involvement of endoplasmic reticulum (er) stress in podocyte injury induced by excessive protein...
TRANSCRIPT

Kidney International, Vol. 68 (2005), pp. 2639–2650
Involvement of endoplasmic reticulum (ER) stress in podocyteinjury induced by excessive protein accumulation
REIKO INAGI, MASAOMI NANGAKU, HIROSHI ONOGI, HIROSHI UEYAMA, YASUKO KITAO,KIYOKAZU NAKAZATO, SATOSHI OGAWA, KIYOSHI KUROKAWA, WILLIAM G. COUSER,and TOSHIO MIYATA
Molecular and Cellular Nephrology, Institute of Medical Sciences, Tokai University, Kanagawa, Japan; Division of Nephrology andEndocrinology, University of Tokyo School of Medicine, Tokyo, Japan; Department of Neuroanatomy, Kanazawa UniversityMedical School, Kanazawa, Japan; and Division of Nephrology, University of Washington, Seattle, Washington
Involvement of endoplasmic reticulum (ER) stress in podocyteinjury induced by excessive protein accumulation.
Background. An imbalance between protein load and foldingcapacity is referred to as endoplasmic reticulum (ER) stress. Asa defense mechanism, cells express ER stress inducible chap-erons, such as oxygen-regulated proteins 150 (ORP150) andglucose-regulated proteins (GRPs). While ER stress is impor-tant in various diseases, a pathophysiologic role for ER stressin kidney disease remains elusive. Here we investigate expres-sion of ER stress proteins in cultured rat podocytes as well asin our recently developed animal model of abnormal proteinretention within the ER of podocytes (i.e., megsin transgenicrat).
Methods. The expression of ER stress inducible proteins(ORP150, GRP78, or GRP94) in cultured podocytes treatedwith tunicamycin, A23187, SNAP, hypoxia, or hyperglycemia,and the renal tissues or isolated glomeruli from megsin trans-genic rats was analyzed by Western blotting analysis, immuno-histochemistry, or confocal microscopy.
Results. Cultured podocytes demonstrated that treatmentwith tunicamycin, A23187, and SNAP, but not hypoxia or hyper-glycemia, up-regulate expression of ER stress proteins. Extractsof isolated glomeruli from megsin transgenic rats reveal markedup-regulation of ER stress chaperones in podocytes, whichwas supported by immunohistochemical analysis. Confocal mi-croscopy revealed that ER stress in podocytes was associatedwith cellular injury. Podocytes of transgenic rats overexpress-ing a mutant megsin, without the capacity for polymerizationwithin the ER, do not exhibit ER stress or podocyte damage,suggesting a pathogenic role of ER retention of polymerizedmegsin.
Conclusion. This paper implicates a crucial role for the ac-cumulation of excessive proteins in the podocyte ER in theinduction of ER stress and associated podocyte injury.
Key words: serpin, megsin, protein malfolding, podocyte, proteinuria,oxygen/glucose-regulated protein (ORP/GRP).
Received for publication December 10, 2004and in revised form June 6, 2005Accepted for publication July 20, 2005
C© 2005 by the International Society of Nephrology
The endoplasmic reticulum (ER) is an intracellularcompartment that plays a critical role in the processing,folding, and exporting of newly synthesized proteins intothe secretory pathway [1]. All cells regulate the capacityof their ER to process client proteins and adapt to an im-balance between client protein load and folding capacity,recently referred to as ER stress [2, 3]. An impairmentof ER membrane integrity due to stress can be injuriousby causing leakage of calcium ions and other ER luminalcomponents.
ER stress is triggered by various stimuli, including anincrease in synthesis of client proteins, and also occursin the course of various pathophysiologic states [4, 5].The ER stress known as the unfolded protein response(UPR) is currently the best understood model of ERstress signaling, a response conserved from yeast to hu-man [6]. The UPR involves transient attenuation of newprotein synthesis, degradation of misfolded proteins, on-set of apoptosis, and expression of a variety of ER stressproteins, such as oxygen-regulated protein (ORP) 150and glucose-regulated proteins (GRPs). Under normalconditions, these ER stress proteins serve as protein chap-erones, complex with defective proteins, and target themfor degradation. During stress, the induction of ER stressproteins may limit accumulation of abnormal proteins incells [6, 7], allowing cells to tolerate an accumulation ofunfolded proteins.
Previous studies have shown that ER stress is closelyassociated with the neuronal cell injury caused by is-chemia [8], epileptic seizures [8], Alzheimer’s disease [9],Parkinson’s disease [10], and polyglutamine diseases [11].ER stress is also involved in muscle cell injury in sporadicinclusion body myositis [12]. Homocysteine also inducesER stress in vascular endothelial cells [13], suggestingits potential role in thrombotic vascular events. Further-more, ER stress induces apoptosis of pancreatic cells andsubsequent development of diabetes mellitus [14, 15]. On
2639

2640 Inagi et al: ER stress in podocyte injury of megsin Tg rat
the other hand, a pathophysiologic role of ER stress inkidney disease has not been defined.
Accumulating evidence emphasizes an essential roleof podocyte injury in proteinuria and progression of kid-ney failure [16, 17]. Podocytes contain a well-developedGolgi system, prominent lysosomes, many mitochondria,and abundant rough and smooth endoplasmic reticulum[18]. The density of these organelles in the cell body in-dicates a high level of anabolic as well as catabolic activ-ity of podocytes, stimulating us to study susceptibility ofpodocytes to ER stress. Adaptive responses of podocytesto ER stress are crucial because podocytes are terminallydifferentiated cells that are unable to proliferate, and lossof these cells can lead to glomerulosclerosis and end-stagekidney failure [19, 20].
In the present study, we investigated expression of ERstress proteins in cultured rat podocytes as a defensivemechanism in vitro. We also employed our recently de-veloped megsin transgenic rat to study expression of ERstress proteins in vivo. Megsin is a new member of the ser-pin (serine protease inhibitor) superfamily cloned froma human mesangial cDNA library [21–26]. An increasedsynthesis of megsin led to flooding of the protein han-dling system and subsequent accumulation of polymeriz-ing megsin within the ER of podocytes [27]. It is likelythat a common structure of reactive site loop and b-sheet, shared by serpins, leads to the aberrant intermolec-ular linkage and polymerization of megsin. These animalsdevelop periodic acid Schiff (PAS)–positive, diastase-resistant megsin inclusions within the ER of podocytesin association with proteinuria and deterioration of renalfunction [27].
