jane gao janegao003@gmail - oregon state university
TRANSCRIPT

1
Effects of Woolsey Fire on Nesting Territories of Southern California Red-Tailed Hawks (Buteo
jamaicensis)
Jane Gao

2
ABSTRACT: The relationship between fire and wildlife habitat is complex. Fires can create
favorable conditions for some species and simultaneously extirpate entire populations of other
species. Red-tailed hawks choose nesting habitats according to resources available. Natural
disturbances such as wildfires change the availability of those resources. In late 2018, the
Woolsey fire burned much of Ventura County, California, changing composition of the flora and
fauna in areas affected by the fire. Nest site use for Red-tailed hawks (Buteo jamaicensis) has
been tracked in the Santa Monica Mountains of Ventura county from the 1970s to 2019. The
Woolsey fire area is surrounded by mountain ranges as well as suburban areas, providing hawks
with a variety of nesting habitats consisting of both native and nonnative trees. The Woolsey fire
affected these varied habitats differently. The area burned by the Woolsey fire had not been
largely affected by fire since the 2003 Simi fire. The fact that this area has been free from
damaging stochastic events for over a decade made it a prime area for studying the immediate
effects of fire on raptor nesting. We investigated how fire affected nest habitability and whether
several variables factored into nest persistence within the 2018 Woolsey fire area. Nest
persistence in this context was defined by the presence of a nesting hawk. We hypothesized that
the magnitude of the burn and the species of the tree in which the nest was built were correlated
to nest persistence. In addition, we proposed that geographic position relative to the fire border,
and not just local destruction of nests, was a key variable in nest persistence. While proximity to
the edge of the fire’s border and magnitude of burn severity were found to be correlated with
nest persistence, tree species was found to have no discernible correlation.

3
INTRODUCTION
Wildfires have always been a part of Southern California’s ecosystems (Cheney et al. 1997).
They engender a patchwork effect on local habitats. Some of the positive outcomes of wildfires
include the release of nutrients back into the soil, clearing space for new seedlings, and removing
nonnative species. Negative outcomes include the destruction of habitat, death of native species,
and removal of erosion-preventing plants (Cheney et al. 1997). While the impact that wildfires
have on California’s ecosystems has both positive and negative aspects, the overall increase in
both number of fires and fire severity in recent decades has been the subject of major study.
In more recent decades human suppression of fire increased, causing a buildup of vegetative fuel
in fire-prone areas (Bowman 2010). This suppression was meant to limit the number of fires
under the notion that fewer fires would create a safer environment for human settlements.
Additionally, climatic controls have changed and altered size, severity, and fire frequency
(Hurteau et al. 2014). Climate change created a longer fire season with more powerful winds and
higher ambient temperatures (Westerling et al. 2008). Therefore, when fires do occur, they
spread quickly and over a larger amount of terrain (Keeley et al. 2001, Syphard et al. 2007). The
change in fire regime can have a profound change on the ecosystem, including vegetation shifts,
invasive species, a decrease in biodiversity, and population shifts (Westerling et al. 2008,
Hurteau et al. 2014, Williams et al. 2019). California’s growing population and increase in fire
activity present a new stressor on Red-tailed hawk nesting habits. The goal of this study is to see
how Red-tailed hawk nesting sites were affected by the 2018 Woolsey fire. Studying wildfire
disturbance effects on apex predators can help future land management make informed
management decisions.

4
In order to study how fires affect Red-tailed hawk nesting habits, it is prudent to understand
California’s historical fire regime. The last few decades have seen a drastic rise in the number of
acres burned in California wildfires (Figure 1). This increase in burned land area corresponds to
a marked increase in California’s average temperature (Figure 2) and in California’s total
number of fires per year (Figure 3). These longitudinal trends of increased intensity and
frequency of California wildfires represent a significant threat to California’s wildlife. One group
that can be especially affected by an increase in wildfire size and frequency is birds of prey.
Fires can change how raptors use available habitat via direct and indirect cascading effects.
While the fire itself may not kill the raptors themselves, increased wildfires may drive raptor
prey to extinction or drive away animal species in lower trophic levels. These changes in the
raptor’s food chain cascade up the trophic levels by simplifying the food web, thus limiting the
raptor’s prey availability. Fire creates a mosaic of habitat patches with variable biotic and abiotic
conditions. For example, fire disturbances can produce early seral plant communities that support
different animal communities compared to pre-fire communities (Keane 2002). The new
vegetative growth provides food and cover for early seral species that colonize the area (Keane
2002). In areas where fires are the most common disturbances, plants and animals have evolved
various adaptations that allow them to survive or benefit from fire. Desert plants evolved
pyriscence or flammable oils with fire-activated seeds (Brown 2000). Other flora such as oak
trees and pine trees have adapted to fire with either more resistant bark or legacy sprouting
(Barton 1999). One subgroup of fauna that is both affected by and even utilizes fire is raptors.
Raptors take advantage of wildfires by searching for flushed out prey seeking to escape the
flames (Sahores et al. 2004, Steenhof et al. 1999). After the wildfire is over, raptors scavenge
burned areas for prey, taking advantage of the razed vegetation and consequent lack of cover

5
(Sahores et al. 2004, Steenhof et al. 1999). Raptors’ ability to capitalize on fire as a natural
disturbance can maximize their productivity in an environment with variable resources.
Figure 1. The number of acres burned per year in California since 1950
Source: Cal Fire. 2020 Number of acres burned per year (in millions)
https://www.fire.ca.gov/stats-events/

6
Figure 2. The average temperature in California shown from 1895 to 2018.
Source: NOAA. 2019 Average temperature in California from January through October each
year, 1895-2018. https://www.ncdc.noaa.gov/cag/
Figure 3. The number of fires reported per year in California starting from 1913 to 2013.
Source: Cal Fire. 2020 Number of Fires Reported Per Year in California.
https://www.fire.ca.gov/stats-events/

7
Scientists in the Southern California National Park Service (NPS) and non-government
organizations (NGOs) have been tracking the effects of urbanization on raptors in the Ventura
County area since 1972. Red-tailed hawks (Buteo jamaicensis) were chosen for the study due to
their position as apex avian predator, their wide trophic niche, and their large population size.
These qualities made them good candidates for urban and wild comparative studies (Marti et al.
1993).
The 2018 fire season was the deadliest and most destructive season on record in California (Cal
Fire 2018, Figure 1, Figure 2, Figure 3). The Woolsey Fire was the largest fire in Southern
California in 2018 and burned 96,949 acres of land, making the landscape more susceptible to
invasive weeds and grasses (LA County 2018). The buildup of non-native vegetation before the
fire increased the vegetative fuels which led to a more fire susceptible landscape, thus creating a
feedback loop (NPS 2018). While the fire as a whole was severe, the severity varied greatly
within Ventura County’s local microenvironments. Some habitats were completely razed to the
ground, while others remained wholly unburned. This variation in severity within the study area
created a mosaic of local environments.
Overall, larger animals had more success in escaping the Woolsey fire, while smaller animals
were more likely to succumb to the smoke and heat (NPS 2018). The Woolsey fire’s blaze
occurred in November, before the Red-tailed hawk nesting season, which starts in early Spring
(Cooper 2019). Red-tailed hawks would have been able to escape the fire easily, but many of
their prey species populations declined. Perch sites necessary for Red-tailed hawks’ hunting also
declined in burned areas. The circumstances of the Woolsey fire present a unique opportunity to
study how the loss of perches, vegetation, and prey affect the nesting habits of the Red-tailed
hawk species.

