isolation and biochemical characterization of streptomyces clavuligerus mutants in the biosynthesis...
TRANSCRIPT

Appl Microbiol Biotechnol (1988) 27:510--516 Applied Microbiology
Biotechnology © Springer-Verlag 1988
Isolation and biochemical characterization of Streptomyces clavuliyerus mutants in the biosynthesis of clavulanic acid and cephamycin C*
Jorge Romero, Paloma Liras, and Juan F. Martin
Departamento de Microbiologia, Facultad de Biologia, Universidad de Le6n, Le6n, Spain
Summary. Seven mutants of Streptomyces clavuli- gerus blocked in the biosynthesis of clavulanic acid, cephamycin C, or both antibiotics, have been isolated and characterized. Mutants ncal and nca2 were unable to synthesize clavulanic acid but produced cephamycin C. Mutants ncel and nce2 were completely blocked in cephamycin C production but formed clavulanic acid. A third group (mutants nccl, ncc4 and ncc5) failed to pro- duce both antibiotics. Arginase activity (forming ornithine) was very low in mutants nccl and ncc5. All the mutants blocked in clavulanic acid biosyn- thesis showed a normal ornithine-6-aminotrans- ferase activity. Mutant nccl, blocked in cephamy- cin biosynthesis, lacked completely lysine-e-ami- notransferase (forming a-aminoadipic acid) and isopenicillin N synthase. Two other mutants (nce2 and nce5) lacked isopenicillin N synthase. There was a good correlation between the isopenicillin N synthase and the lysine-e-aminotransferase ac- tivities of the nca mutants and the ability of those strains to produce cephamycin C. The condensing enzyme involved in the formation of the clavu- lanic acid nucleus appears to be different from the isopenicillin N synthase.
Introduction
The biosynthesis of clavulanic acid, a potent fl- lactamase inhibitor produced by Streptomyces cla- vuligerus (Higgens and Kastner 1971) has been in- vestigated by feeding labelled precursors to the
* Dedicated to Professor H.-J. Rehm on the occasion of his 60th birthday
Offprint requests to: J. F. Martin
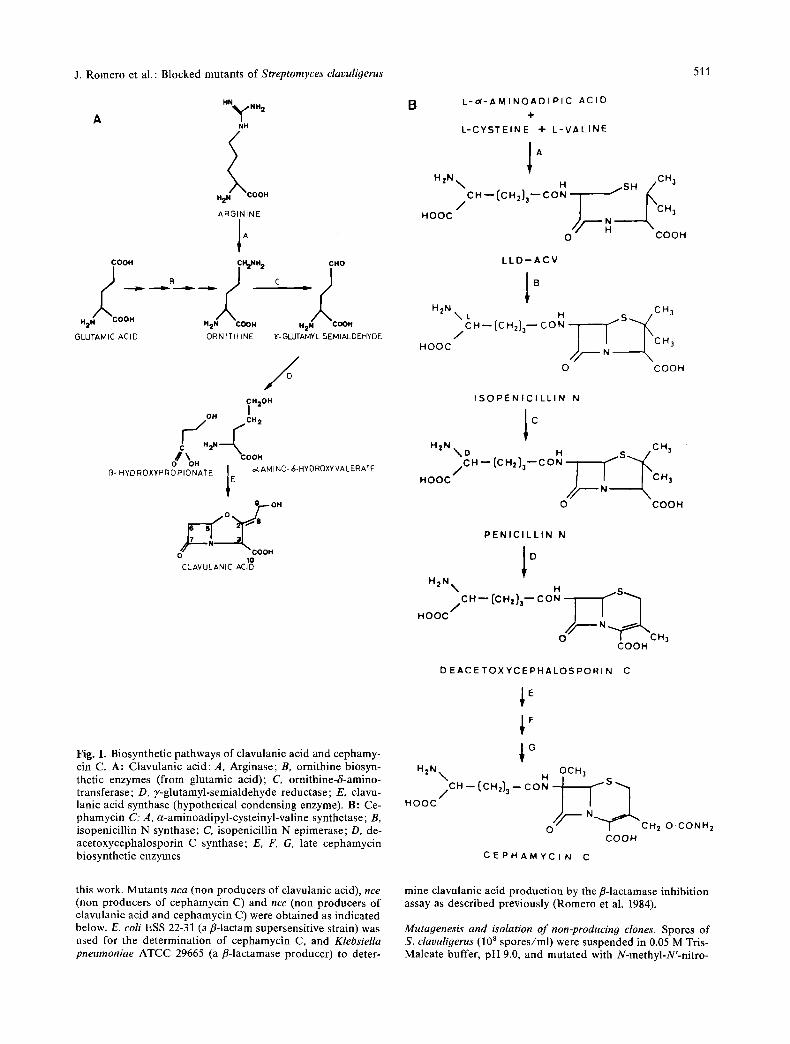
fermentation (Elson and Oliver 1978; Elson et al. 1982). It appears that the three carbon atoms of the fl-lactam ring (C-5, C-6 and C-7) (the C3 unit), are derived from glycerol via a fl-hydroxypropion- ate intermediate (Gutman et al. 1985; Townsend and Ho 1985b) (Fig. 1). The five other carbon atoms (the C5 unit) of clavulanic acid (C-2, C-3, C-8, C-9 and C-10) derive from ornithine, which in turn originates from arginine (Townsend and Ho 1985a; Romero et al. 1986).
