intracellular transport and maturation pathway of human ... · the maturation pathway of human...
TRANSCRIPT

vebpttspappiaGrapaE1
SF
Virology 257, 460–471 (1999)Article ID viro.1999.9699, available online at http://www.idealibrary.com on
0CA
Intracellular Transport and Maturation Pathway of Human Herpesvirus 6
Maria Rosaria Torrisi,*,†,1 Massimo Gentile,* Giorgia Cardinali,*,† Mara Cirone,* Claudia Zompetta,*Lavinia V. Lotti,* Luigi Frati,*,‡ and Alberto Faggioni*
*Dipartimento di Medicina Sperimentale e Patologia, Universita di Roma La Sapienza, 00161 Rome; †Istituto Dermatologico San Gallicano,00153, Rome; and ‡Istituto Neurologico Mediterraneo Neuromed, 86077 Pozzilli, Italy
Received November 10, 1998; returned to author for revision January 21, 1999; accepted March 10, 1999
A peculiar characteristic of cells infected with human herpesvirus 6 (HHV6) is the absence of viral glycoproteins on theplasma membrane, which may reflect an atypical intracellular transport of the virions and/or the viral glycoproteins, differentfrom that of the other members of the herpesvirus family. To investigate the maturation pathway of HHV-6 in the human Tlymphoid cell line HSB-2, we used lectin cytochemistry and immunogold labeling combined with several electron micro-scopical techniques, such as ultrathin frozen sections, postembedding, and fracture-label. Immunolabeling with anti-gp116and anti-gp82-gp105 monoclonal antibodies revealed that the viral glycoproteins are undetectable on nuclear membranesand that at the inner nuclear membrane nucleocapsids acquire a primary envelope lacking viral glycoproteins. Afterde-envelopment, cytoplasmic nucleocapsids acquire a thick tegument and a secondary envelope with viral glycoproteins atthe level of neo-formed annulate lamellae or at the cis-side of the Golgi complex. Cytochemical labeling using helix pomatialectin revealed that the newly acquired secondary viral envelopes contain intermediate forms of glycocomponents, suggest-ing a sequential glycosylation of the virions during their transit through the Golgi area before their final release into theextracellular space. Immunogold labeling also showed that the viral glycoproteins, which are not involved in the buddingprocess, reach and accumulate in the endosomal/lysosomal compartment. Pulse-chase analysis indicated degradation of thegp116, consistent with its endosomal localization and with the absence of viral glycoproteins on the cell surface of theinfected cells. © 1999 Academic Press
1ntmecozcWZTta
tlecoGtappat
INTRODUCTION
Herpesviruses are large DNA-containing envelopediruses that replicate in the nucleus and acquire theirnvelope through budding at the inner nuclear mem-rane into the perinuclear space (Roizman, 1996). Theathways followed by either virions and viral glycopro-
eins during their maturation and intracellular transporto the plasma membranes along the exocytic route aretill controversial: the viral envelope glycoproteins arerocessed to their mature forms by the addition of sug-rs during the intracellular transit, and glycosylation ap-ears necessary for the exit of the virus from the cells. Aossible model proposes that the virions are assembled
n a single budding event at the inner nuclear membranend transported inside vesicles and vacuoles to theolgi complex and then to the cell surface for final
elease by exocytosis to the extracellular space. Thebove mechanism was first suggested for herpes sim-lex type 1 (HSV-1) (Roizman and Furlong, 1974; Johnsonnd Spear, 1982); our subsequent results on HSV-1 andpstein–Barr virus (EBV) infected cells (Torrisi et al.,989, 1992; Campadelli et al., 1993; Di Lazzaro et al.,
1 To whom reprint requests should be addressed at Dip. Medicinaperimentale e Patologia, Viale Regina Elena 324, 00161 Roma, Italy.ax: 396–4468450 or 4452850. E-mail: [email protected].
042-6822/99 $30.00opyright © 1999 by Academic Pressll rights of reproduction in any form reserved.
460
995) were also consistent with this pathway. An alter-ative model, which involves an initial envelopment at
he inner nuclear membrane, followed by de-envelop-ent by fusion with the outer nuclear membrane or with
ndoplasmic reticulum cisternae, and re-envelopment ofytoplasmic naked nucleocapsids at the trans cisternaef the Golgi complex has been described for varicella-oster virus (VZV), pseudorabies virus (PrV) and humanytomegalovirus (HCMV) (Jones and Grose, 1988;healy et al., 1991; Card et al., 1993; Gershon et al., 1994;
hu et al., 1995; Radsak et al., 1996; Granzow et al., 1997).his mechanism has been also proposed by some au-
hors for HSV-1 and EBV (Gong and Kieff, 1990; Browne etl., 1996).
Among human herpesviruses, little is known regardinghe maturation pathway of human herpesvirus 6, a T-ymphotropic herpesvirus that is the etiological agent ofxanthem subitum and is highly suspected to play aofactorial role in the etiopathogenesis and progressionf acquired immunodeficiency syndrome (Lusso andallo, 1995). Few reports have described the ultrastruc-
ure characteristics of HHV-6 (Biberfeld et al., 1987; Nii etl., 1990; Roffman et al., 1990; Cirone et al., 1992). Mor-hologically, HHV-6 is characterized by the presence of arominent tegument, which has been proposed to becquired in a specialized region of the nuclear area of
he infected cell (Roffman et al., 1990). In addition, HHV-

6iviwltvaHtfHn1oaOtta
H
cftcmcsmootln(1wtosletwdavseoc
enpstaap
ccoitortccmcsoioa3o3bcmcofcrsqagtifa
IH
ppmrit
461HHV-6 INTRACELLULAR TRANSPORT
-infected cells display at least two peculiar character-stics that we have recently described: the absence ofiral glycoproteins over the plasma membrane of virus-
nfected cells (Biberfeld et al., 1987; Cirone et al., 1994),hich might reflect an atypical pathway of viral intracel-
ular transport different from that of all other members ofhe herpesvirus family, and the de-novo induction, uponiral infection, of cytoplasmic annulate lamellae, whichppear densely labeled with antibodies directed againstHV-6 glycoproteins (Cardinali et al., 1998). Thus to fur-
her approach the issue of HHV-6 maturation, we per-ormed an ultrastructural analysis of HHV-6 infected
SB-2 cells, using an immunoelectron microscopic tech-ique, called fracture-label (Torrisi and Pinto da Silva,984; Torrisi and Mancini, 1996), which we have previ-usly used to study the maturation pathways of HSV-1nd EBV (Torrisi et al., 1989, 1992; Di Lazzaro et al., 1995).ther approaches, such as gold immunolabeling of ul-
rathin frozen sections or biochemical assays to studyhe fate of viral glycoproteins, were also consistent with
novel mechanism of viral egress.