8
The study area that was affected by fire is undergoing early stages of post-fire succession that
includes an influx of colonizing plants such as Lupines (Lupinus sp.) and Blue dicks
(Dichelostemma capitatum). Early-seral plant species take advantage of the nutrient influx and
open understory, which creates a more biodiverse landscape with pioneer plant species now able
to grow (Arno 2000). These new plant species engender more biodiversity by replacing pre-fire
habitats dominated by a few species with a variety of pioneering early seral species (Arno 2000).
Plant biodiversity then invites R-select animal species (Arno 2000). Forest legacies such as Coast
live oak and Sycamore have also started to sprout and will continue to grow and spread in the
burned area, eventually outcompeting the pioneering species.
The 2018 Woolsey Fire offers an opportunity to studyhow fire affects nest selection for Red-
tailed hawks. We used previously published works (Lee 2004) of historical nest site locations,
territory activity, nesting activity, and tree species in which the nest was located as a basis for
our own data collection. By cross-referencing the old dataset with new data collected after the
fire damage, we can investigate how the Woolsey Fire affected the Red-tailed hawk species in
terms of nesting sites. Our objectives are to quantify how the Woolsey fire affected these
historical Red-tailed hawk nest sites in Ventura County and to determine how the severity of the
microenvironment’s burn contributed to the level of nesting activity. The main variable
considered was nest persistence, which is defined as a nest that is still in use by a Red-tailed
hawk and is still structurally stable following the 2018 fire. We hypothesized that the burn
severity around the nest site, the tree species in which the nest was located, and the nest’s
position relative to the fire’s edge would all affect the nest persistence. The geospatial
relationship between nesting territories was also mapped and analyzed to gain clarity into how
the relative positioning of the nests affected persistence.

9
METHODS
Study area:
Our study site comprised of the entirety of Ventura County, California. It is just north of Los
Angeles County, running along the Pacific coast, and thus along the California Current. The
California Current is an oceanic current that runs southward along the Pacific coast of North
America. It creates summer fog off the coast and produces a Mediterranean climate, with warm,
dry summers just inland, and mild winters (NCDC 2010). Precipitation exceeds 127 cm per year,
with floods occurring during periods of heavy rainfall (WRCC 2010). The topography varies
from 15 m along the coast to 940 m in the Santa Monica Mountains, with both heavily urbanized
areas, open space landscapes, and managed forests (VCTM 2010). Main vegetation communities
include grasslands, chaparral and coastal sage scrub, oak woodland, and urbanized, ornamental
landscapes.
Previously, wildfires have not been a seasonal threat to the study area, historically occurring
every hundred years (NPS 2018). Due to changes in the fire regime, there has been an uptick of
fires in the Ventura County area with over nine fires since 2003 (Keane 2002, Robert E. et al.
2009). The current fire frequency of more than one every 20 years represents a stark acceleration
from previous decades (NPS 2018). The most devastating fire in the Ventura County area in
2018 was the Woolsey Fire. The Woolsey Fire burned 39,234 ha in both Los Angeles and
Ventura counties, extending southwest from the west to the Pacific Ocean and north to south
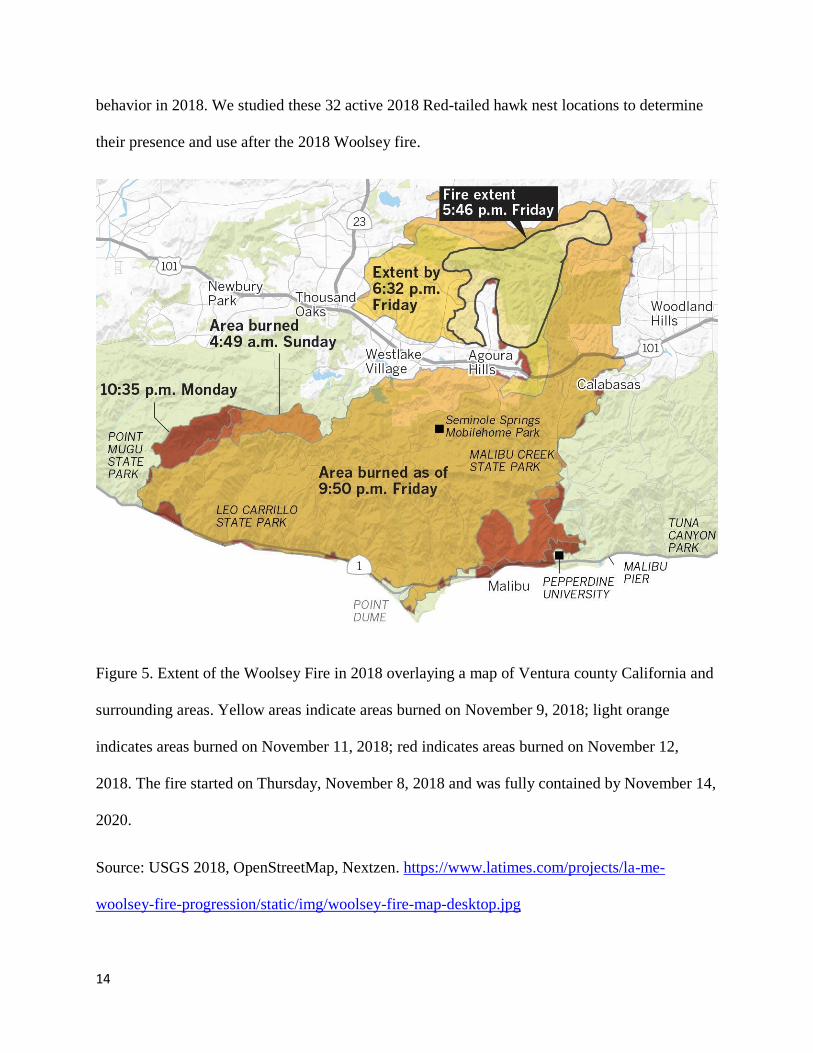
from Point Mugu State Park to Calabasas (Figure 5).
There are many native trees in the Ventura County area which covers the riparian, mountainous,
and coastal strips of the county (Trees for Ventura CA 2018). These trees are adapted for the

10
conditions that historically occurred in Ventura. The Western Sycamore (Platanus racemosa) is
a native tree that grows in canyons, floodplains, and along streams (Trees for Ventura CA 2018).
The Valley Oak (Quercus lobata) ranges over hotter temperature areas of California and grows
quickly (Trees for Ventura CA 2018). The Coast live oak (Quercus agrifolia) are oak trees that
are extremely fire tolerant and also appear in Ventura County (Holland 1986). The pine trees
found in Ventura are one of the most varied and widely spread genus (Pine Trees of California
Need year). Most pine trees are not fire tolerant, the exception being the Pondera pine (Pine
Trees of California). Poplar trees line up California with many native and non-native species.
The native Fremont Cottonwood poplar (Populus fremontii) grows along riparian areas requiring
moist soil and plenty of sunlight (California Native Plant Society 2007) The Eucalyptus trees
came from Australia, introduced to California due to high demand for fast growing wood during
the Goldrush era (Rowland et al. 2019). These trees became a focal point in this study, since they
are one of the factors we hypothesized would determine if the Red-tailed hawk nests persisted.
Study Species:
The Red-tailed hawk is a bird of prey found throughout most of North America (Figure 4). This
species is one of the largest in the genus Buteo, weighing from 690 to 1600 grams (Ferguson-Lee
and Christie 2001).

