journal of vol ,268, no. issue of june 5, pp. 12207-12212 ... · the journal of biological...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY (0 1993 by The American Society for Biochemistry and Molecular Biology, h e .
Vol ,268, No. Issue of June 5, pp. 12207-12212,1993 Printed in U. S.A.
Purification of a Novel Phospholipase C Isozyme from Bovine Cerebellum*
(Received for publication, September 11, 1992, and in revised form, December 24, 1992)
Do Sib Min, Dong Myung Kim, Young Han Lee, Jeongkon Seo, Pann-Ghill Suh, and Sung Ho RyuS From the Department of Life Science, Pohang Institute of Science and Technology, Hyoja-Dong, Pohnag, Korea 790-600
An isozyme of phosphoinositide-specific phospholi- pase C (PLC) was purified to near homogeneity from bovine cerebellum by a combination of several column chromatography procedures. Approximately 80 pg of pure enzyme were obtained from 4 kg of bovine cere- bellum, with a final specific activity of 7.5 pmol/min/ mg protein in the presence of 0.1% deoxycholate. The enzyme is specific for phosphatidylinositol and phos- phatidylinositol4,5-bisphosphate but does not hydro- lyze phosphatidylcholine. The molecular weight of the enzyme determined by SDS-polyacrylamide gel elec- trophoresis is approximately 97,000. Polyclonal anti- bodies to previously characterized PLC isozymes, PLC- 81, -82, -71, -72, and -61, did not cross-react with the purified cerebellar enzyme. Moreover, polyclonal an- tibodies prepared against the cerebellar enzyme did not react with purified PLC-81, -82, -71, -72, or -61. However, the cerebellar enzyme was recognized by two antibodies generated against peptide sequences com- mon to mammalian PLC isozymes. Comparison of par- tial amino acid sequences of the purified cerebellar enzyme with the deduced amino acid sequence of each known PLC isozyme shows that the cerebellar enzyme is a novel PLC, which could be classified as a PLC-8- type isozyme. Thus, we have designated this enzyme PLC-84.
Phosphoinositide-specific phospholipase C (PLC)’ plays a key role in the signal transduction of many calcium mobilizing agonists by triggering the formation of two second messen- gers, diacylglycerol and inositol 1,4,5-trisphosphate (1-4). A number of distinct PLC isozymes, which vary with respect to molecular weight and enzymatic properties, have been puri- tied from a wide range of mammalian tissues (5-19), and several forms have been molecularly cloned and sequenced (20-26). Comparison of the deduced amino acid sequences of the cloned PLCs suggested that there exist at least three principal types of PLC isozyme (PLC-p, -7, and -6 ) , and each type contains several distinct subtypes (27). In mammalian brain, at least five immunologically different PLC isozymes (PLC-01, -71, -61, -62, and - 6 ) were described (8, 10, 22, 26).
~ ~ ~ ~ ~~~~
* This work was supported in part by grants from Korea Science and Engineering Foundation and from Pohang Institute of Science and Technology. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed: POSTECH, De- partment of Life Science, P.O. Box 125 Ponhang, Korea 790-600. Tel.: 82-562-79-2292; Fax: 82-562-79-2199.
The abbreviations used are: PLC, phosphoinositide-specific phos- pholipase C; DTT, dithiothreitol; HPLC, high performance liquid chromatography; MES, 2-(N-morpholino)ethanesulfonic acid; PI, phosphatidylinositol; PIPZ, phosphatidylinositol 4,5-bisphosphate; PAGE, polyacrylamide gel electrophoresis.
The overall amino acid sequence similarity between the iso- zymes is low. However, a significant sequence similarity is apparent in two regions, which were designated the X- and Y-domains, and have been proposed to constitute a putative catalytic site (14, 28, 29).
The diversity in primary structure together with different cellular expression patterns of PLC isozymes provides the assumption that each type of PLC is activated in response to different receptors signal by distinct mechanisms (14). Many efforts have been concentrated on the elucidation of activation mechanisms for each PLC isozyme. PLC-71 and PLC-72 possess SH2 and SH3 domains, which were first identified in the regulatory domains of the nonreceptor tyrosine kinases (14, 20, 21, 24, 25, 27), and appear to mediate the association with the growth factor receptor tyrosine kinase activity (28- 35). Evidence suggests that G-proteins may couple certain PLC isozymes to cell surface receptors (36-39). The a subunit of the pertussis toxin-insensitive G-proteins, Gaq and Gall, which are known to be coupled to muscarinic acetylcholine, thromboxane Az, bradykinin, angiotensin, histamine, and va- sopressin receptors (40-42), have been shown to activate PLC-pl(43-47). Although two additional members of the Gaq class, Gal4 and GaI6, have been reported (48), and cDNAs for PLC-p2 and PLC-p3 have been isolated from promyelocytic HL60 cells and WI-38 fibroblast cells, respectively (27), acti- vation of PLC isozymes by GaI4 and Ga16 has not yet been reported.
We now report purification of a PLC isozyme that appears to be distinct from the mammalian isozymes purified so far. From comparison of the partial amino acid sequence of this isozyme with those of the known PLCs we suggest the new enzyme be designated PLC-04.
EXPERIMENTAL PROCEDURES
Materink 3-Ph0sphatidyl[2-~H]inositol (17.9 Ci/mmol) and 3-phosphati-
dyl[2-3H]inositol 4,5-bisphosphate (1.0 Ci/mmol) were purchased from Amersham International, United Kingdom. Wheat germ phos- phatidylinositol, bovine brain phosphatidylinositol 4,5-bisphosphate, and formalin-fixed Staphylococcus aureus cells (Pansorbin) were ob- tained from Calbiochem. Gel matrices for the column chromatographs were purchased as follows: DE52-cellulose from Bodman Chemicals, Aston, PA; Butyl-Toyopearl 650 M from Tosoh, Tokyo; and Spectra- gel Blue from Spectrum, Los Angeles. TSK DEAEdPW, TSK Phenyl-5PW, and TSK Heparin-5PW HPLC columns were obtained from Tosoh, and Mono S and Mono Q columns were from Pharmacia, Sweden. PLC-PI, -71, or -61 were purified from bovine brain as described previously (8). PLC-p2 and -72 were obtained from Dr. Sue Goo Rhee, National Heart, Lung, and Blood Institute (Bethesda, MD). The polyclonal antiserum against each PLC isozyme was pre- pared in New Zealand White rabbits by immunization according to a standard method (49). Antipeptide antibodies generated against the PLC sequence Gly-Cys-Arg-Cys-Ile-Glu-Leu-Asp-Cys-Trp in the X- domain and against the sequence Leu-Ser-Arg-Ile-Tyr-Pro-Lys-Gly in the Y-domain were also obtained from Dr. Rhee.
