ip3-activated channels and tip growth - esalq.usp.br · introduction fungi, the major decomposers...
TRANSCRIPT

IntroductionFungi, the major decomposers in terrestrial environments,explore new territory by polarized cellular growth (hyphalelongation). This growth trait is shared by members of otherkingdoms, but unlike other organisms, tip growth is thedominant form of growth in fungi.
Although Ca2+ gradients are an important component in alltip-growing organisms examined so far, fungi are notable inutilizing a different mechanism to generate and maintain thetip-high cytoplasmic Ca2+ gradient. Measurements of ionfluxes have identified a tip-localized Ca2+ influx from theexternal environment during growth in the oomyceteSaprolegnia ferax (Lew, 1999), root hairs (Felle and Hepler,1997) and pollen tubes (Pierson et al., 1996), but not in thefungus Neurospora crassa (Lew, 1999; Silverman-Gavrila andLew, 2000). Therefore, plasma membrane stretch-activatedCa2+ channels, identified as the mechanism of Ca2+ entry in S.ferax (Garrill et al., 1993), or another type of Ca2+ channel(Very and Davies, 2000) cannot be the mechanism generatingthe tip-high Ca2+ gradient in N. crassa. Because microinjectionof a Ca2+ chelator 1,2 bis(ortho-aminophenoxy)ethane-N,N,N′,N′-tetrapotassium acetate (BAPTA) into growing N.crassa hypha inhibits growth, and dissipates the tip highcytoplasmic Ca2+ gradient, there is a clear requirement for atip-high Ca2+ gradient during growth. The gradient must begenerated and maintained by internal mechanisms that are
likely to be shared by other fungi, and may be phylogeneticallyunique to the fungal kingdom (Silverman-Gavrila and Lew,2000).
Evidence is accumulating that supports the hypothesis thatthe fungal tip-high Ca2+ gradient is generated and maintainedinternally by inositol (1,4,5)-trisphosphate (IP3)-induced Ca2+
release from tip-localized vesicles and Ca2+ sequestration inthe ER behind the tip (Silverman-Gavrila and Lew, 2001) (Fig.1A). This is based upon inhibitor effects on hyphal growth andmorphology, correlated with cytoplasmic Ca2+ distributionsthat were imaged using dual dye ratioing.
In animal cells, the major site for IP3 receptors is the ER(Berridge and Irvine, 1989). In plants (and fungi), the vacuolarmembranes appear to contain Ca2+ channels, which can beactivated by IP3. For example, IP3 induces Ca2+ release fromplant hypocotyl microsomes (Drobak and Ferguson, 1985), andfrom vacuoles and tonoplast vesicles (Alexandre et al., 1990;Allen et al., 1995). The plant IP3 receptor was characterized indetail by Biswas (Biswas et al., 1995). In fungi, IP3 activatesCa2+ release from vacuoles of Candida albicans(Calvert andSanders, 1995), Saccharomyces cerevisiae(Belde et al., 1993)and N. crassa(Cornelius et al., 1989). InN. crassa, phosphategroups at the 1 and 5 positions of the inositol ring are the onlyessential arrangement required for receptor binding to releaseCa2+ from vacuoles, a unique characteristic compared withanimal IP3 receptors (Schultz et al., 1990). There are reports of
5013
Hyphal extension in fungi requires a tip-high Ca2+
gradient, which is generated and maintained internallyby inositol (1,4,5)-trisphosphate (IP3)-induced Ca2+
release from tip-localized vesicles and subapical Ca2+
sequestration. Using the planar bilayer method wedemonstrated the presence of two types of IP3-activatedCa2+ channels in Neurospora crassamembranes withdifferent conductances: one low (13 picosiemens), the otherhigh (77 picosiemens). On sucrose density gradients thelow conductance channel co-localized with endoplasmicreticulum and plasma membrane, and the highconductance channel co-localized with vacuolarmembranes. We correlated the effect of inhibitors onchannel activity with their effect on hyphal growth andCa2+ gradients. The inhibitor of IP3-induced Ca2+ release,2-aminoethoxidiphenylborate (2-APB), inhibits bothchannels, while heparin, 8-(N,N-diethylamino)-octyl-3,4,5-trimethoxybenzoate, hydrochloride (TMB-8) and
dantrolene inhibit only the large conductance channel.Because 2-APB inhibits hyphal growth and dissipates thetip-high cytosolic [Ca2+] gradient, whereas heparinmicroinjection, TMB-8 and dantrolene treatments do notaffect growth, we suggest that the small conductancechannel generates the obligatory tip-high Ca2+ gradientduring hyphal growth. Since IP3 production must becatalyzed by tip-localized phospholipase C, we show that anumber of phospholipase C inhibitors [neomycin, 1-[6-((17β-3-methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-1H-pyrrole-2,5-dione (U-73122) (but not the inactivepyrrolidine U-73343), 3-nitrocoumarin] inhibit hyphalgrowth and affect, similarly to 2-APB, the location ofvesicular Ca2+ imaged by chlortetracycline staining.
Key words: Fungal tip growth, IP3 receptor, Ca2+ gradient,Phospholipase C, Ca2+ channels, Planar lipid membrane
Summary
An IP3-activated Ca 2+ channel regulates fungal tipgrowthLorelei B. Silverman-Gavrila and Roger R. Lew*Biology Department, York University, 4700 Keele Street, Toronto, Ontario, M3J 1P3, Canada*Author for correspondence (e-mail: [email protected])
Accepted 23 September 2002Journal of Cell Science 115, 5013-5025 © 2002 The Company of Biologists Ltddoi:10.1242/jcs.00180
Research Article

5014
IP3-sensitive Ca2+ stores other than vacuoles in cauliflowerinflorescences (Muir and Sanders, 1997) and in growing pollentubes: in the nuclear-rough ER region (Franklin-Tong et al.,1996; Malho, 1998a) and from vesicles or ER (Zheng andYang, 2000). These observations suggest that plant and fungalvacuoles function as a major IP3-regulated intracellular Ca2+
store equivalent to ER and sarcoplasmic reticulum in animalcells, but IP3-activated Ca2+ release may also involve otherorganelles.
Enzymes and metabolites of the phosphoinositide cycle arepresent in plants (Stevenson et al., 2000), yeast and filamentousfungi. There are indications that one function of IP3 signalingis regulation of morphological transitions. IP3 is involved ingerm tube formation in the yeast-mycelium transition inOphiostoma ulmi (Brunton and Gadd, 1991). In C. albicans,there is an increase in IP3 during germ tube formation of theyeast to hyphal morphological transition (Gadd and Foster,1997). Ins(1,4,5)P3 kinase activity is higher during germ tubeformation compared with the yeast morph. Prior et al. (Prioret al., 1993; Prior et al., 1994) and Lakin-Thomas (Lakin-Thomas, 1993a; Lakin-Thomas, 1993b) identified the keycomponents of the phosphoinositide signal transduction in N.crassa, showing that the number of isomers of bis and trisinositol phosphates present in fungi is far more complex thanin animals and plants. They failed to demonstrate stimulationof phosphoinositide turnover by a variety of external stimuli.However, Kallies (Kallies et al., 1998) found that normal IP3concentration within growing hyphae of N. crassaincreasedtwo to fivefold to trigger Ca2+ release during the heat shockresponse. IP3 signaling may be involved in the regulation ofendogenous metabolism and differentiation since inhibitors ofphosphoinositide turnover led to lower extension rates andincreased hyphal branching (Hosking et al., 1995), includingLi+ inhibition of inositol metabolism and hyphal growth(Hanson, 1991). Besides fungal organisms, there is evidenceof IP3-mediated morphogenesis in tip-growing pollen tubes(Malho, 1998a; Kost et al., 1999).
Our evidence for the presence of an IP3 receptor in fungi wasbased upon an in vivo pharmacological signature: increase incytoplasmic Ca2+ caused by IP3 and depletion of tip-localized[Ca2+]cyt by 2-aminoethoxidiphenylborate (2-APB), whileother known inhibitors of IP3 receptors [8-(N,N-diethylamino)-octyl-3,4,5-trimethoxybenzoate, hydrochloride (TMB-8),dantrolene, heparin] had no effect on hyphal growth(Silverman-Gavrila and Lew, 2001). This inhibitor signaturewas used to assess ion channel activities from endomembranesand to establish the presence of an IP3 receptor required forhyphal growth. Such an IP3 receptor would be novel andunique to fungi as neither the N. crassa nor yeast genomes havesequences homologous to animal IP3 receptors based onBLAST searches. To demonstrate the presence of a functionalIP3 receptor in N. crassa, we used the bilayer lipid membrane(BLM) technique (Fig. 1B).
