isolation, characterization and molecular phylogeny of ... that the quality of ganga water of two...
TRANSCRIPT

Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
ENVIRONMENTAL CHEMISTRY, POLLUTION & WASTE MANAGEMENT | RESEARCH ARTICLE
Isolation, characterization and molecular phylogeny of multiple metal tolerant and antibiotics resistant bacterial isolates from river Ganga, Varanasi, IndiaNiveshika1, Savita Singh1, Ekta Verma1 and Arun Kumar Mishra1*
Abstract: The present study was focused on the isolation of multiple metal tolerant and antibiotics resistant bacterial strains from water samples of five different Ghats of river Ganga, Varanasi, India. These strains were biochemically characterized and their phyloge-netic relatedness was assumed using amplified ribosomal DNA restriction analysis finger-printing and 16S ribosomal gene sequencing. The presence of heterogeneous groups of bacteria belonging to alpha, beta, gamma proteobacteria, and bacilli was noticed. Some of the bacterial strains like Pseudomonas, Serratia, Enterobacter, and Proteus vulgaris were mainly found at the Dashashwamedh Ghat and the Assi Ghat showing minimum inhibi-tory concentration 200–300 mg/L for copper, nickel, lead, and chromium. Comamonas sp. mainly isolated from the Samne Ghat and the Rajendra Prasad Ghat was able to grow at very high concentration of lead viz. 400 mg/L. Some of the strains showed multidrug resis-tant property against 10 different antibiotics which are of most serious concern because these drugs are frequently used against various bacterial infections.
Subjects: Environmental & Agriculture; Bioscience; Environmental Studies & Management
Keywords: bacterial isolates; multiple metal tolerant; antibiotics resistant; amplified ribosomal DNA restriction analysis; phylogeny
*Corresponding author: Arun Kumar Mishra, Laboratory of Microbial Genetics, Department of Botany, Banaras Hindu University, Varanasi 221005, IndiaE-mails: [email protected], [email protected]
Reviewing editor:Lian Pin Koh, University of Adelaide, Australia
Additional information is available at the end of the article
ABOUT THE AUTHORSNiveshika is a research scholar in Department of Botany, Banaras Hindu University, Varanasi, India. Her research area includes Molecular Phylogeny of bacteria and cyanobacteria.
Savita Singh, completed her PhD from Department of Botany, Banaras Hindu University, Varanasi, India. Presently she is working as a senior research fellow at Indian Council of Agriculturally Important Micro-organisms, Mau, Uttar Pradesh, India. She has expertise in the field of Salt Stress Biology of cyanobacteria.
Ekta Verma is working as a research scholar in Department of Botany, Banaras Hindu University, Varanasi, India. Her research area includes Transcriptomics and Proteomics of cyanobacteria.
Arun Kumar Mishra, professor at Department of Botany, Banaras Hindu University, Varanasi, India. He has 17 years of teaching and 25 years of research experience with specialization in Microbial Physiology, Biochemistry, and Genetics.
PUBLIC INTEREST STATEMENTMixing of sewage along with industrial effluents into the Ganga river perturb water quality. Polluted water became a habitat of various pathogenic bacteria having dual properties in rendering multiple heavy metals tolerant capacity as well as antibiotics resistance. The present study was focused on the isolation of multiple metal tolerant and antibiotics resistant bacterial strains from water samples of five different Ghats of river Ganga, Varanasi, India. These strains were biochemically characterized and their phylogenetic relatedness was assumed using amplified ribosomal DNA restriction analysis fingerprinting and 16S ribosomal gene sequencing. Collaboration of physiological, biochemical, and molecular techniques could provide complete information about bacterial identification and characterization. Presence of harmful bacteria in the river Ganga invites serious attention from human health perspective since Ganga water is also used for drinking, holy bathing and irrigation purposes.
Received: 30 June 2016Accepted: 14 December 2016First Published: 23 December 2016
Page 1 of 17
Niveshika
© 2017 The Author(s). This open access article is distributed under a Creative Commons Attribution (CC-BY) 4.0 license.

Page 2 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
1. IntroductionVaranasi, India (25°20′ N, 83°00′ E) lies 8.77 m above sea level in the eastern Gangetic plains of India has at least 84 Ghats. Ghats are riverfront steps leading to the banks of river Ganga. The holy river Ganges and its Ghats are an abode for thousands of natives besides being a place visited by 300 people every hour in the mornings, and on festival days 2,500 people arrive per hour. According to World Health Organization (WHO) reports, in developing countries like India, almost 80% of the total water is being polluted by discharge of domestic sewage and untreated industrial effluents (Sangu & Sharma, 1987). The WHO reported (World Health Organization [WHO], 2002) that infectious dis-ease like diarrhea, caused due to the intake of unsafe and extremely polluted water, is responsible for about 1.7 million deaths a year worldwide. Recently Times of India New Delhi September 2014 reported that the quality of Ganga water of two holy places Varanasi and Allahabad where millions take a dip everyday are most unfit for bathing purpose.
Water of river Ganga at Varanasi region is mainly polluted by industrial effluents, domestic sew-age, and disposal of dead bodies (Mishra & Tripathi, 2007; Pandey, Shubhashish, & Pandey, 2010). About 1,503 industries (Chemical industries, metal processing industries, leather, and textile indus-tries) are located in and around Varanasi (Anonymous, 2006; Industrial Directory, 1981; Rai & Tripathi, 2008). Escalating anthropogenic activities, indiscriminate urbanization, and accelerated industrialization gave birth to the river water pollution particularly heavy metals and microbial pol-lution (Rai, Mishra, & Tripathi, 2010). Either directly or indirectly metals have been known to play a major role in almost all metabolic processes, growth, and development of the microorganisms (Bergey & Breed, 1994; Beveridge & Doyle, 1989). However, these organisms are forced to adapt the increasing concentrations of metals beyond their normal tolerance level. Therefore, microbes have developed various biological mechanisms like complexation, metal efflux systems, utilization or re-duction of the metal during anaerobic respiration in order to tolerate heavy metal stress (Ehrlich & Brierley, 1990). Metal resistant bacteria have the ability to grow under high concentrations of metals and can play an incredible role in the bioremediation of those metals (Bolan et al., 2014).
Introduction of antimicrobial agents such as antibiotics into the river and streams from industrial pollution as metals, emerged as an ever increasing health hazard due to spread of antibiotic resistant organism in water environment (Baquero, Martínez, & Cantón, 2008; Levy, 1997). Ability of different species of bacteria in acquiring multidrug resistance (MDR) has led to severe complication for human beings (Tenover & McGowan, 1996). A large proportion of MDR was found to contain plasmid borne mobile resistance genes that can transmit through bacterial populations (Kumarasamy et al., 2010).
Molecular techniques have initiated a new era of microbiology and may prove to be helpful in de-termining the inhabitants of freshwater microbial communities (Mukherjee, Kumar, Kumar, & Chakraborthy, 2013). Sequences based on 16S ribosomal gene reveal numerous cultured as well as uncultured microorganisms in diverse environments as 16S ribosomal RNA (16S rRNA) molecules contain both highly conserved as well as variable regions (Woese, 1987). In the recent few years, amplified ribosomal DNA restriction analysis (ARDRA) has become a useful tool for assessment of microbial community structure changes as it is a simple, fast and reliable method for evaluation of bacterial diversity in different environments (Martínez-Murcia, Acinas, & Rodriguez-Valera, 1995).
In the present communication, the water quality of river Ganga has been analyzed using physico-chemical properties. Isolation and biochemical characterization of multiple metal tolerant and anti-biotics resistant bacteria have been investigated. Molecular characterization of isolates based on 16S rDNA gene sequences and ARDRA fingerprints were performed. The significance of the proposed study has been ascertained by applying statistical parameters such as pearson correlation/regres-sion and principal component analysis (PCA) analysis.

Page 3 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
2. Materials and methods
2.1. Sample collection and physicochemical analysisThe areas selected for the present study are based on the human use and pollution. Water samples were collected from the bank of five Ghats (5 km area) viz. Dhashashwamedh ghat, Harishchandra ghat, Samne ghat, Rajendra Prasad Ghat and Assi Ghat. Water samples (1 L) were collected monthly during the first week of each month from January 2014 to December 2014 from 15 cm depth at 8.00–10.00 am near bank of each Ghat in three replicate in 1 L acid washed glass bottles. Temperature, pH, and conductivity were measured on the spot using a thermometer, a pH-meter and a conductiv-ity meter. Samples were then transported to the laboratory under sterile condition for isolation of bacteria and analysis of dissolved oxygen (DO), biochemical oxygen demand (BOD), total hardness (TH), total alkalinity (TA), calcium (Ca2+), magnesium (Mg2+), sulfate (SO4
−), total dissolved solids (TDS), total nitrogen (TN) and total phosphorus (TP), the latter estimated using the American Public Health Association [APHA] (1998) method. Estimation of TN content was performed by following the micro Kjeldahl technique using a Gerhardt automatic analyser (model KB8S Kjeldatherm, Germany). Titrimetric methods were for testing DO, BOD and TA whereas a gravimetric method was used for TDS (Vogel, 1978). Turbidometric method was used for estimation of SO4
−2 (Vogel, 1978). An ethylen-ediaminetetraacetic acid titrimetric method was used to estimate Ca2+, Mg2+, and TH (Vogel, 1978). An atomic absorption spectrophotometer (Model 2380, Perkin Elmer, Inc. Norwalk, CT, USA), was used for determination of Cr, Cd, Ni, Pb, Zn, and Cu. A Pearson correlation matrix was constructed by using SPSS software version 16.0 in order to study correlation among physicochemical parameters at five Ghats of river Ganga Varanasi, India.