12207
12208 Phospholipase C Isozyme
Phospholipase C Assay
Enzyme activity was quantitated in the 200-pl assay mixture con- taining 150 pM PI (20,000 cpm [3H]PI), 1 mM EGTA, 10 mM CaC12, 0.1% sodium deoxycholate, 1 mg/ml bovine serum albumin, and 50 mM HEPES, pH 6.8. For the measurement of pH- or Caz+-dependent activities, unilamellar vesicles of [3H]PI or [3H]PIP2 were separately prepared as described elsewhere (50). The reaction mixture was incubated at 37 “C with enzyme and terminated as previously de- scribed (8).
Purification Procedure
Two kg of frozen bovine cerebellum obtained from a local slaughter house were homogenized in a homogenizer (Brinkman, Westbury, NY) with 6 liters of buffer A (20 mM Tris-HC1, pH 7.6, 1 mM EDTA, 1 mM EGTA, and 0.1 mM DTT) containing 0.2 mM phenylmethyl- sulfonyl fluoride. The homogenate was centrifuged at 13,000 X g for 1 h, and the supernatant was adjusted to pH 7.6 with a 1 M Tris solution.
Step 1. DE52-cellulose Anion-exchange Chromatography-The su- pernatant was applied to a DE52-cellulose column (10 X 37 cm) preequilibrated with buffer A, and proteins were eluted with a 6-liter linear gradient from 0 to 1 M KC1 in buffer A. Most activity eluted at 60-140 mM KC1, and the corresponding fractions were pooled for the next step.
Step 2. Butyl-Toyopearl Hydrophobic Chromatography-Two batches of the pooled fractions from step 1 were combined, adjusted to 2 M KC1 by adding solid KCl, and applied to a Butyl-Toyopearl column (10 X 16 cm) preequilibrated with buffer B (20 mM HEPES, pH 7.0, 1 mM EGTA, and 1 mM EDTA) containing 2 M KC1. The column was eluted with 3 liters of decreasing gradient from 2 to 0 M KCl. A broad peak of activity eluted between 1.5 and 0.7 M KC1, and the corresponding fractions were pooled and dialyzed against buffer B.
Step 3. Spectragel Blue Chromatography-The dialyzed sample from step 2 was centrifuged at 13,000 X g for 30 min to remove denatured proteins, and the supernatant was applied to a Spectragel Blue column (5 X 20 cm) preequilibrated with buffer B. The column was eluted with 0.8 liter of a gradient from 0 to 1 M KC1, and the active fractions eluted at 350-500 mM KC1 were pooled. The pooled fractions were concentrated to 20 ml in an Amicon filtration appa- ratus (M, cut off, 30,000) and dialyzed against buffer A.
Step 4. DEAE Anion-exchange HPLC-Each of the two halves of the dialyzed sample from step 3 was loaded separately on a TSK DEAE-5PW column (21.5 X 150 mm) preequilibrated with buffer A at a flow rate of 5 ml/min. Elution was continued at 5 ml/min with a KC1 gradient, from 0 to 0.3 M for 45 min and 0.3 to 1 M for 10 min. The major active peak eluted at 130-170 mM KC1 was pooled and concentrated to 20 ml. The concentrated samples from both column applications were pooled.
Step 5. Phenyl Hydrophobic HPLC-The combined fraction from step 4 was concentrated to 20 ml in an Amicon filtration apparatus (M, cut off, 30,000). Solid KC1 was added into the concentrated sample to 2 M followed by centrifugation at 13,000 X g for 30 min to remove the denatured proteins. The supernatant solution was injected at a flow rate of 5 ml/min into a TSK Phenyl-5PW column (21.5 X 150 mm) preequilibrated with buffer C (20 mM HEPES, pH 7.0, 1 mM EDTA, 1 mM EGTA and 0.1 mM DTT) containing 2 M KC1. Elution was continued at 5 ml/min with a decreasing KC1 gradient from 2 to 0.8 M for 10 min, 0.8 to 0 M for 20 min, and 0 M for 20 min. The active peak eluted between 1.1 and 0.6 M KC1 was concentrated to 10 ml in an Centriprep 30 (Amicon) and dialyzed against buffer C.
Step 6. Heparin HPLC-The dialyzed sample from step 5 was injected at a flow rate of 1 ml/min into a TSK Heparin-5PW column (7.5 X 75 mm) preequilibrated with buffer C. Elution was continued at 1 ml/min with a KC1 gradient from 0 to 0.5 M for 45 min. The active fractions eluted at 90-130 mM KC1 were pooled and washed in an Centriprep 30 with buffer D (20 mM MES, pH 6.0, 1 mM EDTA, 1 mM EDTA, and 0.1 mM DTT).
Step 7. Mono S Cation-exchange Chromatography-The sample from step 6 was injected at a flow rate of 1 ml/min into a Mono S column (5 X 50 mm) preequilibrated with buffer D. Elution was continued at 1 ml/min with a NaCl gradient from 0 to 0.3 M for 20 min. The active fractions eluted between 75 and 100 mM NaCl were collected and dialyzed against buffer A.
Step 8. Mono Q Anion-exchange Chromatography-The protein solution from step 7 was injected at a flow rate of 1 ml/min into a Mono Q column (5 X 50 mm) preequilibrated with buffer A. Elution was continued at 1 ml/min with a KC1 gradient from 0 to 0.4 M for
20 min. Peak fractions eluted at 0.2 M KC1 were collected manually and used for the characterization.
Production of Polyclonal Antiserum
A female BALB/c mouse (7-8 weeks old) was immunized intraper- itoneally with 20 pg of the purified PLC isozyme in complete Freund’s adjuvant three times at 2-week intervals. Three days before death, the mouse was further boosted by intraperitoneal injection of 20 pg of the same enzyme in phosphate-buffered saline (10 mM phosphate, pH 7.2, 150 mM NaCl). Blood was collected from the heart of the mouse and the antiserum was obtained by centrifugation at 10,000 X g for 10 min.
