issn 2321-9122 studies on production, purification and
TRANSCRIPT

~ 1 ~
European Journal of Biotechnology and Bioscience 2014; 1 (5): 1-6
ISSN 2321-9122 EJBB 2014; 1 (5): 1-6 Received 03-04-2014 Accepted: 26-04-2014
M.P.Kusuma R.B.V.R.R Womens College of Pharmacy, Osmania University, Andhra Pradesh, India
M.Vijaya Bhargavi R.B.V.R.R Womens College of Pharmacy, Osmania University, Andhra Pradesh, India.
Correspondence:
M.P.Kusuma Assistant Professor, R.B.V.R.R Womens College of Pharmacy, Osmania University, Andhra Pradesh, India Email: [email protected] Tel: +919441517420
Studies on production, purification and molecular characterisation of polygalacturonase from
Bacillus subtilis
M.P.Kusuma, M.Vijaya Bhargavi
ABSTRACT Polygalacturonases are responsible for the degradation of the long and complex molecules called pectin that occur as structural polysaccharides in the middle lamella and the primary call walls of young plant cells. Pectinases are an integral part of fruit juice and textile industries as well as having various biotechnological applications. Polygalacturonase was produced by SMF using an isolate Bacillus subtilis. The enzyme was purified and the purity was judged by SDS PAGE. A preliminary structural and physico- chemical characterization of polygalacturonase has been performed by N terminal sequencing, accurate molecular mass determination by MALDI-TOF mass spectrometry and secondary structure analysis by circular dichroism.
Keywords: Polygalacturonase, N terminal sequencing and Circular dichroism.
1. Introduction Pectinases were some of the first enzymes to be used in homes. Their commercial application was first observed in 1930 for the preparation of wines and fruit juices. Only in the 1960s did the chemical nature of plant tissues become apparent and with this knowledge, scientists began to use a greater range of enzymes more efficiently. As a result, pectinases are today one of the upcoming enzymes of the commercial sector. Primarily, these enzymes are responsible for the degradation of the long and complex molecules called pectin that occur as structural polysaccharides in the middle lamella and the primary call walls of young plant cells. Pectinases are now an integral part of fruit juice and textile industries as well as having various biotechnological applications. Chemically, pectic substances are complex colloidal acid polysaccharides, with a backbone of galacturonic acid residues linked by a (1±4) linkages. The side chains of the pectin molecule consist of L-rhamnose, arabinose, galactose and xylose. The carboxyl groups of galacturonic acid are partially esterified by methyl groups and partially or completely neutralized by sodium, potassium or ammonium ions. Based on the type of modifications of the backbone chain, pectic substances are classified into protopectin, pectic acid, pectinic acid and pectin (Miller, 1986). Polygalacturonases (PG). Catalyze hydrolysis ofa-1, 4-glycosidic linkages in pectic acid (polygalacturonic acid). In this work polygalacturonase was produced by SMF using an isolate Bacillus subtilis. The enzyme was purified and the purity was judged by SDS PAGE. A preliminary structural and physico-chemical characterization of polygalacturonase has been performed by N terminal sequencing, accurate molecular mass determination by MALDI-TOF mass spectrometry and secondary structure analysis by circular dichroism.
2. Material and methods 2.1 Bacterial isolation The bacteria was isolated from the soil and pure cultures were prepared. The isolated pure cultures were screened for extracellular polygalacturonase using agar media containing pectin as a substrate.

~ 2 ~
European Journal of Biotechnology and Bioscience
2.2 Polygalacturonase assay Polygalacturonase activity was assayed by the colorimetric method of Miller (1959). 0.5 mL of supernatant was incubated with 1.0% of pectin at 40 0C for 10 min under static conditions. After adding 3 mL of di nitro salicylic acid, the mixture was boiled for 15 min and finally diluted to 5 mL with distilled water. The absorbance was measured at 540 nm.