2.2. Isolation and maintenance of bacterial isolatesIn order to get all types of bacterial strains, water samples from each Ghats were concentrated through centrifugation at 10,000 rpm for 15 min. Pellet was dissolved in 100 μL sterilized distilled water and spread on nutrient agar (NA) media containing 0.5% peptone, 0.3% beef extract/yeast extract, 1.5% agar, 0.5% NaCl, and incubated at 37°C. Numerous colonies were obtained on NA plates. The single bacterial colonies showing unique morphology were isolated and subcultured at least five times to ascertain purity of the isolates. The purity of the isolates was monitored routinely by microscopic analysis. Purified bacterial isolates were subjected to the biochemical and molecular identification tests.
2.3. Metal tolerance assays
2.3.1. Minimum inhibitory concentration of metalsThe isolated pure bacterial cultures were checked for their respective minimum inhibitory concen-trations (MICs) towards all the metal ions. Different concentrations of metals ions such as Zn, Cd, Cu, Ni, Pb, and Cr (0, 25, 50, and 100 mg/L) were added separately against each bacterial isolates incu-bated in solid NA medium and then kept for 24–48 h at 37°C in incubator. Those bacterial isolates, in which growth was visible up to 100 mg/L were further grown at higher metal concentration ranging from 150, 200, 250, 300, 350 to 400 mg/L. At that metal concentration where bacterial isolates failed to grow on plates even after 7 days of incubation were considered as the MIC for that particu-lar bacterial strain (Narasimhulu, Rao, & Venu Vinod, 2010).
2.4. Antibiotics resistance assayBacterial strains were checked for antibiotics resistance pattern against ampicillin (Amp), chloram-phenicol (Chl), nalidixic acid (Nal), tetracyclin (Tet), vancomycin (Van), ciprofloxacin (Cip), imipenem (Imi), erythromycin (Ery), streptomycin (Str), and polymixin (Pol) (Himedia, India) according to Kirby-Bauer disk diffusion method (Bauer, Kirby, Sherris, & Turck, 1966). A volume of 5 mL of nutrient broth was prepared for inoculating bacterial strains which were incubated overnight at 37°C. The pure broth cultures were then swabbed over the sterilized 20 mL Mueller Hinton pre-made agar plates. Disks of 6 mm diameter were prepared from sterilized Whatman filter paper. The 10 antibiotics at concentrations of 30 mg/L were delivered in 10 disks which were then dispensed on each of the

Page 4 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
swabbed plates at appropriate distances from each other. Plates were then incubated at 37°C for 24 h. Zones of inhibition were measured in diameter across the centre of each zone in millimeters.
2.5. Physiological characterization of isolated bacteriaGram staining was performed for differentiating Gram-positive and Gram-negative bacteria based on the differences in bacterial cell wall peptidoglycan layer (thick layer present in gram positive).
2.6. Biochemical assay
2.6.1. Citrate testA volume of 100 mL Simmon citrate agar medium (15 g/L agar, 0.2 g/L ammonium dihydrogen phosphate, 0.08 g/L bromothymol blue, 0.8 g/L disodium ammonium phosphate, 0.2 g/L magnesium sulfate heptahydrate, 5 g/L sodium chloride, 2 g/L trisodium citrate and 15 g/L agar) pH 7.0 was prepared and then autoclaved. The sterilized Simmon citrate agar medium was poured in different glass tubes and placed obliquely to prepare slants. After that, the slants were streaked with different bacterial isolates and kept for 48 h in the incubator. The change in color from dark-green to blue was noticed after 48 h (Claus, 1989; Jawetz, 1989).
2.6.2. Catalase testA small inoculum of bacterial isolates was transferred to a clean dry glass slide and few drops of hydrogen peroxide solution (3%) were added. Rapid evolvement of oxygen bubbles gave the positive result for catalase (Facklam & Elliott, 1995).
2.6.3. Oxidase testOne to two drops of Kovács oxidase reagent (1% tetra-methyl-p-phenylenediamine dihydrochloride, in water) (Sigma-Aldrich, USA) were placed on freshly grown bacteria on Nutrient agar media (Kovacs, 1956). Color change to dark purple gave positive test for oxidase.
2.6.4. Indole testAbout 100 mL of tryptone broth was prepared and autoclaved. 7 mL of the medium was added in 20 test tubes. The bacterial colonies were inoculated in the test tubes exclusive of blank. These test tubes were kept at room temperature for 48 h. After 48 h 10 drops of Kovac’s reagent was added and kept for the color change (Isenberg & Sundheim, 1958).
2.6.5. Urease testThe bacterial isolates were inoculated in the urea agar media containing pH indicator phenol red and kept under 37°C under incubator for 24–48 h. Color change to deep pink color gave the positive reac-tion for the presence of urease (Seeliger, 1955).
2.7. Molecular characterizations
2.7.1. Isolation of genomic DNADNA was extracted from the cultures growing on nutrient agar media from 24 h, 37°C using Himedia Bacterial DNA purification kit (MB505). 16S ribosomal gene was amplified using 16S rDNA bacteria specific primers 27F forward (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-TACGGTTACCTTGTTACGACTT-3′). The PCR amplification of 16S rDNA was performed in 25 μL ali-quots containing 20–50 μg DNA template, 0.4 μmol/L of each primers, 1.5 μmol MgCl2, 200 μmol dNTPs and 1U/μL Taq Polymerase. The PCR products were analyzed on a 1.5% agarose gel stained with ethidium bromide in 1X TBE buffer and visualized under gel documentation system.
2.7.2. Cloning and sequencingThe amplified 16S ribosomal RNA was purified from agarose (1.2% w/v) using QIA quick Gel extrac-tion Kit according to manufacturer’s instruction. The purified PCR products were cloned in pGEM-T vector and transformed into competent E. coli DH5α cells using standard protocols. The plasmid

Page 5 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
insert were amplified using vector specific primer SP6 (5′-GATTTAGGTGACACTATA-3′) and T7 (5′-TAATACGACTCACTATAG-3′′). PCR was performed in 25 μl aliquots containing 20–50 μg DNA tem-plates, 0.4 μM of each primers, 1.5 μM MgCl2, 200 μM dNTPs and 1U/μl Taq Polymerase. The PCR conditions comprised an initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 30 s and extension at 72°C for 50 s with final extension at 72°C for 20 min. The PCR products were analyzed on a 1.5% agarose gel stained with ethidium bromide in 1X TBE buffer and visualized under gel documentation system. The bands were cut and eluted using Qiagen quick Gel Extraction kit and eluted amplified products sent directly for sequencing (Sci Genome Cochin, Kerala, India).
2.7.3. ARDRAARDRA was performed prior to sequencing in order to determine number of genotypes and genetic diversity among bacterial strains. Generated amplified fragments of 16S ribosomal RNA were di-gested with restriction endonuclease AluI, HaeIII (Arya, Joshi, Gupta, Kumar, & Raturi, 2015) and AfaI/MspI (Zeng, Liu, Li, Yu, & Chen, 2007) in accordance with manufacturer’s instruction. The restric-tion products were resolved in 2% agarose (w/v solution of agarose in TBE buffer containing 1 μg/ mL of ethidium bromide). Gels were visualized using Bio Rad Gel Documentation system. All reactions were repeated at least three times for each restriction enzyme to ascertain the reproducibility of banding pattern.
2.7.4. Phylogenetic analysis of ARDRA fingerprintsThe presence and absence of distinct and reproducible bands in each of the individual DNA finger-printing pattern generated by AluI, HaeIII, and AfaI/MspI profiles were converted into binary data, and the pooled binary data were used to construct composite dendrogram respectively. The Biodiversity Pro software (Version 2) was used to perform the phylogenetic analysis using Bray-Curtis Cluster analysis.
2.7.5. Principal component analysisDNA fingerprints obtained using all the 4 restriction endonucleases viz. AluI, HaeIII, and AfaI/MspI were utilized for the PCA. The software Sigma plot 11 was used to generate the graphical represen-tation of the values generated by Biodiversity Pro software version 2.0.