The BLM technique is an invaluable method for theinvestigation of the properties of ion channels fromintracellular membranes that are inaccessible to patch clampelectrodes. In this technique, channel-containing vesicles areincorporated into a bilayer membrane so that ion flux throughthe channel can be studied under voltage clamp conditions,permitting the measurement of the channel conductance. Theelectrical activity of the channel is measured under well-
controlled aqueous conditions. Thus, in solutions containingonly Ca2+ as a permeant ion, only Ca2+ permeant channels willbe observed. In addition to biophysical characterization of ionchannels, the technique can be used to determine the directeffect of agonists and antagonists on ion channel conductanceand kinetics. Thus, IP3 activation of Ca2+ channels can beobserved directly, as well as inhibitor effects. With thistechnique, we were able to demonstrate IP3-activated Ca2+
channels in N. crassa membranes.
Materials and Methods Germling preparationLarge scale conidial harvests were prepared by inoculatingEhrlenmeyer flasks containing 20 ml Voegel’s medium (Voegel, 1959)plus 2% agar and 2% sucrose with N. crassa RL 21a (FGSC no. 2219)wild-type conidia. The cultures were grown for 2 days at 30°C in thedark, then at room temperature in continuous light for at least 3 days.Conidia were harvested and resuspended in Voegel’s medium plus 2%sucrose at a final concentration of 106 conidia/ml. Conidia weregerminated by incubation at 37°C on a shaker rotating at 250 rpm. Tomaximize isolation of membranes from growing hyphal tips comparedwith subapical regions, the germlings were collected after 5-6 hoursof growth (germ tube length of 100 to 200 µm) by centrifugation at160 g for 10 minutes.
Microsomal membrane preparationGermlings were disrupted manually by vigorous grinding at 4°C in amortar and pestle using glass sand (150-212 µm) in ice-coldhomogenizing buffer (250 mM sucrose, 10 mM Na2EDTA, 5 mMMgSO4, 25 mM MES, 2.5 mM dithiothreitol, pH adjusted to 7 with1 N KOH). BSA (1% w/v) was added just prior to homogenization.The homogenate was decanted and left on ice for about 1 minute tosediment the sand and cell debris. The supernatant was centrifuged at40,000 g for 15-30 minutes to spin down the mitochondria, cell walls,nuclei and remove traces of sand using a Beckman SW-27 swingingbucket rotor in a Beckman ultracentrifuge or a Sorvall SS-34 fixedangle rotor in a Sorvall RC 5B plus centrifuge. Then the supernatantwas centrifuged in a SW-28 swinging bucket rotor using a BeckmanOptima ultracentrifuge for 1 hour at 80,000 g. The microsomal pelletwas resuspended in 200 µl resuspension medium: either 1 M sucrose,the homogenizing medium without BSA or in 40% (1.17 g ml–1)sucrose and 10 mM MOPS (pH 6.4). The resuspension (about 1 mg/mlprotein) was aliquoted and stored at –20°C. The aliquots (8 µl) weresufficient for 2-3 experiments; by using small aliquots we avoidedmembrane damage caused by freeze-thaw cycles. Proteinconcentration was determined colorimetrically using a modifiedBradford protein assay (Bradford, 1976) with a BSA standard.
Subcellular fractionation in discontinuous sucrose densitygradientsSpecific membrane fractions from N. crassa were isolated bysubcellular fractionation using density gradient centrifugation.Germlings were disrupted by grinding in homogenizing buffer (250mM sucrose, 10 mM EGTA, 10 mM MgSO4, 25 mM MES, 2.5 mMdithiotreitol pH 7.0 with KOH). The 80,000 g microsomal pellet wasresuspended in 40-200 µl resuspension buffer (250 mM sucrose, 2.5mM MES, 2.5 mM dithiotreitol adjusted to pH 7 with solid BTP) andlayered onto a discontinuous three step sucrose gradient [5 ml of 16%,29%, 39% (w/w) sucrose buffered to pH 7.2 with 5 mM MOPS/BTP]and centrifuged for 2 hours at 100,000 g at 4°C. This method wasdesigned to partition the microsomal fraction into ‘vacuolar’ (16%)(Vaughn and Davis, 1981), ER (16/29%) (Borgeson and Bowman,1983) and plasma membrane (29/39%) (Bowman et al., 1981)
Journal of Cell Science 115 (24)

5015IP3-activated channels and tip growth
interfaces. The actual vacuolar membrane density has been reportedto vary from 1.06 to 1.30 g cm–3(Vaughn and Davis, 1981), dependingon the details of homogenization and osmolarity. Because we usedyoung germlings, which should not have had time to accumulatevacuolar polyphosphates and arginine, and homogenization atrelatively low osmolarity (Vaughn and Davis, 1981), with EGTApresent (Bowman and Bowman, 1982), we expect the vacuoles to havea low density, similar to plant vacuoles. To assure minimalcontamination with the ER, we chose a cut-off density of 1.06 g cm–3
(16% w/w sucrose). The membranes from interfaces were collectedand aliquoted in small Eppendorf tubes. Membranes were fusedto planar lipid bilayers and channels assayed for IP3 activation,conductance and inhibitor effects.
Planar bilayer measurements of channel activityPlanar lipid bilayers were formed from a lipid mixture of 1-palmitoyl-2-linoleyl-sn-glycero-3-phosphatidylcholine:1-palmitoyl-2-linoleyl-sn-glycero-3-phosphatidylethanolamine:1-palmitoyl-2-linoleyl-sn-glycero-3-phosphatidyl-L serine (sodium salts):cholesterol (50:10:30:10 parts,w/w) (Avanti Polar Lipids, Alabaster, AL), stored in benzene underN2 at –20°C. The lipid mixture was chosen to ensure a lipidenvironment similar to the naturally occurring fatty acids,phospholipids and sterol:phospholipid ratio (Aaronson et al., 1982),except that cholesterol was used instead of ergosterol. Vesicles andreagents (IP3 for receptor activation, inhibitors) were added to the ciscompartment (Fig. 1B). The sign of the potential difference refers tothe trans compartment; a positive current corresponds to a cation fluxfrom trans to cis or an anion flux from cis to trans compartment andis shown as an upward deflection from zero current. Thus, a positive
current is positive charge flux from the lumenal side to thecytoplasmic side, the direction of Ca2+ flux that is physiologicallyrelevant.
Vesicle fusion with the bilayerAfter a stable bilayer membrane had formed, 4-32 µl microsomalvesicles (1 mg/ml protein) were added to the cis chamber. The cissolution (1 M KCl, 10 mM CaCl2, 200 mM MOPS/BTP, pH 7.2)was hyperosmotic to the trans solution {either 50 mM Ca(OH)2 [anactivity of 30 mM based on the activity coefficient for Ca(ClO4)2(Zaytsev and Aseyev, 1992)] or Ba(OH)2 or D-Gluconic acid (hemi-magnesium salt) and 200 mM MOPS/BTP, pH 7.2}, and thevesicles were hyperosmotic to the cis compartment solution toenhance the frequency of vesicle fusion. In other experiments weused 100 µM Ca(OH)2 and 200 mM MOPS/BTP (pH 7.2)(calculated activity 95.9 µM) in trans. The current was monitoredat various transmembrane holding potentials (±50 to 100 mV) forthe appearance of K+ and/or Cl– channel activity as indicator ofvesicle fusion to the bilayer.
Activation of the channel by IP3
Once channel incorporation into the bilayer membrane was detected,the cis solution was perfused away with 1 mM EGTA, 200 mMMOPS/BTP pH 7.2 to prevent further vesicle fusion and to leave Ca2+
on the trans side as the only ion present in the system. Free Ca2+ wascalculated based on a Turing program implementation of thealgorithm and binding constants from Goldstein (Goldstein, 1978). Inthe absence of any permeant ion except Ca2+, only Ca2+ channels can
A.
B.
Ca2+
3IP
fusionvesicle
Ca2+
stretch-activatedphospholipase C
calciumrelease
3IP receptor
Ca2+
sequestrationcalcium
ADP + Pi
+ATP
channelCa2+
Ca2+
IP3
trans cis
voltage clamp
fusion
-+
Fig. 1. (A) Internal generation and maintenance of the Ca2+ gradient ingrowing hyphae of N. crassa. This model is based upon extensivescreening of inhibitors of Ca2+ signaling and PLC effects on hyphalgrowth, morphology, cytoplasmic and vesicular Ca2+ gradients, as well asdirect evidence of IP3-activated Ca2+ channels using the BLM technique.Treatment with the Ca2+ ATPase inhibitor cyclopiazonic acid inhibitedgrowth (hyphal widening was observed) and increased cytoplasmic [Ca2+]behind the apex, consistent with Ca2+ sequestration into endoplasmicreticulum behind the growing apex (Silverman-Gavrila and Lew, 2001).Microinjection of IP3 receptor agonists (IP3 and Adenophostin A) behindthe tip inhibited growth transiently, caused subapical branching, andaffected the tip-high cytoplasmic Ca2+ gradient; these effects were notobserved after microinjection of the biologically inactive L-IP3(Silverman-Gavrila and Lew, 2001). IP3-induced subapical branching wassimilar to subapical branching induced by ionophoretic injection of Ca2+
(Silverman-Gavrila and Lew, 2000), suggesting that IP3-activated Ca2+
release occurs in growing hyphae. An inhibitor of the IP3 receptor, 2-APBinhibited hyphal elongation and dissipated the tip high cytoplasmic [Ca2+]gradient. Thus, tip-localized IP3 production due to a stretch-activatedphospholipase C could activate vesicular Ca2+ channels at the growingapex to generate the tip-high [Ca2+] gradient. The Ca2+ would inducefusion of wall vesicles at the apex, before being sequestered behind the tipvia the Ca2+ ATPase. (B) The BLM technique used to identify andmeasure IP3-activated Ca2 channel activity in membrane vesicles isolatedfrom N. crassa.Hyphae are first homogenized to release theendomembranes. Following a series of centrifugations, subcellularfractions of membranes are isolated. Vesicles are added to the cis-chamber and fused to the lipid bilayer formed across an aperture in aseptum that separates two chambers: cis and trans. The electronics areconfigured as a high gain current to voltage converter capable ofmeasuring picoAmpere currents through ion channels. With only onepermeant ion, Ca2+, only Ca2+ channels will be observed. Agonistaddition to the cis compartment would activate channels oriented withtheir ligand binding site facing the cis compartment.
