inhibition of rat mammary gland chemical...
TRANSCRIPT

(CANCER RESEARCH 51. 481-486. January 15, 1991]
Inhibition of Rat Mammary Gland Chemical Carcinogenesis by DietaryDehydroepiandrosterone or a Fluorinated Analogue ofDehydroepiandrosterone1
Thomas A. Ratko,2 Carol J. Detrisac, Rajendra G. Mehta, Gary J. Kelloff, and Richard C. Moon3
Laboratory of Paihophysiology, Life Sciences Division, I1T Research Institute, Chicago, Illinois 60616 [T. A. R., R. G. M., C. J. D., and R. C. M.], and National CancerInstitute, Chemoprevention Branch, Bethesda, Maryland 20892 [G. J. K.J
ABSTRACT
The chemopreventive efficacy of p.o. administered dehydroepiandros-terone (DHEA), DHEA plus Ar-(4-hydroxyphenyl)retinamide (4-HPR),or 16a-fluoro-5-androsten-17-one (DHEA analogue 8354) was examinedin rats treated with A'-methyl-A'-nitrosourea (MNU; 50 mg/kg body-weight, i.v.) at 50 days of age. Semipurified diet (AIN-76A) containingeach steroid alone, or DHEA plus 4-HPR, was administered duringinitiation (—1week to +1 week post-MNU), promotion/progression (+1week post-MNU to termination), or both phases (—1week post-MNU
to termination) of the carcinogenic process. Neither DHEA nor DHEAanalogue 8354 (0.2%, w/w) significantly affected the initiation of mammary cancer when administered alone; however, DHEA (0.2%, w/w) plus4-HPR (1 mmol/kg diet) significantly reduced cancer multiplicity (26%)when given during initiation. All three treatments were strongly effectivewhen given during promotion/progression, significantly reducing mammary cancer multiplicity by 77% (DHEA), 84% (DHEA/4-HPR), and66% (DHEA analogue 8354), relative to carcinogen controls. Cancerincidence was significantly inhibited by DHEA (33% inhibition) andDHEA/4-HPR (24% reduction) during promotion/progression. However,the most effective chemopreventive treatment encompassed both phasesof carcinogenesis. Thus, under these conditions, DHEA (0.2% or 0.1%,w/w) reduced cancer incidence (52% and 32% reductions, respectively)and multiplicity (91% and 86% reductions, respectively). Further reduction in mammary cancer incidence was observed in animals that receivedDHEA (both doses) plus 4-HPR (1 and 0.5 mmol/kg diet, respectively).DHEA analogue 8354 (0.2% or 0.1%, w/w) given for the duration of thestudy reduced only cancer multiplicity (61% and 56% reductions, respectively). Tumor-related mortality was significantly lower in rats thatreceived long-term treatment with DHEA or DHEA/4-HPR, when compared with carcinogen controls. Except for a slight, but significant,postcarcinogen decrease in the mean body weights of rats treated concomitant 1\ with DHEA (plus or minus 4-HPR) and MNU, additional grossmanifestations of steroid-induced toxicity were not observed.
INTRODUCTION
DHEA4 is an abundantly secreted adrenal steroid that is an
intermediate in the biosynthesis of other hormones, includingtestosterone and estradici-170 (1). Although a physiologicalrole for DHEA has yet to be defined, a growing body ofevidence, both epidemiológica! and experimental, suggests astrong inverse relationship between alteration in the serum
Received 3/1/90; accepted 10/24/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by contract N01-CN-45912-14 and Grant CA-34664 (R.
G. M.) from the National Cancer Institute.2Present address: American Medical Association, 515 North State Street,
Chicago. IL 60610.3To whom requests for reprints should be addressed, at IIT Research Institute,
10 West 35th Street, Chicago, IL 60616.4The abbreviations used are: DHEA, dehydroepiandrosterone; DHEA ana
logue 8354, 16a-fluoro-5-androsten-17-one; MNU, N-methyl-N-nitrosourea; 4-HPR, /V-(4-hydroxyphenyl)retinamide; DMBA, 7,12-dimethylbenzanthracene;ANOVA. analysis of variance; NCW, net carcass weight; G6PDH, glucose-6-phosphate dehydrogenase; TPA, 12-O-tetradecanoylphorbol-13-acetate.
levels or excretion of this steroid or its metabolites and anumber of disease syndromes including cancer (2).
In women, plasma concentrations of DHEA (present almostexclusively as the sulfate conjugate, DHEA sulfate) show acontinual decline from peak levels that occur in the seconddecade of life (3, 4). Conversely, the incidence of breast cancerincreases with age (5). An early report by Bulbrook et al. (6)showed that preoperative urinary concentrations of 11-deoxy-17-ketosteroids were subnormal in women afflicted with primary breast cancer, and suggested that this alteration may be apredictor of risk for the subsequent appearance of the disease.Subsequent to that report, a prospective study showed a definiteinverse correlation between the urinary excretion of etiocholan-olone and androsterone, the two main urinary metabolites ofadrenal androgens, and the development of breast cancer inwomen (7). More recently, it was shown that 24-h mean plasmalevels of DHEA and DHEA sulfate were subnormal in womenwith premenopausal breast cancer, whereas postmenopausalpatients had supranormal plasma levels of these compounds(8), thus providing a potential biochemical marker for thedichotomization of this disease relative to menopausal status(5).
The in vivo relevance of DHEA to the genesis of neoplasticdisease has been experimentally explored in a number of animalmodels. Schwartz (9) has shown that p.o. administration ofDHEA inhibited the appearance of spontaneous mammarycancer in female C3H(A'J) mice and reduced the incidence and
multiplicity of lung tumors induced in A/J mice by eitherDMBA or urethan (10). In CD-I mice, topical application ofeither DHEA or the synthetic derivative 3/3-methylandrost-5-en-17-one significantly reduced the number and incidence ofDMBA-induced skin papillomas and carcinomas (11). Dietary
administration of DHEA also resulted in significant reductionsin the rate of appearance and frequency of all stages (atypicalhyperplasia, carcinoma in situ, invasive carcinoma) of colonielesions induced in female BALB/c mice with 1,2-dimethylhy-drazine (12). Finally, postcarcinogen dietary administration ofDHEA significantly inhibited the development of thyroid tumors and the frequency of putative neoplastic precursor lesionsin the livers of male F344 rats that were treated with dihydroxy-di-n-propylnitrosamine (13).
