isolation and characterization of chromatin and dna … · isolation and characterization of...
TRANSCRIPT

J. Cell Sci. i i , 131-152 (1972) 13EPrinted in Great Britain
ISOLATION AND CHARACTERIZATION OF
CHROMATIN AND DNA FROM THE SPERM
OF THE SPIDER CRAB, LIBINIA EMARG1NATA
JACK C. VAUGHN AND GERTRUDE W. HINSCHDepartment of Zoology, Miami University, Oxford, Ohio, U.S.A., and Institute ofMolecular Evolution, University of Miami, Coral Gables, Florida, U.S.A., and TheMarine Biological Laboratory, Woods Hole, Mass., U.S.A.
SUMMARYAn isolation technique is described which yields highly purified sperm from various regions
of the reproductive tract of the crab Libinia emarginata. Sperm chromatin was isolated andanalysed with respect to ultrastructure, ultraviolet absorption spectrum, thermal melting profile,behaviour in CsCl gradients, and chemical composition.
Sperm-isolated chromatin not stabilized with uranyl acetate prior to ethanol dehydration,like prokaryotic chromatin, has aggregated fibres with a diameter in the range 30-95 nm ormore. With uranyl acetate stabilization, only very fine fibres are seen. Pronase treatment of thethick fibres leads to formation of very fine fibres, showing that protein is associated with thisisolated chromatin. Polyacrylamide electrophoretic analysis of the chromatin protein shows thatit is not basic. Standard basic protein staining of sperm taken from testes, vas deferens, andseminal receptacles after various periods of storage there gives a negative nuclear reaction in allcases. This suggests the absence of sperm nuclear histones and protamines. Sperm chromatinprotein is salt-dissociable and has a ratio of basic to acidic amino acids of 0-43-0-48. The-protein/DNA ratio is 2-3-3-2, and the ratio EJBO/EJ^, is 0-50-0-67. Thermal denaturationcurves are identical for sperm chromatin and purified sperm DNA, supporting the other datashowing lack of nuclear basic proteins. An (A + T)-rich satellite DNA component comprisesabout 7—9 % of the total sperm genome. It is suggested that the genes which specify gamete-typenuclear basic proteins are either permanently repressed or deleted in decapod Crustacea.
INTRODUCTION
It is now well established that histones complexed to DNA may prevent transcrip-tion (for reviews, see Georgiev, 1969; Phillips, 1971), and that protamines and othergamete-type basic proteins with a very high net positive charge are even bettei generepressors than are the histones (Skalka, Fowler & Hurwitz, 1966). These observationshave led to the hypothesis that the biological significance of protamines and othergamete-type basic proteins may be the complete repression of genes, and perhaps therestoration of the genome to a state devoid of the developmental history of the indi-vidual (Olins, Olins & von Hippel, 1968).
Several recent reports, utilizing a number of crab species and generally indirectapproaches, have concluded that decapod crustacean sperm chromatin is devoid ofhistones and protamines (Chevaillier, 1966, 1967, 1968; Vaughn, 1968; Vaughn et aL1969; Langreth, 1969), although none have isolated and characterized the chromatinitself. The presence of a group of relatively advanced organisms whose sperm are
9-2

132 J. C. Vaughn and G. W. Hinsch
lacking in these proposed gene repressors has been viewed with some scepticism. Wehave examined this question by the most direct approach - isolation and characteriza-tion of the crab sperm chromatin.
MATERIALS AND METHODSThe method for obtaining sperm cells varied slightly, depending upon the region in the
reproductive tract. AH isolations were done on fresh adult Libinia emarginata at 0-4 °C. Crabswere obtained from the supply department at the Marine Biological Laboratory, Woods Hole,Mass., during summer. Purified sperm were used for extraction of total sperm basic proteinsand DNA.
Abbreviations used. SSW, 0-25 M sucrose-filtered seawater; SSC/10, 0-015 M NaCl-o-ooi5 Msodium citrate, pH 7-0; saline-disodium EDTA, 0-05 M NaCl-o-io M disodium ethylene-diaminetetra-acetate, pH 8-o.
Isolation of testis sperm
Testes were freed from other tissues and homogenized with two or three strokes in a Ten-Broeck ground-glass homogenizer in several volumes of SSW. The homogenate was thenfiltered through x 4 cheesecloth and cells pelleted by centrifugation in the 870 rotor of an Inter-national B-20 centrifuge at 1500 rev/min for 15 min at 4 °C. The pellet was resuspended andwashed repeatedly on the centrifuge in SSW until the supernatant was clear. Cells wereresuspended in SSW, layered over 1 M sucrose-seawater and centrifuged in an Internationalclinical centrifuge at full speed for 7 min. The interface contained intact cells and debris, andwas discarded. Sperm pelleted, and were resuspended in SSW, layered over a 50-ml preformedsucrose gradient (1-0-2-2 M sucrose in 3 mM MgClt), and allowed to stand for 24 h at 1 g.Sperm formed a sharp band, which was removed by aspiration with a syringe needle. Purity oftestis sperm isolated in this manner was determined by direct count to be at least 95 % spermand 5 % somatic cells.
Isolation of vas deferens sperm
The vas deferens was arbitrarily divided into proximal (nearest the testes) and distal regions.It was found that sperm within the proximal region occur in 2 forms: free and spermatophore-enclosed. Spermatophores in this species are spherical non-cellular capsules containing fromone to several sperm (Hinsch, 1969) and are not easily broken by methods of homogenizationwhich are sufficiently gentle to prevent disruption of the freed sperm cells.
For sperm cell isolation, the appropriate region of the vas deferens was removed into SSW,minced with scissors, and stirred magnetically for 15 min. This gentle method of freeing spermdiminished contamination by the somatic cells of the vas deferens walls. The suspension wasfiltered through x 4 cheesecloth to remove residual vas deferens material, and sperm andspermatophores were pelleted from the filtrate and washed as described above. Pellets wereresuspended in SSW and layered over a sucrose gradient for fractionation at unit gravity, as fortestis sperm. This yielded 2 sharp bands: sperm and spermatophores. Purity of sperm (withrespect to contamination by somatic cells) was determined by direct counts to be at least 99 %,and always contained a few small spermatophores, whereas spermatophore preparations alwayscontained some free sperm.
Isolation of seminal receptacle sperm
Observation of mating habits in this species (Hinsch, 1968) has shown that females usuallymate just before or just following release of a batch of zoeae larvae, and rarely to never whilefreshly fertilized eggs (bright orange coloured) or developing embryos are being carried. Five' stages' of seminal receptacle sperm were isolated according to the stage of embryonic develop-ment following the procedure described for isolation of vas deferens sperm. Purity of these