2.3 Purification of polygalacturonase 150 mL of cell free supernatant was fractionated by precipitation with ammonium sulphate between 30-90% of saturation. The saturated solution was left overnight at 4 0C, centrifuged at 12,000 rpm for 20 min, dissolved in minimal amount of 10 mm Tris HCl buffer at pH 8 and dialyzed against the same buffer for 24 h at 4 0C [2]. The enzyme sample was subjected Ion exchange chromatography [3] using anion exchanger, DEAE Sephacel was packed into a glass column (15 X 0.55 mm, 10 mL bed volume). The column was equilibrated with Tris HCl buffer (20 mm, pH7.5) and 3.0 mL sample was loaded onto it. The column was washed with Tris HCl buffer containing 20, 30, 40, 50, 100 and 150 mm NaCl concentration. Fractions of 2.5 mL volume were collected. The protein content of the fractions was measured spectrophotometrically at 280 nm and the polygalacturonase activity was assayed by the method described earlier. The fraction showing polygalacturonase activity was pooled, concentrated and saved for further analysis. Sodium do decy l sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out in 3 mm slab gel of 6% acrylamide in Tris borate buffer at pH7.1 containing 0.1% SDS. The gels were stained with Coomassie brilliant blue R-250 and destained by keeping in destaining solution [4].
2.4 N-Terminal Amino Acids Sequencing by Edman Degradation PG, separated by SDS-PAGE, was transferred onto a PVDF membrane and directly subjected to Edman degradation [5, 6].
2.5 MALDI-TOF Mass Spectrometry of Native Protein Mass spectrometry analysis of native protein was performed with a MALDI-TOF micro MX (Waters Co., Manchester, UK) equipped with a pulsed nitrogen laser (_=337 nm) [7]. The instrument source voltage was set to 12 kV. The pulse voltage was optimized at 1999 V and the detector voltage was set to 5200 V. Measurements were performed in the mass range m/z 10000-60000, with a suppression mass gate set to m/z 3000 to prevent detector saturation from matrix cluster peaks and with an extraction delay of 600 ns. Prior to the acquisition of spectra, 1L of protein solution (10 pmol/_L) was mixed with 1L of saturated sinapinic acid matrix solution [10 mg/mL in acetonitrile: water containing 0.1% trifluoroacetic acid (1: 1.5; v: v)] and a droplet of the resulting mixture (1L) placed on the mass spectrometers. Once the liquid was completely evaporated, the sample was loaded into the mass spectrometer and analyzed in positive acquisition linear mode. The instrument was calibrated with a three points external calibration by using an appropriate mix of 10 pmol/L of insulin, cytochrome-c and trypsinogen as standard proteins (Sigma, St. Louis, MO, USA). A mass accuracy near to the nominal (300 ppm) was achieved for each standard. All spectra were processed and analyzed using the
Mass Lynx 4.1 software (Waters, Milford, MA USA).
2.6 Homology Studies Amino acid sequences of PG isolated from different species were retrieved and analyzed by using the BLAST p software and the NCBI taxonomy browser available on-line at the NBCI web site (http://blast.ncbi.nlm.nih.gov/ Blast.cgi). The following amino acid sequences were selected for the analysis: PG from Bordetella avium 197N, AC: 187479388; PG from Bordetella petrii DSM 12804, AC: 163854958; PG from Beutenbergia cavernae DSM 12333, AC: 229821715 PG from Methylibium petroleiphilum PM1, AC: 124268476; PG from Verminephrobacter aporrectodeae, AC347818708; PG from Frankia alni ACN14a AC: 111225564; PG from Variovorax paradoxus EPS, AC: 319793145; PG from Ramlibacter tataouinensis, AC: 337279172; The alignment was performed by using the ClustalW software in the default set-up (http://www.ebi.ac.uk/Tools/ msa/clustalw2/). The SYSTERS Protein Family Database was searched for protein family classification (http:// systers. molgen.mpg.de/) [8].
2.7 Circular Dichroism Spectroscopy CD spectra were obtained at room temperature on a Jasco J-815 dichrograph [Jasco Europe, Cremella (LC) Italy] [10, 11]. For spectra in the near-and far UV, measures were performed with a protein concentration of 0.1 mg/mL in 100 mm Na-acetate buffer, pH 4.6, using a 0.1 cm path-length quartz cuvette. The K2D2 curve-fitting programme software (http://www.ogic.ca/projects/k2d2/) was used to estimate the percentages of secondary structural elements [12].
3. Results and Discussion 3.1 Bacterial isolation A total of 66 colonies were isolated from four soil samples, on the basis of macroscopic characters and microscopic observations, eliminating those that appeared close to each other. All 66 isolated colonies were tested for their polygalacturonase potential using plate assay. Of the 66 isolated colonies tested, only eight bacterial strains shown the polygalacturonase potential. Of the 8 positive isolates, C4 was showing relatively high activity. After performing microscopic and biochemical characterization the isolate was confirmed to be Bacillus subtilis. This isolate was used for production of polygalacturonase by submerged fermentation.