An hypothetical cyclization reaction appears to be involved in the condensation of the C3 and C5 units to form the fl-lactam-oxazolidine nucleus of clavulanic acid (Fig. 1), probably in a similar way to the cyclization reaction carried out by the isopenicillin N synthase that forms the fl-lactam- thiazolidine nucleus of isopenicillin N from the cysteine and valine residues of the tripeptide 6- (L-a-aminoadipyl)-L-cysteinyl-D-valine (Jensen et al. 1986). Both, clavulanic acid and cephamycin are produced simultaneously by S. clavuligerus and the same cyclase might carry out the cycliza- tion leading to the nucleus of both compounds. To advance in our understanding of the steps in- volved in clavulanic acid and cephamycin biosyn- thesis it was convenient to isolate and character- ize mutants blocked in the biosynthetic pathways. We describe in this paper the isolation and char- acterization of a series of mutants blocked in the biosynthesis of either clavulanic acid or cephamy- cin or both antibiotics and provide evidence that the cyclization reactions involved in the formation of the nucleus of both compounds are different.
Materials and methods
Strains. Single colony isolates of S. clavuligerus NRRL 3585, producer of clavulanic acid and cephamycin C were used in
J. Romero et al. : Blocked mutants of Streptomyces clavuligerus
A
HN y NH
NH
H2N~COOH ARGININE
ooo. ix.., /.o 2 c ;
H2 0 H2N COON H2 N COOH GI-UTAMIC ACIO ORN ffHINE "~-GLUTAMYL SEMIALDEHYDE
/ / ~ H20H
OH / CH2
I \ coo. O OH / ~L-AMI NO- $-HYD ROXY VA LERATE
r}- HYD ROXYP fRO PIONATE E
t
r-(°-¢. /~---~ ~k. oo. 0
111 CLAVULANfC ACID
Fig. 1. Biosynthetic pathways of clavulanic acid and cephamy- cin C. A: Clavulanic acid: A, Arginase; B, ornithine biosyn- thetic enzymes (from glutamic acid); C, ornithine-S-amino- transferase; D, 7-glutamyl-semialdehyde reductase; E, clavu- lanic acid synthase (hypothetical condensing enzyme). B: Ce- phamycin C: A, a-aminoadipyl-cysteinyl-valine synthetase; B, isopenicillin N synthase; C, isopenicillin N epimerase; D, de- acetoxycephalosporin C synthase; E, F, G, late cephamycin biosynthetic enzymes
this work. Mutants nca (non producers of clavulanic acid), nce (non producers of cephamycin C) and ncc (non producers of clavulanic acid and cephamycin C) were obtained as indicated below. E. coli ESS 22-31 (a fl-lactam supersensitive strain) was used for the determination of cephamycin C, and Klebsiella pneumoniae ATCC 29665 (a fl-lactamase producer) to deter-
511
B
HOOC
L-~-AMINOAOIPlC ACID +
L-CYSTEINE + L-VALINE
HzN~ H f S H t CH3 /CH--{CH2}3--CON J ~ ' H N CH3
O COOH
LLD-ACV
H2N CH 3
CH--(CH~I~-- CON
HOOC / CH]
O COOH
I S O P E N I C I L L I N N
l C H z N ~ o H S CH~
/ c . - Cc.,),-c °N-T---F" .ooc ;, ' L " '
O COOH
PENICILLIN N
1 ° H2N\ H
HOoc/CH--[CH:,la--CON / ~ N . ~ /
0 I C H 3 COOH
D EACE TOXYCEPHALOSPORI N C
HzN,~ OCH 3 /CH__ (CHzla__ CON S
HOOC 0 CHz O CONH2
COOH CEPHAMYCI N C
mine clavulanic acid production by the fl-lactamase inhibition assay as described previously (Romero et al. 1984).