11
Figure 4. Geographical distribution of the Red-tailed hawk. Orange represents areas that Red-
tailed hawks migrate to for breeding purposes. Purple represents areas that the Red-tailed hawk
remain in year-round. Blue represents areas that the Red-tailed hawk remain only while not
breeding.
Source: BirdLife International 2006
When surveying for them, a bird can be identified as a Red-tailed hawk visually using its
distinctive form. This species of hawk soars with wings often in a dihedral, with a sleek body
(Ballam 1984). Their markings include a distinctive red auburn tail with black patagium
markings on its wings (Ferguson-Lees and Christie 2001). Their distinctive body shape of broad
rounded wings and short wide tails also aided in their identification. Vocalization would also

12
lead to identification with their distinctive cry of two to three second hoarse rasping screams that
begin at high pitches and skew downwards (Ferguson-Lees and Christie 2001).
Red-tailed hawks of Santa Monica Mountains choose nesting sites in both suburban and
wilderness environments. In urban environments they frequently choose sites that are close to
condo complexes and houses (Strahlberg 1999). This anthropogenic pressure creates new free
ecological niches that raptors such as the Red-tailed hawk readily colonize (Luniak 2004).
Raptors’ heuristics for choosing nest sites largely depend on geomorphologic habitat
characteristics as well as prey availability (Poirazidis 2007). Red-tailed hawks frequently inhabit
and defend steep mountain ranges as preferential territory (Boal 1998; Preston 2000). Raptors
frequently choose Western sycamores (Planus racemosa), coastal live oaks (Quercus agrifola),
pine trees (Pinus), and Eucalyptus trees (Eucalyptus globulus) as nesting sites (Cooper 2019).
The structurally sound crowns of the trees along with the high compressive strength of their thick
trunks lend good structural integrity to the raptors’ nesting locations. While these trees are
commonly used by raptors for nesting sites, they are by no means the only structures used;
transmission towers, planters, and even masts of sunken boats have been used (Ellis 2009).
Study Design and Field Sampling:
Raptor nest surveying was the main data collection process for analyzing the effects of the fire
on raptor habitation. The process of nest surveying involves searching visually for raptor nests,
recording their positions, and ascertaining their structural integrity. Raptor nests were identified
and visually assessed. A mated raptor pair often construct or re-use nests made from sticks and
other materials in a large tree between 4 to 21 m from the ground (Stotz 1994). Their nesting
material consists of twigs, bark, pine needles, and plant matter (Preston 2009). The nest is

13
generally 71 to 97 cm in diameter (Stotz 1994). The absence of trash excluded nests that were
likely to be crow or squirrel nests rather than raptor nests (Large Nest Identification 2020). We
also recorded whether nests were built in mature trees, built in a supportive crotch of a tree, or
built using sticks.
In this study, we chose to focus on Red-tailed hawks due to the quantity of data we gathered.
Red-tailed hawks were the only species that had a large enough sample size to be considered
statistically significant, with 32 active nest sites in 2018. Other species studied (such as the
Cooper’s hawk, Red-shouldered hawk, and American kestrel) had fewer than 20 active nests in
2018 and were thus not considered to have a high enough sample size for studying. This study’s
scope focuses on active nests found in 2018 before the Woolsey Fire with the comparison of
nesting survey data in 2019 post-Woolsey Fire.
The entirety of Ventura County has been the subject of raptor nest surveying since the 1970’s
(Lee 2004). The goal of previous raptor surveys was primarily to track population sizes and to
determine the geographic locations of raptor habitat. Lee (2004) and Cooper compiled a database
of historic nest sites from Ventura County, each labeled with a unique name and recorded with
the nearest watershed, geographic location, years of activity, and the species of tree in which the
nest was found. These legacy Red-tailed hawk nest locations formed the basis for our study.
Cooper selected study sites by surveying nesting locations identified by Lee (2004), whose
database was used by the National Park Service (NPS) to understand local raptor population
nesting habits. In 2018, Cooper recorded 32 active Red-tailed hawk nests through the entirety of
Ventura County (2018). Cooper’s collected data included GPS coordinates for these historical
nests, along with tree type, nearby watersheds, and some field notes on the raptor’s nesting
14
behavior in 2018. We studied these 32 active 2018 Red-tailed hawk nest locations to determine
their presence and use after the 2018 Woolsey fire.
Figure 5. Extent of the Woolsey Fire in 2018 overlaying a map of Ventura county California and
surrounding areas. Yellow areas indicate areas burned on November 9, 2018; light orange
indicates areas burned on November 11, 2018; red indicates areas burned on November 12,
2018. The fire started on Thursday, November 8, 2018 and was fully contained by November 14,
2020.
Source: USGS 2018, OpenStreetMap, Nextzen. https://www.latimes.com/projects/la-me-
woolsey-fire-progression/static/img/woolsey-fire-map-desktop.jpg

15
From February 2019 through June 2019, we surveyed Ventura County’s 32 Red-tailed hawk
nests that were recorded as active in 2018. These months were chosen because they encompass
when Red-tailed hawks nest and mate in this area (Cooper 2019). We surveyed nests during
selected periods from sunrise until 11:00 am and from 3:00 pm to sunset. These hours were
chosen because Red-tailed hawks are usually less active and visible during the hotter hours of
midday (Cooper 2019). We spent at least 30 minutes for each visit, waiting to see if a Red-tailed
hawk would appear at the nest site (Anderson et al. 1985). In 2019, each site was surveyed by
myself a minimum of three times over the course of the season; in 2018, a minimum of two visits
were made by Cooper.
An active nest was categorized as a raptor in the nest or on the branch containing the nest. We
relied on raptor behavior as an initial indicator of active nest sites. Territorial behavior included
raptors flying erratically close to us, startled or evasive behavior, or breeding behavior such as
copulation or tandem flights. We also listened for Red-tailed hawk alarm calls and watched for
behavioral displays (e.g., puffing up of feathers in order to appear larger) which served as
indications that nests were in close vicinity. We scanned the area within a 200-meter radius of
each nest site for raptors. For each nest (historical and current), the tree species, raptor species,
and GPS coordinates were recorded. We used a u-Blox M8 GNSS receiver with a ceramic patch
antenna to record the position data. We used the GPS coordinates of historic nest sites and
assessed visually, using Nikon Prostaff 3S Binoculars, whether the nest was still present. If
present, the nest activity was marked as positive, and tree species was recorded.
We visually characterized sampling sites as no burn, lightly burned, moderately burned, and
heavily burned (Table 1). To assess sampling sites, the amount of surrounding vegetation was
visibly scanned in a 200 m radius of the nest (distances were measured using a Calculated

16
Industries measuring wheel). This 200 m radius yields an area of approximately 125,664 m2.
This area was broken into 30 m2 segments. If any of these segments had visibly burned soil or
vegetation, it was counted as a burned segment. The number of burned segments within each
study area was counted and divided by the total number of segments in the area. This resulted in
the burn percentage values that inform the burn categories (Table 1). This methodology was
developed based on a similar measure of categorical characteristics used in forest management
(Leuschner 1990).
Table 1. Types of burn areas defined via percentage of surface area charred within 200 m radius
of nesting site
Burn Types
Percent Burned (per
30m2)
No burn 0%
Lightly burned 1-25%
Moderately burned 26-49%
Heavily burned >50%
Data Analysis: All data were analyzed and graphed using MATLAB numerical computing
software. We first used the total number of nests for each as a gross metric to compare the nest
distribution of the raptors before and after the Woolsey fire. This value presents a succinct
summary of how the fire affected the nesting habits.