5016
be observed. We did not observe spontaneous Ca2+ channel openings.Ca2+ channels were observed only after the addition of IP3, Na2ATPand CaCl2 to the cis compartment sequentially or together to attain afinal concentration of 16 µM IP3, 100 µM Na2ATP and 1 mM CaCl2.
After IP3 activation of the Ca2+ channels, the effects of theinhibitors heparin, 2-APB, TMB-8 and dantrolene (Calbiochem, LaJolla, CA) on channel activity were examined.
Channel conductanceCurrent-voltage measurements were made with a BC-525 C bilayerclamp amplifier (Warner Instruments Corp); holding potentials rangedfrom –100 to +100. Channel currents were filtered at 1 KHz (cornerfrequency) with an 8 pole low-pass Bessel filter and recorded with amodified (pulse code modulation) VCR (DAS/VCR 900; Dagan Corp,Minneapolis, MN). All recordings were carried out at room temperature.Data acquisition and analysis were performed using AxoScopepCLAMP software version 1.1 (Axon Instruments, Burlingame, CA).Recording playbacks were filtered at 0.5 kHz with a 4-pole Bessel filter(LPF 100B, Warner Instruments) and digitized at 1 kHz. For computeranalysis, the recorded voltage clamp signals were not filtered prior torecording and digitization. Current amplitudes were measured manuallyusing cursors, choosing clear channel opening/closing events. Theconductance and the reversal potential were determined from the slopeand X-intercept of the current-voltage curve.
PLC inhibitors: treatments and growth measurementsN. crassawas grown in 35 mm tissue culture dishes on solid substrate
Journal of Cell Science 115 (24)
Aa
b
c
d
e
closed
2 pA
2 sec
Fusion
Wash-ou t
D-IP3
Wash-ou t
L-IP3
B
b
c
d
a Before
IP3
TMB-8
2-APB
2 pA20 sec
Table 1. Relative abundance of small and largeconductance channels in microsomal membrane isolations
Total experiments 138Successful fusions 101/138 (73.2%)IP3 activation 38/101 (37.6%)Small conductance 26 experiments (11.9±5.1 pS, n=16)
68.4% of total IP3-activated channelsLarge conductance: 12 experiments (82.9±69.7 pS, n=5)
31.6% of total IP3-activated channels
Some experiments could be scored as small or large conductance basedupon amplitude measurements at a single positive voltage. Linear regressionto calculate conductance values was possible in a subset of experiments.
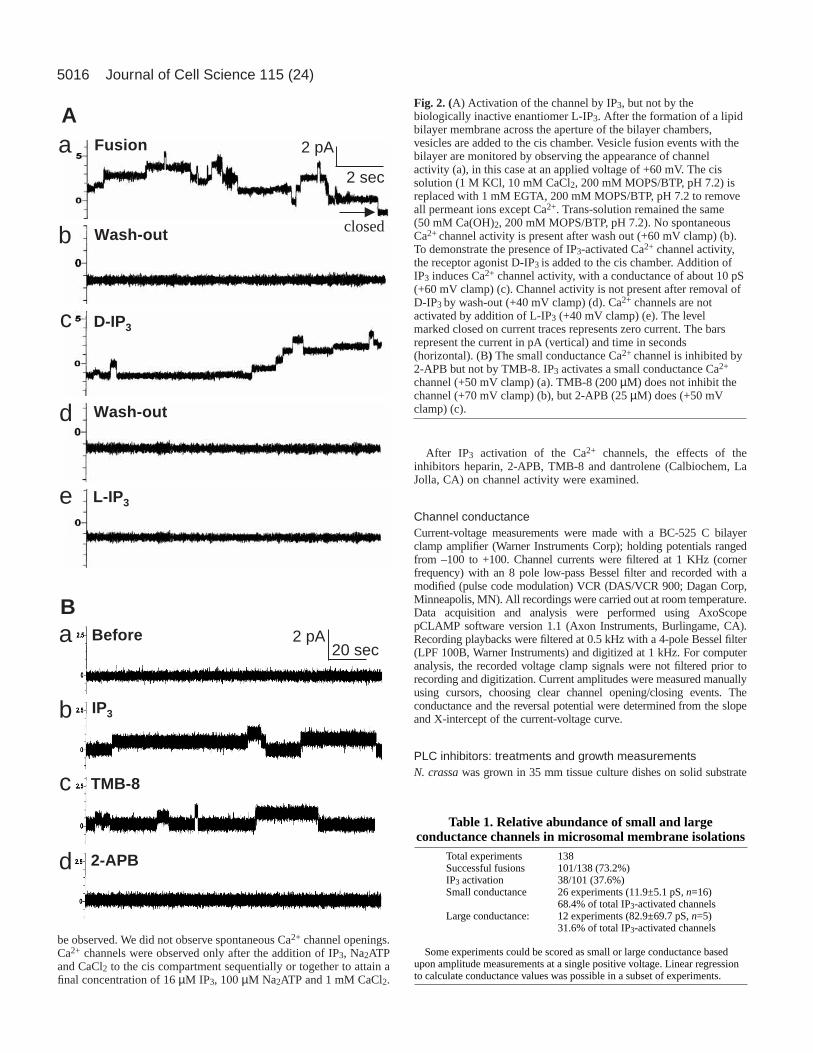
Fig. 2. (A) Activation of the channel by IP3, but not by thebiologically inactive enantiomer L-IP3. After the formation of a lipidbilayer membrane across the aperture of the bilayer chambers,vesicles are added to the cis chamber. Vesicle fusion events with thebilayer are monitored by observing the appearance of channelactivity (a), in this case at an applied voltage of +60 mV. The cissolution (1 M KCl, 10 mM CaCl2, 200 mM MOPS/BTP, pH 7.2) isreplaced with 1 mM EGTA, 200 mM MOPS/BTP, pH 7.2 to removeall permeant ions except Ca2+. Trans-solution remained the same(50 mM Ca(OH)2, 200 mM MOPS/BTP, pH 7.2). No spontaneousCa2+ channel activity is present after wash out (+60 mV clamp) (b).To demonstrate the presence of IP3-activated Ca2+ channel activity,the receptor agonist D-IP3 is added to the cis chamber. Addition ofIP3 induces Ca2+ channel activity, with a conductance of about 10 pS(+60 mV clamp) (c). Channel activity is not present after removal ofD-IP3 by wash-out (+40 mV clamp) (d). Ca2+ channels are notactivated by addition of L-IP3 (+40 mV clamp) (e). The levelmarked closed on current traces represents zero current. The barsrepresent the current in pA (vertical) and time in seconds(horizontal). (B) The small conductance Ca2+ channel is inhibited by2-APB but not by TMB-8. IP3 activates a small conductance Ca2+
channel (+50 mV clamp) (a). TMB-8 (200 µM) does not inhibit thechannel (+70 mV clamp) (b), but 2-APB (25 µM) does (+50 mVclamp) (c).

5017IP3-activated channels and tip growth
(2% w/v gellan gum) containing 2% sucrose and Voegel‘s minimalmedium. After incubation at 28-30°C for 14 hours, the culture wasflooded with buffer solution (BS) [10 mM KCl, 1 mM CaCl2, 1 mMMgCl2, 10 mM PIPES (pH adjusted to 5.8 with KOH) and the osmolalityadjusted to 260 mosmol/kg with sucrose] (Levina et al., 1995).