RESULTS
HV-6 envelopment and tegument acquisition
Morphological analysis of HHV-6 maturation has beenarried out on conventional thin sections of HHV-6-in-
ected HSB2 cells. Ultrastructural examination revealedhe presence of numerous intranuclear naked nucleo-apsids (Figs. 1a, 1d, and 1e, large arrows). Envelop-ent of the nucleocapsids by budding at the inner nu-
lear membranes was also frequently observed (nothown), and enveloped virions lacking a visible tegu-ent were present in the space between the inner and
uter nuclear membranes (Fig. 1a, small arrow). A sec-nd type of naked nucleocapsids, characterized by a
hick tegument, was evident in the cell cytoplasm andocalized mainly in close proximity or inside stacks ofeoformed (Cardinali et al., 1998) annulate lamellae (AL)
Fig. 1c, arrowheads) and in the Golgi area (Figs. 1e andf, arrowheads). Enveloped virions with a thick tegumentere contained in vesicles either surrounded by AL cis-
ernae (Figs. 1b and 1c, small arrows) or in proximity andccasionally in continuity with Golgi cisternae (Fig. 1d,mall arrows). Closer inspection of the enveloped virions
ocated in the perinuclear space compared with thenveloped virions located in cytoplasmic vesicles during
heir transit to the plasma membrane and compared alsoith extracellular enveloped virions clearly show strikingifferences in size and in the presence of a tegumentnd of spike projections on the envelope: envelopedirions in the intranuclear space (Fig. 2a) were in factmaller in size (130 nm), lacking tegument, and theirnvelope appeared devoid of spikes, whereas envel-ped virions in cytoplasmic locations, such as thoseontained in vesicles surrounded by AL (Fig. 2b), and
xtracellular virions (Fig. 2c) were larger in size (170–180m) and showed a thick tegument and numerous spikesrojecting from their envelopes. The presence or ab-ence of visible tegument and spikes seems to suggest
hat the two types of enveloped virions are assembled byt least two different budding events, the first occurringt the inner nuclear membrane and the second in cyto-lasmic locations such as AL or Golgi cisternae.
To verify if the two envelopes differ in viral glycoproteinontent, we performed an immunoelectron microscopi-al analysis on ultrathin frozen sections using a mixturef anti-gp116 and anti-gp82-gp105 monoclonal antibod-
es directed against the two major envelope glycopro-eins of HHV-6. No gold immunolabeling was observedver the cell plasma membranes (Fig. 3d), as previously
eported (Cirone et al., 1994). To rule out the possibilityhat absence of labeling over the plasma membraneould be a generalized phenomenon, affecting other hostell glycoproteins, analysis of the expression of CD5embrane antigen on HHV-6-infected HSB-2 cells was
arried out by immunofluorescence. Full-ring-membranetaining by Leu-1 mAb was equally detectable in ;70%f infected and uninfected cells (data not shown). Gold
mmunolabeling of gp116 and gp82-gp105 was densever extracellular virions (Figs. 3c and 3d, large arrows)nd on enveloped virions in cytoplasmic vacuoles (Figs.a and 3d, small arrows). The membranes of the vacu-les (Figs. 3a, 3b, and 3d) and of the Golgi cisternae (Fig.b) appeared also strongly labeled. The nuclear mem-ranes (Figs. 3a and 3e) and the enveloped virions lo-ated in the space between the inner and outer nuclearembranes (Fig. 3e) were virtually unlabeled. Only oc-
asionally (;7% of the total intracellular enveloped viri-ns, as assessed by random analysis of 50 ultrathin
rozen sections) did virions inside vesicles in the perinu-lear area also appear unlabeled (Fig. 3b). Totally similaresults were obtained using an HHV-6-positive humanerum (data not shown). Thus the viral envelopes ac-uired by budding at the inner nuclear membrane do notppear to contain major viral glycoproteins, such asp116 and gp82-gp105, which in contrast are present on
he envelopes of extracellular virions or of virions locatedn peripheral cytoplasmic structures. This finding con-irms the morphological observation of the presence orbsence of spikes on the virions.
ntracellular transit and sequential glycosylation ofHV-6-enveloped virions
Lectin cytochemistry has been widely applied in theast in combination with fracture-label technique, whichrovides full accessibility to the labeling of intracellularembranes exposed by the freeze-fracture process (Tor-
isi and Pinto da Silva, 1984). Recently lectin cytochem-stry in the fracture-label method has been used to de-ermine the glycosylation stages of HSV-1 envelope com-