Crab sperm chromatin analysis 133
sperm often exceeded 99 %, as determined by direct counting. The 5 seminal receptacle spermstages were arbitrarily designated (in order of increasing age): orange, red, red-brown, earlybrown, and late brown ('zoeae').
Isolation of sperm chromatin and acrosomes
The complete vas deferens was removed into ice-cold saline-disodium EDTA, minced,stirred magnetically for 15 min to free the spermatophores and sperm, filtered through x 4cheesecloth and centrifuged in the 870 rotor of an International B-20 centrifuge at 800 rev/minfor 10 min at 4 °C. The pellet was resuspended and washed twice more on the centrifuge insaline-disodium EDTA. The pellet containing spermatophores and free sperm was resuspendedin 50 ml of saline-disodium EDTA containing 0-25 ml 2-octanol to prevent foaming. Tissueswere homogenized in a model 45 VirTis homogenizer at 85 V for 45 s and centrifuged in theInternational B-20 centrifuge at 6000 rev/min for 15 min. The pellet contained acrosomes andbroken spermatophore walls and the supernatant the chromatin. The pellet was resuspended insaline-disodium EDTA and washed twice more on the centrifuge. The supernatants werepooled for chromatin purification; the pellets were saved for acrosome isolation.
Our technique for chromatin purification is modified from Bonner et al. (1968). The pooled6000-rev/min supernatants were centrifuged in the 50 Ti Spinco rotor at 45 000 rev/min for 1 hto pellet broken membranes and tiny debris, which was discarded. The supernatant was quiteviscous, and was decanted and pelleted by recentrifugation at 45000 rev/min for 14 h. Theresulting translucent gelatinous pellet of crude chromatin was resuspended in saline-disodiumEDTA with the aid of aTen-Broeck ground-glass homogenizer and again pelleted at 45 000 rev/min for 14 h. The washed pellet was twice resuapended in 0-05 M Tris, pH 8, stirred mag-netically for 1 h and pelleted. The pellet was then resuspended in o-oi M Tris, pH 8, stirredmagnetically 4 h, dialysed overnight against this, and sheared with the VirTis homogenizer at25 V for 90 s. After centrifugation in the International B-20 centrifuge at 12500 rev/min for 30min, the water-clear supernatant (sheared, purified chromatin) was dialysed against severalchanges of SSC/10 for 24 h, with stirring. This chromatin preparation was utilized for electronmicroscopy and biochemical analysis.
The acrosome and spermatophore-containing pellet was resuspended in saline-disodiumEDTA and homogenized in the VirTis at 25 V for 90 8. Centrifugation at 2000 rev/min for 10min pelleted the spermatophores, while acrosomes remained in the supernatant. The isolatedacrosomes were pelleted at 6000 rev/min, 15 min, and washed with 0-05 M Tris, pH 8, and001 M Tris, pH 8. These were resuspended in 0-25 M sucrose-o-oi M Tris, pH 8, layered over a50-ml preformed sucrose gradient (0-25-1-7 M sucrose in o-oi M Tris, pH 8) and allowed to sitfor 18-24 h at 4 °C. A band of acrosomes formed near the top of the tube. This acrosomefraction was removed by aspiration with a syringe needle and utilized for electron microscopyand biochemical analysis.
Isolation of DNADNA was isolated from testes and sperm using the methods of Smith (1963, 1964) as modified
by Skinner (1967).For recovery of DNA from sperm chromatin, sperm chromatin was isolated by the centri-
fugation method outlined above, except that o-io M NaCl-o-2o M disodium EDTA, pH 8, wasused throughout, to minimize degradation by DNase in this crab material, which is rich indivalent cations (Skinner, Marsh & Cook, 1965). Following the final dialysis against SSC/10,chromatin (1-5 O.D.JM units) was treated with pancreatic RNase (Worthington, 50 /tg/ml,made DNase free by heating to 80 °C, 30 min) for 30 min at 37 °C, followed by pronase(Calbiochem) which had been made DNase-free (Hotta & Bassel, 1965) by incubating 50 /*g/mlat room temperature for 8-12 h with gentle agitation. DNA was then separated from theenzymes by centrifugation in the 50 Ti Spinco rotor at 45 000 rev/min for 24 h, which pelletedthe DNA. DNA was then resuspended in SSC/10, dialysed against this for 24 h, and used foranalysis.

134 J- C Vaughn and G. W. Hinsch
Analysis of DNA and chromatin
DNA was estimated by the diphenylamine technique (Dische, 1955) and protein by theLowry method (Lowry, Rosebrough, Fair & Randall, 1951), using bovine serum albumin(Sigma) as the standard. Ultraviolet absorption spectra for DNA, chromatin and proteinpreparations were manually determined with a Zeiss PMQ II spectrophotometer.
Chromatin and CsCl (Schwarz-Mann, optical grade) in SSC/10 were mixed to a final con-centration of 209 M in CsCl and 0-25-1-5 mg DNA per 5-ml centrifuge tube, as described byBonner et al. (1968). These tubes were then centrifuged to equilibrium for 48 h at 20 CC in theSW-39 Spinco rotor at 36000 rev/min, which results in a gradient with a density ranging fromabout 120 to 1-40. DNA, with a density of about 170, pellets. Each tube was then punctured,taking care to by-pass the pellet of DNA, and 5-drop fractions were collected. An equal volumeof 0-25 N HC1 was added to each tube and the absorbance determined at 230 nm.
Sperm chromatin protein, after dialysis against 025 N HC1 to remove CsCl, was precipitatedby the addition of 2 volumes of 100 % ethanol, and washed twice in 100 % ethanol, followed by3 washes with acetone. The air-dried powder was used for amino acid analysis, and also forelectrophoresis.
Protein samples (1-6 mg) were hydrolysed individually in sealed evacuated tubes containing6 ml of 6 N HC1 for 24 h, and dried in a rotary evaporator at 40 °C. Distilled water was added3 times to each sample and they were redried. The dried hydrolysates were then dissolved in anappropriate volume (1-1-4-2 ml) of pH 22 sodium citrate buffer and applied to the columns ina Phoenix model K 5000 amino acid analyser, using standard procedures.
Melting profiles of DNA and chromatin were done manually on a Zeiss PMQ II spectro-photometer using a tightly stoppered silica water-jacketed cuvette, monitored with a tele-thermometer (YSI model 42SC) and coupled to a constant-temperature recirculating water bath(Haake model FJ). Meltings were performed in SSC/10, pH 70, following 10 min of specimendegassing under reduced pressure. At each temperature setting, 10 min were allowed fortemperature equilibrium before reading the absorbance, and each reading was double-checked.Melting profiles were taken in the DNA concentration range of 0-6—1-2 O.D.M0 units. Theprocedures of Mandel & Marmur (1968) were followed, although no correction was made forconcentration dilution caused by solvent expansion at the higher temperatures.
Protein extraction and polyacrylamide electrophoresis
Purified sperm from various regions of the reproductive tract were resuspended in ice-cold025 N HC1 and stirred overnight. Isolated and purified acrosomes were also extracted. Follow-ing centrifugation in the International B-20 centrifuge at 12500 rev/min for 30 min, thesupernatant was utilized for electrophoretic analysis. A crude preparation of histones was alsoprepared from the isolated nuclei of zoeae larvae in the same manner. These nuclei wereprepared by homogenization of zoeae in 025 M sucrose-i mM MgCl2, followed by repeatedlow-speed washings on the centrifuge. These washed crude nuclei were then layered over a pre-formed sucrose gradient (10-2-2 M sucrose in 3 mM MgCl^ and centrifuged in an Internationalclinical centrifuge at full speed for 5 min. A sharp band containing purified nuclei was removedby aspiration with a syringe needle.
Proteins in 025 N HC1 were analysed according to the polyacrylamide urea gel electrophoresistechnique described by Bonner et al. (1968), with modifications. The running gel acrylamideconcentration was 11 %.
Electron microscopy
Preparations of isolated chromatin in SSC/10 (5 O.D.j^, units) were streaked on to Formvarcarbon-coated grids, fixed in Karnovsky's (1965) paraformaldehyde-glutaraldehyde mixture,rinsed in distilled water, dehydrated in 100% ethanol and air-dried after passing through iso-amyl acetate (Ris, 1966). Alternatively, grids containing streaked unfixed chromatin werefloated on 1 % aqueous uranyl acetate for 1 h, fixed, dehydrated, and dried. Other grids werestreaked and floated on either DNase-free pronase (o-i mg/ml in SSC/10) or DNase (01 mg/mlin 3 O-OOM MgSO4) for 1 h before fixation, dehydration, and drying. In some cases streaked

