j. lipid res.-2015-sahu-osen-109-21
TRANSCRIPT

This article is available online at http://www.jlr.org Journal of Lipid Research Volume 56, 2015 109
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
Supplementary key words adipocytes • adipose tissue • adipose tri-glyceride lipase • Chanarin Dorfman syndrome • lipase • lipid drop-lets • lipolysis • perilipin
The control of lipolysis in adipocytes of vertebrates is a carefully orchestrated process. Lipolysis, or the hy-drolysis of stored triacylglycerols, releases fatty acids, monoacylg lycerols, and diacylglycerols that serve as sub-strates for energy production and the synthesis of phospho-lipids used for membrane synthesis and repair. The enzymes that catalyze triacylglycerol hydrolysis in adipocytes include adipose triglyceride lipase (ATGL), hormone-sensitive li-pase, and monoacylglycerol lipase, to cleave the fi rst, sec-ond, and third ester bonds, respectively ( 1–4 ). Adipose lipolysis is initiated by a variety of signaling cascades and the ensuing hydrolytic reactions are fi ne-tuned by an assort-ment of cytosolic and lipid droplet-associated proteins.
The best-characterized signaling pathway that initiates adipose lipolysis occurs when catecholamines bind to � -adrenergic receptors on adipocyte plasma membranes. Hormone binding triggers a G protein-mediated cascade that activates adenylyl cyclase, increasing levels of cAMP, in
Abstract CGI-58/ABHD5 coactivates adipose triglyceride lipase (ATGL). In adipocytes, CGI-58 binds to perilipin 1A on lipid droplets under basal conditions, preventing interac-tion with ATGL. Upon activation of protein kinase A (PKA), perilipin 1A is phosphorylated and CGI-58 rapidly disperses into the cytoplasm, enabling lipase coactivation. Because the amino acid sequence of murine CGI-58 has a predicted PKA consensus sequence of RKYS 239 S 240 , we hypothesized that phosphorylation of CGI-58 is involved in this process. We show that Ser239 of murine CGI-58 is a substrate for PKA using phosphoamino acid analysis, MS, and immuno-blotting approaches to study phosphorylation of recombi-nant CGI-58 and endogenous CGI-58 of adipose tissue. Phosphorylation of CGI-58 neither increased nor impaired coactivation of ATGL in vitro. Moreover, Ser239 was not re-quired for CGI-58 function to increase triacylglycerol turn-over in human neutral lipid storage disorder fi broblasts that lack endogenous CGI-58. Both CGI-58 and S239A/S240A-mutated CGI-58 localized to perilipin 1A-coated lipid drop-lets in cells. When PKA was activated, WT CGI-58 dispersed into the cytoplasm, whereas substantial S239A/S240A-mutated CGI-58 remained on lipid droplets. Perilipin phosphory-lation also contributed to CGI-58 dispersion. PKA-mediated phosphorylation of CGI-58 is required for dispersion of CGI-58 from perilipin 1A-coated lipid droplets, thereby increas-ing CGI-58 availability for ATGL coactivation. —Sahu-Osen, A., G. Montero-Moran, M. Schittmayer, K. Fritz, A. Dinh, Y-F. Chang, D. McMahon, A. Boeszoermenyi, I. Cornaciu, D. Russell, M. Oberer, G. M. Carman, R. Birner-Gruenberger, and D. L. Brasaemle. CGI-58/ABHD5 is phosphorylated on Ser239 by protein kinase A: control of subcellular localiza-tion. J. Lipid Res. 2015. 56: 109–121.
This work was supported by National Institutes of Health Grants R01 DK054797 (to D.L.B.) and R01 GM050679 (to G.M.C.), Austrian Science Fund (FWF) Project P26074 and the doctoral school “DK Metabolic and Cardio-vascular Disease (W1226) (to R.B-G.), and FWF Project P22170, the SFB LIPOTOX (F30), and the doctoral school “DK Molecular Enzymology” (W901-B05) (to M.O.)
������� Author’s Choice —Final version full access. Manuscript received 22 September 2014 and in revised form 20 November 2014.
Published, JLR Papers in Press, November 24, 2014 DOI 10.1194/jlr.M055004
CGI-58/ABHD5 is phosphorylated on Ser239 by protein kinase A: control of subcellular localization
Anita Sahu-Osen , 1, * Gabriela Montero-Moran , 1,2,†,§ Matthias Schittmayer , * Katarina Fritz , * Anna Dinh , †,§ Yu-Fang Chang , †, ** Derek McMahon , †,§ Andras Boeszoermenyi , †† Irina Cornaciu , †† Deanna Russell , †,§ Monika Oberer , †† George M. Carman , †, ** Ruth Birner-Gruenberger , 3, * and Dawn L. Brasaemle 3,†,§
Research Unit Functional Proteomics and Metabolic Pathways, Institute of Pathology, Medical University of Graz, Graz, Austria A-8036, and Omics Center Graz,* BioTechMed-Graz, Graz, Austria A-8010; Rutgers Center for Lipid Research, † Departments of Nutritional Sciences § and Food Science,** Rutgers, The State University of New Jersey , New Brunswick, NJ 08901; and Institute of Molecular Biosciences, †† University of Graz , Graz, Austria A-8010
Abbreviations: ATGL, adipose triglyceride lipase; IBMX, isobutyl-methylxanthine; NLSD, neutral lipid storage disorder; PKA, protein kinase A or cAMP-dependent protein kinase; PVDF, polyvinylidene difl uoride; TCEP, Tris(2-carboxyethyl)phosphine; TEV, tobacco etch virus .
1 A. Sahu-Osen and G. Montero-Moran contributed equally to this work.
2 Present address of G. Montero-Moran: Centro de BioCiencias, Universidad Autonoma de San Luis Potosi, Km 14.5 Carretera San Luis Potosi, Matehuala, Ejido Palma de la Cruz, CP78321, Soledad de Graciano Sanchez, San Luis Potosi, Mexico.
3 To whom correspondence should be addressed. e-mail: [email protected] (D.L.B.); [email protected] (R.B-G.)
The online version of this article (available at http://www.jlr.org) contains supplementary data in the form of three fi gures.
��� Author's Choice
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

110 Journal of Lipid Research Volume 56, 2015
were purchased from Amersham Biosciences. Protein A-Sepha-rose was from GE Healthcare. All remaining chemicals, scintilla-tion counting supplies, and solvents were purchased from Fisher Scientifi c or Sigma.
Expression and purifi cation of recombinant mouse CGI-58, S239A-, S240A-, S239A/S240A-, S239D-, and S239E-mutated CGI-58
Preparation of mouse CGI-58 cDNA subcloned into the pET-28a(+) bacterial expression vector was described previously ( 19 ); recombinant CGI-58 included two tandem N-terminal 6-histidine fusion sequences. The cDNA for the S239A-, S240A-, and S239A/S240A-mutated variants of CGI-58 were prepared using PCR to produce two overlapping fragments from the template 12-His-CGI-58 cDNA; fragments were then joined by PCR for subcloning into the pET-28a(+) vector. The mutagenic oligonucleotide primers used to prepare the cDNA for the S239A-mutated variant of CGI-58 were: forward (5 ′ -AAGTACGCGTCTATGTTTGAA-GATGACAC-3 ′ ), and reverse (5 ′ -ACATAGACGCGTACTTCC-GCTTGAAATCAGG-3 ′ ) primers. Mutagenic oligonucleotide primers used to prepare the cDNA for the S240A-mutated variant of CGI-58 were: forward (5 ′ -AAGTACTCCGCAATGTTTGAA-GATGACACGG-3 ′ ) and reverse (5 ′ -AAACATGCGGAGTACTT-CCGCTTGAATCAGG-3 ′ ) primers. The cDNA for the S239A/SS240A-mutated variant of CGI-58 was similarly prepared. All mutations were confi rmed by DNA sequencing. The 12-His-tagged WT and mutated recombinant S239A, S240A, and S239A/S240A variants of CGI-58 were expressed and purifi ed from Esch-erichia coli cell extracts with nickel-nitrilotriacetic acid agarose, as described ( 19 ). All steps for protein purifi cation were performed at 4°C. Enzyme preparations were stored at � 20°C.
Additionally, the coding sequences of mouse CGI-58 and mouse ATGL cDNAs were amplifi ed as described ( 20 ) for sub-cloning into the pProEX HTb vector (Addgene, Cambridge, MA), from which the CGI-58 cDNA was subcloned into the His-pSumo vector (kindly provided by Dr. Christopher D. Lima, Sloan Kettering Institute) with a disrupted BamH1 cleavage site . The forward (5 ′ -CGAAGCAGAGAGCTCGAAAACCTGTATT TT-CAGG-3 ′ ) and reverse (5 ′ -GGAACCCTCGAGTCATCAGTCTACT-GTGTGGC-3 ′ ) oligonucleotide primers included endonuclease cleavage sites for subcloning and a 5 ′ tobacco etch virus (TEV) cleavage site from the pProEX HTb vector. cDNAs were ampli-fi ed with PCR using either the Phusion polymerase kit (New Eng-land Biolabs) or the FailSafe™ PCR system (Epicenter Biotech-nologies, Madison, WI). The S239D and S239E mutations were introduced into the mouse CGI-58 cDNA using the QuikChange ® site-directed mutagenesis kit (Stratagene) with forward (5 ′ -CCT-GATTTCAAGCGGAAGTACGACTCTATGTTTGAAGATGACA-C G-3 ′ ) and reverse (5 ′ -CGTGTCATCTTCAAACATAGAGTCGTA-CTTCCGCTTGAAA TCAGG-3 ′ ) mutagenic primers for S239D CGI-58 and forward (5 ′ -CCTGATTTCAAGCGGAAGTACGA-GTCTATGTTTGAAGATGACACG-3 ′ ) and reverse (5 ′ -CGT G-TC ATCTTCAAACAT AGACTCGTACTTCCGCTTGAAATCAGG-3 ′ ) mutagenic primers for S239E. The mouse ATGL cDNA encoding amino acids 1–288 was subcloned into His-pSumo with 5 ′ TEV site using forward (5 ′ -GCTATGGATCCATGTTCCCGAGGG-3 ′ ) and the reverse (5 ′ -GGCGCTCGAGTCATTTTTCGAACTGCGG-GTGGCTCCAATCCTCCTCT CCAGC-3 ′ ) oligonucleotide prim-ers including endonuclease cleavage sites and a stop codon fol-lowing the nucleotide sequence for D288, followed by the se-quence encoding a Strep-tag ® (IBA, Goettingen, Germany). Mutations were confi rmed by DNA sequencing (Integrated DNA Technologies, Coralville, IA and Agowa, Berlin, Germany).