3.2 Purification The extracted enzyme was subjected to partial purification which included ammonium sulphate precipitation, dialysis, ion exchange chromatography and affinity chromatography. The cell free supernatant polygalacturonase activity was found to be in the 30- 40% salt saturation fraction. Specific activity of polygalacturonase increased from 33.68U/mL to 93.05 U/mL. Further the fraction was loaded onto anion exchanger, DEAE Sephacel equilibrated with 20 mm Tris HCl (pH 7.5 buffer). The proteins were eluted with NaCl gradient. 9.84 fold purification of the enzyme was achieved and its specific activity was found to be 915.6U/Ml. The homogeneity of the polygalacturonase was judged by SDS-PAGE. It was found that PGs had a molecular weight ranging between 43 kDa and 66 kDa (Fig. 1).
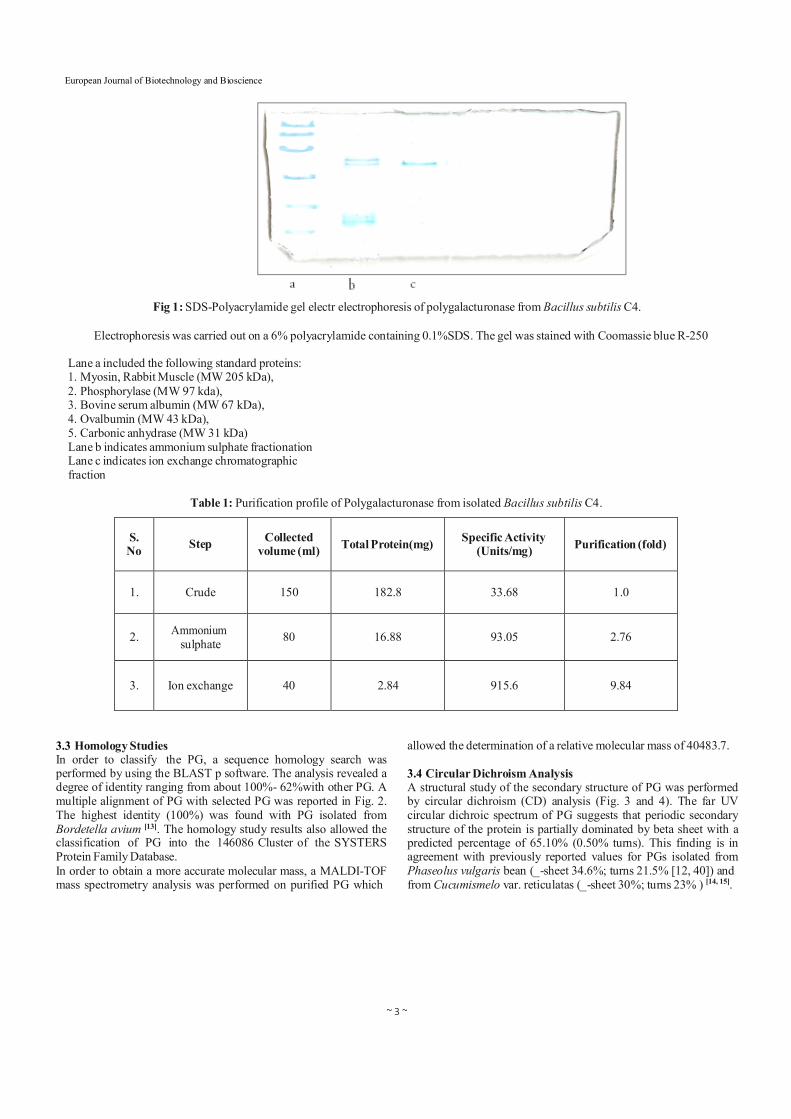
~ 3 ~
European Journal of Biotechnology and Bioscience
Fig 1: SDS-Polyacrylamide gel electr electrophoresis of polygalacturonase from Bacillus subtilis C4.
Electrophoresis was carried out on a 6% polyacrylamide containing 0.1%SDS. The gel was stained with Coomassie blue R-250
Lane a included the following standard proteins: 1. Myosin, Rabbit Muscle (MW 205 kDa), 2. Phosphorylase (MW 97 kda), 3. Bovine serum albumin (MW 67 kDa), 4. Ovalbumin (MW 43 kDa), 5. Carbonic anhydrase (MW 31 kDa) Lane b indicates ammonium sulphate fractionation Lane c indicates ion exchange chromatographic fraction
Table 1: Purification profile of Polygalacturonase from isolated Bacillus subtilis C4.