Individual growth rates were measured every minute for 10 minutesin BS with a Zeiss Axioskop microscope and a 60× water immersionobjective. Then, PLC inhibitors {neomycin, 3-nitrocoumarin, 1-[6-((17β-3-methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-1H-pyrrole-2,5-dione (U-73122)} or, as a control, the biologically inactiveanalog 1-[6-((17β-3-methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-pyrrolidine-2,5-dione (U-73343) were perfused through the chamber (2minutes were required for solution exchange) and growth ratesmeasured for 10 minutes, followed by wash out and measurement ofgrowth rates in BS alone for another 10 minutes. For inhibitorsthat required DMSO solubilization (U-73122, U-73343 and 3-nitrocoumarin) pre-and post-treatments in BS included DMSO at thefinal concentration used during the treatment. Growth rates in BS plusDMSO were very similar to growth rates in BS alone. The number ofexperiments for any given treatment ranged from 7 to 9. Whenmorphological changes at the growing tip were observed during or aftertreatment, other hyphae from the plate were also examined to confirmthat the effect was general.
Chlortetracycline imagingIn preliminary experiments we determined the optimal concentration ofchlortetracycline (CTC) (Sigma), which did not affect hyphal growthbut still provided good fluorescence signal staining. Similar growth rateswere found for control untreated hyphae and hyphae-treated with CTCat 25-50 µM concentration range. After CTC addition, hyphal growthand organellar Ca2+ fluorescence were measured, and then the hyphaewere treated with 2-APB, cyclopiazonic acid, neomycin or U-73122(Calbiochem, La Jolla, CA) and growth and fluorescence measured. Thefluorescence was detected using the same settings before and afterinhibitor treatments. Both confocal (krypton-argon laser Bio Rad MRC,Nikon Optiphot microscope, 40× water immersion lens, BHS, filterpower NDF=0, excitation at 488, emission <515 nm, direct and Kalmanfiltering) and conventional fluorescence (Zeiss 60× water immersionobjective, FITC filter cube, Hammamatsu C4742-95 digital camera)microscopy were used and gave similar results.
ResultsInhibitor effects on growth and the Ca2+ gradient implicatedIP3-activated Ca2+ release from internal stores as the generatorof the gradient. IP3 microinjection inhibits growth transientlyand modifies the tip-high Ca2+ gradient in growing fungalhyphae. It does not affect the trans-plasma membrane electricalpotential (data not shown). Thus the putative IP3 receptor wasexpected to be localized to endomembranes, so we usedcellular membrane fractionations and the BLM technique (Fig.1B) to identify directly IP3-activated Ca2+ channels.
IP3 activates small and large conductance Ca2+
channels in Neurospora crassaTo obtain tip-enriched membranes we used conidial germlings.From a starting material of 30 g germlings, we obtained about0.24 mg total membrane protein. The appearance of channelactivity was used to confirm successful fusion of membranevesicles to the planar bilayer (Fig. 2A). In initial experimentswith total membranes, successful fusions were observed in101/138 trials (73.2%) (Table 1). Following the incorporation ofvesicles into the planar lipid bilayer, the KCl-containing cis
solution was replaced with EGTA and MOPS/BTP so that transCa2+ was the only channel-permeant ion. No spontaneous Ca2+
channel activity was observed, with very rare exceptions (4 outof 79), which could be due to incomplete wash-out of the KCl.Subsequently, D-IP3, ATP and CaCl2 were added to the ciscompartment and activation of the receptor was observed in 38out of 101 successful fusions (37.6%) (Table 1). As a control,the biologically inactive IP3 enantiomer, L-IP3 did not activatethe channel (n=2) (Fig. 2A). Other control experiments were
A
B
a
b IP3
c Heparin
Befo re
1 pA 1 sec
b IP3
c Dantrolene
a 5 sec1 pABefo re
Fig. 3. (A) Heparin does not inhibit the small conductance channel.After vesicle fusion, no spontaneous Ca2+ channels are observed inthe absence of IP3 (40 mV clamp) (a). D-IP3 activates Ca2+ channels(12 pS) (+40 mV clamp) (b). The competitive IP3 channel blockerheparin (50 µM) has no effect on channel activity even after a secondtreatment (40 mV clamp) (c).(B) Dantrolene does not inhibit thesmall conductance channel. Prior to IP3 addition, no spontaneousCa2+ channels are present (+60 mV clamp) (a). IP3 opens a smallconductance channel (11 pS) (+100 mV clamp) (b). Dantrolene doesnot inhibit the channel (+100 mV clamp) (c).

5018
performed by adding Ca2+ and/or ATP before IP3; in the absenceof IP3 they did not activate the channel (n=2, data not shown).ATP was usually added along with IP3 because it is required forfull activation (increased open probability, unchangedconductance) of the animal IP3 receptor (Bezprozvanny andEhrlich, 1993). To assess the requirement for ATP for channelactivity, IP3-induced Ca2+ release was studied in the absence andpresence of varying [ATP]. ATP is not required for channelactivation, nor does subsequent ATP addition increasesignificantly channel activity (n=4, data not shown).
In asymmetric cis/trans solutions with 50 mM Ca(OH)2 (30mM activity) and 200 mM MOPS/BTP pH 7.2 in the transcompartment, and 1 mM [Ca2+] in the cis compartment(calculated Ca2+ activity of 7.1 µM), IP3 activated two channelswith mean conductances of 11.9±5.1 pS (n=16) and 82.9±69.7pS (n=5) measured at potentials clamped between –100 and+100 mV (Table 1).
Inhibitor characterization of the small and largeconductance channels and correlation with hyphalgrowthHyphal growth and cytosolic Ca2+ gradients are affected by 2-
APB, but not by heparin, TMB-8 or dantrolene. This inhibitorsignature was used in preliminary characterization of the IP3-activated Ca2+ channels. The small conductance channel wasinhibited by 2-APB (25 µM) (Fig. 2B), but was unaffected by200 µM TMB-8 (Fig. 2B), 50 µM heparin and 100 µMdantrolene (Fig. 3A,B). After 2-APB inhibition, addition of IP3(16 µM) and ATP (0.1 mM) did not restore channel activity(data not shown). The large conductance channel was inhibitedby 2-APB (data not shown), TMB-8 (Fig. 4A), heparin (Fig.4B) and dantrolene (Fig. 4C). The latter three inhibitors do notaffect hyphal growth or inhibit the small conductance channel.
Journal of Cell Science 115 (24)
Table 2. Effects of Ca2+ regulators on IP3-activated Ca2+
channels, hyphal growth and morphology in N. crassaSmall conductance Large conductance Growth and
Ca2+ regulator channel activity channel activity morphology
2-APB Inhibition (n=7) Inhibition (n=3) Inhibition*Heparin No inhibition (n=4) Inhibition (n=5) No effectTMB-8 No inhibition (n=3) Inhibition (n=3) No effectDantrolene No inhibition (n=2) Inhibition (n=2) No effect
*Hyphal widening at low concentration, apical hyperbranching upon washout.
A B
C
a
b
c
d
2 pA
5 sec
Fusion
Wash-ou t
IP3
TMB-8
b
1 pA2 seca IP3
Heparin
a
b
IP3
Dantrolene
0.1 sec20 pA
Fig. 4. (A) TMB-8 inhibits the IP3-activated large conductance Ca2+ channel. Channel activity signals vesicle fusion (+100 mV clamp) (a). Nospontaneous Ca2+ channels are observed after wash out (+45 mV clamp) (b). IP3 activates a large conductance channel (41 pS) (+70 mV clamp)(c). TMB-8 (200 µM) inhibits channel activity (+60 mV clamp) (d). (B) Heparin inhibits the large conductance channel. Representative channelcurrent recordings show Ca2+ channel activation in response to the application of IP3 (+50 mV clamp) (a). The channels are inhibited byheparin (50 mV clamp) (b). (C) Dantrolene inhibits completely the large conductance channel. IP3-activated Ca2+ channel (131 pS) (+80 mVclamp) (a). Inhibition by 100 µM dantrolene (+80 mV clamp) (b).

5019IP3-activated channels and tip growth
Thus, the small conductance channel is implicated as thegenerator of the tip-high Ca2+ gradient required during hyphalgrowth (Table 2).
Ionic selectivity of IP3 channelsCationic selectivity was determined for Ba2+, Ca2+ and Mg2+
using Ba(OH)2, Ca(OH)2 and Mg gluconate, respectively, in thetrans compartment. For the small conductance IP3 channelsimilar conductances were observed with either 50 mM Ba(OH)2or Ca(OH)2, but the channel was not observed with Mggluconate (n=3) (Fig. 5). The large channel transports Ba2+ andCa2+, and Mg2+ but with a smaller channel amplitude (n=2, notshown). Using constant 50 mM [Ca2+] in trans, the channelconductance was determined at different [Ca2+] in cis. Theconductance decreased slightly with increased [Ca2+]. A cis[Ca2+] of 40 mM does not inhibit the channel (data not shown).