Crab sperm chromatin analysis 135
grids were enzyme treated, floated upon uranyl acetate, then fixed, dehydrated, and dried.Dried grids were postfixed for 1 h with osmium vapour at 60 °C. Grids were examined with anHitachi model 8 electron microscope.
Sperm and isolated acrosomes were pelleted and fixed in Karnovsky's mixture (1965). Afterwashing, materials were postfixed in 1 % OsO4 in phosphate buffer, dehydrated in acetone, andembedded in Spurr (1969). In some cases, fixed pellets were treated for 1 h with 05 % uranylacetate before dehydration. Thin sections were cut on a Porter-Blum MT-2, stained withuranyl acetate (Watson, 1958) and lead citrate (Venable & Coggleshall, 1965) and viewed with aJEM-100B electron microscope.
Cytochemistry
The following cytochemical methods were applied to sperm from testis, vas deferens, and the5 different stages of seminal receptacle sperm, to assure detection of newly arising nuclear basicproteins (if any). We were especially interested in examining sperm at all stages, to examine thepreliminary report of Fenwick & Hinsch (1969) that seminal receptacle sperm of this speciescontain nuclear protamines.
Purified sperm cell preparations in SSW were fixed in solution for an hour (or longer) byadding an equal volume of 8 % formaldehyde—seawater. Fixed cells were smeared on togelatinized glass slides, air-dried and postfixed in 4 % formaldehyde-seawater for 2 h, rinsedseveral hours in running tap water, and air-dried.
DNA was localized by a modification of the Feulgen technique (Bloch & Godman, 1955) andhistones and other basic proteins with the pH 81 fast green method (Alfert & Geschwind, 1953)or the picric acid-bromphenol blue technique (Bloch & Hew, i960). Lysine residues wereoccasionally blocked by deamination (Van Slyke, 1911) or acetylation (Monne & Slautterback,1951), to test for presence of high protein arginine/lysine ratios, using the modificationsdescribed by Bloch (1966).
RESULTS
Sperm cell isolation and ultrastructure
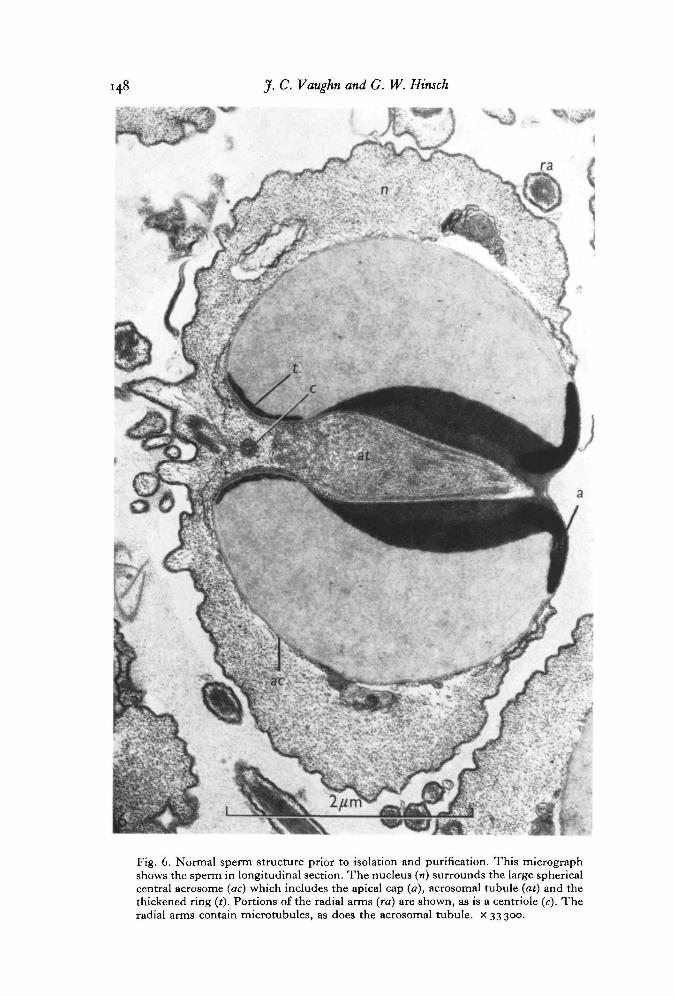
The typical morphology and terminology of Libinia sperm cell components is intro-duced in Figs. 5-6.
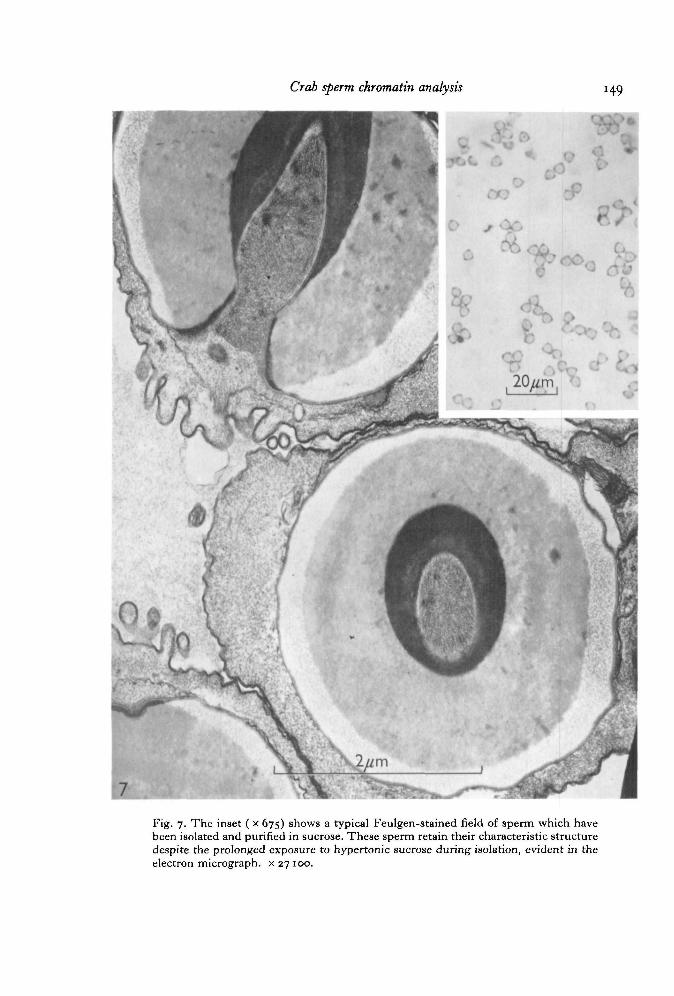
Sperm which have been purified by repeated washings in SSW (Fig. 7) followed bybanding in a sucrose gradient retain the typical ultrastructure characteristic of thesecells, as described by Hinsch (1969). Microtubules are seen in the acrosomal tubuleand also within the nuclear radial arms. The acrosomal components appear to bereasonably intact, so that regions of different densities are seen within the main body ofthe acrosome, and the apical cap, walls of the acrosomal tubule, and thickened ring arerecognizable. Chromatin remains typically non-condensed.
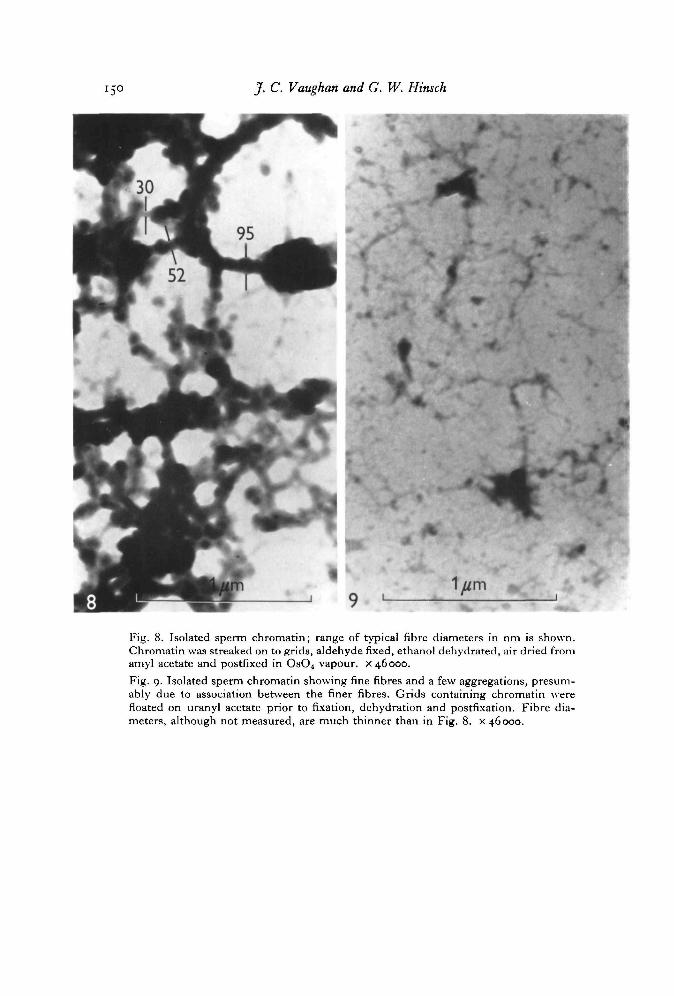
Sperm-isolated chromatin which was not stabilized by uranyl ions (Fig. 8) showstangled masses of fibres having irregular thickness (about 30-95 nm, although someregions are much thicker). In the vicinity of these clumps of fibres are seen manyfiner fibres. Following pronase treatment of non-stabilized chromatin the thick fibresare no longer seen, suggesting that protein is responsible for the clumping behaviour ofthis histoneless chromatin.
Sperm chromatin stabilized with uranyl acetate appears as very fine fibres (diameterunmeasured), Fig. 9. Aggregation of these thinner fibres to form thicker ones isevident.
A typical field of isolated acrosomes is shown in Fig. 10. These were apparently free