METHODS
In vitro rat podocyte experiments
Primary cultured rat podocytes were isolated accord-ing to methods described elsewhere [28] In brief, thecells were initially established on Matrigel-coated tis-sue culture dishes (BD Biosciences, Bedford, MA, USA)and the cells were maintained onto a collagen type I-coated tissue culture dishes in K-1 medium supplementedwith 2% NuSerum (Collaborative Biomedical Products,Bedford, MA, USA), 5 mg/mL insulin, 5 mg/mL trans-ferrin, 5 ng/mL selenium (Sigma-Aldrich, St. Louis, MO,USA), 100 U/mL penicillin, and 100 mg/mL streptomycin(Invitrogen, Carlsbad, CA, USA) under humidified at-mosphere containing 5% CO2. Rat podocytes used in thisstudy showed a cobblestone-like appearance and werepositive for megalin, cytokeratin, and podocalyxin, butnot factor VIII or Thy-1.1, suggesting that the cells weused in this study retained the properties for podocytes.
To evaluate the exposure to several stimuli includingabnormal protein retention, calcium store depletion, ni-
trosative stress, hypoxia, and hyperglycemia, podocyteswere treated with 5 lg/mL tunicamycin (Sigma-Aldrich),a reagent to cause an accumulation of unfolded pro-teins in the ER, 10 lg/mL A23187, a calcium ionophore,110 lg/mL S-nitroso-N-acetyl-D,L-penicillamine (SNAP;Dojin, Kumamoto, Japan), a NO donor, or hypoxia uti-lizing Anaerocult A (<0.1% oxygen; Merck, Darmstadt,Germany) for 24 hours. For exposure to glycemic stress,podocytes were cultured in the presence of 4.5 mg/mLD- or L-glucose for two weeks. In glycemic stress we em-ployed a long exposure time (2 weeks) to mimic chronicderangement of carbohydrate metabolism in diabeticpatients.
Western blot analysis
Proteins (10 mg) from cell lysates or glomeruli isolatedaccording to a conventional sieving method [29] were ho-mogenized in 100 lL of 0.35 mol/L Tris-HCl (pH 6.8) con-taining 10% sodium dodecyl sulfate (SDS), 36% glycerol,5% b-mercaptoethanol, and 0.012% bromophenol blueand centrifuged at 5000 g for 15 minutes. Three to five llof the obtained test samples were electrophoresed onsodium dodecyl sulfate-polyacrylamide gel electrophore-sis (SDS-PAGE), and transferred to polyvinylidene fluo-ride (PVDF) membrane.
To detect ER stress inducible proteins, rabbit poly-clonal antibodies to ORP150 (purified IgG fraction, 0.5lg/mL) [30] or GRP94 (0.4 lg/mL; Stressgen Biotech-nologies, Victoria, BC, Canada), or goat anti-GRP78polyclonal antibody (20 lg/mL; Santa Cruz Biotechnol-ogy, Inc., Santa Cruz, CA, USA) were used as the firstantibody and horseradish peroxidase–conjugated goatanti-rabbit antibodies (1:5000 dilution; Bio-Rad, Her-cules, CA, USA) or alkaline phosphatase-conjugated rab-bit anti-goat IgG (H+L) (1:5000 dilution; Jackson Im-munoResearch Lab, Inc., West Grove, PA, USA) as thesecondary antibody. Rabbit anti-actin polyclonal anti-body (3.5 lg/mL; Sigma-Aldrich) was also used as the firstantibody. The detection for ORP150, GRP94, or actin wasperformed by the enhanced chemiluminescence (ECL)system (Amersham Biosciences, Piscataway, NJ, USA).For detection of GRP78, p-nitro blue tetrazolium chlo-ride and 5-bromo-4-chloro-3-indolyl-phosphate solution(Bio-Rad) was used. In some experiments, quantificationof the intensity of each band was performed by densit-ometry utilizing ATTO lane analyzer software (ATTO,Tokyo, Japan) and normalized by the band intensity ofactin. The experiment was repeated three times.
Megsin transgenic rats
To generate the human megsin transgene construct,the entire coding sequence of megsin cDNA was sub-cloned in the sense orientation into the pBsCAG-2 [24].

Inagi et al: ER stress in podocyte injury of megsin Tg rat 2641
For production of mutant megsin (megsinT334R) trans-genic rats, the site-directed mutagenesis at the position ofP14 (substitution of Thr for Arg at 334 residue) in the re-active site loop of human megsin gene was performedutilizing a standard two-step megaprimer polymerasechain reaction (PCR) with mutated oligonucleotides, andmegsinT334R cDNA was subcloned into the pBsCAG-2 [27]. The megsin transgenes isolated by digestion ofpBsCAG-2 containing megsin cDNA were microinjectedinto one pronucleus of fertilized Wistar rat eggs, fol-lowed by transfer into the oviducts of pseudopregnantrats. Rat genomic DNA extracted from the tail was usedto identify megsin transgenic rats by PCR as describedpreviously [24]. Kidneys from homozygote megsin andmegsinT334R transgenic rats at the age of 8 weeks wereused for PAS staining, immunohistochemistry, and West-ern blot analysis. Animals were treated in accordancewith the guidelines of the Committee on Ethical AnimalCare and Use of Tokai University.
Histology
For light microscopic analysis, kidneys were fixed in4% neutral buffered formaldehyde, embedded in paraf-fin, and sectioned at 4 lm thickness, followed by PASstaining.
Immunohistochemistry
For detection of ORP150, GRP78, or desmin as amarker of podocyte injury [31], tissue sections (2 lm)of kidneys fixed with methyl Carnoy’s were used. Thesections were first incubated with rabbit polyclonal anti-ORP150 antibodies (5 lg/mL purified IgG fraction) [30]or goat polyclonal anti-GRP78 antibodies (1:100 dilu-tion; Santa Cruz Biotechnology, Inc.) followed by bi-otinylated goat anti-rabbit IgG (1:200 dilution; ZymedLaboratories, Inc, South San Francisco, CA, USA) orrabbit anti-goat IgG (1:800 dilution; DakoCytomation,Kyoto, Japan), or incubated with mouse monoclonal an-tibodies to desmin (11.5 lg/mL; DakoCytomation) fol-lowed by reaction with biotinylated horse anti-mouseIgG polyclonal antibody (1:400 dilution; Vector Labo-ratories, Inc., Burlingame, CA, USA). Development wasperformed with peroxidase-conjugated avidine (VectorLaboratories, Inc.) and 3,3′-diaminobenzidine tetrahy-drochloride (Wako, Osaka, Japan). Finally, the tissueswere counterstained with hematoxylin. In some exper-iments, normal rabbit IgG, goat IgG, or mouse IgG pur-chased from DakoCytomation were used as controls.