Partial Amino Acid Sequencing
Peptides used for the sequence analysis were prepared from the purified enzyme by two different methods. The first method involved was by the partial digestion of the purified enzyme with protease V8 and separation on SDS-polyacrylamide gel. Purified PLC (150 pg) was subjected to SDS-PAGE on an 8% polyacrylamide gel and a 97- kDa band was excised. Limited proteolysis with protease V8 and separation of the digested peptides on a 15% SDS-polyacrylamide gel were performed as described elsewhere (53). Electrophoretic transfer of the resolved peptides onto a polyvinylidene fluoride membrane for the sequence analysis were performed as previously described (54). In the second method, the peptides were prepared by tryptic digestion and purified by HPLC. Purified PLC (100 pg) in 0.2 ml of 20 mM Tris, pH 7.6, 1 mM EGTA, 1 mM EDTA, 2.5 mM DTT, and 0.2 M KC1 was denatured by heating for 5 min at 95 “C and was dispersed by brief sonication in a bath-type sonicator. The solution was adjusted
20 40ml 20
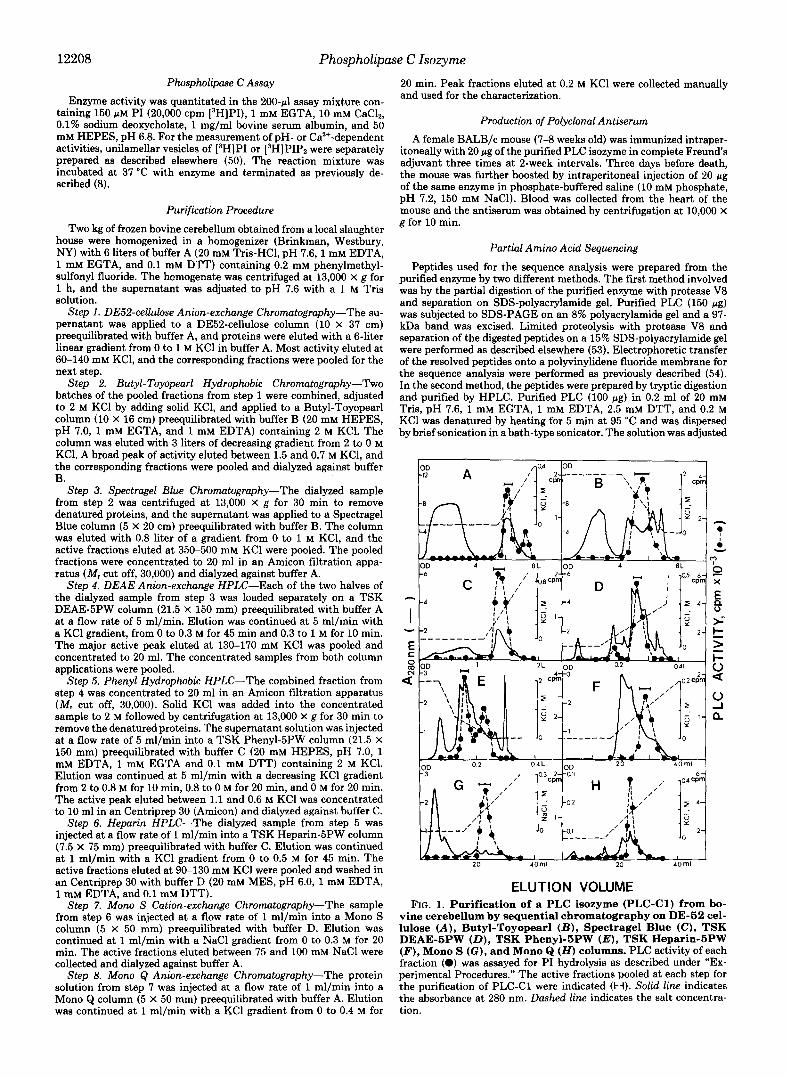
ELUTION VOLUME FIG. 1. Purification of a PLC isozyme (PLC-C1) from bo-
vine cerebellum by sequential chromatography on DE-52 cel- lulose (A), Butyl-Toyopearl ( B ) , Spectragel Blue (C), TSK DEAE-5PW (D) , TSK Phenyl-5PW ( E ) , TSK Heparin-5PW (F), Mono S (G), and Mono Q (H) columns. PLC activity of each fraction (a) was assayed for PI hydrolysis as described under “Ex- perimental Procedures.” The active fractions pooled at each step for the purification of PLC-C1 were indicated (H). Solid line indicates the absorbance at 280 nm. Dashed line indicates the salt concentra- tion.