Reversal potentials and channel conductancemeasurements at defined Ca2+ activitiesIn initial experiments we chose trans and cis [Ca2+] to maximizeour ability to observe measurable currents through IP3-activatedCa2+ channels that release Ca2+ from the lumenal side of fusedvesicles (the trans compartment). The concentration in transwas 50 mM (calculated activity 30 mM). With cis Ca2+ activityat 7.09 µM, the Nernst potential for Ca2+ was –105 mV. Underthese conditions, the reversal potentials for channels weredifficult to measure because the membrane was not stable atnegative voltages, and negative channel current amplitudescould not be measured reliably. To assess ion-selectivity of thechannels, the [Ca2+] in the trans compartment was decreased to100 µM (calculated Ca2+ activity of 96 µM) to bring theexpected reversal potential to less negative values and to studychannel activity under more physiological conditions. In N.crassa, Ca2+-containing vesicles are estimated to have aconcentration of at least 26 µM (Torralba et al., 2001) and thetip-localized cytoplasmic Ca2+ activity was found to be in thenanomolar range (400-600 nM) (Silverman-Gavrila and Lew,2000; Silverman-Gavrila and Lew, 2001). We used a higher cisCa2+ activity of 7.09 µM. With Ca2+ as the only charge carrier,the average conductances (small channel: 15±3 pS (n=8); largechannel: 153.5±90.7 (n=6) were similar to the conductancesobserved with cis and trans Ca2+ activities of 7.09 µM and 30mM (13.4±5.6 pS (n=24) and 77.0±61.1 pS (n=8)), respectively(Fig. 6, Table 3). The theoretical reversal potential for Ca2+ was
calculated to be –33 mV and for Cl– ~–200 mV. The observedreversal potential was –26.30±14.18 (n=8) for the smallconductance channel and –18.59±5.72 (n=6) for the largeconductance channel. As the reversal potential was close to theNernst potential for Ca2+, we can consider the channels to beCa2+ selective with no Cl– permeability. Voltage-dependentactivation was not observed at cis and trans Ca2+ activities of7.09 µM and 96 µM, respectively.
The small conductance channel correlates with ER andplasma membrane, the large conductance with vacuolarmembrane in subcellular gradient fractionationsTo identify possible origins of the IP3-activated Ca2+ channels,we fractionated microsomal endomembranes on sucrosedensity step gradients (Table 4). The mean conductance for thesmall channel was 15.11± 6.93 pS (n=6) and the largeconductance channel 67.12±35.66 pS (n=3). Relativeabundance of channels were 66:34% (27:14) small:largeconductance channel, similar to the results obtained usingmicrosomal endomembranes. Since Ca2+ containing vesiclesdestined for fusion at the tip are produced in the ER, and causeplasma membrane ‘expansion’, we expect enrichment in thesetwo fractions. Indeed, the small conductance channel isenriched in the plasma membrane (78%) and ER fractions(80%). The high conductance channel is enriched in thevacuolar membrane fraction (88%).
Table 3. Reversal potential and conductance of small andlarge conductance channels with low Ca2+ in trans
chamberTotal experiments 36Successful fusions 29/36 (80%)IP3 activation 14/29 (48.27%)Small channel
Conductance 15.0±3.1 pS (n=8)Reversal potential –26.3±14.2 mV (n=8)
Large channelConductance 153.5±90.7 pS (n=6)Reversal potential –18.6±5.8 mV (n=6)
Calculated Nernst potential for Ca2+ was –33 mV.
a
b
c
d
Before
IP3+ Mg2+
2-APB
IP3+ Ca2+
2 pA
2 sec
Fig. 5. The small conductance channel conducts Ca2+, but not Mg2+.After vesicle fusion, no spontaneous Ca2+ channels are present (+60mV clamp) (a). After IP3 addition, the small conductance channeldoes not conduct Mg2+ (+60 mV clamp) (b), but does conduct Ca2+
(12 pS) (+60 mV clamp) (c). 2-APB inhibits completely the channel(+50 mV clamp) (d).

5020
Inhibitors of the IP3-activated Ca2+ channel and PLCaffect similarly growth rates and Ca2+ containing vesicledistributionIf vesicular IP3-activated Ca2+ channels generate the tip-highCa2+ gradient required for tip growth, IP3 production catalyzed
by PLC must be localized at the hyphal tip. Three putative PI-specific PLC genes have been identified in N. crassa(Jung etal., 1997). Initially, we screened PLC inhibitors for their effecton growth rates and morphology. Neomycin inhibits growthcompletely at 400 µM and slightly at 100 µM. Growth rates
Journal of Cell Science 115 (24)
Fig. 6. Current voltage curves of IP3-activated Ca2+
channels in N. crassamembranes. Current-voltagemeasurements reveal two distinct Ca2+channels: asmall conductance channel (circles) and a largeconductance channel (triangles). The current isrecorded at different membrane potentials, then thevoltage dependence of channel current is used toobtain current-voltage relations. Conductance andreversal potential are determined by linear regression.The calculated Nernst potential for Ca2+ is –33 mVfor 100 µM Ca(OH)2 (calculated activity of 96 µM)in trans and 1 mM CaCl2 (calculated activity 7.09µM) in the cis compartment. Note the difference inscale for small (left) and large (right) conductancechannels. In this example, the channel conductancedetermined from the slope of current-voltages curveswas 13 pS for the small channel and 111 pS for thelarge channel. Amplitude versus voltage was linearand reversed near –25 mV for the small channel and–16 mV for the large channel. Compiled data arepresented in Table 3.
Fig. 7. Effects of PLC inhibitors on N. crassagrowth rates. (A) Neomycin treatment. (Upper panel) Control experiments for treatment withexternal added inhibitors show that solution exchange (BS was exchanged with BS) does not affect hyphal growth. Middle and lower panelsillustrate growth rates of hyphae treated with neomycin (as marked). Neomycin inhibits hyphal growth, partially at 100 µM and completely at400 µM. (B) 3-nitrocoumarin causes a dose dependent inhibition of hyphal elongation: complete at 40 µg/ml, very efficient at 20 µg/ml andslight at 4 µg/ml. (C) Growth inhibition with increasing concentration of U-73122. (Upper panel) Control experiments. No influence on growthrates is observed for hyphae treated with the inactive analog U-73343. (Middle and lower panel) U-73122 inhibits growth: efficiently at 10 µMand completely at 400 µM within 4 minutes after its addition. Thin lines show individual experiments. Thicker lines and symbols showmeans±s.e.
5021IP3-activated channels and tip growth
are unaffected in control experiments with buffer solution (BS)(Fig. 7A). A known inhibitor of yeast phosphatidylinositol-specific PLC, 3-nitrocoumarin, inhibited growth (Fig. 7B, Fig.8C) with a concentration dependence very similar to in vitroinhibition of PLC (half-maximal inhibition at 10µg/ml) (Tisiet al., 2001). After washing out the inhibitor 10 minutes aftertreatment, hyphae recovered and restarted growth, showing that
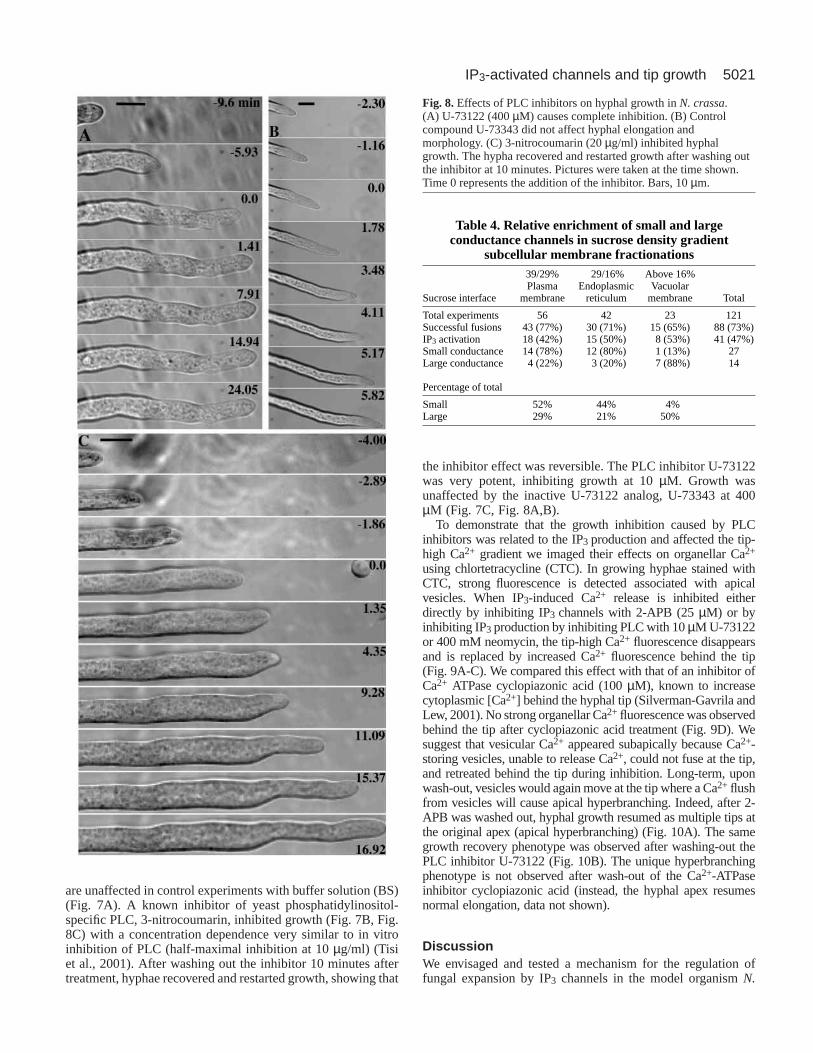
the inhibitor effect was reversible. The PLC inhibitor U-73122was very potent, inhibiting growth at 10 µM. Growth wasunaffected by the inactive U-73122 analog, U-73343 at 400µM (Fig. 7C, Fig. 8A,B).
