interconnections among nuclei of the subcortical visual shell: the intergeniculate leaflet is a...
TRANSCRIPT

Interconnections Among Nucleiof the Subcortical Visual Shell:
The Intergeniculate Leaflet Is a MajorConstituent of the Hamster Subcortical
Visual System
L.P. MORIN* AND J.H. BLANCHARD
Department of Psychiatry, Health Science Center, Stony Brook University, Stony Brook,New York 11794
ABSTRACTThe intergeniculate leaflet (IGL), a major constituent of the circadian visual system, is
one of 12 retinorecipient nuclei forming a ‘‘subcortical visual shell’’ overlying the diencephalic–mesencephalic border. The present investigation evaluated IGL connections with nuclei of thesubcortical visual shell and determined the extent of interconnectivity between these nuclei.Male hamsters received stereotaxic, iontophoretic injections of the retrograde tracer, choleratoxin b fragment, or the anterograde tracer, Phaseolus vulgaris-leucoagglutin, into nuclei ofthe pretectum (medial, commissural, posterior, olivary, anterior, nucleus of the optic tract,posterior limitans), into the superior colliculus, or into the visual thalamic nuclei (lateralposterior, dorsal lateral geniculate, intergeniculate leaflet, ventral lateral geniculate). Retro-gradely labeled cell bodies identified nuclei with afferents projecting to the site of injection,whereas the presence of anterogradely labeled fibers with terminals revealed brain nucleitargeted by neurons at the site of injection. The IGL projects bilaterally to all nuclei of thevisual shell except the lateral posterior and dorsal lateral geniculate nuclei. The IGL also hasafferents from the same set of nuclei, except the nucleus of the optic tract. The extensivebilateral efferent projections distinguish IGL from the ventral lateral geniculate nucleus. Thesuperior colliculus, commissural pretectal, olivary pretectal, and posterior pretectal nucleialso project bilaterally to the majority of subcortical visual nuclei. The IGL has a well-established role in circadian rhythm regulation, but there is as yet no known function for it inthe larger context of the subcortical visual system, much of which is involved in oculomotorcontrol. J. Comp. Neurol. 396:288–309, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: pretectum; lateral posterior nucleus; dorsal lateral geniculate; ventral lateral
geniculate; superior colliculus; suprachiasmatic nucleus
The intergeniculate leaflet (IGL) is a recognized divisionof the lateral geniculate complex (Card and Moore, 1989).It is situated between the dorsal (DLG) and ventral lateralgeniculate (VLG) nuclei and extends from a point ventralto the rostral DLG caudally to the lateral terminal nucleus,a distance of about 2 mm in the hamster (Morin et al.,1992; Morin and Blanchard, 1995). The IGL is retinorecipi-ent throughout its length and contains neuropeptideY–immunoreactive (NPY-IR) neurons which are not foundelsewhere in the adult geniculate complex (Botchkina andMorin, 1995a,b). Many of the NPY-IR neurons project tothe hypothalamic suprachiasmatic nucleus (SCN), site ofthe circadian clock (for reviews, see Meijer and Rietveld,1989; Morin, 1994). This characteristic has led to the
inclusion of the IGL as part of the circadian visual system(Card and Moore, 1989; Morin et al., 1992; Morin andBlanchard, 1995). Numerous functional studies validatethat view (see Morin, 1994, for a review).
As anatomical study of the IGL has progressed, itsdefinition has enlarged from the original description by
Grant sponsor: NINDS; Grant number: NS22168.*Correspondence to: Dr. Lawrence P. Morin, Dept. of Psychiatry, Health
Science Center, SUNY, Stony Brook, NY 11794.E-mail: [email protected]
Received 7 October 1997; Revised 21 January 1998; Accepted 22 Febru-ary 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 396:288–309 (1998)
r 1998 WILEY-LISS, INC.

Hickey and Spear (1976), to one based on several diversecriteria including the developmental migration of NPY-IRneurons, transformation and translocation of radial gliainto IGL astrocytes (Botchkina and Morin, 1995b), andlocation of neurons projecting to the pretectum or contralat-eral IGL (Morin and Blanchard, 1995). The potential IGLcontribution to the non-circadian visual system has alsoexpanded (Klooster et al., 1995; Morin and Blanchard,1995, 1997). An increasing number of reports indicate arelationship between the IGL and other subcortical visualnuclei. A robust projection to the contralateral IGL is wellestablished (Card and Moore, 1989; Morin and Blanchard,1995). Anterograde studies of rat IGL efferents haveidentified projections to the anterior pretectal nucleus(Mikkelsen, 1992). In the hamster, both NPY and enkepha-lin (ENK) neurons of the IGL project to the posteriorlimitans nucleus as well as to more medial areas in thepretectum (Morin and Blanchard, 1995). Neurons of theIGL are reported to have both efferent and afferent connec-tions with the superior colliculus (SC) in rat and frog(Taylor et al., 1986; Montgomery et al., 1991; Kunzle,1996). In addition, the rat IGL has afferent connectionswith the olivary pretectal nucleus (OPT; Klooster et al.,1995).
We have recently documented the extensive retinalinnervation of hamster pretectum and have identified,according to criteria related to visual innervation andpeptide content, seven pretectal nuclei (Morin and Blan-chard, 1997). These mesencephalic nuclei, together withthe lateral posterior (LP), DLG, VLG, and IGL of thediencephalon and the mencephalic SC constitute a ‘‘visualshell’’ of contiguous nuclei overlying the juncture of themesencephalon and diencephalon (Morin and Blanchard,1997). Preliminary investigations indicated that the IGLmight project to many of the nuclei in the subcorticalvisual shell. Therefore, we have used both anterogradeand retrograde methods to evaluate IGL projections andhave additionally studied the extent of interconnectionsbetween all the nuclei of the subcortical visual shell. Theresults show a broad, strongly bilateral pattern of connec-tions between nuclei of the subcortical visual system andsupport the conclusion that the IGL has a larger visualsystem function beyond circadian rhythm regulation.
MATERIALS AND METHODS
Adult, outbred male golden hamsters (Charles River,Wilmington, MA) were maintained with free access to foodand water under a 14-hour light:10-hour dark photoperiodin individual polypropylene cages. All surgery and perfu-sions occurred during the light phase of the photoperiod inanimals deeply anesthetized with sodium pentobarbital(Anpro Pharmaceutical, Arcadia, CA; 100 mg/kg BW). Theanimal use protocol for this research was approved by theInstitutional Animal Care and Use Committee of StonyBrook University.
Surgery
Animals received injections of cholera toxin, b fragment(CT-b; 1%; product #104, List Biological Labs., Inc., Camp-bell, CA) or Phaseolus vulgaris-leucoagglutinin (PHA-L;2.5%) administered iontophoretically (2 µA for CT-b and5–7 µA for PHA-L pulsed 7 seconds on, 7 seconds off forabout 4 minutes). Each nucleus of the subcortical ‘‘visualshell’’ (Figs. 1–3) was accurately targeted at least once foreach tracer. One animal received a CT-b injection intosecondary visual cortex area 18a. Retrograde transport ofCT-b or anterograde transport of PHA-L was permitted for3–5 days. At the end of the transport period, each animalwas deeply anesthetized with pentobarbital and perfusedtranscardially with physiological saline followed by 4%paraformaldehyde in 0.1 M phosphate buffer, pH 7.4, with0.01 M sodium m-periodate and 0.075 M lysine added(McLean and Nakane, 1974).
Histological procedures
Each brain was removed, postfixed for 1 hour, cryopro-tected in a series of sucrose solutions to 30% sucrose inphosphate buffer, then frozen and serially sectioned (30µm) in the coronal plane. Sections were collected in 0.01 Mphosphate-buffered saline (PBS), pH 7.4, with 0.05% so-dium azide. All immunohistochemical reactions were per-formed using free-floating sections. Immunoperoxidasereactions were done using the ABC technique (Hsu et al.,1981; Vector kit) and diaminobenzidine as the chromogen.Antisera to CT-b (goat; List Biological Labs., Inc., Camp-bell, CA) and PHA-L (goat; Vector, Burlingame, CA orrabbit; DAKO, Carpinteria, CA) were used.
Abbreviations
APT anterior pretectal nucleusAPTd anterior pretectal nucleus, dorsal divisionAPTv anterior pretectal nucleus, ventral divisionCPT commissural pretectal nucleusCP cerebral pedunclecsc commissure of the superior colliculusDLG dorsal lateral geniculate nucleusDpG deep gray layer, superior colliculusDpWh deep white layer, superior colliculusDTN dorsal terminal nucleusfr fasciculus retroflexusHb habenulaIGL intergeniculate leafletIMA intramedullary areaInG internal gray layer, superior colliculusInWh internal white layer, superior colliculusLP lateral posterior nucleusLTN lateral terminal nucleusMCPC magnocellular nucleus of the posterior commissureMG medial geniculate nucleusMPT medial pretectal nucleus
MTN medial terminal nucleusNOT nucleus of the optic tractOP optic layer, superior colliculusOPT olivary pretectal nucleusot optic tractPBg parabigeminal nucleuspc posterior commissurePLi posterior limitans nucleusPo posterior thalamic nucleusPPN peripeduncular nucleusPPT posterior pretectal nucleusPrC precommissural nucleusRt reticular thalamic nucleusSC superior colliculusSCN suprachiasmatic nucleusSG superficial gray layer, superior colliculusSubG subgeniculate nucleusVLG ventral lateral geniculate nucleusZI zona incerta3 third ventricle
SUBCORTICAL VISUAL CONNECTIONS 289

Fig. 1. Schematic representation of all CT-b injection sites, exceptthose in the superior colliculus (SC), contributing to the results. Forthe sake of uniformity, all injection sites are shown on the same side ofthe diagrams, although the actual side varied across individuals.
Shapes and sizes of the injections are indicated by each shaded region.The plane of section in which an injection site is identified represents asection through the approximate middle of the site. See Figure 3 forsites in the SC. For abbreviations, see list.
290 L.P. MORIN AND J.H. BLANCHARD
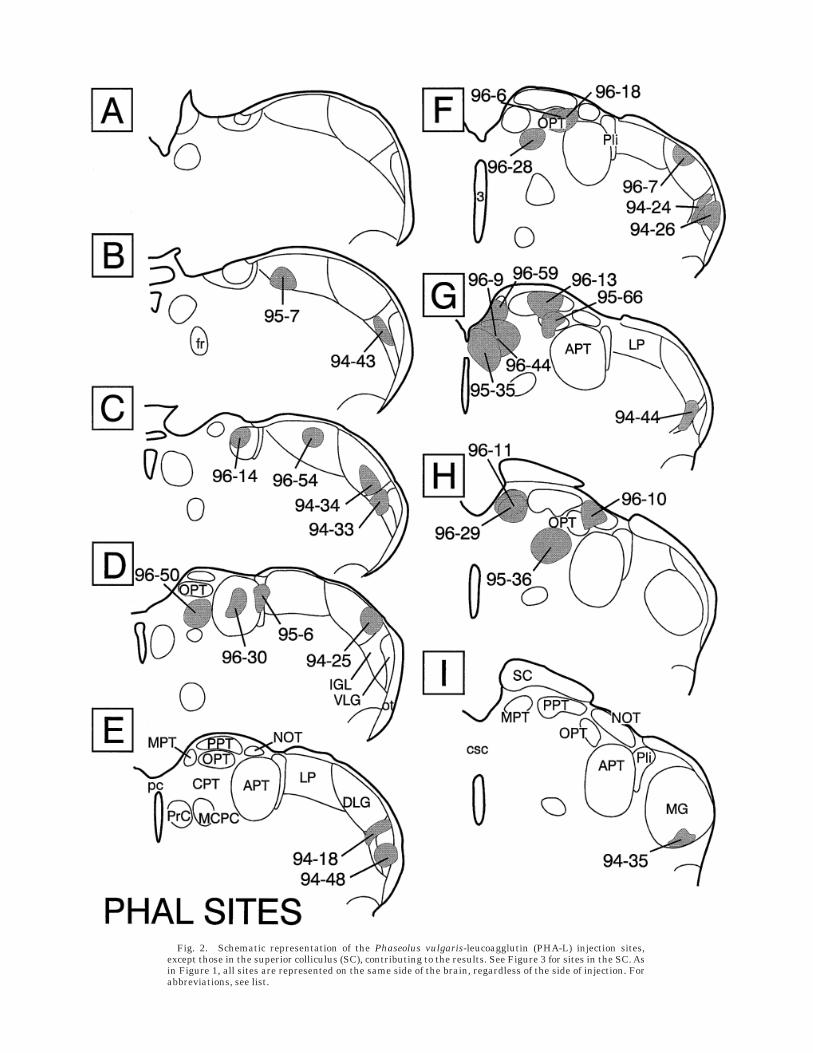
Fig. 2. Schematic representation of the Phaseolus vulgaris-leucoagglutin (PHA-L) injection sites,except those in the superior colliculus (SC), contributing to the results. See Figure 3 for sites in the SC. Asin Figure 1, all sites are represented on the same side of the brain, regardless of the side of injection. Forabbreviations, see list.
Microscopy
Brain sections from PHA-L– and CT-b–injected animalswere evaluated with a Nikon Optiphot microscope usingbrightfield and darkfield techniques. Only the results frombrains with well-targeted tracer placement are reported. Acamera lucida was utilized for making brain drawings.PHA-L injections label a cluster of neurons around theinjection site. The volume described by the cluster wasconsidered the region of effective label uptake (Gerfen andSawchenko, 1984; Morin et al., 1994). The center of eachCT-b injection has a slightly more brilliant appearancewhen viewed using darkfield microscopy. It is surroundedby a region of reaction product that is distinctly denserthan slightly more distal areas. The volume within theboundary of this denser region is considered the zone ofeffective CT-b uptake.