12209
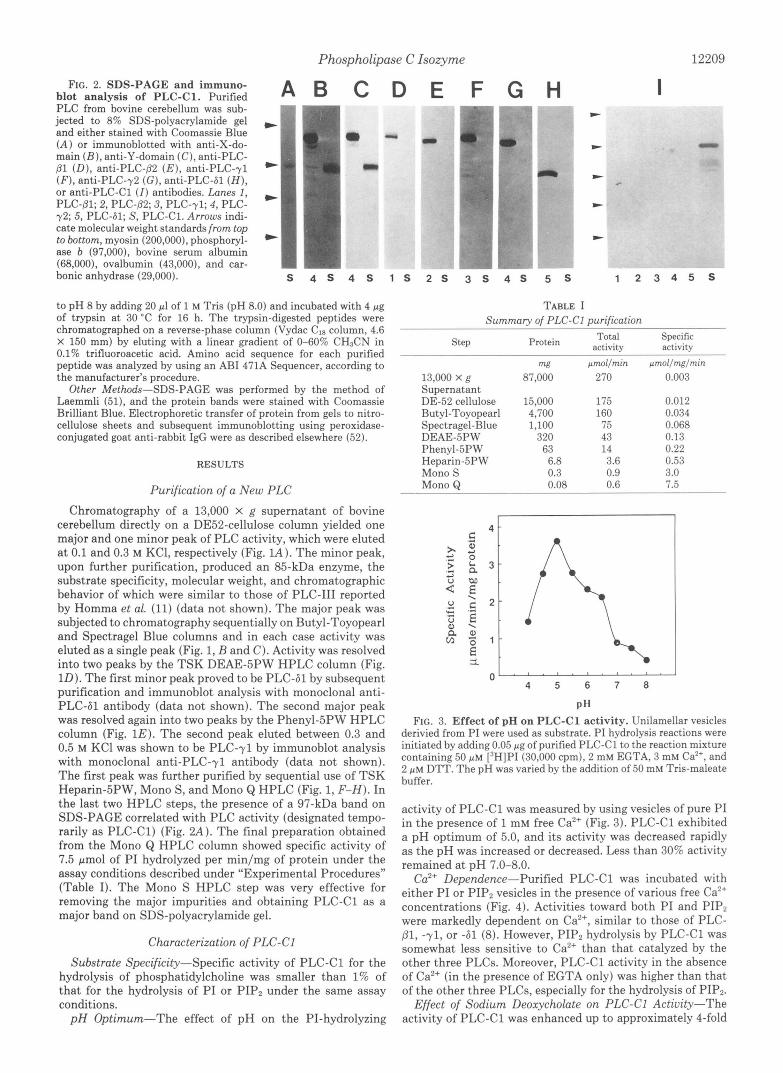
FIG. 2. SDS-PACE and immuno- blot analysis of PLC-Cl. Purified PLC from bovine cerebellum was sub- jected to 8% SDS-polyacrylamide gel and either stained with Coomassie Blue (A) or immunoblotted with anti-X-do- main ( B ) , anti-Y-domain (C), anti-PLC- (31 (D), anti-PLC-PZ ( E ) , anti-PLC-yl (F), anti-PLC-y2 (G), anti-PLC-61 ( H ) , or anti-PLC-C1 (I) antibodies. Lanes 1,
72; 5, PLC-61; S, PLC-C1. Arrows indi- cate molecular weight standards from top to bottom, myosin (200,000), phosphoryl- ase b (97,000), bovine serum albumin (68,000), ovalbumin (43,000), and car- bonic anhydrase (29,000).
PLC-81; 2, PLC-(32; 3, PLC-71; 4, PLC-
to pH 8 by adding 20 pl of 1 M Tris (pH 8.0) and incubated with 4 pg of trypsin at 30 "C for 16 h. The trypsin-digested peptides were chromatographed on a reverse-phase column (Vydac CI8 column, 4.6 X 150 mm) by eluting with a linear gradient of 0-60% CH&N in 0.1% trifluoroacetic acid. Amino acid sequence for each purified peptide was analyzed by using an AB1 471A Sequencer, according to the manufacturer's procedure.
Other Methods-SDS-PAGE was performed by the method of Laemmli (51), and the protein bands were stained with Coomassie Brilliant Blue. Electrophoretic transfer of protein from gels to nitro- cellulose sheets and subsequent immunoblotting using peroxidase- conjugated goat anti-rabbit IgG were as described elsewhere (52).
RESULTS
Purification of a New PLC Chromatography of a 13,000 x g supernatant of bovine
cerebellum directly on a DE52-cellulose column yielded one major and one minor peak of PLC activity, which were eluted a t 0.1 and 0.3 M KCl, respectively (Fig. lA). The minor peak, upon further purification, produced an 85-kDa enzyme, the substrate specificity, molecular weight, and chromatographic behavior of which were similar to those of PLC-I11 reported by Homma et al. (11) (data not shown). The major peak was subjected to chromatography sequentially on Butyl-Toyopearl and Spectragel Blue columns and in each case activity was eluted as a single peak (Fig. 1, B and C). Activity was resolved into two peaks by the TSK DEAE-5PW HPLC column (Fig. 1D). The first minor peak proved to be PLC-61 by subsequent purification and immunoblot analysis with monoclonal anti- PLC-61 antibody (data not shown). The second major peak was resolved again into two peaks by the Phenyl-5PW HPLC column (Fig. 1E). The second peak eluted between 0.3 and 0.5 M KC1 was shown to be PLC-71 by immunoblot analysis with monoclonal anti-PLC-yl antibody (data not shown). The first peak was further purified by sequential use of TSK Heparin-5PW, Mono S, and Mono Q HPLC (Fig. 1, F-H). In the last two HPLC steps, the presence of a 97-kDa band on SDS-PAGE correlated with PLC activity (designated tempo- rarily as PLC-C1) (Fig. 2 A ) . The final preparation obtained from the Mono Q HPLC column showed specific activity of 7.5 pmol of PI hydrolyzed per min/mg of protein under the assay conditions described under "Experimental Procedures" (Table I). The Mono S HPLC step was very effective for removing the major impurities and obtaining PLC-C1 as a major band on SDS-polyacrylamide gel.
Characterization of PLC-CI
Substrate Specificity-Specific activity of PLC-C1 for the hydrolysis of phosphatidylcholine was smaller than 1% of that for the hydrolysis of PI or PIP, under the same assay conditions.
