledonia. In, Landman, N. H., Davis, R. A. and Mapes, R. H. eds., Cepha-lopods Present and Past. New Insights and Fresh Perspectives, p. 144-155. Topics in Geobiology Book series, Kluwer Academic/Plenum Publishers, New York.
Woodruff, D.S., Carpenter, M.P., Saunders, W.B. and Ward, p. D., 1987: Genetic variation and phylogeny in Nautilus. In, Saun-deers, W.B. and Landman, N.H. eds., Nautilus: The Biology and Paleobiology of a Living Fossil, p. 65-83. Plenum, New York.
Wray, C.G., Landman, N.H., Saunders, W.B. and Bonacum, J., 1995: Genetic divergence and geographic diversification in Nautilus. Paleobiology, vol. 21, p. 220-228.
Young, R. E., and Sweeney, M. J., 2002: Taxa Associated with the Family Spirulidae Owen, 1836. Tree of Life, web project, http://tolweb.org/accessory/Spriulidae_Taxa?acc_id=2337