His-pSumo-CGI-58, His-pSumo-ATGL 1-288, and the His-pSumo vector encoding 6-His-smt protein were transformed into BL21(DE3) RIPL CodonPlus E. coli (Agilent Technologies).
turn, activating protein kinase A (PKA). The sub sequent PKA-mediated phosphorylation of multiple proteins enables lipolysis. These proteins include, but are not limited to, perili-pin 1A and hormone-sensitive lipase ( 2, 5, 6 ).
Perilipin 1A at the surfaces of lipid droplets is a master regulator of adipose lipolysis. Under basal (fed) condi-tions, perilipin 1A provides a protective barrier against li-polysis of stored triacylglycerols ( 7 ) and binds CGI-58 (also called ABHD5) ( 8, 9 ), a coactivator of ATGL ( 10 ). When CGI-58 is sequestered on the perilipin scaffold, it cannot interact with or activate ATGL ( 11 ). When catecholamines stimulate adipose lipolysis, perilipin 1A is multiply phos-phorylated by PKA. The phosphorylation of three N-termi-nal serine residues of perilipin 1A facilitates the docking of PKA-phosphorylated hormone-sensitive lipase through a protein-protein interaction with the perilipin scaffold ( 12 ), thus bringing the lipase to its substrate lipids. The PKA-mediated phosphorylation of two carboxyl-terminal serine residues of perilipin 1A facilitates the release of CGI-58 from perilipin 1A, enabling interaction of CGI-58 with ATGL ( 11 ), in turn, activating the lipase ( 10 ).
CGI-58 was identifi ed as an important factor in cellular triacylglycerol homeostasis when mutations in CGI-58 were established as the cause of Chanarin Dorfman syn-drome ( 13 ), a neutral lipid storage disorder (NLSD) char-acterized by excessive storage of triacylglycerols in many cells and tissues ( 13–18 ). CGI-58 was later identifi ed as a coactivator of ATGL ( 10 ), thus explaining its role in triac-ylglycerol turnover; however, the mechanism by which CGI-58 activates ATGL has not yet been elucidated. In this study, we asked whether CGI-58 is a substrate for PKA. We demonstrate that CGI-58 is indeed a substrate for PKA, and show that its phosphorylation is important for the sub-cellular traffi cking of CGI-58 in adipocytes.
MATERIALS AND METHODS
Materials All chemicals were reagent grade. Growth media for cultured
cells were obtained from Mediatech, Inc. (Herndon, VA) or Sigma. FBS, isobutylmethylxanthine (IBMX), forskolin, and pro-tease and phosphatase inhibitor cocktails were purchased from Sigma. The DNA purifi cation kit and nickel-nitrilotriacetic acid agarose matrix were purchased from Qiagen. Coomassie Plus protein assay reagent was purchased from Thermo Scientifi c/Pierce; Bradford and DC protein assays were purchased from Bio-Rad. PfuUltra high-fi delity DNA polymerase was purchased from Stratagene, Inc. (La Jolla, CA). BODIPY 439/503, Alexa Fluor 546 goat-anti rabbit IgG, Alexa Fluor 546 donkey-anti goat IgG, and Alexa Fluor 488 donkey-anti rabbit IgG were purchased from Molecular Probes. Radioactive compounds were purchased from Perkin Elmer Biosciences. Oligonucleotide primers were purchased from Operon BioTechnologies, Inc. (Huntsville, AL). Phospholipids were purchased from Avanti Polar Lipids, Inc. (Alabaster, AL) and Sigma. Bovine heart PKA catalytic subunit and modifi ed trypsin were purchased from Promega. Peptides were synthesized and purifi ed by Bio-Synthesis, Inc. (Lewisville, TX). P81 phosphocellulose paper was from Whatman International Ltd. Hybond-P polyvinylidene difl uoride (PVDF) membranes
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

CGI-58/ABHD5 is phosphorylated by protein kinase A 111
to acid hydrolysis with 6 N HCl ( 22 ). Hydrolysates were dried under vacuum and applied to cellulose TLC plates (EM Science) with standard phosphoamino acids, phosphoserine, phos-phothreonine, and phosphotyrosine. Amino acids were sepa-rated by 2D electrophoresis using formic acid:acetic acid:water (50:156:1,794, v/v) in the fi rst dimension and acetic acid:pyridine:water (100:10:1,890, v/v) in the second dimension ( 23 ). Following electrophoretic separation, the TLC plates were dried and subjected to phosphorimaging analysis with a Molecular Dy-namics Storm phosphorimager. Standard phosphoamino acids were visualized by spraying the plate with 0.25% ninhydrin in acetone.
MS Purifi ed recombinant CGI-58 was phosphorylated in vitro by
incubation with PKA and nonradioactive ATP. Phosphorylated residues of CGI-58 were identifi ed by LC-MS/MS after reduction, alkylation, and enzymatic digestion with AspN, modifi ed trypsin (Promega), or chymotrypsin (Roche). Digests of 0.5 � g CGI-58 were acidifi ed with 0.5% trifl uoroacetic acid and separated by nano-HPLC (Dionex Ultimate 3000) equipped with a � -precolumn (C18, 5 � m, 100 Å, 5 × 0.3 mm) and an Acclaim Pep-Map RSLC nanocolumn (C18, 2 � m, 100 Å, 150 × 0.075 mm) (all Thermo Fisher Scientifi c, Vienna, Austria). Samples were con-centrated on the enrichment column for 2 min at a fl ow rate of 20 � l/min with 0.5% trifl uoroacetic acid as an isocratic solvent. Separation was carried out on the nanocolumn at a fl ow rate of 300 nl/min using the following gradients, where solvent A is 0.3% formic acid in water and solvent B is a mixture of 80% ace-tonitrile in water containing 0.3% formic acid: 0–2 min 4% B, 2–35 min 4–28% B, 35–47 min 28–50% B, 47–48 min 50–95% B, 48–58 min 95% B, 58–58.1 min 95–4% B, 58.1–70 min 4% B, or 0–2 min 4% B, 2–180 min 4–28% B, 180–255 min 28–50% B, 255–260 min 50–95% B, 260–279 min 95% B, 279–280 min 95–4% B, 280–300 min 4% B. The sample was ionized in the nanospray source equipped with stainless steel emitters (ES528, Thermo Fisher Scientifi c). It was analyzed in an Orbitrap Velos Pro™ mass spectrometer (Thermo Fisher Scientifi c, Waltham, MA) operated in positive ion mode, applying alternating full scan MS ( m/z 400–2,000) in the ion cyclotron and MS/MS by high energy collision-induced dissociation of the 10 most intense peaks in the Orbitrap with dynamic exclusion enabled or neutral loss MS 3 scanning for the loss of phosphate in the ion trap.
The LC-MS/MS data were analyzed by searching the mamma-lian SwissProt public database with Proteome Discoverer 1.4 (Thermo Fisher Scientifi c) and Mascot 2.4 (MatrixScience, Lon-don, UK). Carbamidomethylation on Cys was entered as a fi xed modifi cation. Oxidation on methionine and phosphorylation on serine or threonine was entered as a variable modifi cation. A pre-cursor mass error tolerance of 10 ppm and a product mass error tolerance of 0.7 Da were used. A maximum false discovery rate of 5% using decoy database search and a Mascot ion score cut off of 30 were chosen as identifi cation criteria.
Measurement of CGI-58 coactivation of ATGL in vitro Purifi ed recombinant CGI-58 was phosphorylated in vitro by
incubation with PKA and nonradioactive ATP. Both phosphory-lated and nonphosphorylated proteins were incubated with an Sf9 insect cell lysate expressing recombinant mouse ATGL ( 24 ) and a radioactive triacylglycerol substrate emulsifi ed with phos-pholipids, as described previously ( 10 ). Fatty acids were extracted from the reaction mixture with solvents and quantifi ed by scintil-lation counting ( 10 ).
In other experiments, recombinant CGI-58, S239D CGI-58, and S239E CGI-58 were expressed in E. coli , followed by disruption of the cells by sonication and removal of cell debris by centrifugation
Expression of proteins was induced with 0.5 mM isopropyl � -D-1-thiogalactopyranoside when cells reached an optical density of 0.6 at 600 nm. Induced His-pSumo-CGI-58 cells were grown for 9–12 h, while His-pSumo-ATGL 1-288 and His-pSumo cells were grown for 4 h at 30°C. Expression of the proteins was confi rmed with SDS-PAGE.
E. coli expressing recombinant CGI-58 in His-pSumo were dis-rupted by probe sonication in buffer-1 [20 mM Tris-HCl, 500 mM NaCl, 30 mM imidazole, 0.1% NP-40, 3.5 mM � -mercaptoethanol (pH 7.8)] supplemented with protease inhibitor cocktail (Com-plete, EDTA-free Tabs-Roche), 750 U benzonase nuclease HC (Novagen), and 1 mg/ml lysozyme. Cell debris was removed by centrifugation at 30,000 g for 40 min and soluble recombinant CGI-58 was purifi ed with affi nity chromatography using a 5 ml His-Trap FF column (GE Healthcare). The column was washed extensively with buffer-1 and eluted with a linear gradient of 10 column volumes with buffer-2 [20 mM Tris-HCl, 500 mM NaCl, 250 mM imidazole, 10% glycerol, 3.5 mM � -mercaptoethanol (pH 7.8)]. Cleavage with tobacco etch virus protease was per-formed at room temperature for 4 h and the cleaved CGI-58 was further purifi ed on a Superdex 200 (Sigma-Aldrich) column equilibrated in buffer-3 [20 mM Tris-HCl, 300 mM NaCl, 1 mM DTT, 1 mM EDTA (pH 7.8)]. Residual 6-His-smt was removed by reverse affi nity chromatography with a 5 ml His-Trap FF column equilibrated in buffer-3. Recombinant CGI-58 was then dialyzed into the PKA assay buffer.
E. coli expressing recombinant ATGL 1-288 and 6-His-smt pro-tein were disrupted by sonication in buffer-4 [50 mM Tris-HCl, 300 mM NaCl, 10% glycerol, 20 mM imidazole, 1 mM benzami-dine, 1 mM Tris(2-carboxyethyl)phosphine (TCEP) (pH 7.8)] supplemented with 0.1 mM phenylmethanesulfonylfl uoride and protease inhibitor cocktail. Cell debris was removed by centrifu-gation and soluble protein was loaded onto a 1 ml His-Trap FF column. The column was washed extensively with buffer-4 and the proteins were eluted with a gradient of 12 column volumes of buffer-5 [50 mM Tris-HCl, 300 mM NaCl, 10% glycerol, 500 mM imidazole, 1 mM benzamidine, 1 mM TCEP (pH 7.8)]. Subse-quently, ATGL 1-288 was dialyzed into buffer-6 [50 mM Tris-HCl, 300 mM NaCl, 10% glycerol, 1 mM EDTA, 1 mM TCEP (pH 7.8)] and used directly in the triacylglycerol hydrolase activity assay.