RESULTS
A total of 186 animals received injections which pro-duced good reaction product. Of these, 66 cases hadinjection sites sufficiently accurate to permit inclusion inthe present study. The posterior pretectal nucleus (PPT)and nucleus of the optic tract (NOT) were particularlydifficult to inject. Most attempts missed dorsally or in-cluded adjacent ventral nuclei as well. One major cause offaulty injections is related to the fact that the NOT, theolivary pretectal nucleus (OPT), the anterior pretectalnucleus (APT), the posterior limitans nucleus (PLi), andthe LP have a general orientation angling obliquely awayfrom the midline. That is, the rostral ends of the nuclei arenear the midline, but the caudal ends are considerablymore lateral. These nuclei can appear large and adjacentto one another when viewed in the coronal plane (e.g., APTor LP in Fig. 1D–G). However, serial reconstructions(Scalia and Arango, 1979) or horizontal sections (Morinand Blanchard, 1997) reveal the nuclei in an overlapping,front-to-back relationship. Because of this, injections thatappeared to be properly centered in a target site (Figs. 1–3)when viewed at one coronal plane often proved to extendinto adjacent nuclei at a slightly more rostral or caudalplane. Figures 1 and 2 provide the most thorough represen-
tations of size and location of each subcortical visualnucleus (according to definitions in Morin and Blanchard,1997) and should be used to guide interpretation ofmaterial shown in the photomicrographs.
In this study, a projection from one nucleus to another isnot described unless such an interconnection was demon-strated with both anterograde and retrograde methods.Exceptions to this rule are specifically noted. Table 1summarizes the interconnections between the subcorticalvisual nuclei.
Intergeniculate leaflet
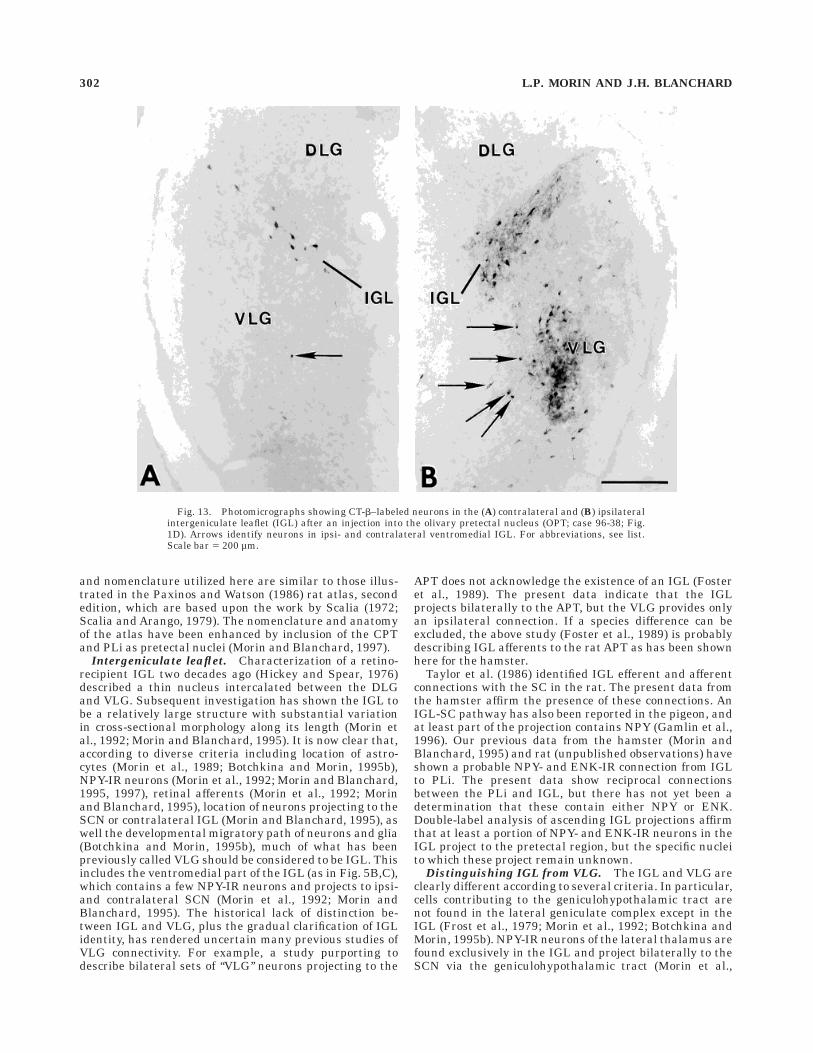
Each of the three IGL injections of CT-b (Fig. 1B,D)included a small amount of spread into adjacent VLG.Numerous neurons are labeled in the IGL and bothintermediate and lateral divisions of the VLG. Neuronsafferent to the IGL are present in each of the ipsilateralsubcortical visual nuclei except the NOT, DLG and LP(Figs. 4–6). The nuclei of the medial pretectum (OPT, PPT,medial pretectal nucleus [MPT], and commissural pretec-tal area [CPT]) also project to the contralateral IGL. IGLefferent projections are even more widespread, terminat-ing in each of the ipsilateral subcortical visual nucleiexcept the DLG and LP (Fig. 7). In addition, IGL efferentprojections terminate in the same nuclei on the contralat-eral side. In each case for which there are efferent projec-tions to another subcortical visual nucleus demonstratedby CT-b injection, labeled neurons are present in both theipsi- and contralateral ventromedial aspect of the IGL aswell as in the ‘‘leaflet’’ portion (Figs. 5B,C, 12, 13). Asstated above, the patterns of innervation observed in thebrains injected with retrograde label were confirmed fromthe anterograde labeling of fibers and terminals in the IGLafter PHA-L injection into each of the subcortical visualnuclei (Figs. 2, 3). The anterograde label reveals numerousfibers from the IGL crossing the midline in the supraoptic,posterior, and superior colliculus commissures.
Ventral lateral geniculate nucleus
Two cases with CT-b injected into the VLG (Fig. 1E,F)produced a pattern of retrogradely labeled neurons verysimilar to that seen after IGL injections (Fig. 5). Four
Fig. 3. Schematic representation of the CT-b and Phaseolus vulgaris-leucoagglutin (PHA-L) injectionsites in the superior colliculus (SC). For abbreviations, see list.
292 L.P. MORIN AND J.H. BLANCHARD

differences between IGL and VLG afferent cell patternswere noted. Labeled cells are present in the ipsilateralNOT, present in the contralateral SC, and not present inthe contralateral MPT or in the precommissural nucleusafter VLG injections. The opposite results are true afterIGL injections. With respect to efferent projections, theVLG is the same as the IGL on the ipsilateral side.However, the most noteworthy difference between theinnervation patterns of the two nuclei is evident in the
distribution of efferents to contralateral subcortical visualnuclei. The VLG projects contralaterally only to the CPT,unlike the IGL which also projects to the contralateralIGL, PLi, APT, NOT, OPT, APT, MPT, and SC (Table 1).
The two CT-b injections differed in location and pro-duced different results in the NOT. Analysis of PHA-L–1Rfibers and terminals in the VLG after a NOT injection (Fig.2H; case 96-10) revealed two distinct regions. A morerostral, central zone essentially devoid of fibers with
TABLE 1. Projections From Nuclei in the Subcortical Visual Shell to Other Nuclei in the Visual Shell Determined From the Combined Analysis of Boththe Retrograde CT-b and Anterograde PHA-L Data1
Targets withafferentterminals
Location of cell bodies
VLG IGL DLG LP PLi APT OPT NOT PPT MPT CPT SC
VLG */2 1/1 2/2 2/2 1/2 1/2 1/1 1/2 1/1 1/2 1/1 1/1IGL 1/2 1/1 2/2 2/2 1/2 1/2 1/1 2/2 1/1 1/1 1/1 1/2DLG 2/2 2/2 2/2 2/2 2/2 2/2 1/2 2/2 2/2 2/2 2/2 1/2LP 2/2 2/2 2/2 */2 2/2 2/2 1/2 2/2 1/2 1/2 2/2 1/1PLi 1/2 1/1 2/2 2/2 1/2 1/2 1/2 2/2 1/2 1/2 1/1 1/1APT 1/2 1/1 2/2 2/2 1/2 1/2 1/2 2/2 1/2 1/2 1/1 1/2OPT 1/2 1/1 2/2 2/2 1/2 1/2 1/1 1/2 1/1 1/2 1/1 1/1NOT 1/2 1/1 2/2 2/2 1/2 1/2 1/1 1/1 1/1 1/2 1/2 1/1PPT 1/2 1/1 2/2 2/2 1/2 1/2 1/1 1/2 1/1 1/2 1/1 1/1MPT 1/2 1/1 2/2 2/2 1/2 1/2 1/2 2/2 1/2 */1 1/1 1/1CPT 1/1 1/1 2/2 2/2 1/1 1/1 1/1 1/2 1/2 1/1 1/1 1/1SC 1/2 1/1 2/2 2/2 1/2 1/2 1/1 1/1 1/1 1/1 1/1 1/1
11 or 2 5 presence or absence of a projection; left of slash 5 ipsilateral; right of slash 5 contralateral.*Could not be determined because the area of interest was obscured by reaction product.
Fig. 4. Brightfield photomicrographs showing neurons labeled inthe ipsilateral (A,C) and contralateral (B,D) midbrain after a CT-binjection into the intergeniculate leaflet (IGL). A,B: Dorsal pretectalregion of case 94-52. The plane of section is approximately that of
Figure 1G. C,D: Dorsocaudal pretectum and rostral superior colliculus(SC), at a level approximately 480 µm caudal to level (A) of case 94-39.See Figure 11 for delineations of brain nuclei. Scale bar 5 500 µm.
SUBCORTICAL VISUAL CONNECTIONS 293
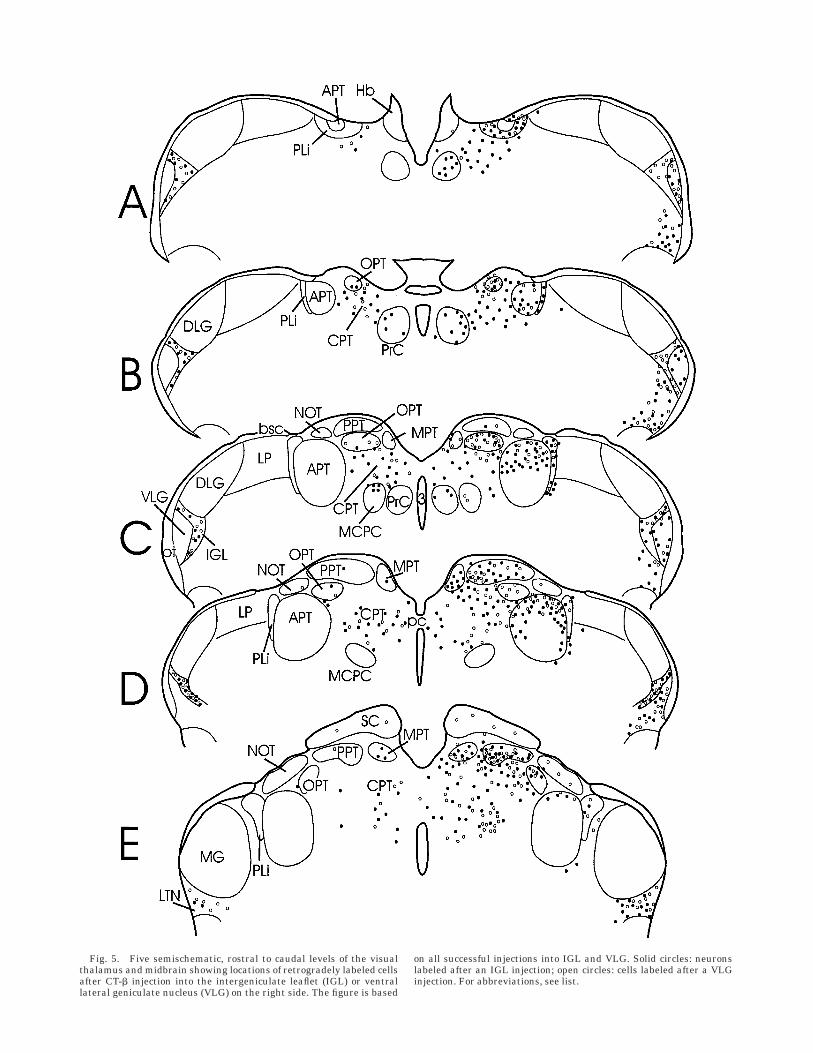
Fig. 5. Five semischematic, rostral to caudal levels of the visualthalamus and midbrain showing locations of retrogradely labeled cellsafter CT-b injection into the intergeniculate leaflet (IGL) or ventrallateral geniculate nucleus (VLG) on the right side. The figure is based
on all successful injections into IGL and VLG. Solid circles: neuronslabeled after an IGL injection; open circles: cells labeled after a VLGinjection. For abbreviations, see list.
terminals (Fig. 8A) is generally surrounded (particularlycaudoventrally) by a terminal field (Fig. 8B). The lattercorresponds to the CT-b injection site (Fig. 1F; case 97-30),yielding labeled neurons in the NOT. In contrast, the morerostral and central CT-b site of case 94-55 (Fig. 1E)corresponds to the central zone and is not associated withlabeled neurons in the NOT. Thus, both the anterograde
and retrograde data suggest regional specificity within theVLG, at least with respect to innervation by the NOT.CT-b–labeled neurons were also found in somewhat differ-ent parts of the SC, depending upon the VLG injection site(Fig. 6). Both injections labeled neurons in the optic andinternal gray layers, but only the more rostral and centralinjection (case 94-55) labeled neurons in the superficialgray layer.