To demonstrate that the growth inhibition caused by PLCinhibitors was related to the IP3 production and affected the tip-high Ca2+ gradient we imaged their effects on organellar Ca2+
using chlortetracycline (CTC). In growing hyphae stained withCTC, strong fluorescence is detected associated with apicalvesicles. When IP3-induced Ca2+ release is inhibited eitherdirectly by inhibiting IP3 channels with 2-APB (25 µM) or byinhibiting IP3 production by inhibiting PLC with 10 µM U-73122or 400 mM neomycin, the tip-high Ca2+ fluorescence disappearsand is replaced by increased Ca2+ fluorescence behind the tip(Fig. 9A-C). We compared this effect with that of an inhibitor ofCa2+ ATPase cyclopiazonic acid (100 µM), known to increasecytoplasmic [Ca2+] behind the hyphal tip (Silverman-Gavrila andLew, 2001). No strong organellar Ca2+fluorescence was observedbehind the tip after cyclopiazonic acid treatment (Fig. 9D). Wesuggest that vesicular Ca2+ appeared subapically because Ca2+-storing vesicles, unable to release Ca2+, could not fuse at the tip,and retreated behind the tip during inhibition. Long-term, uponwash-out, vesicles would again move at the tip where a Ca2+ flushfrom vesicles will cause apical hyperbranching. Indeed, after 2-APB was washed out, hyphal growth resumed as multiple tips atthe original apex (apical hyperbranching) (Fig. 10A). The samegrowth recovery phenotype was observed after washing-out thePLC inhibitor U-73122 (Fig. 10B). The unique hyperbranchingphenotype is not observed after wash-out of the Ca2+-ATPaseinhibitor cyclopiazonic acid (instead, the hyphal apex resumesnormal elongation, data not shown).
DiscussionWe envisaged and tested a mechanism for the regulation offungal expansion by IP3 channels in the model organism N.
Fig. 8.Effects of PLC inhibitors on hyphal growth in N. crassa.(A) U-73122 (400 µM) causes complete inhibition. (B) Controlcompound U-73343 did not affect hyphal elongation andmorphology. (C) 3-nitrocoumarin (20 µg/ml) inhibited hyphalgrowth. The hypha recovered and restarted growth after washing outthe inhibitor at 10 minutes. Pictures were taken at the time shown.Time 0 represents the addition of the inhibitor. Bars, 10 µm.
Table 4. Relative enrichment of small and largeconductance channels in sucrose density gradient
subcellular membrane fractionations39/29% 29/16% Above 16%Plasma Endoplasmic Vacuolar
Sucrose interface membrane reticulum membrane Total
Total experiments 56 42 23 121Successful fusions 43 (77%) 30 (71%) 15 (65%) 88 (73%)IP3 activation 18 (42%) 15 (50%) 8 (53%) 41 (47%)Small conductance 14 (78%) 12 (80%) 1 (13%) 27Large conductance 4 (22%) 3 (20%) 7 (88%) 14
Percentage of total
Small 52% 44% 4%Large 29% 21% 50%

5022
crassa (Fig. 1A). To our knowledge, this is the first directdemonstration of IP3-activated Ca2+ channels in fungi,including evidence that one channel type is involved in tipgrowth during hyphal expansion.
It may be significant that we did not observe spontaneousCa2+ channel activity prior to IP3 addition. In preliminaryexperiments demonstrating the presence of a folic acid-activated Ca2+ channel of about 100 pS conductance inDictyostelium discoideum, we did observe a spontaneous 12 pSCa2+ channel prior to folate addition. Whether or notspontaneous Ca2+ channels are absent completely in fungi mustawait further analysis.
In the ascomycete N. crassa, we identified two types of IP3-activated Ca2+ channels that could be differentiated by theirconductance: one small and one large. Reversal potentialmeasurements confirm that both are Ca2+ selective. In additionto different conductances, ion permeability is also different, inthat the small channel is impermeable to Mg2+.
We had already discovered a well defined inhibitorsignature for hyphal growth, in which only a subset of knowninhibitors of IP3 receptors affected growth and the cytoplasmictip-high Ca2+ gradient required for growth (Silverman-Gavrilaand Lew, 2001). We used this inhibitor signature to identifythe channel that functions in tip growth. Inhibitors that inhibitonly the large conductance channel do not affect tip-growth;for example, heparin microinjection, dantrolene, or TMB-8
treatments (Silverman-Gavrila and Lew, 2001). Thus, thelarge conductance IP3-activated Ca2+ channel does not playa role in hyphal growth. In contrast, 2-APB inhibitscompletely both the small and large conductance channel andhyphal growth and dissipates the tip-high Ca2+ gradient(Silverman-Gavrila and Lew, 2001). Both dissipation of thecytoplasmic calcium gradient and inhibition of hyphalgrowth occur within 1-2 minutes. The inhibition of growth,gradient dissipation and channel inhibition all occur atsimilar 2-APB concentrations (10-25 µM). In someorganisms, the mode of action of 2-APB may not necessarilyinvolve inhibition of IP3-activated Ca2+ channels. In animalcells, it is reported to inhibit Ca2+ entry (Kukkonen et al.,2001; Diver et al., 2001). In higher plants, capacitative Ca2+
entry has been postulated to play a role in pollen tube growth(Malho, 1998b). However, in the fungus N. crassa it isknown that Ca2+ entry does not play a role in generation ofthe tip-high Ca2+ gradient. There are no tip-localized inwardCa2+ currents (Lew, 1999), modifying the electrical drivingforce for Ca2+ entry by voltage clamp has no effect on hyphalgrowth (Silverman-Gavrila and Lew, 2001), and IP3microinjection does not affect the plasma membraneelectrical potential, an expected consequence of Ca2+ entry
(L.B.S.-G. and R.R.L., unpublished). We cannot discountcompletely the possibility of multiple sites of action by 2-APB. However, the direct demonstration of 2-APB inhibitionof the IP3-activated Ca2+ channel, correlated with 2-APBinhibition of growth and dissipation of the tip-high Ca2+
gradient, suggest that the small conductance channel generatesthe tip-high Ca2+ gradient during growth. We propose that thesmall conductance IP3 –activated Ca2+ channel originates inthe ER/Golgi body system. If the vesicles fuse with the plasmamembrane at the apex the channel must be inactivated becauseIP3 microinjection has no effect on the plasma membranepotential.
The difference in inhibitor signatures indicates that thelarge conductance channel is localized in vacuoles as TMB-8, dantrolene and heparin are widely reported to inhibit theIP3-induced Ca2+ release from vacuoles in S. cerevisiae[TMB-8 and dantrolene (Belde et al., 1993)], in N. crassa[dantrolene (Cornelius et al., 1989)], in red beet root [heparinand TMB-8 (Brosnan and Sanders, 1990; Alexandre et al.,1990)], and C. albicans[partial inhibition by heparin (Calvertand Sanders, 1995)]. The localization of the largeconductance channel on step gradients confirms its origin asvacuolar. The vacuolar IP3-activated Ca2+ channel may beimportant in signaling during environmental stress. Heatshock causes rapid increases in intracellular second
Journal of Cell Science 115 (24)
Fig. 9. Effects of Ca2+ and PLC inhibitors on organellar Ca2+
imaged with chlortetracycline. In normal growing hyphaesignificant fluorescence is detected associated with vesicles at thetip. (A) During Ca2+ imaging typical response (reduction offluorescence at the tip, and an increase behind) is observed after theaddition of 25 µM 2-APB at time 0 (n=18). At the same timehyphal elongation stops. (B) Similar results are obtained aftertreatment with 10 µM U-73122 (n=15). (C) Following the treatmentwith 400 µM neomycin similar effects occur (n=9). (D) In hyphaetreated with cyclopiazonic acid the tip high Ca2+ is dissipated, butno increase in fluorescence behind the tip is observed (n=15). Bars,10 µm.

5023IP3-activated channels and tip growth
messengers, cAMP and inositol phosphates (fivefold), as wellas Ca2+ release from isolated N. crassa vacuoles (Kallies etal., 1998). In addition, vacuoles may regulate the homeostasisof cytosolic Ca2+ providing a storage reserve or detoxificationsystem to prevent the effects of excessive Ca2+ in thecytoplasm. Cornelius and Nakashima isolated mutants withlow rates of vacuolar Ca2+ uptake that were inhibited bynormal [Ca2+] in medium (Cornelius and Nakashima, 1987).The vacuolar origin of this channel precludes a role in tipgrowth, since vacuoles are absent in the first 100 µm behindthe hyphal apex (Zalokar, 1959).