Mutagenesis and isolation of non-producin 9 clones. Spores of S. elavuligerus (108 spores/ml) were suspended in 0.05 M Tris- Maleate buffer, pH 9.0, and mutated with N-methyl-N'-nitro-
512 J. Romero et al. : Blocked mutants of Streptomyces clavuligerus
N-nitrosoguanidine (NTG) (1 mg/ml) for 20 min at 30°C (Delic et al. 1970). Under these conditions the mortality after 20 min was about 50%. The mutated spores were washed twice with sterile distilled water and plated after adequate dilutions in solid MEY medium (containing in g per liter, yeast extract, 4; maltose, I0; agar, 20). After 4 days, the colonies were trans- ferred to the same medium in well separated patches, and agar plugs were cut and incubated at 28°C for 3 days. The clavu- lanic acid and/or cephamycin produced were assayed using agar plugs (3 mm diameter) on lawns of K. pneumoniae or E. coli as test strains. Those clones that failed to synthesize anti- biotic were tested again twice and later incubated in liquid GSPG medium (Romero et al. 1984) for clavulanic acid and cephamycin production,
Catabolism of amino acids. To test the utilization of amino acids as carbon sources for growth or antibiotic production, plates containing minimal medium (Aharonowitz and Demain 1977) without glycerol, were supplemented with the adequate amino acids (1 mM). To study the utilization of amino acids as a nitrogen source, ammonium chloride was omitted and the plates were supplemented with glycerol (100 raM).
Cell free extracts. Cell free extracts were prepared by sonica- tion of mycelium of S. clavuligerus grown in GSPG medium for 36 h (Romero et al. 1984).
(L-a-Aminoadipyl)-eysteinyl-valine determination, a-Aminoadi- pyl-cysteinyl-valine (ACV) was determined in cells grown in GSPG medium for 36 h. Mycelium from a 50 ml culture was washed with saline solution, centrifuged and suspended in 5 ml of 10% trichloroacetic acid (TCA). After standing for 2 h at room temperature the pH was adjusted to 7.0 and the TCA was removed by extracting three times with diethylether, The cell extracts were lyophylized, resuspended in 500 p,1 of dis- tilled water and the peptides (without oxidation to their sul- fonic acid derivatives) were detected by thin layer electrophor- esis at pH 3.5 (L6pez-Nieto et al. 1985). Pure samples of a- aminoadipyl-cysteine, ACV and glutation were used as stand- ards.
Enzyme assays. Isopenicillin N synthase, isopenicillin N epi- merase and deacetoxycephalosporin C synthase were assayed as described by Jensen et al. (1983, 1985, 1986). Ornithine-6- aminotransferase (OAT), lysine-e-aminotransferase (LAT) and arginase assays were carried out as indicated by Harwood and Baumberg (1977).
Results
Utilization of antibiotic precursers by the wild type and the blocked mutants
Seven mutant clones that consistently did not pro- duce either cephamycin C (nee mutants) clavu- lanic acid (nca mutants) or both (ncc mutants) by the agar plug method were isolated and tested for auxotrophies and for antibiotic production in liquid medium.
All the non-producer mutants behaved as pro- totrophs, except strain nee5 which was a trypto- phan auxotroph unable to grow on indol-supple- mented minimal medium.
Utilization of amino acids (1 raM) that are precursors of either cephamycin C (e.g. lysine) (Kern et al. 1980) or clavulanic acid (ornithine, ar- ginine, glutamic acid, proline) (Romero et al. 1986) as sole nitrogen or carbon source for growth of S. clavuligerus and the non producer mutants was tested. Lysine, aspartate, arginine, glutamate and proline (at neutral pH) were all used as ni- trogen sources, but only proline was used as car- bon source for growth.