Computer-assisted morphometry
The sections stained immunohistochemically with anti-ORP150 or GRP78 antibodies were used for computer-
assisted morphometry. Each staining picture was scannedusing a 3CCD camera (Olympus Optical Co., Tokyo,Japan), and positive area for ORP150 or GRP78 expres-sion per glomerulus was measured in a blinded mannerusing the software NIH ImageJ (NIH, Bethesda, MD,USA). Twenty consecutive glomeruli in the midcortexwere measured. Glomerular cross sections containingonly a minor portion of the glomerular tuft (<20 discretecapillary segments per cross-section) were not used. Inorder to avoid examiner’s bias, both the largest and thesmallest glomerulus in the 20 glomeruli were excluded.Eighteen glomeruli from each section were examined andaverages were expressed as mean ± SD.
Immunofluorescence studies
Rats were perfused with 4% paraformaldehyde un-der deep anesthesia, and the kidneys were cryostatsectioned (5 lm). Immunofluorescence studies wereperformed using either purified rabbit IgG to humanORP150 (5 lg/mL) [30] or human megsin (P2: 5 lg/mL)[23], goat polyclonal anti-GRP78 antibodies (1:100 di-lution; Santa Cruz Biotechnology Inc.), or mouse mono-clonal antibody to anti-synaptopodin (Progen BiotechnikGmbH, Heidelberg, Germany) followed by visualizationof the binding sites of primary antibodies by the im-munofluorescence method [30]. Images were then digi-tally overlapped.
To investigate localization of megsin/ORP150 indesmin-positive injured podocytes, immunofluorescencestudies with antibodies to human ORP150 and desminfollowed by confocal microscopy were performed. The 4-lm frozen sections were fixed with ethanol:ether (1:1),blocked with 2% bovine serum albumin (BSA) inphosphate-buffered saline (PBS), and incubated with pu-rified rabbit IgG to human ORP150 (5 lg/mL) and mousemonoclonal antibody to desmin (11.5 lg/mL; DakoCy-tomation), followed by incubation with FITC-conjugatedsheep anti-mouse IgG (1:50 dilution; DakoCytoma-tion) and TRITC-conjugated swine anti-rabbit IgG(1:50 dilution; DAKO). The nuclei were counterstainedwith Hoechst Dye 33342 (Calbiochem-Novabiochem, LaJolla, CA, USA), and then the optical tomographic imag-ing (Z-images) for three-dimensional reconstructed im-ages of glomeruli was performed utilizing 510 META(Carl Zeiss, Jena, Germany) [32].
Statistical analysis
Data were expressed as mean ± SD. Analysis of vari-ance (ANOVA) was utilized to evaluate the statistical sig-nificance of various differences. If the analysis detecteda significant difference, the Scheffe’s t test was used tocompare results obtained from the experimental animals.Values are considered significant at P < 0.05.

2642 Inagi et al: ER stress in podocyte injury of megsin Tg rat
ORP150
ORP150
GRP94
GRP94
Actin
8.0
6.0
4.0
2.0
0.0
Fol
d in
duct
ion
4.0
3.0
2.0
1.0
0.0
Fol
d in
duct
ion
****
*
**
**
Contro
l
Tunic
amyc
in
A2318
7
SNAP
< 0.
1% o
xyge
n
High g
lucos
e
Tunic
amyc
in
A2318
7
SNAP
< 0.
1% o
xyge
n
High g
lucos
e
+ – + – + – + – + –
A
B
Fig. 1. Expression of ER stress inducibleproteins in podocytes. Podocytes werecultured in the presence of various stimuliincluding 5 lg/mL tunicamycin to cause anaccumulation of unfolded proteins in the ER,10 lg/mL A23187 as a calcium ionophore,110 lg/mL SNAP as a NO donor, <0.1%oxygen as hypoxia, or 4.5 mg/mL glucoseto induce hyperglycemia. Expression levelsof ER stress inducible chaperone proteins,ORP150 and GRP94, were assessed byWestern blot analysis followed by densito-metry. (A) Representative data from threeexperiments are shown. (B) Quantification ofthe intensity of the bands was performed bydensitometry and normalized to the intensityof actin bands. The mean ratio of ORP150or GRP94 expression in the presence ofvarious stimuli over that in the absence ofstimuli was calculated for each experiment,and the average of the three experiments isexpressed. Treatment with tunicamycin orA23187 markedly up-regulated expressionof ORP150 and slightly of GRP94. SNAPtreatment slightly up-regulated expressionof both ORP150 and GRP94. By contrast,neither high glucose nor hypoxia influencedexpression of ORP150 and GRP94. ∗P <
0.05 vs. control; ∗∗P < 0.01 vs. control. Doseand time dependency of up-regulation ofOPR150 or GRP94 expression in podocytestreated with tunicamycin (C) or A23187(D). Quantification of the intensity of thebands was performed by densitometry andnormalized to the intensity of actin bands.The mean ratio of ORP150 or GRP94expression in the presence of tunicamycin orA23187 over that in the absence of stimuliwas calculated for each experiment, andthe average of the three experiments isexpressed. Treatment with tunicamycin orA23187 markedly up-regulated expressionlevels of ORP150 or GRP94 in a dose-and time-dependent manner. ∗P < 0.05 vs.control, ∗∗P < 0.01 vs. control.

Inagi et al: ER stress in podocyte injury of megsin Tg rat 2643
C8.0
6.0
4.0
2.0
0.0
Fol
d in
duct
ion
3.0
2.0
1.0
0.0
Fol
d in
duct
ion
Contro
l 0.1 3 5 0 6 12 24
Tunicamycin, µg/mL Tunicamycin, hours
Contro
l 0.1 3 5 0 6 12 24
Tunicamycin, µg/mL Tunicamycin, hours
GPR94
ORP150
*
*
*
*
****
**
Fig. 1. (continued)
RESULTS
Expression of ER stress inducible proteins in podocytes
To examine induction of ER stress in podocytes, wesubjected cultured cells to various stimuli, including ab-normal protein retention, calcium store depletion, ni-trosative stress, hypoxia, and hyperglycemia. Expressionlevels of ER stress inducible chaperone proteins, ORP150and GRP94, were assessed by Western blot analysis fol-lowed by densitometry.