462 TORRISI ET AL.

ptmiLsolfFslbtugcwsHpbVlbt
vvlp14c1clos(Tipraot
Ig
g
HmscaN
swem
463HHV-6 INTRACELLULAR TRANSPORT
onents during the intracellular transit of the virions inhe infected cells and to demonstrate a progressive viral
aturation occurring by sequential transport through ornteraction with Golgi cisternae (Torrisi et al., 1992; Diazzaro et al., 1995). We decided therefore to use theame approach to investigate the extent of glycosylationf HHV-6 envelope components in different intracellular
ocations. HSB-2 cells, infected with HHV-6, were fixed,reeze-fractured, and labeled with lectin-gold conjugates.racture-labeled samples were then processed for thinection electron microscopy (for a review on the fracture-
abel method, see Torrisi and Mancini, 1996). Lectin la-eling was also performed on resin-embedded thin-sec-
ioned cells. Helix pomatia lectin, which binds terminalnsubstituted GalNAc, recognizes intermediate forms oflycoconjugates after O-linked addition of the sugar inis-Golgi cisternae (Roth, 1984). Wheat germ agglutinin,hich is specific for sialic acid, binds terminally glyco-
ylated components (Bhavandan and Kalic, 1979). InSB-2 cells, infected with HHV-6, the enveloped virionsresent in the perinuclear space appeared negative foroth HPL (Fig. 4b, arrowhead) and WGA (not shown).irions located inside vesicles surrounded by annulate
amellae were positively labeled by HPL (Fig. 4b, arrows)ut unlabeled by WGA (Fig. 4a). Enveloped virions posi-
ive for HPL were also observed in vesicles or larger
FIG. 1. Morphological analysis of the intracellular maturation of HHSB-2 cells, intranuclear naked nucleocapsids are frequently visible (aembranes can be found in the perinuclear space (a, small arrow). C
tacks of annulate lamellae (c, arrowheads) and in the Golgi complex arharacterized by a thick tegument, which is absent on the intranuclearre seen either surrounded by annulate lamellae (b and c, small arrowsu, nucleus; NM, nuclear membrane; G, Golgi complex; M, mitochond
FIG. 2. Ultrastructural examination of enveloped virions in different locpace possess an envelope smooth and devoid of surface projections. Theith the extracellular viral particles. (b) Virions contained within vesiclesnvelope. A thick tegument is visible and the size of the virions is comparembrane are characterized by an envelope enriched of clearly distingui
acuoles in the Golgi area (Fig. 4c, arrows). Extracellularirions appeared unlabeled by HPL (Fig. 4d) but densely
abeled by WGA (Fig. 4e). HPL-gold particles were alsoresent on AL cisternae (Fig. 4b and Cardinali et al.,998) as well as on Golgi membranes and vacuoles (Fig.c). Quantitation of HPL-gold labeling, determined byounting the number of gold particles per mm2 6 SEM in0 images of AL areas and of virions surrounded by AL,ompared with cytosolic areas for background aspecific
abeling assessment, revealed that density of labelingver the virions (300.71 6 33.6 gold particles/mm2) wasignificantly higher (P , 0.05) than that over AL areas
160.14 6 17.34) and over cytosolic areas (11.9 6 1.32).hus enveloped virions carrying glycoconjugates at an
ntermediate step of glycosylation, as assessed by HPLositive and WGA negative labeling, are detectable al-
eady in close proximity or inside the AL network as wells the Golgi complex, implying progressive glycosylationf envelope components during the intracellular transit of
he virions through the Golgi area.
ntracellular fate and degradation of viral envelopelycoproteins in infected cells
To analyze the intracellular route of traffic of the virallycoproteins not involved in the envelopment process
HSB-2-infected cells. In conventional thin sections of HHV-6-infectedd e, large arrows) and enveloped virions budded at the inner nuclearmic naked nucleocapsids are also evident in proximity of prominent
nd f, arrowheads). These cytoplasmic nucleocapsids (arrowheads) arecapsids (large arrows). Enveloped virions inside cytoplasmic vesiclessociated with Golgi cisternae (d, small arrows). AL, annulate lamellae;ars: 0.5 mm.
during their intracellular transport. (a) Virions located in the intranucleartegument is evident and the size of the virions appears smaller comparednded by annulate lamellae show numerous spikes projecting from thethat of extracellular particles. (c) Extracellular virions close to the plasmaspikes. PM, plasma membrane. Bars: 0.1 mm.
V-6 in, d, anytoplasea (e anucleo) or asrium. B
ationslack ofsurrou
able toshable

affwa
brbr(
Sovpm
464 TORRISI ET AL.
nd to follow their final destiny inside the cell, we per-ormed immunolabeling with anti-gp116 mAb on freeze-ractured infected cells. Dense gold labeling of gp116
as observed on AL cisternae (Fig. 5a and Cardinali etl., 1998), Golgi membranes (Fig. 5d) and on the mem-
FIG. 3. Immunogold labeling of gp116 and gp82-gp105 viral envelections were incubated with a mixture of anti-gp116 and anti-gp82-gpn virions located inside intracellular vacuoles (a and d, small arrows)acuoles, which contain the gold decorated virions (a, b, and d), and therinuclear space (e) appear unlabeled. Occasionally, virions inside vesembranes (a and e) and plasma membranes (d) are virtually unlabel
ranes of large peripheral vacuoles (Fig. 5c); again (Ci-one et al., 1994; Cardinali et al., 1998), nuclear mem-ranes and endoplasmic reticulum cisternae (not shown)
evealed none or little labeling, and plasma membranesFig. 5c) appeared virtually unlabeled.
coproteins on ultrathin cryosections of HHV-6-infected HSB-2 cells.Abs, followed by protein A-colloidal gold. Dense labeling is observedthe extracellular space (c and d, large arrows). The membranes of thernae of the Golgi complex (b) are also strongly labeled. Virions in the
the perinuclear area (b, arrowheads) also appear unlabeled. Nuclearacuoles. Bars: a, b, and d: 0.5 mm; c and e: 0.1 mm.
ope gly105 m
and ine cisteicles in
ed. V, v

vpluHa
465HHV-6 INTRACELLULAR TRANSPORT
FIG. 4. Lectin labeling of HHV-6-infected HSB-2 cells: (a) WGA-gold (18-nm gold particles) labeling of freeze-fractured cells showed that theirions (arrow) inside vesicles surrounded by annulate lamellae are unlabeled. (b and c) Labeling with HPL-gold conjugates (10-nm goldarticles) on resin-embedded, thin-sectioned cells reveals that several enveloped virions (arrows) inside vesicles or vacuoles are positively
abeled, either when located in the area of annulate lamellae (b) or in the Golgi area (c), whereas virions in the perinuclear space appearnlabeled (b, arrowhead). Annulate lamellae (b), cisternae of the Golgi complex (c), and intracellular vacuoles in the Golgi area (c) arePL-positively labeled. (d and e) Extracellular virions and the cell plasma membranes are negative for HPL (d) and positive for WGA (e). Bars:, 0.1 mm; b–e, 0.5 mm.