Dorsal lateral geniculate nucleus
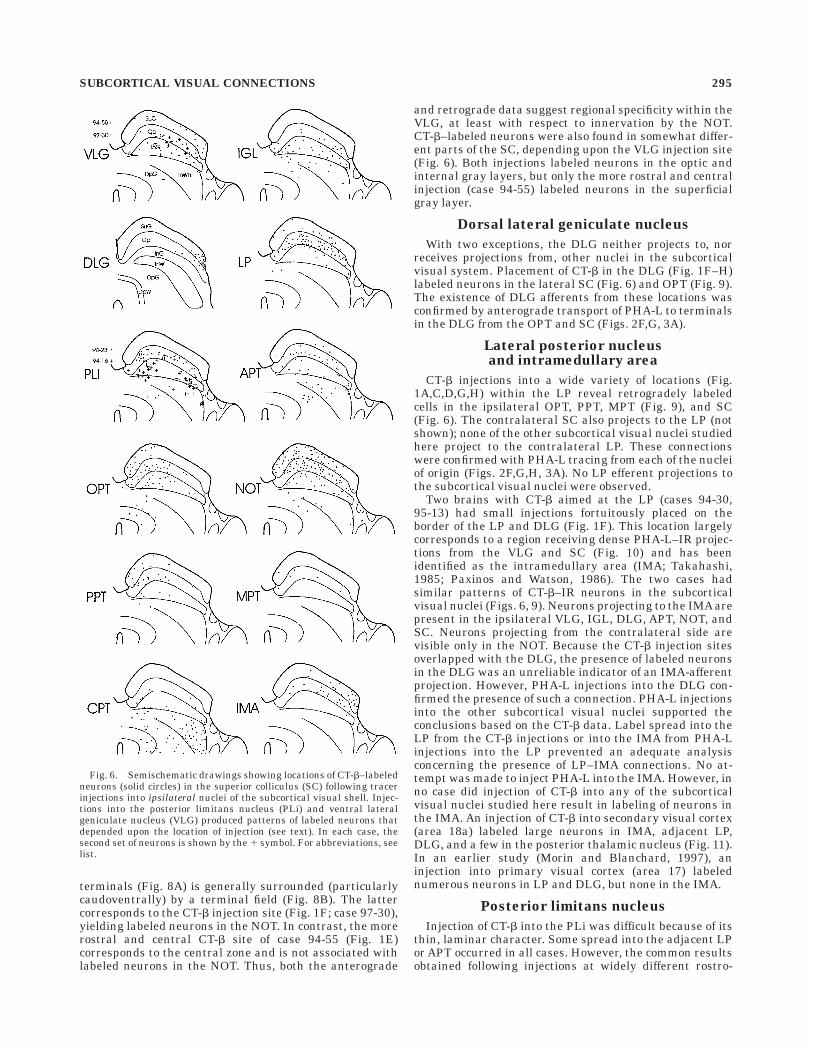
With two exceptions, the DLG neither projects to, norreceives projections from, other nuclei in the subcorticalvisual system. Placement of CT-b in the DLG (Fig. 1F–H)labeled neurons in the lateral SC (Fig. 6) and OPT (Fig. 9).The existence of DLG afferents from these locations wasconfirmed by anterograde transport of PHA-L to terminalsin the DLG from the OPT and SC (Figs. 2F,G, 3A).
Lateral posterior nucleusand intramedullary area
CT-b injections into a wide variety of locations (Fig.1A,C,D,G,H) within the LP reveal retrogradely labeledcells in the ipsilateral OPT, PPT, MPT (Fig. 9), and SC(Fig. 6). The contralateral SC also projects to the LP (notshown); none of the other subcortical visual nuclei studiedhere project to the contralateral LP. These connectionswere confirmed with PHA-L tracing from each of the nucleiof origin (Figs. 2F,G,H, 3A). No LP efferent projections tothe subcortical visual nuclei were observed.
Two brains with CT-b aimed at the LP (cases 94-30,95-13) had small injections fortuitously placed on theborder of the LP and DLG (Fig. 1F). This location largelycorresponds to a region receiving dense PHA-L–IR projec-tions from the VLG and SC (Fig. 10) and has beenidentified as the intramedullary area (IMA; Takahashi,1985; Paxinos and Watson, 1986). The two cases hadsimilar patterns of CT-b–IR neurons in the subcorticalvisual nuclei (Figs. 6, 9). Neurons projecting to the IMA arepresent in the ipsilateral VLG, IGL, DLG, APT, NOT, andSC. Neurons projecting from the contralateral side arevisible only in the NOT. Because the CT-b injection sitesoverlapped with the DLG, the presence of labeled neuronsin the DLG was an unreliable indicator of an IMA-afferentprojection. However, PHA-L injections into the DLG con-firmed the presence of such a connection. PHA-L injectionsinto the other subcortical visual nuclei supported theconclusions based on the CT-b data. Label spread into theLP from the CT-b injections or into the IMA from PHA-Linjections into the LP prevented an adequate analysisconcerning the presence of LP–IMA connections. No at-tempt was made to inject PHA-L into the IMA. However, inno case did injection of CT-b into any of the subcorticalvisual nuclei studied here result in labeling of neurons inthe IMA. An injection of CT-b into secondary visual cortex(area 18a) labeled large neurons in IMA, adjacent LP,DLG, and a few in the posterior thalamic nucleus (Fig. 11).In an earlier study (Morin and Blanchard, 1997), aninjection into primary visual cortex (area 17) labelednumerous neurons in LP and DLG, but none in the IMA.
Posterior limitans nucleus
Injection of CT-b into the PLi was difficult because of itsthin, laminar character. Some spread into the adjacent LPor APT occurred in all cases. However, the common resultsobtained following injections at widely different rostro-
Fig. 6. Semischematic drawings showing locations of CT-b–labeledneurons (solid circles) in the superior colliculus (SC) following tracerinjections into ipsilateral nuclei of the subcortical visual shell. Injec-tions into the posterior limitans nucleus (PLi) and ventral lateralgeniculate nucleus (VLG) produced patterns of labeled neurons thatdepended upon the location of injection (see text). In each case, thesecond set of neurons is shown by the 1 symbol. For abbreviations, seelist.
SUBCORTICAL VISUAL CONNECTIONS 295
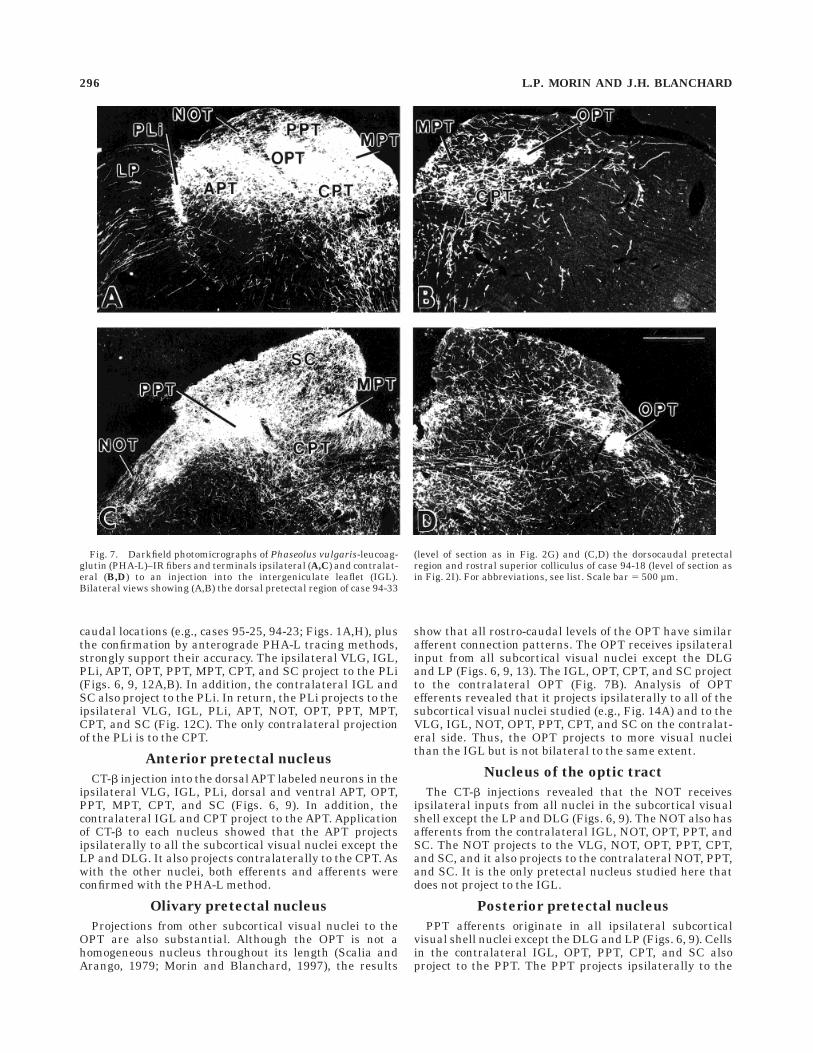
caudal locations (e.g., cases 95-25, 94-23; Figs. 1A,H), plusthe confirmation by anterograde PHA-L tracing methods,strongly support their accuracy. The ipsilateral VLG, IGL,PLi, APT, OPT, PPT, MPT, CPT, and SC project to the PLi(Figs. 6, 9, 12A,B). In addition, the contralateral IGL andSC also project to the PLi. In return, the PLi projects to theipsilateral VLG, IGL, PLi, APT, NOT, OPT, PPT, MPT,CPT, and SC (Fig. 12C). The only contralateral projectionof the PLi is to the CPT.
Anterior pretectal nucleus
CT-b injection into the dorsal APT labeled neurons in theipsilateral VLG, IGL, PLi, dorsal and ventral APT, OPT,PPT, MPT, CPT, and SC (Figs. 6, 9). In addition, thecontralateral IGL and CPT project to the APT. Applicationof CT-b to each nucleus showed that the APT projectsipsilaterally to all the subcortical visual nuclei except theLP and DLG. It also projects contralaterally to the CPT. Aswith the other nuclei, both efferents and afferents wereconfirmed with the PHA-L method.
Olivary pretectal nucleus
Projections from other subcortical visual nuclei to theOPT are also substantial. Although the OPT is not ahomogeneous nucleus throughout its length (Scalia andArango, 1979; Morin and Blanchard, 1997), the results
show that all rostro-caudal levels of the OPT have similarafferent connection patterns. The OPT receives ipsilateralinput from all subcortical visual nuclei except the DLGand LP (Figs. 6, 9, 13). The IGL, OPT, CPT, and SC projectto the contralateral OPT (Fig. 7B). Analysis of OPTefferents revealed that it projects ipsilaterally to all of thesubcortical visual nuclei studied (e.g., Fig. 14A) and to theVLG, IGL, NOT, OPT, PPT, CPT, and SC on the contralat-eral side. Thus, the OPT projects to more visual nucleithan the IGL but is not bilateral to the same extent.
Nucleus of the optic tract
The CT-b injections revealed that the NOT receivesipsilateral inputs from all nuclei in the subcortical visualshell except the LP and DLG (Figs. 6, 9). The NOT also hasafferents from the contralateral IGL, NOT, OPT, PPT, andSC. The NOT projects to the VLG, NOT, OPT, PPT, CPT,and SC, and it also projects to the contralateral NOT, PPT,and SC. It is the only pretectal nucleus studied here thatdoes not project to the IGL.
Posterior pretectal nucleus
PPT afferents originate in all ipsilateral subcorticalvisual shell nuclei except the DLG and LP (Figs. 6, 9). Cellsin the contralateral IGL, OPT, PPT, CPT, and SC alsoproject to the PPT. The PPT projects ipsilaterally to the
Fig. 7. Darkfield photomicrographs of Phaseolus vulgaris-leucoag-glutin (PHA-L)–IR fibers and terminals ipsilateral (A,C) and contralat-eral (B,D) to an injection into the intergeniculate leaflet (IGL).Bilateral views showing (A,B) the dorsal pretectal region of case 94-33
(level of section as in Fig. 2G) and (C,D) the dorsocaudal pretectalregion and rostral superior colliculus of case 94-18 (level of section asin Fig. 2I). For abbreviations, see list. Scale bar 5 500 µm.
296 L.P. MORIN AND J.H. BLANCHARD
VLG, IGL, LP, and all pretectal nuclei. It also projects tothe contralateral VLG, IGL, NOT, OPT, PPT, and SC.
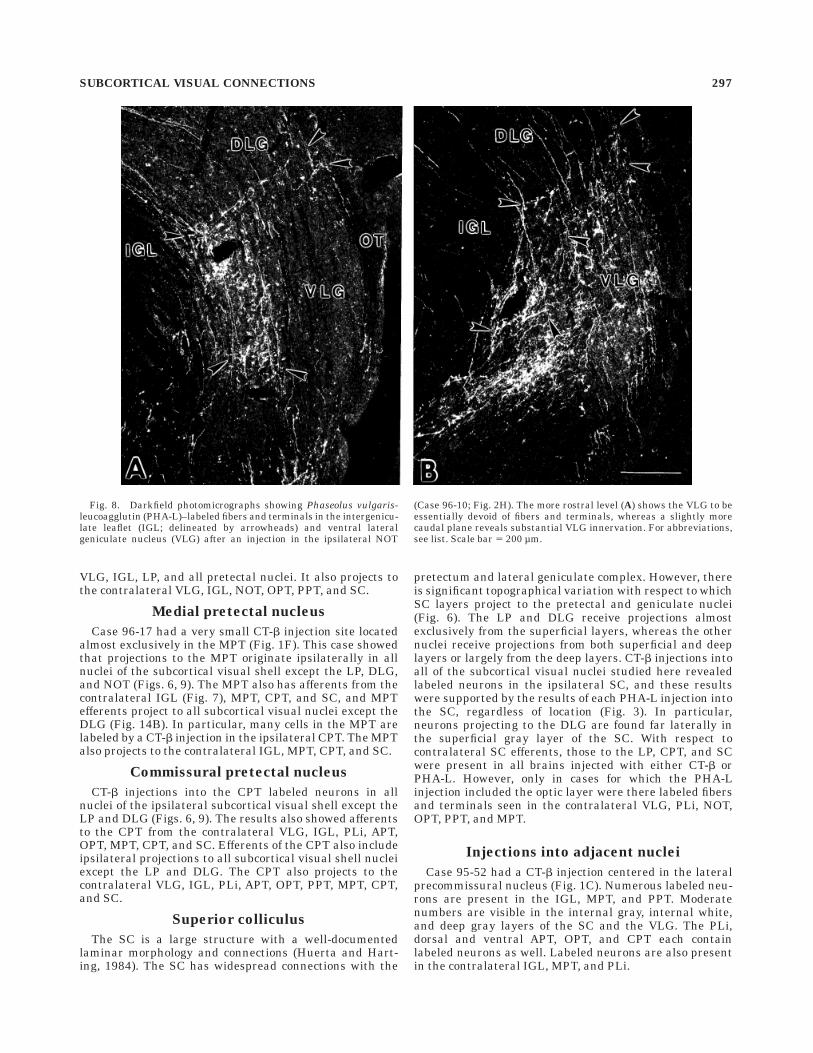
Medial pretectal nucleus
Case 96-17 had a very small CT-b injection site locatedalmost exclusively in the MPT (Fig. 1F). This case showedthat projections to the MPT originate ipsilaterally in allnuclei of the subcortical visual shell except the LP, DLG,and NOT (Figs. 6, 9). The MPT also has afferents from thecontralateral IGL (Fig. 7), MPT, CPT, and SC, and MPTefferents project to all subcortical visual nuclei except theDLG (Fig. 14B). In particular, many cells in the MPT arelabeled by a CT-b injection in the ipsilateral CPT. The MPTalso projects to the contralateral IGL, MPT, CPT, and SC.