p H Optimum-The effect of pH on the PI-hydrolyzing
G H 1 " "" *
I
4 s 5 s 1 2 3 4 5 s
TABLE I Summary of PLC-CI purification
Step Total Specific activity Protein activity
13,000 x g Supernatant DE-52 cellulose Butyl-Toyopearl Spectragel-Blue
Phenyl-5PW Heparin-5PW Mono S Mono Q
DEAE-5PW
w 87,000
15,000 4,700 1,100
320 63 6.8 0.3 0.08
prnol/rnin 270
175 160 75 43 14 3.6 0.9 0.6
prnol/rng/rnin 0.003
0.012 0.034 0.068 0.13 0.22 0.53 3.0 7.5
1
0 " " " " " " 4 5 6 7 0
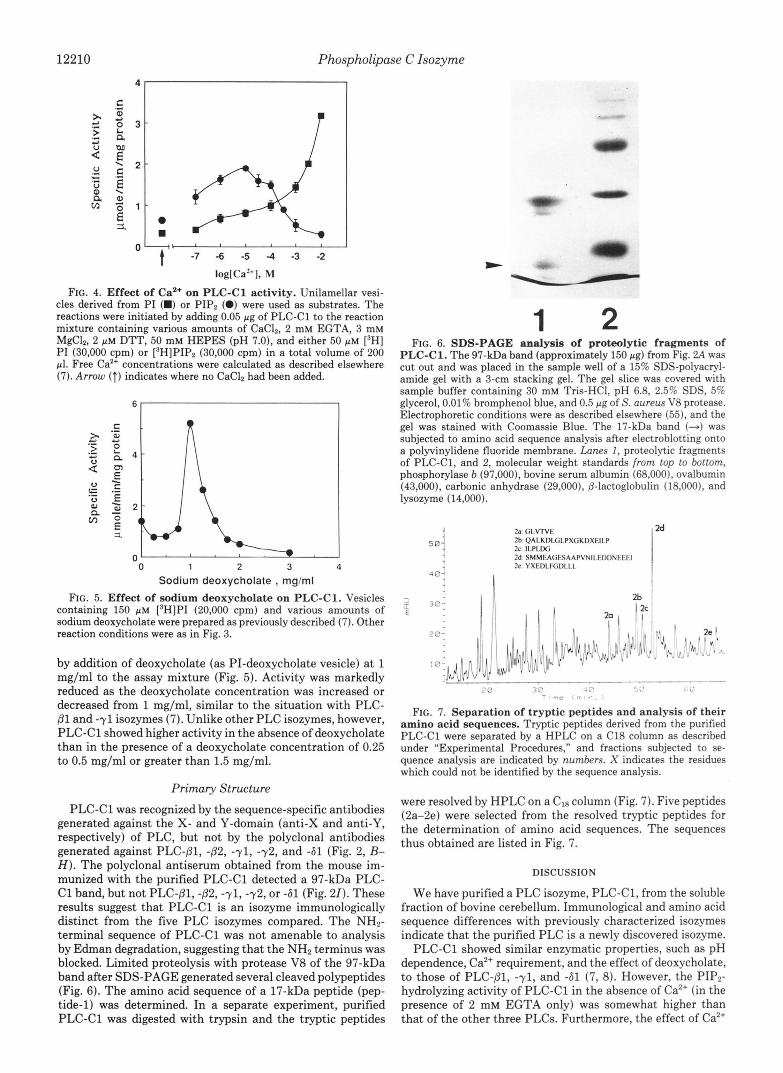
P H FIG. 3. Effect of pH on PLC-C1 activity. Unilamellar vesicles
derivied from PI were used as substrate. PI hydrolysis reactions were initiated by adding 0.05 pg of purified PLC-C1 to the reaction mixture containing 50 p~ ['HIPI (30,000 cpm), 2 mM EGTA, 3 mM Ca", and 2 p~ DTT. The pH was varied by the addition of 50 mM Tris-maleate buffer.
activity of PLC-C1 was measured by using vesicles of pure PI in the presence of 1 mM free Ca2+ (Fig. 3). PLC-C1 exhibited a pH optimum of 5.0, and its activity was decreased rapidly as the pH was increased or decreased. Less than 30% activity remained at pH 7.0-8.0.
Cu2+ Dependence-Purified PLC-C1 was incubated with either PI or PIP2 vesicles in the presence of various free Ca2+ concentrations (Fig. 4). Activities toward both PI and PIP, were markedly dependent on Ca", similar to those of PLC- Dl, -71, or -61 (8). However, PIPz hydrolysis by PLC-C1 was somewhat less sensitive to Caz+ than that catalyzed by the other three PLCs. Moreover, PLC-C1 activity in the absence of Caz+ (in the presence of EGTA only) was higher than that of the other three PLCs, especially for the hydrolysis of PIP,.
Effect of Sodium Deoxycholate on PLC-Cl Activity-The activity of PLC-C1 was enhanced up to approximately 4-fold
12210 Phospholipase C Isozyme
i o k i ' ' I t -7 -6 -5 -4 -3 -2
log[Ca*+], M
FIG. 4. Effect of Ca2+ on PLC-C1 activity. Unilamellar vesi- cles derived from P I (M) or PIPz (0) were used as substrates. The reactions were initiated by adding 0.05 pg of PLC-C1 to the reaction mixture containing various amounts of CaClZ, 2 mM EGTA, 3 mM MgClz, 2 p M DTT, 50 mM HEPES (pH 7.0), and either 50 p M [3H] PI (30,000 cpm) or [3H]PIPz (30,000 cpm) in a total volume of 200 pl. Free Caz+ concentrations were calculated as described elsewhere (7). Arrow (T) indicates where no CaClZ had been added.
0 1 2 3 4
Sodium deoxycholate , mg/ml FIG. 5. Effect of sodium deoxycholate on PLC-C1. Vesicles
containing 150 p~ [3H]PI (20,000 cpm) and various amounts of sodium deoxycholate were prepared as previously described (7). Other reaction conditions were as in Fig. 3.
by addition of deoxycholate (as PI-deoxycholate vesicle) at 1 mg/ml to the assay mixture (Fig. 5). Activity was markedly reduced as the deoxycholate concentration was increased or decreased from 1 mg/ml, similar to the situation with PLC- 81 and -71 isozymes (7). Unlike other PLC isozymes, however, PLC-C1 showed higher activity in the absence of deoxycholate than in the presence of a deoxycholate concentration of 0.25 to 0.5 mg/ml or greater than 1.5 mg/ml.
Primary Structure PLC-C1 was recognized by the sequence-specific antibodies
generated against the X- and Y-domain (anti-X and anti-Y, respectively) of PLC, but not by the polyclonal antibodies generated against PLC-81, -82, -71, -72, and -61 (Fig. 2, B- H). The polyclonal antiserum obtained from the mouse im- munized with the purified PLC-C1 detected a 97-kDa PLC- C1 band, but not PLC-p1, $2, -71, -72, or -61 (Fig. 21). These results suggest that PLC-C1 is an isozyme immunologically distinct from the five PLC isozymes compared. The NH2- terminal sequence of PLC-C1 was not amenable to analysis by Edman degradation, suggesting that the NH2 terminus was blocked. Limited proteolysis with protease V8 of the 97-kDa band after SDS-PAGE generated several cleaved polypeptides (Fig. 6). The amino acid sequence of a 17-kDa peptide (pep- tide-1) was determined. In a separate experiment, purified PLC-C1 was digested with trypsin and the tryptic peptides
1 2 FIG. 6. SDS-PAGE analysis of proteolytic fragments of
PLC-C1. The 97-kDa band (approximately 150 pg) from Fig. 2A was cut out and was placed in the sample well of a 15% SDS-polyacryl- amide gel with a 3-cm stacking gel. The gel slice was covered with sample buffer containing 30 mM Tris-HC1, pH 6.8, 2.5% SDS, 5% glycerol, 0.01% bromphenol blue, and 0.5 pg of S. aureus V8 protease. Electrophoretic conditions were as described elsewhere (551, and the gel was stained with Coomassie Blue. The 17-kDa band (-4 was subjected to amino acid sequence analysis after electroblotting onto a polyvinylidene fluoride membrane. Lanes I, proteolytic fragments of PLC-C1, and 2, molecular weight standards from top to bottom, phosphorylase b (97,000), bovine serum albumin (68,000), ovalbumin (43,000), carbonic anhydrase (29,000), @-lactoglobulin (18,000), and lysozyme (14,000).