Phosphorylation of recombinant mouse CGI-58 and synthetic peptides with PKA
Peptides of 10 amino acids surrounding and including the RKYSS sequence, with peptides containing RKYSS, RKYSA, RKYAS, and RKYAA, were tested for phosphorylation by PKA. Ad-ditionally, the PKA-mediated phosphorylation of comparable variants of purifi ed recombinant CGI-58 was examined. Both syn-thetic peptides and variants of purifi ed recombinant CGI-58 ( � 0.5–0.8 � g) were incubated with the indicated concentrations of PKA and 50 � M [ � 32 P]ATP ( � 230,000–470,000 cpm/nmol) in 10 mM MgCl 2 , 60 mM DTT, 50 mM Tris-HCl (pH 8.0); reactions were incubated for 10 min at 30°C. Reactions were stopped by the addition of 4× Laemmli’s sample buffer ( 21 ), followed by SDS-PAGE, immunoblot analysis, and autoradiography, or by spotting the reaction mixtures onto Whatman P81 phosphocel-lulose fi lters, followed by repeated treatments of the fi lters with 75 mM phosphoric acid and drying. All phosphorylation reac-tions were performed in triplicate. A unit of PKA was defi ned as the amount of enzyme that catalyzed the formation of 1 nmol of product/min. For some experiments, the PKA phosphorylation reaction was performed with nonradioactive ATP.
Phosphoamino acid analysis For phosphoamino acid analysis, a portion of a PVDF mem-
brane containing 32 P-labeled recombinant CGI-58 was subjected
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

112 Journal of Lipid Research Volume 56, 2015
PBS for 20 min, followed by a PBS wash step, and prepared for microscopy, as described previously ( 31 ). Cells expressing perili-pin 1A and CGI-58 were probed with goat polyclonal antisera raised against an N-terminal peptide of perilipin 1A (kindly do-nated by Dr. Constantine Londos, formerly of National Institutes of Health, Bethesda, MD, deceased) and rabbit polyclonal anti-sera raised against recombinant mouse CGI-58 ( 8 ), followed by anti-goat Alexa Fluor 546 and anti-rabbit Alexa Fluor 633. Nuclei were stained with Hoechst 33422 and lipid droplets were stained with BODIPY 493/503 (Life Technologies) ( 32 ), each at 0.1 � g/ml in PBS for 10 min. Cells were viewed with either a Nikon Eclipse E800 fl uorescence microscope equipped with a Photometrics CoolSNAP EZ digital camera or a LSM510 Meta confocal laser scanning microscope (Zeiss, Oberkochen, Germany) using a 63× oil immersion lens. Images were processed using Zen 2008 and ImageJ software. Cells were manually scored for protein localiza-tion patterns by at least two observers blinded to sample identity and automatically scored using the ImageJ JACoP plugin ( 33 ) to determine Manders coeffi cients; more than 50 cells were counted for each determination.
Measurement of cellular triacylglycerol Human NLSD fi broblasts were transduced with adenovirus
to drive the expression of WT CGI-58 or S239A/S240A-mutated CGI-58. At various times after transduction, cells were harvested and lysed in hypotonic lysis solution (10 mM Tris-HCl, 10 mM NaF, 1 mM EDTA, 10 � g/ml leupeptin, 100 � M 4-(2-aminoethyl)ben-zenesulfonylfl uoride hydrochloride, and 500 � M benzamidine) and homogenized by probe sonication (Branson Sonifi er) for 10 s. Lipids were extracted from a portion of samples using isopropanol:hexane:water (80:20:2), and the lipid phase was col-lected and evaporated. Lipid extracts of corn oil were used as tria-cylglycerol standards. The cellular triacylglycerol content was determined using an enzymatic assay from Thermo Electron Trace adapted for use with cultured cells ( 34 ). Absorbance of samples was determined at 540 nm using a VersaMax microplate reader (Molecular Devices). The triacylglycerol measurements were ex-pressed relative to cellular protein content measured by the Bio-Rad DC Protein Assay.
In one experiment, 6 h after transduction of NLSD fi broblasts with intact or S239A/S240A-mutated CGI-58, 10 � M forskolin and 0.5 mM IBMX were added to increase adenylyl cyclase activ-ity, in turn, activating PKA. Cells were collected at 0, 0.5, 1, and 2 h for the determination of triacylglycerol levels.
Adipose tissue collection Animal care and handling were performed in accordance with
the standards established by the Austrian Federal Ministry of Sci-ence and Research, Division of Genetic Engineering and Animal Experiments (Vienna, Austria) and protocols were approved by an institutional review board. Mice had ad libitum access to food and water under a 12 h light/12 h dark cycle in a temperature-controlled environment. Male C57Bl/6 mice (age 8–12 weeks, body weight 20–30 g) were fed a chow diet (Ssniff ® , Soest, Ger-many) and fasted during the daytime for 6 h prior to euthaniza-tion. Gonadal white adipose tissues were collected and cut into small pieces. Tissue samples were incubated in low glucose DMEM supplemented with 2% fatty acid-free BSA, with half of the tissue fragments additionally incubated with 10 � M isoprotere-nol and 0.5 mM IBMX for 20 min at 37°C with gentle shaking. Tissues were then lysed by sonication in ice-cold PBS (pH 7.4), with protease and phosphatase inhibitors. Lysates were centri-fuged at 10,000 g for 10 min at 4°C. Protein content of superna-tants was determined by Bradford Assay.
for 10 min at 2,700 g and determination of the protein concen-tration of the extracts by Bradford assay. Twenty-six micrograms of the supernatants with variants of recombinant CGI-58 were used in triacylglycerol hydrolase assays with 8 � g of partially puri-fi ed recombinant ATGL 1-288, as described ( 25 ).
Generation of recombinant adenovirus Adenoviral expression vectors driving the expression of mouse
CGI-58 and � -galactosidase were described previously ( 19 ). Ade-noviral expression vectors driving expression of the S239A/S240A-mutated variant of CGI-58 and perilipin 1A were prepared by ligating the cDNA for S239A/S240A-mutated CGI-58, mouse perilipin 1A, or mutated perilipin 1A lacking serine residues in six consensus sites for PKA phosphorylation into the shuttle vector for the AdEasy XL adenoviral vector system (Stratagene), and then following the manufacturer’s protocols for recombination of the shuttle vector to make adenoviral expression vectors and for assembly of virions in cultured AD293 cells. The mutated perili-pin 1A cDNA encoded alanine substitutions for serine residues at positions 81, 222, 276, 433, and 492 to prevent phosphorylation by PKA, and a glutamate substitution for serine 517 to permit tar-geting of the mutated perilipin 1A to lipid droplets ( 26 ). Adeno-viral preparations were purifi ed over cesium chloride gradients.
Cell culture and adenoviral transduction Normal human skin fi broblasts (WS1) were obtained from
American Type Culture Collection (Manassas, VA); human skin fi broblasts from an individual with NLSD ( 27–29 ) were gener-ously provided by Dr. Rosalind A. Coleman (University of North Carolina, Chapel Hill, NC). Normal and NLSD fi broblasts were cultured as described previously ( 27–29 ). NLSD fi broblasts were transduced with purifi ed adenoviral preparations for expression of intact (WT) mouse CGI-58 (WT) or S239A/S240A-mutated CGI-58; cells were collected at various times following transduc-tion for the determination of triacylglycerol levels.
For other experiments, purifi ed adenoviruses were used to transduce Cos-7 cells or NIH3T3CAR � fi broblasts. Cos-7 cells were cultured in DMEM supplemented with 10% FBS, 100 units/ml penicillin, and 100 � g/ml streptomycin. NIH3T3CAR � fi bro-blasts are NIH3T3 fi broblasts stably expressing a truncated ver-sion of the Coxsackie and adenovirus receptor (CAR � ) lacking the cytoplasmic signaling domain ( 30 ) to increase the uptake of adenoviral vectors. The cells were cultured in DMEM sup-plemented with 10% calf serum, 100 units/ml penicillin, and 100 � g/ml streptomycin, with 800 � g/ml G418 to maintain se-lection of cells expressing CAR � . Cells were transduced with ad-enoviruses for the expression of either WT mouse perilipin 1A or the mutated variant of mouse perilipin 1A that lacks six serine residues in PKA consensus sequences. At the same time, the cells were also transduced with either WT CGI-58 or S239/S240A CGI-58 and incubated for 48 h before processing the cells for immu-nofl uorescence microscopy.
Immunofl uorescence microscopy Twenty-four hours after transduction, Cos-7 and NIH3T3CAR �
cells expressing perilipin 1A or mutated perilipin 1A and CGI-58 or S239A/240A-mutated CGI-58 were transferred to glass cover-slips. Thirty-six hours after transduction, 200 � M oleic acid com-plexed to fatty acid-free BSA at a 4:1 molar ratio ( 7 ) was added to increase the synthesis and storage of triacylglycerol in lipid droplets. Forty-eight hours after transduction, cells were incubated with 10 � M forskolin and 0.5 mM IBMX in culture medium supplemented with 1% fatty acid-free BSA for 30 min for stimulated conditions, or in culture medium with 1% fatty acid-free BSA for 30 min for basal conditions. Cells were fi xed with 4% paraformaldehyde in
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

CGI-58/ABHD5 is phosphorylated by protein kinase A 113
of CGI-58 from vertebrate species, but not in the sequences of CGI-58 orthologs from Caenorhabditis elegans or Drosoph-ila melanogaster ( Fig. 1A ). To test the hypothesis that CGI-58 is a substrate for PKA, we studied the phosphorylation of recombinant mouse CGI-58 that was purifi ed from E. coli lysates ( 19 ). Recombinant CGI-58 was incubated with mammalian PKA and [ � - 32 P]ATP in vitro; the incorpora-tion of radioactive phosphate into CGI-58 was monitored by phosphorimaging analysis of blots and revealed that re-combinant CGI-58 is a substrate for PKA ( Fig. 1B ). Phos-phoamino acid analysis of the 32 P-labeled protein showed that PKA phosphorylates CGI-58 at a serine residue ( Fig. 1C ).
The kinetics of the in vitro phosphorylation reaction were studied in detail. Phosphorylation of CGI-58 by PKA displayed saturation kinetics and was dependent on time ( Fig. 2A ), concentration of PKA ( Fig. 2B ), concentration of ATP ( Fig. 2C ), and concentration of recombinant CGI-58 ( Fig. 2D ). Analysis of the data using the Michaelis-Menten equation yielded apparent K m values for ATP and recombi-nant CGI-58 of 14.0 ± 2.8 � M and 71.5 ± 13.9 � g/ml, respectively.