2.7.6. Sequence analysis and construction of phylogenetic treeSequences of all the 20 isolates were validated by comparing them with sequences available in a public database (Ribosomal Database Project release 9.43 [RDP]) (Cole et al., 2005). The 16S rDNA sequences obtained were then subjected to nucleotide basic algorithm search tool (Blastn) (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and aligned with the most similar bacterial species and then submit-ted to the NCBI database using the submission tool Sequin. Sequences exhibiting close relatedness in a BLAST search were used for construction of phylogenetic tree by neighbor joining algorithm method with the aim to study their evolutionary history (Saitou & Nei, 1987). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) showed above the branches (Felsenstein, 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Jukes-Cantor method (Jukes & Cantor, 1969) and are in the units of the number of base substitutions per site. All positions containing gaps and missing data were eliminat-ed. There were a total of 120 positions in the final data-set. Evolutionary analyses were conducted by using MEGA 6 software (Tamura, Stecher, Peterson, Filipski, & Kumar, 2013).
3. Results and discussionIn the present study, attempts have been made to isolate different metal tolerant as well as antibi-otic resistant bacterial strains from the banks of Ghats of river Ganga and characterize them using various biochemical and modern molecular techniques. In order to analyze water quality, physico-chemical characteristics of Ganga water near Ghats was performed and found to be higher than maximum permissible limit set by Indian Standard Specification for Drinking Water (IS: 10500)

Page 6 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
throughout the year (Table 1). Out of five selected Ghats, Assi ghat and Dashashwamedh ghat were found to be most polluted Ghats. Water quality of the Assi ghat has drastically deteriorated in the recent past as Assi Nala (Sewage) is directly connected to the Assi Ghat which is the mixing point of untreated waste water, domestic water and industrial pollutants with the river water. But in Dashashwamedh ghat, extreme level of human interference is possibly the major reason of increased water pollution. Level of nutrients viz. total nitrogen and total phosphorus were found to be higher in the Harishchandra ghat which is mainly due to the combustion of human bodies on this particular ghat. Other factors like total alkalinity and total dissolved solids were also high in the same ghat be-cause washing of clothes is also very frequent. The level of heavy metals was found much beyond the safe limit at all the Ghats of river Ganga. According to Indian Standard Specifications for Drinking Water IS: 10500 safe limit (Indian Standard Specifications For Drinking Water IS: 10500, 2012), Cd concentrations beyond 0.01 mg/L becomes toxic, Cr above limit of 0.05 becomes carcinogenic and Pb beyond 0.1 could cause blood cancer in human. Atmospheric deposition of metals emitted from ve-hicles, direct anthropogenic activities and discharge of untreated industrial effluents as well as do-mestic sewages into the river could be the possible reason of metal load in river Ganga (Pandey et al., 2010). The higher level of Cu in water might be due to corrosion of pipes, fittings, and utensils.
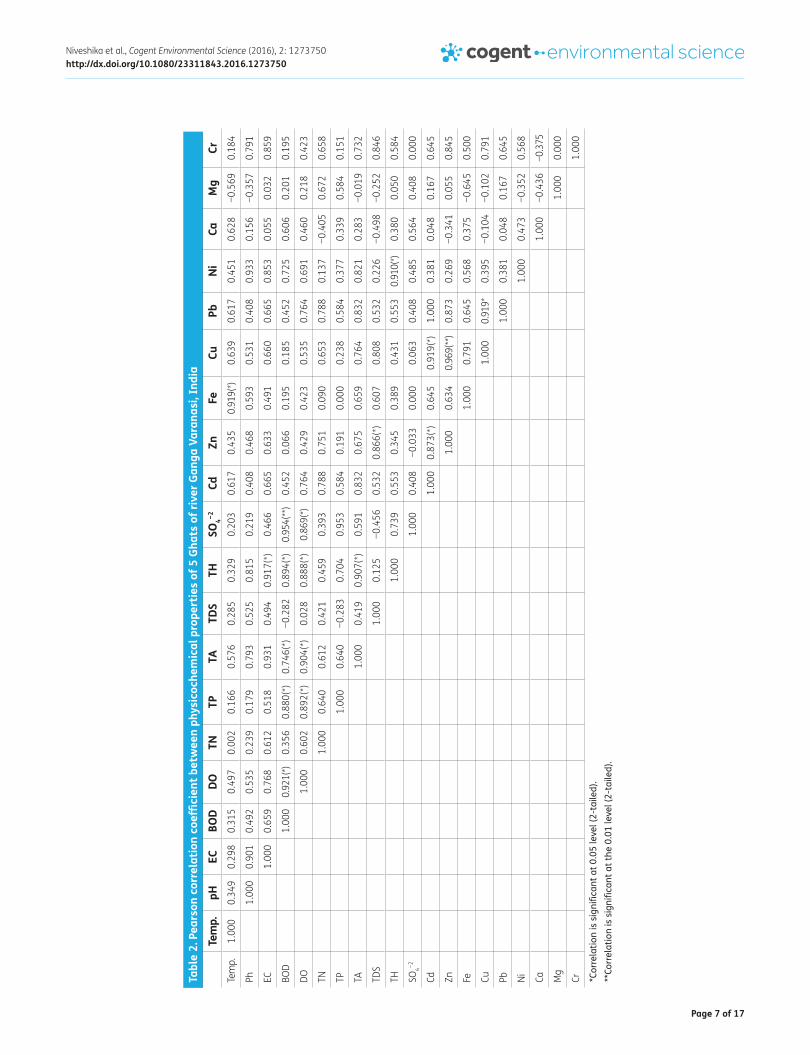
A marked correlation between physico-chemical properties of water collected from different Ghats was observed after performing the statistical analysis using Pearson’s formula of correlation and regression (Table 2). Metals like Zn, Fe, and Ni showed significant correlation with different abi-otic factors. At higher concentrations, metals exert toxic effects by forming complexes with organic compounds and toxic effects of metals can change by changing metals structure (Das et al., 2013).
Table 1. Physicochemical characteristics of 5 Ghats of river Ganga Varanasi
Notes: ± Represents mean standard deviation with three replicates; note that the values are minimum and maximum; Site 1-Dashashwamedh Ghat, Site 2-Harishchandra Ghat, Site 3-Samne Ghat, Site 4-Rajendra Prasad Ghat, Site5-Assi Ghat; EC-Electrical conductivity, BOD-Biological oxygen demand, DO-Dissolved oxygen, TN-Total nitrogen, TP-Total phosphorus, TA-Total alkalinity, TDS-Total dissolved solid, TH-Total hardness.
Parameters Site 1 Site 2 Site 3 Site 4 Site 5pH 7.6 ± 1 7.6 ± 0.5 7.7 ± 0.5 7.4 ± 0.5 7.7 ± 1
Temperature (°C) 27.9 ± 15 27.8 ± 16 27.8 ± 15 27.6 ± 16 27.9 ± 15
EC (μmho/cm) 297 ± 1 296 ± 5 289 ± 3 290 ± 3 299 ± 1
BOD 11.7 ± 2 9.8 ± 1 9.9 ± 1 10.7 ± 1 12.3 ± 2
DO 5.9 ± 0.1 5.6 ± 0.01 5.2 ± 0.1 5.8 ± 0.1 6.1 ± 0.1
TN (mg/L) 3.28 ± 0.1 4.21 ± 0.1 4.34 ± 0.1 3.19 ± 0.1 3.39 ± 0.1
TP (mg/L) 0.15 ± 0.1 0.14 ± 0.01 0.16 ± 0.1 0.14 ± 0.01 0.17 ± 0.1
TA (mg/L) 236 ± 5 223 ± 4 221 ± 5 229 ± 3 245 ± 2
TDS (mg/L) 456 ± 5 439 ± 5 459 ± 5 439 ± 6 457 ± 4
TH (as CaCO3) (mg/L) 235 ± 1 230 ± 1 239 ± 2 229 ± 1 238 ± 2
Sulfate (mg/L) 9.1 ± 0.1 7.6 ± 0.1 6.1 ± 0.1 6.7 ± 0.1 9.8 ± 0.1
Magnesium (mg/L) 1.25 ± 0.1 1.11 ± 0.01 1.18 ± 0.01 1.28 ± 0.1 1.38 ± 0.1
Calcium (mg/L) 37.2 ± 0.1 29.6 ± 0.1 28.1 ± 0.1 33.5 ± 0.1 38.5 ± 0.1
Cadmium (mg/L) 0.08 ± 0.01 0.02 ± 0.01 0.02 ± 0.01 0.05 ± 0.01 0.15 ± 0.01
Zinc (mg/L) 0.25 ± 0.01 0.21 ± 0.02 0.20 ± 0.1 0.25 ± 0.1 0.27 ± 0.1
Iron (mg/L) 0.59 ± 0.3 0.64 ± 0.05 0.68 ± 0.4 0.59 ± 0.3 0.61 ± 0.2
Copper (mg/L) 0.05±0.03 0.02 ± 0.02 0.03 ± 0.02 0.03 ± 0.02 0.08 ± 0.01
Lead (mg/L) 0.09 ± 0.01 0.06 ± 0.01 0.17 ± 0.01 0.19 ± 0.01 0.09 ± 0.02
Nickel (mg/L) 0.31 ± 0.1 0.27 ± 0.1 0.22 ± 0.1 0.31 ± 0.5 0.35 ± 0.5
Chromium (mg/L) 0.06 ± 0.1 0.03 ± 0.01 0.08 ± 0.02 0.05 ± 0.02 0.09 ± 0.01
Page 7 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
Tabl
e 2.