136 J. C. Vaughn and G. W. Hinsch
of contaminating nuclear material, as the electron micrograph (Fig. 11) shows. It isapparent that much of the substance of these acrosomes has been extracted during theisolation, presumably due to the Tris buffer washings.
Polyacrylamide electrophoresis
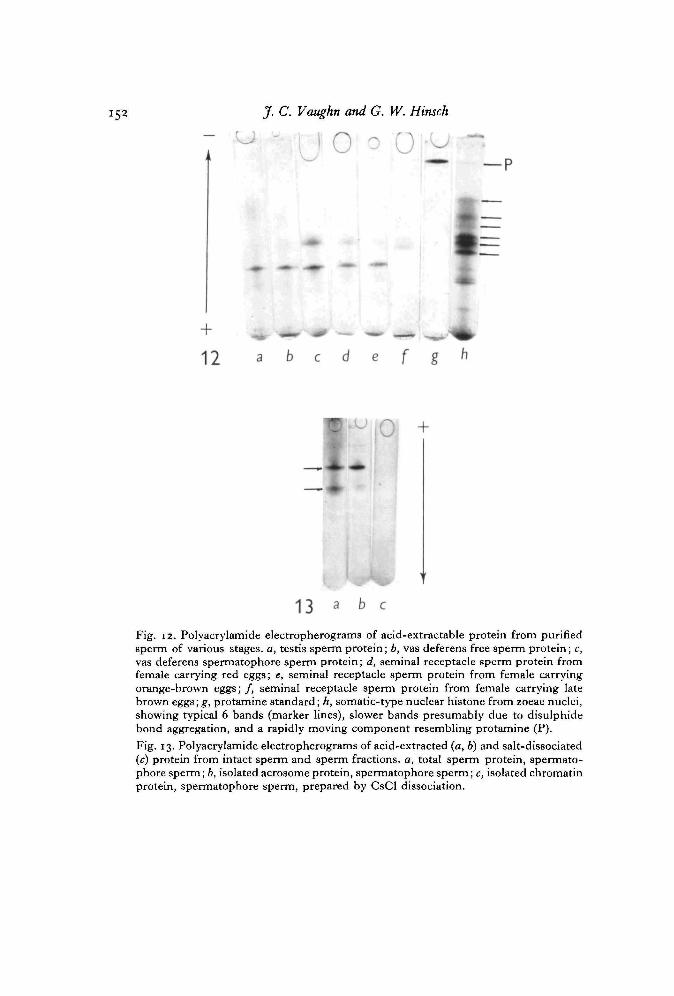
Electropherograms for acid-extracted protein from isolated sperm at various stagesare shown in Fig. 12. The pattern for zoeae nuclear histone (a control) closelyresembles that reported for plant (Fambrough & Bonner, 1969) and animal (Senshu& Iwai, 1967) nuclear histones, and includes at least 6 major bands. Presence of a bandwhich moves like protamine was observed not only in zoeae nuclear preparations, butalso in nuclei isolated from hepatopancreas and testis somatic cells. The validity andsignificance of this finding awaits further study. Sperm acid-extractable proteinincludes at least 2 bands at every stage except possibly long-retained seminal receptaclesperm, where the slower migrating component is not seen. One of these 2 bands movesslower than a typical histone, while the other moves about like a histone. No protamine-like band is present in any sperm stage, although in preparations highly contaminatedwith somatic cells such a band was seen (in addition to faint somatic histone bands).
Analysis of acid-extracted proteins from isolated acrosomes, whose purity wasestablished by electron microscopy (Fig. 13), shows the identical 2 bands seen inanalysis of whole-sperm extracts, while the CsCl-dissociable chromatin protein doesnot move toward the cathode. This shows that sperm chromatin does not containacid-extractable basic proteins (or salt-dissociable basic proteins), and that acrosomescontain at least 2 acid-extractable basic proteins but not a typical somatic-type histonepattern. The precise localization of these acrosomal basic proteins within the spermhas not been established, although stainable acrosomal basic proteins are found in theapical cap, the walls of the acrosomal tubule, and in the thickened ring (Table 3,P- 139)-
Nature of chromatin DNA-protein bonding
Isopycnic centrifugation in 2-09 M CsCl dissociates DNA from ionically bondedhistones and pellets the DNA in other systems (Bonner et al. 1968). In the presentexperiments the puncturing needle was always allowed to penetrate the tube farenough to by-pass the DNA pellet by a few fraction numbers. Protein was recoveredfrom at least one peak (Fig. 1) which was in a tube position removed from the positionof the DNA (demonstrated by DNA analysis and u.v. spectral absorption curves),showing that the protein and DNA are salt dissociable. This protein fraction wasutilized for electrophoretic and amino acid composition analysis.
Sperm chromatin chemical analysis
A typical u.v. absorption spectrum for sperm chromatin and isolated sperm DNAis given in Fig. 2. Presence of protein which co-sediments with sperm DNA in theisolated sperm chromatin was shown by demonstrating that this DNA is obtainable bytreating the chromatin with the proteolytic enzyme pronase, followed by resedimenta-tion of the DNA. The ratio of absorption values at 230:260:280 nm for this DNA is

Crab sperm chromatin analysis
1-50 r
1-00 -
D
60-50 -
0-0020
Fig. i. Caesium chloride density-gradient centrifugation of sperm deoxyribonucleo-protein, showing banding (separation) of protein from DNA (which pellets). Thetechnique of sample collection largely excludes DNA from the fractions. Although notshown, gradient density at completion of the run is through the range i-2C—1-40.
100 r-
Q
OE
Bx
260 290Wavelength, nm
Fig. 2. Spectral absorption curve for isolated sperm chromatin (I) and DNA (II). Bothshow a peak at about 260 nm, whereas chromatin also absorbs relatively heavily at 230and 280 nm.