olmi
ghlvc
pgieIHi
466 TORRISI ET AL.
The intermediate compartment, morphologically rec-gnizable as a tubulo-vesicular structure, was also un-
abeled (Fig. 5b). Therefore viral glycoproteins wereostly present on AL, Golgi cisternae, and vacuoles. To
dentify these vacuoles containing high amount of viral
FIG. 5. (a–c) Immunolabeling of gp116 on freeze-fractured HHV-6-infeeripheral vacuoles (c), whereas intermediate compartment (b) and plasmaolds) and human cathepsin D (small golds): dense labeling for gp116 i
dentified by their positive immunoreaction for human cathepsin D (e, f, andndosomal structures. When exposed to the labeling, virions inside transp
n (e), a tegumented naked capsid is visible in the cell cytoplasm (arrowheaHV-6-infected HSB-2 cells: enveloped virions (arrows) can be frequen
ntermediate compartment; E, endosomes; Ly, lysosomes. Bars: a–c, e, f,
lycoproteins, we performed double-labeling with anti-uman cathepsin D, a marker of late endosomes and
ysosomes; the gold labeling was associated with theseacuolar structures (Figs. 5e–5h) and allowed us to con-lude that gp116 reach and accumulate in late endoso-
B-2 cells: dense gold labeling is present on annulate lamellae (a) andranes (c) appear unlabeled. (d–g) Double immunolabeling of gp116 (largent on Golgi complexes (d) and observed in late endosomal structures,
ons are frequently seen in close proximity to (f, arrows) or inside (h) theseles (g) and in endosomes are strongly labeled by anti-gp116 antibody (h).
d k) Anti-cathepsin D immunolabeling on resin-embedded, thin- sectionednd inside densely labeled endosomal or prelysosomal structures. IC,: 0.5 mm; d, g, and h: 0.1 mm.
cted HSmemb
s preseh). Viri
ort vesicd). (i antly fou
and i–k
mdpqemobfmd
paidAta
nomiespsw
wmaomtcifa
eo(etcsebntipsmmslcpct
mmptocieegfcostavmv
st
pca
467HHV-6 INTRACELLULAR TRANSPORT
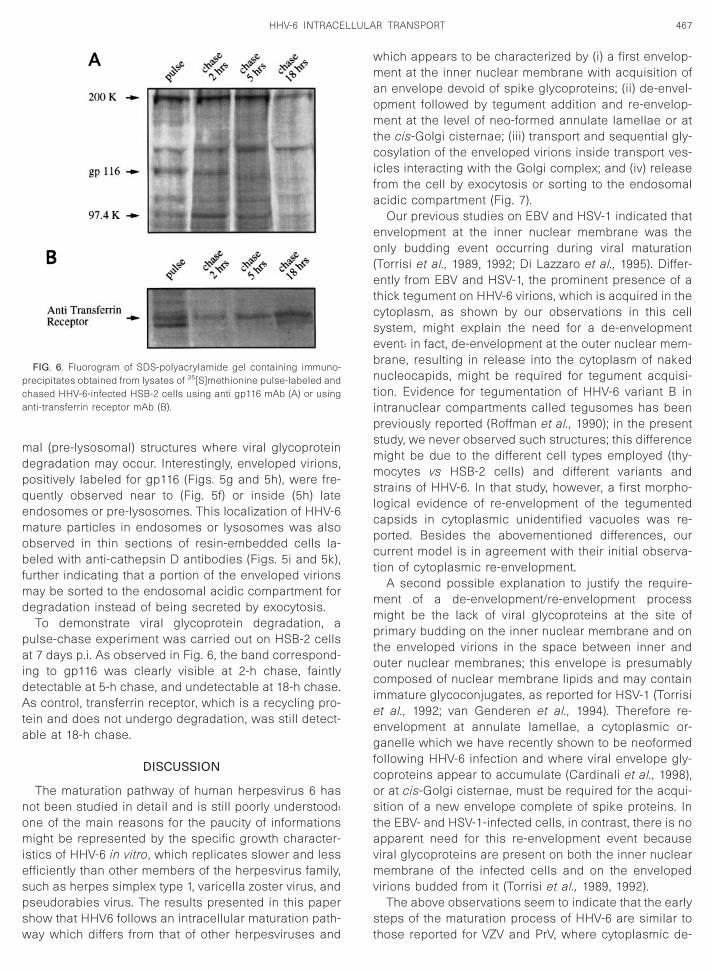
al (pre-lysosomal) structures where viral glycoproteinegradation may occur. Interestingly, enveloped virions,ositively labeled for gp116 (Figs. 5g and 5h), were fre-uently observed near to (Fig. 5f) or inside (5h) latendosomes or pre-lysosomes. This localization of HHV-6ature particles in endosomes or lysosomes was also
bserved in thin sections of resin-embedded cells la-eled with anti-cathepsin D antibodies (Figs. 5i and 5k),
urther indicating that a portion of the enveloped virionsay be sorted to the endosomal acidic compartment for
egradation instead of being secreted by exocytosis.To demonstrate viral glycoprotein degradation, a
ulse-chase experiment was carried out on HSB-2 cellst 7 days p.i. As observed in Fig. 6, the band correspond-
ng to gp116 was clearly visible at 2-h chase, faintlyetectable at 5-h chase, and undetectable at 18-h chase.s control, transferrin receptor, which is a recycling pro-
ein and does not undergo degradation, was still detect-ble at 18-h chase.