PKA phosphorylates CGI-58 at Ser239 The predicted PKA consensus sequence of mouse CGI-58
has potential phosphorylation sites at Ser239 and Ser240 ( Fig. 1A ). To determine whether these serine residues are
Immunoprecipitations Forskolin-treated and untreated control white adipose tissue
lysates and lysates from NIH3T3CAR � fi broblasts expressing intact or 239A/S240A-mutated CGI-58 were precleared with Pro-tein A-Sepharose beads. CGI-58 antiserum ( 8 ) was cross-linked to Protein A-Sepharose using dimethyl pimelimidate. Precleared protein lysates (1 mg protein) were incubated with cross-linked antiserum for 16 h at 4°C; control immunoprecipitations were conducted using control IgG. Immunoprecipitates were col-lected by centrifugation at 1,000 g for 1 min at 4°C. After washing the beads three times with immunoprecipitation buffer [50 mM Tris-HCl (pH 7.4), 300 mM NaCl, 0.1% Triton X-100, 5 mM EDTA, 50 mM NaF, 0.02% NaN 3 , with protease and phosphatase inhibitors], proteins were eluted using hot SDS-PAGE XT (Bio-Rad) sample buffer for 10 min at 90°C. Proteins were separated by electrophoresis on 4–20% gradient gels for SDS-PAGE (Bio-Rad) and transferred to nitrocellulose membranes. Membranes were immunoblotted with anti-CGI-58 (1:50,000) ( 8 ) or phos-pho-S/T-PKA substrate antibody (1:1,000) (#9621; Cell Signaling Technology, Danvers, MA) in 5% (w/v) BSA in TBS with 0.1% Tween-20 overnight at 4°C. Blots were incubated with Clean-Blot IP detection reagents (Thermo Scientifi c) (1:2,500) in 5% (w/v) nonfat milk in TBS with 0.02% Tween-20 for 2 h at 25°C, fol-lowed by detection with SuperSignal West Femto chemilumines-cent substrate (Thermo Scientifi c).
SDS-PAGE, 2D electrophoresis, and immunoblot analysis SDS-PAGE, 2D gel electrophoresis, and immunoblotting of
proteins on nitrocellulose membranes were performed using standard procedures. For 2D gel electrophoresis, proteins were separated in the fi rst dimension on immobilized nonlinear pH gradient strips (pH 3–10) (Bio-Rad) by a gradual increase in volt-age from 200 V to 8,000 V at the rate of 4 V/min (28 kVh in to-tal). The second dimensional separation was performed using 4–12% gradient SDS-PAGE gels (Bio-Rad). Antibodies used for immunoblotting were rabbit polyclonal anti-mouse CGI-58 antiserum ( 8 ), rabbit polyclonal anti- � -galactosidase antibody (#ab616; Abcam, Cambridge, MA), rabbit polyclonal anti-cal-nexin antibody (#SPA-865; StressGen, Victoria, BC, Canada), phospho-S/T-PKA substrate antibody (#9621; Cell Signaling Technology, Danvers, MA), and polyclonal antisera raised against an N-terminal recombinant peptide of perilipin A ( 35 ). Proteins were detected using peroxidase-conjugated goat anti-rabbit IgG (Sigma) and enhanced chemiluminescence reagents (Amer-sham Biosciences or Thermo Scientifi c).
Data and statistical analyses Kinetics data were analyzed according to the Michaelis-Menten
equation using the GraphPad Prism kinetic model-fi tting pro-gram. Data are reported as mean ± standard deviation or stan-dard error and were analyzed using two-way ANOVA and Tukey’s post hoc test with SPSS software. Differences between samples were considered signifi cant at P < 0.05.
RESULTS
Recombinant CGI-58 is phosphorylated by PKA in vitro Analysis of the predicted amino acid sequence of mouse
CGI-58 using PROSITE ( 36 ) revealed a PKA consensus se-quence of RKYSS containing two potential targets for ser-ine phosphorylation, Ser239 and Ser240. This consensus sequence is highly conserved in the amino acid sequences
Fig. 1. CGI-58 is a substrate for in vitro phosphorylation by PKA. A: Amino acid sequences of a portion of CGI-58 are aligned for human (NP_057090), chimpanzee (XP_516397), mouse (NP_080455), rat (NP_997689), C. elegans (hypothetical protein C37H5.2, NP_504299.2), and D. melanogaster (CG1882, isoform B, NP_724609.1). The sequences show conservation of the RKYSS consensus site for PKA phosphorylation in vertebrates, but not in-vertebrates. B: Recombinant CGI-58 (1 � g) was incubated with PKA (0.32 unit/ml) and 50 � M [ � - 32 P]ATP (236,000 cpm/nmol) for 15 min, followed by SDS-PAGE, transfer to PVDF, and phospho-rimaging analysis. C: A PVDF membrane slice containing 32 P-labeled CGI-58 was hydrolyzed with 6 N HCl, followed by 2D electropho-retic analysis. The positions of the standard phosphoamino acids, phosphoserine (P-Ser), phosphothreonine (P-Thr), and phospho-tyrosine (P-Tyr), are indicated. The data are representative of two independent experiments.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

114 Journal of Lipid Research Volume 56, 2015
CGI-58 with the intact RKYSS sequence, and S240A-mutated CGI-58 are good substrates for PKA ( Fig. 3 ). These data sug-gest that the major site of phosphorylation is Ser239.
MS was used to further study the in vitro phosphoryla-tion of CGI-58. Recombinant mouse CGI-58 was purifi ed from E. coli lysates, incubated with PKA and ATP, re-duced, alkylated and digested with trypsin, chymotrypsin, or AspN. Digests were then analyzed by LC-MS/MS using different fragmentation and scanning techniques. A search
substrates for PKA, 10-amino acid peptides containing the intact PKA consensus sequence of RKYSS or comparable peptides with alanine substitutions for these serine resi-dues both individually and in combination (S239A, S240A, S239A/S240A) were synthesized. We examined whether these peptides serve as substrates for PKA in vitro. Both the peptide with the intact RKYSS consensus sequence and the S240A peptide were highly phosphorylated ( Table 1 ). In contrast, the S239A peptide was a poor substrate for PKA, showing only 6% of the activity of the peptide with intact RKYSS. The S239A/S240A peptide was not phos-phorylated by PKA in vitro. These data indicate that Ser239, in the context of the local amino acid environ-ment, serves as a substrate for PKA in vitro.
We then extended these studies to full-length recombi-nant CGI-58. Recombinant variants of CGI-58 with the in-tact RKYSS sequence, or comparable mutations (S239A, S240A, S239A/S240A), were expressed and purifi ed from E. coli lysates, and then incubated with PKA and [ � - 32 P]ATP in vitro. Autoradiography of SDS-PAGE gels containing the re-combinant proteins revealed that the S239A-mutated variant of recombinant CGI-58 is a poor substrate for PKA, whereas
Fig. 2. Kinetics of the in vitro phosphorylation of recombinant CGI-58 by PKA. A: Time dependence. Re-combinant CGI-58 (1 � g) was incubated with PKA (0.32 unit/ml) and 50 � M [ � - 32 P]ATP for the indicated time intervals. B: Concentration of PKA. Recombinant CGI-58 (1 � g) was incubated with the indicated con-centrations of PKA and 50 � M [ � - 32 P]ATP for 10 min. C: Concentration of ATP. Recombinant CGI-58 (1 � g) was incubated with PKA (0.32 unit/ml) and the indicated concentrations of [ � - 32 P]ATP for 10 min. D: Concen-tration of CGI-58. The indicated concentrations of recombinant CGI-58 were incubated with PKA (0.32 unit/ml) and 50 � M [ � - 32 P]ATP for 10 min. After the phosphorylation reactions, the samples were spotted onto What-man P81 paper, washed with 75 mM phosphoric acid, and radioactivity determined by scintillation counting. The data for each panel are the mean ± standard deviation of triplicate samples from one representative experiment out of two. When error bars are not visible, they are contained within the symbol.
TABLE 1. PKA-mediated phosphorylation of peptides of CGI-58
Peptide Sequence PKA Activity ( � mol/min/mg)
S239/S240 (WT) FKRKYSSMFE 43.1 ± 2.1S240A FKRKYSAMFE 45.8 ± 4.7S239A FKRKYASMFE 2.54 ± 0.32S239A/S240A FKRKYAAMFE 0.04 ± 0.02
Ten-amino acid peptides (0.5 mM) were incubated with PKA (0.32 unit/ml) and 50 � M [ � - 32 P]ATP for 10 min and then spotted onto Whatman P81 phosphocellulose fi lters, followed by rinses with 75 mM phosphoric acid and drying. Incorporation of phosphate into peptides was determined by scintillation counting. Data are mean ± standard deviation of triplicate reactions.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

CGI-58/ABHD5 is phosphorylated by protein kinase A 115
observed in the PKA-stimulated tissue when compared with control tissue ( Fig. 4A ). Additionally, CGI-58 was im-munoprecipitated from the stimulated and control tissues and immunoblotted with an anti-phospho-S/T PKA sub-strate antibody or CGI-58 antiserum ( Fig. 4B ). Only the stimulated tissue showed a positive signal for phosphoryla-tion, while similar amounts of CGI-58 were detected in im-munoprecipitates from both the stimulated and control samples. Finally, to confi rm that Ser239 is the major site for phosphorylation of CGI-58 by PKA in intact cells, CGI-58 was immunoprecipitated from stimulated and control NIH3T3CAR � cells expressing intact CGI-58 (WT) or S239A/S240A-mutated CGI-58 ( Fig. 4C ). As expected, only stimulated cells expressing intact CGI-58 showed de-tectable phosphorylation.