Pea
rson
cor
rela
tion
coeffi
cien
t bet
wee
n ph
ysic
oche
mic
al p
rope
rtie
s of
5 G
hats
of r
iver
Gan
ga V
aran
asi,
Indi
a
*Cor
rela
tion
is s
igni
fican
t at 0
.05
leve
l (2-
taile
d).
**Co
rrel
atio
n is
sig
nific
ant a
t the
0.0
1 le
vel (
2-ta
iled)
.
Tem
p.pH
ECBO
DDO
TNTP
TATD
STH
SO4−2
CdZn
FeCu
PbNi
CaM
gCr
Tem
p.1.
000
0.34
90.
298
0.31
50.
497
0.00
20.
166
0.57
60.
285
0.32
90.
203
0.61
70.
435
0.91
9(*)
0.63
90.
617
0.45
10.
628
−0.5
690.
184
Ph1.
000
0.90
10.
492
0.53
50.
239
0.17
90.
793
0.52
50.
815
0.21
90.
408
0.46
80.
593
0.53
10.
408
0.93
30.
156
−0.3
570.
791
EC1.
000
0.65
90.
768
0.61
20.
518
0.93
10.
494
0.91
7(*)
0.46
60.
665
0.63
30.
491
0.66
00.
665
0.85
30.
055
0.03
20.
859
BOD
1.00
00.
921(
*)0.
356
0.88
0(*)
0.74
6(*)
−0.2
820.
894(
*)0.
954(
**)
0.45
20.
066
0.19
50.
185
0.45
20.
725
0.60
60.
201
0.19
5
DO1.
000
0.60
20.
892(
*)0.
904(
*)0.
028
0.88
8(*)
0.86
9(*)
0.76
40.
429
0.42
30.
535
0.76
40.
691
0.46
00.
218
0.42
3
TN1.
000
0.64
00.
612
0.42
10.
459
0.39
30.
788
0.75
10.
090
0.65
30.
788
0.13
7−0
.405
0.67
20.
658
TP1.
000
0.64
0−0
.283
0.70
40.
953
0.58
40.
191
0.00
00.
238
0.58
40.
377
0.33
90.
584
0.15
1
TA1.
000
0.41
90.
907(
*)0.
591
0.83
20.
675
0.65
90.
764
0.83
20.
821
0.28
3−0
.019
0.73
2
TDS
1.00
00.
125
−0.4
560.
532
0.86
6(*)
0.60
70.
808
0.53
20.
226
−0.4
98−0
.252
0.84
6
TH1.
000
0.73
90.
553
0.34
50.
389
0.43
10.
553
0.91
0(*)
0.38
00.
050
0.58
4
SO4−2
1.00
00.
408
−0.0
330.
000
0.06
30.
408
0.48
50.
564
0.40
80.
000
Cd1.
000
0.87
3(*)
0.64
50.
919(
*)1.
000
0.38
10.
048
0.16
70.
645
Zn1.
000
0.63
40.
969(
**)
0.87
30.
269
−0.3
410.
055
0.84
5
Fe1.
000
0.79
10.
645
0.56
80.
375
−0.6
450.
500
Cu1.
000
0.91
9*0.
395
−0.1
04−0
.102
0.79
1
Pb1.
000
0.38
10.
048
0.16
70.
645
Ni1.
000
0.47
3−0
.352
0.56
8
Ca1.
000
−0.4
36−0
.375
Mg
1.00
00.
000
Cr1.
000

Page 8 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
A total of 115 culturable bacteria were isolated on the basis of their morphological and biochemi-cal properties. Of these, 20 were selected for further studies due to their ability to tolerate toxicity of six different metals (Zn, Cd, Cu, Ni, Pb, and Cr) and their resistance against 10 different antibiotics. Bacterial isolates viz. Enterobacter sp., Serratia sp., Pseudomonas fluorescens and Pseudomonas pseudoalcaligenes were able to grow at very high concentrations of various heavy metals such as Cu, Ni, Pb, and Cr. The minimum growth inhibitory concentrations of these metals were in between 300 and 400 mg/L (Table 3). Comamonas has recovered mainly from Samne ghat and Rajendra Prasad ghat where the concentration of Pb was considerably high. Industrial effluents are thought to be the major source of high Pb in these sites. The abundance of Comamonas in these extremely Pb polluted sites might be due to their genomic adaptation and subsequent increased tolerance to Pb. Occurrence and activity of Comamonas strains have already been reported in Pb contaminated soils (Konopka et al., 1999). Stoppel and Schlegel, (1995) also reported metal tolerant strain of Comamonas sp. Although many Comamonas strains has been isolated from clinical samples and regarded as op-portunistic pathogens (Willems et al., 1991). The persistent presence of Serratia, Pseudomonas pseu-doalcaligenes, Enterobacter sp., Proteus vulgaris and Shewanella sp. in the water sample collected from the Assi ghat and Dashashwamedh ghat was observed throughout year. Along with high MIC towards heavy metals, these isolates also showed MDR against Amp, Chl, Nal, Tet, Van, Imi, Cip, Ery, Pol, and Str (Table 4). Zone of inhibition of different antibiotics (Amp, Chl, Nal, Tet, Van, Imi, Cip, Ery, Pol, and Str) against sensitive bacterial strains were found in the range of 5.02 to 11.23 mm. Except Serratia, Pseudomonas pseudoalcaligenes, Comamonas, and Proteus vulgaris, all the other strains were sensitive towards imipenem, a β-lactam carbapenem (Yong et al., 2002). Maximum zone of inhibition produced by imipenem in Aeromonas hydrophila, and Ochrobactrum intermedium viz.11.03 and 11.23 mm, respectively. Therefore, the ability of these bacterial strains to overcome the toxic effects of different drugs poses a serious problem because these drugs are frequently used against bacterial infection. Imipenem is a high end broad spectrum drug mainly used to combat nosocomial
Table 3. Antibiotics resistance patterns of 20 bacterial isolates
Notes: “S”-Sensitive “R”-Resistant; Amp-Ampicillin, Chl-Chloramphenicol, Nal-Nalidixic acid, Tet-Tetracyclin, Van-Vancomycin, Imi-Imipenem, Cip-Ciprofloxacin, Ery-Erythromycin, Pol-Polymixin, Str-Streptomycin.
Bacterial isolates Conc. (mg/L) Amp Chl Nal Tet Van Imi Cip Ery Pol StrProvidencia 30 S S R S R R R S S S
Shewanella sp. 30 R S R R R R S R S R
Shewanella sp. 30 S R S S S R R S S S
Shewanella decolorationis 30 S R R S S R S S S R
Brucella sp. 30 R S R R R R S S S S
Enterobacter sp. 30 R S R S R R R R S R
Serratia marcescens sp. 30 R R R R R R R R R R
Kurthia sp. 30 S S R S S S S S S S
Kurthia gibsonii 30 R R R S S S S S S S
Rummellibacillus sp. 30 R S R S S S R R S S
Comamonas sp. 30 R R R S S R S S S S
Comamonas terrigena 30 R R R R S R S S S R
Proteus vulgaris 30 R R R R R S R R R R
Exiguobacterium 30 S R S S S S S R S S
Aeromonas sp. 30 S S S R S S S R S S
Strenotrophomonas koreensis 30 R S R S S S S S S S
Aeromonas hydrophila 30 R S R R S S R S S S
Pseudomonas fluorescens 30 R R R R R R R R R R
Pseudomonas pseudoalcaligenes 30 R R R R R R R R R R
Ochrobactrum intermedium 30 S S S S R R R R S S

Page 9 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
infections. In the Assi ghat, city municipal waste and untreated sewage are the pervasive source of water pollution and also a suitable environment for the growth of human pathogenic bacteria. These bacteria are known for colonizing the respiratory and urinary tract and have also the ability to cause meningitis, arthritis and wound infection. In Dashashwamedh ghat, uncontrolled human interfer-ences and other various activities like boat landing have created water logging condition where harmful bacteria grow in copious amount. Aeromonas, Kurthia, Ochrobactrum, and Brucella were the usual bacterial strains in all the sites throughout year. Morphological and biochemical characteris-tics of bacterial strains are mentioned in Table 5.