DISCUSSION
The maturation pathway of human herpesvirus 6 hasot been studied in detail and is still poorly understood:ne of the main reasons for the paucity of informationsight be represented by the specific growth character-
stics of HHV-6 in vitro, which replicates slower and lessfficiently than other members of the herpesvirus family,uch as herpes simplex type 1, varicella zoster virus, andseudorabies virus. The results presented in this paperhow that HHV6 follows an intracellular maturation path-ay which differs from that of other herpesviruses and
FIG. 6. Fluorogram of SDS-polyacrylamide gel containing immuno-recipitates obtained from lysates of 35[S]methionine pulse-labeled andhased HHV-6-infected HSB-2 cells using anti gp116 mAb (A) or usingnti-transferrin receptor mAb (B).
hich appears to be characterized by (i) a first envelop-ent at the inner nuclear membrane with acquisition of
n envelope devoid of spike glycoproteins; (ii) de-envel-pment followed by tegument addition and re-envelop-ent at the level of neo-formed annulate lamellae or at
he cis-Golgi cisternae; (iii) transport and sequential gly-osylation of the enveloped virions inside transport ves-
cles interacting with the Golgi complex; and (iv) releaserom the cell by exocytosis or sorting to the endosomalcidic compartment (Fig. 7).
Our previous studies on EBV and HSV-1 indicated thatnvelopment at the inner nuclear membrane was thenly budding event occurring during viral maturation
Torrisi et al., 1989, 1992; Di Lazzaro et al., 1995). Differ-ntly from EBV and HSV-1, the prominent presence of a
hick tegument on HHV-6 virions, which is acquired in theytoplasm, as shown by our observations in this cellystem, might explain the need for a de-envelopmentvent: in fact, de-envelopment at the outer nuclear mem-rane, resulting in release into the cytoplasm of nakeducleocapids, might be required for tegument acquisi-
ion. Evidence for tegumentation of HHV-6 variant B inntranuclear compartments called tegusomes has beenreviously reported (Roffman et al., 1990); in the presenttudy, we never observed such structures; this differenceight be due to the different cell types employed (thy-ocytes vs HSB-2 cells) and different variants and
trains of HHV-6. In that study, however, a first morpho-ogical evidence of re-envelopment of the tegumentedapsids in cytoplasmic unidentified vacuoles was re-orted. Besides the abovementioned differences, oururrent model is in agreement with their initial observa-
ion of cytoplasmic re-envelopment.A second possible explanation to justify the require-
ent of a de-envelopment/re-envelopment processight be the lack of viral glycoproteins at the site of
rimary budding on the inner nuclear membrane and onhe enveloped virions in the space between inner anduter nuclear membranes; this envelope is presumablyomposed of nuclear membrane lipids and may contain
mmature glycoconjugates, as reported for HSV-1 (Torrisit al., 1992; van Genderen et al., 1994). Therefore re-nvelopment at annulate lamellae, a cytoplasmic or-anelle which we have recently shown to be neoformed
ollowing HHV-6 infection and where viral envelope gly-oproteins appear to accumulate (Cardinali et al., 1998),r at cis-Golgi cisternae, must be required for the acqui-ition of a new envelope complete of spike proteins. In
he EBV- and HSV-1-infected cells, in contrast, there is nopparent need for this re-envelopment event becauseiral glycoproteins are present on both the inner nuclearembrane of the infected cells and on the enveloped
irions budded from it (Torrisi et al., 1989, 1992).The above observations seem to indicate that the early
teps of the maturation process of HHV-6 are similar tohose reported for VZV and PrV, where cytoplasmic de-

amrmeGG1fatoclppH
se
matpsics(esvd1tpsfat
468 TORRISI ET AL.
nd re-envelopment have been described. However, aajor difference with the above proposed models is
epresented by the intracellular site where re-envelop-ent seems to occur: whereas for VZV and PrV re-
nvelopment at post-Golgi compartments, such as trans-olgi network, has been described (Whealy et al., 1991;ershon et al., 1994; Harson and Grose, 1995; Zhu et al.,
995; Granzow et al., 1997), our observations are inavour of a re-envelopment occurring at the level ofnnulate lamellae or at the cis-Golgi cisternae. In fact,
he detection of intermediate forms of glycocomponentsn enveloped virions, as revealed by our lectin cyto-hemical analysis, suggests that the HHV-6 virion enve-
ope undergoes a progressive maturation of glycocom-onents by sequential interaction with the Golgi com-lex. This is in agreement with our previous results withSV-1 (Di Lazzaro et al., 1995).Finally, whereas most of the mature enveloped virions
eem to exit the cell by exocytosis to be released in thextracellular space, the double-immunolabeling experi-
FIG. 7. Proposed model for the pathway of intracel
ents with anti-cathepsin D and with anti-gp116 mAbs,s well as the pulse–chase analysis, strongly indicate
hat the viral glycoproteins not involved in the buddingrocess accumulate in late endosomal or pre-lysosomaltructures where degradation might occur. These find-
ngs may account for the previously described peculiarharacteristic of HHV-6-infected cells, i.e., the virtual ab-ence of viral glycoproteins on the plasma membrane
Cirone et al., 1994). Previously reported time-coursexperiments on the expression of HHV-6 gp116 demon-trated that also at early time points after viral infectioniral gps appeared only intracellularly and could notetected over the cell plasma membrane (Cardinali et al.,998 and data not shown). Although we cannot excludehat their absence on the plasma membrane and theirresence in the endosomal compartment might be con-equent to their rapid internalization from the cell sur-
ace, as described for the varicella zoster virus gE (Olsonnd Grose, 1997), this possibility appears unlikely due to
he absence of endocytic signals in gp116 cytoplasmic
nsport and maturation of HHV-6 in infected cells.
lular tra
tattaor
C
p(iBftwmab4pwac
U
7lr3ftmgalBgMtacdlmCttcPsb
P
tdsHCastwcUacsa
F
tffwwlc(fifmCnccpftogaw4wntpI
P
fw
469HHV-6 INTRACELLULAR TRANSPORT
ail. Furthermore because complete enveloped virionsre also frequently observed, along with viral glycopro-
eins, inside endosomal-lysosomal structures, similarlyo what has been recently reported for HSV-1 (Brunetti etl., 1998), the significance of a possible divergent sortingf some virions to the endosomal compartment for deg-
adation remains to be clarified.