17
We hypothesized that the fire, and not pure chance, had altered the nest persistence in Ventura
County. We predicted that there would be significantly fewer nests post-fire than pre-fire. After
collecting the data, we calculated how many active nests in 2018 were still active in 2019. We
then used a Wilcoxon signed-rank test to test our hypothesis and evaluate if the change in nest
persistence was statistically significant. Statistical significance in this context means being sure
that the change in nest persistence was due to the fire, and not due to random chance.
Our hypothesis was that the predicted decrease in active nests would also change the nest
positioning in Ventura County. We first calculated a central nesting coordinate, which we
defined simply as the arithmetic average latitude and longitude position of all the active nests for
each year. We then calculated the average distance of all the nests from the central nesting
coordinate of all other nests. The distance between each nest and the center location was found
by solving the indirect geodetic problem using an iterative error reduction method (Sjöberg,
2006). This algorithm finds the curvilinear distance between two points on Earth’s surface using
their latitude and longitude coordinates. It assumes a WGS-84 ellipsoidal Earth coordinate
system and finds the arc length along the Earth’s surface by projecting the geodetic coordinates
onto a two-dimensional plane. This curvilinear distance was found for each nest and then
averaged for the set of nests pre-Woolsey fire and post-Woolsey fire, respectively. This metric is
significant, as the larger the average distance away from the central location, the more spread-out
the nests are.
While average distance from the reference location characterizes the gross size of the nesting
territory, it does not fully illustrate the distribution of nests within this environment. This
distribution is important to quantify, as it shows how evenly spread the nests are within the area.
Thus, a second metric developed was the standard deviation of the distances of each nest from

18
the central location. This metric illustrates how spread-out the nests are from the average
distance, effectively showing how spread out the nests are from each other. The higher the
standard deviation of distances from the central reference coordinate, the fewer “clumps” of
nests exist and more evenly spread the nests are within the studied area. A lower standard
deviation indicates that the nests are all within a similar distance from the central location.
Nest locations were plotted on a map overlaid with the boundary of the Woolsey fire (Woolsey
Fire GIS Data & Applications 2019). The boundary was downloaded as a shape file, opened
using ArcGIS software, and converted to data points using Python. This allowed it to be plotted
alongside the collected raptor data, using MATLAB graphing software.
In addition to simply plotting the GPS coordinates within the Woolsey fire perimeter, nests were
categorized as being either in an edge zone or outside an edge zone. An edge zone is defined as a
change in community structure that occurs at a boundary of two or more habitats (Levin 2009).
To determine which nests were in the edge zone, the distance between the fire border and each
nest was calculated. Any nest within a territorial radius of the border was classified as within the
edge zone. Red-tailed hawk territory is usually around 2.33 ± 0.09 km2 (Janes, 1984). This
territory is approximated as a circle whose center is the nest. The distance from the nest to the
edge of the territory is a distance we have termed the territorial radius, as it is the distance from
the center of the circle to its circumference. Given the area of the circle is equal to 𝜋𝑟2, where r
is the radius, we can determine the average territorial radius to be around 861 ± 17 m. To be
conservative, the territorial radius used in the calculation was 844 m. We tested the change in
proportion of nests in each burn severity zone by using a chi-square test with Yates continuity
correction to compare the 2018 active nests and 2019 nests. We used a Yates continuity

19
correction in our chi-square test to account for the sample size lower than 40 (Beukelman, et al,
2016).
In addition to studying the geographic locations and level of burn of the nests, we identified the
species of tree in which the nests were made. Tree species is a key component of the nest’s
microenvironment. Different tree species have different leaf types, growth rates, and leaf cover.
These properties may have contributed to the nests’ persistence. For this study, we compared the
composition of trees in which the Red-tailed hawks nested pre-fire to the composition of trees
used post-fire. To determine whether or not tree species is correlated with nest survival, a chi-
squared test was performed on the tree data. The six tree species were the independent
categorical data in the chi-squared test, with the binary option of persistence or loss being the
dependent variable. In addition, we used a Yates continuity correction in our chi-square test to
account for the sample size lower than 40 (Beukelman, et al, 2016).
RESULTS
We surveyed 32 Red-tailed hawk nests within the study area in 2019 and compared it to data
from 2018. Many of the active nests in 2018 were gone in 2019. In addition to seeing how many
active 2018 nests were still active in 2019, we wanted to see if the persistence of nests after the
Woolsey fire was affected by the fire itself. In any population of active nests, some will be
destroyed or go into disrepair over the course of a year. Such standard nest degradation may be
caused by stochastic events over the course of the year that may affect any number of nests.
Our initial analysis quantified how many of the active 2018 nests remained active in 2019. Of the
original 32 nests surveyed, only 12 persisted into 2019. A Wilcoxon signed-rank test comparing
the 2018 and 2019 nest data yielded a test statistic W of 21, with a reduced Nr value of 20. The
critical test statistic value for a two-tailed significance level of 0.01 is 38. Since the test value is

20
lower than the critical value, we can say, with a 99% confidence level, that the degradation of
nests from 2018 to 2019 was not random but rather had one or more influential factors causing
the nests to become inactive. The pre-fire and post-fire nests (Table 2) were mapped (Figure 6),
along with the border of the Woolsey Fire.
Figure 6. Latitude and longitude (degrees) of Red-tailed hawk nests found pre-fire and nests that
persisted after the Woolsey Fire in Ventura County during raptor mating season overlaid with
Woolsey Fire boundaries. Blue colored dots indicate nests that were only found in 2018 and did
not persist post-fire, having been destroyed. Red colored dots indicate nests that were found in
both 2018 and 2019, persisting after the Woolsey Fire.