PKA-mediated phosphorylation of Ser239 does not alter CGI-58 function in either coactivation of ATGL in vitro or turnover of triacylglycerol in NLSD fi broblasts
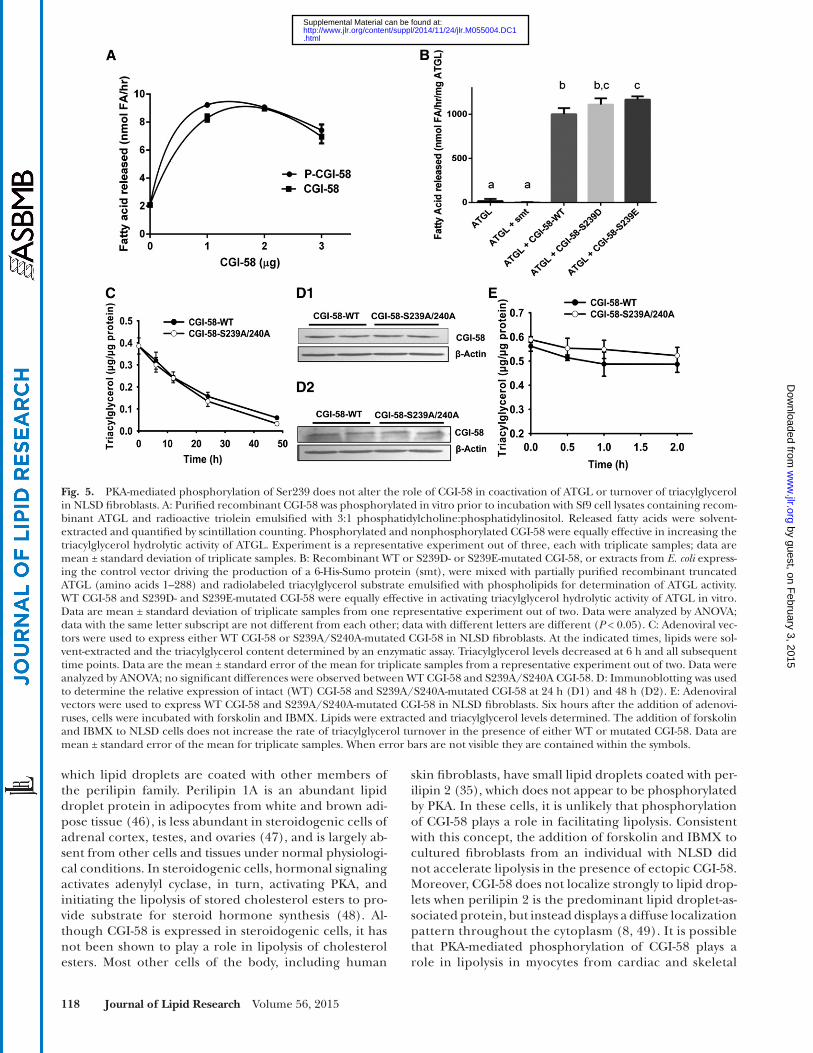
To gain understanding of the function of CGI-58 phos-phorylation, CGI-58-mediated coactivation of the triacylg-lycerol hydrolase activity of ATGL was assessed in vitro. Purifi ed recombinant CGI-58 was incubated with PKA and nonradioactive ATP prior to incubation of phosphory-lated and nonphosphorylated CGI-58 with Sf9 insect cell lysates containing recombinant mouse ATGL and an emulsifi ed radioactive triacylglycerol substrate. Phosphor-ylated and nonphosphorylated CGI-58 showed equivalent capacity to activate the triacylglycerol hydrolase activity of ATGL over hydrolysis catalyzed by ATGL alone ( Fig. 5A ). Similar results were obtained when CGI-58 with S239D or S239E phosphomimetic mutations were used to activate a partially purifi ed truncated variant (amino acids 1–288) of recombinant ATGL ( Fig. 5B ). The data indicate that the phosphorylation of CGI-58 neither facilitates nor impedes the coactivation of ATGL.
To test CGI-58 function in intact cells, adenoviral ex-pression vectors were employed to drive the expression of either intact CGI-58 (WT) or S239A/S240A-mutated CGI-58 in cultured human NLSD fi broblasts that lack func-tional CGI-58 ( 10 ), and have perilipin 2-containing lipid droplets (not shown). Immunoprecipitations revealed that NLSD cells express ATGL (data not shown), yet are unable to turn over triacylglycerol normally in the absence of CGI-58 ( 27 ). Without ectopic expression of CGI-58, NLSD cells expressing � -galactosidase as a control protein show approximately 15-fold higher triacylglycerol levels relative to fi broblasts from control humans cultured in the same media (supplementary Fig. 1). By 6 h after transduction with adenoviruses for either the intact (WT) or mutated variant of CGI-58, the triacylglycerol content of NLSD cells decreased signifi cantly (relative to untransduced cells; t = 0 h), and continued to decrease over 48 h ( Fig. 5C ). The rate of triacylglycerol turnover was similar for cells express-ing WT and mutated CGI-58. Immunoblotting of cell ly-sates for CGI-58 revealed that 6 h is suffi cient to detect ectopic CGI-58 (data not shown); levels of CGI-58 increased with longer time of protein expression. More-over, equivalent levels of intact and mutated CGI-58 were
of the mammalian SwissProt database unambiguously identifi ed Ser239 as the major phosphorylation site in CGI-58 peptides derived from three different digests ( Table 2 ). The majority of peptides containing Ser239 were phosphorylated.
To determine whether CGI-58 is a substrate for PKA in intact cells or tissues, two types of experiments were con-ducted. Murine white adipose tissue was stimulated ex vivo with isoproterenol and IBMX to activate adenylyl cyclase, sustain elevated levels of cAMP, and, in turn, activate PKA. Immunoblots of 2D gels of the tissue lysates revealed three to four distinct spots detected by the CGI-58 antiserum that likely represent CGI-58 isoforms. Because phosphory-lation adds negative charge, the isoelectric point of a phos-phorylated protein is shifted to an acidic pH, as was
Fig. 3. CGI-58 is phosphorylated in vitro by PKA on Ser239. Re-combinant CGI-58 (WT) or S240A-, S239A-, or S239A/S240A-mutated CGI-58 (1 � g) were incubated with PKA (0.32 unit/ml) and 50 � M [ � - 32 P]ATP before SDS-PAGE, transfer to PVDF, and phosphorimaging analysis (A). B: Coomassie-stained gel prior to protein transfer.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

116 Journal of Lipid Research Volume 56, 2015
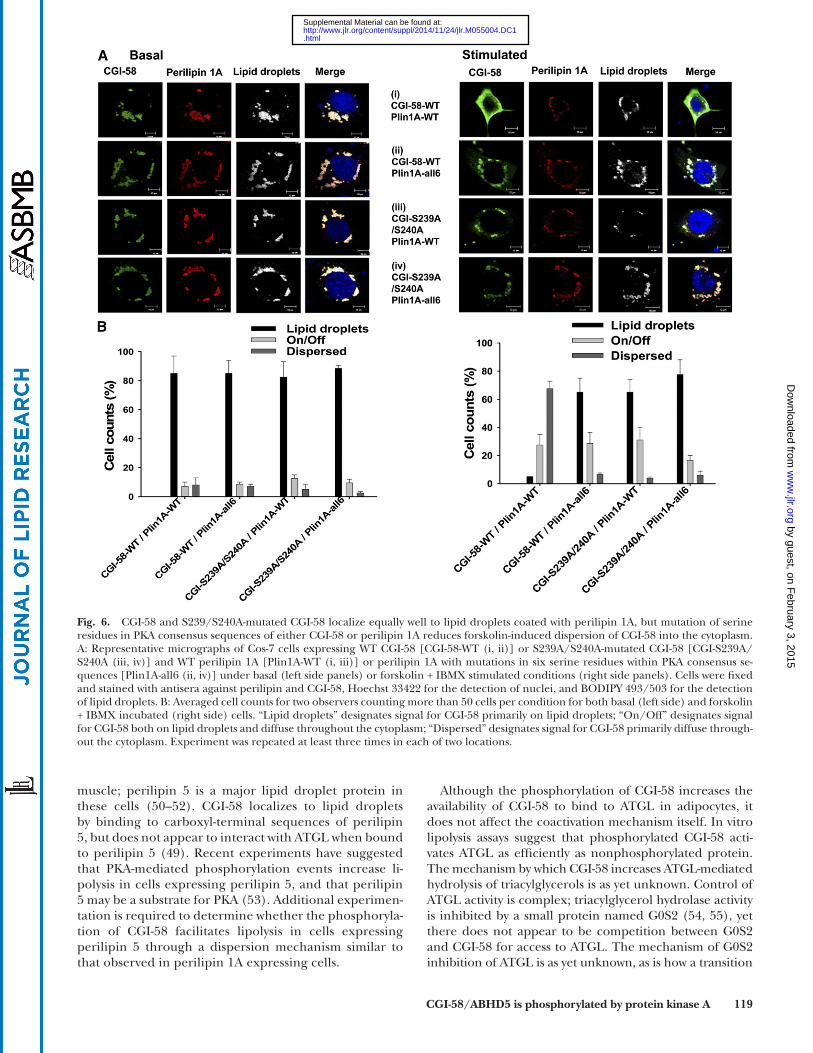
1A lacking serine residues in six consensus PKA sites. Ad-enoviral titers were adjusted to drive equivalent levels of protein expression for both variants of perilipin 1A and both variants of CGI-58 (supplementary Fig. 2). For 12 h prior to starting the experiment, the cells were incubated with oleic acid to increase triacylglycerol synthesis and lipid droplet formation, and consequently, stabilize perili-pin 1A ( 38 ). Cells were then incubated for 30 min with forskolin and IBMX, or under basal conditions, before fi xation and immunostaining of the cells for perilipin and CGI-58. The data show that both intact (WT) and S239A/S240A-mutated CGI-58 localized to perilipin-coated lipid droplets under basal conditions, whether WT or mutated variants of perilipin 1A were expressed ( Fig. 6A, B ; Table 3 ). In contrast, the mutation of PKA-site serine residues in CGI-58 and perilipin 1A decreased the dispersion of CGI-58 into the cytoplasm following incubation of the cells with forskolin and IBMX, whether only one protein was mutated or both were mutated in combination. Thus, the PKA-mediated phosphorylation of both CGI-58 and perili-pin 1A contribute to the dispersion of CGI-58 from perili-pin 1A-coated lipid droplets.
We conducted a similar experiment using cells express-ing S239D CGI-58 and perilipin 1A. Under basal condi-tions, S239D CGI-58 localized effi ciently to perilipin 1A-coated lipid droplets (supplementary Fig. 3), suggest-ing that the addition of negative charge to amino acid 239 of CGI-58 is insuffi cient to cause dispersion of CGI-58 from perilipin 1A-coated lipid droplets without the activa-tion of PKA. When cells were stimulated with forskolin and IBMX, S239D CGI-58 effi ciently dispersed into the cy-toplasm (supplementary Fig. 3). These data suggest that negative charge at position 239 is important for CGI-58 dispersion when PKA has been activated.
DISCUSSION
In this study, we employed a variety of approaches to demonstrate that CGI-58 is phosphorylated on Ser239 by PKA. The fi nding is supported by data from the phosphor-ylation of recombinant CGI-58 in vitro, immunoblotting of immunoprecipitated proteins from mammalian tissue and cells in which phosphorylation occurred in intact cells, and MS. All of our studies were conducted using mouse CGI-58; however, human CGI-58 is likely phosphory-lated, given the complete conservation of the PKA consen-sus sequence in chordates.
detected ( Fig. 5D ). In summary, S239A/S240A-mutated CGI-58 was as effective as WT CGI-58 in facilitating the turnover of triacylglycerols in NLSD cells.