The advent of tools in molecular biology has revolutionized the process of strain identification and saved money, time and energy too. In this study ARDRA was used as a model to examine the capac-ity of restriction-based techniques for bacterial strain identification, and the possibility of generating phylogenetic information from ARDRA-based dendrograms. Approx. 1.3 kb 16S ribosomal RNA am-plified product was observed. ARDRA concerning the 16S ribosomal RNA amplified product of bacte-rial isolates digested by AluI, HaeIII and AfaI/MspI were reproducible yielded 2–5, 3–7, and 2–4 bands respectively. Band pattern of each isolates represented particular ARDRA genotypes. AluI showed similar banding pattern for Aeromonas hydrophila and Aeromonas sp., Shewanella decolora-tionis, and Shewanella, Comamonas terrigena and Comamonas sp., Pseudomonas fluorescens, and Pseudomonas pseudoalcaligenes as shown in Figure 1a. Also double restriction endonuclease diges-tion AfaI/MspI was failed to differentiate most of the bacterial isolates and yielded lowest number of genotypes showing in Figure 2a. However, HaeIII discriminated all the strains on the basis of band pattern and generated species specific patterns as shown in Figure 3a which was further confirmed by sequencing data that all isolates belong to different species. Therefore, it was found that out of all the restriction endonuclease, HaeIII could able to detect interspecies and interstrain variability of cultivable fresh water bacterial strains Restriction endonuclease viz. HaeIII has also been used for bacterial identification and diversity assessment by many workers (Heyndrickx, Vauterin, Vandamme, Kersters, & De Vos, 1996; Sun et al., 2008)
Table 4. Minimum inhibitory concentration (MIC) of heavy metals (mg/L) against bacterial isolatesBacterial isolates Zn (mg/L) Cd (mg/L) Cu (mg/L) Ni (mg/L) Pb (mg/L) Cr (mg/L)Providencia sp. 150 150 200 200 150 150
Shewanella sp. 150 150 150 150 150 150
Shewanella sp. 150 150 150 150 200 150
Shewanella decolorationis 200 200 300 200 200 200
Brucella sp. 200 150 150 200 200 300
Enterobacter sp. 200 200 300 350 350 350
Serratia sp. 200 200 350 400 350 300
Kurthia sp. 200 200 200 200 150 150
Kurthia gibsonii 200 200 200 300 200 150
Rummellibacillus 150 100 150 200 150 150
Comamonas sp. 150 150 150 200 400 150
Comamonas terrigena 150 150 200 200 400 150
Proteus vulgaris 150 150 200 200 200 150
Exiguobacterium sp. 200 150 150 200 200 150
Aeromonas sp. 150 100 150 150 150 150
Strenotrophomonas koreensis 150 100 150 150 150 150
Aeromonas hydrophila 150 100 150 150 150 150
Pseudomonas fluorescens 200 200 300 350 300 250
Pseudomonas pseudoalcaligenes 200 200 200 350 300 300
Ochrobactrum intermedium 100 100 150 100 100 150

Page 10 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
Tabl
e 5.
Mor
phol
ogic
al a
nd b
ioch
emic
al c
hara
cter
istic
s of
bac
teria
l iso
late
sM
orph
olog
ical
Bioc
hem
ical
Bact
eria
l iso
late
sCo
lony
Col
orCo
lony
text
ure
Shap
eGr
am n
atur
eCa
tala
seOx
idas
eCi
trat
eIn
dole
Urea
sePr
ovid
enci
a sp
.Du
ll gr
ayO
paqu
eRo
d sh
ape
G−+
−+
++
Shew
anel
la s
p.Br
owni
shRo
und
Rod
shap
eG−
++
−−
−
Shew
anel
la s
p.Pa
le w
hite
Roun
dRo
d sh
ape
G−+
−+
−−
Shew
anel
la d
ecol
orat
ioni
sPa
le w
hite
Roun
dRo
d sh
ape
G−+
++
+−
Bruc
ella
sp.
Ligh
t gra
yPu
ncta
teCo
ccob
acill
iG−
++
−−
+
Ente
roba
cter
sp.
Shin
y ta
nRo
und
Rod
shap
eG−
−−
+−
−
Serra
tia m
arce
scen
sSh
iny
red
Roun
dRo
d sh
ape
G−+
−−
−+
Kurt
hia
sp.
Crea
mRo
und
Rod
shap
eG+
++
+−−
−
Kurt
hia
gibs
onii
Crea
mRo
und
Rod
shap
eG+
++
+−
−
Rum
mel
libac
illlu
s sp
.Cr
eam
Roun
dRo
d sh
ape
G++
++
−−
Com
amon
as s
p.Cr
eam
Roun
dRo
d sh
ape
G−+
++
−−
Com
amon
as te
rrige
naCr
eam
Roun
dRo
d sh
ape
G−+
++
−−
Prot
eus v
ulga
risGr
ayRo
und
Rod
shap
eG−
+−
−+
−
Exig
uoba
cter
ium
sp.
Pale
yel
low
Roun
dRo
d sh
ape
G++
−+
−−
Aero
mon
as s
p.W
hite
Roun
dRo
d sh
ape
G−+
++
+−
Stre
notro
phom
onas
kor
eens
isCr
eam
Roun
dRo
d sh
ape
G−+
+−
−−
Aero
mon
as h
ydro
phila
Crea
mRo
und
Rod
shap
eG−
++
++
−
Pseu
dom
onas
fluo
resc
ens
Gray
Roun
dRo
d sh
ape
G−+
++
−+
Pseu
dom
onas
pse
udoa
lcal
igen
esGr
ayRo
und
Rod
shap
eG−
++
+−
−
Ochr
obac
trum
inte
rmed
ium
Pale
Roun
dRo
d sh
ape
G−+
+−
−+
Page 11 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
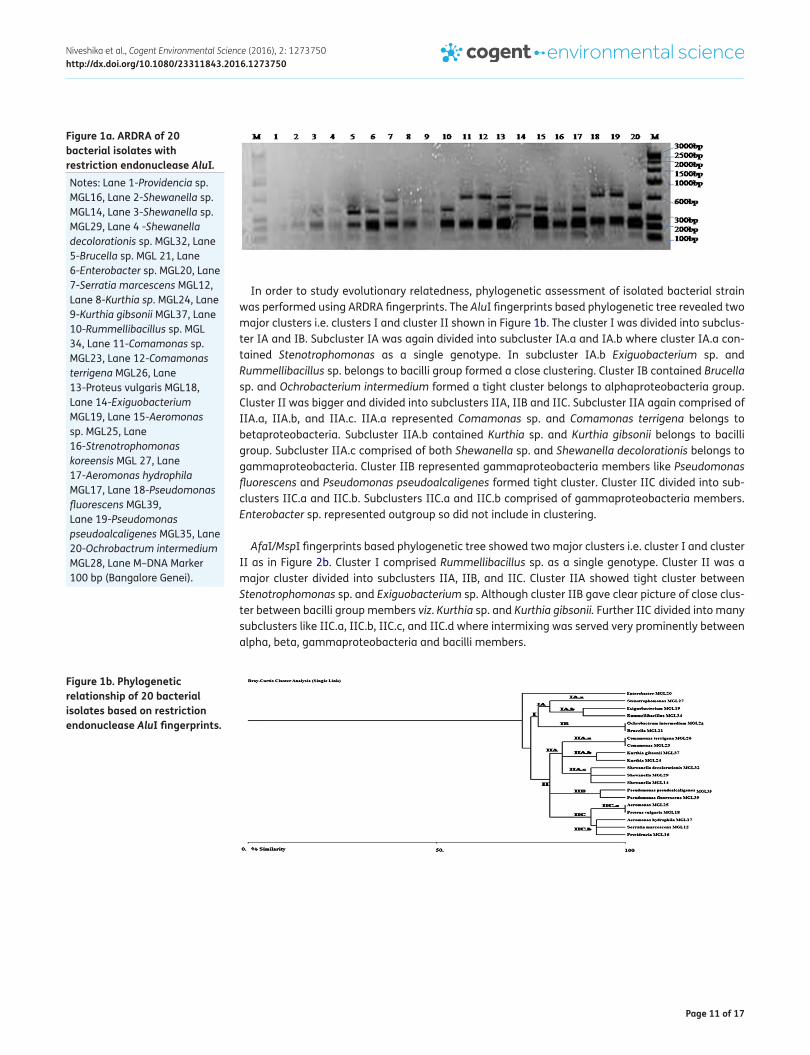
In order to study evolutionary relatedness, phylogenetic assessment of isolated bacterial strain was performed using ARDRA fingerprints. The AluI fingerprints based phylogenetic tree revealed two major clusters i.e. clusters I and cluster II shown in Figure 1b. The cluster I was divided into subclus-ter IA and IB. Subcluster IA was again divided into subcluster IA.a and IA.b where cluster IA.a con-tained Stenotrophomonas as a single genotype. In subcluster IA.b Exiguobacterium sp. and Rummellibacillus sp. belongs to bacilli group formed a close clustering. Cluster IB contained Brucella sp. and Ochrobacterium intermedium formed a tight cluster belongs to alphaproteobacteria group. Cluster II was bigger and divided into subclusters IIA, IIB and IIC. Subcluster IIA again comprised of IIA.a, IIA.b, and IIA.c. IIA.a represented Comamonas sp. and Comamonas terrigena belongs to betaproteobacteria. Subcluster IIA.b contained Kurthia sp. and Kurthia gibsonii belongs to bacilli group. Subcluster IIA.c comprised of both Shewanella sp. and Shewanella decolorationis belongs to gammaproteobacteria. Cluster IIB represented gammaproteobacteria members like Pseudomonas fluorescens and Pseudomonas pseudoalcaligenes formed tight cluster. Cluster IIC divided into sub-clusters IIC.a and IIC.b. Subclusters IIC.a and IIC.b comprised of gammaproteobacteria members. Enterobacter sp. represented outgroup so did not include in clustering.