S. No
Step
Collected
volume (ml)
Total Protein(mg)
Specific Activity
(Units/mg)
Purification (fold)
1.
Crude
150
182.8
33.68
1.0
2. Ammonium
sulphate
80
16.88
93.05
2.76
3.
Ion exchange
40
2.84
915.6
9.84
3.3 Homology Studies In order to classify the PG, a sequence homology search was performed by using the BLAST p software. The analysis revealed a degree of identity ranging from about 100%- 62%with other PG. A multiple alignment of PG with selected PG was reported in Fig. 2. The highest identity (100%) was found with PG isolated from Bordetella avium [13]. The homology study results also allowed the classification of PG into the 146086 Cluster of the SYSTERS Protein Family Database. In order to obtain a more accurate molecular mass, a MALDI-TOF mass spectrometry analysis was performed on purified PG which
allowed the determination of a relative molecular mass of 40483.7.
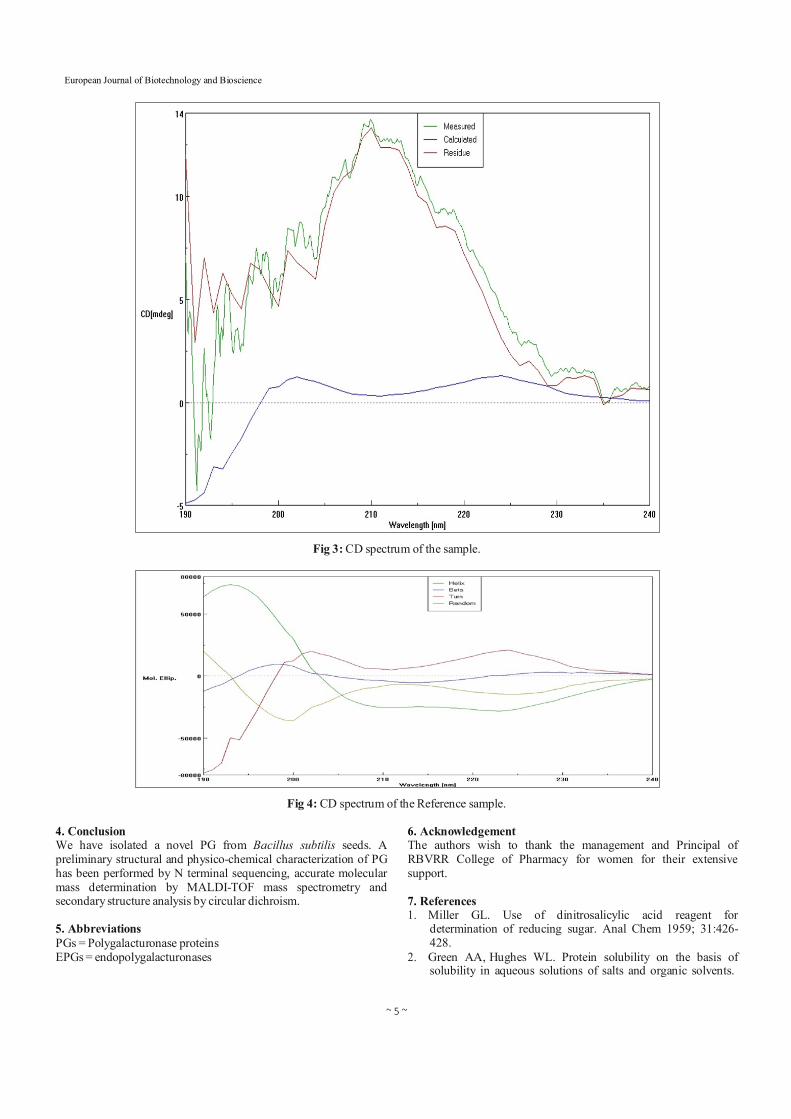
3.4 Circular Dichroism Analysis A structural study of the secondary structure of PG was performed by circular dichroism (CD) analysis (Fig. 3 and 4). The far UV circular dichroic spectrum of PG suggests that periodic secondary structure of the protein is partially dominated by beta sheet with a predicted percentage of 65.10% (0.50% turns). This finding is in agreement with previously reported values for PGs isolated from Phaseolus vulgaris bean (_-sheet 34.6%; turns 21.5% [12, 40]) and from Cucumismelo var. reticulatas (_-sheet 30%; turns 23% ) [14, 15].