The results described above indicate that DHEA has significant chemopreventive efficacy against experimental carcinogenesis at a diverse group of target sites. However, with theexception of breast cancer, there is no clear precedent relatingspecific cancer risk in humans to alterations in circulating levelsof this steroid or the production and excretion of its metabolites(2). For this reason, and given the lack of previous experimentaldata regarding chemopreventive efficacy of DHEA in mammarycarcinogenesis, we have undertaken to explore the temporalspecificity of DHEA as an inhibitor of MNU-induced mammary
481
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from

DEHYDROEPIANDROSTERONE ANTICARCINOGENESIS
gland carcinogenesis in female rats. The potential clinical significance in using this breast cancer model is its similarity tothe human disease in terms of histopathology (14) and responseto hormonal manipulations (15-17). In addition, the use ofMNU, a direct acting mammary gland carcinogen (18), toinduce cancer effectively rules out complications related toDHEA-mediated effects on carcinogen metabolism and disposition (2, 10). Previous work demonstrated that the chemopre-ventive efficacy of retinoids (including 4-HPR) in experimentalbreast cancer is enhanced when they are combined with varioushormonal treatment modalities (19, 20). The present study wasdesigned to evaluate the potential steroid modulating effects ofDHEA, administered alone and in combination with the synthetic retinoid 4-HPR in rat mammary carcinogenesis. Finally,the chemopreventive activity of a fluorinated derivative ofDHEA, denoted DHEA analogue 8354 (21), was examined ina separate study. This analogue is a more potent inhibitor thanDHEA of several biochemical processes considered central tocarcinogenesis. The analogue apparently does not elicit theandrogenic and estrogenic effects associated with the parentsteroid (21, 22). Thus, comparison of these agents in chemo-prevention studies may lead to insights as to the mechanism(s)by which these compounds suppress carcinogenesis.
MATERIALS AND METHODS
Experimental Animals. Virgin female Sprague-Dawley [Hsd: (SD)BR] rats were received from Harlan/Sprague-Dawley (Indianapolis, IN)at 28 days of age and maintained in isolation for 2 weeks. A total of390 rats were used in the reported studies. Animals were housed ingroups of 2 to 3 in polycarbonate cages containing hardwood bedding.Cage materials were replaced twice weekly. The animal rooms wereilluminated for 12 h each day and maintained at a temperature of 22 ±1°C(SE) and 50% relative humidity. Animals were allowed free access
to food and water throughout each study.Diets and Chemopreventive Agents. The basal diet for each study was
modified AIN-76A semipurified diet (adjusted corn starch) TD 85449(Teklad, Madison, WI). 4-HPR was obtained from Cilag AG, Schaff-house, Switzerland. The required vehicle for 4-HPR consisted of 12.5g of ethanol, 36.9 g of corn oil, and 0.6 g of Tenox 20 (20% tertiarybutylhydroquinone, 10% citric acid, 70% propylene glycol; EastmanChemicals, Kingsport, TN) per kg diet. Control animals (carcinogenand carcinogen vehicle groups) in the study with DHEA received abasal diet containing the 4-HPR vehicle. DHEA was purchased fromSigma Chemical Co. (St. Louis, MO). The DHEA analogue 8354 wasa gift from Fort Washington Resources (Hatboro, PA). Its synthesisand chemical properties were described previously (21). Fresh batchesof diet were prepared weekly (high doses) or every 3 weeks (low doses),as more animals received the high-dose diets, and stored at —20°C
before use.As analyzed by high-pressure liquid chromatography (23), 4-HPR
was completely stable under the storage conditions used and when dietsupplemented with 4-HPR was left at room temperature for 4 days.The stability and content of DHEA and DHEA analogue 8354 in thebasal diet were assessed by high-pressure liquid chromatography usingextraction and Chromatographie methodology developed in our laboratory.5 Each steroid was extracted from the diet with 100% methanol
(50 ml/5 g of diet) by occasionally swirling the mixture over a periodof 2 h at ambient temperature. After gravity filtration to removepaniculate material, the volume of each filtrate was adjusted as necessary to 50 ml, and 10-^1aliquots were injected directly onto a WhatmanPartisi! ODS-2 column. The steroids were eluted from the columnusing 100% methanol at a flow rate of 1.2 ml/min. The DHEA peakwas visualized by UV absorption spectroscopy at a wavelength of 210nm; DHEA analogue 8354 was detected at 215 nm. Standard curves
*T. A. HuitÃn,unpublished observations.
were prepared by spiking 5 g of basal diet with known amounts ofmaterial, extracting and quantitating as described above. Recoveries ofDHEA were generally >90%; recoveries of DHEA analogue 8354 weregenerally >84%. As determined by the described method, both steroidswere stable for 4 days at room temperature when mixed in the basaldiet at the levels used in the studies. All diet materials were replacedtwice weekly.
DHEA/4-HPR Combination Chemoprevention Study. Two hundredsixty rats were randomized by weight into 12 groups at 43 days of age.With the exception of the carcinogen control group, which received adiet containing 4-HPR vehicle (30 rats), the other carcinogen-treatedexperimental groups (8 additional) each contained 25 animals. Groups(3 total) that received MNU vehicle (0.85% NaCl solution) consistedof 10 rats each. Provision of diets supplemented with steroid plusretinoid was also begun at this time according to 1 of 3 schedules, toencompass the periods of initiation (1 week before through 1 week afterMNU treatment, i.e., -1 week to +1 week), promotion/progression (1week after MNU treatment, continued to the end of the study, i.e., +1week to the end), or both (1 week before MNU treatment, continuedto the end of the study, i.e., —1week to the end). As determined inpreliminary feeding studies, DHEA was well tolerated when fed aloneat 0.2% or 0.1% (w/w) of diet (6.93 and 3.47 mmol/kg diet, respectively); combination diets contained the same doses of DHEA plus 4-HPR at 1 or 0.5 mmol (391 or 195.5 mg/kg diet, respectively).
