interaction of glucocorticoids with glucagon and ... · in the control of gluconeogenesis and...
TRANSCRIPT

THE JOURNAL OF BICJLOG~CAL CHEMISTRY Vol. 247, No. 11, Issue of June 10, pp. 3579-3588, 1972
Printed in U.S.A.
Interaction of Glucocorticoids with Glucagon and Epinephrine in the Control of Gluconeogenesis and Glycogenolysis in Liver and of Lipolysis in Adipose Tissue*
(Received for publication, February 7, 1972)
JOHN H. EXTON,$ NAOMI FRIEDMANN,~ ELLEN HEE-AIK WONG,~ J.PARKE BRINEAUX,JACKIE D. CORBIN,
ANDCHARLES R. PARK
From the Department of Physiology, Vanderbilt University, Nashville, Tennessee 372232
SUMMARY
The effects of glucagon and epinephrine on gluconeogenesis and glycogenolysis in liver and of catecholamines and adreno- corticotrophic hormone on lipolysis in adipose tissue were examined in normal and adrenalectomized rats.
Glucagon or epinephrine stimulation of glucose synthesis from lactate was markedly reduced in isolated perfused livers from fasted, adrenal-deficient rats. Administration in uiuo of the glucocorticoid dexamethasone 30 min prior to perfusion restored the gluconeogenic response to glucagon. Cortisol and dexamethasone were also effective when added to the perfusion medium, but had no effect in the absence of glu- cagon.
Cyclic AMP (adenosine 3’ ,5’-monophosphate) accumu.lation was normal or increased in livers from adrenalectomized rats perfused without hormones or with glucagon or epinephrine. Gluconeogenesis in livers from fasted, adrenal-deficient rats was relatively unresponsive to cyclic AMP or dibutyryl cyclic AMP added to the perfusion medium. Hypophysectomized rats also showed an impairment of cyclic AMP activation of gluconeogenesis which was corrected by treatment with cortisol but not growth hormone.
High concentrations of epinephrine, glucagon, and cyclic AMP stimulated glucose production and lactate gluconeo- genesis in livers from fed, adrenalectomized rats. However, more nearly physiological concentrations of the hormones did not activate glucose release and phosphorylase normally in such livers. Livers from fed, adrenalectomized rats also showed diminished activation of gluconeogenesis and phos- phorylase by exogenous cyclic AMP.
Omission of Ca+f from the perfusion medium did not alter control or cyclic AMP-stimulated glycogenolysis or gluco- neogenesis in normal rats. Addition of ethyleneglycol-bis
* This investigation was supported by program project Grant AM-07462 and Training Grant GM-00423 from the National In- stitutes of Health, United States Public Health Service. Portions of this work are taken from the Ph.D. thesis submitted by J. D. C. to Vanderbilt University, August, 1968. Preliminary reports of some of this work have appeared (l-4).
$ Investigator of the Howard Hughes Medical Institute. § Present address, Department of Biochemistry, University of
Pennsylvania,Philadelphia, Pa. f Present address, Department of Biochemistry, University of
Singapore, Singapore.
(P-aminoethyl ether)-N, N’-tetraacetic acid to Ca+f-free medium reduced the control rates of glycogenolysis and gluconeogenesis but did not abolish the stimulation by cyclic AMP. Lowering the Ca++ level did not accentuate the impairment of cyclic AMP-stimulated glycogenolysis in fed, adrenalectomized rats nor did increasing the Ca++ level relieve the impairment of cyclic AMP-stimulated gluco- neogenesis. It is therefore concluded that variations in plasma Ca+f are not important in the regulation of hepatic glycogenolysis or gluconeogenesis.
Epinephrine stimulation of free fatty acid and glycerol release is impaired in epididymal fat pads or isolated fat cells from adrenalectomized rats. Cyclic AMP accumulation in response to epinephrine is, however, increased in fat pads from such animals. Activation of lipolysis by norepinephrine, adrenocorticotrophic hormone, or cyclic AMP is also reduced in fat cells from adrenal-deficient rats.
These studies indicate that adrenal glucocorticoids are re- quired for normal activation of gluconeogenesis, glycogen- olysis, and lipolysis by epinephrine and other hormones. Part of the function of the steroids appears to be to maintain normal sensitivity of these processes to cyclic AMP. The possibility is discussed that the corticoids exert their permis- sive effects by preserving a normal intracellular ionic en- vironment.
The classical observations of Long, Katzin, and Fry (5) es- tablished that adrenal secretions were required for the main- tenance of blood glucose levels in fasting mice. Adrenal cortical hormones appear to act, in part, by maintaining the supply of amino acids from peripheral tissues to the liver where they are partly utilized for glucose synthesis. In addition, adrenal steroids exert effects on the gluconeogenic process in the liver. This is revealed by the observation that the gluconeogenic re- sponse to fasting, diabetes, or glucagon is reduced or abolished in livers from adrenalectomized rats (1, 2, 4).
Adrenal steroids also influence lipolysis in adipose tissue (6). Acting with growth hormone they increase the release of fatty acid and glycerol from isolated fat cells by a mechanism appar- ently involving protein synthesis (7). Adrenalectomy has also
3579
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from

3580 Interaction of Glucocorticoids with Glucagon Vol. 247, No. 11
been shown to reduce the lipolytic response of adipose tissue to epinephrine or ACTHi (3, 8-12).
The purpose of the present study has been to elucidate the role of glucocorticoids in the control of glucose production in the liver and of lipolysis in fat tissue. Preliminary observations of the interaction of glucagon and adrenal cortical hormones in the regulation of gluconeogenesis (1) have been extended. It has also been found that the glycogenolytic response to glucagon or epinephrine is impaired in livers from adrenal-deficient rats con- firming observations in tivo (13) and resembling the situation recently described in hearts from adrenalectomized rats (14).
The results indicate that gluconeogenesis, glycogenolysis, and lipolysis become less sensitive to activation by cyclic AMP in the absence of glucocorticoids. A general mechanism for the “permissive effect” of adrenal steroids is proposed.
MATERIALS AND METHODS
Male Sprague-Dawley rats fed ad Zibitum with Purina labora- tory chow or fasted 18 to 22 hours were used. Their body weights were 100 to 150 g in the liver studies and 150 to 200 g in the fat tissue studies. Bilateral adrenalectomy was performed 5 to 7 days prior to study and adrenalectomized animals were given 0.5% NaCl solution to drink. Hypophysectomized rats were obtained from Hormone Assay Laboratories, Inc., Chicago. The technique of liver perfusion has been described in detail previously (15). The perfusion medium consisted of Krebs- Henseleit bicarbonate buffer containing 3% bovine serum albu- min (Cohn Fraction V, Nutritional Biochemicals) and 15 to 20% bovine erythrocytes (16).
Liver glycogen and perfusate glucose were measured and the production of [“Clglucose and [14C]glycogen from [14C]lactate were determined as described previously. Some liver samples frozen in an aluminum clamp (17) at the end of perfusion were pulverized (18) and assayed for phosphorylase in the presence of 0.2 mM AMP (19). Levels of cyclic AMP were kindly deter- mined in the frozen liver powder by Doctors R. W. Butcher and G. A. Robison using the liver phosphorylase activation method (20).
In the studies with intact adipose tissue, epididymal fat pads were excised from rats under nembutal anesthesia, divided into distal, medial, and proximal portions, and distributed at random into 25-ml flasks so that each received two portions. The flasks contained 5 ml of Krebs-Henseleit bicarbonate buffer gassed with 02-COZ (95:5) and made up with half the prescribed calcium and, in some experiments, 5 y. bovine serum albumin (Nutritional Biochemicals). Fat pads were incubated at 37“ in a Dubnoff metabolic shaker for 15 min and then transferred to fresh buffer and incubated under test conditions for 30 or 60 min. Experi- ments were terminated by removing the pieces of fat and drop- ping them into an operating Waring Blendor containing 17 ml of 0.05 M HCl and 0.1 ml of purified tritiated cyclic AMP (about 10,000 dpm). The homogenates were heated in a boiling water bath for 15 min, then fractionated and assayed for cyclic AMP according to the procedure of Butcher et al. (20).
Two milliliters of the incubation medium were heated in a boiling bath and centrifuged. Glycerol was determined in the supernatant fluid by the method of Wieland (21). Some samples of medium were assayed for free fatty acids according to the
1 The abbreviations used are: ACTH, adrenocorticotrophic hormone; cyclic AMP, adenosine 3’,5’-monophosphate; EGTA, ethyleneglycol-his@-aminoethyl ether)-N, N’-tetraacetic acid.
method of Dole (22). DNA was determined by the method of Ceriotti (23) in two fat pads taken at random from the normal and adrenalectomized groups and extracted with chloroform- methanol (2: 1).
Isolated fat cells were prepared by the method of Rodbell (24). Aliquots (0.5 to 2.0 ml) of the cells suspended in Krebs-Henseleit bicarbonate buffer containing 4% albumin were distributed into 25-ml polyethylene-counting vials containing sufficient albumin- bicarbonate medium with or without additions to give a final volume of 2.5 ml. After regassing, the vials were incubated for 30 min at 37” with shaking. The experiments were terminated by addition of HC104 to a final concentration of 3%. Glycerol was determined (21) in the supernatant fluid after centrifugation.
Dry weights were determined by filtering aliquots of cell sus- pensions through tared 0.45 p Millipore filters and reweighing the filters after drying in vucuo. DNA was also measured in cell suspensions by the method of Burton (25).
n-[U-14C]Lactate and nn-[2-14C]lactate were obtained from Amersham-Searle. Sodium n-lactate solution (1 M) was pre-
pared from n-lactic acid from Mann. n-Epinephrine hydro- chloride and ACTH were from Parke-Davis Co., Detroit, and glucagon was a kind gift from Eli Lilly Co., Indianapolis. L-
Norepinephrine bitartrate was from Winthrop Laboratories, New York, N. Y. Solutions of the catecholamines and glucagon were prepared in NaCl-albumin solution as described previously (26). Cyclic AMP was from Schwarz and N6,0P’-dibutyryl cyclic AMP was from Boehringer. Cortisol was from Mann, cortisol sodium succinate (Solu-Cortef) from the Upjohn Co., Kalamazoo, Mich., and dexamethasone sodium phosphate (deca- dron phosphate) from Merck, Sharp and Dohme, Rahway, N. J. Growth hormone (bovine) was a kind gift from the Endocrinology Study Section of the National Institutes of Health. Collagenase used in the preparation of fat cells was from Worthington.
RESULTS
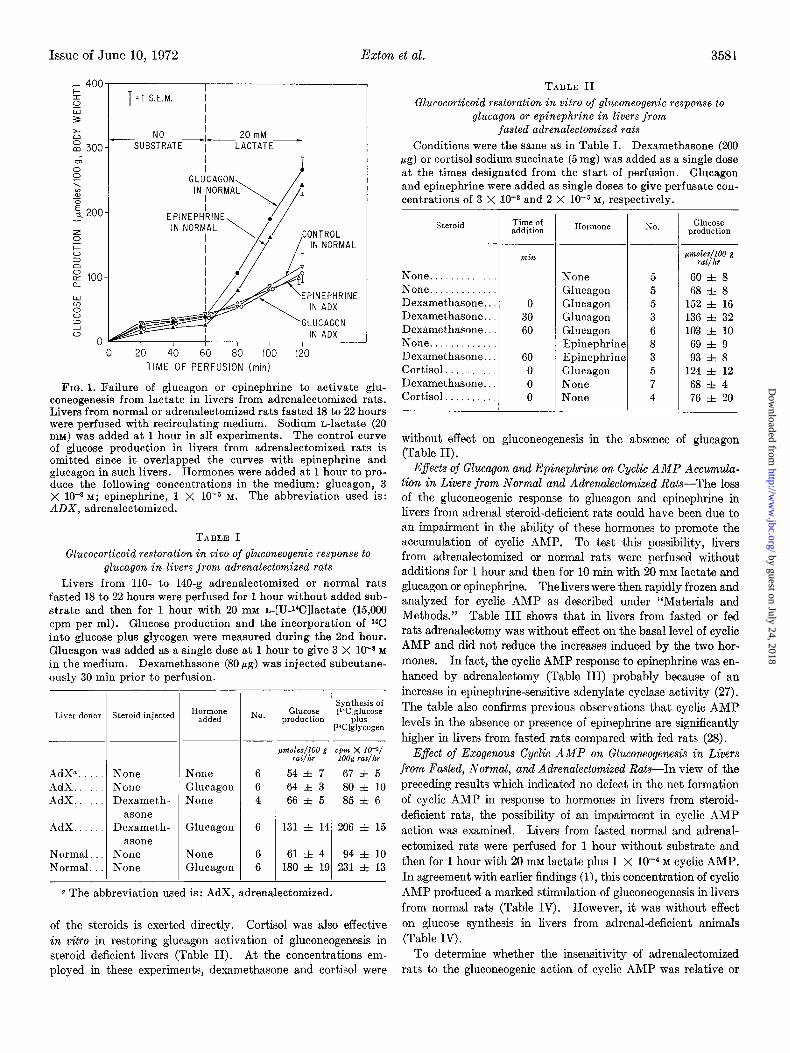
Impaired Activation of Gluconeogenesis by Glucagon and Epineph- rine in Livers from Fasted Adrenalectomized Rats--In order to test the dependence of the gluconeogenic responses to glucagon and epinephrine on adrenal secretions, livers from fasted normal or adrenalectomized rats were initially perfused for 1 hour without substrate and then for a further 1 hour with 20 mM L-lactate in the absence or presence of glucagon or epinephrine. As shown in earlier studies (26), hormonal activation of gluconeogenesis from lactate was very marked in livers from normal rats (Fig. 1). In livers from adrenal-deficient rats, however, neither glucagon nor epinephrine activated the synthesis of glucose from lactate under these conditions (Fig. 1).
Subcutaneous injection of the glucocorticoid dexamethasone 30 min prior to perfusion restored glucagon activation of gluco- neogenesis as revealed by increased glucose production and syn- thesis of [r4C]glucose plus [r4C]glycogen from [14C]lactate (Table I). Dexamethasone treatment per se produced a small increase in [14C]hexose synthesis (Table I).
To determine whether glucocorticoids acted directly on the liver to restore the glucagon response, dexamethasone or cortisol was added to the medium perfusing livers from adrenalectomized rats at various times. When present from the start of perfusion, dexamethasone increased the gluconeogenic response to glucagon about 2-fold (Table II). Added at 30 or 60 min it was also ef- fective in restoring the response to glucagon or epinephrine (Table II). These data indicate that at least part of the action
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
Issue of June 10, 1972 Exton et al. 358 1
T =I S.E.M. /
__ NO -/-- 20 mM SUBSTRATE / LACTATE-
0 20 40 60 80 too 120 TIME OF PERFUSION (min)
FIG. 1. Failure of glucagon or epinephrine to activate glu- coneogenesis from lactate in livers from adrenalectomized rats. Livers from normal or adrenalectomized rats fasted 18 to 22 hours were perfused with recirculating medium. Sodium n-lactate (20 mM) was added at 1 hour in all experiments. The control curve of glucose production in livers from adrenalectomized rats is omitted since it overlapped the curves with epinephrine and glucagon in such livers. Hormones were added at 1 hour to pro- duce the following concentrations in the medium: glucagon, 3 X 10-s M; epinephrine, 1 X 10e5 M. The abbreviation used is: ADX, adrenalectomized.
TABLE I Glucocorticoid restoration in vivo of gluconeogenic response to
glucagon in livers- from adrenalectomized rats
Livers from llO- to 140-g adrenalectomized or normal rats fasted 13 to 22 hours were perfused for 1 hour without added sub- strate and then for 1 hour with 20 mM L-[U-Wllactate (15,000 cpm per ml). Glucose production and the incorporation of 1% into glucose plus glycogen were measured during the 2nd hour. Glucagon was added as a single dose at 1 hour to give 3 X 10V8 M in the medium. Dexamethasone (80 rg) was injected subcutane- ously 30 min prior to perfusion.
Liver donor
AdXa.. AdX.. AdX..
AdX..
Normal. Normal.
Steroid injected
None None Dexameth-
asone Dexameth-
asone None None
HOlXllOll~ added NO.
None 6 Glucagon 6 None 4
Glucagon 6
None 6 Glucagon 6
Synthesis of GlllUXe
production [~~C]glucose
plus [‘qglycogen
54 f 7 67 f 5 64 f 3 80 rk 10 66 f 5 85 f 6
131 f 14 206 f 15
61 f 4 94 f 10 180 f 19 231 f 13
a The abbreviation used is: AdX, adrenalectomized.
of the steroids is exerted directly. Cortisol was also effective in vitro in restoring glucagon activation of gluconeogenesis in steroid deficient livers (Table II). At the concentrations em- ployed in these experiments, dexamethasone and cortisol were
TABLE II Glucocorticoid restoration in vitro of gluconeogenic response to
glucagon or epinephrine in livers from fasted adrenalectomized rats
Conditions were the same as in Table I. Dexamethasone (200 pg) or cortisol sodium succinate (5 mg) was added as a single dose at the times designated from the start of perfusion. Glucagon and epinephrine were added as single doses to give perfusate con- centrations of 3 X 10-s and 2 X 10-j M, res
Steroid Time of addition
None. None. Dexamethasone. Dexamethasone. Dexamethasone. None. . Dexamethasone. Cortisol . Dexamethasone. Cortisol
min
1 0 30 60
60 0
0 0
None Glucagon Glucagon Glucagon Glucagon Epinephrine Epinephrinc Glucagon None None
ectively.
Glucose production
5 5 5 3 6 8 3 5 7 4
my;;0 s
60 f 8 68 f 8
152 f 16 136 f: 32 103 f 10 69 f 9 93 f 8
124 f 12 68 f 4 76 f 20
without effect on gluconeogenesis in the absence of glucagon (Table II).
E$ects of Glucagon and Epinephrine on Cyclic AMP Accumula- tion in Livers from Normal and Adrenalectomized Rats-The loss of the gluconeogenic response to glucagon and epinephrine in livers from adrenal steroid-deficient rats could have been due to an impairment in the ability of these hormones to promote the accumulation of cyclic AMP. To test this possibility, livers from adrenalectomized or normal rats were perfused without additions for 1 hour and then for 10 min with 20 mM lactate and glucagon or epinephrine. The livers were then rapidly frozen and analyzed for cyclic AMP as described under “Materials and Methods.” Table III shows that in livers from fasted or fed rats adrenalectomy was without effect on the basal level of cyclic AMP and did not reduce the increases induced by the two hor- mones. In fact, the cyclic AMP response to epinephrine was en- hanced by adrenalectomy (Table III) probably because of an increase in epinephrine-sensitive adenylate cyclase activity (27). The table also confirms previous observations that cyclic AMP levels in the absence or presence of epinephrine are significantly higher in livers from fasted rats compared with fed rats (28).
Effect of Exogenous Cyclic AMP on Gluconeogenesis in Livers from Fasted, Normal, and Adrenalectomized Rats-In view of the preceding results which indicated no defect in the net formation of cyclic AMP in response to hormones in livers from steroid- deficient rats, the possibility of an impairment in cyclic AMP action was examined. Livers from fasted normal and adrenal- ectomized rats were perfused for 1 hour without substrate and then for 1 hour with 20 mM lactate plus 1 X 10m4 M cyclic AMP. In agreement with earlier findings (1)) this concentration of cyclic AMP produced a marked stimulation of gluconeogenesis in livers from normal rats (Table IV). However, it was without effect on glucose synthesis in livers from adrenal-deficient animals (Table IV).
To determine whether the insensitivity of adrenalectomized rats to the gluconeogenic action of cyclic AMP was relative or
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
3582 Interaction of Glucocorticoids with Glucagon Vol. 247, No. 11
absolute, the effects of higher concentrations of exogenous nu- response in the steroid-deficient livers whereas injection of the cleotide were tested. Table IV shows that 2 x 10m4 and 1 X hormone into the medium in the oxygenation chamber as a single lop3 M nucleotide produced significant but submaximal stimula- dose (as in the experiments of Fig. 1) was ineffective. tion of glucose synthesis in the steroid-deficient livers. Higher Impaired Activation of Glucose Production and Gluconeogenesis levels were not tested since these frequently inhibited gluconeo- Hormones and Cyclic AMP in Livers from Fed Adrenalectomized genesis in normal livers. The more potent derivative of cyclic Rats-Activation of gluconeogenesis by glucagon and epineph- AMP, N6,02’-dibutyryl cyclic AMP also stimulated gluconeo- rine can be shown in livers from fed rats by measuring the con- genesis in livers from adrenalectomized rats at 2 X 10m4 M con- version of [14C]lactate to [14C]glucose plus [14C]glycogen (26). centration (Table IV). Initial experiments with livers from fed adrenalectomized rats
Since high concentrations of cyclic AMP induced a significant failed to show any defect in the activation of hexose synthesis by activation of gluconeogenesis in the adrenalectomized rats, the glucagon, epinephrine, or cyclic AMP (Table V, top four lines). effects of high levels of glucagon were examined. Table IV However, when the concentration of cyclic AMP was reduced, shows that glucagon infused at a rate of 0.4 nmole per min into an impairment in the sensitivity of gluconeogenesis to activation the influent medium did produce a significant but submaximal became apparent in the steroid-deficient livers (Table V, bottom
four lines). TABLE III
Cyclic AMP levels in livers from adrenalectomized or normal rats TABLE IV
perfused with glucagon or epinephrine Effect of adrenalectomy on gluconeogenic response to cyclic AMP Livers from fed or fasted, normal or adrenalectomized rats and high concentrations of glucagon in livers from fasted rats
were perfused for 1 hour without hormones and then for 10 min with glucagon or epinephrine at the concentrations shown.
Livers from fasted normal or adrenalectomized rats were per- fused for 1 hour without substrate and then for 1 hour with 20 -
-
-
(
-
-
NO. Cyclic AMP mM L-lactate and the designated concentrations of glucagon or cyclic AMP or its NC, W-dibutyryl derivative (DB cyclic AMP).
Treatment of rat
Normal, fasted.. AdX,O fasted. . Normal, fasted. AdX, fasted. Normal, fasted. AdX, fasted..
Normal, fed. AdX, fed. Normal, fed. . AdX, fed. . . Normal, fed. AdX, fed. Normal, fed. AdX, fed.
ZoncentratioI
M
1 x 10-s 1 x 10-s 1 x 10-e 1 x 10-6
3 x 10-s 3 X 10-S 2 x 10-r 2 x 10-T 1 x 10-e 1 x 10-e
HOIItlOlVZ
None None
Glucagon Glucagon Epinephrine Epinephrine
None None
Glucagon Glucagon Epinephrine Epinephrine Epinephrine Epinephrine
nmoleslg liner
6 0.87 f 0.03 7 0.92 f 0.05 3 21.2 f 0.4 3 25.7 f 1.9 6 3.4 f 0.6 7 6.3 f 1.6
-
( 3uconeogenesis from lactate
Addition Jo. of pair! loncentratiol
1 x 10-d 2 x 10-4 1 x 10-Z 2 x 10-d 5 x lo-'" 3 X lo-Sb
Kormal rats Adrenalecto- mized rats
/&n&s g1uc0st?/100 g rat/h
None. . . . . . Cyclic AMP. Cyclic AMP. . Cyclic AMP. DB Cyclic AMP.. Glucagon. . Glucagon..........
67 f 7 209 f 18 180 f 19 206 f 13 188 f 20 233 f 40 217 f 18
62 f 4 62 f 6 91 f 9
129 f 8 117 f 6 103 f 8
74 f 10
7 0.59 f 0.02 7 0.61 f 0.03 4 28.7 f 1.4 4 28.7 f 3.0 7 0.80 f 0.04 7 0.91 f 0.03 -L I - 3 0.95 f 0.01 o Glucagon was infused at constant rate to produce this con-
4 1.90 f 0.58 centration at 2 hours assuming no degradation. b Glucagon was added as a single dose at 1 hour to produce this
. . a The abbreviation used is: AdX, adrenalectomlzed. concentration.
TABLE V
E$ects of glucagon, epinephrine, and cyclic AMP on glucose production and gluconeogenesis in lactate-perfused livers from fed normal and adrenalectomized rats
Livers from normal or adrenalectomized rats fed ad libitum were perfused without substrate for 1 hour and then for a further hour with 20 mM L-lactate, D,L-[2-‘%]lactate (15,000 cpm/ml), plus the agents noted. Glucose and [Wlglucose plus [“Clglycogen formation were measured over the 2nd hour.
Glucose production
NOlTl~l I AdX”
Synthesis of [Wlglucose plus [Wlglycogen
Normal I
AdX
@m X lo-‘/lo0 g rot/hr
65 f 4 55 f 3 130 f 10 90 f 8 105 f 9 102 f 9 108 f 15 105 f 7 120 f 17 57 f 10
90 f 6 58 f 2 90 f 13 57 f 11 82 f 7 51 f 4
Concentration Agent No. of pairs
M ,moles/100 g rat/hr
None. Glucagon. Epinephrine. Cyclic AMP. Cyclic AMP. Cyclic AMP. Cyclic AMP. Cyclic AMP.
132 f 8 495 f 58 421 f 29 410 f 8 466 f 61 444 f 31 307 f 35 273 f 26
73 f 4 246 f 52 156 f 36 222 f 51 130 f 37 103 f 11
74 f 8 82 f 5
4 X 10-S 1 x 10-b 1 x 10-s 2 x 10-4 1 x 10-4
4 x 10-s 2 x 10-b
a The abbreviation used is: AdX, adrenalectomized.
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
Issue of June 10, 1972 Exton et al. 3583
Glucose production in the livers from the fed adrenalectomized rats was stimulated by high concentrations of hormones and cyclic AMP (Table V, top five lines), but low levels of the nucleo- tide elicited impaired responses (Table V, bottom three lines). This suggested that an impairment of the activation of glyco- genolysis by cyclic AMP also existed in the steroid-deficient livers. To test this possibility more directly, livers from fed normal or adrenalectomized rats were perfused without added substrate and the absence or presence of different concentrations of glucagon, epinephrine, or cyclic AMP. Table VI shows that glucose mobilization in livers from adrenalectomized rats was subnormal or absent in response to low concentrations of these agents. Although glycogen levels were lower than normal in these livers (Table VII), the defect lay in the failure of phos- phorylase to be activated normally by low levels of all three agents (Table VII). As expected from the results of Table VI the steroid-deficient livers showed full activation of the enzyme in the presence of high levels of glucagon or epinephrine (data not shown). These results indicate that the phosphorylase ac- tivation system is relatively insensitive to cyclic AMP in livers from adrenalectomized rats.
TABLE VI
Glycogenolysis in livers from fed normal and adrenalectomized rats perfused with hormones or cyclic AMP
Livers from normal or adrenalectomized rats fed ad libitum were perfused for 1 hour without hormones and then for 1 hour with the agents noted. Glucose production was measured over the 2nd hour. Hormones were infused at constant rate during the 2nd hour to produce the final concentrations noted (assuming no degradation). No substrate was added.
Agent Glucose production
Concentration No. of pairs Normal AdX“
24 ,molcs/100 g rat/hr
None. . 11 Epinephrine. 1 x 10-e 8 Epinephrine. . 2 x 10-S 8 Glucagon.. . . . . 1 X lo-l0 4
Glucagon.. . . . 4 X lo-10 4 Cyclic AMP. . 1 X 10e4 15
0 The abbreviation used is: AdX, adrenalectomized.
41 f 5 283 f 31 350 f 22 237 f 27 408 f 13 364 f 22
13 f 2 65 f 16
270 f 26 79 f 22
319 f 38 57 f 10
Effects of Varying Medium Ca++ on Glycogenolysis and Gluco-
neogenesis in Livers from Normal and Adrenulectomized Rats- Miller et al. (14) have shown a defect in epinephrine activation of phosphorylase in perfused hearts from adrenalectomized rats and have localized the impairment to the phosphorylase b kinase reaction. This reaction requires Ca++ ions (29, 30) and Miller et al. (14) have found that the phosphorylase defect can be re- paired by increasing the Ca++ concentration of the perfusion medium. It was therefore of interest to test whether changes in medium Ca++ could alter glycogenolysis or gluconeogenesis in the perfused liver.
Removal of Ca++ from the perfusion medium did not reduce basal or cyclic AMP-stimulated glucose production in lactate- perfused livers from normal fasted or fed rats (Table VIII).
TABLE VIII
E$ects of EGTA and low Cat+ on glycogenolysis and gluconeogenesis in livers from fed or fasted normal rats
Livers from normal rats fed ad libitum or fasted 18 to 22 hours were perfused for 10 min without recirculation, then for 50 min with recirculation. At 60 min, L-[UJ%]lactate was added to give 20 mM and 15,000 cpm per ml, and glucose and [l%]glucose formation were measured over the next 30 min. Cyclic AMP when present was added at 60 min to give 1 X lo+ M. Ca++-free medium was used in all experiments. It was prepared by wash- ing bovine erythrocytes twice in Ca++-free Krebs-Henseleit bicarbonate buffer containing 3% bovine serum albumin (Frac- tion V, Pentex) and resuspending them in this buffer. EGTA (ethyleneglycol-bis(&aminoethyl ether)-N,N’-tetraacetic acid) was 1 mM and Ca+ was 2.5 mM.
Additions Glucose production
Nutritional No. of state
$ g pairs
s w I I -cAMPa +cAMP
-I ’ 1 pm&s/100 g rat/30 min
Fed...... + - Fed...... - - Fed...... - + Fasted.. + - Fasted.. -/- Fasted..
[W]Glucose synthesis
-CAMP +cAMP
cpm x lo-~/loo g rat/30 lnin
- +
3 67 f 10 274 f 2( 3 63 i 3 238 f 1: 3 47 f 4 221 f 24 5 39 f 2 58 f 1 5 46 f 4 58 f 4 5 17 f 3 30 f 3
45 f 4 63 f 1 46 f 4 58 f 4 23 f 2 33 f 4
a The abbreviation used is: CAMP, cyclic AMP.
TABLE VII
Glycogen levels and phosphorylase activzty in livers from fed normal and adrenalectomized rats perfused with hormones or cyclic AMP
Final glycogen levels and phosphorylase activities (measured in the presence of 0.2 mM AMP) were determined in the experi- ments of Table VI.
Agent I Concentration
None. ................. Epinephrine ............ Epinephrine. ........... Glucagon .............. Cyclic AMP. ..........
‘w
1 x 10-6 2 x 10-6 1 x lo-‘” 1 x lo-’
Final liver glycog& Phosphorylase activity
NOIllld I
AdXb Normal I
AdX
pm&s/100 g rat nzg Pi released/g liuer/30 min
648 f 124 204 f 44 1.35 f 0.08 1.07 f 0.06 512 f 183 119 f 50 3.46 f 0.27 2.13 f 0.10 776 f 144 204 f 80 4.86 f 0.22 3.78 f 0.25 316 f 88 232 f 44 4.90 f 0.51 2.16 f 0.09 586 f 126 78 f 26 2.46 f 0.17 1.03 f 0.15
a Hexose equivalents. b The abbreviation used is: AdX, adrenalectomized.
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
3584 Interaction of Glucocorticoids with Glucagon Vol. 247, No. 11
TABLE IX E$ects of raising and lowering medivm Ca++ on glztconeogenesis and
glycogenolysis in livers from adrenalectomized rats perfused with cyclic AMP
Livers from fasted adrenalectomized rats were perfused for 1 hour without substrate and then for 1 hour with 20 mM lactate plus 0.1 mM cyclic AMP. Livers from fed adrenalectomized rats were perfused for 1 hour without additions and then for 1 hour with 0.1 mM cyclic AMP or 2 X 10-e M epinephrine. Glucose production refers to the 2nd hour.
Nutritional state ICalcium knell Addition No.
Fasted. . 2.5 Fasted. . 5.0 Fed. . . . 2.5 Fed..... . 1.25 Fed. . . . . . . 2.5 Fed.. . . . . 1.25
7n.M
Cyclic AMP 6 Cyclic AMP 12 Cyclic AMP 8 Cyclic AMP 8 Epinephrine 3 Epinephrine 3
Glucose production
pnoles/100 g ratlhr
61 f 7 65 f 5 46 f 9 62 i 17
250 f 24 291 f 48
TABLE X
Effects of cortisol and growth hormone treatment on gluconeogenesis in livers from hypophysectomized rats
Livers from fasted hypophysectomized rats were perfused for 1 hour without substrate and then for 1 hour with 20 mM L-[U-W]- lactate without or with 0.1 mM cyclic AMP. Glucose and [r4C]- glucose formation refer to the 2nd hour. Cortisol (2.5 mg) or growth hormone (100rg) was injected subcutaneously twice daily for 2 days prior to perfusion.
Treatment
None. . . Cortisol.. . Growth hor-
mone.....
-
-
-
Glucose production [WlGIucose synthesis No. of
pairs -cAMPa +cAMP -CAMP +cAMP
,moles/lOO g ratlhr cpm X 10-a/100 g rat/ hr
4 52 f 6 46 f 4 73 f 10 63 f 7 6 189 f 17 401 i 33 151 i 19 203 f 19
4 55 f 2 57 i 6 73 f 1 72 f 7
TABLE XI Effects of steroid treatment on glucagon activation of gluconeogeqesis
in livers from normal rats
Livers from fed or fasted normal rats were perfused for 1 hour with 20 mM L-[U-Wllactate (15,000 cpm per ml). Glucagon was infused the rate of 14 pmoles per min which would have produced a final concentration of 2 X lo-* M assuming no degradation. Cortisol treatment was 2.5 mg injected subcutaneously twice daily for 2 days prior to perfusion.
Nutritional state
Fed. Fed.. Fasted. Fasted.
Treatment
None Cortisol None Cortisol
\To. of Glucose production [WIGlucose synthesis
pairs -Glucagon +Glucagon -Glucagon +Glucagon
jmoles/100 g rai/hr cih x lo-~/loo g rat/hr
6 211 f 11552 f 16 73 f 3 145 % 8 6 324f12666f22 122i5187fll 6 181 f 12 242 f 10 122 f 4 199 i 5 2 248, 262 718, 751 106, 139 157, 181
TABLE XII Effect of epinephrine on free fatty acid release in epididymal fat
pads from normal and adrenalectomized rats
Epididymal fat pads from fed or fasted normal or adrenalec- tomized rats were incubated for 10 min in albumin-buffer as de- scribed under “Materials and Methods” and then transferred to fresh albumin-buffer without or with 11 pM epinephrine. Free fatty acid release was,measured over the next hour.
Additions
Fed.. None 4 Fed. . Epinephrine 4
Fasted. . None 4 Fasted. . . Epinephrine 4
No. of pairs
Free fatty acid release
Normal rats AdX rat?
aeq/g wet tissue/hr
0.15 i 0.02 -0.12 i 0.09 1.15 f 0.23 0.75 f 0.12
0.30 f 0.05 0.17 f 0.03 1.32 f 0.22 0.56 f 0.18
a The abbreviation used is: AdX, adrenalectomized.
a The abbreviation used is: CAMP, cyclic AMP.
However, addition of the Ca +-chelating agent ethyleneglycol- bis(Saminoethy1 ether)-A7 ,N’-tetraacetic acid to Ca++-free me- dium did significantly reduce the control rates of gluconeogenesis and glycogenolysis (Table VIII). Despite these changes, ac- tivation of glycogenolysis by cyclic AMP was not impaired and stimulation of gluconeogenesis from lactate was preserved (Table VIII). These data render it very unlikely that variations in blood Ca* are important in the basal or hormonal control of hepatic glucose production.
Further experiments were carried out to determine whether changes in the Ca++ concentration of the medium altered the sensitivity of livers from adrenalectomized rats to cyclic AMP or hormones. Table IX shows that glucose production in livers from fasted adrenal-deficient rats was not increased in the pres- ence of 1 x 10m4 M cyclic AMP when the Ca++ concentration of the medium was increased from 2.5 to 5 mM. Furthermore, glucose output of livers from fed or fasted adrenalectomized rats was not further reduced below normal in the presence of 1 x lo-’ M cyclic AMP or 2 X 10-G M epinephrine when the Ca++ level was reduced to 1.25 mM (Table IX).
Examination for Permissive Role of Growth Hormone in Cyclic AMP Action in Hypophysectomized Rats-Growth hormone and cortisol display synergism in their actions on lipolysis in adipose tissue (7) and in the induction of insulin insensitivity in hearts from hypophysectomized rats (31). It was therefore of interest to see whether growth hormone or glucocorticoid alone would be capable of restoring normal responses to cyclic AMP in hy- pophysectomized rats.
Table X shows that livers from fed or fasted hypophysecto- mized rats had a similar impairment of activation of gluconeo- genesis by 1 x 10m4 M cyclic AMP as livers from adrenalecto- mized rats. Treatment of fasted hypophysectomized rats with cortisol in vivo 2 days prior to perfusion greatly increased the basal rate of gluconeogenesis and restored to normal the activa- tion by 1 x lop4 M cyclic AMP,2 whereas growth hormone ad- ministration by itself was without effect (Table X). These re- sults indicate that growth hormone is not required for the action
2 The steroid replacement in these experiments was of longer duration than in the experiments with adrenalectomized rats in Table I and caused glycogen accumulation. Glycogenolysis was probably largely responsible for the high rates of glucose produc- tion observed in the absence and presence of cyclic AMP.
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
Issue of June 10, 1972 Exton et al. 3585
TABLE XIII Effect of epinephrine on glycerol release and cyclic AMP levels in
epididymal fat pads from normal and adrenalectomized rats Epididymal fat pads were incubated for 15 min in buffer as
described under “Materials and Methods” and then transferred to fresh buffer and incubated for 30 min without or with epineph- rine (11 PM) plus caffeine (1 mM). At the end of incubation, pads were rapidly fixed for cyclic AMP assay and media were assayed for Plvcerol.
TABLE XIV
Efects of epinephrine, norepinephrine, and ACTH on glycerol release from isolated fat cells from normal
and adrenalectomized rats Fat cells from fed normal and adrenalectomized rats were incu-
bated in albumin buffer for 30 min at 37”. The reaction was stopped with HClOa and glycerol was measured in the protein- free supernatant.
I I I
Additions
None. Epinephrine
+ caffeine.
2. Glycerol output Tissue cyclic AMP
pa’rs Normal rats AdX’” rats Normal rats AdX rats
pmoles/mg DNA/30 min nmoles/mg DNA
4 0.79 f 0.400.62 f 0.180.3 f 0.10.4 f 0.1
6 3.46 f 0.302.25 f 0.375.7 f 1.69.7 f 2.0
a The abbreviation used is: AdX, adrenalectomized.
of cortisol. Whether or not administration of growth hormone would have reduced the dose of steroid required for restoration was not studied.
Effects of Cortisol Treatment on Cyclic AMP Responses in Nor- mal Ruts-The preceding results suggested that administration of glucocorticoids to normal rats might enhance the effects of
Glycerol release Lipolytic agent Concentration No. of
pairs Normal rats AdXa rats
pmles/mg DNA/30 nzin
None. 20 6.2 f 1.2 4.0 f 1.0 Epinephrine. 3 x 10-M 4 20.6 f 6.2 12.8 f 3.4 Epinephrine. 2 x 1o-7 M 17 43.2 f 8.6 23.0 f 4.6 Epinephrine. 5 x lo-’ M 17 70.2 f 9.4 33.2 f 5.0 Epinephrine. . 5 x lo- M 13 60.2 f 10.032.8 f 4.8
Norepinephrine. 1.6 X 10es M 8 32.6 f 2.8 15.8 f 3.0 Norepinephrine. 3 x 1o-6 M 8 59.4 f 7.6 40.0 f 4.6
ACTH. . 1 milliunits/ml 5 45.6 f 11.614.2 f 1.8 ACTH. . . 4 milliunits/ml 5 97.4 f 15.865.4 f 10.4
a The abbreviation used is: AdX. adrenalectomized.
glucagon or cyclic AMP on glycogenolysis or gluconeogenesis. To test this possibility, fed or fasted rats were injected with 2.5 mg of cortisol twice daily for 2 days and their livers were then perfused with 20 mM [14C]lactate in the absence or presence of z 60-
glucagon. Table XI shows that the steroid treatment increased 2
basal glucose production in both the fed and fasted animals. Z
The increase was attributable to higher glycogen levels (data not shown). [K!]Glucose synthesis was also enhanced by cor- NORMAL RATS
tisol in the fed rats but not in the fasted rats. Steroid treatment thus increased glycogen breakdown and gluconeogenesis in fed animals and promoted the breakdown of glycogen deposited in fasted animals. Glucose production in the presence of glucagon was also higher in fed or fasted steroid-treated rats, but [14C]glu- ADRENALECTOMIZED RATS
case synthesis in glucagon-perfused livers was increased by steroid treatment only in the fed rats.
Effects of Epinephrine on Lipolysis and Cyclic AMP Accumula- tion in Epididymal Fat Pads from Normal and Adrenalectomized o-̂ ” I I I I I A” I Ruts-In agreement with the results of previous investigations (g-12), free fatty acid release by fat pads from fed or fasted adrenalectomized rats incubated with epinephrine was lower than normal (Table XII). Release in control incubations was also reduced by adrenalectomy. In another set of experiments, adrenal deficiency reduced glycerol release, expressed per mg of DNA, from fat pads incubated with epinephrine plus caffeine (Table XIII). Expressed on the same basis, control cyclic AMP levels were unchanged by adrenalectomy and were higher in the steroid-deficient pads incubated with epinephrine plus caffeine (Table XIII). Thus, the impairment in the lipolytic response was not due to reduced net cyclic AMP formation.
A
0 3 x10- 1x10-8 3x10-u 1x10‘~ 3x10-' 3x10-6 NOREPINEPHRINE CONCENTRATION (MI
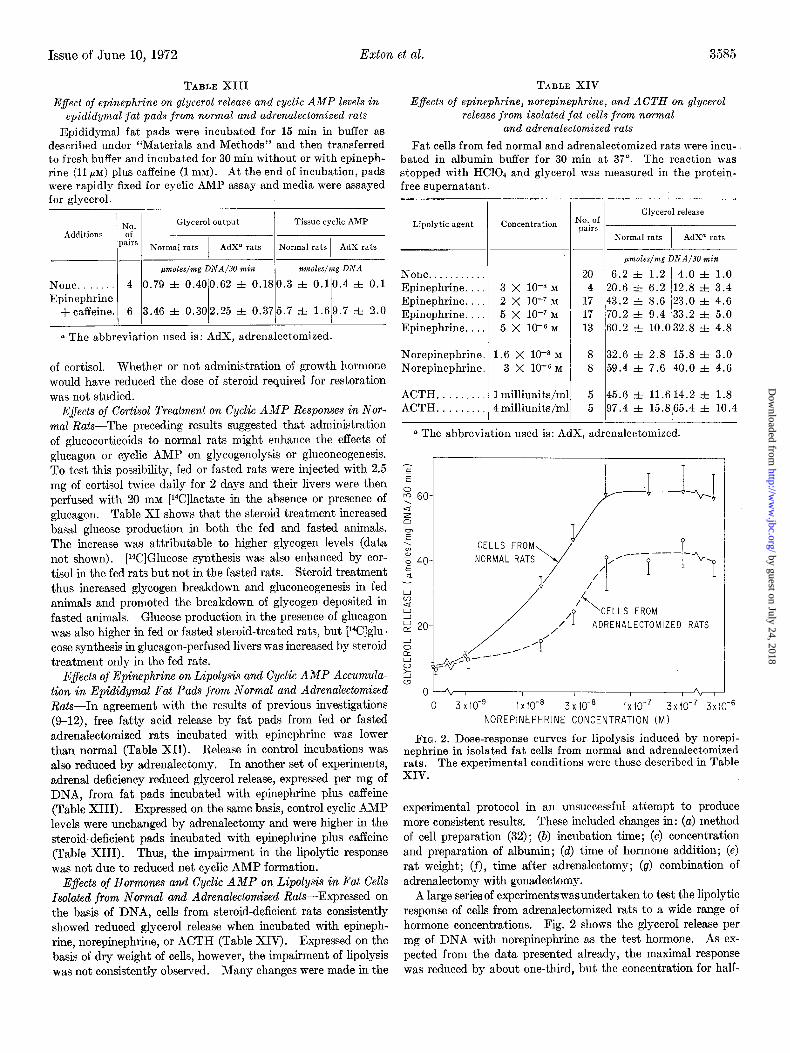
FIG. 2. Dose-response curves for lipolysis induced by norepi- nephrine in isolated fat cells from normal and adrenalectomized rats. The experimental conditions were those described in Table XIV.
E$ects of Hormones and Cyclic AMP on Lipolysis in Fat Cells Isolated from Normal and Adrenalectomized Rats-Expressed on the basis of DNA, cells from steroid-deficient rats consistently showed reduced glycerol release when incubated with epineph- rine, norepinephrine, or ACTH (Table XIV). Expressed on the basis of dry weight of cells, however, the impairment of lipolysis was not consistently observed. Many changes were made in the
experimental protocol in an unsuccessful attempt to produce more consistent results. These included changes in: (a) method of cell preparation (32) ; (b) incubation time; (c) concentration and preparation of albumin; (d) time of hormone addition; (e) rat weight; (f), time after adrenalectomy; (g) combination of adrenalectomy with gonadectomy.
A large series of experimentswasundertaken to test the lipolytic response of cells from adrenalectomized rats to a wide range of hormone concentrations. Fig. 2 shows the glycerol release per mg of DNA with norepinephrine as the test hormone. As ex- pected from the data presented already, the maximal response was reduced by about one-third, but the concentration for half-
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from
3586 Interaction of Glucocorticoids with Glucagon
TABLE XV E$ects of exogenous cyclic AMP and dibutyryl cyclic AMP on glycerol release from
fat cells isolated from normal and adrenalectowmed rats
Cells were incubated for 30 min as described in Table XIV.
Vol. 247, No. 11
Agent 1 Concentration / No. of pairs 1
None. Cyclic AMP. Cyclic AMP. . Cyclic AMP. DBcyclicAMP...... DB cyclic AMP.. . DB cyclic AMP.. . . .I
1)L.W
10 20 40
0.5 1 5
20 2 2
20 8 9
17
~mmoles/mg DNA
8.5 f 1.3 5.5 f 1.0
45.0 f 8.0 11.9 f 2.0 10.0 i 2.0 5.9 f 1.7 43.7 f 11.0 15.7 f 4.2
109.0 f 17.0 63.0 f 6.0
Glycerol release
NClrlllt31 I
ACK
p?wles/g dry wt
3.7 f 0.5 3.9 f 0.8 1.4, 14.0 1.4, 6.2 2.4, 14.0 2.2, 6.2
19.5 f 3.1 8.5 f 1.4b 4.3 f 0.9 4.2 f 1.2
19.0 f 4.4 11.3 f 2.8" 47.6 f 7.3 44.0 f 4.8
a The abbreviations used are: AdX, adrenalectomized; DB cyclic AMP, Ne,02’-dibutyryl cyclic AMP. b P < 0.005 compared with normal cells. E P > 0.1 compared with normal cells.
maximal response was largely unaltered. Essentially similar results were obtained with ACTH and epinephrine (Table XIV).
Fat cells from normal and adrenalectomized rats were also incubated with different concentrations of cyclic AMP or di- butyiyl cyclic AMP. Table XV shows that the concentration of exogenous cyclic AMP had to be raised to 40 mM to elicit a consistent lipolytic response. Expressed on the basis of cell DNA or dry weight, the response was reduced in steroid-deficient cells incubated with this concentration of nucleotide. As ex- pected, dibutyryl cyclic AMP promoted lipolysis at lower con- centrations than cyclic AMP (Table XV). Lipolysis in cells from adrenalectomized animals incubated with 1 or 5 mM di- butyryl cyclic AMP was subnormal on the basis of DNA but not significantly changed on the basis of dry weight.
glucagon in heart, liver, or adipose tissue is unchanged or in- creased by adrenal deficiency, other investigators have reported reduced cyclic AMP responses to these hormones (34, 35). It should be noted that only maximally effective concentrations of hormones were tested in the present study and that cyclic AMP levels were measured at only one time point (10 min). It is possible that the cyclic AMP response is impaired with more physiological levels of hormones or has an altered time course (28).
DISCUSSION
The present findings confirm and extend earlier observations indicating impairments of cyclic AMP-mediated hormone actions in adrenalectomized animals. Defects have been observed in
uivo and in vitro in free fatty acid and glycerol release from adipose tissue (8-12, 33-36)) glycogenolysis and gluconeogenesis in liver (1, 4, 13, 3335), and glycogenolysis in heart and skeletal muscle (14, 33, 34, 37). There is evidence that the following cyclic AMP-mediated hormone effects may also be dependent on ad- renal steroids. (a) Catecholamine actions on the cardiovascular system (3840), and on skin in response to cold (33-35). (b) Vasopressin action on water and Na+ flux in the toad bladder (41). (c) Glucagon stimulation of ureogenesis in the perfused liver (42) and induction of phosphoenolpyruvate carboxykinase in fetal liver explants (43).
There is additional indirect evidence that glucocorticoids may increase cyclic AMP formation in adipose tissue. For example, Fain (46) and Goodman (47) have observed that growth hormone and dexamethasone markedly increase the lipolytic response of fat cells or tissue t’o theophylline. More direct information has been obtained by Moskowitz and Fain (48) who observed that dexamethasone increased the accumulation of radioactive cyclic AMP in fat cells whose adenine nucleotides had been labeled with [3H]adenine. To date, no measurements of adenylate cyclase activity in fat cell membranes or homogenates from adrenalectomized animals have been reported. Senft et al. (49) have reported decreased phosphodiesterase activity in fat and other tissues treated with glucocorticoid.
The role of the adrenal steroids in these situations has beeit termed “permissive” (44) since it is to permit other hormones to produce normal responses. The present and other studies sug- gest the existence of a general mechanism for the supportive action of corticoids. In all the situations examined, the steroids appear t.o be required for maintenance of normal responsiveness of processes to cyclic AMP and not primarily for normal cyclic AMP accumulation in response to hormones activating adenylate cyclase.
In the cases of livers from fed rats and hearts from fed or fasted animals, the decreased responses induced by adrenalectomy were due to diminished sensitivity of the glycogenolytic or gluconeo- genie process to cyclic AMP. They did not reflect decreased levels of phosphorylase or of the limiting enzymes of gluconeo- genesis since normal responses could be achieved if sufficient hormone was administered. In the case of livers from fasted rats the impaired activation of gluconeogenesis by glucagon or cate- cholamine could have been due in part to a reduced maximum rate of the pathway (50) since administration of dibutyryl cyclic AMP or high concentrations of glucagon did not induce a normal response. With respect to adipose tissue, limitation of the lipolytic pathway as well as decreased sensitivity to cyclic AMP appear to be involved. Expressed on the basis of DNA, the maximum lipolytic responses to epinephrine, norepinephrine, ACTH, cyclic AMP, and dibutyryl cyclic AMP were consistently lower in steroid-deficient tissue.
Although the present and previous (14) observations indicate The nature and sit.e of the defect in phosphorylase activation that the accumulation of cyclic AMP induced by epinephrine or induced by adrenal insufficiency has been explored in the perfused
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from

Issue of June 10, 1972 Exton et al. 3587
heart (14). The impairment appears to reside in the activation of phosphorylase b since phosphorylase kinase activation by protein kinase is apparently normal (14). The action of phos- phorylase kinase from muscle has been shown to require Ca++ (30) and it has been suggested that the defect in adrenalectomy may be due to an intracellular deficiency of this cation, particu- larly since it can be remedied by raising the Ca++ concentration of the perfusion medium (14).
It is possible that the defect in the phosphorylase activation system in the steroid-deficient liver resembles that in the heart. It is currently being explored further. Preliminary results in- dicate unimpaired protein kinase activity in perfused livers from adrenal-insufficient rats as measured by a2P incorporation into Fl-histone in the presence and absence of glucagon or cyclic AMP.a
The role of adrenal steroids in the activation of gluconeogenesis by cyclic AMP remains obscure in the absence of precise informa- tion regarding the mechanism of action of cyclic AMP on this process. As in the case of hepatic glycogenolysis and in contrast to renal gluconeogenesis (52), the present findings do not indicate a major role for external Ca++ in the control of hepatic gluco- neogenesis. However, as noted below, the experiments do not exclude possible shifts in intracellular Ca++. Friedmann and Rasmussen (53) have observed that tetracaine, an anesthetic known to alter Ca++ binding and transport, blocks calcium efflux induced by cyclic AMP in the perfused liver and also the stimula- tion of glycogenolysis and gluconeogenesis.
Although the present data do not support a role for changes in intracellular metabolism of Ca++ in the “permissive” action of steroid on gluconeogenesis, they do not exclude it. This is be- cause it is uncertain to what extent changes in perfusate Ca++ alter the intracellular level of free Ca++. Although the data of Table VIII indicate that cyclic AMP activation of gluconeo- genesis is retained in perfusions with Ca++-free medium or EGTA, the magnitude of the activation is decreased. It is also possible that the alterations in the Ca++ level of the perfusate in the experiments of Table IX and those reported in Reference 28 did not change gluconeogenesis because they did not suffi- ciently alter the intracellular concentration of Ca++.
The nature of the defect in the lipolytic response to cyclic AMP also remains to be defined. Preliminary observations indicate that protein kinase activity is unimpaired in adipose tissue from adrenalectomized rats4 resembling the situation in heart and liver. Since it has been reported that Ca++ is required for lipase activa- tion (54), it is tempting to postulate that adrenal steroids might act by increasing free intracellular Ca++ in adipose tissue as has been proposed for heart (14).
In the general context of possible interrelationships between corticoids, cyclic AMP, and ions, the following observations should be noted. (a) Extremely small changes in intracellular ions, e.g. Ca++ are capable of exerting large effects on physiologi- cal processes, e.g. muscle contraction and phosphorylase k&se action (55, 56). (b) Ions are not distributed evenly within cells frequently being accumulated in mitochondria and endo- plasmic vesicles (57). (c) Cyclic AMP, or hormones which in- crease its accumulation, stimulates ion mo\-ement across cellular and subcellular membranes (51, 57-59). (d) Adrenal steroids
3 L. E. Mallette, T. A. Langan, and J. H. Exton, unpublished observations.
4 J. D. Corbin, unpublished observations.
affect calcium transport across intestine (60) and calcium accum- ulation in rat liver mitochondria (61). It is also well known that they influence Na+ and K+ movement in many cell types, and changes in the distribution of these ions across the plasma membrane may alter Ca++ flux (59, 62, 63). (e) Ca++, Na+, and K+ are known to influence many cyclic AMP-mediated hor- mone actions (29, 30, 52-54, 59, 60, 64).
These considerations suggest that the alterations in cyclic AMP responsiveness induced by steroids could be related to changes in intracellular ion metabolism as proposed by Rasmus- sen and Tenenhouse (65). As discussed above, the perfusions with EGTA and varying Ca++ levels did not rule out an impor- tant role for this ion in the control of hepatic glycogenolysis and gluconeogenesis since crucial shifts in Ca++ may still have oc- curred in these experiments. The possible roles of Na+ and K+ were not explored.
Current knowledge of steroid hormone action indicates that these agents stimulate the synthesis of specific proteins probably by increasing messenger RNA synthesis in the nucleus. It is possible that glucocorticoids may act in this way to maintain the levels of transport proteins involved in ion transfer across cellular and subcell&r membranes. Such a mechanism has been pro- posed for aldosterone. The “permissive” action of adrenal steroids might therefore involve the maintenance of a normal intracellular ionic environment so that the sensitivity of phys- iological processes to cyclic AMP is preserved.
Acknowledgments-We are very greatly indebted to Doctors G. A. Robison and R. W. Butcher for performing the measure- ments of cyclic AMP in liver reported in this study. We also thank Sandy Harper for her highly skilled assistance in the ex- periments with fat cells.
5.
6.
7.
8.
9.
10. 11.
12.
13.
14.
15.
16.
REFERENCES FRIEDMANN, N., EXTON, J. H., AND PARK, C. R. (1967) Biochem.
Biophys. &es: Commkn. 29,‘113 ’ FRIEDMANN, N., EXTON, J. H., AND PARK, C. R. (1968) Fed.
Proc. 27, 625 CORBIN, J. D., AND PARK, C. R. (1969) Fed. Proc. 28, 702 EXTON. J. H.. MALLETTE. L. E.. JEFFERSON. L. S.. WONG.
E. g. A., F&EIDMANN, k.; MILLER, T. B., Jo., AND P.&RI<; C. R. (1970) Recent Progr. Hormone Res. 26, 411
LONQ, C. N. H., KATZIN, B., AND FRY, E. G. (1940) Endo- crinology 26,309
JEANRENAUD, B., AND RENOLD, A. E. (1960) J. Biol. Chem. 236, 2217
FAIN, J. N., KOVACEV, V. P., AND Scow, R. 0. (1965) J. Biol. Chem. 240, 3522-3529
SCHOTZ, M. C., MASSON, G. M. G., AND PAGE, I. H. (1959) Proc. Sot. EXD. Biol. Med. 101, 159
SHAFRIR, E., S&SMAN, K. E., -IT’D STEINBERG, D. (1959) J. Lipid Res. 1, 459
RESHEF, L., AND SHAPIRO, B. (1960) Metabolism 9, 551 MAICKEL, R. P., AND BRODIE, B. B. (1963) Ann. N. Y. Acad.
Sci. 104, 1059 SHAFRIR, E., AND KERPEL, S. (1964) Arch. Biochem. Biophys.
106, 237 SCHAEFFER, L. D., CHENOITETH, M., AND DCNN, A. (1969)
Biochim. Biophys. Acta 192, 292-303 MILLER, T. B., EXTON, J. H., AND PARK, C. R. (1971) J. Biol.
Chem.’ 246, 3672-3678 EXTON. J. H.. AND PARIC. C. R. (1967) J. Biol. Chem. 242.
2622~i636 ’ \ I
MALLETTE, L. E., EXTON, J. H., AND PARIC, C. R. (1969) J. Biol. Chem. 244, 5713-5723
17. WOLLENBERGER, A., RISTAU, O., AND SCHOFFA, G. (1960) Pjluegers Arch. Gesamte Physiol. Menschen. Tiere 270, 399
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from

3588 Interaction of Glucocorticoids with Glucagon Vol. 247, No. 11
18. NEELY, J. R., LIEBERMEISTER, H., AND MORGAN, H. E. (1967) 41. HANDLER, J. S., PRESTON, A. S., AND ORLOFF, J. (1969) J. CZin. Amer. J. Physiol. 212, 815 Invest. 48. 823
19. CORNBLATH, M., RANDLE, P. J., PARMEGQIANI, A., AND MOR- 42. MILLER, L. L. (1961) Recent Progr. Hormone Res. 17, 539 GAN, H. E. (1963) J. Biol. Chem. 238, 1592-1597 43. WICKS, W. D. (1971) J. BioZ. Chem. 246, 217-223
20. BUTCHER, R. W., Ho, R. J., MENO, H. C., AND SUTHERLAND, 44. INGLB, D. J. (1952) J. Endocrinol. 8, XXIII E. W. (1965) J. Biol. Chem. 240, 4515-4523 45. PAUK, G. L., AND REDDY, W. J. (1971) Diabetes 20, 129
31. WIELAND, 0. (1963) in H. U. BERQMEYER (Editor), Methods of 46. FAIN, J. N. (1968) Endocrinology 82, 825-830 enzymatic analysis, p. 211, Academic Press, New York 47. GOODMAN, H. M. (1968) Endocrinology 82, 1027-1034
22. DOLE, V. P. (1956) J. Clin. Invest. 36, 150 48. MOSKOWITZ, J., AND FAIN, J. N. (1970) J. BioZ. Chem. 246, 23. CERIOTTI. G. (1952) J. Biol. Chem. 198, 297 1101-1107 24. 25. 26. 27.
28.
29.
30.
31.
32. 33.
34.
35.
36. 37.
38. 39.
40.
RODBELL; M. (1964) J. Biol. Chem. 239; 375-380 BURTON, K. (1956) Biochem. J. 62, 315 EXTON, J. H., AND PARIC, C. R. (1968) J. Biol. Chem. 243, 4189 BITENSKY, M. W., RUSSELL, V., AND BLANCO, J. (1970) Endo-
crinology 86, 154-159 EXTON, J. H., ROBISON, G. A., SUTHERLAND, E. W., ANDPARK,
C. R. (1971) J. BioZ. Chem. 246, 61666177 MEYER, W. L., FISCHER, E. H., AND KREBS, E. G. (1964) Bio-
chemistry 3, 1033 BROSTROM, C. O., HUNI~ELER, F. L., AND KREBS, E. G. (1971)
J. BioZ. Chem. 246, 1961-1967 HENDERSON, 111. J., MORGAN, H. E., AND PARK, C. R. (1961)
J. BioZ. Chem. 236, 2157-2161 KONO, T. (1969) J. BioZ. Chem. 244, 1772-1778 BRODIE, B. B., MAICI~EL, R. P., AND STERN, D. N. (1965) in
A. E. RENOLD AND G. F. CAHILL, JR. (Editors), Handbook of physiology, Section 5: Adipose tissue, p. 583, American Physiological Society, Washington, D. C.
BRODIE, B. B., DAVIES, J. I., HYNIE, S., KRISHNA, G., AND WEISS, B. (1966) Pharmacol. Rev. 18, 273
MAICICEL, R. P., STERN, D. N., TAKABATAKE, E., AND BRODIE, B. B. (1967) J. Pharmacol. Exp. Ther. 167, 111
GOODMAN, H. M. (1970) Endocrinology 86, 1064-1074 SHAEFFER, L. D., CHENOWIBTH, M., AND DUNN, A. (1969)
Biochim. Biophys. Acta 192, 304-309 FRITZ, I., AND LEVINE, R. (1951) Amer. J. Physiol. 166, 456 BROWN, F. K., AND REMINGTON, J. W. (1955) Amer. J. Physiol.
182, 279 LEFER, A. M., AND SUTFIN, D. C. (1964) Amer. J. Physiol. 206,
1151
49.
50.
51. FRIEDMANN, N., AND PARIZ, C. R. (1970) Proc. Nat. Acad. Sci. U. S. A. 66, 368
52. NGIATA, N., AND RASMUSSEN, H. (1970) Biochim. Biophys. Acta 216, 1
53. FRIEDMANN, N.. AND RASMUSSEN. H. (1970) Biochim. Biophys.
54. 55. 56.
57. 58.
59.
60.
61.
62. 63.
64.
65.
SENFT, G., SCHULTZ, G., MUNSKE, K., AND HOFFMANN, M. (1968) Diabetologia 4, 330
EXTON, J. H., AND PARIC, C. R. (1965) J. BioZ. Chem. 240, PC955-957
Acta 222,hl ’ ,
RIZACIC. M. A. (1964) J. Biol. Chem. 239. 392-395 BIANCH;, C. P. (1968) Fed. Proc. 27, 126’ OZAWA, E., AND EBASHI, S. (1967) J. Biochem. (Tokyo) 62,
285 RASMUSSEN, H. (1970) Science 170, 404 WAKABAYASHI, K., KAMBERI, I. A., AND MCCANN, S. M. (1969)
Endocrinology 86, 594 CL.~USEN, T. (1970) in B. J~ANRENAUD AND D. HEPP (Editors),
Adipose tissue: regulation and metabolic functions, Academic Press, New York p. 66.
HARRISON, H. E., AND HARRISON, H. C. (1960) Amer. J. Phys- iol. 199, 265
KIMBERG, D. V., AND GOLDSTEIN, S. A. (1966) J. Biol. Chem. 241, 95-103
REUTER, H., AND SEITZ, N. (1968) J. Physiol. 196, 451 BLAUSTEIN, M. P., AND HODOICIN, A. L. (1969) J. Physiol. 200,
431 NAMM, D. H., MAYER, S. E., AND MALTBIE, M. (1968) Mol.
Pharmacol. 4, 522 RASMUSSEN, H., AND TENENHOUSE, A. (1968) Proc. Nat. Acad.
Sci. U. S. A. 69, 1364
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from

Corbin and Charles R. ParkJohn H. Exton, Naomi Friedmann, Ellen Hee-Aik Wong, J. Parke Brineaux, Jackie D.Gluconeogenesis and Glycogenolysis in Liver and of Lipolysis in Adipose TissueInteraction of Glucocorticoids with Glucagon and Epinephrine in the Control of
1972, 247:3579-3588.J. Biol. Chem.
http://www.jbc.org/content/247/11/3579Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/11/3579.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 24, 2018http://w
ww
.jbc.org/D
ownloaded from