innate immunity importance? - front line of defense - required to instruct adaptive responses why is...
TRANSCRIPT

Innate Immunity
Importance?
- front line of defense- required to instruct adaptive responses
Why is it interesting?- the recognition problem- host-pathogen arms race

Innate immunity:
Cells – macrophages, neutrophils, dendritic cells, mast cells, NK cells, epithelial cells, etc.
Immediate destruction of pathogens.
Mechanisms (e.g.’s):- low pH (gut)- defensins- complement fixation- phagocytosis, followed by killing mediated by:
- antimicrobial peptides, defensins- proteases- ROS, RNS

Questions:
1) What does the innate immune system actually recognize?2) How are subsequent responses coordinated and controlled?

PAMPs ‘n DAMPs How does the innate immune system recognize pathogens?
“Approaching the Asymptote? Evolution and Revolution in Immunology” CA Janeway,Cold Spring Harb. Symp Quant Biol 1989
“The Immunologist’s Dirty little secret”
“Pattern Recognition Receptors (PRR).”
- Distinguish “infectious non-self from non-infectious self”- Non-clonally distributed, germline encoded- recognize conserved microbial products, required for microbe function, not found in host (“Pathogen-associated molecular patterns” , or PAMPs) - evolutionarily ancient recognition system

Innate immunity instructs adaptive responses
Innate and adaptive responses are intertwined.(e.g. IFN-gamma activates macrophage killing function)
microbe
TCRMHCII
B7 CD28
PatternRecognitionreceptors
APCs
cytokines

Toll-like Receptors
1989: Janeway proposes PRRs1996: Toll shown to be involved in innate immunity in Drosophila1997: Janeway lab identifies human homologue of Toll (TLR4) 1998 : Beutler lab demonstrates that missense mutation in TLR4 underlies insensitivity to LPS in C3HeJ mouse

TLRs and some of their PAMP ligands:
TLR2/1: diacyl lipopeptidesTLR2/6: triacyl lipopeptidesTLR3: dsRNATLR4: LPSTLR5: flagelllinTLR7,8: ssDNATLR9: unmethylated CpG DNA

(Gay and Gangloff,Ann Rev Biochem 2007)
, ssDNA
TLRs and theirligands
TLR2
TLR4

Outcomes: inflammatory cytokine production, upregulation of APC costimulatory
molecules
TLR signalling
Kawai and Akira, Semin Immunol 19:24-32 (2007)
“(toll-like OR tlr*) AND review AND (signalling OR signaling)”->1216 refs.1462
(toll-like[ti] OR tlr*[ti]) AND review and (signaling OR signalling)-> 580 refs

An Alternative View: The Danger Hypothesis(Polly Matzinger)
Not so much non-self as “Danger” that is recognized. Danger signals can be derived from infection, or from damaged/altered self.

Stranger or Danger?

Is the “Danger” model tautological?
Recognition of host damage may have arisen in order to see:- damage as a consequence of infection- damage as something that will lead to infection- damage as something that requires similar
responses as infection (e.g. leukocyte recruitment)

CLEC9A – a C-type lectin
Danger receptor?
Mincle, another C-type lectin, has also been implicated in responses to necrotic cells (as well as to pathogenic fungi)
(Yamasaki et al, Nat. Immunol 2008)
(Nature 2009)

Toll-like receptors are not the only innate PRRs:
(Ann Rev Immunol 2005)

“Complexity in action”
The challenge – to make sense of it all
(Underhilland Ozinsky,Ann Rev Imm2002)

One source of complexity: cooperation between differentreceptors.
Receptor cooperation:
1) direct
e.g.s?

TLRs use coreceptors
TLR4/MD-2 uses CD14 for recognition of LPS. (Also, to a lesser extent CD14 for TLR2). Necessary for TRIF-dependent signaling in response to rough LPS.
TLR2/6 and TLR2/1 heterodimerize for recognition of different ligands.
TLR2 uses CD36 for recognition of Gm+ve bacteria

(Triantafilou, M et al, J Biol Chem 2006)
Ligand-induced and constitutive association of TLR2 with coreceptors
FRET
TLR2 Receptor X
FRET e
ffici
en
cySpatial organization of innate immune recognition?

2) intersecting signaling cascades
Pi

3) Indirect or sequential interactions:

Responses due to engaging multiple receptors can be
- synergistic- antagonistic- emergent (1+2=C)
Three examples of interacting signaling pathways:
1) FcR/TLR: Type II macrophage activation2) DC-SIGN/TLR: inhibition of TLR responses3) TLR/TLR : synergistic IL-12 production

+LPS
FcR cross-linking + LPS -> “type II activated macrophage”IL-10 and IL-12 responses to LPS reversed -> Th2, not Th1 bias
(Anderson and Mosser, J. Leuk.Biol. 2002)
1)

(J Immunol. 2001)
Injecting “Type II” activated macrophages intomice protected them from otherwise lethal dose of endotoxin.