AfaI/MspI fingerprints based phylogenetic tree showed two major clusters i.e. cluster I and cluster II as in Figure 2b. Cluster I comprised Rummellibacillus sp. as a single genotype. Cluster II was a major cluster divided into subclusters IIA, IIB, and IIC. Cluster IIA showed tight cluster between Stenotrophomonas sp. and Exiguobacterium sp. Although cluster IIB gave clear picture of close clus-ter between bacilli group members viz. Kurthia sp. and Kurthia gibsonii. Further IIC divided into many subclusters like IIC.a, IIC.b, IIC.c, and IIC.d where intermixing was served very prominently between alpha, beta, gammaproteobacteria and bacilli members.
Figure 1b. Phylogenetic relationship of 20 bacterial isolates based on restriction endonuclease AluI fingerprints.
Figure 1a. ARDRA of 20 bacterial isolates with restriction endonuclease AluI. Notes: Lane 1-Providencia sp. MGL16, Lane 2-Shewanella sp. MGL14, Lane 3-Shewanella sp. MGL29, Lane 4 -Shewanella decolorationis sp. MGL32, Lane 5-Brucella sp. MGL 21, Lane 6-Enterobacter sp. MGL20, Lane 7-Serratia marcescens MGL12, Lane 8-Kurthia sp. MGL24, Lane 9-Kurthia gibsonii MGL37, Lane 10-Rummellibacillus sp. MGL 34, Lane 11-Comamonas sp. MGL23, Lane 12-Comamonas terrigena MGL26, Lane 13-Proteus vulgaris MGL18, Lane 14-Exiguobacterium MGL19, Lane 15-Aeromonas sp. MGL25, Lane 16-Strenotrophomonas koreensis MGL 27, Lane 17-Aeromonas hydrophila MGL17, Lane 18-Pseudomonas fluorescens MGL39, Lane 19-Pseudomonas pseudoalcaligenes MGL35, Lane 20-Ochrobactrum intermedium MGL28, Lane M–DNA Marker 100 bp (Bangalore Genei).
Page 12 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
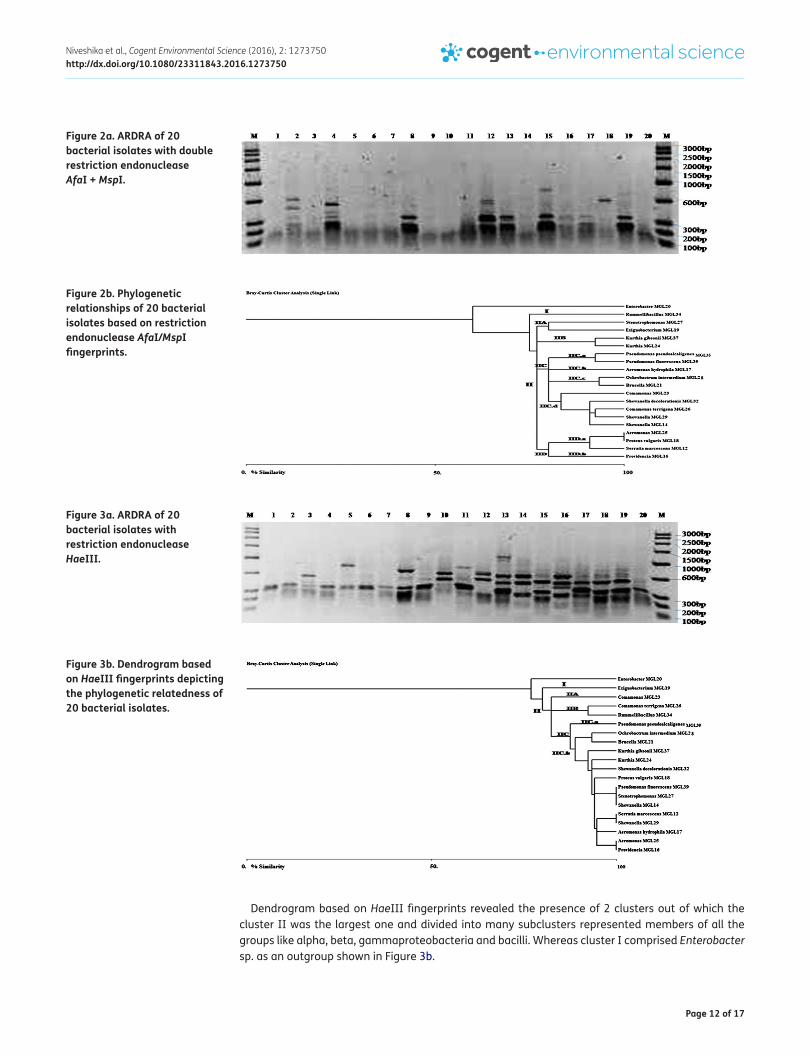
Dendrogram based on HaeIII fingerprints revealed the presence of 2 clusters out of which the cluster II was the largest one and divided into many subclusters represented members of all the groups like alpha, beta, gammaproteobacteria and bacilli. Whereas cluster I comprised Enterobacter sp. as an outgroup shown in Figure 3b.
Figure 2b. Phylogenetic relationships of 20 bacterial isolates based on restriction endonuclease AfaI/MspI fingerprints.
Figure 3a. ARDRA of 20 bacterial isolates with restriction endonuclease HaeIII.
Figure 3b. Dendrogram based on HaeIII fingerprints depicting the phylogenetic relatedness of 20 bacterial isolates.
Figure 2a. ARDRA of 20 bacterial isolates with double restriction endonuclease AfaI + MspI.

Page 13 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
The dendrogram generated after combining all DNA fingerprints obtained from ARDRA using AluI, HaeIII and AfaI/MspI showed two clusters i.e. cluster I and cluster II as depicted in Figure 4a. Cluster I revealed members of alphaproteobacteria. Cluster II was a major cluster divided into cluster IIA, IIB, IIC and IID. Cluster II A further subdivided into subcluster IIA.a, IIA.b and IIA.c. Members of betaproteobacteria showed a tight clustering in cluster II A.a. Cluster IIA.b comprised members of bacilli group viz. Kurthia sp. and Kurthia gibsonii whereas in cluster IIA.c included members of gam-maproteobacteria. Cluster IIB comprised Pseudomonas fluorescens and Pseudomonas pseudoalcali-genes. Cluster IIC represented Stenotrophomonas sp. as a single genotype. Cluster IID subdivided into subcluster IID.a, IID.b and IID.c. Subcluster IID.a included bacillli group members whereas sub-cluster IID.b and IID.c comprised members of gammaproteobacteria.
Based on the result of ARDRA fingerprinting and phylogenetic characterization of bacterial iso-lates, we could say that ARDRA is a robust means which can generate species specific pattern for rapid assessment of microbial diversity. Representative strain of each ARDRA fingerprints could be chosen for 16S ribosomal RNA sequencing for obtaining information about what type of microorgan-ism present in the sample. The reliability of ARDRA technique in congruence with the phylogenetic analysis for microbial diversity study was also proved by many workers (Arzu, Nilgun, Birgul, & Cumhur, 2012; Baik et al., 2008). Vaneechoutte and Heyndrickx (2001) mentioned ARDRA technique as a new approach for bacterial identification, phylogeny and taxonomy study. Moyer, Tiedje, Dobbs, and Karl (1996) compared phylogenetic trees based on ARDRA profiles and 16S rDNA sequences and therefore concluded that using different types of restriction endonucleases could be able to yield 76–100% success in obtaining phylogenetic affiliations accurately. Although, more studies on effi-ciency of ARDRA with other restriction endonucleases required with respect to fresh water bacterial community.