138 J. C. Vaughn and G. W. Hinsch
Table i. Analytical data on isolated sperm chromatinfrom Libinia emarginata
Table 2
Amino acid
LysHisArgAspThrSerGluProGlyAlaICysValMetHeLeuT y rPheT r pN H ,
DNA, /tg/O.D. ISOProtein, /ig/O.D.2,0 1Protein/DNAAbsorption 280Absorption 260
n m
n m
. Amino acid composition
L. emarginata\sperm chromatin
protein
I
8-52i-6o3-25
11-87I I - 8 I
6 4 41581S-856-126-33—6-92—3-624-68o-8o0 9 0——
Unknown peak 5-42
Basic/acidic: 048
II
7 7 41-092-41
11-4111-486 7 0
14-726 2 86 055'55—6 0 50 9 23-264 7 51 3 2
6 1 5——
4 i 3
°-43
I II
490 52-6137 169323 3'2
059 067
of various nucleoproteins*
Whole calfjthymushistone
16-0
7-9
4 75'5S'28-o5-o8-3
1 5 1o-o7 00 7
3-57 62 20 0
o-o
2 O O
E. coli§DNA
associatedprotein
6 31-7
6-i
io-o574 3
U S4-2
n - 8IO-Io-66-9i -6
5'38-62-33- io-o——
o-66
• Data are expressed as mol per 100 mol of amino acid. Dashes indicate the amino acid wasnot tested for or the amount was not reported,
f Values not corrected for hydrolytic losses.X Data taken from Rasmussen, Stroier, Murray & Luck (1962).§ Data taken from Zubay & Watson (1959).
0-39:1 -oo: 0-51; for the isolated sperm chromatin it is 0-96:1 -oo: 0-59. The absorptionincreases in sperm chromatin at 230 and 280 nm are presumably due to bound protein.
Direct chemical analyses of sperm chromatin for protein and DNA (Table 1) showa protein/DNA ratio of 2-3-3-2. The amino acid analysis of this chromatin protein(Table 2) shows that it is highly acidic, with a ratio of basic/acidic amino acids equal to0-43-0-48.
Crab sperm chromatin analysts
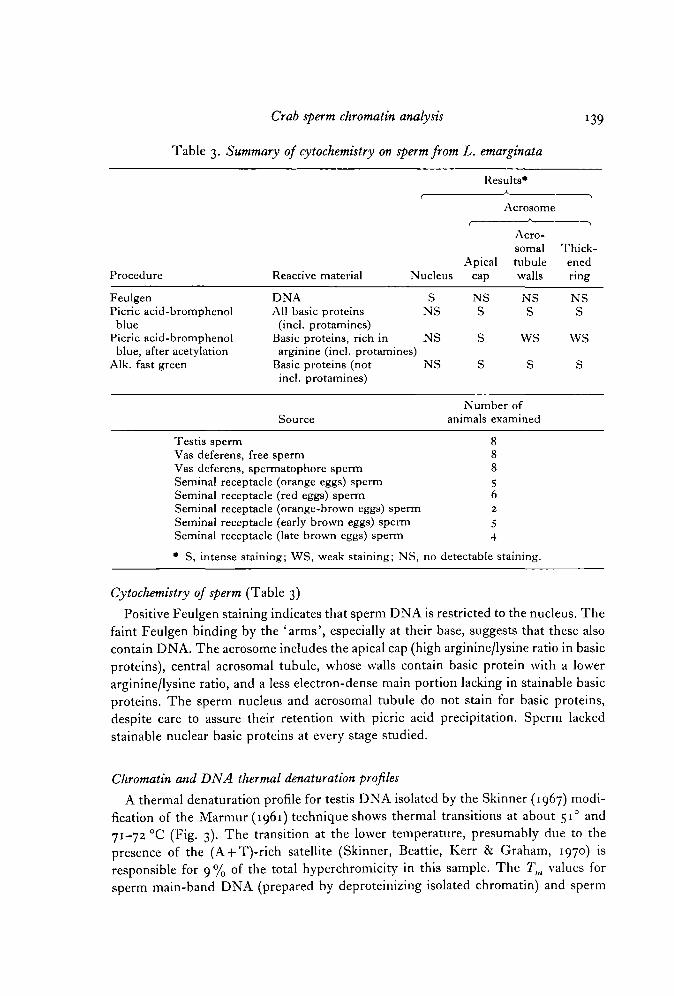
Table 3. Summary of cytochemistry on sperm from L. emarginata
139
Results*
t
ApicalProcedure Reactive material Nucleus cap
Feulgen DNAPicric acid-bromphenol All basic proteinsblue (incl. protamines)
Picric acid-bromphenol Basic proteins, rich inblue, after acetylation arginine (incl. protamines)
Alk. fast green Basic proteins (notincl. protamines)
Source
Testis spermVas deferens, free spermVas deferens, spermatophore spermSeminal receptacle (orange eggs) spermSeminal receptacle (red eggs) spermSeminal receptacle (orange-brown eggs) spermSeminal receptacle (early brown eggs) spermSeminal receptacle (late brown eggs) sperm
• S, intense staining; WS, weak staining; NS,
S NSNS S
NS S
NS S
Acrosome
Acro-somaltubulewalls
NSS
WS
s
Number ofanimals examined
88856
i 2
54
no detectable sitaining.
•i
Thick-enedring
NSS
WS
s
Cytochemistry of sperm (Table 3)
Positive Feulgen staining indicates that sperm DNA is restricted to the nucleus. Thefaint Feulgen binding by the 'arms', especially at their base, suggests that these alsocontain DNA. The acrosome includes the apical cap (high arginine/lysine ratio in basicproteins), central acrosomal tubule, whose walls contain basic protein with a lowerarginine/lysine ratio, and a less electron-dense main portion lacking in stainable basicproteins. The sperm nucleus and acrosomal tubule do not stain for basic proteins,despite care to assure their retention with picric acid precipitation. Sperm lackedstainable nuclear basic proteins at every stage studied.
Chromatin and DNA thermal denaturation profiles
A thermal denaturation profile for testis DNA isolated by the Skinner (1967) modi-fication of the Marmur (1961) technique shows thermal transitions at about 51° and71-72 °C (Fig. 3). The transition at the lower temperature, presumably due to thepresence of the (A + T)-rich satellite (Skinner, Beattie, Kerr & Graham, 1970) isresponsible for 9 % of the total hyperchromicity in this sample. The Tm values forsperm main-band DNA (prepared by deproteinizing isolated chromatin) and sperm

140 J. C. Vaughn and G. W. Hinsch
1001—
8.1
25 35 45 55 65 75Temperature, °C
85
Fig. 3. Melting profile of testis DNA in SSC/10, isolated accordingto the modified Marmur technique.
chromatin in SSC/10 (Fig. 4) are within 1 °C of each other, suggesting that thechromatin protein is not histone or protamine, as these should protect the DNA frommelting (Shih & Bonner, 1970; Olins, 1969). This expectation has recently beenestablished for Libinia somatic cell chromatin (Vaughn, 1971). This sperm chromatinprotein behaves like the acidic proteins which comprise part of the chromatin of othereukaryotes, and which do not alter the Tm in those systems (Bonner et al. 1968). Thehyperchromicity of this crab sperm DNA and chromatin is about 38 %, suggestingthat little nucleic acid denaturation occurred during the isolation procedures. Absenceof an (A + T)-rich satellite transition in the region of 51 °C in SSC/10 melts of spermDNA obtained by deproteinization of isolated chromatin is due to loss of this com-ponent during the sperm chromatin isolation procedure, as sperm DNA isolateddirectly by the Marmur procedure shows a typical transition at about 51 °C, in additionto the main band transition at about 71 °C (Vaughn, 1971).