At 50 days of age, all carcinogen-treated rats received a single i.v.injection of freshly prepared MNU solution (50 mg/kg body weight)via the jugular vein as described previously (18). Crystalline MNU(Ash-Stevens, Detroit, MI) was dissolved to a concentration of 12.5mg/ml in 0.85% NaCl solution acidified to pH ~5.0 with glacial acetic
acid. Control animals received an i.v. injection of the NaCl solutiononly.
Commencing 4 weeks after receiving MNU, animals were palpatedweekly to monitor mammary tumor appearance. The date of appearanceand location of every palpable tumor were recorded. The body weightsof all rats were recorded once a week for the duration of each study.All rats were observed twice daily for any indications of agent-inducedtoxicity. At no time during the experiments were the estrous cycles ofany rats monitored.
After 180 days of chemopreventive treatment, surviving rats in allgroups were sacrificed by CO2 asphyxiation. During each study, animalsthat appeared moribund were killed by CO2 asphyxiation. All rats thatwere killed or found dead were promptly given thorough postmortemexamination. Mammary tumors were coded by location, removed,measured, and weighed. All tumors and any other grossly abnormal-appearing tissues were removed and fixed in 10% buffered formalin.Sections were stained with hematoxylin and eosin and classified histo-pathologically. Mammary tumor pathology was defined according tothe criteria of Young and Hallowes (24).
DHEA Analogue 8354 Chemoprevention Study. One hundred thirtyrats were randomized by weight into 7 groups at 43 days of age. Eachgroup that received MNU (5 total) consisted of 20 animals; in addition,a control group that received steroid-supplemented diet and carcinogenvehicle also contained 20 rats. Another vehicle-treated group thatreceived AIN-76A basal diet contained 10 animals. Dose schedules forchemopreventive treatment were identical to those described above forthe DHEA/4-HPR combination study. The DHEA analogue 8354 wasfed to treated rats at 0.2% or 0.1% (w/w) of diet (6.89 and 3.44 mmol/kg diet, respectively). All procedures and observations were performedas described above for the DHEA/4-HPR combination study.
Statistical Analysis. Tumor incidence curves were generated by thelife table method and compared by log rank analysis (25). The statisticalsignificance of differences between mean tumor multiplicities was assessed using one-way ANOVA. Individual tumor numbers from theDHEA/4-HPR combination study were transformed by the square rootmethod to normalize the data before comparison by ANOVA (26).Differences in group total mean body weights at termination of theDHEA analogue 8354 study were tested for statistical significance byANOVA, using untransformed individual weights. Due to significantunresolved heterogeneity among the group variances, total mean body
482
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from

DEHYDROEPIANDROSTERONE ANTICARCINOGENESIS
Table 1 Effect of dietary DHEA on MNV-induced mammary gland cardnogenesis infernale Sprague-Dawley ratsVirgin, female Sprague-Dawley rats (50 days old) received single injections of MNU (50 mg/kg body weight) dissolved in a vehicle of 0.85% NaCI solution (pH
~5.0), or vehicle alone. Administration of AIN-76A semipurified diet (basal diet) supplemented with 4-HPR vehicle or DHEA at the indicated doses was begun asdescribed below. The study was terminated 180 days after rats received MNU or NaCI solution. Different numbers of asterisks in each column of data denotestatistically significant intergroup differences.
No. ofratsMNU24
+25+25+25+30+10Vehicle10
VehicleDHEA(%
diet,w/w)0.2'o.y0.2'0.1'—
*0.2*ACancerincidence
(%)100*67"48"68"100*Cancers/rat"'*8.8±1.0*2.3±0.6"0.9±0.2"1.4
±0.2"10.1±0.8*Tumors/rat"''9.1
±0.9*2.4±0.6"1.0
±0.2"1.4±0.2**10.7
±0.8*Terminal
survival63*8096"92**57*100100Terminaltotal bodywt(g)a269
±11265±6255
±5"261±5287
±12*265±7'294
±12Terminal
NCW(g)*-'254
±926125125724526529465510712
" Group mean average ±SEM.* Includes all histologically confirmed mammary carcinomas found at necropsy.' Includes all palpable tumors.d NCW represents terminal weight of tumor-bearing rat minus weight of excised tumors.' Diet given from —1week to +1 week post-MNU.^Diet given from +1 week post-MNU to end of study.* Diet given from -1 week post-MNU or vehicle to end of study.* Diet contained 4-HPR vehicle (see text for details).' P < 0.05 versus respective control group.
weights for the DHEA/4-HPR combination study were compared using(as appropriate) a i test for means with significant heterogeneity ofvariances, as suggested by Sokal and Rohlf (26). In the latter case,significance was attributed to P < 0.01. As shown in the "Results." we
also have calculated a NCW for each animal that survived to the endof each study. The NCW represents the total (gross) terminal weightof each tumor-bearing rat minus the total weight of tumor tissue (i.e.,the tumor burden). This parameter (NCW) was also evaluated byANOVA as described above to detect differences between the variousmeans. An appropriate discrimination test for making unplanned comparisons (least significant difference method) was used to define statistically significant intergroup differences in means (26). Differences inpercentage survival and cancer incidences were tested for significanceby Fisher's exact test (2-tailed). In all cases, with the exception of
multiple / tests, statistical significance was ascribed to a comparisononly when a P < 0.05 was attained.
RESULTS
DHEA/4-HPR Combination Study. The chemopreventive activity of DHEA alone, and DHEA plus 4-HPR, were examinedin a combined study using common MNU and MNU vehiclecontrols. However, to facilitate their presentation, the data havebeen separated into two Tables, with the common controlsshown in each.