~ 4 ~
European Journal of Biotechnology and Bioscience
Fig 2: Multiple sequence alignment of PG using CLUSTAL W.
Table 2: Results of the sample spectrum camparing to reference spectrum.
Fraction Ratio
Helix: 3.50%
Turn: 0.50%
Beta: 65.10%
Random: 30.90%
Total: 100.00%
RMS: 38.811
~ 5 ~
European Journal of Biotechnology and Bioscience
Fig 3: CD spectrum of the sample.
Fig 4: CD spectrum of the Reference sample.
4. Conclusion We have isolated a novel PG from Bacillus subtilis seeds. A preliminary structural and physico-chemical characterization of PG has been performed by N terminal sequencing, accurate molecular mass determination by MALDI-TOF mass spectrometry and secondary structure analysis by circular dichroism.
5. Abbreviations PGs = Polygalacturonase proteins EPGs = endopolygalacturonases
6. Acknowledgement The authors wish to thank the management and Principal of RBVRR College of Pharmacy for women for their extensive support.
7. References 1. Miller GL. Use of dinitrosalicylic acid reagent for
determination of reducing sugar. Anal Chem 1959; 31:426- 428.
2. Green AA, Hughes WL. Protein solubility on the basis of solubility in aqueous solutions of salts and organic solvents.

~ 6 ~
European Journal of Biotechnology and Bioscience
Methods Enzymol 1955; 1:67-90. 3. Hayrunnisa N, esen T, Ahmet A, Medine G, Nazan D.
Production of a novel pectin lyase from Bacillus pumilus (P9), purification and characterisation and fruit juice application, Rom. Biotech Lett 2010; 15:5167-5175.
4. Stagemann H. SDS-PAGE, Merits and limits. In: Electrokinetic separation methods. Righetti P G Van Oss CJ and Vanderhoffeds JW (eds), Elsevier North Holland Amsterdam 1979; 313-336.
5. Di Maro A, Terracciano I, Sticco L, Fiandra L, Ruocco M, Corrado G. et al. Purification and characterization of a viral chitinase active against plant pathogens and herbivoresfrom transgenic tobacco J Biotechnol 2010; 147:1-6.
6. Di Maro A, Chambery A, Carafa V, Costantini S, Colonna G, Parente A. Structural characterization and comparative modeling of PD-Ls 1-3, type 1 ribosome-inactivating proteins from summer leaves of Phytolacca dioica L. Biochimie 2009; 91:352-363.
7. Chambery A, Di Maro A, Parente A. Primary structure and glycan moiety characterization of PD-Ss, type 1 ribosomeinactivating proteins from Phytolacca dioica L. seeds, by precursor ion discovery on a Q-TOF mass spectrometer. Phytochemistry 2008; 69:1973-1982.
8. Krause A, Stoye J, Vingron M. Large scale hierarchical clustering of protein sequences. BMC Bioinformatics 2005; 6- 15.
9. Meinel T, Krause A, Luz H, Vingron M, Staub E. The SYSTERS Protein Family Database in 2005. Nucleic Acids Res 2005; 33:226-229.
10. Johnson WC, Jr Protein secondary structure and circular dichroism: a practical guide. Proteins 1990; 7:205-214.
11. Johnson WC, Jr Analysis of circular dichroism spectra. Methods Enzymol 1992; 210:426-447.
12. Perez-Iratxeta C, Andrade-Navarro MA. K2D 2. Estimation of protein secondary structure from circular dichroism spectra. BMC Struct Biol 2008; 8-25.
13. Simpson CG, MacRae E, Gardner RC. Cloning of a Polygalacturonase Inhibiting Protein from Kiwifruit (Actinidia deliciosa). Plant Physiol 1995; 108:1748-1748.
14. Fish WW, Davis AR. The purification, physical/chemical characterization, and c DNA sequence of cantaloupe fruit polygalacturonase-inhibiting protein. Phytopathology, 2004; 94:337-344.
15. Fish WW, Madihally SV. Modeling the inhibitor activity and relative binding affinities in enzyme-inhibitor-protein systems: application to developmental regulation in a PG-PGIP system. Biotechnol Prog 2004; 20:721-727.