Crab sperm chromatin analysts 141
100
75
50
I
25
35 45 55 65 75Temperature, °C
85 95
Fig. 4. Melting profiles of sperm DNA (#) prepared by deproteinization of isolatedsperm chromatin, and of the isolated sperm chromatin (A) prior to deproteinization.Melting was done in SSC/10.
DISCUSSION
Nuclear histone changes
In a review of histone changes occurring during spermiogenesis in a variety ofanimals and a few plants, Bloch (1969) recognized 5 classes of histone changes. Onecalled 'crustacean-type' was the least well understood class. Published accountsdealing with these sperm support the conclusion that they are lacking in nuclear basicproteins, although the experimental support for this has not been, to our way ofthinking, sufficiently rigorous. Chevaillier (1966) reported lack of binding of basicprotein-specific dyes in sperm nuclei of the crab Eupagurus bemhardus, although thesesperm contain acrosomal basic proteins which he hypothesized may representsloughed nuclear histones, which he subsequently named 'decapodine' (Chevaillier,1967). Chevaillier (1968) found that the nuclear chromatin fibres in these sectionedsperm clump unless stabilized by heavy metal ions, after which their diameterapproaches that of DNA. This fibre morphology was not affected by pronase treat-ment but was destroyed by DNase, suggesting that these fibres are naked DNA.Langreth (1969) has reported the absence of stainable nuclear basic proteins in sperm

142 J. C. Vaughn and G. W. Hinsch
nuclei of testes, spermatophores and seminal receptacles in various species of the crabgenus Cancer. In combined fine-structural and cytochemical studies, she observes acorrelation between appearance of very fine testis sperm chromatin fibres (resemblingthose in bacteria, which lack histone, as shown by Raaf & Bonner, 1968) and loss ofnuclear basic protein stainability. Quantitative DNA and histone measurements havedemonstrated a gradual loss of spermatid nuclear somatic-type histone which is notaccompanied by replacement with gamete-type histones or protamines in the sand crabEmerita analoga (Vaughn, 1968; Vaughn et al. 1969; Vaughn & Thomson, 1972).
The present study is the first to directly isolate and analyse a crustacean spermchromatin, applying numerous different approaches to establish the nature of thismaterial. Our results are in general agreement with those obtained by the cytochemicaland fine-structural approaches: sperm chromatin in the crab L. emarginata is devoid ofhistone and protamine. Our results are not consistent with those cytochemical studiesin which crab sperm DNA was found to be protamine-bound (Fenwick & Hinsch,1969) or to be unassociated with any protein whatever (Chevaillier, 1968; Langreth,1969). However, we have not performed experiments to rule out the possibility thata part (or all) of the observed Libinia sperm chromatin protein is artifactual, arisingfrom secondary association with the DNA during chromatin isolation. In a studyposing this very problem, inconclusive results were obtained by Raaf & Bonner (1968)with bacterial chromatin, when it was found that addition of naked DNA to acell homogenate resulted in a DNA-protein conjugate. Our experiments allow onlythe conclusion that Libinia sperm nuclei and isolated chromatin are lacking inDNA-associated basic proteins.
We show that clean acrosome preparations contain stainable and also acid-extract-able basic proteins whose electrophoretic behaviour is different from histones, whereaspreparations contaminated with somatic cells show bands typical of somatic histones.The non-histone nature of the acrosomal basic proteins in L. emarginata is consistentwith results obtained for acrosomes isolated from the marine echiuroid worm Urechiscaupo (Das, Micou-Eastwood & Alfert, 1967). However, these basic proteins werereportedly not acid-extractable.
Numerous reports have appeared relating the state of DNA-histone interaction tothe control of DNA-dependent RNA synthesis in eukaryotes (Huang & Bonner, 1962;Pogo, Allfrey & Mirsky, 1966; Skalka et al. 1966. For review, see Elgin, Froehner,Smart & Bonner, 1971). Histones in eukaryotes may only be non-specific repressors,keeping the genes in a repressed state unless either activated by other informationalmacromoltcules (possibly one of the short-lived nuclear RNAs, Britten & Davidson,1969), or by acetylation, methylation, or phosphorylation of histones. The correlationof net positive charge of the basic proteins with degree of gene repression (Skalka et al.1966), and the generality that during animal spermatogenesis somatic-type histonesare replaced by gamete-type histones or protamines which have a very high netpositive charge (for review, see Bloch, 1966), have led to a common belief (of uncertainorigin) that the appearance of these gamete-type nuclear basic proteins is in some wayrelated to the turning off and continued repression of sperm genes at the transcriptionallevel. This viewpoint is difficult to reconcile with observations that the sperm genes

Crab sperm chromatin analysis 143
generally turn off before the gamete-type basic proteins are even synthesized (Das,Kaufmann & Gay, 1964a, b; Monesi, 1964, 1965, 1967; Vaughn, 1966; DeLeon &Vaughn, 1970), suggesting that the mechanism of sperm gene repression lies else-where. The present report that sperm DNA in Libinia is not at any time associated withbasic protein of any kind may mean merely that a different protein is doing the job of ahistone, or that a unique mechanism is operative in repressing crab sperm genes - onewith little general importance. An alternative possibility would be that a commonmechanism explains repression of sperm genes in all species but that that mechanismis not directly related to histone or protamine. If true, then crab sperm chromatinwould represent a relatively simple system for the study of such gene-repressingsystems.
The idea that gamete nuclear basic proteins may represent functionless, degeneratehistones derived from random genetic drift was originally suggested in a paper byBloch (1969). Absence of all nuclear basic proteins in the decapod crustacean sperm isnot out of keeping with this interesting hypothesis, since a functionless protein mightnot be selected for survival unless some new role were acquired.
Satellite DNA
Crustacean satellite DNA was first described in the crab Cancer borealis (Sueoka,1961), and more recently in several crustacean species by Smith (1964) and by theOak Ridge group (Skinner, 1967; Skinner et al. 1970; Graham & Skinner; 1970).Several of these species contain both an (A + T)-rich and a (G + C)-rich satellite.Libinia dubia dAT satellite was reported to be composed of more than 90 % alternatingadenylate and thymidylate residues, to have a Tm of 52 °C in SSC/10, to comprisesome 6 % of the total nuclear DNA, and to band at a density of 1-675 g/cm3 in CsCl(Skinner et al. 1970). Our finding of a DNA component comprising about 7-9 % of thetotal hyperchromicity and having a Tm of about 51 °C in SSC/10 in the sperm andtestis of L. emarginata is in close agreement with these data. The actual (A + T)-richsatellite content in our study would of course presumably be somewhat less than7-9 % of the total DNA, as the hyperchromicity of these satellites is generally greaterthan analytical ultracentrifuge densitometer tracings (Marmur & Doty, 1959).
Our results suggest that the dAT satellite in L. emarginata sperm is complexed withmacromolecules, presumably protein, which are somehow different from those asso-ciated with the bulk sperm DNA, as our procedure for chromatin isolation by differ-ential centrifugation (in which the pellets were discarded early in the procedure) doesnot recover this component. This interpretation is consistent with other studies(Vaughn, 1971) and with the recent work of Duerksen & McCarthy (1971), whoshowed by controlled differential shearing and centrifugation of testis chromatin of thecrab Cancer magister that the dAT satellite is enriched in the condensed chromatinfraction. The demonstration of this satellite in purified sperm, in which there are noconventional mitochondria, supports the recent reports by Skinner et al. (1970) thatthis satellite is nuclear and not mitochondrial. This conclusion is also supported by theobservation (Vaughn, 1971) that isolated somatic cell chromatin contains the usualsatellite components.