MATERIALS AND METHODS
ells and infection
HSB-2 cells were cultured in RPMI 1640 medium sup-lemented with 10% FCS plus antibiotics. The GS strain
Ablashi et al., 1991) of HHV-6 (variant A) was employedn this investigation and was propagated in HSB-2 cells.riefly, the virus stock (titer 105 TCID50) was obtained
rom 7 days supernatant of infected cells, when .80% ofhe cells showed cytopathic effect. Cell-free culture fluid
as harvested and filtered through a 0.45-mm filter andade into pellets by centrifugation at 25,000 g for 90 min
t 4°C. For infection, 5 3 106 pelleted cells were incu-ated with an appropriate dilution of the virus stock. After
h at 37°C, the cells were washed once and resus-ended in complete medium. For all experiments, cellsere collected after 7 days p.i., when .80% of the cellsppeared infected. Uninfected HSB-2 cells were used asontrols.
ltrathin cryosections
Cells were fixed with 8% paraformaldehyde in PBS pH.4 for 2 h at 4°C, washed, and embedded in 2% agaroseow melting point (LMP) that was solidified on ice. Aga-ose blocks were infused with 2.3 M sucrose in PBS for
h at 4°C, frozen in liquid nitrogen and cryosectionedollowing the method described by Tokuyasu (1973). Ul-rathin cryosections were collected using sucrose and
ethyl cellulose and incubated with a mixture of anti-p82 and anti-gp116 mAbs (Virotech, Rockville, MD). Thentibody against gp82-gp105 binds to linear epitopes,
ocated in the N-terminal end (Pfeiffer et al., 1993, 1995;. Chandran, personal communication). The mAb againstp-116, which corresponds to HHV-6 gB (Chou andarousek, 1992; Ellinger et al., 1993) binds to conforma-
ional epitopes (B. Chandran, personal communication)nd, as shown by immunoprecipitation, recognizes pre-ursor as well as mature forms of the protein (Balachan-ran et al., 1989; our data, not shown). All samples were
abeled with colloidal gold (prepared by the citrateethod) conjugated with protein A (Pharmacia Finehemicals, Uppsala, Sweden) for 3 h at 4°C. Alterna-
ively ultrathin cryosections were incubated with a highitered human serum. All sections were then labeled witholloidal gold (10 nm) conjugated with protein A (1:10 inBS). Finally, ultrathin cryosections were stained with aolution of 2% methyl cellulose and 0.4% uranyl acetateefore EM examination.
ostembedding
HHV6-infected HSB-2 cells were fixed with 0.5% glu-araldehyde in PBS pH 7.4 for 1 h at 4°C, partially dehy-rated in ethanol and embedded in LR White resin. Thinections were collected on nickel grids, labeled withPL colloidal gold (10 nm) conjugates (Sigma Chemicalo., St. Louis, MO) (1:5 in Tris buffer 0.15 M NaCl, 0.5%lbumin, 0.05% Tween 20) for 1 h at 37°C. Control thinections were pre-incubated in 100 mM N-acetyl-galac-
osamine (GalNAc) for 30 min at 37°C. Thin sectionsere also labeled with anti-human cathepsin D poly-
lonal antibodies (kindly provided by Dr. Ciro Isidoro,niversity of Torino, Italy) (1:50 in PBS) for 1 h at 25°Cnd protein A-colloidal gold (18 nm prepared by theitrate method) for 30 min at 25°C. All sections weretained with uranyl acetate and lead citrate before ex-mination with EM.
racture-label
HHV6-infected HSB-2 cells were fixed with 0.5% glu-araldehyde in phosphate-buffered saline (PBS) pH 7.4or 1 h at 4°C, impregnated with 30% glycerol in PBS, androzen in Freon 22 cooled by liquid nitrogen. Frozen cells
ere fractured in liquid nitrogen by repeated crushingith a glass pestle and gradually deglycerinated. For
ectin labeling, fractured cells were incubated with HPLolloidal gold (10 nm) conjugates (Sigma Chemical Co.)
1:5 in PBS 0.15 M NaCl, 0.5% albumin, 0.05% Tween 20)or 1 h at 37°C. Control experiments were pre-incubatedn 100 mM GalNAc for 30 min at 37°C. Alternatively,reeze-fractured cells were incubated in a solution of 1
g/ml of wheat germ agglutinin (WGA, Sigma Chemicalo.) in O.1 M Sorensen’s phosphate buffer-4% polyvi-ylpyrrolidone, pH 7.4, for 1 h at 37°C and labeled witholloidal gold (18 nm prepared by the citrate method)onjugated with ovomucoid for 3 h at 4°C. Control sam-les were preincubated in 0.4 M N-acetyl-D-glucosamine
or 15 min at 37°C, treated with WGA in the presence ofhe competitor sugar for 1 h at 37°C and labeled withvomucoid-coated colloidal gold as above. For immuno-old labeling, fractured samples were incubated withnti-gp116 mAb (1:20 in PBS) for 1 h at 25°C and labeledith colloidal gold conjugated with protein A for 3 h at
°C. In double-labeling experiments, fractured samplesere treated with anti-gp116 mAb and protein A-gold (18m) as above and then incubated with anti-human ca-
hepsin D polyclonal antibodies (1:50 in PBS) followed byrotein A-colloidal gold conjugates (10 nm; British Biocell
nt., Cardiff, UK; 1:10 in PBS, for 30 min at 25°C).