Figure 4b. Principal component analysis based on ARDRA fingerprinting generated by 4 restriction endonucleases viz. AluI, HaeIII and AfaI/MspI profile of 20 isolated bacterial strains.
Figure 4a. Dendrogram based on ARDRA fingerprinting generated by 4 restriction endonucleases viz. AluI, HaeIII and AfaI/MspI profile of 20 isolated bacterial strains.

Page 14 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
Furthermore, the fingerprints obtained using all the 4 restriction endonucleases viz. AluI, HaeIII and AfaI/MspI were utilized for the PCAPCA in order to check whether phylogenetic tree were in co-herence with PCA or not. PCA revealed the presence of seven major clusters and two minor clusters showing in Figure 4b. In the major clusters, Aeromonas sp., Aeromonas hydrophila, Serratia marces-cens, Providencia sp. and Proteus vulgaris represented members of group gammaproteobacteria set-tled into one cluster. Members of betaproteobacteria viz. Comamonas sp. and Comamonas terrigena occupied single cluster, whereas members of bacilli groups were represented into two separate clus-ters. Members of gammaproteobacteria viz. Pseudomonas fluorescens, pseudomonas pseudoalcali-genes, Shewanella sp. and Shewanella decolorationis were occupied two major clusters. In the major clusters, members of alphaproteobacteria viz. Brucella sp. and Ochrobactrum intermedium com-prised in separate cluster. In minor clusters Enterobacter sp. as well as Stenotrophomonas sp. repre-sented outgroup. Thus the phylogenetic tree and the PCA gave a clear picture of evolutionary lineage of bacterial isolates.
After comparing ARDRA fingerprints of all the isolated strains, 16S ribosomal RNA amplified prod-ucts were cloned using competent cell of E.coli DH5α. Blue and white colonies were appeared on the petriplate. White colonies represented the recombinants. The cloned 16S ribosomal RNA were se-quenced and then sequences were submitted to the NCBI databank where accession numbers were assigned (KP994633, KP994626, KR063129, KR063128, KR063114, KP994630, KP994627, KR063119, KR063118, KR063127, KR063115, KR063116, KP994628, KR063129, KR063111, KR063130, KR063124, KR063123, KR063125, KR063121).
The phylogenetic tree was constructed by using neighbor-joining method (Figure 5). Bootstrap values (%) were based on 1,000 replicates and more than 50% bootstrap values were shown at the branch point. The sequenced 20 bacterial strains grouped into four major classes; alpha, beta, gam-maproteobacteria and bacilli. The dominant group was gammaproteobacteria having 60% bacterial isolates, only 10% isolates belongs to alphaproteobacteria, 10% were betaproteobacteria, and 20% were bacilli.
Therefore, it is worth mentioning that combination of physiological, biochemical and molecular methods gives clear cut picture about the existence of multiple metal tolerant and antibiotics resist-ant bacterial community in fresh water system.
Figure 5. Phylogenetic tree showing the relationship on the basis of 16S rDNA gene sequences of 20 bacterial isolates belonging to different genera and the related members from these genera. The tree was constructed by using the neighbor-joining method. Bootstrap values (%) are based on 1,000 replicates and shown at the branch point with more than 50% bootstrap values.
*Representing reference strain.

Page 15 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
4. ConclusionMixing of sewage along with industrial effluents into the Ganga river perturb water quality. Polluted water became a habitat of various pathogenic bacteria having dual properties in rendering multiple heavy metals tolerant capacity as well as antibiotics resistance. Collaboration of physiological, biochemical and molecular techniques could provide complete information about bacterial identification and characterization. Presence of harmful bacteria in the river Ganga invites serious attention from human health perspective since Ganga water is also used for drinking, holy bathing and irrigation purposes.
AcknowledgementsThe Head, Department of Botany, Banaras Hindu University, Varanasi, India is gratefully acknowledged for providing laboratory facilities. Three of us (Niveshika, Savita Singh and Ekta Verma) are thankful to the UGC, New Delhi for financial support in the form of Junior Research Fellowship.
FundingThis work was supported by University Grants Commission, India.
Author details Niveshika1
E-mail: [email protected] Singh1
E-mail: [email protected] Verma1
E-mail: [email protected] Kumar Mishra1
E-mails: [email protected], [email protected] Laboratory of Microbial Genetics, Department of Botany,
Banaras Hindu University, Varanasi 221005, India.
Citation informationCite this article as: Isolation, characterization and molecular phylogeny of multiple metal tolerant and antibiotics resistant bacterial isolates from river Ganga, Varanasi, India, Niveshika, Savita Singh, Ekta Verma & Arun Kumar Mishra, Cogent Environmental Science (2016), 2: 1273750.
ReferencesAmerican Public Health Association [APHA]. (1998). Standard
methods for the examination of water and waste water. Washington, DC: Author.
Anonymous. (2006). Jawaharlal Nehru National Urban Renewal Mission (JNNURM) (p. 69). Varanasi. Retrieved from http://jnnurm.nic.in/wpcontent/uploads/2011/01/PMSpeechOverviewE.pdf
Arya, M., Joshi, G. K., Gupta, A. K., Kumar, A., & Raturi, A. (2015). Isolation and characterization of thermophilic bacterial strains from Soldhar (Tapovan) hot spring in Central Himalayan Region, India. Annals of Microbiology, 65, 1457–1464. http://dx.doi.org/10.1007/s13213-014-0984-y
Arzu, C. C., Nilgun, T., Birgul, O., & Cumhur, C. (2012). The genetic diversity of genus Bacillus and the related genera revealed by 16s rRNA gene sequences and ARDRA analyses isolated from geothermal regions of Turkey. Brazilian Journal of Microbiology, 43, 309–324.
Baik, K. S., Park, S. C., Kim, E. M., Bae, K. S., Ann, J. H., Ka, J. O., … Seong, C. N. (2008). Diversity of bacterial community in freshwater of Woopo wetland. The Journal of Microbiology, 46, 647–655. http://dx.doi.org/10.1007/s12275-008-0135-x
Baquero, F., Martínez, J. L., & Cantón, R. (2008). Antibiotics and antibiotic resistance in water environments. Current Opinion in Biotechnology, 19, 260–265. http://dx.doi.org/10.1016/j.copbio.2008.05.006
Bauer, A. W., Kirby, W. M. M., Sherris, J. C., & Turck, M. (1966). Antibiotic susceptibility testing by a standardized single disc method. American Journal of Clinical Pathology, 45, 493–591.
Bergey, D. H., & Breed, R. S. (1994). Biochemical identification of enterobacteriaceae. Bergey’s Manual of Determinative Bacteriology, 1, 416–417.
Beveridge, T. J., & Doyle, R. (1989). Metal ions and bacteria. New York, NY: Wiley.
Bolan, N., Kunhikrishnan, A., Thangarajan, R., Kumpiene, J., Park, J., Makino, T., … Scheckel, K. (2014). Remediation of heavy metal(loid)s contaminated soils – To mobilize or to immobilize? Journal of Hazardous Materials, 141–166. http://dx.doi.org/10.1016/j.jhazmat.2013.12.018
Claus, G. W. (1989). Understanding microbes: A laboratory textbook for microbiology. New York, NY: W.H. Freeman.
Cole, J. R., Farris, R. J., Wang, Q., Kulam, S. A., McGarrell, D. M., Garrity, G. M., & Tiedje, J. M. (2005). The ribosomal database project (RDP-II): Sequences and tools for high-throughput rRNA analysis. Nucleic Acids Research, 33, 294–296.
Das, S., Patnaik, S. C., Sahu, H. K., Chakraborty, A., Sudarshan, M., & Thatoi, H. N. (2013). Heavy metal contamination, physico-chemical and microbial evaluation of water samples collected from chromite mine environment of Sukinda, India. Transactions of Nonferrous Metals Society of China, 23, 484–493. http://dx.doi.org/10.1016/S1003-6326(13)62489-9
Ehrlich, H. L., & Brierley, C. L. (1990). Microbial mineral recovery. New York, NY: McGraw-Hill.
Facklam, R., & Elliott, J. A. (1995). Identification, classification, and clinical relevance of catalase-negative, gram-positive cocci, excluding the Streptococci and Enterococci. Clinical Microbiology Reviews, 8, 479–495.