Production of clavulanic acid and cephamycin C in liquid GSPG medium
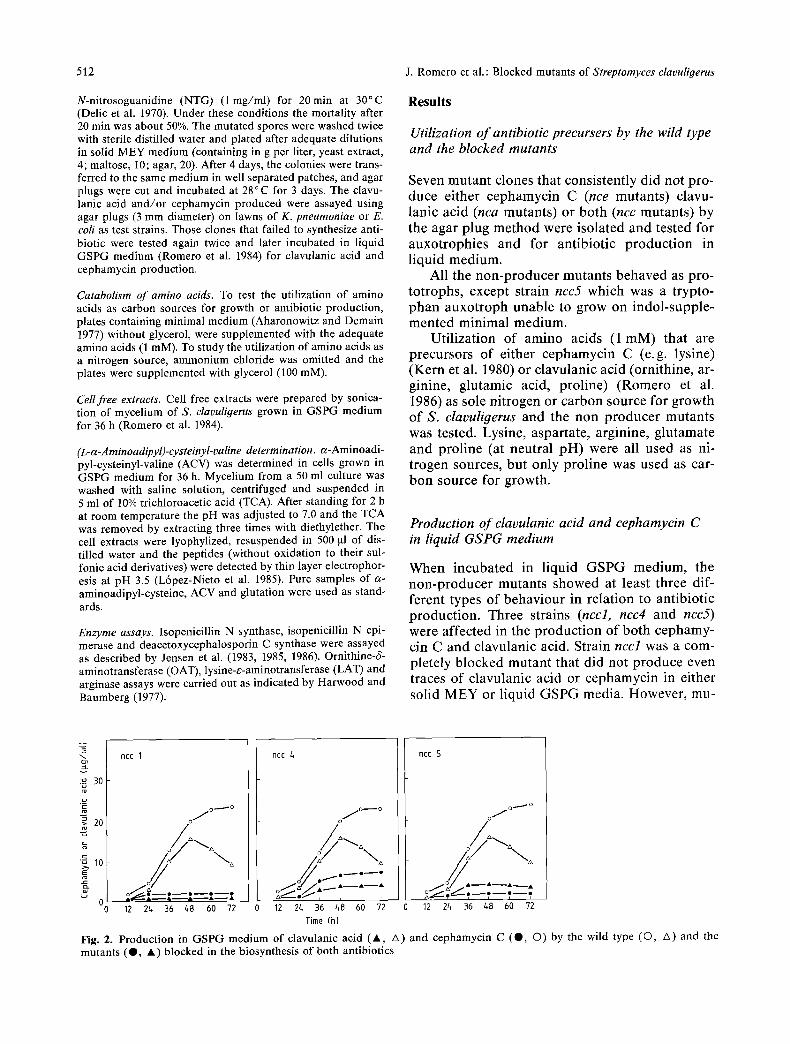
When incubated in liquid GSPG medium, the non-producer mutants showed at least three dif- ferent types of behaviour in relation to antibiotic production. Three strains (nccl, ncc4 and ncc5) were affected in the production of both cephamy- cin C and clavulanic acid. Strain nccl was a com- pletely blocked mutant that did not produce even traces of clavulanic acid or cephamycin in either solid MEY or liquid GSPG media. However, mu-
E
:=L
30 u m
.u g g 2o m o
ro g
0 o
ncc 1
o / O .--.---~ o
12 24 36 48 60 72
ncc 4
o
o ~ ~
12 24 36 /.8 60 72 Time (h)
ncc S
° / o ' ~ °
O ~ S a ~ A ~ & ~ A 12 24 36 48 60 72
Fig. 2. Production in GSPG medium of clavulanic acid ( a , A) and cephamycin C (O, O) by the wild type (©, A) and the mutants (O, a ) blocked in the biosynthesis of both antibiotics
J. Romero et al. : Blocked mutants of Streptomyces clavuligerus 513
E
:=1. 30
._c
u 0 o
nee 1
° / o ~ °
12 2t, 36 t,8 60 72
rice 2 ~ \ _
/o/ V °
~ - - 9 - - e - - 9 12 24 36 48 60 72
Time (h)
50
30
20
10
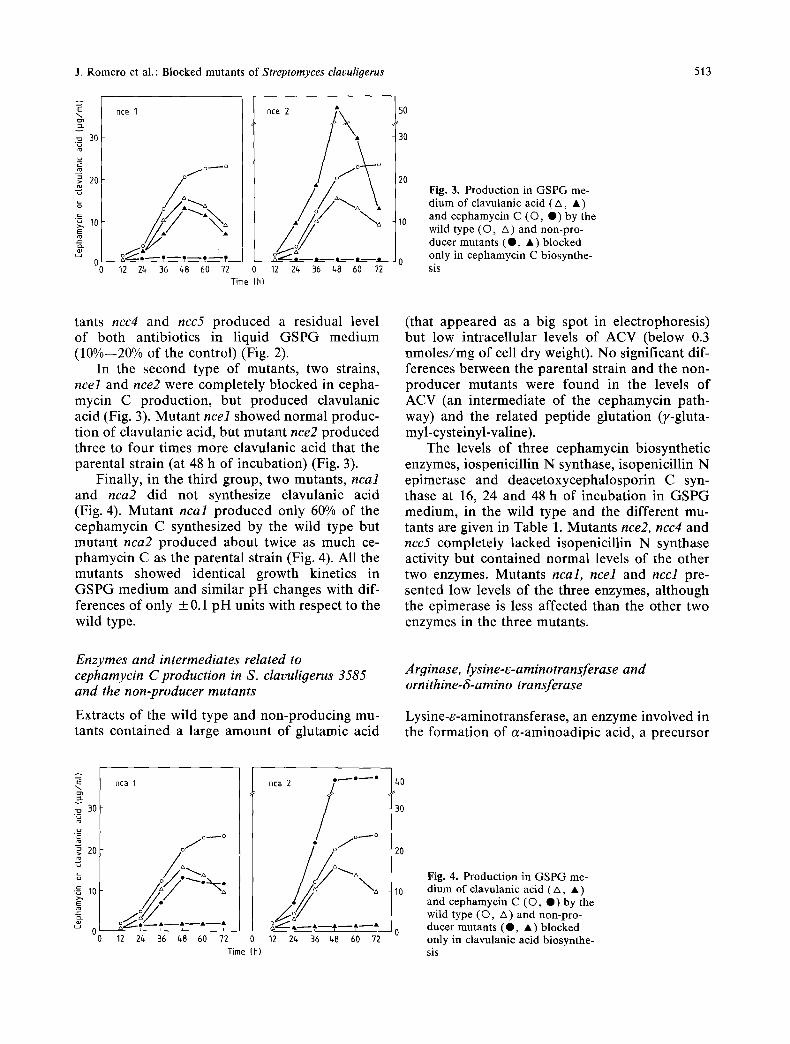