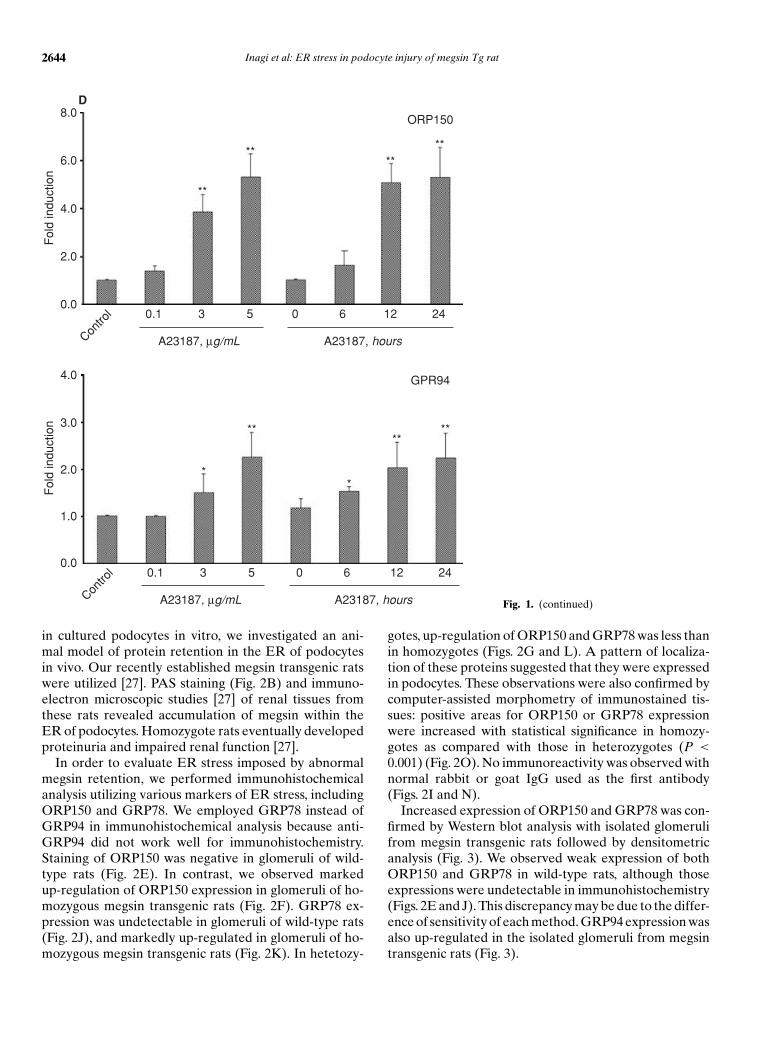
Representative data of Western blot analysis and den-sitometric quantification are shown in Figure 1A and B.Treatment with tunicamycin, a reagent to cause an accu-mulation of unfolded proteins in the ER, or A23187, acalcium ionophore, markedly up-regulated expression ofORP150 (tunicamycin, 5.03- ± 1.56-fold; A23187, 5.26 ±0.97-fold, P < 0.01) and less of GRP94 (tunicamycin,1.40 ± 0.37-fold, P < 0.05; A23187, 2.24 ± 0.53-fold,P < 0.01) in podocytes. We detected a lower molecularweight band of ORP150 in addition to a normal molec-ular weight band of ORP150 in podocyte treated with
tunicamycin. It may be due to abnormal glycosylationby tunicamycin. Up-regulation of ORP150 and GRP94expression was observed in a time- and dose-dependentmanner (Figs. 1C and D). Up-regulation of ORP150 andGRP94 in the podocytes treated with SNAP, a NO donor,was mild but statistically significant (2.32 ± 0.34-fold and1.45 ± 0.1-fold, P < 0.05, respectively). By contrast, nei-ther high D-glucose nor hypoxia influenced expression ofORP150 (high glucose, 0.89 ± 0.12-fold; hypoxia, 1.16 ±0.19-fold) and GRP94 (high glucose, 0.97 ± 0.03-fold; hy-poxia, 1.17 ± 0.20-fold) in podocytes. High concentrationof L-glucose, which serves as a control of high osmolar-ity, did not change expression levels of ORP150 (0.92 ±0.17-fold) or GRP94 (1.06 ± 0.13-fold) in podocytes,either.
Megsin transgenic rats suffered from ER stress
As tunicamycin blocks N-linked glycosylation andcauses retention of unfolded proteins in the ER, andturned out to be a powerful inducer of ER stress proteins
2644 Inagi et al: ER stress in podocyte injury of megsin Tg rat
D8.0
6.0
4.0
2.0
0.0
Fol
d in
duct
ion
4.0
3.0
2.0
0.0
1.0
Fol
d in
duct
ion
Contro
l 0.1 3 5 0 6 12 24
A23187, µg/mL A23187, hours
Contro
l 0.1 3 5 0 6 12 24
A23187, µg/mL A23187, hours
GPR94
ORP150
**
****
**
**
**
****
Fig. 1. (continued)
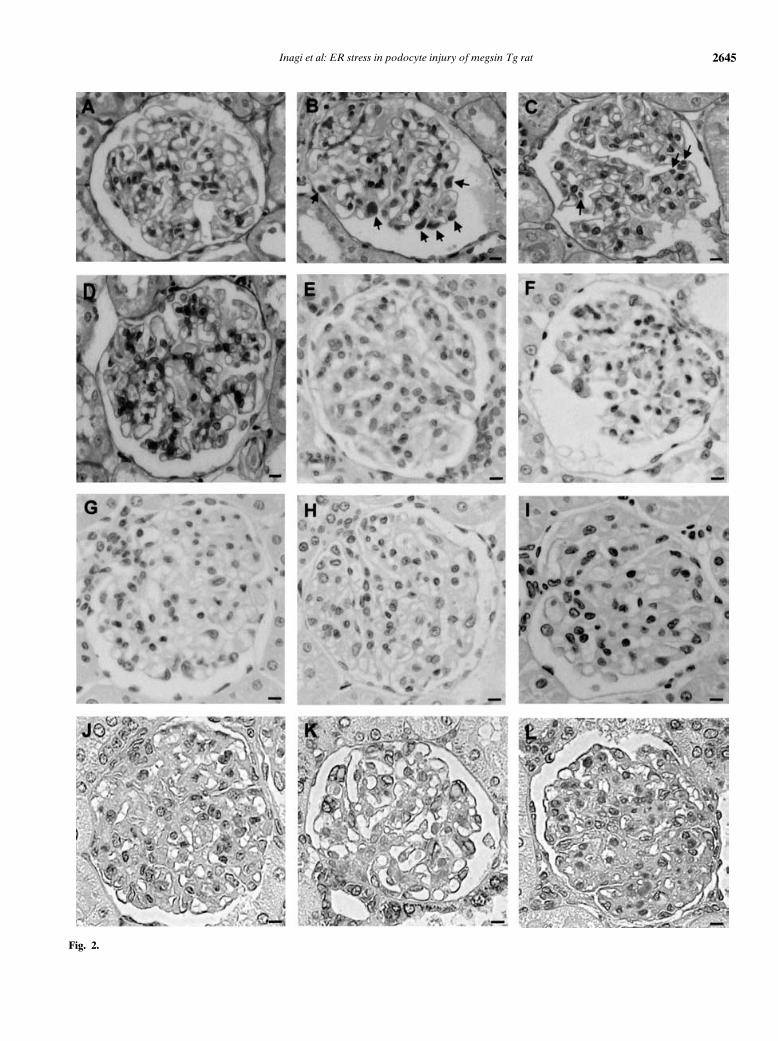
in cultured podocytes in vitro, we investigated an ani-mal model of protein retention in the ER of podocytesin vivo. Our recently established megsin transgenic ratswere utilized [27]. PAS staining (Fig. 2B) and immuno-electron microscopic studies [27] of renal tissues fromthese rats revealed accumulation of megsin within theER of podocytes. Homozygote rats eventually developedproteinuria and impaired renal function [27].