Commissural pretectal nucleus
CT-b injections into the CPT labeled neurons in allnuclei of the ipsilateral subcortical visual shell except theLP and DLG (Figs. 6, 9). The results also showed afferentsto the CPT from the contralateral VLG, IGL, PLi, APT,OPT, MPT, CPT, and SC. Efferents of the CPT also includeipsilateral projections to all subcortical visual shell nucleiexcept the LP and DLG. The CPT also projects to thecontralateral VLG, IGL, PLi, APT, OPT, PPT, MPT, CPT,and SC.
Superior colliculus
The SC is a large structure with a well-documentedlaminar morphology and connections (Huerta and Hart-ing, 1984). The SC has widespread connections with the
pretectum and lateral geniculate complex. However, thereis significant topographical variation with respect to whichSC layers project to the pretectal and geniculate nuclei(Fig. 6). The LP and DLG receive projections almostexclusively from the superficial layers, whereas the othernuclei receive projections from both superficial and deeplayers or largely from the deep layers. CT-b injections intoall of the subcortical visual nuclei studied here revealedlabeled neurons in the ipsilateral SC, and these resultswere supported by the results of each PHA-L injection intothe SC, regardless of location (Fig. 3). In particular,neurons projecting to the DLG are found far laterally inthe superficial gray layer of the SC. With respect tocontralateral SC efferents, those to the LP, CPT, and SCwere present in all brains injected with either CT-b orPHA-L. However, only in cases for which the PHA-Linjection included the optic layer were there labeled fibersand terminals seen in the contralateral VLG, PLi, NOT,OPT, PPT, and MPT.
Injections into adjacent nuclei
Case 95-52 had a CT-b injection centered in the lateralprecommissural nucleus (Fig. 1C). Numerous labeled neu-rons are present in the IGL, MPT, and PPT. Moderatenumbers are visible in the internal gray, internal white,and deep gray layers of the SC and the VLG. The PLi,dorsal and ventral APT, OPT, and CPT each containlabeled neurons as well. Labeled neurons are also presentin the contralateral IGL, MPT, and PLi.
Fig. 8. Darkfield photomicrographs showing Phaseolus vulgaris-leucoagglutin (PHA-L)–labeled fibers and terminals in the intergenicu-late leaflet (IGL; delineated by arrowheads) and ventral lateralgeniculate nucleus (VLG) after an injection in the ipsilateral NOT
(Case 96-10; Fig. 2H). The more rostral level (A) shows the VLG to beessentially devoid of fibers and terminals, whereas a slightly morecaudal plane reveals substantial VLG innervation. For abbreviations,see list. Scale bar 5 200 µm.
SUBCORTICAL VISUAL CONNECTIONS 297
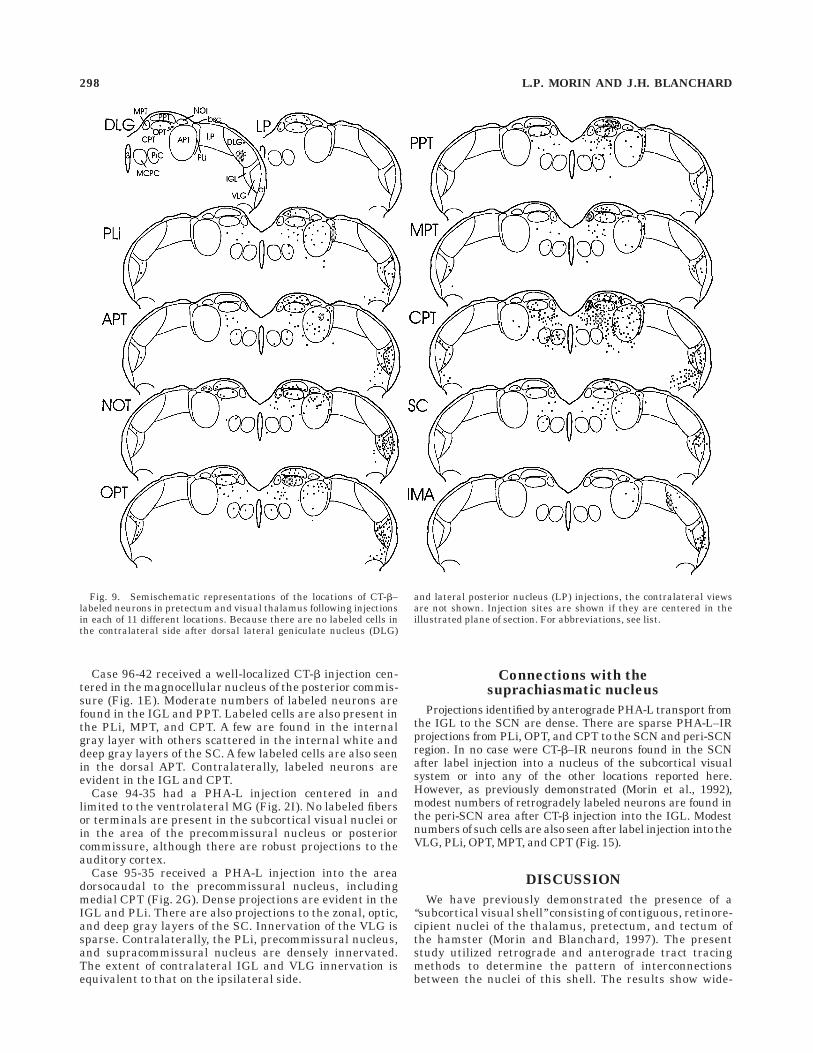
Case 96-42 received a well-localized CT-b injection cen-tered in the magnocellular nucleus of the posterior commis-sure (Fig. 1E). Moderate numbers of labeled neurons arefound in the IGL and PPT. Labeled cells are also present inthe PLi, MPT, and CPT. A few are found in the internalgray layer with others scattered in the internal white anddeep gray layers of the SC. A few labeled cells are also seenin the dorsal APT. Contralaterally, labeled neurons areevident in the IGL and CPT.
Case 94-35 had a PHA-L injection centered in andlimited to the ventrolateral MG (Fig. 2I). No labeled fibersor terminals are present in the subcortical visual nuclei orin the area of the precommissural nucleus or posteriorcommissure, although there are robust projections to theauditory cortex.
Case 95-35 received a PHA-L injection into the areadorsocaudal to the precommissural nucleus, includingmedial CPT (Fig. 2G). Dense projections are evident in theIGL and PLi. There are also projections to the zonal, optic,and deep gray layers of the SC. Innervation of the VLG issparse. Contralaterally, the PLi, precommissural nucleus,and supracommissural nucleus are densely innervated.The extent of contralateral IGL and VLG innervation isequivalent to that on the ipsilateral side.
Connections with thesuprachiasmatic nucleus
Projections identified by anterograde PHA-L transport fromthe IGL to the SCN are dense. There are sparse PHA-L–IRprojections from PLi, OPT, and CPT to the SCN and peri-SCNregion. In no case were CT-b–IR neurons found in the SCNafter label injection into a nucleus of the subcortical visualsystem or into any of the other locations reported here.However, as previously demonstrated (Morin et al., 1992),modest numbers of retrogradely labeled neurons are found inthe peri-SCN area after CT-b injection into the IGL. Modestnumbers of such cells are also seen after label injection into theVLG, PLi, OPT, MPT, and CPT (Fig. 15).
DISCUSSION
We have previously demonstrated the presence of a‘‘subcortical visual shell’’ consisting of contiguous, retinore-cipient nuclei of the thalamus, pretectum, and tectum ofthe hamster (Morin and Blanchard, 1997). The presentstudy utilized retrograde and anterograde tract tracingmethods to determine the pattern of interconnectionsbetween the nuclei of this shell. The results show wide-
Fig. 9. Semischematic representations of the locations of CT-b–labeled neurons in pretectum and visual thalamus following injectionsin each of 11 different locations. Because there are no labeled cells inthe contralateral side after dorsal lateral geniculate nucleus (DLG)
and lateral posterior nucleus (LP) injections, the contralateral viewsare not shown. Injection sites are shown if they are centered in theillustrated plane of section. For abbreviations, see list.
298 L.P. MORIN AND J.H. BLANCHARD

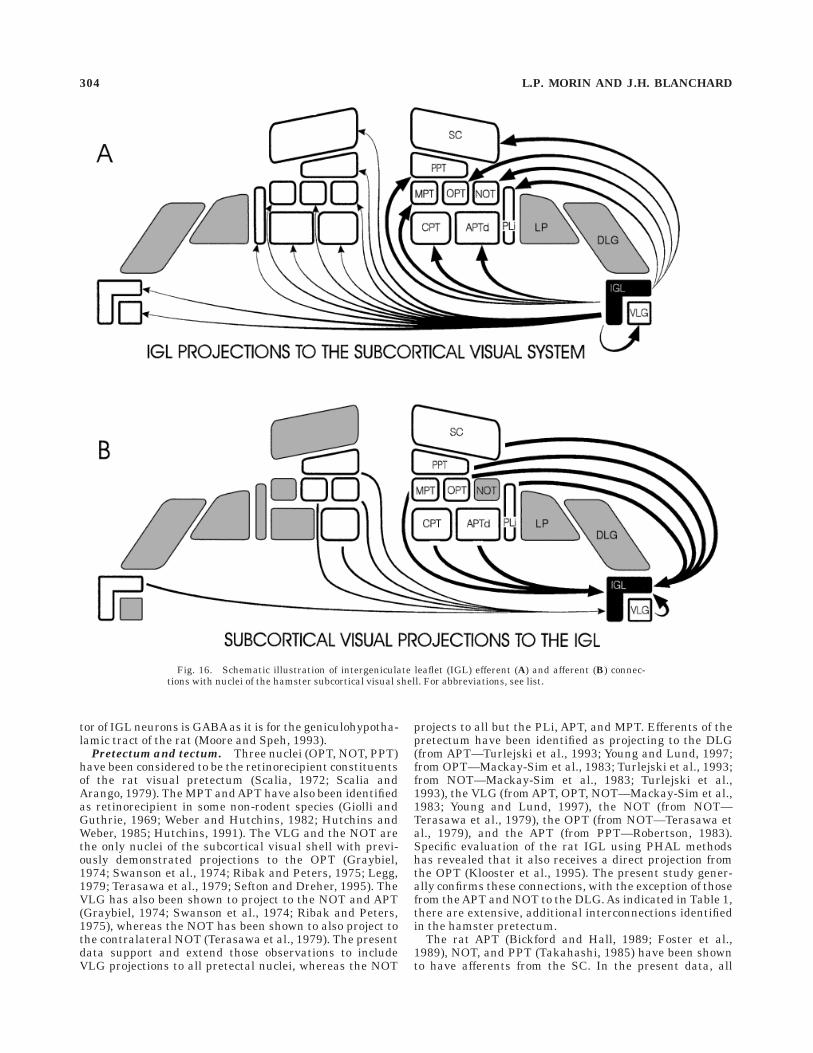
spread interconnectivity of nuclei in the pretectum, tec-tum, and ventral lateral thalamus. With a few exceptions,the nuclei do not connect directly to the cortical relaynuclei of the dorsal and lateral visual thalamus. One majorconclusion is that the IGL, previously considered solely aconstituent of the circadian visual system, is an integralpart of the subcortical visual system, extensively con-nected to the pretectum and tectum by both efferents andafferents which are usually bilateral (Fig. 16).
Technical considerations
The advantages of using both retrograde and antero-grade methods to confirm the presence of internuclearconnections are twofold. First, the experimental errors ofthe two tract tracing methods are unlikely to be the same.One such error might be spurious uptake of label by fibersof passage at the site of injection. Although this type ofproblem can occasionally occur for both CT-b and PHA-L(Cliffer and Giesler, 1988; Rhoades et al., 1989; Chen andAston-Jones, 1995), it is unlikely that the error will occurfor both tracer types in a fashion that will lead to afalse-positive declaration of interconnection between twonuclei. Second, the two tracer types are injected into bothtarget and source nuclei, thereby reducing the likelihood offalse-positive results. The requirement in the presentstudy that both tracer types confirm each projection is aconservative approach which may have reduced the num-ber of identified connections. This is particularly likely forcontralateral projections, which are always less robustthan those which are ipsilateral. Such may be the case forVLG efferents which, in the rat, have been reported toproject bilaterally to the OPT and NOT (Swanson et al.,1974; Ribak and Peters, 1975; Martin et al., 1993). Onlythe ipsilateral projections are confirmed here. On the otherhand, it is possible that the earlier studies of the rat VLGefferents included injection of tracer into the IGL which,according to the present data, does project bilaterally tothese nuclei.
The most serious limitation of the present work is thedifficulty of identifying the precise area within which theapplied tracer is actually incorporated by neurons fortransport. In general, we were unable to limit the denselyimmunoreactive area around an injection site to any of thesmaller target nuclei. On the other hand, several caseswith electrode tips in different brain nuclei which are verynear one another had markedly different patterns oflabeled cells and fibers. The best example is in the region ofthe IGL-DLG-VLG where placement in any one of thethree locations produces a pattern distinct from placementin an adjacent area. These and similar data from thepretectum suggest that the unimpeded radius for thecritical zone of iontophoresis of CT-b or PHA-L was fairlysmall (300–400 µm), with the injection parameters em-ployed here. This is consistent with the view that thecritical region of PHA-L application is that which containsPHA-L–labeled cell bodies (Gerfen and Sawchenko, 1984).In addition, injection sites frequently conformed in shapeand size to nuclear boundary constraints. This phenom-enon was particularly obvious for IGL and PLi injections(e.g., cases 95-6, 94-18; Fig. 2D,E). An additional consider-ation is the fact that anterograde transport of PHA-L canoccur in neurons located quite far from the injection site ifdendrites extend into the critical zone of label uptake(Gerfen and Sawchenko, 1984; Morin et al., 1994).
It is also possible that some of the neurons visualizedthrough the use of CT-b are the consequence of transsynap-tic label transport as has been reported for cholera toxinconjugated to HRP in the rat visual system (Trojanowskiand Schmidt, 1984). However, we have shown elsewherethat after CT-b is injected intraocularly, neurons are notlabeled anywhere in the hamster visual system except inthe oculomotor nucleus (Morin and Blanchard, 1997). Suchlabeling is expected because of terminal uptake and directretrograde transport by oculomotor fibers. The absence ofother labeled neurons in the study of retinal efferentssupports the view that, if transsynaptic labeling occurred
Fig. 10. Darkfield photomicrograph showing Phaseolus vulgaris-leucoagglutin (PHA-L)–labeled fibers and terminals in the intramedul-lary area (IMA) from an injection site in the ipsilateral ventral lateralgeniculate nucleus (VLG; case 94-26). See Figure 2E for a more
complete delineation of the brain nuclei at this level of section.Asterisk indicates a blood vessel. For abbreviations, see list. Scalebar 5 500 µm.