The data summarized in Table 1 indicate that dietary DHEAsignificantly inhibited MNU-induced tumorigenesis in femalerats when given during the +1 week to end schedule. Thus, ratsfed DHEA at 0.2% (w/w) during this period developed cancerat an incidence of 67% and mean (±SEM) multiplicity of 2.3±0.6 cancers/rat, representing significant (P< 0.05) reductionsin these parameters relative to the controls. In contrast, treatment with 0.2% DHEA during the -1 week to +1 week period
had no significant effect on mammary cancer incidence (100%)or mean (±SEM) multiplicity (8.8 ±1.0 cancers/rat) whencompared with controls that received a diet containing 4-HPRvehicle only. This control group had a cancer incidence of 100%and mean (±SEM) multiplicity of 10.1 ±0.8 cancers/rat.Treatment with DHEA over the in-life phase of the study (—1
week to end) significantly reduced mammary cancer incidenceto 48% (0.2% dose) and 68% (0.1 % dose). Mean (±SEM)cancermultiplicity was 0.9 ±0.2 (0.2% dose) and 1.4 ±0.2 (0.1%dose) cancers/rat. However, statistical comparison of the tumordata for the two groups that received DHEA from —1week to
end at 0.1% or 0.2% showed no significant intergroup differences in cancer incidence or mean multiplicity. Finally, log rankanalysis of cancer incidence curves showed significant lengthening of cancer latency in groups treated with DHEA duringthe +1 week to end or —1week to end periods, relative to thosetreated during the —1week to +1 week period or the controls.
As shown in Table 2, combination treatment with DHEAplus 4-HPR was most effective when given from —1week to
the end of the study. Cancer incidence and multiplicity alsowere significantly lower in the groups treated with the combination during the +1 week to end period relative to the controls.Furthermore, cancer multiplicity in the rats that receivedDHEA plus 4-HPR during the —1week to +1 week period was
significantly lower than that of the carcinogen controls. Finally,comparison of the data in Tables 1 and 2 indicates that combined treatment with DHEA plus 4-HPR during the -1 weekto end period was significantly more effective in reducing cancermultiplicity and incidence than was treatment with the steroidalone. When administered alone to rats at 1 mmol/kg AIN-76A diet, 4-HPR has no effect on MNU-induced tumorigenesis.6
The survival data in Tables 1 and 2 show that 57% of thecarcinogen control rats lived until the end of the experiment(187 days); survival of rats that received DHEA alone rangedfrom 63% (-1 week to +1 week) to 96% (-1 week to end,0.2%) (Table 1). As shown in Table 2, survival of carcinogen-treated rats that received DHEA plus 4-HPR ranged from 72-100%. During the experiment, only 4 rats died in a cancer-freestate. In the control groups that received carcinogen vehicleand diet containing DHEA plus or minus 4-HPR, grossly visibleevidence of steroid-induced pathology was not observed atnecropsy.
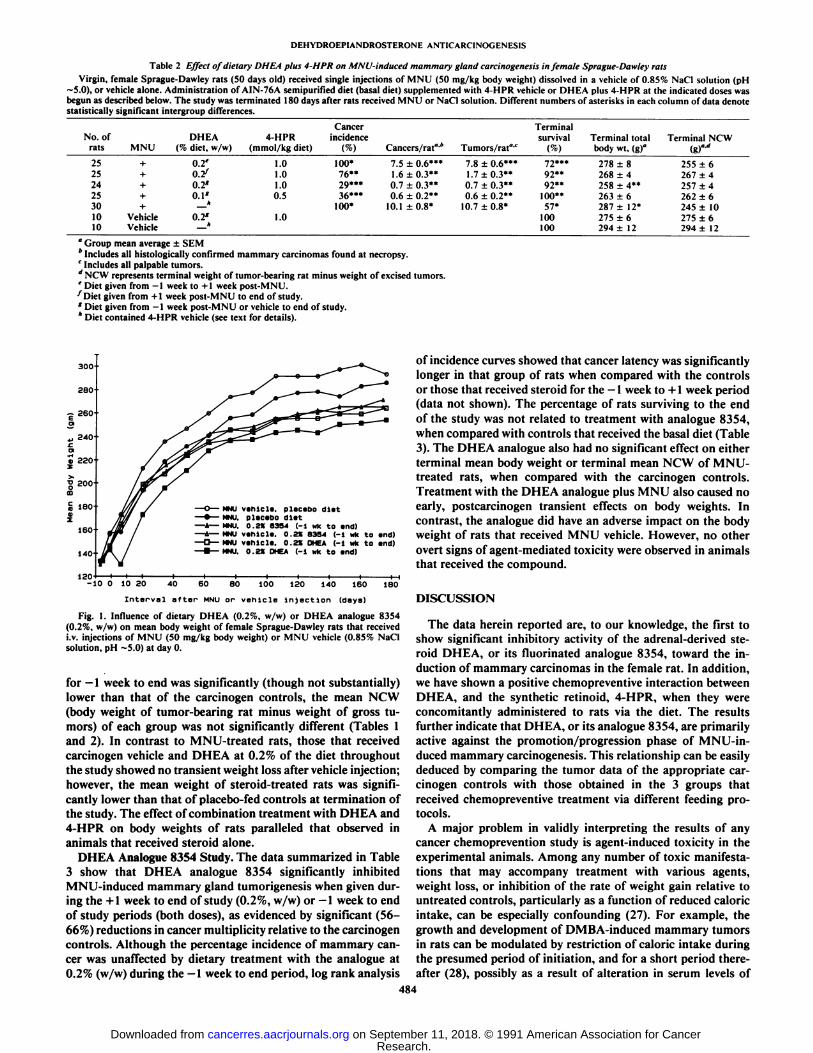
Fig. 1 shows that the mean body weight of rats treated withdietary DHEA at 0.2% (w/w) was approximately 18% lower (P< 0.05) than that of placebo-fed controls within 6 days ofreceiving MNU injections. However, the rate of weight gain inthat group recovered within the subsequent 7-14 days aftercarcinogen exposure, and was comparable with that of thecarcinogen controls for at least 60 additional days. At termination of the study, although the gross mean body weight ofthe MNU-treated group on the diet containing 0.2% DHEA
'T. A. Ratko. C. J. Detrisac, R. G. Mehla, G. J. Kelloff. and R. C. Moon,
unpublished observations.