In order to evaluate ER stress imposed by abnormalmegsin retention, we performed immunohistochemicalanalysis utilizing various markers of ER stress, includingORP150 and GRP78. We employed GRP78 instead ofGRP94 in immunohistochemical analysis because anti-GRP94 did not work well for immunohistochemistry.Staining of ORP150 was negative in glomeruli of wild-type rats (Fig. 2E). In contrast, we observed markedup-regulation of ORP150 expression in glomeruli of ho-mozygous megsin transgenic rats (Fig. 2F). GRP78 ex-pression was undetectable in glomeruli of wild-type rats(Fig. 2J), and markedly up-regulated in glomeruli of ho-mozygous megsin transgenic rats (Fig. 2K). In hetetozy-
gotes, up-regulation of ORP150 and GRP78 was less thanin homozygotes (Figs. 2G and L). A pattern of localiza-tion of these proteins suggested that they were expressedin podocytes. These observations were also confirmed bycomputer-assisted morphometry of immunostained tis-sues: positive areas for ORP150 or GRP78 expressionwere increased with statistical significance in homozy-gotes as compared with those in heterozygotes (P <
0.001) (Fig. 2O). No immunoreactivity was observed withnormal rabbit or goat IgG used as the first antibody(Figs. 2I and N).
Increased expression of ORP150 and GRP78 was con-firmed by Western blot analysis with isolated glomerulifrom megsin transgenic rats followed by densitometricanalysis (Fig. 3). We observed weak expression of bothORP150 and GRP78 in wild-type rats, although thoseexpressions were undetectable in immunohistochemistry(Figs. 2E and J). This discrepancy may be due to the differ-ence of sensitivity of each method. GRP94 expression wasalso up-regulated in the isolated glomeruli from megsintransgenic rats (Fig. 3).
Inagi et al: ER stress in podocyte injury of megsin Tg rat 2645
Fig. 2.

2646 Inagi et al: ER stress in podocyte injury of megsin Tg rat
100
80
60
40
20
0
300
250
200
150
100
50
0
Pos
itive
for
OR
P15
0 ex
pres
sion
(µm
2 )
Pos
itive
for
GP
R78
exp
ress
ion
(µm
2 )
# #
*
*
*
*
Wild
-type
meg
sin T
g (h
omo)
meg
sin T
g (h
eter
o)
T334R
meg
sin T
g
Wild
-type
meg
sin T
g (h
omo)
meg
sin T
g (h
eter
o)
T334R
meg
sin T
g
O
Fig. 2. Induction of ER stress inducible pro-teins in the kidneys of megsin transgenic rats.PAS staining of a wild-type (A), a megsintransgenic (homozygote, B, and heterozygote,C), and a mutated-megsin (megsinT334R)transgenic rat (D) at the age of 8 weeks. PAS-positive deposits (indicated by arrows) wereobserved within the cytoplasm of glomerularepithelial cells in a megsin transgenic rat butnot in a wild-type rat or a megsinT334R trans-genic rat. Immunohistochemical detection ofORP150 (E–I) and GRP78 (J–N) in the kid-ney of a wild-type rat (E and J), a megsin ho-mozygote (F and K), a megsin heterozygote(G and L), and a megsinT334R transgenicrat (H and M). Expression of ORP150 andGRP78 is markedly augmented in glomeruliof megsin transgenic rats, as compared withthose in wild-type and megsinT334R trans-genic rats. No immunoreactivity was observedwith normal rabbit or goat IgG as the firstantibody in a megsin homozygote rat (I andN). The bar indicates 10 lm. (O) Computer-assisted morphometry of the section stainedimmunohistochemically with anti-ORP150 orGRP78. Each section of three rats derivedfrom each group was examined, and aver-ages were expressed as mean ± SD. Pos-itive areas for ORP150 or GRP78 expres-sion were increased with statistical signifi-cance in homozygotes as compared with thosein heterozygotes. ∗P < 0.001 vs. wild-type orT334Rmegsin transgenic rats. #P < 0.001.
Polymerization of megsin and subsequent ER reten-tion play a pathogenic role in development of ER stressin podocytes. Podocytes of heterozygote transgenic ratsoverexpressing mutant megsin (megsinT334R, a muta-tion at the position of P14 corresponding to the proximalhinge region of the reactive site loop), which lacks ser-ine protease inhibitory activity as well as the capacityfor polymerization within the ER, did not have megsindroplets within the cytoplasm (Fig. 2D) [27]. In addi-tion, they did not show increased expression of ER stressmarkers, ORP150, GRP94, or GRP78 by immunohisto-chemistry (Figs. 2H and M) and Western blot analysis forisolated glomeruli (Fig. 3).
ER stress induced in podocytes with excessive megsinaccumulation
We previously demonstrated that accumulation of thetransgene product, megsin, was localized exclusively in
podocytes in glomeruli by immunoelectron microscopicanalysis [27]. Of note, OPR150-expressed podocytes ofmegsin transgenic rats showed accumulation of megsintransgene product (Fig. 4).