Mechanism?
PI3K
Nuclear Ikb-alpha -> inhibition of IL-12 production
TLR FcR
NFkB IkB
ERK
chromatin remodelling at IL-10 locus

C-type Lectins:
-involved in recognizing both endogenous and exogenousglycosylated ligands
TLR4 TLR2
McGreal etal, Curr OpImmunol 2005
-glucansICAM-3HIV, M. Tb
(“Dendritic cell-specific ICAM-grabbing non-integrin”)
2)

Mycobacterial ManLAM binding to DC-SIGN suppresses LPS-induced DC maturation
J. Exp. Med, 2003

TLR DC-SIGN
NFkB
Raf
Raf-mediated acetylation of NFKB and increased IL-10 production. (Gringhuis et al, Immunity 2007)

3) TLRs synergize for IL-12 production
Napolitani, G. et al Nat Imm 2005
Trinchieri and Sher, Nat Rev Imm 2007
IL-12, IL-23synergistically up-regulated

Teleology of these interactions?
Why [telologically] the synergy in IL-12 production?
Napolitani et al proposed a “combinatorial safety code” (a “two-key” model for pathogen detection and IL-12 production)
Why does FcR ligation -> increased IL10, decreased IL12?
Why does DC-SIGN/TLR coengagement -> increased IL-10?

Do PRRs “code” for targeted pathogen responses?
Coding or redundancy?
(Underhill and Ozinsky, Curr Op Immunol 2002)
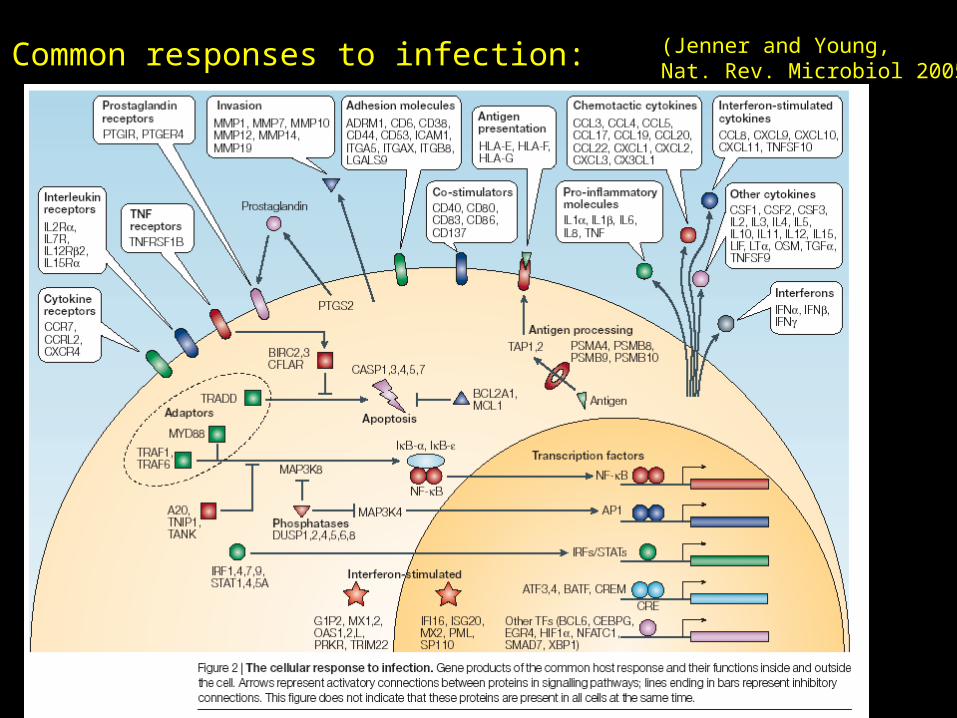
Common responses to infection: (Jenner and Young,Nat. Rev. Microbiol 2005)

RAW 264.7 cells: - mouse “macrophage-like” cell line - cell of choice for Alliance for Cellular Signaling large-scale analysis
The Question : How complex is the network of signaling pathways in a macrophage?
The approach:
- 22 ligands (TLR agonists, cytokines, and more)-What happens when you add them individually? -What happens when you add them in every possible pairwise combination?
(231 combos [=(222-22)/2])
(Nat Cell Biol 2006)


One at a time, now….
The single ligand screen:

The punchline:
(now, red/blue meansZ values, i.e. non-additivity)
(231*147= 33,957 data points)

cluster: TLRs + ISO or PGE
GCSF, IL10, IL6, RANTES
MIP1alpha, TNFalpha
Unbiased clustering of non-additivity:

“A simplified picture of the signalling network”

(around 40 significant clusters)
How many “interaction agents” might there be?
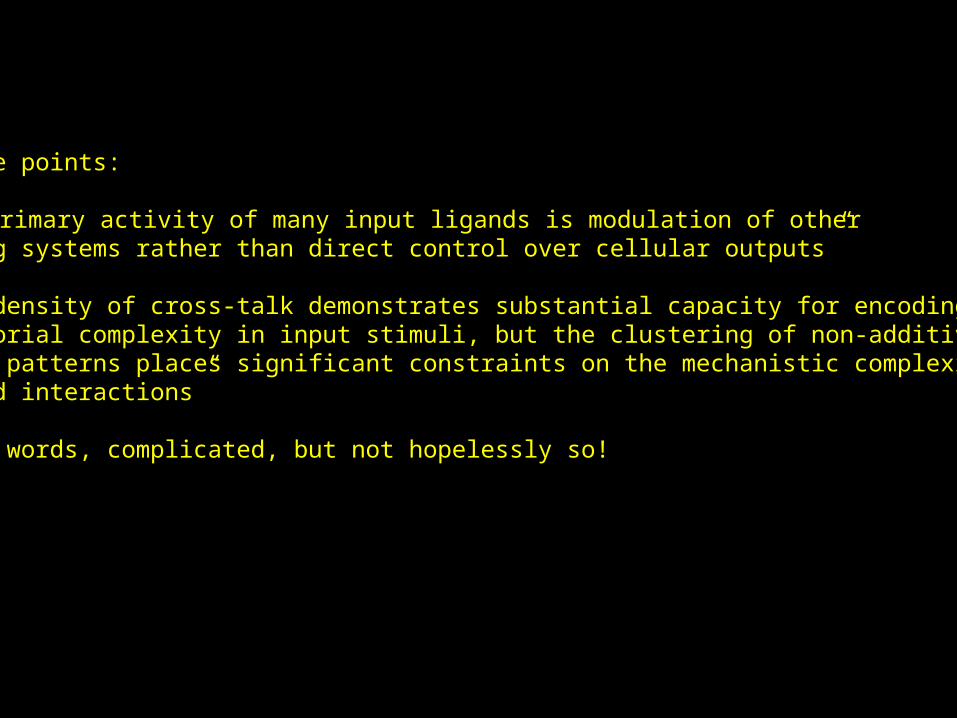
Take home points:
1) “the primary activity of many input ligands is modulation of other signaling systems rather than direct control over cellular outputs”
2) “The density of cross-talk demonstrates substantial capacity for encodingcombinatorial complexity in input stimuli, but the clustering of non-additiveresponse patterns places significant constraints on the mechanistic complexityof ligand interactions”
In other words, complicated, but not hopelessly so!

signaling pathway network
Nat Immunol 10, 327 (April 2009)

Nat Immunol 10, 327 (2009)
(Genome wide RNAi screen in Drosophila)
Cell as single “mega-network?”

Unbiased, global approaches:
Global readouts?
Global interventions?

Different possibilities for cooperation in vivo
(Trinchieri, Nat Rev Imm 2007)

Using ENU mutagenesis, followed by positional cloning.
At around 60,000 F1 mice screened as of several years ago.
Bruce Beutler lab’s ongoing forward genetic screen:
Multiple phenotypic screens, including:
ex vivo: TLR signaling screen ((peritoneal mphi) dsDNA screen
NALP3 Inflammasome screencontrol of virus infection (MCMV,
adenovirus, flu, Rift Valley Fever)in vivo: susceptibility to MCMV and LCMV
DSS-induced colitis

(Beutler,B. Immunogenetics 57:385 (2005)
Some hits :

“To date, 282 transmissible mutations that cause discernable phenotypes have been set aside for positional cloning in the Beutler laboratory; 180 mutations have been mapped to chromosomes, and in 152 instances, molecular identification of the causative mutation has been made. These mutations fall within 105 genes. 154 of the mutations studied affect immunity, and about half of the mutations affecting immunity that are cloned prove to be novel in the sense that no such phenotype had been predicted by knockout mutations, or knockouts had not been created. Only about 50% recessive saturation of the genome has been achieved to date in any given screen; therefore, it is expected that many key discoveries of function lie in waiting. “
(Beutler lab webpage: http://www.scripps.edu/genetics/beutler/research.html)

(Nat Imm 2006)
(Nature, 2003)
(Nature, 2005)
(PNAS 2006)
(Nat Imm 2005)
(Science 2008)
(Nature Genetics 2007)

PNAS 2008
PNAS 2009
PNAS 2009