Fig. 3. Production in GSPG me- dium of clavulanic acid (/x, • ) and cephamycin C ((3, O) by the wild type (O, A) and non-pro- ducer mutants (Q, • ) blocked only in cephamycin C biosynthe- sis
tants ncc4 and ncc5 produced a residual level of both antibiotics in liquid GSPG medium (10%--20% of the control) (Fig. 2).
In the second type of mutants, two strains, ncel and nce2 were completely blocked in cepha- mycin C production, but produced clavulanic acid (Fig. 3). Mutant ncel showed normal produc- tion of clavulanic acid, but mutant nce2 produced three to four times more clavulanic acid that the parental strain (at 48 h of incubation) (Fig. 3).
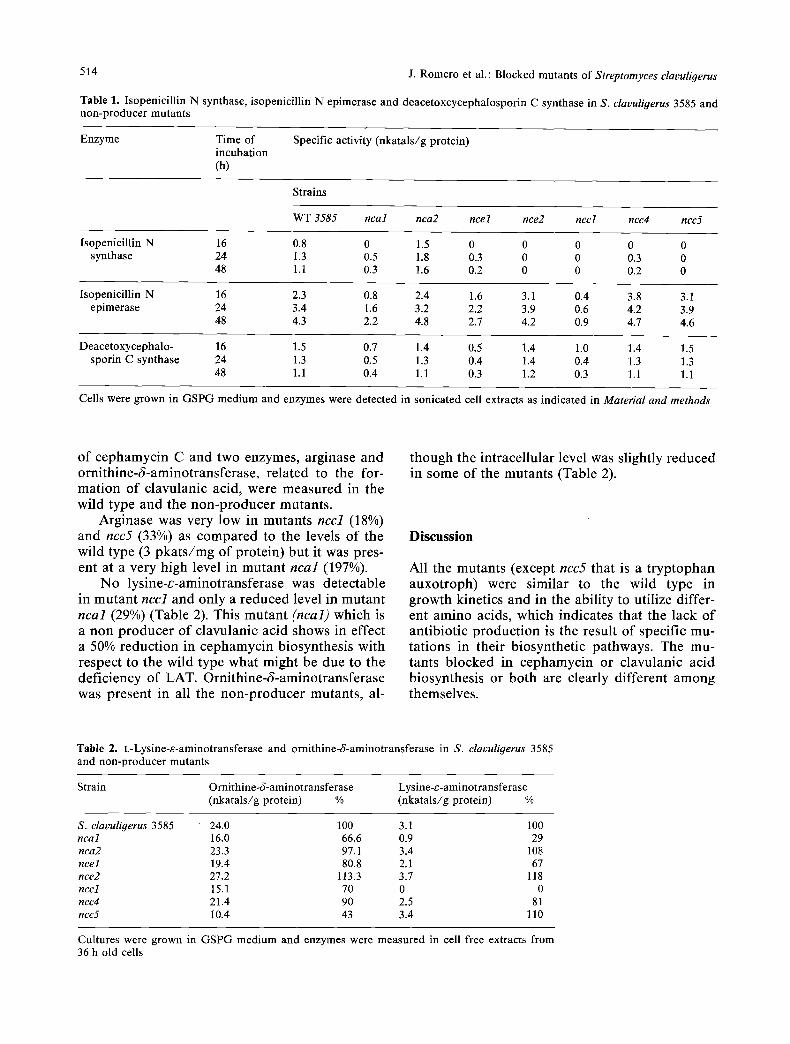
Finally, in the third group, two mutants, ncal and nca2 did not synthesize clavulanic acid (Fig. 4). Mutant ncal produced only 60% of the cephamycin C synthesized by the wild type but mutant nca2 produced about twice as much ce- phamycin C as the parental strain (Fig. 4). All the mutants showed identical growth kinetics in GSPG medium and similar pH changes with dif- ferences of only + 0.1 pH units with respect to the wild type.