SUBCORTICAL VISUAL CONNECTIONS 299
in the present studies, it was insufficient to distinctlyidentify second-order neurons.
Anatomical considerations
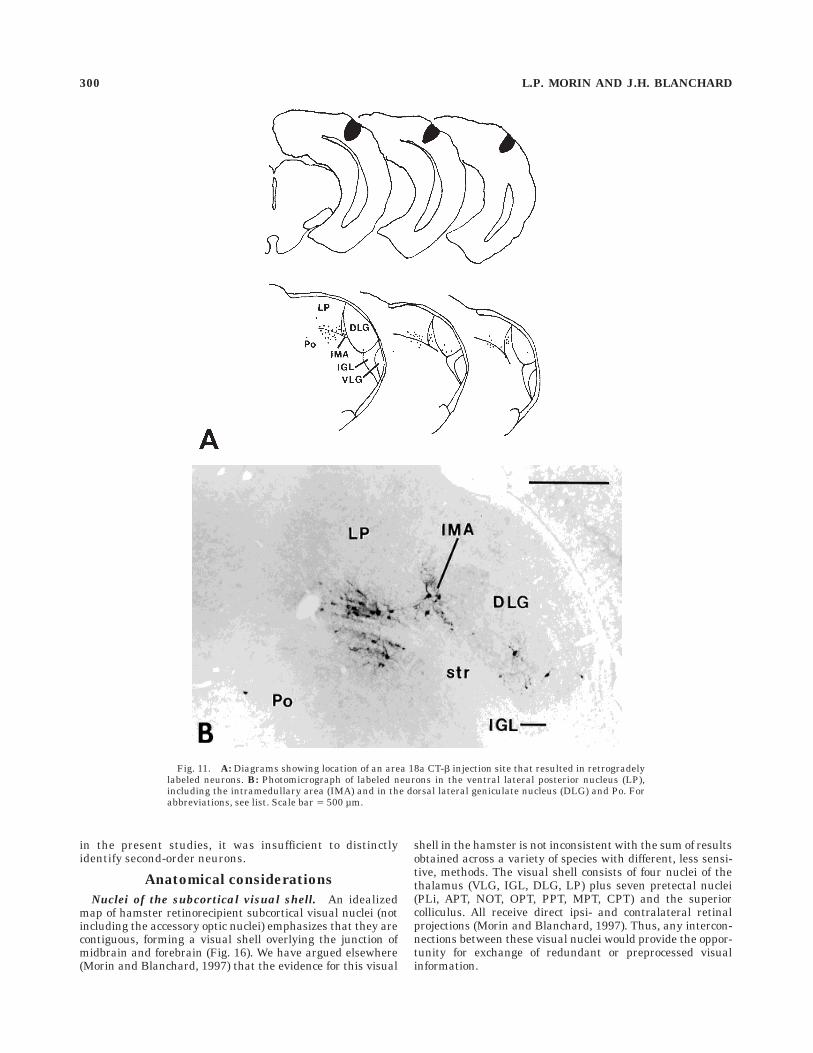
Nuclei of the subcortical visual shell. An idealizedmap of hamster retinorecipient subcortical visual nuclei (notincluding the accessory optic nuclei) emphasizes that they arecontiguous, forming a visual shell overlying the junction ofmidbrain and forebrain (Fig. 16). We have argued elsewhere(Morin and Blanchard, 1997) that the evidence for this visual
shell in the hamster is not inconsistent with the sum of resultsobtained across a variety of species with different, less sensi-tive, methods. The visual shell consists of four nuclei of thethalamus (VLG, IGL, DLG, LP) plus seven pretectal nuclei(PLi, APT, NOT, OPT, PPT, MPT, CPT) and the superiorcolliculus. All receive direct ipsi- and contralateral retinalprojections (Morin and Blanchard, 1997). Thus, any intercon-nections between these visual nuclei would provide the oppor-tunity for exchange of redundant or preprocessed visualinformation.
Fig. 11. A: Diagrams showing location of an area 18a CT-b injection site that resulted in retrogradelylabeled neurons. B: Photomicrograph of labeled neurons in the ventral lateral posterior nucleus (LP),including the intramedullary area (IMA) and in the dorsal lateral geniculate nucleus (DLG) and Po. Forabbreviations, see list. Scale bar 5 500 µm.
300 L.P. MORIN AND J.H. BLANCHARD

There is an extensive literature (e.g., Berman, 1977;Weber and Harting, 1980) describing various efferent andafferent projections of the subcortical visual nuclei, al-though there has been relatively little emphasis on inter-connections between these nuclei. Connectivity of mostnuclei in the rat visual system has been the subject of acomprehensive recent review (Sefton and Dreher, 1995).However, accurate comparison of the present results withthe rat literature is difficult because of 1) the historicallack of distinction between IGL and VLG (see below),
2) nomenclatural confusion that has surrounded studies ofthe pretectum for decades (for a thorough discussion, seeSimpson et al., 1988), 3) absence of information aboutretinal projections to the rat MPT, in contrast to thehamster (Morin and Blanchard, 1997) and other species(Weber and Hutchins, 1982; Hutchins and Weber, 1985),and 4) the discovery that two additional nuclei (CPT andPLi) are significant constituents of the retinorecipientrodent pretectum (Morin and Blanchard, 1997). Except forthe latter two nuclei, the general pretectal organization
Fig. 12. Photomicrographs showing CT-b–labeled cells in (A) thenuclei of the pretectum (level approximates that of Fig. 1H) and (B)the intergeniculate leaflet (IGL) and ventral lateral geniculate nucleus(VLG) after an injection in the posterior limitans nucleus (PLi; case
96-41; Fig. 1B). C: Darkfield photomicrograph of Phaseolus vulgaris-leucoagglutin (PHA-L)–IR fibers and terminals in the IGL and VLGafter a tracer injection in the PLi (case 95-6; Fig. 2D). For abbrevia-tions, see list. Scale bars 5 500 µm in A, 200 µm in C.
SUBCORTICAL VISUAL CONNECTIONS 301
and nomenclature utilized here are similar to those illus-trated in the Paxinos and Watson (1986) rat atlas, secondedition, which are based upon the work by Scalia (1972;Scalia and Arango, 1979). The nomenclature and anatomyof the atlas have been enhanced by inclusion of the CPTand PLi as pretectal nuclei (Morin and Blanchard, 1997).
Intergeniculate leaflet. Characterization of a retino-recipient IGL two decades ago (Hickey and Spear, 1976)described a thin nucleus intercalated between the DLGand VLG. Subsequent investigation has shown the IGL tobe a relatively large structure with substantial variationin cross-sectional morphology along its length (Morin etal., 1992; Morin and Blanchard, 1995). It is now clear that,according to diverse criteria including location of astro-cytes (Morin et al., 1989; Botchkina and Morin, 1995b),NPY-IR neurons (Morin et al., 1992; Morin and Blanchard,1995, 1997), retinal afferents (Morin et al., 1992; Morinand Blanchard, 1995), location of neurons projecting to theSCN or contralateral IGL (Morin and Blanchard, 1995), aswell the developmental migratory path of neurons and glia(Botchkina and Morin, 1995b), much of what has beenpreviously called VLG should be considered to be IGL. Thisincludes the ventromedial part of the IGL (as in Fig. 5B,C),which contains a few NPY-IR neurons and projects to ipsi-and contralateral SCN (Morin et al., 1992; Morin andBlanchard, 1995). The historical lack of distinction be-tween IGL and VLG, plus the gradual clarification of IGLidentity, has rendered uncertain many previous studies ofVLG connectivity. For example, a study purporting todescribe bilateral sets of ‘‘VLG’’ neurons projecting to the
APT does not acknowledge the existence of an IGL (Fosteret al., 1989). The present data indicate that the IGLprojects bilaterally to the APT, but the VLG provides onlyan ipsilateral connection. If a species difference can beexcluded, the above study (Foster et al., 1989) is probablydescribing IGL afferents to the rat APT as has been shownhere for the hamster.
Taylor et al. (1986) identified IGL efferent and afferentconnections with the SC in the rat. The present data fromthe hamster affirm the presence of these connections. AnIGL-SC pathway has also been reported in the pigeon, andat least part of the projection contains NPY (Gamlin et al.,1996). Our previous data from the hamster (Morin andBlanchard, 1995) and rat (unpublished observations) haveshown a probable NPY- and ENK-IR connection from IGLto PLi. The present data show reciprocal connectionsbetween the PLi and IGL, but there has not yet been adetermination that these contain either NPY or ENK.Double-label analysis of ascending IGL projections affirmthat at least a portion of NPY- and ENK-IR neurons in theIGL project to the pretectal region, but the specific nucleito which these project remain unknown.
Distinguishing IGL from VLG. The IGL and VLG areclearly different according to several criteria. In particular,cells contributing to the geniculohypothalamic tract arenot found in the lateral geniculate complex except in theIGL (Frost et al., 1979; Morin et al., 1992; Botchkina andMorin, 1995b). NPY-IR neurons of the lateral thalamus arefound exclusively in the IGL and project bilaterally to theSCN via the geniculohypothalamic tract (Morin et al.,
Fig. 13. Photomicrographs showing CT-b–labeled neurons in the (A) contralateral and (B) ipsilateralintergeniculate leaflet (IGL) after an injection into the olivary pretectal nucleus (OPT; case 96-38; Fig.1D). Arrows identify neurons in ipsi- and contralateral ventromedial IGL. For abbreviations, see list.Scale bar 5 200 µm.
302 L.P. MORIN AND J.H. BLANCHARD
1992; Botchkina and Morin, 1995b). The IGL also receivesbilateral projections from the precommissural nucleus, butthe VLG has no precommissural nucleus afferents. Bilat-eral connectivity is a defining characteristic of the IGLthat fully distinguishes the IGL from the VLG: Everynucleus innervated by the ipsilateral IGL is also inner-
vated by the contralateral IGL (Fig. 16). In contrast, theVLG projects bilaterally only to the CPT. Despite this largedifference, the ipsilateral patterns of subcortical visualsystem innervation by the VLG and IGL are very similar(Table 1). It is particularly striking that there is noipsilateral visual nucleus to which the IGL projects towhich the VLG does not also project.
A set of ENK-IR neurons is present immediately adja-cent to the ventromedial IGL in the medial VLG (Morinand Blanchard, 1995). These do not project to the SCN, butmay project to the OPT. We have previously shown thatinjection of a retrograde label into a pretectal area thatincludes the OPT labels numerous neurons in the IGL andVLG (Morin and Blanchard, 1995). The current datainclude case 96-38 which had a small, precise injectionalmost exclusively in the non-bifurcated part of the rostralOPT. The pattern of retrogradely labeled neurons in theVLG closely mimics the distribution of ENK-IR neuronsshown by Morin and Blanchard (1995; their Fig. 3B–D).This would be consistent with the evidence showing fairlydense enkephalinergic innervation of the OPT (Morin andBlanchard, 1997) and presence of VLG neurons containingENK-IR that project to the region of the OPT. It is also thecase that both NPY- and ENK-IR neurons project to thePLi (Morin and Blanchard, 1995). Whether the peptide-IRneurons giving rise to this projection are located in the IGLor VLG or both has not yet been fully tested. NPY and ENKare obvious candidate peptides to be found in any of theIGL efferent projections to subcortical visual nuclei. It isalso likely, however, that the predominant neuromodula-
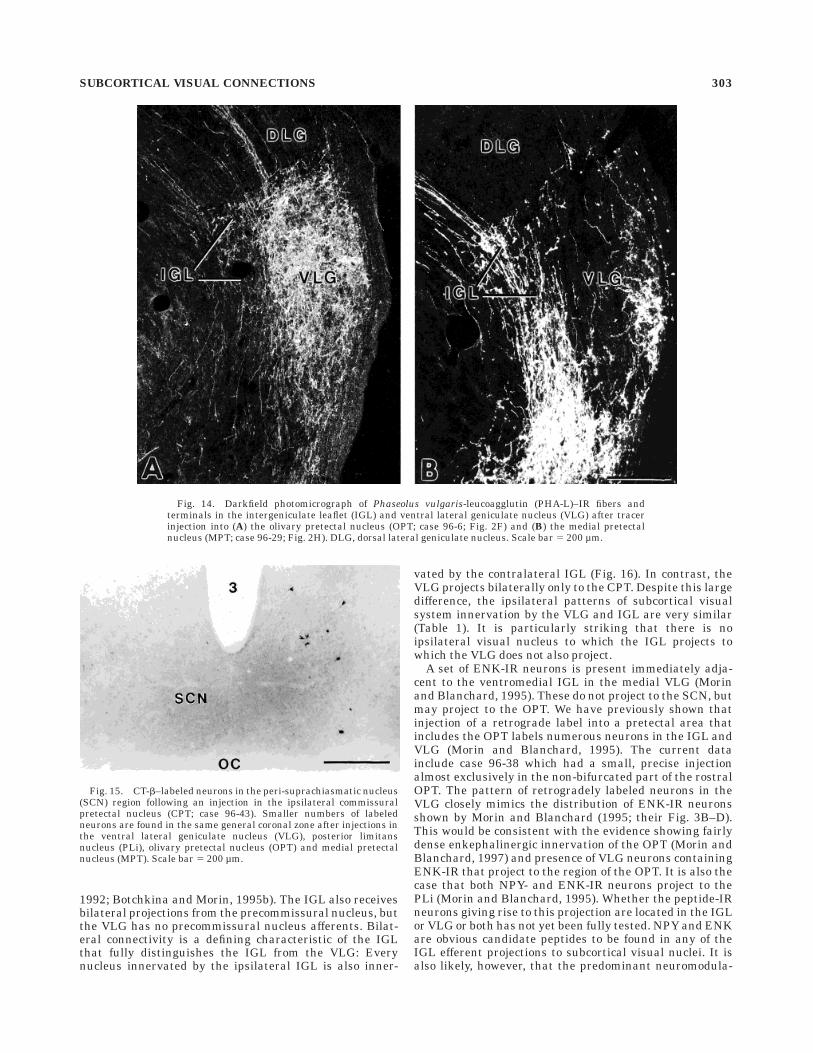
Fig. 14. Darkfield photomicrograph of Phaseolus vulgaris-leucoagglutin (PHA-L)–IR fibers andterminals in the intergeniculate leaflet (IGL) and ventral lateral geniculate nucleus (VLG) after tracerinjection into (A) the olivary pretectal nucleus (OPT; case 96-6; Fig. 2F) and (B) the medial pretectalnucleus (MPT; case 96-29; Fig. 2H). DLG, dorsal lateral geniculate nucleus. Scale bar 5 200 µm.