Because there is no indication of sequences homologous tothe multigenic IP3 receptor families in animals, we do notexpect close relatedness to animal IP3 receptors. However, thechannel does share similarities with type 3 IP3 receptorscompared with types 1 and 2. Type 1 and 2 receptors arestimulated by low [Ca2+] and inhibited by high [Ca2+], we didnot observe Ca2+ inhibition, similar to type 3 receptors(Thrower et al., 2001). Ca2+ release by type 3 is much lessaffected by ATP compared with type 1 (Thrower et al., 2001),similar to our observations. These observations are based onBLM measurements of channel activity, and may not hold truein the complex cytoplasmic milieu, in which other regulatoryproteins may affect channel activity.
Since generation of the cytosolic tip-high Ca2+ gradientoccurs at the growing apex, IP3 must be produced apically. PLCis the upstream step in IP3 signaling that catalyzes theconversion of phosphatidylinositol (4,5)-bisphosphate to IP3and DAG. PLC activity is reported to be higher in hyphal versusyeast forms of C. albicans(Bennett et al., 1998). To confirm anapex-localized role of PLC, we examined the effects of PLCinhibitors on growth and organellar Ca2+. All inhibitors weexamined inhibited growth followed by hyphal widening. It isnotable that the half maximal inhibition of PLC activity by 3-nitrocoumarin, an efficient inhibitor of yeast PLC (Tisi et al.,
2001), was similar to its inhibition of growth in vivo in N.crassa, strongly suggesting that PLC is essential for growth.Although any one of the inhibitors could inhibit otherprocesses besides PLC, the fact that all affect hyphal growthimplicates PLC activity as a central component of hyphalgrowth, producing IP3 at the apex. We then examinedwhether the PLC inhibitors affect organellar Ca2+.
The vesicular Ca2+ gradient has been visualized by CTCfluorescence in growing tips of pollen tubes (Reiss et al.,1985), the oomycetes S. ferax(Yuan and Heath, 1991) andN. crassa(Schmid and Harold, 1988). In N. crassa the CTCgradient is very similar to the steep gradient of apicalvesicles (Collinge and Trinci, 1974) (L.B.S.-G. and R.R.L.,unpublished) suggesting that vesicular Ca2+ originates fromvesicles at the growing apex. Because 2-APB inhibits IP3-activated Ca2+ channels, and thus Ca2+ release fromvesicles, and dissipates the cytosolic [Ca2+] gradient at thetip, we expected increased vesicular Ca2+ fluorescence atthe tip. Instead vesicular Ca2+ appeared sub-apically. It ispossible that Ca2+-containing vesicles, formed via theER/Golgi body system, were unable to release Ca2+, couldnot fuse at the tip, and retreated subapically. Upon inhibitorwash out, they would again move forward, causing anapical hyperbranching phenotype. We observed the samesubapical retreat during PLC inhibitor treatment andsubsequent apical hyperbranching after wash out. As acontrol, cyclopiazonic acid, an inhibitor of ER Ca2+
ATPase, inhibits growth and increases basal cytoplasmic[Ca2+]. The loss of vesicular Ca2+ fluorescence in hyphaetreated with cyclopiazonic acid is consistent with decreasedCa2+ sequestration behind the apex. Cyclopiazonic acid hasalso been reported to decrease vesicular Ca2+ at hyphal apicesbased on pyroantimoniate-staining of a vesicle sub-populationin fixed tissues examined by electron transmission microscopy(Torralba et al., 2001).
In conclusion the present paper reports the first identificationand characterization of IP3-activated Ca2+ channels in a lowereukaryote. One of these channels is implicated strongly as partof the mechanism responsible for the generation of a tip-highCa2+ gradient required for fungal tip growth. Our present andevolving hypothesis is shown in Fig. 1A. Tip-localized PLC isactivated, possibly by stretch (Kinnunen, 2000) and producesIP3. The IP3 activates Ca2+ channels from a subset of tip-localized Ca2+-containing vesicles that release Ca2+ into apicalcytoplasm. The Ca2+ is required for fusion of wall vesicles withplasma membrane to cause tip expansion. The Ca2+ containingvesicles function in a ‘kiss and run’ fashion, shuttling Ca2+
from ER to the growing apex. They accumulate Ca2+ througha Ca2+ ATPase. A ‘kiss and run’ shuttle is consistent with thefact that Ca2+ efflux is not observed at growing tips (Lew,1999).
This work was supported by a NSERC grant to R.R.L. and anOGSST scholarship to L.B.S-G. Special thanks to Enzo Martegani(Milano University, Italy) for the generous gift of 3-nitrocoumarin.
ReferencesAaronson, L. R., Johnston, A. M. and Martin, C. E. (1982). The effects of
temperature acclimation on membrane sterols and phospholipids ofNeurospora crassa. Biochim. Biophys. Acta. 713, 456-462.
Alexandre, J., Lassalles, J. P. and Kado, R. T. (1990). Opening of Ca2+
Fig. 10.Hyperbranching after washing out inhibitors of IP3 receptor andPLC. (A) Within 20 minutes of washing out 25 µM 2-APB, hyphalwidening, which extended over 20-40 µm from the tip, and an apicalhyperbranching phenotype was observed. (B) U-73122 (10 µM) wash-outalso caused hyphal widening and multiple apical branches. The picturewas taken 14 minutes after wash out. (C) Control hypha 16 minutes afterwash out. Hyperbranching was not observed after wash-out ofcyclopiazonic acid (not shown); in this case, hypha resumed normalhyphal elongation, similar to C. Bars, 10 µm.

5024
channels in isolated red beet root vacuole membrane by inositol 1,4,5-triphosphate. Nature343, 567-570.
Allen, G. J., Muir, S. R. and Sanders, D. (1995). Release of Ca2+ fromindividual plant vacuoles by both InsP3 and cyclicADP-ribose. Science268,735-737.
Belde, P. J. M., Vossen, J. H., Borst-Pauwels, G. W. J. and Theuvenet,A. P. R. (1993). Inositol 1,4,5-triphosphate releases Ca2+ from vacuolarmembrane vesicles ofSaccharomyces cerevisiae. FEBS Lett. 323,113-118.
Bennett, D. E., McCreary, C. E. and Coleman, D. C. (1998). Geneticcharacterization of phospholipase C gene from Candida albicans: presenceof homologous sequences in Candida species other than Candida albicans.Microbiology 144, 55-72.
Berridge, M. J. and Irvine, R. F. (1989). Inositol phosphates and cellsignaling. Nature 341, 197-205.
Bezprozvanny, I. and Ehrlich, B. E. (1993). ATP modulates the function ofinositol 1,4,5 triphosphate-gated channels at two sites. Neuron 10, 1175-1184.
Biswas, S., Dalal, B., Sen, M. and Biswas, B. B. (1995). Receptor for myo-inositol trisphosphate from the microsomal fraction of Vigna radiata.Biochem. J. 306, 631-636.
Borgeson, C. E. and Bowman, B. J. (1983). Isolation and characterizationof the Neurospora crassa endoplasmic reticulum. J. Bacteriol. 156, 362-368.
Bowman, E. J. and Bowman, B. J. (1982). Identification and properties ofan ATPase in vacuolar membranes of Neurospora crassa. J. Bacteriol. 151,1326-1337.
Bowman, E. J., Bowman, B. J. and Slayman, C. W. (1981). Isolation andcharacterization of plasma membrane from wild type Neurospora crassa. J.Biol. Chem. 256, 12336-12342.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding.Anal. Biochem. 72, 248-254.
Brosnan, J. M. and Sanders, D. (1990). Inositol trisphosphate-mediatedCa2+ release in beet microsomes is inhibited by heparin.FEBS Lett. 260,70-72.
Brunton, A. H. and Gadd, G. M. (1991). Evidence for an inositol lipid signalpathway in the yeast-mycelium transition of Ophiostoma (Ceratocystis)ulmi, the Dutch elm disease fungus. Mycol. Res. 95, 484-491.
Calvert, C. M. and Sanders, D. (1995). Inositol trisphosphate-dependent andindependent Ca2+ mobilization pathways at the vacuolar membrane ofCandida albicans. J. Biol. Chem. 270, 7272-7280.
Collinge, A. J. and Trinci, A. P. (1974). Hyphal tips of wild- type andspreading colonial mutants of Neurospora crassa. Arch. Microbiol. 99, 353-368.
Cornelius, G. and Nakashima, H. (1987). Vacuoles play a decisive role incalcium homeostasis in Neurospora crassa. J. Gen. Microbiol. 133, 2341-2347.
Cornelius, G., Gebauer, G. and Techel, D. (1989). Inositol trisphosphateinduces calcium release from Neurospora crassavacuoles. Biochem.Biophys. Res. Commun. 162, 852-856.