2h QALKDLCiLPXGKDXEII.P 2a c i l . v r v E
2c II.PI.IXi 2d SMMliAciESAAPVNII.EDDN~~~l 2e YXFDI.FC,I)I.I.I.
I I 2b
I * ! ! I I
172 a ." - . " I I
.. . . -. . . ' ..
FIG. 7. Separation of tryptic peptides and analysis of their amino acid sequences. Tryptic peptides derived from the purified PLC-C1 were separated by a HPLC on a C18 column as described under "Experimental Procedures," and fractions subjected to se- quence analysis are indicated by numbers. X indicates the residues which could not be identified by the sequence analysis.
were resolved by HPLC on a CIS column (Fig. 7). Five peptides (2a-2e) were selected from the resolved tryptic peptides for the determination of amino acid sequences. The sequences thus obtained are listed in Fig. 7.
DISCUSSION
We have purified a PLC isozyme, PLC-C1, from the soluble fraction of bovine cerebellum. Immunological and amino acid sequence differences with previously characterized isozymes indicate that the purified PLC is a newly discovered isozyme.
PLC-C1 showed similar enzymatic properties, such as pH dependence, Ca2+ requirement, and the effect of deoxycholate, to those of PLC-p1, -71, and -61 (7, 8). However, the PIP2- hydrolyzing activity of PLC-C1 in the absence of Ca2+ (in the presence of 2 mM EGTA only) was somewhat higher than that of the other three PLCs. Furthermore, the effect of Ca2+

Phospholipase C Isozyme 12211
on the PIP2-hydrolyzing activity of PLC-C1 was smaller than its effect on the other three PLCs (2-3-fold uersw more than
Previously, Ryu et al. (8) purified PLC-81, -71, and -61 from bovine cerebrum proteins precipitated at pH 5.0, but failed to detect a peak corresponding to PLC-C1. It is not clear whether PLC-C1 might be specifically localized in cer- ebellum or whether Ryu et al. might not have detected PLC- C1, because it was not precipitated at pH 5.0. PLC-yl and - 61 were separated from PLC-C1 by the Phenyl-5PW and DEAE-5PW HPLC columns, respectively, and were identified by immunoblot analysis with anti-PLC-yl or -61 antibody. We could also detect PLC-81 eluted between 60 and 80 mM KC1 on DE52 column chromatography by using the radio- immunoassay described by Suh et al. (55). However, PLC-81 activity could not be detected, possibly because PI-hydrolyz- ing activity of PLC-81 at high calcium concentration (10 mM, in this work) is less than that of PLC-yl or PLC-61 (7).
PLC-C1 was recognized by both the anti-X and anti-Y antibodies. None of the polyclonal antibodies generated
10-fold).
against PLC-Dl, -82, -71, -72, and -61 cross-reacted with PLC-C1, and the anti-PLC-C1 antiserum did not react with these five PLCs (Fig. 2). Among the six peptide sequenced, three (peptide-1, -2c, and -2e) shared some similarity with the sequences of all known PLC isozymes. Besides, as noted from Table 11, the three PLC-C1-derived peptides exhibit higher sequence identities with PLC-8 type enzyme than with PLC- y or PLC-6 types. The highest identity was found in the peptides of the norpA product, a PLC-/3 type enzyme. More convincing evidence for PLC-C1 being a member of PLC-8 type came from the sequences of peptide-2b and -2d. Peptide- 2b showed a significant similiarity to the region that precedes the X-domains of all known PLC-8 type enzymes but not to any regions of other PLC isozymes. Furthermore, peptide-2d showed showed a low but statistically meaningful homology to the region separating the X- and Y-domains of PLC-81 and PLC-82. Amino acid sequence identity outside the X- and Y-domains is negligible between different members but is shown to be significant between the members of the same type PLC (57). For example, PLC-Dl and PLC-82 exhibit a
TABLE I1 Comparison of the partial amino acid sequences of PLC-Cl with those of other PLC isozymes
Isozyme Amino acid sequence" Identity
Peptide-1 PLC-81 731 *
S F V F R K V I L P D L A V L R I A V Y p i V F k K V v L P s L A c L R I A a Y
PLC-82 733 PLC-83'
p F V F e K i l m P e L A s L R v A V m p F d F p K V v L P t L A s L R I A a f
norpA 640 p F V F R K V v L P D L A V L R f g V Y plc21 790 p F V F k K V v L P e L A s i R I A a Y PLC-yl 1144 p F h F q i s n P e f A f L R f v V Y PLC-y2 1117 PLC-61 PLC-62 688 PLC-63' Peptide-2c PLC-81 761
I L P L D G
PLC-82 I L P v q a
763 I i P i n a PLC-83' I L P v s a mrpA 765 I L P L D G plc21 820 PLC-yl 1177
v L P v i G t f P v k G
PLC-y2 1152 PLC-61 720
t y P i k a t i P w n s
PLC-62 693 PLC-63" 699
I L v p e l t L P L s s
Peptide-2ed Y X E D L F G D L L L PLC-81 427 PLC-82
Y c r l i F G D a L L
PLC-PBC Y c r t i F G D m L L Y c r s i F G D a L L
mrpA 429 Y c d D f F G D L L L plc21 426 PLC-71 428
Y c r e i F G D m L L
PLC- y2 h f r k v l G D t L L
420 PLC-61 404
a f k e v F G D L L L
PLC-62 h l r a i l G p i L L
397 PLC-63' 392
h l t e i l G D q L L h l c t i l G D m L v
Peptide-2b Q A L K D L G L P X G K D X E I L P PLC-81 187 PLC-82 183
t A L e a c s L P s s r n d s I p q a A L s a c h L P k G K n d a I n P
PLC-83' t A L e s c G L k f n r s e s I r P norpA 190 plc21
t c i K D a G L P D d K n a t m t k 188 k A i d v t G L P s G K v d s I s v
Peptide-2d' PLC-81 495 PLC-82 499 S p p s A p a v w A g e e g t E l e e E E v e
e k V t f e i y d P n L A f L R f v V Y 687 e F e F e V t v P D L A l v R f m V e
t l c F r i l v P e L A l L R f v V k 667 t l q F q l r a P e L A l v R f v V e
423
S M M E A G E S A A P V N I L E D D N E E E I S v f E p s s p g A g e a d t E s D D d d d d
Identical residues are shown in uppercase letters; nonidentical residues in lowercase letters.