rocessing for electron microscopy
Cells and fracture-labeled samples were processedor thin section electron microscopy as follows: postfixed
ith 1% osmium tetroxide, stained with uranyl acetate (5

m8sbeadam
P
mqwpcwi(bdigr
DLsRfF
A
B
B
B
B
B
C
C
C
C
C
C
D
E
G
G
G
H
J
J
L
N
O
P
P
R
470 TORRISI ET AL.
g/ml), dehydrated in acetone, and embedded in Epon12. In some experiments, samples were additionallytained en bloc with 0.1% tannic acid in Veronal acetateuffer, pH 7.4, for 30 min at 25°C. Thin sections werexamined unstained or poststained with uranyl acetatend lead hydroxide. Density of the immunogold labeling,etermined as gold particles/mm2 and the statisticalnalysis were evaluated using a Sigma Scan Measure-ent System (Jandel Scientific, Corte Madera, CA).
ulse–chase analysis
HHV-6-infected HSB-2 cells (7 days p.i.) were kept inethionine-free RPMI 1640 for 30 min at 37°C. Subse-
uently the cells were pulse-labeled for 30 min at 37°Cith 100 mCi/ml of 35[S]methionine. After labeling, oneortion of the cells was frozen (pulse). The remainingells were washed twice in RPMI 1640 supplementedith 10% FCS, suspended in the same medium and the
ncubation at 37°C was continued for 2, 5 and 18 hchase). Cells (3 3 106/point) were solubilized in RIPAuffer (0,01 M Tris–HCl pH 7.4, 0,15 M NaCl, 1% sodiumeoxycholate, 1% Triton X-100, 0.1% SDS, 1 mM PMSF),
mmunoprecipitated with monoclonal antibodies againstp 116 (Virotech International, Inc.) or against transferrin
eceptor (Becton Dickinson) and analyzed by SDS–PAGE.
ACKNOWLEDGMENTS
We thank Dr. C. Isidoro for the generous gift of anti-human cathepsinpolyclonal antibodies. We also thank Mr. Giuseppe Lucania and Ms.
ucia Cutini for excellent technical assistance. This work was partiallyupported by grants from MURST, from Associazione Italiana per laicerca sul Cancro (AIRC), from Ministero della Sanita, Progetto AIDS,
rom CNR (Target Project on Biotechnology) and from Istituto Pasteurondazione Cenci-Bolognetti, Universita di Roma La Sapienza.
REFERENCES
blashi, D. V., Balachandran, N., Josephs, S. F., Hung, C. L., Krueger,G. R., Kramarsky, B., Salahuddin, S. Z., and Gallo, R. C. (1991).Genomic polymorphism, growth properties, and immunologic varia-tions in human herpesvirus 6 isolates. Virology 184, 545–552.
alachandran, N., Amelse, R. E., Zhou, W. W., and Chang, C. K. (1989).Identification of proteins specific for human herpesvirus 6-infectedhuman T cells. J. Virol. 63, 2835–2840.
havandan, V. P., and A. W. Kalic. (1979). The interaction of wheat germagglutinin with sialoglycoproteins. The role of sialic acid. J. Biol.Chem. 254, 4000–4008.
iberfeld, P., Kramarsky, B., Salahuddin, S. Z., and Gallo, R. C. (1987).Ultrastructural characterization of a new human B lymphotropic DNAvirus (human herpesvirus 6) isolated from patients with lymphopro-liferative disease. J. Nat. Cancer Inst. 79, 933–939.
rowne, H., Bell, S., Minson, T., and Wilson, D. W. (1996). An endoplas-mic reticulum-retained herpes simplex virus glycoprotein H is absentfrom secreted virions: Evidence for reenvelopment during egress.J. Virol. 70, 4311–4316.
runetti, C. R., Dingwell, K. S., Wale, C., Graham, F. L., and Johnson,D. C. (1998). Herpes simplex virus gD and virions accumulate inendosomes by mannose 6-phosphate-dependent and -independentmechanisms. J. Virol. 72, 3330–3339.
ampadelli, G., Brandimarti, R., Di Lazzaro, C., Ward, P. L., Roizman, B.,
and Torrisi, M. R. (1993). Fragmentation and dispersal of Golgi pro-teins and redistribution of glycoproteins and glycolipids processedthrough the Golgi apparatus after infection with herpes simplex virus1. Proc. Natl. Acad. Sci. USA 90, 2798–2802.
ard, J. P., Rinaman, L., Lynn, R. B., Lee, B. H., Meade, R. P., Miselis,R. R., and Enquist, L. W. (1993). Pseudorabies virus infection of the ratnervous system: Ultrastructural characterization of viral replication,transport, and pathogenesis. J. Neurosci. 13, 2515–2539.
ardinali, G., Gentile, M., Cirone, M., Zompetta, C., Frati, L., Faggioni, A.,and Torrisi, M. R. (1998). Viral glycoproteins accumulate in newlyformed annulate lamellae following infection of lymphoid cells byhuman herpesvirus 6. J. Virol. 72, 9738–9746.
hou, S., and Marousek, G. I. (1992). Homology of the envelope glyco-protein B of human herpesvirus 6 and cytomegalovirus. Virology 191,523–528.
irone, M., Zompetta, C., Angeloni, A., Ablashi, D. V., Salahuddin, S. Z.,Pavan, A., Torrisi, M. R., Frati, L., and Faggioni, A. (1992). Infection byhuman herpesvirus 6 (HHV-6) of human lymphoid T cells occursthrough an endocytic pathway. AIDS Res. Hum. Retrov. 8, 2031–2037.
irone, M., Campadelli-Fiume, G., Foa-Tomasi, L., Torrisi, M. R., andFaggioni, A. (1994). Human herpesvirus 6 envelope glycoproteins Band H-L complex are undetectable on the plasma membrane ofinfected lymphocytes. AIDS Res. Hum. Retrov. 10, 175–179.
i Lazzaro, C., Campadelli-Fiume, G., and Torrisi, M. R. (1995). Inter-mediate forms of glycoconjugates are present in the envelope ofherpes simplex virions during their transport along the exocyticpathway. Virology 214, 619–623.