Enzymes and intermediates related to cephamycin C production in S. clavuligerus 3585 and the non-producer mutants
Extracts of the wild type and non-producing mu- tants contained a large amount of glutamic acid
(that appeared as a big spot in electrophoresis) but low intracellular levels of ACV (below 0.3 nmoles/mg of cell dry weight). No significant dif- ferences between the parental strain and the non- producer mutants were found in the levels of ACV (an intermediate of the cephamycin path- way) and the related peptide glutation (~'-gluta- myl-cysteinyl-valine).
The levels of three cephamycin biosynthetic enzymes, iospenicillin N synthase, isopenicillin N epimerase and deacetoxycephalosporin C syn- thase at 16, 24 and 48 h of incubation in GSPG medium, in the wild type and the different mu- tants are given in Table 1. Mutants nce2, ncc4 and ncc5 completely lacked isopenicillin N synthase activity but contained normal levels of the other two enzymes. Mutants ncal, ncel and nccl pre- sented low levels of the three enzymes, although the epimerase is less affected than the other two enzymes in the three mutants.
Arginase, lysine-e-aminotransferase and ornithine-6-amino transferase
Lysine-e-aminotransferase, an enzyme involved in the formation of a-aminoadipic acid, a precursor
E
zt
30 m ._u m ~ 2o
c
E ro x= u
0
nca I
° / ° ~ °
12 24 36 48 60 72 Time (h)
nca 2 • • • 40 J ,
30
12 2(* 36 4B 60 72
Fig. 4. Production in GSPG me- dium of clavulanic acid (A, • ) and cephamycin C (O, O) by the wild type (Q, A) and non-pro- ducer mutants (O, &) blocked only in clavulanic acid biosynthe- sis
514 J. Romero et al.: Blocked mutants of Streptomyces clavuligerus
Table 1. Isopenicillin N synthase, isopenicillin N epimerase and deacetoxcycephalosporin C synthase in S. clavuligerus 3585 and non-producer mutants
Enzyme Time of incubation (h)
Specific activity (nkatals /g protein)
Strains
WT3585 ncal nca2 ncel nee2 nccl ncc4 ncc5
Isopenicillin N 16 0.8 0 1.5 0 0 0 0 0 synthase 24 1.3 0.5 1.8 0.3 0 0 0.3 0
48 1.1 0.3 1.6 0.2 0 0 0.2 0
Isopenicillin N 16 2.3 0.8 2.4 1.6 3.1 0.4 3.8 3.1 epimerase 24 3.4 1.6 3.2 2.2 3.9 0.6 4.2 3.9
48 4.3 2.2 4.8 2.7 4.2 0.9 4.7 4.6
Deacetoxycephalo- 16 1.5 0.7 1.4 0.5 1.4 1.0 1.4 1.5 sporin C synthase 24 1.3 0.5 1.3 0.4 1.4 0.4 1.3 1.3
48 1.1 0.4 1.1 0.3 1.2 0.3 1.1 1.1
Cells were grown in GSPG medium and enzymes were detected in sonicated cell extracts as indicated in Material and methods
of cephamycin C and two enzymes, arginase and ornithine-fi-aminotransferase, related to the for- mation of clavulanic acid, were measured in the wild type and the non-producer mutants.