Podocyte damage by ER stress in megsin transgenic rats
We further performed immunohistochemical analy-sis utilizing desmin as a marker of podocyte injury [31]in order to clarify whether ER stress in podocytes ofmegsin transgenic rats was associated with cellular injury.Glomeruli of megsin transgenic rats showed expression ofdesmin in podocytes and the signal intensity was higher inhomozygotes than in heterozygotes (Fig. 5A, panels b andc). Whereas staining for desmin was absent in glomeruliof wild-type (Fig. 5A, panel a). No immunoreactivity wasobserved with normal mouse IgG used as the first anti-body (Fig. 5A, panel e). Confocal microscopic analysis ofdouble staining of desmin and ORP150 localized ORP150

Inagi et al: ER stress in podocyte injury of megsin Tg rat 2647
ORP150
ORP150
GRP78
GRP78
GRP94
GRP94
β-actin
5.0
0.0
1.0
2.0
3.0
4.0
Fol
d in
duct
ion
*
**
Wild
-type
rats
T334R
meg
sin T
g
meg
sin T
g
Wild
-type
rats
T334R
meg
sin T
g ra
ts
meg
sin T
g ra
ts
AB
Fig. 3. Western blot analysis of ER stressinducible proteins. Proteins from isolatedglomeruli derived from wild-type, megsin ho-mozygote transgenic, and mutated megsin(T334Rmegsin) transgenic rats (8-week-old)were analyzed by Western blot analysis usingantibodies to ER stress inducible proteinssuch as ORP150, GRP78, and GRP-94. Poly-clonal antibodies to b-actin were also usedas an internal control. (A) Representativedata from three independent experiments areshown. (B) Quantification of the intensity ofthe bands was performed by densitometry andnormalized to the intensity of actin bands. Themean ratio of ORP150, GRP78, or GRP94expression in megsin or T334Rmegsin trans-genic rats over that in wild-type rats was calcu-lated for each experiment, and the average ofthe three experiments is expressed. Increasedprotein expression of ORP 150, GRP78, andGRP94 was observed in isolated glomerulifrom megsin transgenic rats, as comparedwith those from age-matched wild-type orT334Rmegsin transgenic rats. ∗P < 0.05 vs.wild-type rats.
Megsin ORP150 Merge
Fig. 4. ER stress is induced in podocytes withexcessive megsin accumulation. Localizationof ER stress inducible proteins in glomerulifrom megsin homozygote transgenic rats (8-week-old) was assessed by double stainingusing antibodies to ORP150 and megsin.Frozen kidney sections (5 lm) were incu-bated with rabbit polyclonal IgG to humanORP150 or human megsin followed by reac-tion with FITC- or TRITC-conjugated anti-rabbit IgG. OPR150-expressing podocytes ofmegsin transgenic rats showed accumulationof megsin transgene products. Representa-tive positively stained cells were indicated bywhite arrowheads. The bar indicates 10 lm.
(red) in podocytes with the cytoplasm positive for desmin(green). It suggests that podocyte injury was associatedwith ER stress (Fig. 5B).
We further analyzed podocyte damage by double stain-ing of synaptododin, a marker for podocytes, and GRP78in megsin homozygote transgenic rats. The signal ofsynaptopodin expression adjacent to GRP78 positive sig-nal was relatively faint as compared with that in wild-typerats, which was negative for GRP78 staining (Fig. 5C).
By contrast, in megsinT334R transgenic rats, thedesmin staining was extremely weak (Fig. 5A, panel d).These results, together with the lack of PAS-positive ma-terials composed of megsin and ORP150 in the glomeruliof megsinT334R transgenic rats, clearly suggested thatpodocyte injury was not induced in this model.
DISCUSSION
ER stress in glomerular injury was recently empha-sized by Cybulsky et al [33]. They reported that comple-
ment attack against podocytes induced ER injury, whichwas associated with up-regulation of ER stress proteinssuch as GPR78 (bip) and GRP94 both in vitro and invivo. Our results confirm and extend their findings be-yond complement-induced podocyte injury.
Tunicamycin, which causes retention of unfolded pro-teins in the ER, induced ER stress in podocytes. In con-trast, neither hypoxia nor high glucose resulted in ERstress in these cells. Factors inducing ER stress may be celltype–specific because hypoxia can lead to up-regulationof ER stress proteins in astrocytes [34], vascular smoothmuscle cells [35], monocytes [35], ovary cells [36], andrenal tubular cells [37]. Glomerular cells are relativelyspared in ischemic kidney diseases compared with tubularcells [38], and hypoxia may not be as noxious to podocytesas it is to some other types of cells. In agreement withour in vitro findings, immunohistochemical analysis con-ducted in a rat ischemia-reperfusion of model showedthat ORP150 staining is more prominent in tubular cellsthan in glomeruli [39].

2648 Inagi et al: ER stress in podocyte injury of megsin Tg rat
Synaptopodin GRP78 Merge
C
Fig. 5. Podocyte damage by ER stress in megsin transgenic rats. (A) Immunohistochemical detection of desmin, a marker of podocyte injury, in thekidney of a wild-type (panel a), a megsin transgenic rat (homozygote, panel b, and heterozygote, panel c), and a megsinT334R transgenic rat (paneld) at the age of 8 weeks. The sections (2 lm) were first incubated with anti-desmin monoclonal antibody, followed by reaction with biotinylatedanti-mouse IgG. Expression of desmin was detected in glomeruli of a megsin transgenic rat but not in wild-type or megsinT334R transgenic rats.No immunoreactivity was observed with normal mouse IgG as the first antibody in a megsin homozygote rat (panel e). The bar indicates 10 lm.

Inagi et al: ER stress in podocyte injury of megsin Tg rat 2649
It was not surprising that high glucose similar to what isseen in diabetes mellitus did not induce expression of ERstress proteins because ER stress proteins are known tobe up-regulated by energy depletion under low glucoseconditions [40, 41].
Here we demonstrated a crucial role of accumulationof excessive proteins in the ER in induction of ER stressutilizing our recently established megsin transgenic rats[27]. Megsin is a novel member of the serpin superfam-ily, and serpins are known to form aberrant intermolec-ular linkage in hepatocytes and neurons [42, 43]. To ourknowledge, this is the first report to show that ER stressis associated with excessive protein accumulation in renalcells. It should be noted that the transgene of our megsintransgenic rats coded an intact molecule, in contrast tosome human genetic disorders in which mutations in thecoding region of highly expressed ER client proteins di-rectly perturb their folding [2, 44]. However, previousstudies also showed that unfolding or misfolding of pro-teins can occur under several circumstances, includingmacromolecular crowding, oxidative stress, exposure totoxins, and aging, suggesting that ER stress can play animportant role in a wide range of diseases [8, 45].
Western blotting analysis in this study demonstratedup-regulation of a variety of ER-stress proteins inglomeruli isolated from megsin transgenic rats. We pre-viously described predominant expression of megsin inmesangial cells of normal animals [26, 46], and cannotexclude a possible involvement of mesangial cells, en-dothelial cells, or infiltrating cells in induction of the ERstress proteins. However, immunohistochemical analysisas well as immunoelectron microscopic analysis [27] re-vealed that megsin transgenic rats unexpectedly showedaccumulation of megsin in ER of podocytes. Expressionpatterns of the transgene revealed by in situ hybridizationwere ubiquitous in resident glomerular cells of the trans-genic rats [27]. Accumulation of megsin also varied in dif-ferent organs in spite of similar expression at the mRNAlevel [27]. These observations led us to speculate that indi-vidual cellular susceptibility must influence accumulationof the transgene product in ER, demonstrating predom-inant accumulation of megsin in podocytes of the trans-genic rats. However, this requires further investigation.