Fig. 15. CT-b–labeled neurons in the peri-suprachiasmatic nucleus(SCN) region following an injection in the ipsilateral commissuralpretectal nucleus (CPT; case 96-43). Smaller numbers of labeledneurons are found in the same general coronal zone after injections inthe ventral lateral geniculate nucleus (VLG), posterior limitansnucleus (PLi), olivary pretectal nucleus (OPT) and medial pretectalnucleus (MPT). Scale bar 5 200 µm.
SUBCORTICAL VISUAL CONNECTIONS 303
tor of IGL neurons is GABA as it is for the geniculohypotha-lamic tract of the rat (Moore and Speh, 1993).
Pretectum and tectum. Three nuclei (OPT, NOT, PPT)have been considered to be the retinorecipient constituentsof the rat visual pretectum (Scalia, 1972; Scalia andArango, 1979). The MPT and APT have also been identifiedas retinorecipient in some non-rodent species (Giolli andGuthrie, 1969; Weber and Hutchins, 1982; Hutchins andWeber, 1985; Hutchins, 1991). The VLG and the NOT arethe only nuclei of the subcortical visual shell with previ-ously demonstrated projections to the OPT (Graybiel,1974; Swanson et al., 1974; Ribak and Peters, 1975; Legg,1979; Terasawa et al., 1979; Sefton and Dreher, 1995). TheVLG has also been shown to project to the NOT and APT(Graybiel, 1974; Swanson et al., 1974; Ribak and Peters,1975), whereas the NOT has been shown to also project tothe contralateral NOT (Terasawa et al., 1979). The presentdata support and extend those observations to includeVLG projections to all pretectal nuclei, whereas the NOT
projects to all but the PLi, APT, and MPT. Efferents of thepretectum have been identified as projecting to the DLG(from APT—Turlejski et al., 1993; Young and Lund, 1997;from OPT—Mackay-Sim et al., 1983; Turlejski et al., 1993;from NOT—Mackay-Sim et al., 1983; Turlejski et al.,1993), the VLG (from APT, OPT, NOT—Mackay-Sim et al.,1983; Young and Lund, 1997), the NOT (from NOT—Terasawa et al., 1979), the OPT (from NOT—Terasawa etal., 1979), and the APT (from PPT—Robertson, 1983).Specific evaluation of the rat IGL using PHAL methodshas revealed that it also receives a direct projection fromthe OPT (Klooster et al., 1995). The present study gener-ally confirms these connections, with the exception of thosefrom the APT and NOT to the DLG. As indicated in Table 1,there are extensive, additional interconnections identifiedin the hamster pretectum.
The rat APT (Bickford and Hall, 1989; Foster et al.,1989), NOT, and PPT (Takahashi, 1985) have been shownto have afferents from the SC. In the present data, all
Fig. 16. Schematic illustration of intergeniculate leaflet (IGL) efferent (A) and afferent (B) connec-tions with nuclei of the hamster subcortical visual shell. For abbreviations, see list.
304 L.P. MORIN AND J.H. BLANCHARD

pretectal nuclei receive bilateral projections from the SC,except for the APT, for which it is only ipsilateral. Pretectalefferents to the SC have been identified as originating inthe OPT and NOT (Beitz, 1989). In the present study withthe hamster, all pretectal nuclei are shown to project to theSC, and most of these connections are bilateral (Table 1).Similarly, the data confirm the existence of SC afferentsfrom the VLG and IGL (Graybiel, 1974; Swanson et al.,1974; Ribak and Peters, 1975; Legg, 1979; Taylor et al.,1986) and SC efferents to these locations (Pasquier andVillar, 1982; Mackay-Sim et al., 1983; Takahashi, 1985;Taylor et al., 1986). The SC is also widely known to projectto the DLG and LP (Robson and Hall, 1976; Crain andHall, 1980; Perry, 1980; Mason and Groos, 1981; Pasquierand Villar, 1982; Mooney et al., 1984; Takahashi, 1985;Taylor et al., 1986; Turlejski et al., 1993), and theseconnections are confirmed here. The present studies do notreveal any SC afferents from DLG or LP, in agreementwith the rat literature (Sefton and Dreher, 1995).
The PLi is known to receive afferents from the inferiorcolliculus (Ledoux et al., 1985), and there is a moderatelydense serotonergic projection to it from the dorsal raphenucleus. Other connections have not been described. Theregion of the CPT is unnamed in the Paxinos-Watson atlas(1986), and little is known about it. As is the case for theother pretectal nuclei, the hamster CPT receives directretinal projections and does not project to cortex (Morinand Blanchard, 1997). The present data show that the CPTalso has connectivity attributes similar to the other subcor-tical visual nuclei. In particular, it is reciprocally con-nected to the ipsi- and contralateral IGL. This characteris-tic suggests that the ‘‘mesencephalic pretectal gray’’ regionof the frog may be homologous to the CPT because itprojects bilaterally to the nucleus of Bellonci and anteriorlateral thalamic nucleus, which form the presumptivehomologue of the IGL (Montgomery et al., 1991; Montgom-ery and Fite, 1991). These characteristics reinforce theprevious suggestion that the CPT should be considered oneof the pretectal nuclei.
The MPT has been differentiated from adjacent nucleiaccording to the absence of NPY- and ENK-IR neurons andthe absence of NPY-IR terminals (Morin and Blanchard,1997). The present data further distinguish the MPT,showing that it has a dense concentration of neuronsprojecting specifically to the CPT. In addition, the hamsterMPT is distinguished by the presence of a very denseserotonin-IR terminal plexus which contrasts with a muchless dense level of innervation in adjacent PPT and CPT(Morin and Meyer-Bernstein, unpublished observations).The MPT has not been considered part of the rodent visualpretectum until recently (Morin and Blanchard, 1997),although it is retinorecipient in other species (Weber andHutchins, 1982; Hutchins and Weber, 1985). Additionalanatomical or functional information about the area ap-pears to be absent.
Thalamocortical visual nuclei. The DLG is widelyreported to receive projections from the OPT and SC(Robson and Hall, 1976; Mackay-Sim et al., 1983; Kubotaet al., 1987; Harting et al., 1991; Turlejski et al., 1993;Uhlrich and Manning, 1995), results supported by thepresent data. However, the large number of labeled neu-rons reported in at least four pretectal nuclei after place-ment of a retrograde tracer in the rat DLG (Turlejski et al.,1993) is not consistent with the present data nor is itconsistent with other studies in the rat (Mackay-Sim et al.,
1983) showing a lesser projection from the OPT and NOT,or with a PHA-L anterograde study that failed to identifyOPT projections to the DLG (Klooster et al., 1995). In thepresent study, a distinctively clustered, dense group ofneurons was visible in the lateral superficial gray layer ofthe SC after injection of CT-b into centrocaudal DLG. Thegeneral view that SC projects to DLG was confirmed herewith PHA-L injections into the SC, although the injectionswere largely in the central optic layer with projections toDLG found more rostral than the DLG sites into which theretrograde label was injected. Previously, Harting et al.(1991) applied anterograde tracer to the hamster superfi-cial gray layer (plus some optic layer). This injection wasmore medial than the present PHA-L injection sites andlabeled a dense terminal plexus in rostroventral DLG. Aninjection of retrograde tracer into the rat dorsal DLGlabeled many neurons in the central superficial gray layer(Harting et al., 1991). The differences between the presentresults and the earlier studies with rat and hamsterprobably relate to the specific injection sites and are thusindications of topographic organization of the tectogenicu-late projections.
There have apparently been few previous reports of LPconnections with the pretectum in the rat. A projectionfrom the pretectal region to the LP has been described(Schober, 1981), as well as a specific projection from theNOT to the LP (Mason and Groos, 1981). The present datado not support the existence of an NOT-LP pathway, but doindicate that the LP receives projections from the OPT,PPT, and MPT. No LP efferent projections to the pretectumor SC were seen in the present hamster material, althougha projection to the SC has been reported in the rat (Tayloret al., 1986; Beitz, 1989).
The IMA has received little attention as a distinctsubdivision of the LP. Takahashi (1985) describes it as athin lamination between the rat LP and DLG, originallydesignated the nucleus pulvinaris pars geniculata of therat (Lund and Webster, 1967) and sheep (Rose, 1942). TheIMA receives cortical projections from secondary visualarea 18a, but none from area 17 (Takahashi, 1985).Previous anatomical studies of rat LP nucleus efferents tocortex have not identified the IMA as a separate region,but the data suggest projections to area 18a (Hughes,1975; Coleman and Clerici, 1980; Warton et al., 1988). Thedata support the existence of the IMA and further demon-strate that it projects to area 18a, but not to area 17(present results and Morin and Blanchard, 1997). Thus,the IMA is a thalamic structure in accordance with thegeneral rule that thalamic sensory nuclei project to cortex,but pretectal nuclei do not. In contrast to the report byTakahashi (Takahashi, 1985), however, the present workshows a robust projection from the SC to the IMA. Inaddition, although Karamanlidis and Magras (1972) re-port a retinal projection to the sheep IMA, Takahashi(1985) cites unpublished observations denying such aprojection in the rat. The hamster has a contralateral, butnot an ipsilateral, retinal projection to the IMA (Blanchardand Morin, unpublished observations). The present resultsalso show a limited degree of connectivity between thesubcortical visual nuclei and the LP or IMA. The latterreceives extensive projections from the ventral nuclei(VLG, IGL) as well as the APT and NOT. The LP hasafferents from most of the dorsomedial pretectal nuclei(OPT, PPT, MPT). Only the SC projects to both. Thedifferent patterns of subcortical visual afferents to LP and
SUBCORTICAL VISUAL CONNECTIONS 305

IMA suggest that the IMA should be considered a nucleusdistinct from the LP, rather than a subdivision of it.
Bilaterality of connections between visual nuclei.
As indicated above, all nuclei of the subcortical visual shellto which the IGL projects ipsilaterally also receive innerva-tion from the contralateral IGL. Several other nuclei arealso strongly bilateral in their projections (Table 1). Inrank order, they are the IGL (100%), CPT (90%), SC (75%),OPT (58%), PPT (55%), MPT (36%), NOT (33%), PLi (10%),APT (10%), VLG (10%), LP (0), and DLG (0). Bilateralityappears to be a general property of pretectal and ventralthalamic visual nuclei that contrasts with the absence ofsuch from the thalamic relay nuclei projecting to the visualcortex.
Connections with the suprachiasmatic nucleus. Thegeniculohypothalamic tract projects robustly from the IGLto the SCN (Morin et al., 1992; Morin and Blanchard,1995), but there are few other SCN afferent projectionsfrom the subcortical visual nuclei revealed either byPHA-L tract tracing (present data) or retrograde labelinjected into the SCN (unpublished data). In contrast, therat has a distinct projection from the pretectum to the SCNthat has been identified by both anterograde and retro-grade methods (Mikkelsen and Vrang, 1994). The PHA-Lmethod has revealed the presence of sparse projectionsfrom the SCN to the OPT (Morin et al., 1994) in thehamster. The present data suggest the existence of morerobust projections from the peri-SCN region to the medialpretectal nuclei as well as the ventrolateral geniculateregion. In contrast to the peri-SCN pattern of cells ob-served in the hamster, the rat appears to have intra-SCNneurons projecting to the IGL/VLG (Card and Moore,1989).
Functional considerations
Dorsal thalamus. The DLG contributes to the com-plex operations of visual stimulus analysis (Sherman andGuillery, 1996). Much less information is available concern-ing the function of the LP. A comprehensive evaluation ofthe hamster LP suggests that it lacks topographic organi-zation (Mooney et al., 1984). The LP may relay visualinformation from the tectum to cortex, but not in the samemanner as the DLG (see Mooney et al., 1984). In addition,the hamster LP appears to contain neurons which areexclusively somatosensory as well as others that arevisual-somatosensory (Mooney et al., 1984). The observedinputs from the OPT, MPT, and PPT may contributeinformation from multiple sensory modalities. It has beenproposed that the LP (Mooney et al., 1984), unlike theDLG, may send information regarding the occurrence ofnovel visual stimuli to extrastriate cortex.
Pretectum and tectum. The most thoroughly evalu-ated phenomenon related to pretectal function is thepupillary light reflex (see Young and Lund, 1997, for areview). The OPT is thought to be central to the photicregulation of pupillary constriction (Gamlin and Clarke,1995; Young and Lund, 1997). Retinotopically organized,tonic-on neurons are found throughout the OPT (Scaliaand Arango, 1979), and stimulation of this site, but notothers, causes pupillary constriction (Trejo and Cicerone,1984). A class of luminance detector neurons showinggraded increases in firing rate with increased illuminationhas been found in the OPT, and a second class in the PPTshows similar rate increases, but in response to decreasedillumination (Clarke and Ikeda, 1985). In each case,
change in firing rate is accompanied by change in pupilsize, suggesting that the PPT mediates pupillodilation andthe OPT mediates pupilloconstriction. The pupillary lightreflex is a consensual phenomenon meaning that exposureof one eye to light results in both pupils being constricted.This requires bilateral communication of the photic infor-mation to the constrictor muscles of the iris. Bilateralefferents from the OPT to the nucleus of Edinger-Westphalmay serve this function (Gamlin and Clarke, 1995; Youngand Lund, 1997). However, the present data show a highlycomplex arrangement of bilateral interconnections amongnuclei also receiving bilateral retinal projections. Thus,consensual optomotor responses could conceivably be modu-lated by a variety of nuclei in the subcortical visual shell(cf., Legg, 1975).