21
Of the persisting nests, 75% existed within an edge zone (within 844 m of burned areas). In
contrast, only 60% of the nests that perished existed within the edge zone.
In 2018, the average separation distance of all nests was 3.44 km with a standard deviation of
2.04 km. In 2019, the average separation distance was 5.04 km with a standard deviation of 1.70
km. As there were fewer nests in 2019, these results were expected.
Table 2. Percentage of nests found within each burn category
Used Pre-fire Used Post-fire
High burn 4 (12%) 0 (0%)
Moderately
burned 5 (16%) 1 (8%)
No burn 23 (72%) 11 (92%)
Total 32 (100%) 12 (100%)
Twelve (62%) of the original 32 nests persisted after the fire. For the nests that persisted post-
fire, 0 (0%) were found in the high burn areas, 1 (8%) were found in the moderately burned area,
and 11 (92%) were found in the no burn areas. We see that the proportions of nests in each zone
changed after the fire, as 4 (12%) of the nests active in 2018 were in high burn area, 5 (16%)
were in moderately burned areas, and 23 (72%) were in no burn areas. This change in
proportions can be confirmed with a chi-square test with a Yates continuity correction
(Beukelman, et al, 2016). The chi-square statistic with Yates correction (which accounts for the
small sample size and total lack of active nests in the post-fire and high burn category) was

22
found to be 14.98, which was well above the critical value for two degrees of freedom at 99%
confidence levels. This chi-square value, at two degrees of freedom, accounts for a p-value of
0.0005586. This means that the proportion of nests in each burn category changed significantly
pre-fire and post-fire.
Figure 8. Persistence of nests found in areas with no burn, moderately burned, and highly burned
areas.
In the high burned areas, 0 (0%) nests persisted and 4 (100%) did not persist post-fire. For the
moderately burned areas, 1 (20%) nests persisted and 4 (80%) did not persist post-fire. For the no
burn areas, 11 (48%) nests persisted and 12 (52%) did not persist post-fire.
The distribution of burned nests throughout Ventura County is illustrated in Figure 9.

23
Figure 9. Red-tailed hawk (RTHA) nest locations marked according to burn severity.
The above figure shows the location of nests that did not persist after the Woolsey fire and marks
them according to burn severity (Figure 9). To further delineate how both persisting nests and
non-persisting nests were distributed according to burn severity, the following figures map the
burn level of non-persisting nests and persisting nests, respectively (Figure 10, Figure 11).

24
Figure 10. Burn levels of areas surrounding non-persisting nests.
The figure shows the location of nests that persisted after the Woolsey fire according to burn
severity (Figure 10).
The high burned nests were completely destroyed, where the nest was no longer found at the
GPS coordinates. The nests in the moderate burn and the no burn were still found at their
previous GPS coordinates but were no longer in use.

25
Figure 11. Burn levels of persisting nests.
The figure shows the location of nests that persisted after the Woolsey Fire and their
corresponding burn levels (Figure 11).

26
The figure shows the number of nests that persisted after the Woolsey fire for each tree species
(Figure 12).
Figure 12. Number of pre-Woolsey fire nests that persisted post-Woolsey fire sorted by tree
species.

27
The null hypothesis for studying the connection between tree type and the survival of the nest in
the tree was that there is no correlation. To test whether tree type did play a part in determining
nest survival, a chi-squared test was performed to evaluate the data.
The chi-squared value of the trees, even without a Yates continuity correction, was 2.15, with 5
degrees of freedom. This yields a p-value of 0.8274. With Yates continuity correction (which
accounts for the low number of samples), the chi-square value of 0.4001, yielding a p-value of
0.99. Thus, the composition of trees that had active nests in 2018 was around the same
composition of trees that had active nests in 2019 (after the Woolsey fire).
DISCUSSION
Wildfire activity in California has greatly increased, particularly in western U.S. forests
(Westerling et al. 2006, Figure 1, Figure 2, Figure 3). The trends of longer fire seasons with
increased economic impact are projected to continue under plausible climate change scenarios,
making it a priority to study how wildfires affect various species (National Assessment Synthesis
Team 2000, Houghten et al. 2001, Running 2006). Fire has a myriad of effects on the
environment including soil, water quality, microbial communities, and vegetative dynamics.
With more data and analysis, government agencies and NGOs can potentially create more
comprehensive plans for land management. Although Red-tailed hawks are known to be
attracted to fire due to the flushing of prey, there have been few longitudinal studies on how
populations of Red-tailed hawks change post-fire (Stoddard 1963). It would be prudent to study
their long-term nesting success post-fire because Red-tailed hawks can serve as an indicator of
ecosystem health (Dmowski 1999, Therrien et al. 2012). Historically raptors were used as

28
indicators of heavy metal pollution, since bioaccumulation occurred with their prey (Dmowski
1999). Other studies have also used raptor species to serve as indicators because top predators
are sensitive to ecosystem changes, therefore changes in their numbers may reflect changes in
environmental conditions (Therrien et al. 2012). Because of their status as useful indicators,
keeping Red-tailed hawk population density stable and understanding how wildfires affect their
nesting habits is of broad biological importance.
Multiple variables were considered in analyzing the effect of the Woolsey fire on the Red-tailed
hawk nests in Ventura county. We hypothesized that tree type, geographic position, and relative
position to the fire’s edge zone could influence nest persistence rates. To test this hypothesis,
several statistical tools were employed.
The six different tree types which housed the nests all have different population sizes and
distributions within Ventura County. All six are dry climate trees, which grow well in Southern
California (Trees for Ventura CA 2018, Pine Trees of California 2016, California Native Plant
Society 2007, Rowland et al. 2019). The relative frequencies and population sizes of these tree
species allowed us to use a chi-squared test to determine if there is any association between tree
species and nest persistence. From 2018 to 2019, there was no statistically meaningful change in
tree composition used by the Red-tailed hawk for nests. That means the tree composition
probably had little to no effect on nest persistence.
While tree composition did not change after the Woolsey fire, the proportion of nests found in
each burn severity zone did change significantly. The higher the burn type, the more likely a nest
was to be lost post-fire. This was likely due to the fact that all of the nests in high burn zones
were completely destroyed. Nonetheless, even areas with no burn still lost nests. Of the nests
located in no-burn areas, 11 (48%) became inactive (Figure 8, Figure 9, Figure 10). Furthermore,

29
the percentage of nests lost was largest in the non-burned area (Figure 8, Figure 9, Figure 10).
Since even unburned territory lost a large number of nests, fire can cause nests to be lost through
means other than simply razing territory.
In 2018, most raptor nests found were located inside of the zone burned by the Woolsey fire
(Figure 9). Note that the GIS data only cataloged the perimeter of the fire without accounting for
the intensity of the burn sites. Of the nests that did not persist post-fire, most were within the
Woolsey Fire zone, with only two exceptions (Figure 10). Nine of the nests that persisted post-
fire were on the edge of the fire zone, thus indicating nest position is a significant factor in
determining its persistence. There could be elements of edge habitat and habitat mosaics that
affected nesting decisions that can be further studied. Other raptor species were found to prefer
edge habitats and distribute accordingly (Sanchez et al. 1999).
Raptors are known to be attracted to recently burned areas due to lack of cover for their prey
species (Ran et al. 2005). Smaller species, such as rabbits and mice, increase in density after a
fire due to increased food resources (Tietje et al. 2008, Converse et al. 2006). While species such
as these initially decrease in population size, they accumulate in early seral areas and rapidly
reproduce (Converse 2006). These smaller species make up much of the Red-tailed hawks’ prey,
the post-fire environment may provide greater food availability (Barro and Susan 1991, Lyon et
al. 2000). Since food resources are not a limiting factor in nest persistence, other factors could
play an important role.
The results of this study suggest that high and moderately burned areas will likely cause the nests
to not persist due to viable nest and territory loss or the complete destruction of the actual nests.
In addition, even non-burned areas within the fire’s borders can be lost. Although recently
burned areas attract the raptors, the Red-tailed hawks are still not nesting the following year post-