(B) Localization of ORP150 within injured podocytes. Three-dimensional colored intensity orthogonal section imaging of glomeruli were carried outafter immunofluorescence studies using antibodies to ORP150 (red) and desmin (green), a marker of podocyte injury. Nuclei were poststained withHoechst Dye 33342. X-Y cross-section plane image (blue line square), X-Z cross-section plane image (green line square), Y-Z cross-section planeimage (red line square). ORP150 (red) was localized in podocytes with the cytoplasm positive for desmin (green), suggesting that podocyte injurywas associated with ER stress. The bar indicates 5lm. (C) Double immunostaining of an ER stress inducible protein, GRP78, and synaptopodin, amarker of podocytes, in glomeruli from megsin homozygote transgenic rats (8-week-old). Frozen kidney sections (4 lm) from age-matched wild-type(upper panel) or megsin transgenic rats (lower panels) were incubated with mouse monoclonal antibodies to synaptopodin and goat polyclonal IgGto GRP78, which were detected by TRITC-conjugated anti-mouse IgG and FITC-conjugated anti-goat IgG, respectively. The signal of synaptopodinexpression adjacent to GRP78 positive signal was relatively faint in megsin transgenic rats compared with that in wild-type rats, which was negativefor GRP78 expression, suggesting podocyte damage by ER stress. The bar indicates 10 lm.
The ER stress in podocytes of megsin transgenic ratswas associated with up-regulation of ER stress proteinssuch as ORP150 and GRP78. ORP150 was first purifiedand cloned as a novel stress protein induced by oxy-gen deprivation in cultured rat astrocytes [34]. Glucose-regulated proteins, GRP78 and GRP94, are also majorconstituents of the ER of mammalian cells and serve asa protective mechanism to adapt to stress on the ER [41,47, 48]. While induction of ER stress proteins may be animportant mechanism of protection of podocytes, furtherstudies are warranted to conclude whether the ER stressresponse is protective or is responsible for some patho-logic findings.
CONCLUSION
We reported induction of ER stress by various stimuliin cultured glomerular cells. Furthermore, our transgenicrats with accumulation of megsin in the ER of podocytesdemonstrated ER stress-induced podocyte injury. Ourfindings implicated a crucial role for the accumulationof excessive proteins in the podocyte ER in the inductionof ER stress and associated podocyte injury.
ACKNOWLEDGMENTS
We thank Dr. Johbu Itoh (Laboratories for Structure and FunctionalResearch, Tokai University School of Medicine, Japan) for his tech-nical support with the confocal microscopic analyses. This study wassupported by grants from the New Energy and Industrial TechnologyDevelopment Organization to T.M., from the Ministry of Education,Culture Sports, Science and Technology of Japan (15590861) to R.I.,and from the Research and Study Program of Tokai University Educa-tional System General Research Organization to R.I. and T.M.
Reprint requests to Toshio Miyata, M.D., Ph.D., Molecular andCellular Nephrology, Institute of Medical Sciences, Tokai University,Kanagawa 259-1193, Japan.E-mail: [email protected]
REFERENCES
1. ELLGAARD L, HELENIUS A: ER quality control: Towards an under-standing at the molecular level. Curr Opin Cell Biol 13:431–437,2001
2. LEE AS: The glucose-regulated proteins: Stress induction and clin-ical applications. Trends Biochem Sci 26:504–510, 2001
3. LEE AS: Mammalian stress response: Induction of the glucose-regulated protein family. Curr Opin Cell Biol 4:267–273, 1992

2650 Inagi et al: ER stress in podocyte injury of megsin Tg rat
4. ARIDOR M, BALCH WE: Integration of endoplasmic reticulum sig-naling in health and disease. Nat Med 5:745–751, 1999
5. RON D: Proteotoxicity in the endoplasmic reticulum: Lessons fromthe Akita diabetic mouse. J Clin Invest 109:443–445, 2002
6. KAUFMANN RJ: Stress signaling from the lumen of the endoplasmicreticulum: Coordination of gene transcriptional and translationalcontrols. Genes Dev 13:1211–1233, 1999
7. PAHL HL: Signal transduction from the endoplasmic reticulum tothe cell nucleus. Physiol Rev 79:683–701, 1999
8. PASCHEN W, FRANDSEN A: Endoplasmic reticulum dysfunction—Acommon denominator for cell injury in acute and degenerative dis-eases of the brain? J Neurochem 79:719–725, 2001
9. KATAYAMA T, IMAIZUMI K, SATO N, et al: Presenilin-1 mutationsdownregulate the signaling pathway of the unfolded-protein re-sponse. Nat Cell Biol 1:479–485, 1999
10. IMAI Y, SODA M, INOUE H, et al: An unfolded putative transmem-brane polypeptide, which can lead to endoplasmic reticulum stress,is a substrate of Parkin. Cell 105:891–902, 2001
11. NISHITOH H, MATSUZAWA A, TOBIUME K, et al: ASK1 is essential forendoplasmic reticulum stress-induced neuronal cell death triggeredby expanded polyglutamine repeats. Genes Dev 16:1345–1355, 2002
12. VATTEMI G, ENGEL WK, MCFERRIN J, et al: Endoplasmic reticulumstress and unfolded protein response in inclusion body myositis mus-cle. Am J Pathol 164:1–7, 2004
13. OUTINEN PA, SOOD SK, PFEIFER SI, et al: Homocysteine-inducedendoplasmic reticulum stress and growth arrest leads to specificchanges in gene expression in human vascular endothelial cells.Blood 94:959–967, 1999
14. SOCHA L, SILVA D, LESAGE S, et al: The role of endoplasmic reticulumstress in nonimmune diabetes: NOD.k iHEL, a novel model of betacell death. Ann NY Acad Sci 1005:178–183, 2003
15. HARDING HP, RON D: Endoplasmic reticulum stress and the devel-opment of diabetes: A review. Diabetes 51(Suppl 3):S455–461, 2002
16. MUNDEL P, SHANKLAND SJ: Podocyte biology and response to injury.J Am Soc Nephrol 13:3005–3015, 2002