Visual responsiveness of several NOT neuron classeshas been studied, but the contribution of these classes tovisual function is not known. The NOT appears to partici-pate in the regulation of optokinetic nystagmus (Simpsonet al., 1988; Ilg and Hoffmann, 1996). Its cells respondpreferentially to stimulus movement, orientation, direc-tion, or speed (Collewijn, 1975; Maekawa et al., 1984;Ballas and Hoffmann, 1985; Natal and Britto, 1987; Ilgand Hoffmann, 1996). The retinal projection to these cellsis thought to be primarily from W class ganglion cells(Hoffmann and Stone, 1985). Given the historical confu-sion concerning the anatomical definition of the NOT(Simpson et al., 1988), its potential confusion with the PLi(Morin and Blanchard, 1997), and the general absence ofinformation concerning the PLi, functions attributed tothe NOT should be treated with caution.
We have previously shown direct retinal innervation ofthe dorsal APT in the hamster (Morin and Blanchard,1997). This, plus its connections with the VLG, SC, andzona incerta (present data and Cadusseau and Roger,1991; May et al., 1997) suggest strong primary and second-ary visual modulation of APT function in the hamster. Thisconclusion is supported by the presence of afferents fromthe visual cortex (Nauta and Bucher, 1954; Lent, 1982;Cadusseau and Roger, 1991). The various visual systeminputs enter the dorsal APT in the rat or hamster (presentdata and Cadusseau and Roger, 1991; Morin and Blan-chard, 1997). The present data also show that retrogradelylabeled neurons in the ventral APT project to the dorsalAPT (e.g., Fig. 9). This would suggest interaction of visualinformation with the somatosensory information pro-cessed in the ventral division (Yezierski, 1988). In general,the APT has not been studied as part of the subcorticalvisual system. Numerous reports indicate that it modifiessomatosensory input and has a particular antinociceptivecapability (see Rees and Roberts, 1993, for a review).
Little is known concerning function of the PLi. The fewconnectivity studies suggest that the PLi may processsomasthetic (Ledoux et al., 1987) and acoustic (Ledoux etal., 1985) information. Direct innervation of the hamsterPLi by retinal ganglion cells (Morin and Blanchard, 1997)and IGL neurons (present data) also indicates a visualfunction. This is emphasized by its afferent connectionswith all pretectal nuclei except the NOT and by its efferentprojections to all pretectal nuclei (present data). It appearslikely that the PLi serves multiple stimulus modalities.
The SC is divided into two distinct functional units.Neurons in the superficial laminae (optic layer and above)are considered to be exclusively responsive to visual input,whereas those of the deep laminae (internal gray layer and
306 L.P. MORIN AND J.H. BLANCHARD

below) are responsive to multiple sensory modalities,including visual, auditory, and somatosensory stimuli(Tiao and Blakemore, 1976; Finley et al., 1978; Rhoades,1981; Stein, 1984). The present data show three distinctgroups of subcortical visual nuclei receiving projectionsfrom the SC: 1) projections largely from the superficiallayers (DLG, LP, MPT, PPT), 2) projections largely fromthe deep layers (IGL, CPT), and 3) roughly equal projec-tions from superficial and deep layers (VLG, PLi, APT,OPT, NOT). In general, all the subcortical visual nucleistudied here receive projections from the visual and multi-modal sensory areas of the SC, allowing the possibilitythat information related to multiple sensory modalities isgaining widespread access to the subcortical visual shell.The nuclei of the visual shell may then contribute to motorcontrol of eye movements and orienting behaviors (Finleyet al., 1978; Sparks, 1990).
Intergeniculate leaflet. Perhaps the most fascinatingresult detailed here concerns the widespread efferent andafferent connections between the IGL and the subcorticalvisual system. The IGL is considered one of the two majornuclei of the circadian visual system (see Morin, 1994, for areview). It projects to the SCN, site of the circadian clock,via the geniculohypothalamic tract which, in the hamster,contains NPY and ENK as neuromodulators. A rhythm-regulating function is supported by studies showing that ifthe IGL is stimulated electrically (Rusak et al., 1989) orchemically (Johnson et al., 1989), phase shifts occur whichare consistent with those produced by direct application ofNPY to the SCN (Albers and Ferris, 1984; Rusak et al.,1990). IGL lesions reduce the rate of re-entrainment to ashifted light–dark cycle (Johnson et al., 1989), eliminatethe increase in circadian period length during constantlight (Harrington and Rusak, 1986; Pickard et al., 1987),and eliminate the ability of benzodiazepine to inducecircadian rhythm phase shifts (Johnson et al., 1988; Bielloet al., 1991; Meyer et al., 1993). Cells of the IGL areluminance detectors, although cells may be monotonic ornon-monotonic (Harrington and Rusak, 1989, 1991). Giventhe apparent complexities of IGL efferent and afferentconnections, it is not certain to what extent IGL neuronsrespond directly to light or indirectly through other subcor-tical visual nuclei.
The visual system connections of the IGL beyond thoseto the SCN and contralateral IGL suggest that the leaflethas a broader function than regulation of circadianrhythms. One hint of this role may come from datashowing rhythm shifts in response to novel wheel access orsocial stimuli (Mrosovsky, 1988; Reebs and Mrosovsky,1989a,b). Such phase shifts are blocked by IGL lesions(Janik and Mrosovsky, 1994; Wickland and Turek, 1994)and are also eliminated by depletion of brain NPY (Bielloet al., 1994). The IGL does not contain GABAA/benzodiaz-epine receptors (Michels et al., 1990), suggesting that thedrug and novel wheel-induced phase shifts may provideinformation to the SCN circadian clock through the IGLand geniculohypothalamic tract, but have their primarysite of action elsewhere. Candidates would be the SC orany of the pretectal nuclei with connections afferent to theIGL. The geniculohypothalamic tract connection from IGLto SCN may serve as the final common path by whichmultimodal, species-specific, secondary entraining stimuliexert circadian rhythm phase control.
The widespread innervation of the subcortical visualsystem by the IGL raises substantial questions concerning
non-rhythm functions of the system. Clearly, the IGLconnections to the OPT suggest a capacity to influence thepupillary light reflex. However, there seems to be rela-tively little specificity of the IGL efferent pathways withinthe subcortical visual system. In fact, innervation of thesubcortical visual nuclei by both ipsi- and contralateralIGL projections plus direct input from ipsi- and contralat-eral retina appears to be a major characteristic of thisportion of the system. In a similar vein, the pervasivebilaterality of IGL-efferent connections is a most signifi-cant and interesting feature. These characteristics un-doubtedly have functional importance for circadian rhythmor oculomotor regulation that remain to be determined.Similarly, the emerging pattern of extensive connectionsbetween the IGL and non-visual forebrain and midbrainnuclei (Horvath, 1996; Moore et al., 1996; Vrang andMikkelsen, 1996) is an interesting anatomical feature thatpresently defies functional explanation.
ACKNOWLEDGMENTS
This study was supported by NINDS grant NS22168 toL.P.M.
LITERATURE CITED
Albers, H.E. and C.F. Ferris (1984) Neuropeptide Y: role in light-darkentrainment of hamster circadian rhythms. Neurosci. Lett. 50:163–168.
Ballas, I. and K.-P. Hoffmann (1985) A correlation between receptive fieldproperties and morphological structures in the pretectum of the cat. J.Comp. Neurol. 238:417–428.
Beitz, A.J. (1989) Possible origin of glutamatergic projections to themidbrain periaqueductal gray and deep layer of the superior colliculusof the rat. Brain Res. Bull. 23:25–35.
Berman, N. (1977) Connections of the pretectum in the cat. J. Comp.Neurol. 174:227–254.
Bickford, M.E. and W.C. Hall (1989) Collateral projections of predorsalbundle cells of the superior colliculus in the rat. J. Comp. Neurol.283:86–106.
Biello, S.M., M.E. Harrington, and R. Mason (1991) Geniculo-hypothalamictract lesions block chlordiazepoxide-induced phase advances in Syrianhamsters. Brain Res. 552:47–52.
Biello, S.M., D. Janik, and N. Mrosovsky (1994) Neuropeptide Y andbehaviorally induced phase shifts. Neuroscience 62:273–279.
Botchkina, G.I. and L.P. Morin (1995a) Organization of permanent andtransient neuropeptide Y-immunoreactive neuron groups and fibersystems in the developing hamster diencephalon. J. Comp. Neurol.357:573–602.
Botchkina, G.I. and L.P. Morin (1995b) Specialized neuronal and glialcontributions to development of the hamster lateral geniculate complexand circadian visual system. J. Neurosci. 15:190–201.
Cadusseau, J. and M. Roger (1991) Cortical and subcortical connections ofthe pars compacta of the anterior pretectal nucleus in the rat. Neurosci.Res. 12:83–100.
Card, J.P. and R.Y. Moore (1989) Organization of lateral geniculate-hypothalamic connections in the rat. J. Comp. Neurol. 284:135–147.
Chen, S. and G. Aston-Jones (1995) Evidence that cholera toxin B subunit(CTb) can be avidly taken up and transported by fibers of passage.Brain Res. 674:107–111.
Clarke, R.J. and H. Ikeda (1985) Luminance and darkness detectors in theolivary and posterior pretectal nuclei and their relationship to thepupillary light reflex in the rat. I. Studies with steady luminance levels.Exp. Brain Res. 57:224–232.
Cliffer, K.D. and G.J. Giesler, Jr. (1988) PHA-L can be transportedanterogradely through fibers of passage. Brain Res. 458:185–191.
Coleman, J. and W.J. Clerici (1980) Extrastriate projections from thalamusto posterior occipital-temporal cortex in rat. Brain Res. 194:205–209.
Collewijn, H. (1975) Direction-selective units in the rabbit’s nucleus of theoptic tract. Brain Res. 100:489–508.
Crain, B.J. and W.C. Hall (1980) The normal organization of the lateralposterior nucleus of the golden hamster. J. Comp. Neurol. 193:351–370.
SUBCORTICAL VISUAL CONNECTIONS 307

Finley, J.C.W., S.E. Schneps, K.G. Wilson, and G.E. Schneider (1978)Topography of visual and somatosensory projections to the superiorcolliculus of the golden hamster. Brain Res. 142:223–235.
Foster, G.A., A.R. Sizer, H. Rees, and M.H.T. Roberts (1989) Afferentprojections to the rostral anterior pretectal nucleus of the rat: a possiblerole in the processing of noxious stimuli. Neuroscience 29:685–694.
Frost, D.O., K.-F. So, and G.E. Schneider (1979) Postnatal development ofretinal projections in Syrian hamsters: A study using autoradiographicand anterograde degeneration techniques. Neuroscience 4:1649–1677.
Gamlin, P.D.R. and R.J. Clarke (1995) The pupillary light reflex pathway ofthe primate. J. Am. Optometric Assoc. 66:415–418.
Gamlin, P.D.R., A. Reiner, K.T. Keyser, N. Brecha, and H.J. Karten (1996)Projection of the nucleus pretectalis to a retinorecipient tectal layer inthe pigeon (Columbia livia). J. Comp. Neurol. 368:424–438.
Gerfen, C.R. and P.E. Sawchenko (1984) An anterograde neuroanatomicaltracing method that shows the detailed morphology of neurons, theiraxons and terminals: immunohistochemical localization of an axonallytransported plant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L).Brain Res. 290:219–238.
Giolli, R.A. and M.D. Guthrie (1969) The primary optic projections in therabbit. An experimental degeneration study. J. Comp. Neurol. 36:96–126.
Graybiel, A.M. (1974) Visuo-cerebellar and cerebello-visual connectionsinvolving the ventral lateral geniculate nucleus. Exp. Brain Res.20:303–306.
Harrington, M.E. and B. Rusak (1986) Lesions of the thalamic intergenicu-late leaflet alter hamster circadian rhythms. J. Biol. Rhythms 1:309–325.
Harrington, M.E. and B. Rusak (1989) Photic responses of geniculo-hypothalamic tract neurons in the Syrian hamster. Vis. Neurosci.2:367–375.
Harrington, M.E. and B. Rusak (1991) Luminance coding properties ofintergeniculate leaflet neurons in the golden hamster and the effects ofchronic clorgyline. Brain Res. 554:95–104.
Harting, J.K., M.F. Huerta, T. Hashikawa, and D.P. Van Lieshout (1991)Projection of the mammalian superior colliculus upon the dorsal lateralgeniculate nucleus: Organization of tectogeniculate pathways in nine-teen species. J. Comp. Neurol. 304:275–306.
Hickey, T.L. and P.D. Spear (1976) Retinogeniculate projections in hoodedand albino rats: An autoradiographic study. Exp. Brain Res. 24:523–529.
Hoffmann, K.-P. and J. Stone (1985) Retinal input to the nucleus of the optictract of the cat assessed by antidromic activation of ganglion cells. Exp.Brain Res. 59:395–403.
Horvath, T.L. (1996) Evidence that circadian and visual signals from theSCN and LGN are integrated on neuroendocrine dopamine cells in therat hypothalamus. Soc. Neurosci. 26:31.
Hsu, S.-M., L. Raine, and H. Fanger (1981) Use of avidin-biotin-peroxidasecomplex (ABC) in immunoperioxidase techniques: A comparison be-tween ABC and unlabeled antibody (PAP) procedures. J. Histochem.Cytochem. 29:577–580.
Huerta, M.F., and J.K. Harting (1984) The mammalian superior colliculus:studies of its morphology and connections. In H. Vanegas (ed): Compara-tive Neurology of the Optic Tectum. New York: Plenum, pp. 687–773.
Hughes, H.C. (1975) Anatomical and neurobehavioral investigations con-cerning the thalamo-cortical organization of the rat’s visual system. J.Comp. Neurol. 175:311–336.
Hutchins, B. (1991) Evidence for a direct retinal projection to the anteriorpretectal nucleus in the cat. Brain Res. 561:169–173.
Hutchins, B. and J.T. Weber (1985) The pretectal complex of the monkey: areinvestigation of the morphology and retinal terminations. J. Comp.Neurol. 232:425–442.
Ilg, U.J. and K.P. Hoffmann (1996) Responses of neurons of the nucleus ofthe optic tract and the dorsal terminal nucleus of the accessory optictract in the awake monkey. Eur. J. Neurosci. 8:92–105.
Janik, D. and N. Mrosovsky (1994) Intergeniculate leaflet lesions andbehaviorally-induced shifts of circadian rhythms. Brain Res. 651:174–182.
Johnson, R.F., L. Smale, R.Y. Moore, and L.P. Morin (1988) Lateralgeniculate lesions block circadian phase shift responses to a benzodiaz-epine. Proc. Natl. Acad. Sci. U.S.A. 85:5301–5304.