30
fire. One variable that might explain the variation in persistence of nests in non-burned areas was
the location relative to the fire’s border. This geographic relationship is essential in
understanding Red-tailed hawks’ nesting behavior.
The higher rates of nest persistence at the edge of the Woolsey fire can be illustrated by studying
the layout of the nests relative to each other throughout the study area. In 2018, the nests were
densely clustered on average but had a higher standard deviated distance from the central nesting
location (defined as the average latitudinal and longitudinal position of all the active nests). This
low distance from the average nest location indicates that the nests were close together on
average but were well dispersed throughout Ventura County, as indicated by the higher standard
deviation. In 2019, the nests were more spread out and were on average further from the central
nesting location, with fewer nests going towards the center point. Thus, while the 2019 active
nests were on average further from the average nesting spot, their average distance from the
center was around the same for each nest. This indicates that 2019 nests were active more on the
outskirts of the nesting territory, while 2018 nests were active throughout the nesting territory.
Thus, post-fire nests mostly existed at the outskirts of the fire, in the edge zone area.
One reason why nests persisted more on the edges of the Woolsey Fire may be due to urban Red-
tailed hawks’ availability for greater land cover diversity and patch richness (Stout et al. 2006).
Nests bordering the Woolsey fire’s area of impact would likely have more land cover diversity
due to the fire. Red-tailed hawks are known to nest in open woodlots or edges; the fire helped to
create more edges within the environment, making it a more preferred nesting site (Howell,
James, et al. 1978).
Previous studies have concluded that the Red-tailed hawk species should be classified as a
woodland edge-species, where there is an ecotone between a forest and open land (Bock and

31
Lepthien 1976). Furthermore, Bock and Lepthien (1976) classified Red-tailed hawks as open
land species, which are animals whose preferred habitat would have less understory and cover.
The early seral stages created by the Woolsey fire became the ecotone for Red-tailed hawks.
Results from our study indicate that the Red-tailed hawk species fared better near the fire’s edge,
with 75% of the persisting nests remaining active in the edge habitat of the Woolsey Fire. The
edge habitat is the part of the ecosystem where the non-burned ecosystem met the burned
ecosystem. Edge effects create highly productive areas that are beneficial to wildlife. The
ecotone shares characteristics of both communities which usually creates more species richness
and abundance (Kark 2007). The increase in productivity could explain why the Red-tailed
hawks stayed at their historical nest site if their nest occupied an edge habitat. These birds of
prey are generalists who could take advantage of increased species abundance. Since generalist
species can thrive in wide varieties of environmental conditions, the generalists’ habitats would
be those with high population numbers and species of prey items. Another study concluded
similar habitat preferences in Red-tailed hawks where prey availability is highly correlated with
habitat quality (Janes 1984).
The higher than average persistence of Red-tailed hawk nests that lie within the edge zone,
especially among no burned nests, indicates two key outcomes. First, Red-tailed hawks can
effectively thrive in territories with a varied rate of succession habitat. Second, even nests in
microenvironments that are locally left unburned will likely degrade if the macro-environment is
within the fire’s border.
The poorer habitat quality of non-edge zones could explain why so many of the nests were
abandoned post-fire. The mated pairs could likely have abandoned the nests due to their poorer
quality and relocated to higher quality habitats. Similar studies concluded that their Red-tailed

32
hawks nested at or near the edge of two habitats (Moorman and Bryan 1996). Moorman and
Bryan (1996) also concluded that this species nested closer to forest openings. The nests near the
Woolsey fire’s edge may have persisted thanks to the creation of early seral stage open land
within the fire’s border next to unburned forested areas.
With the increase of fire disturbances and fire intensity in California, Red-tailed hawk survival
could be negatively impacted due to their average nesting success of 80.1% and 1.36 young per
laying pair (Stout et al. 2006). Their slow population growth rate would mean it would take
longer for their populations to rise back to previous levels if there were to be more intense fires.
Fire disturbance from wildfire has a history of altering California’s ecosystems. There has been a
steady increase in number of wildfires, intensity of wildfires, and duration time of wildfires due
to climate change (Union of Concerned Scientists 2013). Due to the increasing effects that fire
disturbances have on California environments, it is important to study the complexity of
feedbacks and quantify both positive and negative influences it can have on the flora and fauna.
The change in fire regimes could have unforeseen consequences on the apex predators and the
disturbance processes (Hovick et al. 2017). This research helps to quantify the effects that post-
wildfire disturbances can have on Red-tailed hawks nesting habits.
Acknowledgements: Special thanks to Lena Lee for providing her previous works to the project.
Big thanks to Dr. Michael Wing and Dr. Susie Dunham with reviewing and assistance in the
project and to Dr. Blaine Vogt for reviewing the project.

33
LITERATURE CITED
Andersen, D. E., Rongstad, O. J., & Mytton, W. R. (1985). Line transect analysis of raptor
abundance along roads. Wildlife Society Bulletin (1973-2006), 13(4), 533-539.
Arno, S. F. (2000). Fire in western forest ecosystems. Wildland fire in ecosystems. Fort Collins,
CO: Rocky Mountain Research Station. USDA Forest Service General Technical Report
RMRS-GTR-42–volume, 2.
Ballam, J. M. (1984). The use of soaring by the Red-tailed Hawk (Buteo jamaicensis). The Auk,
101(3), 519-524.
Barton, A. M. (1999). Pines versus oaks: effects of fire on the composition of Madrean forests in
Arizona. Forest Ecology and Management, 120(1-3), 143-156.
Barro, S. C., & Conard, S. G. (1991). Fire effects on California chaparral systems: an overview.
Environment International, 17(2-3), 135-149.
Bednarz, J. C., & Dinsmore, J. J. (1982). Nest-sites and habitat of Red-shouldered and Red-tailed
hawks in Iowa. The Wilson Bulletin, 31-45.
Beukelman, T., & Brunner, H. I. (2016). Trial Design, Measurement, and Analysis of Clinical
Investigations. In Textbook of pediatric rheumatology (pp. 54-77). WB Saunders.

34
BirdLife International. (2016). Buteo jamaicensis. The IUCN Red List of Threatened Species
2016: e.T22695933A93534834. https://dx.doi.org/10.2305/IUCN.UK.2016-
3.RLTS.T22695933A93534834.en
Boal, C. W., & Mannan, R. W. (1998). Nest-site selection by Cooper's Hawks in an urban
environment. The Journal of wildlife management, 864-871.
Bock, C. E., & Lepthien, L. W. (1976). Geographical ecology of the common species of Buteo
and Parabuteo wintering in North America. The Condor, 78(4), 554-557.
Bonta, M., Gosford, R., Eussen, D., Ferguson, N., Loveless, E., & Witwer, M. (2017).
Intentional fire-spreading by “Firehawk” raptors in Northern Australia. Journal of
Ethnobiology, 37(4), 700-718.
Bosakowski, T. H. O. M. A. S., & Smith, D. G. (1997). Distribution and species richness of a
forest raptor community in relation to urbanization. Journal of Raptor Research, 31, 26-
33.
Bowman, D. M., & Murphy, B. P. (2010). Fire and biodiversity. Conservation Biology for All,
163.
Brack Jr, V., Cable, T. T., & Driscoll, D. E. (1984, January). Food Habits of Urban Amerian
kestrels, Falco sparverius. In Proceedings of the Indiana Academy of Science (Vol. 94,
pp. 607-614).