144 J- c- Vaughn and G. W. Hinsch
The authors are indebted to Miss Marilyn Cayer (University of Miami) for some of theelectron microscopy included in this paper, to C. R. Windsor (University of Miami) for per-forming the amino acid analyses of sperm chromatin protein, and to Dr C. B. Metz (Universityof Miami) and the Fertilization and Gamete Physiology Training Program at the MarineBiological Laboratory, Woods Hole (NIH grant 5-Toi-HDooo26-oo), where this work wasperformed. The first author also acknowledges research support by Miami University.
Preliminary accounts of the findings were presented at the General Meetings, MarineBiological Laboratory, Woods Hole, Mass., August 1970, at the AAAS Annual Meeting,Chicago, Illinois, December 1970, and at the American Society for Cell Biology Meeting, NewOrleans, La., November 1971.
Figs. 5 and 6 are published with the permission of the Academic Press, Inc., which holds thecopyright for these figures.
REFERENCESALFERT, M. & GESCHWIND, I. I. (1953). A selective staining method for the basic proteins of cell
nuclei. Proc. natn. Acad. Sci. U.S.A. 39, 991-999.BLOCH, D. P. (1966). Cytochemistry of the histones. Protoplasmatologia 5 (3d), 1-56.BLOCH, D. P. (1969). A catalog of sperm histones. Genetics, Princeton 61 (suppl.), 93-111.BLOCH, D. P. & GODMAN, G. C. (1955). A microphotometric study of the syntheses of deoxy-
ribonucleic acid and nuclear histone. J. biophys. biochem. Cytol. 1, 17-28.BLOCH, D. P. & HEW, H. Y. C. (i960). Changes in nuclear histones during fertilization, and
early embryonic development in the pulmonate snail, Helix aspersa. J. biophys. biochem.Cytol. 8, 69-81.
BONNER, J., CHALKLEY, G. R., DAHMUS, M., FAMBROUGH, D., FUJIMURA, F., HUANG, R. C,HUBERMAN, J., JENSEN, R., MARUSHIGE, K., OHLENBUSCH, H., OLIVERA, B. & WIDHOLM, J.(1968). Isolation and characterization of chromosomal nucleoproteins. In Methods inEnzymology,vo\. 12, part B (ed. L. Grossman&K. Moldave), pp. 3-65. New York: AcademicPress.
BRITTEN, R. J. & DAVIDSON, E. H. (1969). Gene regulation for higher cells: a theory. Science,N. Y. 165, 349-357-
CHEVAILLIER, P. (1966). Contribution a lYtude du complexe ADN-histone dans le sperma-tozoide du pagure Eupagurus bernhardus L. (Crustace decapode). J. Microscopie 5, 730-758.
CHEVAILLIER, P. (1967). Mise en evidence et etude cytochimique d'une proteine basique extra-nucleaire dans les spermatozoides des crustaces decapodes. J. Cell Biol. 32, 547-556.
CHEVAILLIER, P. (1968). Etude cytochimique ultrastructurale des nucleoproteines dans lespermatozoide du pagure Eupagurus bernhardus L. (Crustace ddcapode). J. Microscopie 7,107-114.
DAS, C. C, KAUFMANN, B. P. & GAY, H. (1964a). Histone-protein transition in Drosophilamelanogaster. I. Changes during spermatogenesis. Expl Cell Res. 35, 507-514.
DAS, C. C, KAUFMANN, B. P. & GAY, H. (1964ft). Autoradiographic evidence of synthesis of anarginine-rich histone during spermiogenesis in Drosophila melanogaster. Nature, Lond. 204,1008-1009.
DAS, N. K., MICOU-EASTWOOD, J. & ALFERT, M. (1967). Cytochemical and biochemicalproperties of basic proteins of Urechis acrosomes. J. Cell Biol. 35, 455-458.
DELEON, R. P. & VAUGHN, J. C. (1970). Histone metabolism during spermatogenesis in thetoad Bufo americanus. Am. Zool. 10, 317.
DISCHE, Z. (1955). Color reactions of nucleic acid components. In The Nucleic Acids, vol. 1 (ed.E. Chargaff & J. N. Davidson), p. 287. New York: Academic Press.
DUERKSEN, J. D. & MCCARTHY, B. J. (1971). Distribution of deoxyribonucleic acid sequences infractionated chromatin. Biochemistry, N.Y. 10, 1471-1478.
ELGIN, S., FROEHNER, S., SMART, J. & BONNER, J. (1971). The biology and chemistry ofchromosomal proteins. In Advances in Cell and Molecular Biology, vol. 1 (ed. E. J. DuPraw),pp. 1-57. New York: Academic Press.
FAMBROUGH, D. M. & BONNER, J. (1969). Limited molecular heterogeneity of plant histones.Biochim. biophys. Ada 175, 113-122.

Crab sperm chromatin analysis 145
FENWICK, R. G. & HINSCH, G. W. (1969). Cytochemical localization of materials within thesperm of Libinia emarginata. Biol. Bull. mar. biol. Lab., Woods Hole 137, 398-399.
GEORGIEV, G. P. (1969). Histones and the control of gene action. A. Rev. Gen. 3, 155-180.GRAHAM, D. E. & SKINNER, D. M. (1970). Conservation of nucleotide sequences in d(G + C)-
rich satellite DNAs of Crustacea. J. Cell Biol. 47, 76 A.HINSCH, G. W. (1968). Reproductive behavior in the spider crab, Libinia emarginata (L.). Biol.
Bull. mar. biol. Lab., Woods Hole 135, 273-278.HINSCH, G. W. (1969). Microtubules in the sperm of the spider crab, Libinia emarginata L.
J. Ultrastruct. Res. 29, 525-534.HOTTA, Y. & BASSEL, A. (1965). Molecular size and circularity of DNA in cells of mammals and
higher plants. Proc. natn. Acad. Sci. U.S.A. 53, 356-362.HUANG, R-C. & BONNER, J. (1962). Histone, a suppressor of chromosomal RNA synthesis.
Proc. natn. Acad. Sci. U.S.A. 48, 1216-1222.KARNOVSKY, M. J. (1965). A formaldehyde-glutaraldehyde fixative of high osmolality for use in
electron microscopy. J. Cell BM. 27, 137 A.LANGRETH, S. G. (1969). Spermiogenesis in Cancer crabs. J. Cell Biol. 43, 575—603.LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measurement
with the Folin phenol reagent. J. biol. Chem. 193, 265-275.MANDEL, M. & MARMUR, J. (1968). Use of ultraviolet absorbance-temperature profile for
determining the guanine plus cytosine content of DNA. In Methods in Enzymology, vol. 12,part B (ed. L. Grossman & K. Moldave), pp. 195-206. New York: Academic Press.
MARMUR, J. (1961). A procedure for the isolation of deoxyribonucleic acid from micro-organisms. J. molec. Biol. 3, 208-218.
MARMUR, J. & DOTY, P. (1959). Heterogeneity in deoxyribonucleic acids. I. Dependence oncomposition of the configurational stability of deoxyribonucleic acids. Nature, Lond. 183,1427-1429.
MONESI, V. (1964). Autoradiographic evidence of a nuclear histone synthesis during mousespermiogenesis in the absence of detectable quantities of nuclear ribonucleic acid. Expl CellRes. 36, 683-688.
MONESI, V. (1965). Synthetic activities during spermatogenesis in the mouse. RNA and protein.Expl Cell Res. 39, 197-224.
MONESI, V. (1967). Ribonucleic acid and protein synthesis during differentiation of male germcells in the mouse. Archs Anat. microsc. Morph. exp. 56, 61-74.
MONNE, L. & SLAUTTERBACK, D. B. (1951). The disappearance of protoplasmic acidophilia upondeamination. Ark. Zool., Ser. 2, i, 455-462.
OLINS, D. E. (1969). Interactions of lysine-rich histones and DNA._7. molec. Biol. 43, 439-460.OLINS, D. E., OLINS, A. L. & VON HIPPEL, P. H. (1968). On the structure and stability of
DNA-protamine and DNA-polypeptide complexes.^, molec. Biol. 33, 265-281.PHILLIPS, D. M. P. (ed.) (1971). Histones and Nticleoliistones. London: Plenum Press.POGO, B., ALLFREY, V. G. & MIRSKY, A. E. (1966). RNA synthesis and histone acetylation
during the course of gene activation in lymphocytes. Proc. natn. Acad. Sci. U.S.A. 55,805-812.
RAAF, J. & BONNER, J. (1968). Properties of Escherichia coli deoxyribonucleoprotein as comparedwith the deoxyribonucleoprotein of higher organisms. Archs Biochem. Biophys. 125, 567-579.
RASMUSSEN, P., STROIER, P., MURRAY, K. & LUCK, J. M. (1962). On the complexity of calfthymus histone. Biochemistry i, 79-89.
Ris, H. (1966). Fine structure of chromosomes. Proc. R. Soc. B 164, 246—257.SENSHU, T. & IWAI, K. (1967). Disc electrophoresis of calf thymus histones. Seikagaku 39,
906-911.SHIH, T. Y. & BONNER, J. (1970). Thermal denaturation and template properties of DNA
complexes with purified histone fractions, y. molec. Biol. 48, 469-487.SKALKA, A., FOWLER, A. & HURWITZ, J. (1966). The effect of histones on the enzymatic syn-
thesis of ribonucleic acid. y. biol. Chem. 241, 588—596.SKINNER, D. M. (1967). Satellite DNA's in the crabs Gecarcinus lateralis and Cancer pagurus.
Proc. natn. Acad. Sci. U.S.A. 58, 103-110.SKINNER, D. M., BEATTIE, W. G., KERR, M. S. & GRAHAM, D. E. (1970). Satellite DNAs in
Crustacea: Two different components with the same density in neutral CsCl gradients.Nature, Lond. 227, 837-839.