Arginase was very low in mutants nccl (18%) and ncc5 (33%) as compared to the levels of the wild type (3 pkats /mg of protein) but it was pres- ent at a very high level in mutant ncal (197%).
No lysine-e-aminotransferase was detectable in mutant nccl and only a reduced level in mutant ncal (29%) (Table 2). This mutant (ncal) which is a non producer of clavulanic acid shows in effect a 50% reduction in cephamycin biosynthesis with respect to the wild type what might be due to the deficiency of LAT. Ornithine-fi-aminotransferase was present in all the non-producer mutants, al-
though the intracellular level was slightly reduced in some of the mutants (Table 2).
Discussion
All the mutants (except ncc5 that is a tryptophan auxotroph) were similar to the wild type in growth kinetics and in the ability to utilize differ- ent amino acids, which indicates that the lack of antibiotic production is the result of specific mu- tations in their biosynthetic pathways. The mu- tants blocked in cephamycin or clavulanic acid biosynthesis or both are clearly different among themselves.
Table 2. L-Lysine-e-aminotransferase and ornithine-~-aminotransferase in S. clavuligerus 3585 and non-producer mutants
Strain Ornithine-fi-aminotransferase Lysine-e-aminotransferase (nkatals /g protein) % (nkatals /g protein) %
S. clavuligerus 3585 24.0 100 3.1 100 heal 16.0 66.6 0.9 29 nca2 23.3 97. ! 3.4 108 neel 19.4 80.8 2.1 67 nee2 27.2 113.3 3.7 118 noel 15. I 70 0 0 nee4 21.4 90 2.5 81 nee5 10.4 43 3.4 110
Cultures were grown in GSPG medium and enzymes were measured in cell free extracts from 36 h old cells

J. Romero et al.: Blocked mutants of Streptomyces clavuligerus 515
The lack of assays for the enzymes involved in clavulanic acid biosynthesis makes it difficult to locate the mutations in the nca and ncc mutants that result in the inability to synthesize clavulanic acid. The step involved in the deamination of or- nithine to 7-glutamate-semialdehyde (an hypothe- tical intermediate in clavulanic acid biosynthesis) (Romero et al. 1986) catalyzed by OAT, is active in all mutants. Mutant nce2 that produces 3 to 4 times as much clavulanic acid as the wild type does have, in effect, the highest OAT activity. However, it is not completely clear whether the deamination of ornithine takes place at this stage (Fig. 1) or whether cyclization takes place using directly ornithine and the resulting amino group in carbon 9 is later deaminated and reduced to the primary alcohol existing in clavulanic acid (Townsend and Ho 1985a).
Arginase activity was very low in mutants nccl and ncc5 which might explain the deficiency of clavulanic acid biosynthesis due to the limitation of ornithine supply (Romero et al. 1986). It is likely that the other mutants blocked in clavulanic acid biosynthesis are altered in the "clavulanic acid synthetase" (cyclase).
The identification of mutants blocked in ce- phamycin biosynthesis has been more successful. The lack of LAT activity in mutant nccl may ex- plain the lack of cephamycin biosynthesis by this mutant. As indicated below this mutant is also de- ficient in isopenicillin N synthase and to a lesser extent in isopenicillin N isomerase and deacetox- ycephalosporin C synthase (expandase) (Table 1). It is interesting to note that another strain, ncal that showed a 29% LAT activity with respect to the wild type contains about 30% of the isopeni- cillin N synthase activity and about 50% of the epimerase and expandase activities of parental strain and produces 50% less cephamycin than the wild type. These results suggest that LAT and the three cephamycin biosynthetic enzymes may be regulated in a concerted manner as shown pre- viously in S. lactamdurans (Castro et al. 1985) and Acremoniurn chrysogenum (Ramos et al. 1986). Mutants nce2, nccl and ncc5 completely lacked isopenicillin N synthase, a specific step of the ce- phamycin biosynthetic pathway. In addition, mu- tants ncel and nce4 showed greatly reduced levels of the same enzyme.
Mutant nca2 (a cephamycin overproducer blocked in clavulanic acid biosynthesis) showed normal levels of the epimerase and expandase and an increased cyclase activity (about 140% to 200%), suggesting that the isopenicillin N syn- thase may be a rate limiting enzyme in the wild
type for cephamycin biosynthesis under these ex- perimental conditions.