35
Brown, J. K., & Smith, J. K. (2000). Wildland fire in ecosystems: effects of fire on flora. Gen.
Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: US Department of Agriculture, Forest
Service, Rocky Mountain Research Station. 257 p., 42.
California Department of Forestry and Fire Protection. (2020). “Cal Fire Stats and Events.”
Organization Image, www.fire.ca.gov/stats-events/.
Cheney, N. P., & Gould, J. S. (1997). Fire growth and acceleration. International Journal of
Wildland Fire, 7(1), 1-5.
Chiang, S. N., Bloom, P. H., Bartuszevige, A. M., & Thomas, S. E. (2012). Home range and
habitat use of Cooper’s Hawks in urban and natural areas. Studies in Avian Biology, 45,
1-16.
Converse, S. J., White, G. C., Farris, K. L., & Zack, S. (2006). Small mammals and forest fuel
reduction: national‐scale responses to fire and fire surrogates. Ecological Applications,
16(5), 1717-1729.
Cooper, D. (2019). Lecture presented in Ventura County. Personal Communication.
Crooks, K. R., Suarez, A. V., & Bolger, D. T. (2004). Avian assemblages along a gradient of
urbanization in a highly fragmented landscape. Biological conservation, 115(3), 451-462.

36
Dmowski, K. (1999). Birds as bioindicators of heavy metal pollution: review and examples
concerning European species. ACTA ORNITHOLOGICA-POLSKA AKADEMIA NAUK-
ORIGINAL EDITION-, 34, 1-26.
Ellis, D. H., Craig, T., Craig, E., Postupalsky, S., LaRue, C. T., Nelson, R. W., ... & Dawson, J.
W. (2009). Unusual raptor nests around the world. Journal of Raptor Research, 43(3),
175-198.
Ferguson-Lees, J.; Christie, D. (2001). Raptors of the World. London: Christopher Helm.
ISBN 978-0-7136-8026-3.
“Fremont Cottonwood, Populus Fremontii.” (2017). California Native Plant Society,
calscape.org/Populus-fremontii-(Fremont-Cottonwood).
Henneman, Carlene. Habitat associations of red-shouldered hawks in central Minnesota
landscapes. (2006). Diss. University of Minnesota Major: Wildlife conservation.
“Home > Trees for Ventura,CA.” (2008). California Native Plant Society, calscape.org/loc-
Ventura,CA/cat-Trees/ord-popular.
Holland, R. F. (1986). Preliminary descriptions of the terrestrial natural communities of
California. California Department of Fish and Game. Unpublished report. Sacramento, CA.
Hovick, T. J., McGranahan, D. A., Elmore, R. D., Weir, J. R., & Fuhlendorf, S. D. (2017). Pyric‐
carnivory: Raptor use of prescribed fires. Ecology and Evolution, 7(21), 9144-9150.

37
Howell, J., Smith, B., Holt Jr, J. B., & Osborne, D. R. (1978). Habitat structure and productivity
in red-tailed hawks. Bird-banding, 162-171.
Houghton JT et al (Eds) (2001) IPCC climate change: the scientific basis. Cambridge University
Press,Cambridge, United Kingdom and New York, NY, USA
Hurteau, M. D., Westerling, A. L., Wiedinmyer, C., & Bryant, B. P. (2014). Projected effects of
climate and development on California wildfire emissions through 2100. Environmental
science & technology, 48(4), 2298-2304.
Hutto, R. L., & Patterson, D. A. (2016). Positive effects of fire on birds may appear only under
narrow combinations of fire severity and time-since-fire. International Journal of
Wildland Fire, 25(10), 1074-1085.
Kark, Salit. (2007). Effects of Ecotones on Biodiversity. 10.1016/B978-012226865-6/00573-0.
Keane, R. E. (2002). Cascading effects of fire exclusion in Rocky Mountain ecosystems: a
literature review. US Department of Agriculture, Forest Service, Rocky Mountain
Research Station.
Keane, R. E., Agee, J. K., Fulé, P., Keeley, J. E., Key, C., Kitchen, S. G., ... & Schulte, L. A.
(2009). Ecological effects of large fires on US landscapes: benefit or catastrophe? A.
International Journal of Wildland Fire, 17(6), 696-712.

38
Keeley, J. E., & Fotheringham, C. J. (2001). Historic fire regime in southern California
shrublands. Conservation Biology, 15(6), 1536-1548.
Kern, C. C., Montgomery, R. A., Reich, P. B., & Strong, T. F. (2013). Canopy gap size
influences niche partitioning of the ground-layer plant community in a northern temperate
forest. Journal of Plant Ecology, 6(1), 101-112.
Los Angeles County “Woolsey Fire Incident Update.” Los Angeles County Fire Department,
www.fire.lacounty.gov/woolsey-fire-incident/.
Smith, J. K., & Lyon, L. J. (2000). Wildland fire in ecosystems: effects of fire on fauna (Vol. 2).
US Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Janes, S. W. (1984). Influences of territory composition and interspecific competition on Red‐
tailed Hawk reproductive success. Ecology, 65(3), 862-870.
“Large Nest Identification.” (2020). US Fish and Wildlife Service,
www.fws.gov/midwest/eagle/Nhistory/nest_id.html.
Lee, L. (2004). [Raptor nest survey]. Unpublished raw data.
Levin, Simon A. (2009). The Princeton Guide to Ecology. Princeton University Press. p. 780.
Leuschner, W. A. (1990). Forest regulation, harvest scheduling, and planning techniques. John
Wiley & Sons, Inc.

39
Luniak, M. (2004, July). Synurbization–adaptation of animal wildlife to urban development. In
Proceedings 4th international urban wildlife symposium (pp. 50-55). University of
Arizona,.
MacLaren, P. A., Anderson, S. H., & Runde, D. E. (1988). Food habits and nest characteristics of
breeding raptors in southwestern Wyoming. The Great Basin Naturalist, 548-553.
Mannan, R. W., Steidl, R. J., & Boal, C. W. (2008). Identifying habitat sinks: a case study of
Cooper’s hawks in an urban environment. Urban Ecosystems, 11(2), 141-148.
Marti, C. D., Korpimäki, E., & Jaksić, F. M. (1993). Trophic structure of raptor communities: a
three-continent comparison and synthesis. In Current ornithology (pp. 47-137). Springer,
Boston, MA.
Minor, W. F., Minor, M., & Ingraldi, M. F. (1993). Nesting of Red-Tailed Hawks and Great
Horned Owls in a Central New York Urban/Suburban Area (Anidamiento de Buteo
jamaicensis y de Bubo virginianus en un area urbana/suburbana de la parte central de
New York). Journal of Field Ornithology, 433-439.
Monamy, V., & Fox, B. J. (2000). Small mammal succession is determined by vegetation density
rather than time elapsed since disturbance. Austral Ecology, 25(6), 580-587.
Moorman, C. E., & Chapman, B. R. (1996). Nest-site selection of red-shouldered and red-tailed
hawks in a managed forest. The Wilson Bulletin, 357-368.