Initial experiments to test CGI-58 function in NLSD fi -broblasts were conducted in the absence of compounds to activate PKA. We next investigated whether the activation of PKA would alter the rate of triacylglycerol turnover in NLSD cells expressing ectopic CGI-58. Adenoviruses driv-ing the expression of either intact (WT) or S239A/S240A-mutated CGI-58 were added to NLSD cells 6 h prior to the addition of forskolin and IBMX. Measurement of the tria-cylglycerol content of cell lysates over the next 2 h revealed no increase in the rate of lipolysis due to the activation of PKA, and no difference in the rate of triacylglycerol turn-over when comparing data from cells expressing WT CGI-58 to those from cells expressing S239A/S240A-mutated CGI-58 ( Fig. 5E ). Thus, PKA is not a major modulator of lipolysis in NLSD fibroblasts, and phosphorylation of Ser239 (or Ser240) is not required for the function of CGI-58 in facilitating triacylglycerol turnover in human skin fi -broblasts. It is important to note that fi broblasts lack both perilipin 1 and hormone-sensitive lipase; hence, the major players in PKA-mediated lipolysis are absent in these cells.
PKA-mediated phosphorylation of Ser239 alters the subcellular localization of CGI-58 in forskolin-treated cells expressing perilipin 1A
We next asked whether the PKA-mediated phosphoryla-tion of CGI-58 affects subcellular localization of the pro-tein. PKA controls lipolysis in adipocytes in part by modulating the subcellular localization of CGI-58 ( 8, 11, 37 ). Under basal conditions, CGI-58 binds to perilipin 1A on adipocyte lipid droplets, but when PKA is activated, CGI-58 disperses into the cytoplasm ( 8, 11 ). The PKA-mediated phosphorylation of carboxyl terminal serine residues of perilipin 1A has been shown to facilitate the dispersion of CGI-58 from lipid droplets ( 11 ); this event is necessary for CGI-58 to gain access to ATGL. We hypoth-esized that the PKA-mediated phosphorylation of CGI-58 might also contribute to the translocation of CGI-58 into the cytoplasm in lipolytically stimulated cells. Experi-ments were conducted in both cultured Cos-7 cells and NIH3T3CAR � fi broblasts; the latter cells lack detectable endogenous CGI-58, as determined by immunoprecipita-tion followed by immunoblotting (Fig. 4C). Using adeno-viral expression vectors, either intact CGI-58 (WT) or S239A/S240A-mutated CGI-58 was expressed with either intact (WT) perilipin 1A or a mutated variant of perilipin
TABLE 2. Phosphorylated peptides of CGI-58 identifi ed by MS
Enzyme Sequence m/z � m (ppm) Ion Score Expect Value
Trypsin KYsSMFEDDTVTEYIYHCNVQTPSGETAFK 1,209.85059 1.09 104 2.9E-009AspN DFKRKYsSmFE 767.32617 0.42 32 7.5E-003Chymotrypsin KRKYsSMF 571.75867 3.16 30 6.9E-003
Purifi ed recombinant CGI-58 was incubated with PKA in vitro, digested with either trypsin, AspN, or chymotrypsin, and analyzed by LC-MS/MS. Peptides were identifi ed by searching the mammalian SwissProt database with Proteome Discoverer and Mascot software. Modifi ed amino acids are shown in small letters (s designates phosphorylated serine, m designates oxidized methionine). The m/z and mass error ( � m) of measured peptides, as well as Mascot ion scores and expected values, are listed.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

CGI-58/ABHD5 is phosphorylated by protein kinase A 117
PKA-mediated phosphorylation of CGI-58 also plays a role in the dispersion of CGI-58 from the perilipin scaffold, to enable CGI-58 interaction with and coactivation of ATGL. A previous study has demonstrated that perilipin 1A se-questration of CGI-58 prevents the interaction of CGI-58 and ATGL, reducing basal lipolysis; human truncation mutations in perilipin 1A that prevent CGI-58 sequestra-tion lead to elevated rates of basal lipolysis ( 45 ). Thus, pre-cise control of the subcellular localization of CGI-58 is an important component of the regulation of lipolysis in adipocytes.
Although the PKA-mediated phosphorylation of CGI-58 is an important step in the activation of lipolysis for cells in which perilipin 1A is the major lipid droplet-associated protein, it is likely less important for cells in
In adipocytes, the PKA-mediated phosphorylation of key mediators of lipolysis is a crucial event in the initiation of lipolysis. Phosphorylation of hormone-sensitive lipase is required to trigger movement of the lipase from a diffuse cytoplasmic localization to the surfaces of lipid droplets ( 39–42 ), and to activate lipase activity in an as yet poorly understood mechanism ( 12, 43, 44 ). The phosphorylation of perilipin 1A is also required to enable hormone-sensitive lipase docking on lipid droplets through protein-protein interactions between the lipase and perilipin ( 12 ). Addi-tionally, phosphorylation of perilipin 1A is required to release CGI-58 from its binding site on a carboxyl termi-nal sequence of perilipin 1A ( 8, 11 ); this redistribution of CGI-58 into the cytoplasm increases the interaction of CGI-58 with ATGL ( 11 ). We now demonstrate that
Fig. 4. CGI-58 is a substrate for PKA in white adipose tissue and intact cells. White adipose tissue sections from C57B1/6 mice were incubated with 0.5 mM IBMX and 10 � M isoproterenol for 20 min at 37°C (A, B). A: Immunoblot of 2D electrophoresis gel of stimulated and control white adipose tissue lysates probed with CGI-58 antiserum. Inset shows an enlargement of the boxed region. B: Immunoblots of CGI-58 immunopre-cipitated from tissue lysates made from stimulated and control white adipose tissue probed with PKA pS/T substrate antibody (P-CGI-58) and CGI-58 antiserum. Control immunoprecipitations with control IgG showed no signal for CGI-58 [lane 1 (i)]; similarly, precipitations conducted without IgG, but with Protein A-Sepharose, showed no signal for CGI-58 [lane 1 (ii)]. C: NIH3T3CAR � cells expressing intact CGI-58 (WT) or S239A/240A-mutated CGI-58 were incubated with 0.5 mM IBMX and 10 � M forskolin for 20 min at 37°C. Immunoblots show CGI-58 immunoprecipitated from cell lysates made from stimulated and control NIH3T3CAR � cells expressing WT CGI-58 or S239A/S240A-mutated CGI-58 probed with PKA pS/T substrate antibody (i) and CGI-58 antiserum (ii). Phosphorylated protein was detected only when WT CGI-58 was expressed in cells and the cells were incubated under stimulated conditions. CGI-58 was not de-tected in immunoprecipitations of nontransduced cells, or when a nonimmune IgG was used for immunoprecipitations.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:
118 Journal of Lipid Research Volume 56, 2015
skin fi broblasts, have small lipid droplets coated with per-ilipin 2 ( 35 ), which does not appear to be phosphorylated by PKA. In these cells, it is unlikely that phosphorylation of CGI-58 plays a role in facilitating lipolysis. Consistent with this concept, the addition of forskolin and IBMX to cultured fi broblasts from an individual with NLSD did not accelerate lipolysis in the presence of ectopic CGI-58. Moreover, CGI-58 does not localize strongly to lipid drop-lets when perilipin 2 is the predominant lipid droplet-as-sociated protein, but instead displays a diffuse localization pattern throughout the cytoplasm ( 8, 49 ). It is possible that PKA-mediated phosphorylation of CGI-58 plays a role in lipolysis in myocytes from cardiac and skeletal
which lipid droplets are coated with other members of the perilipin family. Perilipin 1A is an abundant lipid droplet protein in adipocytes from white and brown adi-pose tissue ( 46 ), is less abundant in steroidogenic cells of adrenal cortex, testes, and ovaries ( 47 ), and is largely ab-sent from other cells and tissues under normal physiologi-cal conditions. In steroidogenic cells, hormonal signaling activates adenylyl cyclase, in turn, activating PKA, and initiating the lipolysis of stored cholesterol esters to pro-vide substrate for steroid hormone synthesis ( 48 ). Al-though CGI-58 is expressed in steroidogenic cells, it has not been shown to play a role in lipolysis of cholesterol esters. Most other cells of the body, including human
Fig. 5. PKA-mediated phosphorylation of Ser239 does not alter the role of CGI-58 in coactivation of ATGL or turnover of triacylglycerol in NLSD fi broblasts. A: Purifi ed recombinant CGI-58 was phosphorylated in vitro prior to incubation with Sf9 cell lysates containing recom-binant ATGL and radioactive triolein emulsifi ed with 3:1 phosphatidylcholine:phosphatidylinositol. Released fatty acids were solvent-extracted and quantifi ed by scintillation counting. Phosphorylated and nonphosphorylated CGI-58 were equally effective in increasing the triacylglycerol hydrolytic activity of ATGL. Experiment is a representative experiment out of three, each with triplicate samples; data are mean ± standard deviation of triplicate samples. B: Recombinant WT or S239D- or S239E-mutated CGI-58, or extracts from E. coli express-ing the control vector driving the production of a 6-His-Sumo protein (smt), were mixed with partially purifi ed recombinant truncated ATGL (amino acids 1–288) and radiolabeled triacylglycerol substrate emulsifi ed with phospholipids for determination of ATGL activity. WT CGI-58 and S239D- and S239E-mutated CGI-58 were equally effective in activating triacylglycerol hydrolytic activity of ATGL in vitro. Data are mean ± standard deviation of triplicate samples from one representative experiment out of two. Data were analyzed by ANOVA; data with the same letter subscript are not different from each other; data with different letters are different ( P < 0.05). C: Adenoviral vec-tors were used to express either WT CGI-58 or S239A/S240A-mutated CGI-58 in NLSD fi broblasts. At the indicated times, lipids were sol-vent-extracted and the triacylglycerol content determined by an enzymatic assay. Triacylglycerol levels decreased at 6 h and all subsequent time points. Data are the mean ± standard error of the mean for triplicate samples from a representative experiment out of two. Data were analyzed by ANOVA; no signifi cant differences were observed between WT CGI-58 and S239A/S240A CGI-58. D: Immunoblotting was used to determine the relative expression of intact (WT) CGI-58 and S239A/S240A-mutated CGI-58 at 24 h (D1) and 48 h (D2). E: Adenoviral vectors were used to express WT CGI-58 and S239A/S240A-mutated CGI-58 in NLSD fi broblasts. Six hours after the addition of adenovi-ruses, cells were incubated with forskolin and IBMX. Lipids were extracted and triacylglycerol levels determined. The addition of forskolin and IBMX to NLSD cells does not increase the rate of triacylglycerol turnover in the presence of either WT or mutated CGI-58. Data are mean ± standard error of the mean for triplicate samples. When error bars are not visible they are contained within the symbols.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:
CGI-58/ABHD5 is phosphorylated by protein kinase A 119
Although the phosphorylation of CGI-58 increases the availability of CGI-58 to bind to ATGL in adipocytes, it does not affect the coactivation mechanism itself. In vitro lipolysis assays suggest that phosphorylated CGI-58 acti-vates ATGL as effi ciently as nonphosphorylated protein. The mechanism by which CGI-58 increases ATGL-mediated hydrolysis of triacylglycerols is as yet unknown. Control of ATGL activity is complex; triacylglycerol hydrolase activity is inhibited by a small protein named G0S2 ( 54, 55 ), yet there does not appear to be competition between G0S2 and CGI-58 for access to ATGL. The mechanism of G0S2 inhibition of ATGL is as yet unknown, as is how a transition
muscle; perilipin 5 is a major lipid droplet protein in these cells ( 50–52 ). CGI-58 localizes to lipid droplets by binding to carboxyl-terminal sequences of perilipin 5, but does not appear to interact with ATGL when bound to perilipin 5 ( 49 ). Recent experiments have suggested that PKA-mediated phosphorylation events increase li-polysis in cells expressing perilipin 5, and that perilipin 5 may be a substrate for PKA ( 53 ). Additional experimen-tation is required to determine whether the phosphoryla-tion of CGI-58 facilitates lipolysis in cells expressing perilipin 5 through a dispersion mechanism similar to that observed in perilipin 1A expressing cells.