Felsenstein, J. (1985). Phylogenies and the comparative method. The American Naturalist, 125, 1–15. http://dx.doi.org/10.1086/284325
Heyndrickx, M., Vauterin, L., Vandamme, P., Kersters, K., & De Vos, P. (1996). Applicability of combined amplified ribosomal DNA restriction analysis (ARDRA) patterns in bacterial phylogeny and taxonomy. Journal of Microbiological Methods, 26, 247–259. http://dx.doi.org/10.1016/0167-7012(96)00916-5
Indian Standard Drinking Water Specification (Second Revision) IS 10500. (2012). Retrieved from http://www.cgwb.gov.in/Documents/WQ-standards.pdf
Industrial Directory. (1981). Government of India Ministry of Micro, Small & Medium Enterprises, Brief Industrial Profile of Varanasi District (updated) 1981. Retrieved from http://dcmsme.gov.in/dips/DIP,%20Varanasi.pdf
Isenberg, H. D., & Sundheim, L. H. (1958). Indole reactions in bacteria. Journal of Bacteriology Bacteriology, 75, 682–690.
Jawetz, E. (1989). Medical microbiology (18th ed.). San Mateo, CA: Appleton and Lange.
Jukes, T. H., & Cantor, C. R. (1969). Evolution of protein molecules. In H. N. Munro (Ed.), Mammalian protein metabolism (pp. 21–132). New York, NY: Academic press. http://dx.doi.org/10.1016/B978-1-4832-3211-9.50009-7

Page 16 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
Konopka, A., Zakharova, T., Bischoff, M., Oliver, L., Nakatsu, C., & Turco, R. F. (1999). Microbial biomass and activity in lead-contaminated soil. Applied and Environmental Microbiology, 65, 2256–2259.
Kovacs, N. (1956). Identification of pseudomonas pyocyanea by the oxidase reaction. Nature, 178, 703. http://dx.doi.org/10.1038/178703a0
Kumarasamy, K. K., Toleman, M. A., Walsh, T. R., Bagaria, J., Butt, F., Balakrishnan, R., Chaudhary, U., … Woodford, N. (2010). Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. The Lancet Infectious Diseases, 10, 597–602. http://dx.doi.org/10.1016/S1473-3099(10)70143-2
Levy, S. B. (1997). Antibiotic resistance: An ecological imbalance. In D. J. Chadwick & J. Goode (Eds.), Antibiotic resistance: Origins, Evolution, Selection and Spread (pp. 1–9). Chichester: Wiley and Sons.
Martínez-Murcia, A. J., Acinas, S. G., & Rodriguez-Valera, F. (1995). Evaluation of prokaryotic diversity by restrictase digestion of 16S rDNA directly amplified from hypersaline environments. FEMS Microbiology Ecology, 17, 247–255. http://dx.doi.org/10.1016/0168-6496(95)00029-A
Mishra, A., & Tripathi, B. D. (2007). Heavy metal contamination of soil and bioaccumulation in vegetables irrigated with treated waste water in the tropical city of Varanasi, India. Toxicological and Environmental Chemistry, 5, 861–871.
Moyer, C. L., Tiedje, J. M., Dobbs, F. C., & Karl, D. M. (1996). A computer- simulated restriction fragment length polymorphism analysis of bacterial small-subunit rRNA genes: Efficacy of selected tetrameric restriction enzymes for studies of microbial diversity in nature. Applied and Environmental Microbiology, 62, 2501–2507.
Mukherjee, S., Kumar, D., Kumar, A. K., & Chakraborthy, R. (2013). 16S rRNA gene sequence analysis of the metagenome derived from water of river Mahananda at Siliguri. Indian Journal of Biotechnology, 12, 80–88.
Narasimhulu, K., Rao, S., & Venu Vinod, A. (2010). Isolation and Identifi cation of Bacterial Strains and Study of their Resistance to Heavy Metals and Antibiotics. Journal of Microbial and Biochemical Technology, 02, 074–076. http://dx.doi.org/10.4172/1948-5948
Pandey, J., Shubhashish, K., & Pandey, R. (2010). Heavy metal contamination of Ganga River at Varanasi in relation to atmospheric deposition. Journal of Tropical Ecology, 51, 365–373.
Rai, P. K., & Tripathi, B. D. (2008). Heavy metals in industrial wastewater, soil and vegetables in Lohta village, India. Toxicological and Environmental Chemistry, 90, 247–257. http://dx.doi.org/10.1080/02772240701458584
Rai, P. K., Mishra, A., & Tripathi, B. D. (2010). Heavy metal and microbial pollution of the River Ganga: A case study of water quality at Varanasi. Aquatic Ecosystem and Health Management, 13, 352–361. http://dx.doi.org/10.1080/14634988.2010.528739
Saitou, N., & Nei, M. (1987). The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4, 406–425.
Sangu, R. P. S., & Sharma, S. K. (1987). An assessment of water quality of River Ganga at Garmukeshwar. Indian Journal of Ecology, 14, 278–281.
Seeliger, H. P. R. (1955). Use of a urease test for the screening and identification of cryptococci. Journal of Bacteriology, 72, 127–131.
Stoppel, R. D., & Schlegel, H. G. (1995). Nickel-resistant bacteria from anthropogenically nickel-polluted and naturally nickel-percolated ecosystems. Applied and Environmental Microbiology, 61, 2276–2285.
Sun, L., Qiu, F., Zhang, X., Dai, X., Dong, X., & Song, W. (2008). Endophytic bacterial diversity in rice (Oryza sativa L.) roots estimated by 16S rDNA sequence analysis. Microbial Ecology, 55, 415–424. http://dx.doi.org/10.1007/s00248-007-9287-1
Tamura, K., Stecher, G., Peterson, D., Filipski, A., & Kumar, S. (2013). MEGA6: Molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution, 30, 2725–2729. http://dx.doi.org/10.1093/molbev/mst197
Tenover, F. C., & McGowan, J. E. (1996). Reasons for the emergence of antibiotic resistance. The American Journal of the Medical Sciences, 311, 9–16. http://dx.doi.org/10.1016/S0002-9629(15)41625-8
Vaneechoutte, M., & Heyndrickx, M. (2001). Application and analysis of ARDRA patterns in bacterial identification, taxonomy and phylogeny. In L. Dijkshoorn, K. J. Towner, & M. Struelens (Eds.), New approaches for the generation and analysis of microbial typing data (pp. 211–247). Amsterdam: Elsevier. http://dx.doi.org/10.1016/B978-044450740-2/50010-1
Vogel, A. I. (1978). A text book of quantitative inorganic analysis including elementary instrumental analysis (4th ed., pp. 837, 328–329, 504–506). London: The English Language Book Society and Langman.
Willems, A., Pot, B., Falsen, E., Vandamme, P., Gillis, M., Kersters, K., & De Ley, J. (1991). Polyphasic taxonomic study of the emended genus comamonas: Relationship to aquaspirillum aquaticum, E. Falsen group 10, and other clinical isolates. International Journal of Systematic Bacteriology, 41, 427–444. http://dx.doi.org/10.1099/00207713-41-3-427
Woese, C. R. (1987). Bacterial evolution. Microbiological Reviews, 51, 221–271.
World Health Organization [WHO]. (2002). The world health report. Geneva: Author.
Yong, D., Lee, K., Yum, J. H., Shin, H. B., Rossolini, G. M., & Chong, Y. (2002). Imipenem-EDTA disk method for differentiation of metallo-beta-lactamase-producing clinical isolates of pseudomonas spp. and acinetobacter spp. Journal of Clinical Microbiology, 40, 3798–3801. http://dx.doi.org/10.1128/JCM.40.10.3798-3801.2002
Zeng, Y., Liu, W., Li, H., Yu, Y., & Chen, B. (2007). Effect of restriction endonucleases on assessment of biodiversity of cultivable polar marine planktonic bacteria by amplified ribosomal DNA restriction analysis. Extremophiles, 11, 685–692. http://dx.doi.org/10.1007/s00792-007-0086-x

Page 17 of 17
Niveshika et al., Cogent Environmental Science (2016), 2: 1273750http://dx.doi.org/10.1080/23311843.2016.1273750
© 2017 The Author(s). This open access article is distributed under a Creative Commons Attribution (CC-BY) 4.0 license.You are free to: Share — copy and redistribute the material in any medium or format Adapt — remix, transform, and build upon the material for any purpose, even commercially.The licensor cannot revoke these freedoms as long as you follow the license terms.
Under the following terms:Attribution — You must give appropriate credit, provide a link to the license, and indicate if changes were made. You may do so in any reasonable manner, but not in any way that suggests the licensor endorses you or your use. No additional restrictions You may not apply legal terms or technological measures that legally restrict others from doing anything the license permits.
Cogent Environmental Science (ISSN: 2331-1843) is published by Cogent OA, part of Taylor & Francis Group. Publishing with Cogent OA ensures:• Immediate, universal access to your article on publication• High visibility and discoverability via the Cogent OA website as well as Taylor & Francis Online• Download and citation statistics for your article• Rapid online publication• Input from, and dialog with, expert editors and editorial boards• Retention of full copyright of your article• Guaranteed legacy preservation of your article• Discounts and waivers for authors in developing regionsSubmit your manuscript to a Cogent OA journal at www.CogentOA.com