The amino acid sequences were provided by Dr. Sue Goo Rhee, National Institutes of Health. X indicates the residue which could not be identified.
* Number indicates the NHn-terminal residue number of the sequence compared.
e Acidic residues were shown in italics.
%
100 65 55 60 80 65 40 40 45 35 30 100 50 33 50 100 50 33 17 17 33 50 100 60 60 60 80 60 40 60 30 40 30 100 28 44 33 39 39 100 26 26

12212 Phospholipase C Isozyme
48% identity in the amino-terminal region preceding the X- domain and a 28% identity in the region separating X and Y. Another characteristic feature of mammalian PLC-8 type is a high proportion of acidic amino acids between the X- and Y-domains (58). The 23-membered peptide-2d contained 8 acidic residues. Sequence homologous to peptide-2a could not be found in any PLC sequences.
PLC-C1 migrated as a 97-kDa band on SDS-PAGE and appeared to have a blocked NHZ terminus. There are no reports of intact PLCs with a molecular mass of 97 kDa. However, 100-kDa PLC proteolytic fragments of the 150-kDa PLC-pl (8) and the 140-kDa PLC-62 have been described.' Both of the fragments could be detected by the polyclonal antibodies generated against the corresponding intact en- zymes (Fig. 2, D and E ) . We could not detect any 140-150- kDa bands after immunoprecipitation of the cerebellar extract with anti-PLC-C1 antiserum (data not shown). It is not clear, however, whether the 97-kDa PLC-C1 is generated from a higher molecular weight enzyme or whether it is an intact enzyme. In this connection, it is important to note that the apparent molecular weight of the product of norpA, which was predicted to encode a 1095-residue protein with a calcu- lated molecular weight of 125,000, was shown to be only 98,000 on SDS-PAGE (16,56). From all of these results, we conclude that the purified PLC-C1 is a different isozyme from those PLC-01, -71, -61, and -62 previously purified from mammalian brain and should be classified as a PLC-p- type isozyme. We therefore suggest that PLC-C1 should be designated PLC-p4, and that at least five distinct isozymes exist in bovine brain. It is not clear whether PLC-C1 is a mammalian homolog of norpA gene product; the norpA product does not contain acidic amino acids between the X- and Y-domains.
Three mammalian PLC-ps (PLC-01, $2, and $3) have been cloned (22,27). However, only PLC-01 has been purified from bovine brain (7), and the existence of PLC-p2 or PLC- p3 in mammalian cells has not yet been reported. PLC-pl has been shown to be coupled to G, and G1l (43-47). Two addi- tional members of G, class, G14 and GI6, have been described (48), but have not yet been shown to associate with a PLC-0 type isozyme. It is possible that one of these two G-proteins will be shown to be associated with PLC-@4.
REFERENCES 1. Nishizuka, Y. (1984) Science 226.1365-1370 2. Majerus, P. W., Connolly, T. M., Dickmyn, H., Ross, T. S., Bross, T. E.,
Ishii. H.. Bansal. V. S.. and Wilson. D. B. (1986) Science 234. 1519- 1526 '
. .
3 Ben1 e M J., and Irvine, R. F. (1989) Nature 341,197-205 4: R a n a s : S., . and Hokin, L. E. (1990) PhysioL Rev. 70,115-164 5. Hofmann, S., and Majerus, P. W. (1982) J. Biol. Chem. 267,6461-6469 6. Ryu, S. H., Cho, K. Y., Lee, K. Y., Suh, P. G., and Rhee, S. G. (1986)
7. Ryu, S. H., Cho, K. S., Lee, K. Y., Suh, P. G., and Rhee, S. G. (1987) J.
8. Ryu, S. H., Suh, P . G., Cho, K. S., Lee, K. Y., and Rhee, S. G. (1987) Proc.
9. Bennett, C. F., and Crooke, S. T. (1987) J. Bid. Chem. 262,13789-13797 10. Katan, M., and Parker, P. J. (1987) Eur. J. Bwchem. 168.413-418 11. Homma, Y., Imakl, J., Nakanishi, O., and Takenawa, T. (1988) J. Biol.