llinger, K., Neipel, F., Foa-Tomasi, L., Campadelli-Fiume, G., and Fleck-enstein, B. (1993). The glycoprotein B homologue of human herpes-virus 6. J. Gen. Virol., 74, 495–500.
ershon, A. A., Sherman, D. L., Zhu, Z., Gabel, C. A., Ambron, R. T., andGershon, M. D. (1994). Intracellular transport of newly synthesizedvaricella-zoster virus: Final envelopment in the trans-Golgi network.J. Virol. 68, 6372–6390.
ong, M., and Kieff, E. (1990). Intracellular trafficking of two majorEpstein–Barr virus glycoproteins, gp350/220 and gp110. J. Virol. 64,1507–1516.
ranzow, H., Weiland, F., Jones, A., Klupp, B. G., Karger, A., and Met-tenleitter, T. C. (1997). Ultrastructural analysis of the replication cycleof pseudorabies virus in cell culture: A reassessment. J. Virol. 71,2072–2082.
arson, R., and Grose, C. (1995). Egress of varicella-zoster virus fromthe melanoma cell: A tropism for the melanocyte. J. Virol. 69, 4994–5010.
ohnson, D. C., and Spear, P. G. (1982). Monensin inhibits the process-ing of herpes simplex virus glycoproteins, their transport to the cellsurface, and the egress of virions from infected cells. J. Virol. 43,1102–1112.
ones, F., and Grose, C. (1988). Role of cytoplasmic vacuoles in vari-cella-zoster virus glycoproteins trafficking and virion envelopment.J. Virol. 62, 2701–2711.
usso, P., and Gallo, R. C. (1995). Human herpesvirus 6 in AIDS.Immunol. Today 16, 67–71.
ii, S., Yoshida, M., Uno, F., Kurata, T., Ikuta, K., and Yamanishi, K. (1990).Replication of human herpesvirus 6 (HHV-6): morphological aspects.Adv. Exp. Med. Biol. 278, 19–28.
lson, J. K., and Grose, C. (1997). Endocytosis and recycling of vari-cella-zoster virus Fc receptor glycoprotein gE: Internalization medi-ated by a YXXL motif in the cytoplasmic tail. J. Virol. 71, 4042–4054.
feiffer, B., Berneman, Z., Naipel, F., Chang, C. K., Tirwatnapong, S., andChandran, B. (1993). Identification and mapping of the gene encodingglycoprotein gp82-gp105 of HHV-6 and mapping of the neutralizingepitope recognized by monoclonal antibodies. J. Virol. 67, 4611–4620.
feiffer, B., Thompson, B., and Chandran, B. (1995). Characterization ofa cDNA derived from multiple splicing that encodes the HHV-6 gp105of the gp82-gp105 complex. J. Virol. 69, 3490–3500.
adsak, K., Eickmann, M., Mockenhaupt, T., Bogner, E., Kern, H., Eis-

R
R
R
R
T
T
T
T
T
V
W
Z
471HHV-6 INTRACELLULAR TRANSPORT
Hubinger, A., and Reschke, M. (1996). Retrieval of human cytomeg-alovirus glycoprotein B from the infected cell surface for virus en-evelopment. Arch. Virol. 141, 557–572.
offman, E., Albert, J., Goff, J., and Frenkel, N. (1990). Putative site for theacquisition of human herpesvirus 6 virion tegument. J. Virol. 64,6308–6313.
oizman, B. (1996). Herpesviridae. In ‘‘Fields Virology’’ (B. Fields, P. M.Knipe, and P. M. Howley, Eds.), 3rd ed., Vol. 2, pp. 2221–2230.Lippincott-Raven, Philadelphia.
oizman, B., and Furlong, D. (1974). The replication of herpesviruses. In‘‘Comprehensive Virology’’ (H. Fraenkel-Conrat and R. R. Wagner,Eds.), Vol. 3, p. 229–403. Plenum Press, New York.
oth, J. (1984). Cytochemical localization of terminal N-acetyl-D-galac-tosamine residues in cellular compartments of intestinal Gobletcells: Implications for the topology of O-glycosilation. J. Cell Biol. 98,399–406.
okuyasu, K. T. (1973). A technique for ultracryotomy of cell suspen-sions and tissues. J. Cell Biol. 57, 551–565.
orrisi, M. R., and Pinto da Silva, P. (1984). Compartmentalization ofintracellular membrane glycocomponents is revealed by fracture-label. J. Cell Biol. 98, 29–34.
orrisi, M. R., Cirone, M., Pavan, A., Zompetta, C., Barile, G., Frati, L., andFaggioni, A. (1989). Localization of Epstein-Barr virus envelope gly-coproteins on the inner nuclear membrane of virus-producing cells.J. Virol. 63, 828–832.
orrisi, M. R., Di Lazzaro, C., Pavan, A., Pereira, L., and Campadelli-Fiume, G. (1992). Herpes simplex virus envelopment and maturationstudied by fracture-label. J. Virol. 66, 554–561.
orrisi, M. R., and Mancini, P. (1996). Freeze-fracture immunogold la-beling. Histochem. Cell Biol. 106, 19–30.
an Genderen, I. L., Brandimarti, R., Torrisi, M. R., Campadelli, G., andvan Meer, G. (1994). The phospholipid composition of extracellularherpes simplex virions differs from that of host cell nuclei. Virology200, 831–836.healy, M. E., Card, J. P., Meade, R. P., Robbins, A. K., and Enquist, L. W.(1991). Effect of brefeldin A on alphaherpesvirus membrane proteinglycosylation and virus egress. J. Virol. 65, 1066–1081.
hu, Z., Gershon, M. D., Hao, Y., Ambron, R. T., Gabel, C. A., andGershon, A. A. (1995). Envelopment of varicella-zoster virus: Target-ing of viral glycoproteins to the trans-Golgi network. J. Virol. 69,7951–7959.