Fig. 6. CGI-58 and S239/S240A-mutated CGI-58 localize equally well to lipid droplets coated with perilipin 1A, but mutation of serine residues in PKA consensus sequences of either CGI-58 or perilipin 1A reduces forskolin-induced dispersion of CGI-58 into the cytoplasm. A: Representative micrographs of Cos-7 cells expressing WT CGI-58 [CGI-58-WT (i, ii)] or S239A/S240A-mutated CGI-58 [CGI-S239A/S240A (iii, iv)] and WT perilipin 1A [Plin1A-WT (i, iii)] or perilipin 1A with mutations in six serine residues within PKA consensus se-quences [Plin1A-all6 (ii, iv)] under basal (left side panels) or forskolin + IBMX stimulated conditions (right side panels). Cells were fi xed and stained with antisera against perilipin and CGI-58, Hoechst 33422 for the detection of nuclei, and BODIPY 493/503 for the detection of lipid droplets. B: Averaged cell counts for two observers counting more than 50 cells per condition for both basal (left side) and forskolin + IBMX incubated (right side) cells. “Lipid droplets” designates signal for CGI-58 primarily on lipid droplets; “On/Off” designates signal for CGI-58 both on lipid droplets and diffuse throughout the cytoplasm; “Dispersed” designates signal for CGI-58 primarily diffuse through-out the cytoplasm. Experiment was repeated at least three times in each of two locations.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

120 Journal of Lipid Research Volume 56, 2015
involvement of CGI-58 mislocalization in Chanarin-Dorfman syn-drome. J. Biol. Chem. 279 : 30490 – 30497 .
10 . Lass , A. , R. Zimmermann , G. Haemmerle , M. Riederer , G. Schoiswohl , M. Schweiger , P. Kienesberger , J. G. Strauss , G. Gorkiewicz , and R. Zechner . 2006 . Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman syndrome. Cell Metab. 3 : 309 – 319 .
11 . Granneman , J. G. , H. P. Moore , R. Krishnamoorthy , and M. Rathod . 2009 . Perilipin controls lipolysis by regulating the interactions of AB-hydrolase containing 5 (Abhd5) and adipose triglyceride lipase (Atgl). J. Biol. Chem. 284 : 34538 – 34544 .
12 . Wang , H. , L. P. Hu , K. Dalen , H. Dorward , A. Marcinkiewicz , D. Russell , D. W. Gong , C. Londos , T. Yamaguchi , C. Holm , et al . 2009 . Activation of hormone-sensitive lipase requires two steps, protein phosphorylation and binding to the PAT-1 domain of lipid droplet coat proteins. J. Biol. Chem. 284 : 32116 – 32125 .
13 . Lefèvre , C. , F. Jobard , F. Caux , B. Bouadjar , A. Karaduman , R. Heilig , H. Lakhdar , A. Wollenberg , J. L. Verret , J. Weissenbach , et al . 2001 . Mutations in CGI-58, the gene encoding a new pro-tein of the esterase/lipase/thioesterase subfamily, in Chanarin-Dorfman syndrome. Am. J. Hum. Genet. 69 : 1002 – 1012 .
14 . Chanarin , I. , A. Patel , G. Slavin , E. J. Wills , T. M. Andrews , and G. Stewart . 1975 . Neutral-lipid storage disease: a new disorder of lipid metabolism. BMJ . 1 : 553 – 555 .
15 . Dorfman , M. L. , C. Hershko , S. Eisenberg , and F. Sagher . 1974 . Ichthyosiform dermatosis with systemic lipidosis. Arch. Dermatol. 110 : 261 – 266 .
16 . Igal , R. A. , J. M. Rhoads , and R. A. Coleman . 1997 . Neutral lipid stor-age disease with fatty liver and cholestasis. J. Pediatr. Gastroenterol. Nutr. 25 : 541 – 547 .
17 . Oberer , M. , A. Boeszoermenyi , H. M. Nagy , and R. Zechner . 2011 . Recent insights into the structure and function of comparative gene identifi cation-58. Curr. Opin. Lipidol. 22 : 149 – 158 .
18 . Slavin , G. , E. J. Wills , J. E. Richmond , I. Chanarin , T. Andrews , and G. Stewart . 1975 . Morphological features in a neutral lipid storage disease. J. Clin. Pathol. 28 : 701 – 710 .
19 . Montero-Moran , G. , J. M. Caviglia , D. McMahon , A. Rothenberg , V. Subramanian , Z. Xu , S. Lara-Gonzalez , J. Storch , G. M. Carman , and D. L. Brasaemle . 2010 . CGI-58/ABHD5 is a coenzyme A-dependent lysophosphatidic acid acyltransferase. J. Lipid Res. 51 : 709 – 719 .
20 . Gruber , A. , I. Cornaciu , A. Lass , M. Schweiger , M. Poeschl , C. Eder , M. Kumari , G. Schoiswohl , H. Wolinski , S. D. Kohlwein , et al . 2010 . The N-terminal region of comparative gene identifi cation-58 (CGI-58) is important for lipid droplet binding and activation of adipose triglyceride lipase. J. Biol. Chem. 285 : 12289 – 12298 .
21 . Laemmli , U. K. 1970 . Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature . 227 : 680 – 685 .
22 . Yang , W. L. , and G. M. Carman . 1996 . Phosphorylation and regula-tion of CTP synthetase from Saccharomyces cerevisiae by protein kinase A. J. Biol. Chem. 271 : 28777 – 28783 .
23 . Boyle , W. J. , P. van der Geer , and T. Hunter . 1991 . Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional sepa-ration on thin-layer cellulose plates. Methods Enzymol. 201 : 110 – 149 .
24 . McMahon , D. , A. Dinh , D. Kurz , D. Shah , G. S. Han , G. M. Carman , and D. L. Brasaemle . 2014 . Comparative gene identifi cation 58/ � / � hydrolase domain lacks lysophosphatidic acid acyltransferase activity. J. Lipid Res. 55 : 1750 – 1761 .
is made between G0S2 inhibition and CGI-58 activation of lipase activity.
We have shown that Ser239 of CGI-58 is phosphorylated by PKA in vitro, as well as in tissue samples and intact cul-tured cells. Interestingly, 2D electrophoresis revealed three to four isoforms of CGI-58, suggesting additional posttranslational modifi cations. Additional studies are needed to address this hypothesis.
The authors thank Dr. Rosalind Coleman for generously providing human fi broblasts from an individual with NLSD; Dr. Dagmar Kratky for generously providing murine adipose tissues; Amy Marcinkiewicz and Dr. Vidya Subramanian for development of the adenoviral expression vector for perilipin; Roland Osen, Julia Kohlbacher, Sean Sullivan, Brittney Suchan, and Stefan Spoerk for expert technical assistance; and Dr. Diana Johnson and Mayda Hernandez for helpful discussions. A portion of this work was conducted while Anita Sahu-Osen was a visiting student in the Brasaemle laboratory.
REFERENCES
1 . Ahmadian , M. , Y. Wang , and H. S. Sul . 2010 . Lipolysis in adipo-cytes. Int. J. Biochem. Cell Biol. 42 : 555 – 559 .
2 . Girousse , A. , and D. Langin . 2012 . Adipocyte lipases and lipid drop-let-associated proteins: insight from transgenic mouse models. Int. J. Obes. (Lond) . 36 : 581 – 594 .
3 . Lass , A. , R. Zimmermann , M. Oberer , and R. Zechner . 2011 . Lipolysis - a highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid Res. 50 : 14 – 27 .
4 . Zechner , R. , P. C. Kienesberger , G. Haemmerle , R. Zimmermann , and A. Lass . 2009 . Adipose triglyceride lipase and the lipolytic ca-tabolism of cellular fat stores. J. Lipid Res. 50 : 3 – 21 .
5 . Brasaemle , D. L. 2007 . The perilipin family of structural lipid drop-let proteins: stabilization of lipid droplets and control of lipolysis. J. Lipid Res. 48 : 2547 – 2559 .
6 . Brasaemle , D. L. , V. Subramanian , A. Garcia , A. Marcinkiewicz , and A. Rothenberg . 2009 . Perilipin A and the control of triacylglycerol metabolism. Mol. Cell. Biochem. 326 : 15 – 21 .
7 . Brasaemle , D. L. , B. Rubin , I. A. Harten , J. Gruia-Gray , A. R. Kimmel , and C. Londos . 2000 . Perilipin A increases triacylglycerol storage by decreasing the rate of triacylglycerol hydrolysis. J. Biol. Chem. 275 : 38486 – 38493 .
8 . Subramanian , V. , A. Rothenberg , C. Gomez , A. W. Cohen , A. Garcia , S. Bhattacharyya , L. Shapiro , G. Dolios , R. Wang , M. P. Lisanti , et al . 2004 . Perilipin A mediates the reversible binding of CGI-58 to lipid droplets in 3T3-L1 adipocytes. J. Biol. Chem. 279 : 42062 – 42071 .