483
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from
DEHYDROEPIANDROSTERONE ANTICARCINOGENESIS
Table 2 Effect of dietary DHEA plus 4-HPR on MNU-induced mammary gland curcinogenests infernale Sprague-Dawley ratsVirgin, female Sprague-Dawley rats (50 days old) received single injections of MNU (50 mg/kg body weight) dissolved in a vehicle of 0.85% NaCI solution (pH
~5.0), or vehicle alone. Administration of A1N-76A semipurified diet (basal diet) supplemented with 4-HPR vehicle or DHEA plus 4-HPR at the indicated doses wasbegun as described below. The study was terminated 180 days after rats received MNU or NaCI solution. Different numbers of asterisks in each column of data denotestatistically significant intergroup differences.
No. ofratsMNU25
+25+24+25+30+10Vehicle10
VehicleDHEA(%
diet,w/w)0.2'o.y0.2*0.1*A0.2*A4-HPR(mmol/kgdiet)1.01.01.00.51.0Cancerincidence100*76**29*"36*"100*Cancers/rat""''7.5
±0.6***1.6+0.3"0.7±0.3"0.6+0.2**10.1±0.8*Tumors/rat°'c7.8
±0.6***1.7+0.3"0.7±0.3**0.6+0.2**10.7±0.8*Terminal
survival72***92"92**100"57*100100Terminaltotalbody wt.(g)°278
+8268±4258
±4"263+6287±12*275+6294±12Terminal
NCW(g)"-'2552672572622452756446106294+
12°Group mean average ±SEM* Includes all histologically confirmed mammary carcinomas found at necropsy.' Includes all palpable tumors.
NCW represents terminal weight of tumor-bearing rat minus weight of excised tumors.e Diet given from —1week to +1 week post-MNU.'Diet given from +1 week post-MNU to end of study.* Diet given from —1week post-MNU or vehicle to end of study.* Diet contained 4-HPR vehicle (see text for details).
220
200
160
120
O—MNU vehicle, plecebo diet•—MNU. plecebo diet•A MNU. 0.2X B354 (-1 Hk to end)•4—MNUvehicle. 0.2X 8354 (-1 Hk to end)O—MNU vehicle. 0.2X DHEA (-1 wk to end)•—MNU. 0.2X DHEA (-1 Hk to end)
-10 0 10 20 40 60 80 100 120 140 160 180
Interval after MNU or vehicle injection (days)
Fig. 1. Influence of dietary DHEA (0.2%, w/w) or DHEA analogue 8354(0.2%, w/w) on mean body weight of female Sprague-Dawley rats that receivedi.v. injections of MNU (50 mg/kg body weight) or MNU vehicle (0.85% NaCIsolution, pH ~5.0) at day 0.
for —1week to end was significantly (though not substantially)
lower than that of the carcinogen controls, the mean NCW(body weight of tumor-bearing rat minus weight of gross tumors) of each group was not significantly different (Tables 1and 2). In contrast to MNU-treated rats, those that receivedcarcinogen vehicle and DHEA at 0.2% of the diet throughoutthe study showed no transient weight loss after vehicle injection;however, the mean weight of steroid-treated rats was significantly lower than that of placebo-fed controls at termination ofthe study. The effect of combination treatment with DHEA and4-HPR on body weights of rats paralleled that observed inanimals that received steroid alone.
DHEA Analogue 8354 Study. The data summarized in Table3 show that DHEA analogue 8354 significantly inhibitedMNU-induced mammary gland tumorigenesis when given during the +1 week to end of study (0.2%, w/w) or -1 week to endof study periods (both doses), as evidenced by significant (56-66%) reductions in cancer multiplicity relative to the carcinogencontrols. Although the percentage incidence of mammary cancer was unaffected by dietary treatment with the analogue at0.2% (w/w) during the —1week to end period, log rank analysis
of incidence curves showed that cancer latency was significantlylonger in that group of rats when compared with the controlsor those that received steroid for the —1week to +1 week period(data not shown). The percentage of rats surviving to the endof the study was not related to treatment with analogue 8354,when compared with controls that received the basal diet (Table3). The DHEA analogue also had no significant effect on eitherterminal mean body weight or terminal mean NCW of MNU-treated rats, when compared with the carcinogen controls.Treatment with the DHEA analogue plus MNU also caused noearly, postcarcinogen transient effects on body weights. Incontrast, the analogue did have an adverse impact on the bodyweight of rats that received MNU vehicle. However, no otherovert signs of agent-mediated toxicity were observed in animalsthat received the compound.
DISCUSSION
The data herein reported are, to our knowledge, the first toshow significant inhibitory activity of the adrenal-derived steroid DHEA, or its fluorinated analogue 8354, toward the induction of mammary carcinomas in the female rat. In addition,we have shown a positive chemopreventive interaction betweenDHEA, and the synthetic retinoid, 4-HPR, when they wereconcomitantly administered to rats via the diet. The resultsfurther indicate that DHEA, or its analogue 8354, are primarilyactive against the promotion/progression phase of MNU-induced mammary carcinogenesis. This relationship can be easilydeduced by comparing the tumor data of the appropriate carcinogen controls with those obtained in the 3 groups thatreceived chemopreventive treatment via different feeding protocols.
A major problem in validly interpreting the results of anycancer chemoprevention study is agent-induced toxicity in theexperimental animals. Among any number of toxic manifestations that may accompany treatment with various agents,weight loss, or inhibition of the rate of weight gain relative tountreated controls, particularly as a function of reduced caloricintake, can be especially confounding (27). For example, thegrowth and development of DMBA-induced mammary tumorsin rats can be modulated by restriction of caloric intake duringthe presumed period of initiation, and for a short period thereafter (28), possibly as a result of alteration in serum levels of
484
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from

DEHYDROEPIANDROSTERONE ANTICARCINOGENESIS
Table 3 Effect of dietary DHEA analogue 8354 on MNU-induced mammary gland carcinogenesis infernale Sprague-Dawley ratsVirgin, female Sprague-Dawley rats (50 days old) received single injections of MNU (50 mg/kg body weight) dissolved in a vehicle of 0.85% NaCI solution (pH
~5.0), or vehicle alone. Administration of AIN-76 semipurified diet (basal diet) supplemented with DHEA analogue 8354 at the indicated doses was begun asdescribed below. The study was terminated 180 days after rats received MNU or NaCI solution. Different numbers of asterisks in each column of data denotestatistically significant intergroup differences.
No. ofratsMNU20
+20+20+20+20+20Vehicle10
VehicleDHEA
8354(% diet,w/w)0.2'o.y0.2'0.1"—
*0.2"ACancer
incidence10095100100100Cancers/rat"'*11.2
±1.0*4.1±0.6**4.8
±1.1**5.4±0.8"12.2±
1.0*Tumors/rat°'r11.5
±1.0*4.2±0.6**4.8±1.0"5.8±0.8"12.6+ 0.9*Terminal
survival6555805070100100Terminaltotal bodywt(g)fl276
±7254±12272
±5265±5266±7266
±5*305±10"Terminal
NCW(g)"'"258
±7239±14259
±6255±6240±6266
±5305±10
a Group mean average ±SEM* Includes all histologically confirmed mammary carcinomas found at necropsy.1Includes all palpable tumors.d NCW represents terminal weight of tumor-bearing rat minus weight of excised tumors.' Diet given from -1 week to +1 week post-MNU.^Diet given from +1 week post-MNU to end of study.* Diet given from —1week post-MNU or vehicle to end of study.* Basal diet (AIN-76A).
prolactin and gonadotropins (29). In the present study, we haveno data showing an effect of dietary DHEA on food consumption, therefore, it is not possible to identify alteration of caloricintake as a specific factor that could be causally related to theearly transient weight loss observed in DHEA-treated ratsimmediately after their exposure to MNU (Fig. 1). Since transient weight loss did not occur in the groups that receivedDHEA and MNU vehicle, it appears that toxicity of the steroidper se was not a sole factor in that phenomenon. Interestingly,a similar effect was documented by Schwartz et al. (30) infemale CD-I mice, which were fed DHEA at 0.4% or analogue8354 at 0.2% and 0.4% in the diet, concomitant with theirexposure to DMBA via topical application.
Thus, our data suggest that the observed anticarcinogenicityof DHEA was specific to some unknown action of the steroid,rather than to inhibition of body weight gain. This interpretation is also supported by the NCW data, which indicate similarbody weights in all rats, regardless of the DHEA treatmentprotocol. However, it is possible, though difficult to quantify,that in the carcinogen control rats, the sizeable mean tumorburden induced significant cachexia, which could have reducedtheir mean NCW, thus giving the appearance that treatmentwith DHEA had no effect on this parameter. This may indeedhave occurred, as evidenced by the lower mean NCW of thecarcinogen controls, although data are not available to supportthis contention.
The present data show that DHEA analogue 8354 did notsignificantly affect weight gain of rats given the steroid at 0.1 %of 0.2% (w/w) via the diet. This result contrasts with datareported by Schwartz et al. (21 ), who showed a significant effectof analogue 8354 on weight gain of male BALB/cJ mice thatreceived the agent at 0.25% in the diet, despite their consumingquantities of food similar to those ingested by controls. However, due to the differences in species and sex of animals, it isdifficult to directly compare our results with those of Schwartzet al. (21).
Examination of the findings from animals receiving DHEAalone or in combination with 4-HPR reveals two beneficialeffects associated with the addition of 4-HPR to the DHEAdiets. At both doses evaluated, mammary cancer incidence wasfurther reduced when the combination diets were given throughout all phases of carcinogenesis. Furthermore, concomitantadministration of these agents significantly lowered cancer mul
tiplicity during the initiation phase. The results of the 4-HPR/DHEA combination study are not without precedent in showinga positive chemopreventive interaction between a retinoid andhormonal manipulation in the MNU-induced mammary carcinogenesis model. For example, in rats that received dietary 4-HPR, s.c. injections of tamoxifen, or 4-HPR plus tamoxifeninjections, the incidence and multiplicity of subsequent tumorsafter surgical excision of the first lesion were reduced to agreater extent by the combination of agents when comparedwith either dietary 4-HPR alone or injections of tamoxifenalone (19). A previous study showed a positive interactionbetween tamoxifen and 4-HPR in the inhibition of MNU-induced tumorigenesis (20). Also, when compared with eithertreatment mode alone, an enhanced inhibitory effect on mammary carcinogenesis was observed in ovariectomized rats thatalso received dietary 4-HPR (31). Thus, the significant interaction between DHEA and 4-HPR, as was previously documented between the retinoid and various other hormonal manipulations, suggests that further investigation of this combination in cancer chemoprevention is warranted.
Both DHEA and its analogue 8354 significantly modify anumber of processes that are considered to be central to tumorpromotion. These effects include the inhibition by DHEA ofDNA synthesis in breast epithelium of C3H mice and phorbolester-stimulated skin of ICR mice (32), reduction of TPA
stimulation of Superoxide production in human neutrophils(33), and inhibition of TPA-induced increases of prostaglandinE2 levels in mouse skin (22). In addition, both DHEA andDHEA analogue 8354 are potent inhibitors of mammalianG6PDH (21), the first enzyme in the hexose-monophosphateshunt (34). This metabolic pathway is the primary extramito-chondrial source of NADPH, which is required in a number ofbiochemical processes including xenobiotic catabolism bymixed function oxidase, and the biosynthesis of fatty acids andsteroid hormones (34). It is also the primary pathway by whichhexose phosphates are converted to the ribose and deoxyribosephosphates required for de novo biosynthesis of nucleotides andnucleic acids (34). Thus, it is apparent that treatment of ratswith either DHEA or its analogue could have profound effectson a metabolically active population of target cells of the youngrat mammary gland, with several possible loci at which tumorpromotion could be blocked or suppressed, perhaps as a function of the inhibition of G6PDH (35).
485
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from

DEHYDROEPIANDROSTERONE ANTICARCINOGENESIS
Chemical modification of the basic DHEA structure has beenused as a strategy by other investigators to reduce the incidenceand severity of side effects engendered in experimental animalsby the parent hormone. Schwartz et al. (21) recently showedthat contrary to the effects seen with DHEA, the DHEA 8354analogue was nonestrogenic in a rat uterine weight assay, andnonandrogenic when assayed using the castrated rat seminalvesicle model. This analogue is also more potent than the parentsteroid in suppressing TPA-induced increases in epidermalDNA synthesis, Superoxide formation, and prostaglandin E2synthesis (22). Our findings demonstrate no increased efficacyof the 8354 analogue over DHEA in the promotion and progression phases of mammary carcinogenesis. This suggests thatthe previously measured parameters that differentiate thesecompounds in vivo are not responsible for these observed chem-opreventive effects. The similarity of the data for DHEA andthe DHEA 8354 analogue in the MNU rat mammary modelsystem thus may be related to the effects on G6PDH activity,which are similar for both compounds (21). The effects ofchronic p.o. administration of DHEA or DHEA analogue 8354on elements of rat reproductive physiology are presently unknown. A study to evaluate blood steroid hormone levels duringsuch chronic conditions is presently underway. Such data willprovide additional avenues for exploration of the chemopreven-tive mechanism of action of these compounds in this model ofhuman breast cancer.
ACKNOWLEDGMENTS
The authors wish to thank our technical staff for their expert performance. Additional thanks are due to Fort Washington Resourcesfor providing the DHEA analogue 8354, Dr. A. G. Schwartz for helpfuldiscussion regarding this project, and P. Moser for excellent secretarialassistance in preparing this manuscript.
REFERENCES
1. Vande Wiele, R., and Lieberman, S. The metabolism of dehydroisoandros-terone. In: G. Pincus and E. Vollmer, (eds.). Biological Activity of Steroidsin Relation to Cancer, pp. 93-110. New York: Academic Press. Inc., 1960.
2. Gordon, G. B., Shantz, L. M., and Talalay, P. Modulalion of growth,differentiation and carcinogenesis by dehydroepiandrosterone. Adv. EnzymeRegul., 26: 355-382, 1987.
3. Orentreich, N., Brind, J. L., Rizer, R. L., and Vogelman, J. H. Age changesand sex differences in serum dehydroepiandrosterone sulfate concentrationsthroughout adulthood. J. Clin. Endocrinol. Metab., 59: 551-555, 1984.
4. Carlstrom, K., Brody, S., Lunnell, N-O.. Lagrelius, A., Mollerstrom. G..Pousette, A., Rannevik, G., Stege, R., and von Schoultz, B. Dehydroepiandrosterone sulphate and dehydroepiandrosterone in serum: differences related to age and sex. Maturitas, 10: 297-306. 1988.
5. DeWaard. F.. DeLaive, J. W. J., and Baanders-Van Halewijn. E. A. On thebimodal age distribution of mammary carcinoma. Br. J. Cancer, 14: 437-448, 1960.
6. Bulbrook, R. D.. Hayward, J. L., Spicer, C. C, and Thomas, B. S. Abnormalexcretion of urinary steroids by women with early breast cancer. Lancet, 2:1238-1240, 1962.
7. Bulbrook, R. D., Hayward, J. L., and Spicer. C. C. Relation between urinaryandrogen and corticoid excretion and subsequent breast cancer. Lancet. 2:395-398, 1971.
8. Zumorff, B., Levin, J., Rosenfeld, R. S., Markham, M., Strain, G. W., andFukushima, D. K. Abnormal 24-hr mean plasma concentrations of dehydro-isoandrosterone and dehydroisoandrosterone sulfate in women with primaryoperable breast cancer. Cancer Res., 41: 3360-3363, 1981.
9. Schwartz, A. G. Inhibition of spontaneous breast cancer formation in femaleC3H (A'Va) mice by long-term treatment with dehydroepiandrosterone.Cancer Res., 39: 1129-1132, 1979.
10. Schwartz, A. G., and Tannen, R. H. Inhibition of 7,12-dimethylbenz[a]anthracene- and urethan-induced lung tumor formation in A/J mice by long-term treatment with dehydroepiandrosterone. Carcinogenesis (Lond.), 2:1335-1337. 1981.
11. Pashko, L. L., Hard, G. C, Rovito. R. J.. Williams, J. R., Sobel, E. L.. andSchwartz. A. G. Inhibition of 7,12-dimethylbcnz(a)anthracene-induced skinpapillomas and carcinomas by dehydroepiandrosterone and 3-fi-methyl-an-drost-5-en-l7-one in mice. Cancer Res.. 45: 164-166. 1985.
12. Nyce, J. W.. Magee. P. N.. Hard, G. C., and Schwartz. A. G. Inhibition of1,2-dimethylhydrazine-induced colon tumorigenesis in BALB/c mice by dehydroepiandrosterone. Carcinogenesis (Lond.), 5: 57-62. 1984.
13. Moore, M. A, Thamavit, W., Tsuda, H., Sato, K.. Ichihara. A., and Ito, N.Modifying influence of dehydroepiandrosterone on the development of dih-ydroxy-di-n-propylnitrosamine-initiated lesions in the thyroid, lung and liverof F344 rats. Carcinogenesis (Lond.), 7: 311-316, 1986.
14. Tseng, M. T. Ultrastructure of the hormone-dependent jV-nitrosomethylurea-induced mammary carcinoma of (he rat. Cancer Res., 40: 3112-3115. 1980.
15. Turcot-Lemay. L., and Kelly, P. A. Characterization of estradiol, progesterone, and prolactin receptors in nitrosomelhylurea-induced mammary tumorsand effect of antiestrogen treatment on the development and growth of thesetumors. Cancer Res., 40: 3232-3240. 1980.
16. Arafah, B. M., Fullino. P. M., Manni. A., and Pearson. O. H. Effect ofovariectomy on hormone receptors and growth of A'-nitrosomethylurea-induced mammary tumors in the rat. Cancer Res., 40:4628-4630, 1980.
17. Gandilhon, P., Melancon, R., Djiane, J., and Kelly. P. A. 7V-Nitroso-Af-methylurea-induced mammary tumors in the rat: role of prolactin and aprolactin-lowering drug. J. Nati. Cancer Inst., 70: 105-109, 1983.
18. McCormick, D. L.. Adamowski. C. B.. Fiks, A., and Moon, R. C. Lifetimedose-response relationships for mammary tumor induction by a single administration of A'-methyl-A'-nitrosourea. Cancer Res., 41: 1690-1694. 1981.
19. Ratko. T. A,. Detrisac, C. J., Dinger, N. M., Thomas, C. F., Kelloff, G. J.,and Moon, R. C. Chemopreventive efficacy of combined retinoid and tamox-ifen treatment following surgical excision of a primary mammary cancer infemale rats. Cancer Res., 49:4472-4476, 1989.
20. McCormick, D. L.. and Moon. R. C. Retinoid-tamoxifen interaction inmammary cancer chemoprevention. Carcinogenesis (Lond.), 7: 193-196,1986.
21. Schwartz, A. G., Lewbart, M. L., and Pashko, L. L. Novel dehydroepiandrosterone analogues with enhanced biological activity and reduced sideeffects in mice and rats. Cancer Res., 48:4817-4822, 1988.
22. Hastings. L. A., Pashko. L. L., Lewbart, M. L., and Schwartz, A. G.Dehydroepiandrosterone and two structural analogs inhibit 12-O-tetradeca-noylphorbol-13-acetale stimulation of prostaglandin E2 content in mouseskin. Carcinogenesis (Lond.). 9: 1099-1102, 1988.
23. HuitÃn,T. A., Mehta, R. G., and Moon, R. C. A simple high-pressure liquidChromatographie method for the separation of retinoids including \ (4hydroxyphenyl)-all-rrans-retinamide. J. Chromatogr., 341: 187-192, 1985.
24. Young, S.. and Hallowes, R. C. Tumours of the mammary gland. In: V. S.Turusov (ed.). Pathology of Tumours in Laboratory Animals, Vol. I, Part I,pp. 31-73. Lyon, France: International Agency for Research on Cancer,1973.
25. Peto, R., Pike, M. C., Armitage. P.. Breslow. N. E., Cox, D. R., Howard, S.V., Mantel, N., McPherson. K.. Peto. J., and Smith, P. G. Design andanalysis of randomized clinical trials requiring prolonged observations ofeach patient. II. Analysis and examples. Br. J. Cancer, 35: 1-39, 1977.
26. Sokal, R. R., and Rohlf, F. J. Biometry. San Francisco: W. H. Freeman andCo., 1981.
27. Tannenbaum. A. Genesis and growth of tumors. II. Effects of caloric restriction per se. Cancer Res., 2: 460-467, 1942.
28. Sylvester, P. W., Aylsworth. C. F., and Meites, J. Relationship of hormonesto inhibition of mammary tumor development by underfeeding during the"critical period" after carcinogen administration. Cancer Res., 41: 1384-
1388, 1981.29. Campbell, G. A.. Kurcz, M., Marshall, S., and Meites. J. Effects of starvation
in rats on serum levels of follicle stimulating hormone, leuteinizing hormone,thyrotropin, growth hormone and prolactin; response to LH-releasing hormone and thyrotropin-releasing hormone. Endocrinology, 100: 580-587,1977.
30. Schwartz, A. G., Fairman, D. K.. Polansky, M., Lewbart, M. L., and Pashko,L. L. Inhibition of 7,12-dimethylben/.|a]-anthracene-initiatedand 12-O-tetra-decanoylphorbol-13-acetate-promoted skin papilloma formation in mice bydehydroepiandrosterone and two synthetic analogs. Carcinogenesis (Lond.),10: 1809-1813. 1989.
31. McCormick, D. L., Mehta, R. G., Thompson, C. A., Dinger, N., Caldwell,J. A., and Moon, R. C. Enhanced inhibition of mammary carcinogenesis bycombined treatment with A'-(4-hydroxyphenyl)retinamide and ovariectomy.Cancer Res., 42: 508-512, 1982.
32. Pashko, L. L., Schwartz. A. G., AbouGharbia, M.. and Swern, D. Inhibitionof DNA synthesis in mouse epidermis and breast epithelium by dehydroepiandrosterone and related steroids. Carcinogenesis (Lond.). 2:717-721. 1981.
33. Whitcomb, J. M., and Schwartz. A. G. Dehydroepiandrosterone and 16«-Br-epiandrosterone inhibit 12-O-tetradecanylphorbol-13-acetate stimulationof Superoxide radical production by human polymorphonuclear leukocytes.Carcinogenesis (Lond.), 6: 333-335, 1985.
34. Lehninger, A. L. Biochemistry'- New York: Worth Publishers. Inc., 1975.35. Raineri, R., and Levy, H. R. On the specificity of steroid interaction with
mammary gland glucose-6-phosphate dehydrogenase. Biochemistry. 9:2233-2243, 1970.
486
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from

1991;51:481-486. Cancer Res Thomas A. Ratko, Carol J. Detrisac, Rajendra G. Mehta, et al. DehydroepiandrosteroneDietary Dehydroepiandrosterone or a Fluorinated Analogue of Inhibition of Rat Mammary Gland Chemical Carcinogenesis by
Updated version
http://cancerres.aacrjournals.org/content/51/2/481
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/51/2/481To request permission to re-use all or part of this article, use this link
Research. on September 11, 2018. © 1991 American Association for Cancercancerres.aacrjournals.org Downloaded from