17. KRIZ W: Podocyte is the major culprit accounting for the progressionof chronic renal disease. Microsc Res Tech 57:189–195, 2002
18. PAVENSTADT H, KRIZ W, KRETZLER M: Cell biology of the glomerularpodocyte. Physiol Rev 83:253–307, 2003
19. KRIZ W: Progressive renal failure—Inability of podocytes to repli-cate and the consequences for development of glomerulosclerosis.Nephrol Dial Transplant 11:1738–1742, 1996
20. FRIES JW, SANDSTROM DJ, MEYER TW, et al: Glomerular hypertro-phy and epithelial cell injury modulate progressive glomeruloscle-rosis in the rat. Lab Invest 60:205–218, 1989
21. YASUDA K, MIYATA T, NANGAKU M, et al: Functional quantitativeanalysis of the genome in cultured human mesangial cells. KidneyInt 53:154–158, 1998
22. MIYATA T, NANGAKU M, SUZUKI D, et al: A mesangium-predominantgene, megsin, is a new serpin upregulated in IgA nephropathy. J ClinInvest 102:828–836, 1998
23. INAGI R, MIYATA T, SUZUKI D, et al: Specific tissue distribution ofmegsin, a novel serpin, in the glomerulus and its up-regulation inIgA nephropathy. Biochem Biophys Res Commun 286:1098–1106,2001
24. MIYATA T, INAGI R, NANGAKU M, et al: Overexpression of the serpinmegsin induces progressive mesangial cell proliferation and expan-sion. J Clin Invest 109:585–593, 2002
25. INAGI R, MIYATA T, NANGAKU M, et al: Transcriptional regulationof a mesangium-predominant gene, megsin. J Am Soc Nephrol13:2715–2722, 2002
26. INAGI R, NANGAKU M, MIYATA T, et al: Mesangial cell-predominantfunctional gene, megsin. Clin Exp Nephrol 7:87–92, 2003
27. INAGI R, NANGAKU M, USUDA N, et al: Novel serpinopathy in ratkidney and pancreas induced by overexpression of megsin. J AmSoc Nephrol 16:1339–1349, 2005
28. NANGAKU M, PIPPIN J, RICHARDSON CA, et al: Beneficial effectsof systemic immunoglobulin in experimental membranous nephro-pathy. Kidney Int 50:2054–2062, 1996
29. SALANT DJ, DARBY C, COUSER WG: Experimental membranousglomerulonephritis in rats. J Clin Invest 66:71–81, 1980
30. KITAO Y, OZAWA K, MIYAZAKI M, et al: Expression of 150 kDa oxy-gen regulated protein (ORP150), a molecular chaperone in the en-doplasmic reticulum, rescues hippocampal neurons from glutamatetoxicity. J Clin Invest 108:1439–145, 2001
31. FLOEGE J, ALPERS CE, SAGE EH, et al: Markers of complement-dependent and complement-independent glomerular visceral ep-ithelial cell injury in vivo. Expression of antiadhesive proteins andcytoskeletal changes. Lab Invest 67:486–97, 1992
32. ITOH J, KAWAI K, SERIZAWA A, et al: Three-dimensional imagingof hormone-secreting cells and their microvessel environment inestrogen-induced prolactinoma of the rat pituitary gland by confocallaser scanning microscopy. Appl Immunohistochem Mol Morphol9:364–370, 2001
33. CYBULSKY AV, TAKANO T, PAPILLON J, et al: Complement C5b-9 membrane attack complex increases expression of endoplasmicreticulum stress proteins in glomerular epithelial cells. J Biol Chem277:41342–41351, 2002
34. KUWABARA K, MATSUMOTO M, IKEDA J, et al: Purification andcharacterization of a novel stress protein, the 150-kDa oxygen-regulated protein (ORP150), from cultured rat astrocytes and itsexpression in ischemic mouse brain. J Biol Chem 271:5025–5032,1996
35. TSUKAMOTO Y, KUWABARA K, HIROTA S, et al: 150-kD oxygen-regulated protein is expressed in human atherosclerotic plaquesand allows mononuclear phagocytes to withstand cellular stress onexposure to hypoxia and modified low density lipoprotein. J ClinInvest 98:1930–1941, 1996
36. ROLL DE, MURPHY BJ, LADEROUTE KR, et al: Oxygen regulated 80kDa protein and glucose regulated 78 kDa protein are identical.Mol Cell Biochem 103:141–148, 1991
37. BANDO Y, OGAWA S, YAMAUCHI A, et al: 150-kDa oxygen-regulatedprotein (ORP150) functions as a novel molecular chaperone inMDCK cells. Am J Physiol Cell Physiol 278:C1172–1182, 2000
38. TRUONG LD, FARHOOD A, TASBY J, et al: Experimental chronic re-nal ischemia: Morphologic and immunologic studies. Kidney Int41:1676–1689, 1992
39. BANDO Y, TSUKAMOTO Y, KATAYAMA T, et al: ORP150/HSP12Aprotects renal tubular epithelium from ischemia-induced cell death.FASEB J 18:1401–1403, 2004
40. MASSA SM, LONGO FM, ZUO J, et al: Cloning of rat grp75, an hsp70-family member, and its expression in normal and ischemic brain. JNeuro Res 40:807–819, 1995
41. LITTLE E, RAMAKRISHNAN M, ROY B, et al: The glucose-regulatedproteins (GRP78 and GRP94): Functions, gene regulation, and ap-plications. Crit Rev Eukaryot Gene Expr 4:1–18, 1994
42. PERLMUTTER DH: Liver injury in alpha1-antitrypsin deficiency:An aggregated protein induces mitochondrial injury. J Clin Invest110:1579–1583, 2002
43. DAVIS RL, SHRIMPTON AE, HOLOHAN PD, et al: Familial dementiacaused by polymerization of mutant neuroserpin. Nature 401:376–379, 1999
44. WANG J, TAKEUCHI T, TANAKA S, et al: A mutation in the insulin 2gene induces diabetes with severe pancreatic beta-cell dysfunctionin the Mody mouse. J Clin Invest 103:27–37, 1999
45. KAUFMAN RJ: Orchestrating the unfolded protein response in healthand disease. J Clin Invest 110:1389–1398, 2002
46. NANGAKU M, MIYATA T, SUZUKI D, et al: Cloning of rodent megsinrevealed its up-regulation in mesangioproliferative nephritis. Kid-ney Int 60:641–652, 2001
47. KOZUTSUMI Y, SEGAL M, NORMINGTON K, et al: The presenceof malfolded proteins in the endoplasmic reticulum signalsthe induction of glucose-regulated proteins. Nature 332:462–464,1988
48. SHIU RP, POUYSSEGUR J, PASTAN I: Glucose depletion accounts forthe induction of two transformation-sensitive membrane proteinsin Rous Sarcoma virus-transformed chick embryo fibroblasts. ProcNatl Acad Sci USA 74:3840–3844, 1977