40
National Assessment Synthesis Team (2000) Climate change impacts on the United States: the
potentialconsequences of climate variability and change. US GCRP, Washington DC
“National Centers for Environmental Information.” VENTURA, CALIFORNIA - Climate
Summary, 2010, wrcc.dri.edu/cgi-bin/cliMAIN.pl?ca9285.
Navarro-Lopez, J., & Fargallo, J. A. (2015). Trophic niche in a raptor species: the relationship
between diet diversity, habitat diversity and territory quality. PloS one, 10(6), e0128855.
NPS. (2018) Woolsey Fire. Retrieved from https://www.nps.gov/samo/learn/management/2018-
woolsey-fire.htm
Palmer, R. S., ed. (1988). Handbook of North American birds. Volume 5 Diurnal Raptors (part
2).
“Pine Trees of California.” (2016). California Native Pine Trees, Pinus,
www.treesforme.com/ca_pinus.html.
Poirazidis, K., Goutner, V., Tsachalidis, E., & Kati, V. (2007). Comparison of nest-site selection
patterns of different sympatric raptor species as a tool for their conservation. Animal
Biodiversity and Conservation, 30(2), 131-145.
Preston, Charles R. (2000). Red-tailed hawk. Stackpole Books.
Preston, C. R. and R. D. Beane. (2009). "Red-tailed Hawk (Buteo jamaicensis), The Birds of
North America.

41
Ran, S. W., Lee, J. W., Paek, W. K., Lee, H. S., Kim, I. K., Hong, G. P., ... & Paek, I. R. (2005).
The effect of forest fire on the raptor habitation. Korean Journal of Environment and
Ecology, 19(4), 385-392.
“Red-Tailed Hawk Identification, All About Birds, Cornell Lab of Ornithology.”, All About
Birds, Cornell Lab of Ornithology, www.allaboutbirds.org/guide/Red-tailed_Hawk/id.
Rottenborn, S. C. (2000). Nest-site selection and reproductive success of urban Red-shouldered
Hawks in central California. Journal of Raptor Research, 34(1), 18-25.
Rowland, Teisha, et al. (2019, March 29). How the eucalyptus came to California. The Santa
Barbara Independent. https://www.independent.com/2011/01/15/how-eucalyptus-came-
california
Rullman, S., & Marzluff, J. M. (2014). Raptor presence along an urban–wildland gradient:
Influences of prey abundance and land cover. Journal of Raptor Research, 48(3), 257-
272.
Running SW (2006) Is global warming causing more, larger wildfires? Science 313:927–928
Sahores, M., & Trejo, A. (2004). Diet shift of Barn Owls (Tyto alba) after natural fires in
Patagonia, Argentina. Journal of Raptor Research, 38, 174-177.
Sánchez‐Zapata, J. A., & Calvo, J. F. (1999). Raptor distribution in relation to landscape
composition in semi‐arid Mediterranean habitats. Journal of Applied Ecology, 36(2), 254-
262.

42
"San Francisco Downtown, California". National Climatic Data Center (NCDC) 1981-2010
Monthly Normals. Western Regional Climate Center (WRCC), National Oceanic and
Atmospheric Administration (NOAA).
Schmutz, J. K., Schmutz, S. M., & Boag, D. A. (1980). "Coexistence of three species of hawks
(Buteo spp.) in the prairie–parkland ecotone". Canadian Journal of Zoology. 58 (6):
1075–1089.
Sjöberg, L. E. (2006). New solutions to the direct and indirect geodetic problems on the
ellipsoid. ZfV-Zeitschrift für Geodäsie, Geoinformation und Landmanagement, (zfv
1/2006).
Steenhof, K., Kochert, M. N., Carpenter, L. B., & Lehman, R. N. (1999). Long-term prairie
falcon population changes in relation to prey abundance, weather, land uses, and habitat
conditions. The Condor, 101(1), 28-41.
Stoddard, H. L. (1963, March). Bird habitat and fire. In Proceedings of the Tall Timbers Fire
Ecology Conference (Vol. 2, pp. 163-175).
Stotz, N. (1994). "Unusual nesting attempt by Red-tailed Hawk". Kingbird. 44 (3): 818–820.
Stout, W. E., Temple, S. A., & Papp, J. M. (2006). Landscape Correlates of Reproductive
Success for an Urban‐Suburban Red‐Tailed Hawk Population. The Journal of wildlife
management, 70(4), 989-997.

43
Stralberg, D. (1999). A landscape-level analysis of urbanization influence and spatial structure
in chaparral breeding birds of the Santa Monica Mountains, CA (Doctoral dissertation).
Swenson, J. J., & Franklin, J. (2000). The effects of future urban development on habitat
fragmentation in the Santa Monica Mountains. Landscape Ecology, 15(8), 713-730.
Syphard, A. D., Radeloff, V. C., Keeley, J. E., Hawbaker, T. J., Clayton, M. K., Stewart, S. I., &
Hammer, R. B. (2007). Human influence on California fire regimes. Ecological
applications, 17(5), 1388-1402.
Therrien, J. F., Goodrich, L. J., Barber, D. R., & Bildstein, K. L. (2012). A long‐term database
on raptor migration at Hawk Mountain Sanctuary, northeastern United States: Ecological
Archives E093‐174. Ecology, 93(8), 1979-1979.
Tietje, W., Vreeland, J., & Weitkamp, W. (2001). Live oak saplings survive prescribed fire and
sprout. California agriculture, 55(2), 18-22.
Tietje, W. D., Lee, D. E., & Vreeland, J. K. (2008). Survival and abundance of three species of
mice in relation to density of shrubs and prescribed fire in understory of an oak woodland
in California. The Southwestern Naturalist, 53(3), 357-369.

44
Union of concerned Scientists. (2013) Infographic: Western Wildfires and Climate Change.
Retrieved from https://www.ucsusa.org/global-warming/science-and-
impacts/impacts/infographic-wildfires-climate-change.html
Ventura County topographic map, elevation, relief. (2010). Retrieved from https://en-
gb.topographic-map.com/maps/zzgt/Ventura-County/
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006). Increases in Western US forest
wildfireassociated with warming and advances in the timing of spring. Science 313:940–
943
Williams, A. P., Abatzoglou, J. T., Gershunov, A., Guzman‐Morales, J., Bishop, D. A., Balch, J.
K., & Lettenmaier, D. P. (2019). Observed impacts of anthropogenic climate change on
wildfire in California. Earth's Future, 7(8), 892-910.
Woolsey Fire GIS Data & Applications. (2019, January 29). Retrieved from
https://egis3.lacounty.gov/dataportal/2018/11/21/woolsey-fire-nov-2018-gis-data-
applications/
WRCC. (n.d.). Western Regional Climate Center. Retrieved from
https://wrcc.dri.edu/Climate/narrative_ca.php