Johnson, R.F., R.Y. Moore, and L.P. Morin (1989) Lateral geniculate lesionsalter activity rhythms in the hamster. Brain Res. Bull. 22:411–422.
Karamanlidis, A.N. and J. Magras (1972) Retinal projection in domesticungulates. I. The retinal projections in sheep and pig. Brain Res.44:127–145.
Klooster, J., G.F.J.M. Vrensen, L.J. Muller, and J.J.L. Van der Want (1995)Efferent projections of the olivary pretectal nucleus in the albino ratsubserving the pupillary light reflex and related reflexes. A lightmicroscopic tracing study. Brain Res. 688:34–46.
Kubota, T., M. Morimoto, T. Kanaseki, and J. Inomata (1987) Projectionfrom the pretectal nuclei to the dorsal lateral geniculate nucleus in thecat: A wheat germ agglutinin-horseradish peroxidase study. Brain Res.421:30–40.
Kunzle, H. (1996) Diencephalic connections of the superior colliculus in thehedgehog tenrec. Exp. Brain Res. 111:36–370.
Ledoux, J.E., D.A. Ruggiero, and D.J. Reis (1985) Projections to thesubcortical forebrain from anatomically defined regions of the medialgeniculate body in the rat. J. Comp. Neurol. 242:182–213.
Ledoux, J.E., D.A. Ruggiero, R. Forest, R. Stornetta, and D.J. Reis (1987)Topographic organization of convergent projections to the thalamusfrom the inferior colliculus and spinal cord in the rat. J. Comp. Neurol.264:123–146.
Legg, C.R. (1975) Effects of subcortical lesions on the pupillary light reflexin the rat. Neuropsychologia 13:373–376.
Legg, C.R. (1979) An autoradiographic study of the efferent projections ofthe ventral lateral geniculate nucleus of the hooded rat. Brain Res.170:349–352.
Lent, R. (1982) The organization of subcortical projections of the hamster’svisual cortex. J. Comp. Neurol. 206:227–242.
Lund, R.D. and K.E. Webster (1967) Thalamic afferents from the dorsalcolumn nuclei. An experimental anatomical study in the rat. J. Comp.Neurol. 130:301–312.
Mackay-Sim, A., A.J. Sefton, and P.R. Martin (1983) Subcortical projectionsto lateral geniculate and thalamic reticular nuclei in the hooded rat. J.Comp. Neurol. 213:24–35.
Maekawa, K., T. Takeda, and M. Kimura (1984) Responses of the nucleus ofthe optic tract neurons projecting to the nucleus reticularis tegmentipontis upon optokinetic stimulation in the rabbit. Neurosci. Res.2:1–25.
Martin, J.R., P. Schoch, F. Jenck, J. Moreau, and W.E. Haefely (1993)Pharmacological characterization of benzodiazepine receptor ligandswith intrinsic efficacies ranging from high to zero. Psychopharmacology111:415–422.
Mason, R. and G.A. Groos (1981) Cortico-recipient and tecto-recipientvisual zones in the rat’s lateral posterior (pulvinar) nucleus: Ananatomical study. Neurosci. Lett. 25:107–112.
May, P.G., W. Sun, and W.C. Hall (1997) Reciprocal connections between thezona incerta and the pretectum and superior colliculus of the cat.Neuroscience 77:1091–1114.
McLean, I.W. and P.K. Nakane (1974) Periodate-lysine-paraformaldehydefixative: a new fixative for immunoelectron microscopy. J. Histochem.Cytochem. 22:1077–1083.
Meijer, J.H. and W.J. Rietveld (1989) Neurophysiology of the suprachias-matic circadian pacemaker in rodents. Physiol. Rev. 69:671–707.
Meyer, E.L., M.E. Harrington, and T. Rahamani (1993) A phase-responsecurve to the benzodiazepine chlordiazepoxide and the effect of geniculo-hypothalamic tract ablation. Physiol. Behav. 53:237–243.
Michels, K.M., L.P. Morin, and R.Y. Moore (1990) GABAA/benzodiazepinereceptor localization in the circadian timing system. Brain Res. 531:16–24.
Mikkelsen, J.D. (1992) The organization of the crossed geniculogeniculatepathway of the rat: A Phaseolus vulgaris-leucoagglutinin study. Neuro-science 48:953–962.
Mikkelsen, J.D. and N. Vrang (1994) A direct pretectosuprachiasmaticprojection in the rat. Neuroscience 62:497–505.
Montgomery, N.M. and K.V. Fite (1991) Organization of ascending projec-tions from the optic tectum and mesencephalic pretectal gray in Ranapipiens. Vis. Neurosci. 7:459–478.
Montgomery, N.M., K.V. Fite, and Z. Li (1991) Anatomical evidence for anintergeniculate leaflet in Rana pipiens. Neurosci. Lett. 133:105–108.
Mooney, R.D., S.E. Fish, and R.W. Rhoades (1984) Anatomical and func-tional organization of pathway from superior colliculus to lateralposterior nucleus in hamster. J. Neurophysiol. 51: 407–431.
Moore, R.Y. and J.C. Speh (1993) GABA is the principal neurotransmitter ofthe circadian system. Neurosci. Lett. 150:112–116.
Moore, R.Y., M.M. Moga, and R. Weis (1996) Intergeniculate leaflet andventral lateral geniculate projections in the rat. Soc. Neurosci. 26:31.
Morin, L.P. (1994) The circadian visual system. Brain Res. Rev. 67:102–127.
308 L.P. MORIN AND J.H. BLANCHARD

Morin, L.P. and J. Blanchard (1995) Organization of the hamster interge-niculate leaflet: NPY and ENK projections to the suprachiasmaticnucleus, intergeniculate leaflet and posterior limitans nucleus. Vis.Neurosci. 12:57–67.
Morin, L.P. and J. Blanchard (1997) Neuropeptide Y and enkephalinimmunoreactivity in retinorecipient nuclei of the hamster pretectumand thalamus. Vis. Neurosci. 14:765–777.
Morin, L.P., R.F. Johnson, and R.Y. Moore (1989) Two brain nucleiregulating circadian rhythms are identified by GFAP immunoreactivityin hamsters and rats. Neurosci. Lett. 99:55–60.
Morin, L.P., J.H. Blanchard, and R.Y. Moore (1992) Intergeniculate leafletand suprachiasmatic nucleus organization and connections in thehamster. Vis. Neurosci. 8:219–230.
Morin, L.P., N. Goodless-Sanchez, L. Smale, and R.Y. Moore (1994) Projec-tions of the suprachiasmatic nuclei, subparaventricular zone andretrochiasmatic area in the golden hamster. Neuroscience 61:391–410.
Mrosovsky, N. (1988) Phase response curves for social entrainment. J.Comp. Physiol. 162:35–46.
Natal, C.L. and L.R.G. Britto (1987) The pretectal nucleus of the optic tractmodulates the direction and selectivity of accessory optic neurons inrats. Brain Res. 419:320–323.
Nauta, V.J.H. and V.M. Bucher (1954) Efferent connections of the striatecortex in the albino rat. J. Comp. Neurol. 100:257–286.
Pasquier, D.A. and M.J. Villar (1982) Specific serotonergic projectins to thelateral geniculate body from the lateral cell groups of the dorsal raphenucleus. Brain Res. 249:142–146.
Paxinos, G. and C. Watson (1986) The Rat Brain in Stereotaxic Coordi-nates. New York: Academic.
Perry, V.H. (1980) A tectocortical visual pathway in the rat. Neuroscience5:915–927.
Pickard, G.E., M.R. Ralph, and M. Menaker (1987) The intergeniculateleaflet partially mediates effects of light on circadian rhythms. J. Biol.Rhythms 2:35–56.
Reebs, S.G. and N. Mrosovsky (1989a) Effects of induced wheel running onthe circadian activity rhythms of Syrian hamsters: Entrainment andphase response curve. J. Biol. Rhythms 4:39–48.
Reebs, S.G. and N. Mrosovsky (1989b) Large phase-shifts of circadianrhythms caused by induced running in a re-entrainment paradigm: Therole of pulse duration and light. J. Comp. Physiol. [A] 165:819–825.
Rees, H. and M.H.T. Roberts (1993) The anterior pretectal nucleus: aproposed role in sensory processing. Pain 53:121–135.
Rhoades, R.W. (1981) Organization of somatosensory input to the deepcollicular laminae in hamster. Behav. Brain Res. 3:201–222.
Rhoades, R.W., S.E. Fish, N.L. Chiaia, C. Bennett-Clarke, and R.D. Mooney(1989) Organization of the projections from the trigeminal brainstemcomplex to the superior colliculus in the rat and hamster: Anterogradetracing with Phaseolus vulgaris leucoagglutinin and intra-axonal injec-tion. J. Comp. Neurol. 289:641–656.
Ribak, C.E. and A. Peters (1975) An autoradiographic study of theprojections from the lateral geniculate body of the rat. Brain Res.92:341–368.
Robertson, R.T. (1983) Efferents of the pretectal complex: Separate popula-tions of neurons project to lateral thalamus and to inferior olive. BrainRes. 258:91–95.
Robson, J.A. and W.C. Hall (1976) Projections from the superior colliculusto the dorsal lateral geniculate nucleus of the grey squirrel (Sciuriuscarolinensis). Brain Res. 113:379–385.
Rose, J.E. (1942) The thalamus of the sheep: Cellular and fibrous structureand comparison with pig, rabbit, and cat. J. Comp. Neurol. 77:469–523.
Rusak, B., J.H. Meijer, and M.E. Harrington (1989) Hamster circadianrhythms are phase-shifted by electrical stimulation of the geniculo-hypothalamic tract. Brain Res. 493:283–291.
Rusak, B., H.A. Robertson, W. Wisden, and S.P. Hunt (1990) Light pulsesthat shift rhythms induce gene expression in the suprachiasmaticnucleus. Science 248:1237–1240.
Scalia, F. (1972) The termination of retinal axons in the pretectal region ofmammals. J. Comp. Neurol. 145:223–258.
Scalia, F. and V. Arango (1979) Topographic organization of the projectionsof the retina to the pretectal region in the rat. J. Comp. Neurol.186:271–292.
Schober, W. (1981) Efferente und afferente Verbindungen des Nucleuslateral posterior thalami (‘‘Pulvinar’’) der Albinoratte. Z. Mikosk. Anat.Forsch. 95:827–844.
Sefton, A.J. and B. Dreher (1995) Visual system. In G. Paxinos (ed): The RatNervous System. New York: Academic, pp. 833–898.
Sherman, S.M. and R.W. Guillery (1996) Functional organization of thala-mocortical relays. J. Neurophysiol. 76:1367–1395.
Simpson, J.I., R.A. Giolli, and R.H.I. Blanks (1988) The pretectal nuclearcomplex and the accessory optic system. Rev. Oculomot. Res. 2:335–364.
Sparks, D.L. (1990) Signal transformations required for the generation ofsaccadic eye movements. Ann. Rev. Neurosci. 13:309–336.
Stein, B.E. (1984) Multimodal representation in the superior colliculus andoptic tectum. In H. Vanegas (ed): Comparative Neurology of the OpticTectum. New York: Plenum, pp. 819–841.
Swanson, L.W., W.M. Cowan, and E.G. Jones (1974) An autoradiographicstudy of the efferent projections of the ventral lateral geniculatenucleus in the albino rat and cat. J. Comp. Neurol. 156:143–164.
Takahashi, T. (1985) The organization of the lateral thalamus of the hoodedrat. J. Comp. Neurol. 231:281–309.
Taylor, A.M., G. Jeffery, and A.R. Lieberman (1986) Subcortical afferent andefferent connections of the superior colliculus in the rat and compari-sons between albino and pigmented strains. Exp. Brain Res. 62:131–142.
Terasawa, K., K. Otani, and Y. Fukuda (1979) Descending pathways of thenucleus of the optic tract in the rat. Brain Res. 173:405–417.
Tiao, Y.-C. and C. Blakemore (1976) Functional organization in the visualcortex of the golden hamster. J. Comp. Neurol. 168:459–482.
Trejo, L.J. and C.M. Cicerone (1984) Cells in the pretectal olivary nucleusare in the pathway for the direct light reflex of the pupil in the rat.Brain Res. 300:49–62.
Trojanowski, J.Q. and M.L. Schmidt (1984) Interneuronal transfer ofaxonally transported proteins: Studies with HRP and HRP conjugatesof wheat germ agglutinin, cholera toxin and the B subunit of choleratoxin. Brain Res. 311:366–369.
Turlejski, K., R.L. Djavadian, and B. Dreher (1993) Parabigeminal, pretec-tal and hypothalamic afferents to rat’s dorsal lateral geniculate nucleus.Comparison between albino and pigmented strains. Neurosci. Lett.160:225–231.
Uhlrich, D.J. and K.A. Manning (1995) Projection of individual axons fromthe pretectum to the dorsal lateral geniculate complex in the cat. J.Comp. Neurol. 363:147–159.
Vrang, N. and J.D. Mikkelsen (1996)Afferent connections of the intergenicu-late leaflet of the golden hamster: A combined retrograde and antero-grade tracing study. Soc. Neurosci. 26:232.
Warton, S.S., S.E. Dyson, and A.R. Harvey (1988) Visual thalamocorticalprojections in normal and enucleated rats: HRP and fluorescent dyestudies. Exp. Neurol. 100:23–39.
Weber, J.T. and J.K. Harting (1980) The efferent projections of the pretectalcomplex: An autoradiographic and horseradish peroxidase analysis.Brain Res. 194:1–28.
Weber, J.T. and B. Hutchins (1982) The demonstration of a retinalprojection to the medial pretectal nucleus in the domestic cat and thesquirrel monkey: An autoradiographic analysis. Brain Res. 232:181–186.
Wickland, C. and F.W. Turek (1994) Lesions of the thalamic intergeniculateleaflet block activity-induced phase shifts in the circadian activityrhythm of the golden hamster. Brain Res. 660:293–300.
Yezierski, R.P. (1988) Spinomesencephalic tract: projection from the lumbo-sacral spinal cord of the rat, cat and monkey. J. Comp. Neurol.267:131–146.
Young, M.J. and R.D. Lund (1997) The anatomical substrates subservingthe pupillary light reflex in rats: origin of the consensual pupillaryresponse. Neuroscience 62:481–486.
SUBCORTICAL VISUAL CONNECTIONS 309