Diver, J. M., Sage, S. O. and Rosado, J. A. (2001). The inositol trisphosphatereceptor antagonist 2-aminoethoxydiphenylborate (2-APB) blocks Ca2+
entry channels in human platelets: cautions for its use in studying Ca2+
influx. Cell Calcium30, 323-329.Drobak, B. K. and Ferguson, I. B. (1985). Release of Ca2+ from plant
hypocotyl microsomes by inositol-1,4,5 trisphosphate. Biochem. Biophys.Res. Commun. 130, 1241-1246.
Felle, H. H. and Hepler, P. K. (1997). The cytosolic Ca2+ concentrationgradient of Sinapis alba root hairs revealed by Ca2+ selective microelectrodetests and fura-dextran ratio imaging. Plant Physiol. 114, 39-45.
Franklin-Tong, V. E., Drobak, B. K., Allan, A. C., Watkins, P. A. C. andTrewavas, A. J. (1996). Growth of pollen tubes of Papaver rhoeasisregulated by a slow-moving calcium wave propagated by inositol 1,4,5-trisphosphate. Plant Cell8, 1305-1321.
Gadd, G. M. and Foster, S. A. (1997). Metabolism of inositol 1,4,5trisphosphate in Candida albicans: significance as a precursor of inositolpolyphosphates and in signal transduction during the dimorphic transitionfrom yeast cells to germ tube. Microbiology 143, 437-448.
Garrill, A., Jackson, S. L., Lew, R. R. and Heath, I. B. (1993). Ion channelactivity and tip-growth: tip-localized stretch-activated channels generate anessential Ca2+ gradient in the oomycete Saprolegnia ferax. Eur. J. Cell Biol.60, 358-365.
Goldstein, D. A. (1978). Calculations of the concentrations of free cations and
cation-ligand complexes in solutions containing multiple divalent cationsand ligands. Biophys. J. 26, 235-342.
Hanson, B. A. (1991). The effects of lithium on the phosphoinositides andinositol phosphate ofNeurospora crassa. Exp. Mycol. 15, 76-90.
Hosking, S. L., Robson, G. D. and Trinci, A. P. J. (1995). Phosphoinositidesplay a role in hyphal extension and branching in Neurospora crassa. Exper.Mycol. 19, 71-80.
Jung, O. J., Lee, E. J., Kim, J. W., Chung, Y. R. and Lee, C.-W. (1997).Identification of putative phosphoinositide-specific phospholipase C genesin filamentous fungi. Mol. Cells 7, 192-199.
Kallies, A., Gebauer, G. and Rensing, L. (1998). Heat shock effects onsecond messenger systems ofNeurospora crassa. Arch. Microbiol. 170, 191-200.
Kinnunen, P. J. K. (2000). Lipid bilayers as osmotic response elements. CellPhysiol. Biochem. 10, 243-250.
Kost, B., Lemichez, E., Spielhofer, P., Hong, Y., Tolias, K., Carpenter, C.and Chua, N.-H. (1999). Rac homologues and compartmentalizedphosphatidylinositol 4,5-bisphosphate act in a common pathway to regulatepolar pollen tube growth. J. Cell Biol. 145, 317-330.
Kukkonen, J. P., Lund, P. E. and Akerman, K. E. (2001). 2-aminoethoxydiphenyl borate reveals heterogeneity in receptor-activatedCa2+ discharge and store-operated Ca2+ influx. Cell Calcium30, 117-129.
Lakin-Thomas, P. L. (1993a). Effects of inositol starvation on the levels ofinositol phosphates and inositol lipids in Neurospora crassa. Biochem. J.292, 805-811.
Lakin-Thomas, P. L. (1993b). Evidence against a direct role for inositolphosphate metabolism in the circadian oscillator and the blue light signaltransduction pathway in Neurospora crassa. Biochem J. 292, 813-818.
Levina, N. N., Lew, R. R., Hyde, G. J. and Heath, I. B. (1995). The rolesof Ca2+ and plasma membrane ion channels in hyphal tip growth ofNeurospora crassa. J. Cell Sci. 108, 3405-3417.
Lew, R. R. (1999). Comparative analysis of Ca2+ and H+ flux magnitude andlocation along growing hyphae of Saprolegnia ferax and Neurospora crassa.Eur. J. Cell Biol. 78, 892-902.
Malho, R. (1998a). Role of 1,4,5-inositol triphosphate-induced Ca2+ releasein pollen tube orientation. Sex Plant Reprod. 11, 231-235.
Malho, R. (1998b). The role of calcium and associated proteins in tip growthand orientation. In Sexual Plant Reproduction and BiotechnologicalApplications (M. Cresti, G. Cai, A. Moscatelli, eds), pp. 253-270. Vienna,New York: Springer Verlag.
Muir, S. R. and Sanders, D. (1997). Inositol 1,4,5-trisphosphate-sensitiveCa2+ release across nonvacuolar membranes in cauliflower.Plant Physiol.114, 1511-1521.
Pierson, E. S., Miller, D. D., Callaham, D. A., van Aken, J., Hackett, G.and Hepler, P. K. (1996). Tip-localized calcium entry fluctuates duringpollen tube growth. Dev. Biol. 174, 160-173.
Prior, S. L., Cunliffe, B. W., Robson, G. D. and Trinci, A. P. (1993).Multiple isomers of phosphatidyl inositol monophosphate and inositol bis-and trisphosphates from filamentous fungi. FEMS Microbiol. Lett. 110,147-152.
Prior, S. L., Robson, G. D. and Trinci, A. P. J. (1994). Phosphoinositideturnover does not mediate the effects of light or choline or the relief ofderepression of glucose metabolism in filamentous fungi. Mycol. Res. 98,291-294.
Reiss, H. D., Herth, W. and Nobiling, R. (1985). Development of membraneand calcium-gradients during pollen germination ofLilium longifluorum.Planta. 163, 84-90.
Schultz, C., Gebauer, G., Metschies, T., Rensing, L. and Jastorff, B. (1990).Cis,cis-cyclohexane 1,3,5-triol polyphosphates release calcium fromNeurospora crassavia an unspecific Ins 1,4,5-P3 receptor. Biochem.Biophys. Res. Commun. 166, 1319-1327.
Schmid, J. and Harold, F. M. (1988). Dual roles for calcium ions in apicalgrowth of Neurospora crassa. J. Gen. Microbiol. 134, 2623-2631.
Silverman-Gavrila, L. B. and Lew, R. R. (2000). Calcium and tip-growth inNeurospora crassa. Protoplasma 213, 203-217.
Silverman-Gavrila, L. B. and Lew, R. R. (2001). Regulation of the tip-high[Ca2+] gradient in growing hyphae of the fungus Neurospora crassa. Eur. J.Cell Biol. 80, 379-390.
Stevenson, J. M., Perera, I. Y., Heilmann, I., Persson, S. and Boss, W.F. (2000). Inositol signaling and plant growth. Trends Plant Sci. 5, 252-258.
Thrower, E. C., Hagar, R. E. and Ehrlich, B. E. (2001). Regulation ofIns(1,4,5)P3 receptor isoforms by endogenous modulators. Trends. Pharm.Sci. 22, 580-586.
Journal of Cell Science 115 (24)

5025IP3-activated channels and tip growth
Tisi, R., Coccetti, P., Banfi, S. and Martegani, E. (2001). 3-Nitrocoumarinis an efficient inhibitor of budding yeast phospholipase-C. Cell Biochem.Funct. 19, 229-235.
Torralba, S., Heath, I. B. and Ottensmeyer, F. P. (2001). Ca2+ shuttling invesicle during tip growth in Neurospora crassa. Fungal Gen. Biol. 33, 181-193.
Vaughn, L. E. and Davis, R. H. (1981). Purification of vacuoles fromNeurospora crassa. Mol. Cell. Biol. 1, 797-806.
Very, A.-A. and Davies, J. M. (2000). Hyperpolarization-activated calciumchannels at the tip of Arabidopsisroot hairs. Proc. Natl. Acad. Sci. USA 97,9801-9806.
Voegel, H. J. (1959). A convenient growth medium for Neurospora. Microb.Genet. Bull. 13, 42-46.
Zalokar, M. (1959). Growth and differentiation of Neurospora crassahyphae.Am. J. Bot. 46, 602-610.
Zheng, Z.-L. and Yang, Z. (2000). The Rop GTPase switch turns on polargrowth in pollen. Trends Plant Sci. 5, 298-303.
Yuan, S. and Heath, I. B. (1991). Chlortetracycline staining patterns of growinghyphal tips of the oomycete Saprolegnia ferax. Exp. Mycol. 15, 91-102.
Zaytsev, I. D. and Aseyev, G. G. (1992). Properties of Aqueous Solutions ofElectrolytes(translated by M. A. Lazarev and V. R. Sorochenko), p. 76. BocaRaton, Ann Harbor, London, Tokyo: CRC Press.