12. Banno, Y., Yada, Y., and Nozawa, Y (1988) J. Biol. Chem. 2 6 3 , 11459-
Bwchem. Bwphys. Res. Commun. 141,137-144
Biol. Chem. 262,12511-12518
Natl. Acad. Set. U. S. A. 84,6649-6653
Chem. 263,6592-6598
11465
Y. H. Lee, I. K. Lee, S. H. Ryu, and P. G. Suh, unpublished observation.
13. Fukui, T., Lutz, R. J., and Lowenstein, J. M. (1988) J. Biol. Chem. 2 6 3 ,
14. Rhee, S. G., Suh, P. G., Ryu, S. H., and Lee, S. Y. (1989) Science 2 4 4 ,
15. Banno, Y., Yu, A., Nakaahima, T., Homma, Y., Takenawa, T., and Nozawa,
16. Toyoshima, S., Masumoto, N., Wang, P., Inoue, H., Yoshika, T., Hotta, Y.,
17. Tomkins, T. A., and Motxarello, M. A. (1991) J. Biol. Chem. 2 6 6 , 4228-
18. Roy, G., Villar, L. M., Lazaro, I., Gonzalez, M., Bootello, A., and Gonzalez-
19. Grlenai;, K . K. Taubman M. B., Akers, M., Mendlowitz, M., and
20. Stahl, M. L., Ferenz, C. R., Kelleher, K. L., Kriz, R. W., and Knopf, J. L.
21. Suh, P. G., Ryu, S. H., Moon, K. H., Suh, H. W., and Rhee, S. G. (1988)
22. Suh, P. G., Ryu, S. H., Moon, K. H., Suh, H. W., and Rhee, S. G. (1988)
17730-17737
546-550
Y. (1990) Biochem. Bwphys. Res. Commun. 167,396-401
and Osawa, T. (1990) J. Biol. Chem. 266,14842-14848
4236
Par ue P (1991) J. Biol. Chem. 266,11495-11501
Alexanier, R. W . (1991) J. Bwl. Chem. 266, 15498-15504
(1988) Nature 332,269-272
Proc. Natl. Acad. Sci. U. S. A. 86,5419-5423
Poll Kd. 1 fil -1 fiR 23. Katan, M., Kriz, R. W., Totty, N., Philp, R., Meldrum, E., Aldape, R. A.,
24. Ohta, 8.: ' Matsue, A., Nozawa, Y., and Kawaka, Y. (1988) FEBS Lett. 2 4 2 ,
".- ", -"- -1- Kno f J L., and Parker, P. J. (1988) CeU 5 4 , 171-177
25. Emori, Y., Homma, Y., Sarimachi, H., Kawasaki H., Nakanishi, O., Suzuki,
26. Meldrum, E., Kriz, R. W., Totty, N., and Parker, P. (1991) Eur. J. Biochem.
27. Kriz, R., Lin, L.-L., Sultzman, L., Ellis, C., Heldin, C.-H., Dawson, T., and
tion Symposium 150) p 112-127, Wiley, Chichester Knopf, J. (1990) in Proto-Oncogenes in Cell Development (Ciba Founda-
28. Wahl, M. I. Nishibe S. &h P. G., Rhee, S. G., and Carpenter, G. (1989) Proc. Nail. Acad. ici. 'U. S . k 86,1568-1572
29. Mar olis, B., Rhee, S. G., Felder, S., Mervic, M., Lyall, R Levitzki, A,, U h c h , A., Zilberstain, A., and Schlessenger, J. (1989) b U 67, 1101-
30. Meisenhelder, J., Suh, P. G., Rhee, S. G., and Hunter, T. (1989) Cell 67, 1107
31. Nishibe, S. Wahl, M. I., Wedegaertner, P. B., Kim, J. J., Rhee, S. G., and 1109-1122
32. Anderson, D., Koch, C. A., Grey, L., Ellis, C., Moran, M. F., and Powson, Carpent&, G. (1990) Proc. Natl. Acad. Sci. U. S. A. 87,424-428
33. Kim, H. K., Kim, J. W., Zilberstein, A., Margolis, B., Kim, J. G., Schlessin- T. (1990) Science 260,979-982
eer. J.. and Rhee. S. G. (1991) Cell 65.435-441
31-35
K., and Takenawa, T. (1989) J. Biol. Chem. 264,21855-21890
196,159-165
34. 35.
36. 37.
38.
R h , ' S.'G. (1991) Trends'B~&hm. Sci. .16, 297-301 Sultzman, L. Ellis, C., Lin, L.-L., Pawson, T., and Knopf, J. (1991) Mol.
Cockcroft, S. (1987) Trends Biochem. Sei. 12,75-78 Morris, A. J., Waldo, G. L.. Downs, C. P., and Harden, T. K. (1990) J. Biol.
Cell. Biol. i l , 2018-2025
Taylor, S. J., Smith, J. A,, and Exton, J. H. (1990) J. Bwl. Chem. 266, Chem. 268,13508-13514
17150-17156 39. Thomas G. M. H., Geny, B., and Cockcroft, S. (1991) EMBO J. 10,2507-
251 2 40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51. 52.
53.
54. 55.
56.
58. 57.
Gutowski, S. Smrcka A., Nowak L., Wu D. Simon, M., and Sternweis,
Shenker, A., Goldsmith, P., Unson, C. G., and Spiegel, A. M. (1991) J. Biol.
Wange, R. L., Smrcka, A. V., Sternweis, P. C., and Exton, J. H. (1991) J.
Taylor, S. J., Chae, H. Z., Rhee, S. G., and Exton, J. H. (1991) Nature 360,
Smrcka, A. V., Helper, J. R., Brown, K. O., and Sternweis, P. C. (1991)
Waldo, G., Boyer, J., Morris, A., and Harden, T. K. (1991) J. Biol. Chem.
Blank, J. L., Ross, A. H., and Exton J. H. (1991) J. Biol. Chem. 2 6 6 ,
Wu, D., Lee, C. H., Rhee, S. G., and Simon, M. I. (1992) J. Biol. Chem.
Simon, M. I., Strathmann, M. P., and Gautam, N. (1991) Science 2 6 2 ,
P. C. (199i) J. BioL'Chem. 266,20519-505i4
Chem. 266,9309-9313
Bwl. Chem. 266,11409-11412
516-518
Science 26 1,804-807
2 6 6 , 14217-14225
18206-18216
2 6 7 , 1811-1817
802-808 Harlow, E., and Lane, D. (1988) Antibodies; A Laboratory Manual, pp. 53-
Hofmann, S. L., and Majerus, P. W. (1982) f: Biol. Chem. 2 6 7 , 14359-
.~~
119, Cold Spring Harbor Laboratory, Cold S ring Harbor, NY
14Rfi4 Laemmii, U. K. (1970) Nature 227,680-685 Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci.
Cleveland, D. W., Fisher, S. G., Kirshiner, M. W., and Laemmli, U. K. Matsudaira P. (1987) J. Biol. Chem. 262,10035-10038 Suh. P. G.. Rvu. S. H.. Choi. W. C.. Lee. K. Y.. and Rhee. S. G. (1988) J.
U. S. A. 76,4350-4354
(1977) J. Biol. Chem. 2 6 2 , 1102-1106
Bloomquist, ~ Schneuwly, S., Perdew, M., Montell,
Rhee. S. G.. and Chol. K. D. 11992) J. Btol. Chem. 267.12393-12396 C., Stellar, H., Ru@n, G.;-and Pak; W: L. (1988) Cell 64,723-733
Park,'D., Jhon, D.-Y.: Kriz, R . , Knopf, J., and Rhee, S , G. (1992) J. Biol. Chem. 267,16048-16055