The isolation of mutants ncel and nce2 which are deficient in the cyclization of ACV carried out by isopenicillin N synthase but still form normal levels of clavulanic acid rules out the hypothesis that the cyclization reactions to form isopenicillin N and clavulanic acid are carried out by the same enzyme. However, the high frequency of isolation of mutants blocked in both clavulanic acid and cephamycin (ncc mutants) might suggest that both biosynthetic processes are somehow related.
Acknowledgements. The authors wish to thank for a grant from the Diputaci6n Provincial of Le6n, and the technical assist- ance of Maria Paz Puertas AndrOs.
References
Aharonowitz Y, Demain AL (1977) Influence of inorganic phosphate and organic buffers on cephalosporin produc- tion by S. clavuligerus. Arch Microbiol 115:169--173
Castro JM, Liras P, Cort6s J, Martin JF (1985) Regulation of c¢-aminoadipyl-cysteinyl-valine, isopenicillin N synthetase isopenicillin N isomerase and deacetoxycephalosporin C synthetase by nitrogen sources in Streptomyces lactamdu- rans. Appl Microbiol Biotechnol 22:32--40
Delic V, Hopwood DA, Friend EJ (1970) Mutagenesis by N- methyl-N-nitrosoguanidine (NTG) in Streptomyces coelico- lor. Mut Res 9:167--182
Elson SW, Oliver RS (1978) Studies on the biosynthesis of cla- vulanic acid. I. Incorporation of C-labelled precursors. J Antibiot 31:586--592
Elson SW, Oliver RS, Bycroft BW, Faruk EA (1982) Studies on the biosynthesis of clavulanic acid. III. Incorporation of DL-3,4 ~4C-glutamic acid. J Antibiot 35:81--86
Gutman AL, Ribon V, Boltanski A (1985) Incorporation of [3- hydroxypropionate in the I~-lactam residue of clavulanic acid. J Chem Commun 1985:1627--1629
Harwood CR, Baumberg S (1977) Arginine hydroxamate re- sistant mutants of Bacillus subtilis with altered control of arginine metabolism. J Gen Microbiol 100:177--188
Higgens CE, Kastner RE (1971) Streptomyces clavuligerus sp. nov., a fl-lactam antibiotic producer. Int J Syst Bacteriol 21:326--331
Jensen SE, Westlake DWS, Wolfe S (1983) Partial purification and characterization of isopenicillin N epimerase ac- tivity from Streptomyces clavuligerus. Can J Microbiol 29:1526--1531
Jensen SE, Westlake DWS, Wolfe S (1985) Deacetoxycepha- losporin C synthetase and deacetoxycephalosporin C hy- droxylase are two separated enzymes in Streptomyces clavu- ligerus. J Antibiot 38:263--265
Jensen SE, Leskiw BK, Vining LC, Aharonowitz Y, Westlake DWS, Wolfe S (1986) Purification of isopenicillin N syn- thetase from Streptomyces clavuligerus. Can J Microbiol 32:953--957
Kern BA, Hendlin D, Inamine E (1980) L-lysine-e-aminotrans- ferase involved in cephamycin C synthesis in Streptomyces lactamdurans. Antimicrob Agents Chemother 17:679--685

516
L6pez-Nieto M J, Ramos FR, Luengo JM, Martin JF (1985) Characterization of the biosynthesis in vivo of a-aminoadi- pyl-cysteinyl-valine in Penieillium chrysogenum. Appl Mi- crobiol Biotechnol 22:343--351
Ramos FR, L6pez-Nieto M J, Martin JF (1986) Coordinate in- crease of isopenicillin N synthetase, isopenicillin N epi- merase and deacetoxycephalosporin C synthetase in a high cephalosporin-producing mutant of Acremonium chrysoge- hum and simultaneous loss of the three enzymes in a non- producing mutant. FEMS Microbiol Lett 35:123-- 127
Romero J, Liras P, Martin JF (1984) Dissociation of cephamy- cin and clavulanic acid biosynthesis in Streptomyces clavu- ligerus. Appl Microbiol Biotechnol 20:318--325
J. Romero et al. : Blocked mutants of Streptomyces clavuligerus
Romero J, Liras P, Martin JF (1986) Utilization of ornithine and arginine as specific precursors of clavulanic acid. Appl Environ Microbiol 52:892--897
Townsend CA, Ho M (1985a) Biosynthesis of clavulanic acid: origin of the C5 unit. J Am Chem Soc 107:1065--1066
Townsend CA, Ho M (1985b) Biosynthesis of clavulanic acid: origin of the C3 unit. J Am Chem Soc 107:1066--1068
Received May 12, 1987/Accepted September 25, 1987