146 J. C. Vaughn and G. W. Hinsch
:SKINNER, D. M., MARSH, D. J. & COOK, J. S. (1965). Physiological salt solution for the landcrab, Gecarcimis lateralis. Biol. Bull. mar. biol. Lab., Woods Hole 129, 355-365.
SMITH, M. (1963). Deoxyribonucleic acids in crabs of the genus Cancer. Biocliem. biophys. Res.Commun. 10, 67-72.
.SMITH, M. (1964). Deoxyribonucleic acids of Crustacea. J. molec. Biol. 9, 17-23.SPURR, A. R. (1969). A low viscosity epoxy resin embedding medium for electron microscopy.
J. Ultrastnict. Res. 26, 31-43.SUEOKA, N. (1961). Variation and heterogeneity of base composition of deoxyribonucleic acids:
a compilation of old and new data. J. molec. Biol. 3, 31-40.VAN SLYKE, D. (191 I) . A method for quantitative determination of aliphatic amino groups.
Applications to the study of proteolysis and proteolytic products. J. biol. Chem. 9, 185-204.VAUGHN, J. C. (1966). Relationship of the 'sphere chromatophile' to the fate of displaced histones
following histone transition in rat spermiogenesis. J. Cell Biol. 31, 257-278.VAUGHN, J. C. (1968). Changing nuclear histone patterns during development. I. Fertilization
and early cleavage in the crab, Emerita analoga. J. Histochem. Cytochem. 16, 473-479.VAUGHN, J. C. (1971). Satellite DNA analysis in the spider crab, Libinia emarginata. Biol. Bull.
mar. biol. Lab., Woods Hole 141, 405.VAUGHN, J. C, CHAITOFF, J., DELEON, R., GARLAND, C. & THOMSON, L. (1969). Changing
nuclear histone patterns during development. II. Isolation and partial characterization of'decapodine' from sperm cells of the crab, Emerita analoga. Expl Cell Res. 54, 362-366.
VAUGHN, J. C. & THOMSON, L. A. (1972). A kinetic study of DNA and basic protein metabolismduring spermatogenesis in the sand crab, Emerita analoga. J. Cell Biol. 52, 322-337.
VENABLE, J. H. & COGGESHALL, R. J. (1965). A simplified lead citrate stain for use in electronmicroscopy. J. Cell Biol. 25, 407-408.
WATSON, M. L. (1958). Staining of tissue sections for electron microscopy with heavy metals. .7.biophys. biochem. Cytol. 4, 475-478.
ZUBAY, G. & WATSON, M. R. (1959). The absence of histone in the bacterium Escherichia coli. I.Preparation and analysis of nucleoprotein extract. .7. biophys. biochem. Cytol. 5, 51-54.
{Received 9 November 1971)

Crab sperm chromatin analysis 147
iFig. 5. Normal sperm structure prior to isolation and purification. The phase-contrastphotomicrograph in the inset ( x 2500) shows a polar view of a sperm, looking downupon the circular apical cap (a). The acrosome is surrounded by the nucleus (n), whichcontains non-condensed chromatin fibres and is continuous with the 4 radial arms (ra)of which 3 are seen. These radial arms contain microtubules. The electron micrographshows a sperm in transverse section through the acrosome (ac) which includes the walls(ui) of the central acrosomal tubule, x 31 650.
148 J. C. Vaughn and G. W. Hinsch
Fig. 6. Normal sperm structure prior to isolation and purification. This micrographshows the sperm in longitudinal section. The nucleus (n) surrounds the large sphericalcentral acrosome (ac) which includes the apical cap (a), acrosomal tubule (at) and thethickened ring (t). Portions of the radial arms (ra) are shown, as is a centriole (c). Theradial arms contain microtubules, as does the acrosomal tubule, x 33300.
Crab sperm chromatin analysis 149
Fig. 7. The inset ( x 675) shows a typical Feulgen-stained field of sperm which havebeen isolated and purified in sucrose. These sperm retain their characteristic structuredespite the prolonged exposure to hypertonic sucrose during isolation, evident in theelectron micrograph, x 27 100.
J. C. Vaughan and G. W. Hinsch
Fig. 8. Isolated sperm chromatin; range of typical fibre diameters in nm is shown.Chromatin was streaked on to grids, aldehyde fixed, ethanol dehydrated, air dried fromamyl acetate and postfixed in OsO4 vapour, x 46000.Fig. 9. Isolated sperm chromatin showing fine fibres and a few aggregations, presum-ably due to association between the finer fibres. Grids containing chromatin werefloated on uranyl acetate prior to fixation, dehydration and postfixation. Fibre dia-meters, although not measured, are much thinner than in Fig. 8. x 46000.

Crab sperm chromatin analysis
f
2firc\
Fig. 10. A typical field of isolated acrosomes, showing both lateral (/) and polar (p)views. The nucleus has apparently been completely removed. The field also contains afew bits of 8permatophore wall, x 570.Fig. 11. Electron micrograph to demonstrate purity of isolated acrosomes, showingcomplete removal of nuclear material. Both transverse and longitudinal views are seen.The washings during purification have also removed much of the solid material fromthe acrosomes, although the acrosomal tubule (at), apical cap (a) and thickened ring (t)are present, xo.100.
JJ1 J. C. Vaughn and G. W. Hinsch
^ . .— —P
1 . i .•
1 2 a b c d e f g h
i: 0
1 3 a b c
Fig. 12. Polyacrylamide electropherograms of acid-extractable protein from purifiedsperm of various stages, a, testis sperm protein; b, vas deferens free sperm protein; c,vas deferens spermatophore sperm protein; d, seminal receptacle sperm protein fromfemale carrying red eggs; e, seminal receptacle sperm protein from female carryingorange-brown eggs; /, seminal receptacle sperm protein from female carrying latebrown eggs; g, protamine standard; h, somatic-type nuclear histone from zoeae nuclei,showing typical 6 bands (marker lines), slower bands presumably due to disulphidebond aggregation, and a rapidly moving component resembling protamine (P).
Fig. 13. Polyacrylamide electropherograms of acid-extracted (a, b) and salt-dissociated(c) protein from intact sperm and sperm fractions, a, total sperm protein, spermato-phore sperm; b, isolated acrosome protein, spermatophore sperm; c, isolated chromatinprotein, spermatophore sperm, prepared by CsCl dissociation.