9 . Yamaguchi , T. , N. Omatsu , S. Matsushita , and T. Osumi . 2004 . CGI-58 Interacts with perilipin and is localized to lipid droplets: possible
TABLE 3. Protein colocalization analysis by determination of Manders coeffi cients
Expressed ProteinsColocalization of CGI-58 with
Perilipin 1A (M2)Colocalization of CGI-58 with Lipid
Droplets (M2)
CGI-58 Perilipin 1A Basal Stimulated Basal Stimulated
WT WT 0.86 ± 0.03 0.46 ± 0.03 0.86 ± 0.02 0.46 ± 0.03WT All6 0.98 ± 0.01 0.65 ± 0.03 0.96 ± 0.01 0.63 ± 0.03239A/240A WT 0.88 ± 0.02 0.65 ± 0.03 0.79 ± 0.03 0.62 ± 0.03239A/240A All6 0.83 ± 0.03 0.84 ± 0.04 0.80 ± 0.03 0.88 ± 0.03
Cos-7 cells expressing WT CGI-58 ( Fig. 6Ai, ii ; CGI-WT) or S239A/S240A-mutated CGI-58 ( Fig. 6Aiii, iv ; CGI-S239A/S240A), and WT perilipin 1A (Plin1A-WT) or perilipin 1A with mutations in six serine residues within PKA consensus sequences (Plin1A-all6) were incubated under basal or stimulated conditions before fi xation and staining for CGI-58, perilipin 1, nuclei, and lipid droplets. Micrographs were analyzed automatically with the ImageJ JACoP plugin. Colocalization of CGI-58 with either perilipin 1A or lipid droplets was assessed by determination of Manders coeffi cients 2 (M2) (mean ± standard error of the mean). An M2 of 0.00 indicates no colocalization, while 1.00 indicates complete colocalization.
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at:

CGI-58/ABHD5 is phosphorylated by protein kinase A 121
25 . Schweiger , M. , T. O. Eichmann , U. Taschler , R. Zimmermann , R. Zechner , and A. Lass . 2014 . Measurement of lipolysis. Methods Enzymol. 538 : 171 – 193 .
26 . Marcinkiewicz , A. , D. Gauthier , A. Garcia , and D. L. Brasaemle . 2006 . The phosphorylation of serine 492 of perilipin a directs lipid droplet fragmentation and dispersion. J. Biol. Chem. 281 : 11901 – 11909 .
27 . Igal , R. A. , and R. A. Coleman . 1996 . Acylglycerol recycling from triacylglycerol to phospholipid, not lipase activity, is defec-tive in neutral lipid storage disease fi broblasts. J. Biol. Chem. 271 : 16644 – 16651 .
28 . Igal , R. A. , P. Wang , and R. A. Coleman . 1997 . Triacsin C blocks de novo synthesis of glycerolipids and cholesterol esters but not recycling of fatty acid into phospholipid: evidence for functionally separate pools of acyl-CoA. Biochem. J. 324 : 529 – 534 .
29 . Igal , R. A. , and R. A. Coleman . 1998 . Neutral lipid storage disease: a genetic disorder with abnormalities in the regulation of phospho-lipid metabolism. J. Lipid Res. 39 : 31 – 43 .
30 . Orlicky , D. J. , J. DeGregori , and J. Schaack . 2001 . Construction of stable coxsackievirus and adenovirus receptor-expressing 3T3-L1 cells. J. Lipid Res. 42 : 910 – 915 .
31 . Blanchette-Mackie , E. J. , N. K. Dwyer , T. Barber , R. A. Coxey , T. Takeda , C. M. Rondinone , J. L. Theodorakis , A. S. Greenberg , and C. Londos . 1995 . Perilipin is located on the surface layer of intra-cellular lipid droplets in adipocytes. J. Lipid Res. 36 : 1211 – 1226 .
32 . Gocze , P. M. , and D. A. Freeman . 1994 . Factors underlying the variability of lipid droplet fl uorescence in MA-10 Leydig tumor cells. Cytometry . 17 : 151 – 158 .
33 . Bolte , S. , and F. P. Cordelieres . 2006 . A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 224 : 213 – 232 .
34 . Schwartz , D. M. , and N. E. Wolins . 2007 . A simple and rapid method to assay triacylglycerol in cells and tissues. J. Lipid Res. 48 : 2514 – 2520 .
35 . Brasaemle , D. L. , T. Barber , N. E. Wolins , G. Serrero , E. J. Blanchette-Mackie , and C. Londos . 1997 . Adipose differentiation-related protein is an ubiquitously expressed lipid storage droplet-associated protein. J. Lipid Res. 38 : 2249 – 2263 .
36 . Hulo , N. , A. Bairoch , V. Bulliard , L. Cerutti , E. De Castro, P. S. Langendijk-Genevaux, M. Pagni, and C. J. Sigrist. 2006 . The PROSITE database. Nucleic Acids Res. 34 : D227 – D230 .
37 . Granneman , J. G. , H. P. Moore , R. L. Granneman , A. S. Greenberg , M. S. Obin , and Z. Zhu . 2007 . Analysis of lipolytic protein traffi ck-ing and interactions in adipocytes. J. Biol. Chem. 282 : 5726 – 5735 .
38 . Brasaemle , D. L. , T. Barber , A. R. Kimmel , and C. Londos . 1997 . Post-translational regulation of perilipin expression. Stabilization by stored intracellular neutral lipids. J. Biol. Chem. 272 : 9378 – 9387 .
39 . Brasaemle , D. L. , D. M. Levin , D. C. Adler-Wailes , and C. Londos . 2000 . The lipolytic stimulation of 3T3–L1 adipocytes promotes the translocation of hormone-sensitive lipase to the surfaces of lipid storage droplets. Biochim. Biophys. Acta . 1483 : 251 – 262 .
40 . Egan , J. J. , A. S. Greenberg , M. K. Chang , S. A. Wek , M. C. Moos , Jr ., and C. Londos . 1992 . Mechanism of hormone-stimulated lipolysis in adipocytes: translocation of hormone-sensitive lipase to the lipid storage droplet. Proc. Natl. Acad. Sci. USA . 89 : 8537 – 8541 .
41 . Su , C. L. , C. Sztalryd , J. A. Contreras , C. Holm , A. R. Kimmel , and C. Londos . 2003 . Mutational analysis of the hormone-sensitive lipase translocation reaction in adipocytes. J. Biol. Chem. 278 : 43615 – 43619 .
42 . Sztalryd , C. , G. Xu , H. Dorward , J. T. Tansey , J. A. Contreras , A. R. Kimmel , and C. Londos . 2003 . Perilipin A is essential for the trans-location of hormone-sensitive lipase during lipolytic activation. J. Cell Biol. 161 : 1093 – 1103 .
43 . Krintel , C. , P. Osmark , M. R. Larsen , S. Resjo , D. T. Logan , and C. Holm . 2008 . Ser649 and Ser650 are the major determinants of protein kinase A-mediated activation of human hormone-sensitive lipase against lipid substrates. PLoS ONE . 3 : e3756 .
44 . Krintel , C. , M. Morgelin , D. T. Logan , and C. Holm . 2009 . Phosphorylation of hormone-sensitive lipase by protein kinase A in vitro promotes an increase in its hydrophobic surface area. FEBS J. 276 : 4752 – 4762 .
45 . Gandotra , S. , K. Lim , A. Girousse , V. Saudek , S. O'Rahilly , and D. B. Savage . 2011 . Human frame shift mutations affecting the carboxyl terminus of perilipin increase lipolysis by failing to sequester the ad-ipose triglyceride lipase (ATGL) coactivator AB-hydrolase-containing 5 (ABHD5). J. Biol. Chem. 286 : 34998 – 35006 .
46 . Greenberg , A. S. , J. J. Egan , S. A. Wek , N. B. Garty , E. J. Blanchette-Mackie , and C. Londos . 1991 . Perilipin, a major hormonally regulated adipocyte-specifi c phosphoprotein associated with the periphery of lipid storage droplets. J. Biol. Chem. 266 : 11341 – 11346 .
47 . Servetnick , D. A. , D. L. Brasaemle , J. Gruia-Gray , A. R. Kimmel , J. Wolff , and C. Londos . 1995 . Perilipins are associated with cho-lesteryl ester droplets in steroidogenic adrenal cortical and Leydig cells. J. Biol. Chem. 270 : 16970 – 16973 .
48 . Kraemer , F. B. , V. K. Khor , W. J. Shen , and S. Azhar . 2013 . Cholesterol ester droplets and steroidogenesis. Mol. Cell. Endocrinol. 371 : 15 – 19 .
49 . Granneman , J. G. , H. P. Moore , E. P. Mottillo , and Z. Zhu . 2009 . Functional interactions between Mldp (LSDP5) and Abhd5 in the control of intracellular lipid accumulation. J. Biol. Chem. 284 : 3049 – 3057 .
50 . Dalen , K. T. , T. Dahl , E. Holter , B. Arntsen , C. Londos , C. Sztalryd , and H. I. Nebb . 2007 . LSDP5 is a PAT protein specifi cally ex-pressed in fatty acid oxidizing tissues. Biochim. Biophys. Acta . 1771 : 210 – 227 .
51 . Wolins , N. E. , B. K. Quaynor , J. R. Skinner , A. Tzekov , M. A. Croce , M. C. Gropler , V. Varma , A. Yao-Borengasser , N. Rasouli , P. A. Kern , et al . 2006 . OXPAT/PAT-1 is a PPAR-induced lipid droplet protein that promotes fatty acid utilization. Diabetes . 55 : 3418 – 3428 .
52 . Yamaguchi , T. , S. Matsushita , K. Motojima , F. Hirose , and T. Osumi . 2006 . MLDP, a novel PAT family protein localized to lipid droplets and enriched in the heart, is regulated by peroxisome proliferator-activated receptor alpha. J. Biol. Chem. 281 : 14232 – 14240 .
53 . Wang , H. , M. Bell , U. Sreenevasan , H. Hu , J. Liu , K. Dalen , C. Londos , T. Yamaguchi , M. A. Rizzo , R. Coleman , et al . 2011 . Unique regulation of adipose triglyceride lipase (ATGL) by perilipin 5, a lipid droplet-associated protein. J. Biol. Chem. 286 : 15707 – 15715 . [Erratum. 2013. J. Biol. Chem. 288: 10952.]
54 . Lu , X. , X. Yang , and J. Liu . 2010 . Differential control of ATGL-mediated lipid droplet degradation by CGI-58 and G0S2. Cell Cycle . 9 : 2719 – 2725 .
55 . Yang , X. , X. Lu , M. Lombes , G. B. Rha , Y. I. Chi , T. M. Guerin , E. J. Smart , and J. Liu . 2010 . The G(0)/G(1) switch gene 2 regulates adipose lipolysis through association with adipose triglyceride li-pase. Cell Metab. 11 : 194 – 205 .
by guest, on February 3, 2015
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2014/11/24/jlr.M055004.DC1Supplemental Material can be found at: