interaction between adenosine and angiotensin ii in renal...
TRANSCRIPT

ACTAUNIVERSITATISUPSALIENSISUPPSALA2007
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 237
Interaction between Adenosineand Angiotensin II in RenalAfferent Arterioles of Mice
ENYIN LAI
ISSN 1651-6206ISBN 978-91-554-6822-4urn:nbn:se:uu:diva-7702


To my family, especially to Lingli and Siyun


LIST OF PAPERS
The thesis is based on the following papers, which will be refereed to in
the text by their Roman numerals:
I. Andreas Patzak, En Yin Lai, Ralf Mrowka, Andreas Steege,
Pontus B. Persson, A. Erik G. Persson AT1 receptors mediate an-
giotensin II induced release of nitric oxide in afferent arterioles
Kidney International, Vol. 66 (2004), pp.1949-1958.
II. En Yin Lai, Andreas Patzak, Andreas Steege, Ralf Mrowka,
Russell Brown, Nadine Spielmann, Pontus B. Persson, Bertil B.
Fredholm, A. Erik G. Persson Contribution of adenosine recep-
tors in the control of arteriolar tone and adenosine-angiotensin II
interaction Kidney International, (2006) 70, 690-698.
III. En Yin Lai, Andreas Patzak, A. Erik G. Persson Low dose angio-
tensin II enhances cytosolic calcium concentration and contractile
responses of afferent arterioles to adenosine Manuscript.
IV. En Yin Lai, Peter Martinka, Michael Fähling, Ralf Mrowka,
Andreas Steege, Adrian Gericke, Mauricio Sendeski, Pontus B.
Persson, A. Erik G. Persson, Andreas Patzak Adenosine restores
angiotensin II-induced contractions by receptor-independent en-
hancement of calcium sensitivity in renal arterioles Circulation
Research 2006 99: 1117 - 1124.
Reprints were made with permission from the publisher.


CONTENTS
INTRODUCTION ........................................................................................11Renal blood supply...................................................................................12Glomerular filtration rate .........................................................................12Juxtaglomerular apparatus........................................................................12Tubuloglomerular feedback .....................................................................13Renin-angiotensin system ........................................................................13Ang II and NO..........................................................................................13Adenosine.................................................................................................13
AIMS ............................................................................................................15Study I: .....................................................................................................15Study II:....................................................................................................15Study III: ..................................................................................................15Study IV: ..................................................................................................15
MATERIAL AND METHODS....................................................................16Reagents ...................................................................................................16Animals ....................................................................................................16Dissection and perfusion ..........................................................................17Measurement of isotonic contraction .......................................................17Confocal microscopy and NO measurement............................................17Quanticell 900 system and Calcium imaging...........................................18Isolation of vessels ...................................................................................18Statistics ...................................................................................................18
PROTOCOLS, RESULTS & DISCUSSIONS .............................................20Study I ......................................................................................................20
Protocols ..............................................................................................20Results .................................................................................................20Discussion............................................................................................20
Study II.....................................................................................................21Protocols ..............................................................................................21Results .................................................................................................22Discussion............................................................................................22
Study III ...................................................................................................23Protocols ..............................................................................................23

Results .................................................................................................23Discussion............................................................................................24
Study IV ...................................................................................................24Protocols ..............................................................................................24Results .................................................................................................25Discussion............................................................................................25
FIGURES AND LEGENDS .........................................................................27
CONCLUSIONS ..........................................................................................36Study I ......................................................................................................36Study II.....................................................................................................36Study III ...................................................................................................36Study IV ...................................................................................................36
PAPER LIST OF PHD STUDY ...................................................................38Cover Page Stories ...................................................................................38Journal Publications .................................................................................39
ACKNOWLEDGEMENTS..........................................................................41
REFERENCES .............................................................................................43

ABBREVIATIONS
A1AR
A1(-/-)
A2AR
Ado
Af
Ang II
AT1AR
AT2 AR
CLSM
[Ca2+]i
DAF-FM DA
EC
Ef
Fura-2 AM
GFR
Adenosine A1-receptor
Adenosine A1-receptor knockout
mice
Adenosine A2-receptor
Adenosine
Afferent arteriole
Angiotensin II
Angiotensin II type 1 receptor
Angiotensin II type 2 receptor
Confocal laser scanning microscopy
Intracellular calcium concentration
4-amino-5-methylamino-2´,7´-
difluorofluorescein diacetate
Endothelial cell
Efferent arteriole
5-Oxazolecarboxylic acid, 2-(6-
(bis(2-((acetyloxy)methoxy)-2-
oxoethyl)amino)-5-(2-(bis(2-
((acetyloxy) methoxy)-2-
oxoethyl)amino)-5-
methylphenoxy)ethoxy)-2-
benzofuranyl)-, (acetyloxy)methyl
ester
Glomerular filtration rate

JGA
L-NAME
MD
NE
NO
NOS
QC 900
RBF
RAAS
TGF
VSMC
Juxtaglomerular apparatus
N -nitro-L-arginine methyl ester
Macula densa
Norepinephrine
Nitric oxide
Nitric oxide synthase
Applied imaging Quanticell 900
Renal blood flow
Renin-angiotensin-aldosterone sy-
stem
Tubuloglomerular feedback
Vascular smooth muscle cell

11
INTRODUCTION
The autoregulation of renal perfusion and filtration guarantees a continuous and efficient function of the kidney in the sense of plasma clearance of me-tabolites and control of water and salt balance. Renal autoregulation is mainly based on the myogenic responses of renal vessels and the tubu-loglomerular feedback (TGF) [1, 2]. The TGF mechanism consists of a se-quence of events linking salt load in the distal tubule to the vascular tone of afferent and possibly efferent arterioles, resulting in short-term changes to the filtration rate and a fast control of the sodium balance.
The present work focuses on the role of adenosine (Ado), angiotensin II (Ang II), nitric oxide (NO) and the renal vascular regulation. These vasoac-tive substances contribute significantly to the modulation and resetting of the TGF. Ang II is a very potent vasoconstrictor and increases renal vascular resistance. It reduces renal blood flow (RBF) and glomerular filtration rate (GFR) [3, 4]. It has been shown using microdialysis that Ang II induces re-lease of NO in the kidney [5]. Further, studies in isolated perfused glomeru-lar arterioles and in the macula densa also indicate an Ang II induced NO release [6-8]. NO ranks highly among the most effective vasodilators in the kidney [9]. It is produced by constitutive NO synthases (NOS), which are expressed in the endothelium (eNOS) and in macula densa cells (nNOS) [10]. Results of in vivo studies suggest a significant influence of NO on arte-riolar tone and on the TGF [11-14]. Although there is a general agreement in regard to an Ang II induced NO release, the role of Ang II receptors in this context is not clear.
Ado plays an important role in the blood flow control of organs. The main effect of Ado is vasodilation mediated by the Ado type 2 receptors (A2AR) [15-19]. In contrast, vasoconstrictor abilities of Ado have been observed in the kidney. This is due to a significant expression of the type 1 receptor (A1AR), mediating vasoconstriction. This vasoconstrictor action of Ado via A1AR is related to its proposed function as a mediator of TGF [20-22].
Studies suggest an interaction between Ado and Ang II in the control of renal perfusion, filtration and TGF. Inhibition or lack of A1AR as well as inhibition of the angiotensin converting enzyme reduce the TGF [23]. This hints at a modulatory role of Ang II in the TGF. On the other hand, Ado can modulate the action of Ang II [15, 19]. Ado and Ang II interact mainly syn-ergistically in the kidney. However, the mechanisms of this interaction and the contribution of Ado and Ang II receptors are not clear. Considering the

12
importance of Ado and Ang II for renal function, increase in the knowledge about the underlying mechanisms is relevant for understanding of the patho-genesis of renal dysfunctions.
Renal blood supply Human renal blood flow normally is about 1100 ml/min for both kidneys. The renal artery leads blood into the kidney through the interlobar arteries, arcuate arteries, interlobular arteries, afferent arterioles and the glomerular capillaries. High hydrostatic pressure of 60 mmHg in the glomerular capil-laries causes rapid fluid filtration into the tubular system. Kidneys regulate hydrostatic pressures in both glomerular and peritubular capillaries by ad-justing the resistances of afferent and efferent arterioles which is very impor-tant for the GFR and RBF [24].
Glomerular filtration rate The GFR is determined by the balance of hydrostatic and colloid osmotic forces acting across the capillary membrane and the capillary filtration coef-ficient, the product of the permeability and filtering surface area of the capil-laries. The glomerular capillaries have a higher filtration rate than most other capillaries because of a higher glomerular hydrostatic pressure and a larger capillary filtration coeffient. GFR is about 125 ml/min in an adult [25].
Juxtaglomerular apparatus The juxtaglomerular apparatus (JGA) [26-28] consists of the macula densa (MD) [26], the afferent and efferent arterioles and the mesangium. JGA is found at the hilus of the renal glomerulus where the thick ascending limbs (TAL) of the Henle loop thus to the glomerulus of origin. The same nephron changes its morphologic characteristics as it comes in contact with the vas-cular pole. JGA permits interactions between the tubular epithelial cells and the arteriolar smooth muscle cells. The MD senses the fluid load, on the basis of the concentrations of sodium chloride in distal tubule, and then ad-justs the afferent arteriolar tone and renin release [29]. The juxtaglomerular granular cells synthesize, store and release renin.

13
Tubuloglomerular feedback The TGF [20, 29] is a very important mechanism which operates as a nega-tive feedback. TGF activates a response to the change in GFR by altering afferent arteriolar tone and renin release from the granular cells. The TGF mechanism refers to the changes of Na+, Cl- and K+ concentrations in the TAL tubular fluid that are sensed by the macula densa cells. Increasing lu-minal sodium concentration increases the magnitude of the TGF response.
Renin-angiotensin system The most important function of the renin-angiotensin system is to normalize the changes in extracellular fluid volume and arterial pressure when the amount of sodium intake changes. Kidneys play a role in long-term regula-tion of arterial pressure by excreting variable amounts of sodium and water. Kidneys contribute also to short-term arterial pressure regulation by secret-ing renin that ultimately leads to formation of the vasoconstrictor Ang II. The renin-angiotensin system is an automatic feedback mechanism that helps normalize the arterial pressure at different salt intake levels. The renin is synthesized and stored in an inactive form prorenin in the juxtaglomerular cells (JG cells) of the kidneys. A decrease in arterial pressure causes many of the prorenin molecules in the JG cells to split and release renin. The renin can remain around JG cells as well as the smooth muscle cells of afferent and efferent arterioles modulating RBF and TGF through the action of Ang II.
Ang II and NO Afferent arterioles (Af) are key elements in controlling the preglomerular resistance, glomerular hemodynamics and GFR. The juxtaglomerular cells are the main source of renin production and the high level of Ang II in Af. To balance the Ang II effect, strong activities of vasodilatory systems are assumed. Af pretreated with the NO synthase inhibitor L-NAME, and the endothelial NO synthase knockout mice, demonstrate increased Ang II sensi-tivity increased, which indicates the activation of the NO release by Ang II [7].
AdenosineAdenosine is involved in the regulation of intracellular metabolism and func-tion. It acts predominantly on extracellular receptor subtypes. Adenosine

14
receptors belong to the G protein coupling receptors superfamily which has A1, A2A, A2B and A3 subtypes [30-33]. A1AR in the rodent kidney are present in Af, glomeruli including mesangial cells, JG cells, et al.[34-38]. The Ado vasoconstrictory action in Af and its cell metabolism are the main reasons that scientists investigated Ado as a mediator of TGF [20, 39-41]. Activation of A1AR leads to constriction of the Af. The recent observation of a com-plete loss of TGF activity in micropuncture studies of A1(-/-) knockout mice strongly supports the contention that Ado is a mediator in this mechanism [21, 22]. It is important for TGF that the A1AR mediate vasoconstriction in the preglomerular arterioles [42, 43]. Adenosine A2AR family consists of A2AAR and A2BAR subtypes [44]. A2AAR in the rodent kidney is present in glomerular epithelial cells and capillaries. A2BAR in the rodent kidney is present in preglomerular microvessels. Both of A2AAR and A2BAR have vasodilatory effect. The arterioles regulate RBF in conjunction with the dila-tory A2AAR [45, 46]. The A3 receptor is present in the whole kidney of ro-dent.

15
AIMS
The general aim of the thesis was to investigate the interaction between adenosine and angiotensin II in Af of mice. The following questions were in the center of interest.
Study I: Is the Ang II induced NO release in Af specific for Ang II? Which Ang II receptors are involved? What is the physiological importance of Ang II in-duced NO release in the juxtaglomerular apparatus?
Study II: How do isolated perfused Af respond to Ado? How does Ang II modulate the Ado effect on Af? How does Ado modulate Ang II responses? Which receptors are involved?
Study III: What is the role of cytosolic calcium in the interaction between Ado and Ang II? Which receptors are involved in the interaction?
Study IV: Does Ado influence the desensitization of Ang II responses in mice? What are the mechanisms of the re-sensitization of Ang II responses? What is the physiological significance of this Ado action?

16
MATERIAL AND METHODS
ReagentsDMEM/F12 (Invitrogen AB, Lidingö, Sweden), K+-solution, angiotensin II (Ang II), adenosine (Ado), norepinephrine (NE), CGS21680, CPA, 8-cyclopentyltheophylline (CPT), 4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol (ZM241385), inosine, PD123.319, ZD7155, 8-(p-sulfophenyl)theophylline (8-SPT), S-(4-nitrobenzyl)-6-thioinosine (NBTI), ethyleneglycol-bis( -aminoethyl)-N,N,N ,N -tetraacetic acid (EGTA), (4-(4-Fluorophenyl)-2-(4-methylsulfinyl)-5-(4-pyridyl)1H-imidazole) (SB203580), 1,2-Dimethoxy-12-[1,3]benzodioxolo[5,6-c] phe-nanthridinium chloride (Chelerythrine), (+)-(R)-trans-4-(1-aminoethyl)-N-(4-pyridyl) cyclohexanecarboxamide dihydrochloride monohydrate (Y-27632), L-NAME, DAF-FM, and 8,8'-[Carbonylbis[imino-3,1-phenylenecarbonylimino(4-methyl-3,1-phenylene)carbonylimino]] bis-1,3,5-naphthalenetrisul-fonic acid hexasodium (suramin) were from Sigma-Aldrich (Stockholm, Sweden). Fura-2 Calcium Imaging Calibration Kit (F-6774) and 1-[6-Amino-2-(5-carboxy-2-oxazolyl)-5-benzofuranyloxy]-2-(2-amino-5-methylphenoxy)ethane-N,N,N',N'-tetraacetic acid (Fura-2 AM) were from Calbiochem (VWR International AB, Stockholm, Sweden). Plu-ronic F-127 was from Molecular Probes (AB Göteborgs Termometerfabrik, Sweden). N-(4-acethylphenyl)-2-[4-(2,3,6,7-tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purin-8-yl)phenoxy]-acetamide (MRS1706) was from Tocris Cookson Ltd. (Bristol, United Kingdom).
AnimalsC57BL6 mice, Ado A1AR knockout A1(-/-) and their controls A1(+/+) mice were used in the experiments. A1(-/-) and A1(+/+) were generated as de-scribed by Johansson et al.[47]. Mice were fed with standard chow and al-lowed free access to water.

17
Dissection and perfusion Dissections and perfusions [7, 17, 48, 49]: Mice were killed by cervical dis-location and the kidneys were removed and sliced along the corticome-dullary axis. The Af were dissected using sharpened forceps under a stereo-scopic microscope at 4°C in albumin enriched Dulbecco´s Modified Eagle Medium (DMEM). The Af were identified by preparation of the arterial tree such as the interlobular artery and the attached glomerulus. The Af with their glomeruli intact were transferred into a thermoregulated chamber fixed on the stage attached to one of the specific setups including a laser confocal microscope, an inverted microscope and an Applied Imaging Quanticell 900 system (QC 900). The Af were perfused using a perfusion system which allowed adjustment of holding and perfusion pipettes. The holding pipette had a 26 µm aperture at the tip and aspirated the Af. The perfusion pipette had a 5 µm opening which was put into the lumen of Af. The perfusion pres-sure was 100 mmHg at the pressure head which was expected as the physio-logical pressure and flow.
Measurement of isotonic contraction The experiments were recorded on video tape using super video system (SVHS) from a Nikon water immersion objective lens and a digital camera. Video sequences were digitized using a frame grabber card. The vessel lu-minal diameters were determined using customized software. Basal control values were obtained at the end of the adaptation period. All series were measured for statistical analysis of steady state responses at the last 10 sec-onds of a 2 minutes treatment period.
Confocal microscopy and NO measurement The confocal laser scanning microscopy (CLSM) systems is based upon a simple optical principle. In conventional fluorescence microscopy, the image quality suffers from fluorescence emission from parts of the specimen out-side the plane of focus. In CLSM the specimen is illuminated in the focal plane by using suited optics. A tiny diaphragm is placed in front of pho-tomultiplier tubes to ensure that all out of focus fluorescence light does not reach the detector. By proper optical alignment of the illumination and de-tection light paths only emitted light from the optical plane is collected. The optical slices can be cut through the specimen by adjusting the microscope.
The NO indicator [50, 51] DAF-FM DA was used for detecting the NO production in Af. DAF-FM diacetate is cell permeable and diacetylated by intracellular esterases to become DAF-FM. It reacts with NO in presence of

18
oxygen forming green-fluorescent triazolofluoresceins. DAF-FM DA was loaded from the luminal side for 45 minutes. Measurements were carried out using a confocal system. The relative changes of the DAF-FM fluorescent product concentration were determined by calculating the ratio of the fluo-rescence intensities.
Quanticell 900 system and Calcium imaging Afferent arterioles were perfused and loaded with Fura-2 AM in the bath solution for 45 min. The perfusion system was mounted on the inverted mi-croscope stage. Fluorescence was measured using QC 900. The arterioles were excited alternately at 340 nm and 380 nm. The emission was measured at 510 nm. The ratio of 340/380 was used to determine the intracellular cal-cium concentration after calibration experiments using Fura-2 Calcium Im-aging Calibration Kit (F-6774) in our setup. Intracellular calcium concentra-tions were calculated from the obtained calibration formula:
[Ca2+]i = Kd x Beta x (R-Rmin)/(Rmax-R)(Kd = 224, Beta = 3.193, Rmin = 0.485, Rmax = 3.12, Calcium unit: µM) [52].
Isolation of vessels Mice preglomerular arterioles were isolated using a modified iron oxide-sieving technique. Mice were anaesthetized and the left kidney ventricle was cannulated and perfused with ice-cold iron-oxide suspension containing 1% Fe2O4. The kidneys were decapsulated and the cortex was dissected with a razor blade. The tissue was transferred to the PBS solution and magnetized for separating renal vessels. The iron oxide loaded tissue was resuspended in PBS and passed three times through decreasing size needles of diameters from 0.9 to 0.6 mm. Then the vascular suspension was filtered through a sieve with 80 - 100 m pores. The remaining vessels were immediately stored at -80°C for PCR investigation.
StatisticsThe Brunner test for analysis of longitudinal data was used for dose depend-ent changes in the arteriolar diameters and the differences between two groups in the amount of diameters in concentration-response-curves, and for differences in the dose dependent changes in diameter between the groups. The Mann-Whitney U-test was applied for the comparison of drug effect due to bolus application, the comparison of fluorescence measurements, and the comparison of luminal diameters of the different groups in the steady state

19
during the control situation. Analysis of variance (ANOVA) for repeated measurements (non-parametric Brunner test) was used to test time-dependent changes in the arteriolar diameter and to check differences be-tween the groups. Post hoc comparisons were performed using Tuckey’s test. Wilcoxon tests were applied for the comparison of treatment effects on basal diameter and the comparison of control diameters between groups. Data are presented as mean SEM. The confidence level p value was set to 0.05 for all statistical tests.

20
PROTOCOLS, RESULTS & DISCUSSIONS
Study I Protocols Dose response curves for Ang II (10-12 - 10-6 mol/l) were obtained in iso-lated, perfused Af. To test the effects of the Ang II receptors, the dose re-sponse curves for Ang II were measured after AT1 antagonist ZD7155 and AT2 antagonist PD123.319 pretreatment.
To investigate whether Ang II activates NO and which receptor subtype is involved. ZD7155 and PD123.319 were applied from the luminal side. This protocol allows the investigation of the desensitization of Ang II action on NO release. To block NO production, Af were pretreated with L-NAME before applying Ang II.
ResultsAng II reduced Af luminal diameter about 30% at 10-8 mol/l and 40% at 10-6
mol/l. PD123.319 pretreatment did not change the Ang II sensitivity of Af. To test a putative vasodilatory action of Ang II via AT2AR, Af were precon-stricted with NE. Ang II did not change the luminal diameter when it was applied in concentrations from 10-12 to 10-6 mol/l when AT1AR were blocked with ZD7155. The time dependent changes in the luminal diameter by Ang II (10-8 mol/l) bolus application with or without L-NAME pretreatment are shown in Fig. 1. Bolus application of Ang II led to a strong contraction which was enhanced by L-NAME pretreatment.
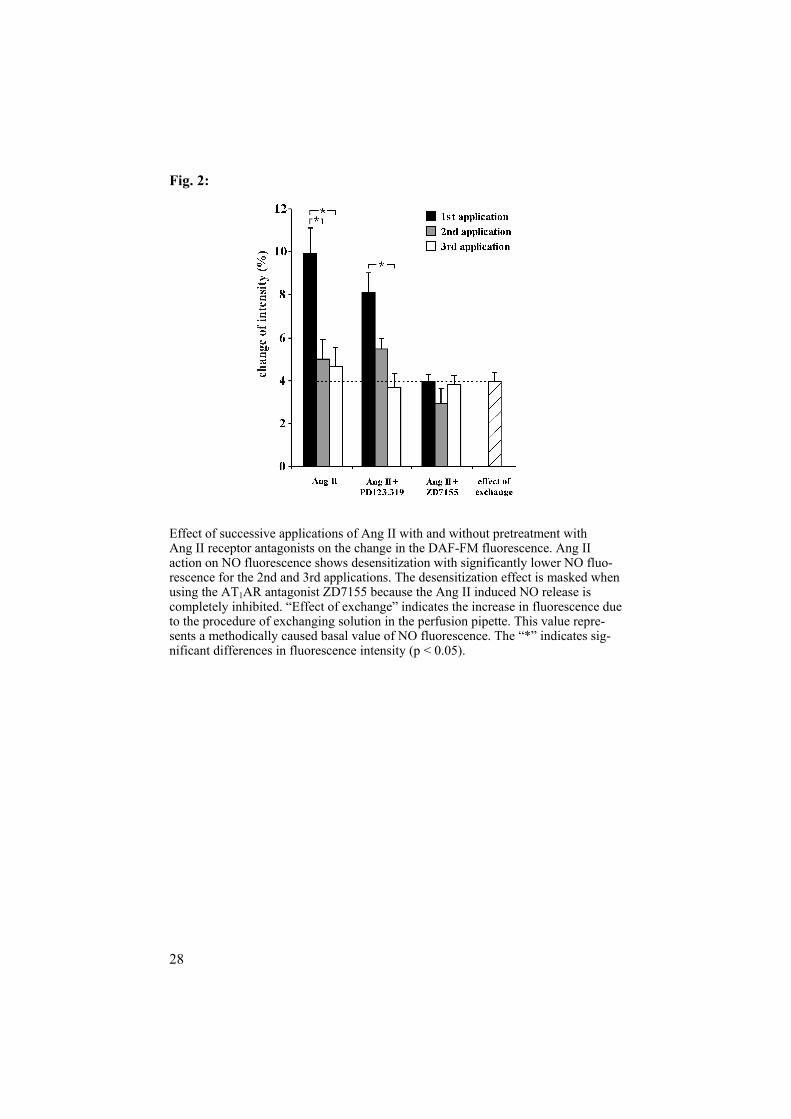
NO imaging: Ang II (10-7 mol/l) increased the fluorescence intensity about 10% when applied from the luminal and bath side. PD123.319 pre-treatment did not influence the intensity, but ZD7155 reduced the intensity and thus the NO release. There were no differences among the first, second and third applications of Ang II when pretreated with ZD7155 (Fig. 2). L-NAME pretreatment abolished the Ang II induced NO release.
DiscussionThe nonspecific NO synthases inhibitor L-NAME enhanced the contractile response to a bolus application of Ang II [7]. Ang II activates NOS to pro-

21
duce NO which counteracts the vasoconstrictory effect of the peptide. The AT1AR are involved in the Ang II induced NO release. Ang II increased the fluorescence intensity by about 10%. The specific AT2 inhibitor PD123.319 did not affect the increase of the fluorescence. L-NAME pretreatment abol-ished not only the specific Ang II effect but also the effect of exchange. The results demonstrate the AT1AR mediated NO release from the isolated, per-fused Af of mice [53-56]. Some investigators have suggested that AT2 ARare important mediators for the vasodilatory action of Ang II. In the rat kid-ney, AT2AR stimulation has been associated with the increased production of bradykinin, NO, and cGMP [57, 58]. A study in rabbits suggests that AT2AR activation counteracts AT1 mediated vasoconstriction in the renal cortex, but counteracts AT1-mediated vasodilatation in vessels controlling medullary perfusion [59]. In our experiments, the second and the third appli-cations of Ang II were significantly less effective in enhancing the fluores-cence intensity. The data show a clear desensitization of the Ang II induced enhancement of fluorescence intensity while AT1AR were intact. Desensiti-zation [60, 61] is well known for the G-protein coupled receptors. The de-sensitization of the Ang II induced increase in fluorescence supports the hypothesis of AT1 mediated NO release.
Study II Protocols Concentration response curves were measured in the perfused Af of C57BL6, A1 (-/-) and A1 (+/+) mice. Ado concentration-response-curves were tested from 10-11 to 10-4 mol/l. A series of concentration-response-curves were measured for CPA (selective A1AR-agonist), CGS21680 (selec-tive A2AAR-agonist), CPT (selective A1AR-antagonist), ZM241385 (selec-tive A2AAR-antagonist) and 8-SPT (non-selective Ado receptor antagonist). Functional characterization of Ado receptors was also done by treatment with CPT (10-5 mol/l), CGS21680 (10-12-10-4 mol/l), ZM241385 (10-7 mol/l), the combination of CPT and ZM241385 as well as ZM241385 and MRS1706 (selective A2BAR-antagonist, 10-6 mol/l), 8-SPT (5x10-5 mol/l), and a following parallel application together with Ado (10-11 - 10-4 mol/l). A possible function of A3AR [62] was tested by application of inosine (10-11 - 10-4 mol/l).
The concentration-response of Ang II (10-14 - 10-6 mol/l) was measured in combination with Ado (10-8 or 10-5 mol/l). We tested the role of Ado-receptors in the interaction by applying CPT with Ado 10-8 mol/l, and ZM241385 with MRS1706 10-6 mol/l together with Ado 10-5 mol/l, respec-tively. Furthermore, to check the influence of Ang II on the response of the

22
Af to Ado, Ang II (10-10 mol/l) and Ado (10-11 to 10-4 mol/l) were applied simultaneously.
ResultsAf responded biphasically to increasing Ado concentrations (Fig. 3). The degree of the constrictions for a single experiment did not exceed 15% of the control luminal diameter. Ado in the concentration range of 10-10 to 10-6
mol/l caused a 7% reduction in luminal diameter. Concentrations higher than 10-6 mol/l dilated the Af. The A1AR agonist CPA reduced diameters by 12% at 10-6 mol/l. The A2AAR agonist CGS21680 increased diameter by 12% at 10-7 mol/l. There was no change detected by the applications of CPT (A1AR antagonist), ZM241385 (A2AAR antagonist) and 8-SPT (non-selective Ado receptor antagonist). Pretreatment with CPT abolished the constrictory re-sponse of Ado. In A1(-/-) mice, no constrictory response was observed. The Af diameter of A1(-/-) mice increased continuously with higher Ado concen-tration. The response differed significantly compared to the A1(+/+) mice (Fig. 4). The Ado concentration-response-curve had a biphasic shape and corresponded to the C57BL6 mice.
DiscussionStudy II showed that A1AR, A2AAR and A2BAR subtypes were involved in the control of afferent arteriolar tone. Physiological concentrations of Ado activated vasoconstractor A1AR as well as vasodilator A2AR. Thus all, the response to Ado was rather small. A1AR mediated actions could demonstrate the selective A1AR agonist CPA application which decreased luminal diame-ters concentration dependently with a 12% reduction. This suggests that the A1AR mediated vasoconstriction is moderate. The specific A1AR antagonist CPT did not change luminal diameters significantly. A2AAR activator CGS21680 increased 12% diameter. The constrictory phase of the concen-tration-response to Ado was completely abolished in the A1(-/-) mice ex-periments. The concentration-response of Af in A1(+/+) did not differ from that of the C57BL6 strain. The dilatory effect of CGS21680 points to a sig-nificant expression of A2AAR, which could be involved in the dilatory re-sponse at lower Ado concentrations. While affinity constants of A2AAR for Ado are in the range of 10-8 mol/l, the affinity of A2BAR is lower at about 10-
5 mol/l [63]. Ado plays a role as a mediator in the TGF [64, 65]. The constrictory ef-
fect of Ado on Af tone seems to be modest. However, we estimate that a 10% diameter reduction may increase vascular resistance by about 50%. The constrictory power is sufficient to be responsible for the changes in arteriolar diameter related to the TGF. Ado administered at low concentration (10-8
mol/l) to the bath solution amplifies the Ang II induced constriction of the

23
Af. High concentration of Ado (10-5 mol/l) did not change the Ang II re-sponse of Af. It is assumed that the interaction between Ado and Ang II is due to synergistic action of AT1AR and A1AR on phospholipase C [66]. This hypothetic mechanism may be interfered by the activation of A2AR at high Ado concentration. The Ang II induced constriction in Af of rabbits was enhanced by Ado in excess of an additive effect [19]. The idea of the interaction between Ado and Ang II is supported by results of a recent study showing a reduced response of Af in mice lacking A1AR upon Ang II [16]. The deletion of AT1AR or angiotensin-converting enzyme restrained TGF responses in these knockout mice [67, 68]. Altogether, these studies indicate that synergistic interaction between Ado and Ang II is important for renal blood flow control and TGF.
Study III Protocols Ang II was applied to the perfused Af from bath solution in the concentra-tion of 10-12, 10-10 and 10-8 mol/l, respectively, for 16 min. The Ado concen-tration response was measured by cumulative bath application in concentra-tions from 10-11 to 10-4 mol/l together with Ang II (10-12, 10-10 and 10-8 mol/l, respectively) and without Ang II. The Ang II concentration response curve was measured by cumulative application of Ang II (10-11 to 10-6 mol/l, each for 2 min) with or without the selective AT1AR antagonist ZD7155, 10-7
mol/l. Fluorescence was measured using QC 900 to test the dose related effects
of Ang II on the cytosolic calcium concentration of the perfused Af. The Af were loaded with Fura-2 AM. The cytosolic calcium concentrations were measured in the arteriolar wall during cumulative application of Ang II (10-11
to 10-6 mol/l) with or without ZD7155. Changes in the cytosolic calcium concentration were measured the response to bolus applications of Ado (10-8
and 10-5 mol/l) and Ado + Ang II 10-10 mol/l with or without ZD7155 in the arteriolar wall. Further, the Af were treated with CPT while applying Ado 10-8 + Ang II 10-10 mol/l and the combination of ZM241385 and MRS1706 was used to determine contribution of A2AR during application of Ado 10-5
+ Ang II 10-10 mol/l on calcium transients.
ResultsAdo (10-11 - 10-4 mol/l) induced modest changes in arteriolar diameter. How-ever, treatment with Ang II in concentrations of 10-12, 10-10 and 10-8 mol/l, respectively, increased the contractile response significantly compared to Ado alone (Fig. 5). Ang II (10-11 - 10-6 mol/l) led to a concentration depend-

24
ent constriction of the Af. Treatment with the AT1AR antagonist ZD7155 (10-7 mol/l) prevented the response to cumulative applications. Cumulative application of Ang II (10-11 - 10-6 mol/l) increased cytosolic calcium concen-trations dose dependent. The maximum increase in the calcium concentra-tion was observed at 10-7 mol/l Ang II (Fig. 6).
DiscussionStudy III shows a significant influence of Ang II on the vasoconstrictor re-sponse of Ado in isolated, perfused Af of mice. Ang II enhances Ado in-duced vasoconstriction even in very low concentration of 10-12 mol/l. This is noteworthy, because Ado itself induces only modest changes in arteriolar diameters in our model. The results agree with the contention that Ang II modulate the TGF in the kidney [45]. Our data further indicate that the TGF mechanism depends on plasma and renal Ang II concentrations which are not only able to constrict arterioles, but also can increase the sensitivity and strength of this mechanism.
Ado (10-8 mol/l) increased cytosolic calcium concentrations which is in line with its vasoconstrictory action in Af. Higher concentrations of Ado (10-
5 mol/l) further increased calcium transients. The Ado induced increases in cytosolic calcium were significantly enhanced by simultaneous application of Ang II (10-10 mol/l). Ang II induced calcium increase could be effectively inhibited by ZD7155 which indicates a role of AT1AR in mediation of cyto-solic calcium changes. The conclusion is supported by the fact that the cal-cium response to cumulative increasing Ang II concentrations was also sig-nificantly diminished by ZD7155.
Changes in cytosolic calcium transients do not explain completely why significant constriction occurs during combined application of Ado (10-8
mol/l) and Ang II (10-12 mol/l). GPCR can activate the Rho/Rho kinase pathway leading to increased calcium sensitivity of the contractile machinery [15-19]. A recent study suggests a specific interaction between Ado and Ang II via increased p38 MAPK and MLC20 phosphorylation by Ado and subsequent enhancement of calcium sensitivity in Af [47].
Study IV Protocols For testing desensitization, 5 applications of 10-8 mol/l Ang II were repeat-edly applied from bath side in Af for 2 min, 10 min washout between appli-cations. For testing resensitization, Ado 10-5 mol/l was applied during the washout period (10 min) and then removed from the bath immediately be-fore the following Ang II application. The Ca2+-imaging experiments were

25
performed following the same protocols for testing the intracellular Ca2+-concentration in the wall of Af upon the Ang II applications.
Ang II induced contractions were tested using the nonspecific Ado recep-tors antagonist 8-SPT (5x10-5 mol/l), the inhibitor of Ado transporters NBTI (3x10-7 mol/l), the nonspecific P2 receptor antagonist suramin (10-4 mol/l) during Ado treatment, inhibitors of Rho-kinase (Y-27632, 10-6 mol/l), PKC (chelerythrine, 10-5 mol/l) and p38 MAPK (SB203580, 10-5 mol/l).
KCl induced constrictions were measured at very low Ca2+-concentrationsas produced by addition of EGTA to the bath solution (5x10-3 mol/), and after addition of CaCl2 (5 x 10-4 mol/l in the bath solution), for testing Ca2+-sensitivity of the contractile apparatus of Af. To investigate the Ca2+-sensitivity, Ado was added using the same protocol.
ResultsSuccessive applications of Ang II from bath side resulted in a clear desensi-tization of the Ang II induced constriction. Ado treatment restored the con-tractile Ang II response (Fig. 7). The Ca2+-imaging experiments showed increased intracellular Ca2+-concentration in Af upon the Ang II application. These Ca2+-transients diminished during subsequent Ang II applications. Ado treatment did not change Ca2+-transients significantly (Fig. 8).
Ca2+-sensitivity of the contractile apparatus: KCl (100 mmol/l) induced strong and transient constrictions of perfused Af, which did not desensitize. KCl induced constrictions were very weak at very low Ca2+-concentrationsproduced by addition of EGTA to the bath solution, and became stronger after addition of CaCl2. Ado aggravated the KCl induced constriction after treatment with EGTA and after addition of CaCl2 (Fig. 9A). Ado increased the Ca2+-sensitivity of Af significantly (Fig. 9B).
DiscussionStudy IV shows that Ado completely restores the response upon repeated applications of Ang II in Af. The restoration is caused by increasing the Ca2+-sensitivity of the contractile apparatus in the vascular smooth muscle. Restored Ang II response of the arterioles may be important in the control of arteriolar tone and consequently for the control of renal perfusion and filtra-tion during ischemic and postischemic periods. This mechanism may poten-tially play a role for the TGF.
Successive applications of Ang II resulted in a decrease of the contractile response and a concomitant attenuation of the cytosolic Ca2+-transients. The desensitization is most likely attributable to the phosphorylation and endocy-tosis of AT1AR with the consequence of a reduction of Ca2+-mobilization[69]. Although treatment with Ado between the successive applications of

26
Ang II completely restored the contractile response, intracellular Ca2+-transients were not re-established.
Ado acts on A1AR and A2AR, which mediate constriction and dilation in Af, respectively. The nonspecific Ado receptor antagonist 8-SPT did not support the idea of a receptor-mediated increase of the Ca2+-sensitivity and concomitant re-sensitization of the response to Ang II. Although it was re-cently shown that 8-SPT completely inhibits the action of Ado on A1AR and A2AR in Af [17]. However, the Ado transporter NBTI inhibited the resensi-tion in vascular smooth muscle cells. This strongly suggests that Ado need to cross the cell membrane to be effective in regard to the restoration of the contraction. Ado is produced intracellularly by sequential dephosphorylation of ATP to Ado. The increase in the Ca2+-sensitivity of vascular smooth mus-cle cells may be important in several pathophysiological and physiological situations.

27
FIGURES AND LEGENDS
Fig. 1:
Effects of Ang II bolus application on luminal diameter with and without pretreat-ment with L-NAME. This figure illustrates the significant difference on the time courses of contractions. (Brunner test, p < 0.05).
28
Fig. 2:
Effect of successive applications of Ang II with and without pretreatment with Ang II receptor antagonists on the change in the DAF-FM fluorescence. Ang II action on NO fluorescence shows desensitization with significantly lower NO fluo-rescence for the 2nd and 3rd applications. The desensitization effect is masked when using the AT1AR antagonist ZD7155 because the Ang II induced NO release is completely inhibited. “Effect of exchange” indicates the increase in fluorescence due to the procedure of exchanging solution in the perfusion pipette. This value repre-sents a methodically caused basal value of NO fluorescence. The “*” indicates sig-nificant differences in fluorescence intensity (p < 0.05).

29
Fig. 3:
Cumulative concentration-response-curve for Ado in C57Bl6 mice. The absolute and the percentage changes of the diameter are given (mean SEM). The “*” indi-cates a significant concentration-response.

30
Fig. 4:
Concentration-response-curves for Ado in A1AR knockout A1(-/-) and wild types A1(+/+) mice. The absolute and the percentage changes of the diameter are given (mean SEM). The “§” indicates a significant points compared to the control. The “*” indicates a significant concentration-response, and the “#” indicates differences in the concentration-response-curves between A1(-/-) and A1(+/+).

31
Fig. 5:
Effect of Ang II on the Ado concentration response. * indicates significant differ-ences for Ado + Ang II 10-8 mol/l (n=6) compared to Ado without Ang II treatment (n=13), # indicates significant differences compared to Ado + Ang II 10-12 (n=6), and § to Ado + Ang II 10-10 mol/l (n=8).

32
Fig. 6:
Changes in cytosolic calcium during application of increasing concentrations of Ang II in Af. The selective AT1AR antagonist ZD7155 clearly inhibits the effect of Ang II on the calcium concentration. * indicates significant differences compared to Ang II 10-11 mol/l, # indicates significant differences compared to Ang II without ZD7155.

33
Fig. 7.
Desensitization of Ang II induced constrictions (relative luminal diameter changes) in Af (black bars). Ado treatment in between the successive applications restored the response almost completely (grey bars). (*) indicates significant differences com-pared to the 1st application, (#) indicates significant differences between Ado treated and untreated arterioles.

34
Fig. 8:
Relative changes in the intracellular Ca2+-concentration ([Ca2+]i) in the afferent arte-riolar wall. [Ca2+]i decrease upon successive Ang II applications (black bars). There was no effect of Ado treatment in between successive Ang II applications on [Ca2+]itransients except for the 5th application (grey bars). (*) indicates significant differ-ences compared to the 1st Ang II application, (#) indicates significant differences between Ado treated and untreated arterioles.

35
Fig. 9:
Fig. 9A: KCl (100 mmol/l) induced constrictions of Af presented as relative changes of the luminal diameter over time. Measurements were performed in the presence of EGTA (5x10-3 mol/l in bath), which presumably reduced extracellular Ca2+-concentration ([Ca2+]e) to a very low level (indicated by “EGTA” in 9B), and after increase of [Ca2+]e by addition of CaCl2 (5x10-4 mol/l) to the bath solution (indicated by “EGTA+CaCl2”). Ado treatment increased the responses significantly, both with and without CaCl2. B: Relative changes of the luminal diameter for at 24 s after KCl administration (data from Fig. 9A) demonstrate higher Ca2+-sensitivity of Ado treated arterioles. (*) indicates differences between Ado treated and untreated groups.

36
CONCLUSIONS
Study I Study I demonstrates the Ang II induced activation of the NO system in Af. This mechanism can prevent the strong vasoconstrictor influence of Ang II, which occurs by high renin and angiotensin converting enzyme activity in the JGA. AT1AR play a prominent role in mediating the Ang II induced NO release.
Study II Study II demonstrates the expression of A1AR, A2AAR and A2BAR mRNA in Af and their physiological role in the control of afferent arteriolar tone. The response of Af upon Ado at physiological concentrations results from the action of vasoconstrictor A1AR and vasodilator A2AR, whose balance is changed in favor of A1AR at lower, and in favor of A2AR at higher, Ado concentrations. This behavior confirms the studies at the organ level. Ado given at low concentrations potentiated the Ang II-induced constriction, which may be an important mechanism in the mediation of TGF by Ado.
Study III Study III demonstrates an enhancing effect of Ang II in low non constrictor concentrations on the contractile response to Ado in Af. The stronger con-traction may not only be due to an increase in cytosolic calcium. Data sug-gest the contribution of calcium independent mechanisms in the more than additive vasoconstrictory action of Ado and Ang II. The concerted action of Ang II and Ado on Af is an important determinant for the TGF in the kidney.
Study IV Study IV demonstrates that exogenously applied Ado restores Ang II in-duced contractions, which are subjected to desensitization during repeated applications of Ang II. Ado increases calcium sensitivity of the Af, along

37
with augmented phosphorylation of the regulatory unit of the myosin light chain. Ado exerts its effect via intracellular action. Enzymes p38 MAPK, RhoA/Rho-kinase and PKC pathway are involved in the increase of MLC20 phosphorylation. This mechanism may contribute to the mediation of the TGF by ado in the JGA of the kidney.

38
PAPER LIST OF PHD STUDY
Cover Page Stories 1. Andreas Patzak, En Yin Lai, Ralf Mrowka, Andreas Steege, Pontus B. Persson, A. Erik G. Persson AT1 receptors mediate angiotensin II-induced release of nitric oxide in afferent arterioles Kidney International 2004 Nov;66(5):1949-58.
2. En Yin Lai, Peter Martinka, Michael Fähling, Ralf Mrowka, Andreas Steege, Adrian Gericke, Mauricio Sendeski, Pontus B. Persson, A. Erik G. Persson, Andreas Patzak Adenosine restores angiotensin II-induced contrac-tions by receptor-independent enhancement of calcium sensitivity in renal arterioles Circulation Research 2006 99: 1117 - 1124; Epub 2006 Oct 12.

39
Journal Publications 1. A. Patzak, F. Kleinmann, E.Y. Lai, E. Kupsch, A. Skelweit, R. Mrowka Nitric oxide counteracts angiotensin II induced contraction in efferent arte-rioles in mice Acta Physiol Scand 2004 Aug; 181(4):439-44. 2. Andreas Patzak, En Y Lai, Ralf Mrowka, Andreas Steege, Pontus B. Persson, A. Erik G. Persson AT1 receptors mediate angiotensin II-induced release of nitric oxide in afferent arterioles Kidney International 2004 Nov;66(5):1949-58. (Paper I)3. Patzak A, Lai E, Persson PB, Persson AE. Angiotensin II-nitric oxide interaction in glomerular arterioles Clin Exp Pharmacol Physiol 2005 May-Jun;32(5-6):410-4. Review. 4. Andreas Patzak, Julia Bontscho, EnYin Lai, Eckehardt Kupsch, Angela Skalweit, Claus-Michael Richter, Mathias Zimmermann, Christa Thöne-Reineke, Olaf Joehren, Michael Godes, Andreas Steege and Berthold Hocher Angiotensin II sensitivity of afferent glomerular arterioles in endothelin-1 transgenic mice Nephrol Dial Transplant 2005 Dec;20(12):2681-9. Epub 2005 Sep 27. 5. En Yin Lai*, Andreas Patzak*, Andreas Steege, Ralf Mrowka, Russell Brown, Nadine Spielmann, Pontus B. Persson, Bertil B. Fredholm, A. Erik G. Persson (*equal contribution) Contribution of adenosine receptors in the control of arteriolar tone and adenosine-angiotensin II interaction Kidney International 2006 Aug;70(4):690-8. Epub 2006 Jun 28. (Paper II)6. En Yin Lai, Peter Martinka, Michael Fähling, Ralf Mrowka, Andreas Steege, Adrian Gericke, Mauricio Sendeski, Pontus B. Persson, A. Erik G. Persson, Andreas Patzak Adenosine restores angiotensin II-induced contrac-tions by receptor-independent enhancement of calcium sensitivity in renal arterioles Circulation Research 2006 99: 1117 - 1124; Epub 2006 Oct 12. (Paper IV)7. En Yin Lai, Leif Jansson, Andreas Patzak, A. Erik G. Persson Vascular reactivity in arterioles from normal and alloxan-diabetic mice: studies on single perfused islets Diabetes 2007 Jan;56(1):107-12. 8. En Yin Lai*, A Erik G Persson, Birgitta Bodin, Örjan Källskog, Arne Andersson, Ulrika Pettersson, Peter Hansell, Leif Jansson (*Correspondence) Endothelin-1 and pancreatic islet vasculature: studies in vivo and on isolated, vascularly perfused pancreatic islets Am J Physiol Endocrinol Metab. 2007 Feb 6; [Epub ahead of print]9. Andreas Patzak*, En Yin Lai*, Pontus B. Persson, A. Erik G. Persson (*equal contribution) Long term effect of adenosine on the contractile re-sponse to angiotensin II in afferent arterioles Submitted for publication 10. Peter Martinka, En Yin Lai, Michael Fähling, Sebastian Schmidt, An-dreas Steege, Adrian Gericke, Vera Jankowski, Joachim Jankowski, Rudolf Schubert, Matthias Gaestel, A. Erik G. Persson, Pontus B. Persson, Andreas Patzak Adenosine increases calcium sensitivity via receptor-independent

40
activation of p38 MAPK/MK2 pathway in isolated mesenteric arteries Sub-mitted for publication 11. En Yin Lai, Andreas Patzak, A. Erik G. Persson Low dose angiotensin II enhances cytosolic calcium concentration and contractile responses of affer-ent arterioles to adenosine Manuscript (Paper III)12. Lina Nordquist*, En Yin Lai*, Mats Sjöquist, Andreas Patzak, Leif Jansson, A Erik G Persson (*equal contribution) Proinsulin C-peptide con-stricts glomerular afferent arterioles in diabetic mice: A potential reno-protective mechanism ofaction Manuscript.

41
ACKNOWLEDGEMENTS
This study was carried out at the Department of Medical Cell Biology, Divi-sion of Integrative Physiology, Faculty of Medicine, Uppsala University, Sweden. It was also financially supported by the Swedish Medical Research Council (Project number: K99-14X-03522-28D, K2002-14X-03522-31D, K2003-04X-03522-32A and K2006-04X-03522-35-3), the Swedish Heart and Lung Fundation (Project number: 2004-0645 and 2006-0329), the Wallenberg Foundation, the Ingabritt and Arne Lundberg Foundation and the Wenner-Gren-Foundation.
This thesis is the result of the collaborations with a number of people. I would like to express my sincere gratitude to all those people whom have helped me in one way or the other. Especially thanks to:
Professor A. Erik G. Persson, my excellent supervisor, thanks for intro-ducing me to the very interesting field of renal physiology, for creating an excellent research lab and for your great support and encouragement all the time. Thanks for making me aim higher with your “not only a PhD but a scientist” approach. Thanks for nice dinners and Christmas gifts every year.
Professor Peter Hansell and Associate Professor Ruisheng Liu, my co-supervisors, for introducing me to microperfusion, good discussions and always kind help.
Associate Professor Andreas Patzak, Humboldt University, for teaching me physiology and scientific research, for sharing your excellent technique and broad knowledge, for your quick and never-ending help, for your excel-lent and friendly discussions via hundreds of e-mails and phone-calls during the last 5 years, for our nice friendship and your very kind family and friends in Berlin, for all of our nice trips in China, Europe and USA, for many nice dinners together in the world.
Professor Leif Jansson, for guiding me in the broad field of diabetes, for our islet arteriole perfusion and diabetes paper. It is fantastic to work with you, you are a true gentleman.
Professor Örjan Källslog, for teaching me physiology, especially during the first year. You are so kind and enthusiasm.
Professor Lena Holm, for your kind help, nice talks and dinners. Professor Mats Sjöquist, for nice talks. Professor Carl-Henrik Heldin and Associate Professor Paraskevi
Heldin, Ludwig institute, Uppsala University, for nice dinners and friend-ship.

42
Professor Pontus B. Persson, and All friends at Humboldt University, It was great time during my many visits in the Berlin lab.
Professor Bertil B. Fredholm, Karolinska Institute, for supplying A1knockout mice.
Professor Godfried M. Roomans, Professor Arne Andersson, Professor Stellan Sandler, Agneta Bäfve, Karin Öberg, Marianne Ljungkvist andKärstin Flink, for your nice administrative help.
Dr. Carina Carlsson, Dr. Michael Broman, and Dr. P-O Carlsson fornice talks.
All co-authors, for our friendly, productive, perfect team. All colleagues of our B1 corridor, Louise Rügheimer, Lina Nordquist,
Angelica Fasching, Mattias Carlström, Johan Sällström, Fredrik Palm, Magnus Johansson, Mia Phillipson, Michael Hultström, Johanna Hen-riksnäs, Russell Brown, Anna Ollerstam, Olof Schreiber, Johan Olerud, Ulrika Pettersson, Nina, Andrei, Joel, Joey, Malou, Jenny, Sara, Åsa, Gunno Nilsson, Göran Ståhl, Britta Isaksson and Annika Jägare, for our “Pig Lab”, teaching team and great research team, for your kind help, for our “café time”, “cake time”, “lunch time” and “presentation + cake time”, for having a nice 5 years period and being good friends.
Thanks to all my Chinese friends in Uppsala for having very good time and great friendship, especially: Hongmei and Yu Jiang, Zhang Jing and Shouting, Zhao Ying and Ruisheng, Li Jingping, Zhang Xiaoqun, Hu Guozhen, Zhou Xiaofu, Zhao Xiaomin, Xu Ning, Ma Ruixin, Zhou Chuanzheng, Ma Zufu, Lei Pengxiang, Xue Bo and Liu Aiqin, Li Yue-juan, Li Zhanchun and Zhao Lijun, Jianman and Su Chao, Lu Lingge and Yuming, Cheng Tingzu and Tan Huirong.
All my Chinese friends in China, for never-ending friendship.Muyun, Weiwei, et al. for all of our next excellent generation, I am
proud of you.My parents, parents-in-law, sisters and brothers, for your love and en-
couragement. Siyun, my son, for being my excellent teacher and friend, for your great
achievements.Lingli, my lovely wife, for your deepest love and great support all the
time.

43
REFERENCES
1. Guyton AC, Langston JB, Navar G: Theory For Renal Autoregula-tion By Feedback At The Juxtaglomerular Apparatus. Circ Res15:SUPPL:187-197, 1964
2. Thurau KW: Autoregulation Of Renal Blood Flow And Glomerular Filtration Rate, Including Data On Tubular And Peritubular Capillary Pres-sures And Vessel Wall Tension. Circ Res 15:SUPPL:132-141, 1964
3. Ichikawi I, Harris RC: Angiotensin actions in the kidney: renewed insight into the old hormone. Kidney Int 40:583-596, 1991
4. Rosivall L, Rinder DF, Champion J, et al.: Intrarenal angiotensin I conversion at normal and reduced renal blood flow in the dog. Am J Physiol245:F408-415, 1983
5. Siragy HM, Carey RM: The subtype 2 (AT2) angiotensin receptor mediates renal production of nitric oxide in conscious rats. J Clin Invest100:264-269, 1997
6. Ito S, Johnson CS, Carretero OA: Modulation of angiotensin II-induced vasoconstriction by endothelium-derived relaxing factor in the iso-lated microperfused rabbit afferent arteriole. J Clin Invest 87:1656-1663, 1991
7. Patzak A, Mrowka R, Storch E, et al.: Interaction of angiotensin II and nitric oxide in isolated perfused afferent arterioles of mice. J Am Soc Nephrol 12:1122-1127, 2001
8. Liu R, Persson AE: Angiotensin II stimulates calcium and nitric ox-ide release from Macula densa cells through AT1 receptors. Hypertension43:649-653, 2004
9. Majid DS, Navar LG: Nitric oxide in the control of renal hemody-namics and excretory function. Am J Hypertens 14:74S-82S, 2001
10. Bachmann S, Bosse HM, Mundel P: Topography of nitric oxide synthesis by localizing constitutive NO synthases in mammalian kidney. AmJ Physiol 268:F885-898, 1995

44
11. Ollerstam A, Pittner J, Persson AE, et al.: Increased blood pressure in rats after long-term inhibition of the neuronal isoform of nitric oxide syn-thase. J Clin Invest 99:2212-2218, 1997
12. Baumann JE, Persson PB, Ehmke H, et al.: Role of endothelium-derived relaxing factor in renal autoregulation in conscious dogs. Am J Physiol 263:F208-213, 1992
13. Sigmon DH, Beierwaltes WH: Influence of nitric oxide derived from neuronal nitric oxide synthase on glomerular filtration. Gen Pharmacol34:95-100, 2000
14. Manning RD, Jr., Hu L, Mizelle HL, et al.: Role of nitric oxide in long-term angiotensin II-induced renal vasoconstriction. Hypertension21:949-955, 1993
15. Hall JE, Granger JP, Hester RL: Interactions between adenosine and angiotensin II in controlling glomerular filtration. Am J Physiol 248:F340-346, 1985
16. Hansen PB, Hashimoto S, Briggs J, et al.: Attenuated renovascular constrictor responses to angiotensin II in adenosine 1 receptor knockout mice. Am J Physiol Regul Integr Comp Physiol 285:R44-49, 2003
17. Lai EY, Patzak A, Steege A, et al.: Contribution of adenosine recep-tors in the control of arteriolar tone and adenosine-angiotensin II interaction. Kidney Int 70:690-698, 2006
18. Osswald H, Schmitz HJ, Heidenreich O: Adenosine response of the rat kidney after saline loading, sodium restriction and hemorrhagia. PflugersArch 357:323-333, 1975
19. Weihprecht H, Lorenz JN, Briggs JP, et al.: Synergistic effects of angiotensin and adenosine in the renal microvasculature. Am J Physiol266:F227-239, 1994
20. Osswald H, Muhlbauer B, Schenk F: Adenosine mediates tubu-loglomerular feedback response: an element of metabolic control of kidney function. Kidney Int Suppl 32:S128-131, 1991
21. Brown R, Ollerstam A, Johansson B, et al.: Abolished tubuloglome-rular feedback and increased plasma renin in adenosine A1 receptor-deficient mice. Am J Physiol Regul Integr Comp Physiol 281:R1362-1367, 2001
22. Sun D, Samuelson LC, Yang T, et al.: Mediation of tubuloglomeru-lar feedback by adenosine: evidence from mice lacking adenosine 1 recep-tors. Proc Natl Acad Sci U S A 98:9983-9988, 2001

45
23. Schnermann J: Micropuncture analysis of tubuloglomerular feed-back regulation in transgenic mice. J Am Soc Nephrol 10:2614-2619, 1999
24. Guyton AC, Hall JE: Guyton and Hall. Textbook of Medical Physi-ology Tenth edition:279-294, 2000
25. Guyton AC, Hall JE: Guyton and Hall. Textbook of Medical Physi-ology Tenth edition:295-312, 2000
26. Ruyter J: Uber einen merkwurdigen Abschnitt der vasa afferentia der Mäuseniere. Z. Zellforsch 2:242-248, 1925
27. Goormaghtigh N: Les segments neuro-myo-artériels, juxtaglomerulaires du renin. Arch. Biol 43:575-591, 1932
28. Zimmerman KW: Uber den Bau des Glomerulus der Säugerniere. Ztschr. f. mikr. anat. Forsh. 32:176-278, 1933
29. Schnermann J, Persson AE, Agerup B: Tubuloglomerular feedback. Nonlinear relation between glomerular hydrostatic pressure and loop of henle perfusion rate. J Clin Invest 52:862-869, 1973
30. Feoktistov I, Biaggioni I: Adenosine A2B receptors. Pharmacol Rev49:381-402, 1997
31. Fishman P, Bar-Yehuda S: Pharmacology and therapeutic applica-tions of A3 receptor subtype. Curr Top Med Chem 3:463-469, 2003
32. Fredholm BB, AP IJ, Jacobson KA, et al.: International Union of Pharmacology. XXV. Nomenclature and classification of adenosine recep-tors. Pharmacol Rev 53:527-552, 2001
33. Fredholm BB, Cunha RA, Svenningsson P: Pharmacology of adeno-sine A2A receptors and therapeutic applications. Curr Top Med Chem 3:413-426, 2003
34. Townsend-Nicholson A, Baker E, Schofield PR, et al.: Localization of the adenosine A1 receptor subtype gene (ADORA1) to chromosome 1q32.1. Genomics 26:423-425, 1995
35. Weber RG, Jones CR, Palacios JM, et al.: Autoradiographic visuali-zation of A1-adenosine receptors in brain and peripheral tissues of rat and guinea pig using 125I-HPIA. Neurosci Lett 87:215-220, 1988
36. Toya Y, Umemura S, Iwamoto T, et al.: Identification and charac-terization of adenosine A1 receptor-cAMP system in human glomeruli. Kid-ney Int 43:928-932, 1993

46
37. Tang Y, Zhou L: Characterization of adenosine A1 receptors in hu-man proximal tubule epithelial (HK-2) cells. Receptors Channels 9:67-75, 2003
38. Mahan LC, McVittie LD, Smyk-Randall EM, et al.: Cloning and expression of an A1 adenosine receptor from rat brain. Mol Pharmacol 40:1-7, 1991
39. Franco M, Bell PD, Navar LG: Effect of adenosine A1 analogue on tubuloglomerular feedback mechanism. Am J Physiol 257:F231-236, 1989
40. Pflueger AC, Osswald H, Knox FG: Adenosine-induced renal vaso-constriction in diabetes mellitus rats: role of nitric oxide. Am J Physiol276:F340-346, 1999
41. Schnermann J: Effect of adenosine analogues on tubuloglomerular feedback responses. Am J Physiol 255:F33-42, 1988
42. Joyner WL, Mohama RE, Myers TO, et al.: The selective response to adenosine of renal microvessels from hamster explants. Microvasc Res35:122-131, 1988
43. Weihprecht H, Lorenz JN, Briggs JP, et al.: Vasomotor effects of purinergic agonists in isolated rabbit afferent arterioles. Am J Physiol263:F1026-1033, 1992
44. Townsend-Nicholson A, Baker E, Sutherland GR, et al.: Localiza-tion of the adenosine A2b receptor subtype gene (ADORA2B) to chromo-some 17p11.2-p12 by FISH and PCR screening of somatic cell hybrids. Ge-nomics 25:605-607, 1995
45. Osswald H: Renal effects of adenosine and their inhibition by theo-phylline in dogs. Naunyn Schmiedebergs Arch Pharmacol 288:79-86, 1975
46. Premen AJ, Hall JE, Mizelle HL, et al.: Maintenance of renal auto-regulation during infusion of aminophylline or adenosine. Am J Physiol248:F366-373, 1985
47. Johansson B, Halldner L, Dunwiddie TV, et al.: Hyperalgesia, anxi-ety, and decreased hypoxic neuroprotection in mice lacking the adenosine A1 receptor. Proc Natl Acad Sci U S A 98:9407-9412, 2001
48. Lai EY, Martinka P, Fahling M, et al.: Adenosine restores angio-tensin II-induced contractions by receptor-independent enhancement of cal-cium sensitivity in renal arterioles. Circ Res 99:1117-1124, 2006
49. Patzak A, Lai EY, Mrowka R, et al.: AT1 receptors mediate angio-tensin II-induced release of nitric oxide in afferent arterioles. Kidney Int66:1949-1958, 2004

47
50. Kojima H, Nakatsubo N, Kikuchi K, et al.: Detection and imaging of nitric oxide with novel fluorescent indicators: diaminofluoresceins. Anal Chem 70:2446-2453, 1998
51. Kojima H, Urano Y, Kikuchi K, et al.: Fluorescent Indicators for Imaging Nitric Oxide Production. Angew Chem Int Ed Engl 38:3209-3212, 1999
52. Grynkiewicz G, Poenie M, Tsien RY: A new genertaion of Ca2+ indicators with greatly improved fluorescence properties. J Biol Chem260:3440-3450, 1985
53. Boulanger CM, Caputo L, Levy BI: Endothelial AT1-mediated re-lease of nitric oxide decreases angiotensin II contractions in rat carotid artery. Hypertension 26:752-757, 1995
54. Pueyo ME, Arnal JF, Rami J, et al.: Angiotensin II stimulates the production of NO and peroxynitrite in endothelial cells. Am J Physiol274:C214-220, 1998
55. Saito S, Hirata Y, Emori T, et al.: Angiotensin II activates endothe-lial constitutive nitric oxide synthase via AT1 receptors. Hypertens Res19:201-206, 1996
56. Thorup C, Kornfeld M, Goligorsky MS, et al.: AT1 receptor inhibi-tion blunts angiotensin II-stimulated nitric oxide release in renal arteries. JAm Soc Nephrol 10 Suppl 11:S220-224, 1999
57. Abadir PM, Carey RM, Siragy HM: Angiotensin AT2 receptors di-rectly stimulate renal nitric oxide in bradykinin B2-receptor-null mice. Hy-pertension 42:600-604, 2003
58. Siragy HM: AT(1) and AT(2) receptors in the kidney: role in dis-ease and treatment. Am J Kidney Dis 36:S4-9, 2000
59. Duke LM, Eppel GA, Widdop RE, et al.: Disparate roles of AT2 re-ceptors in the renal cortical and medullary circulations of anesthetized rab-bits. Hypertension 42:200-205, 2003
60. Bohm SK, Khitin LM, Grady EF, et al.: Mechanisms of desensitiza-tion and resensitization of proteinase-activated receptor-2. J Biol Chem271:22003-22016, 1996
61. Casey PJ: Protein lipidation in cell signaling. Science 268:221-225, 1995
62. Jin X, Shepherd RK, Duling BR, et al.: Inosine binds to A3 adeno-sine receptors and stimulates mast cell degranulation. J Clin Invest 100:2849-2857, 1997

48
63. Fredholm BB, Abbracchio MP, Burnstock G, et al.: Nomenclature and classification of purinoceptors. Pharmacol Rev 46:143-156, 1994
64. Osswald H, Vallon V, Muhlbauer B: Role of adenosine in tubu-loglomerular feedback and acute renal failure. J Auton Pharmacol 16:377-380, 1996
65. Schnermann J: Juxtaglomerular cell complex in the regulation of re-nal salt excretion. Am J Physiol 274:R263-279, 1998
66. Schnermann J, Levine DZ: Paracrine factors in tubuloglomerular feedback: adenosine, ATP, and nitric oxide. Annu Rev Physiol 65:501-529, 2003
67. Traynor T, Yang T, Huang YG, et al.: Inhibition of adenosine-1 re-ceptor-mediated preglomerular vasoconstriction in AT1A receptor-deficient mice. Am J Physiol 275:F922-927, 1998
68. Traynor T, Yang T, Huang YG, et al.: Tubuloglomerular feedback in ACE-deficient mice. Am J Physiol 276:F751-757, 1999
69. Freedman NJ, Lefkowitz RJ: Desensitization of G protein-coupled receptors. Recent Prog Horm Res 51:319-351; discussion 352-313, 1996


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 237
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Medicine”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-7702
ACTAUNIVERSITATISUPSALIENSISUPPSALA2007