inhibition of cytotoxic responses by prostaglandin e2 in the presence of interleukin 2
TRANSCRIPT

CELLULAR IMMUNOLOGY 72, 286-293 (1982)
Inhibition of Cytotoxic Responses by Prostaglandin E2 in the Presence of lnterleukin 2
MICHAEL WOLF AND WULF DROEGE
Institut fir Immunologic und Genetik. Deutsches Krebsforschungszentrum, D-69 Heidelberg, Federal Republic of Germany
Received May 7, 1982; accepted July 6, I982
Experiments are described which provide direct evidence for a strong suppressive effect of prostaglandin Er (PGE2) on the activation of cytotoxic T lymphocytes (CTL). The experiments also tested the hypothesis that PGEz inhibits cytotoxic responses exclusively by preventing the helper T cells from producing interleukin 2 (IL-2). In support of this hypothesis we found that indomethacin enhanced cytotoxic responses in the absence but not in the presence of con- canavalin A-activated spleen cell supernatant (CSCS) indicating that the endogenous/y pro- duced prostaglandin(s) inhibited primarily the production of IL-2. However, the addition of PGEz to microcultures was found to inhibit in a dose-dependent manner the activation of cytotoxic responses against I-region compatible stimulator cells even in the presence of CSCS. This inhibition was not observed in the presence of I-region incompatible stimulator cells, indicating that the inhibitory effect of PGEz did not result from a general toxic effect on the culture. The results argue against the notion that PGEz inhibits cytotoxic responses exclusively by preventing the production of IL-2.
INTRODUCTION
Several lines of indirect evidence have previously suggested that prostaglandins inhibit the activation of cytotoxic T lymphocytes (CTL):’ Prostaglandins have been shown to inhibit proliferative responses of lymphoid cells (1-4, reviewed in (5, 6)); and indomethacin and other inhibitors of prostaglandin synthesis have recently been shown to enhance cytotoxic responses against allogeneic cells if added during the induction but not during the effector phase of the cytotoxic response (3). Prosta- glandin E, and E2 have also been reported to inhibit the production of interleukin 2 (IL-2) (4, 7), a soluble mediator which is produced by activated helper T cells and is required for the activation of CTL precursor cells as a second signal (for review see (8, 9)).
Our present experiments provide direct evidence for a strong inhibitory effect of PGE2 on cytotoxic responses; and they indicate that this inhibitory effect is not exclusively explained by a suppression of the production of IL-2.
MATERIALS AND METHODS
Animals. The congenic strains of mice were obtained from a stock at the German Cancer Research Center. All other strains of mice were obtained from Bomholtgard,
’ Abbreviations: CSCS, concanavalin A-activated spleen cell supematant; CTL, cytotoxic T lympho- cyte(s); IL-2, interleukin 2; PGE2, prostaglandin Er.
286
0008-8749/82/140286-08$02.00/O Copyright Ca 1982 by Academic Press, Inc. All rights of reproduction in any form resCNcd.

REGULATION OF CYTOTOXIC RESPONSES BY PROSTAGLANDIN(S) 287
Ry, Denmark. Most of the experiments were performed with 8- to 12-week-old male animals.
The preparation of concanavalin A-activated spleen cell supernatant (CSCS). The CSCS preparations were obtained by culturing 3 X lo8 spleen cells from CBA mice in 30 ml Gibco medium 199 containing 0.15 mg concanavalin A (Gibco), 10% fetal calf serum (Paesel), and 3 x 10e5 M 2-mercaptoethanol for 24 hr. The supernatants were isolated by centrifugation at 300g for 10 min and stored frozen. a-Methyl- mannoside at a final concentration of 1 mM (0.194 mg/ml) plus HEPES (5 X 10m3 M) was added to the supernatant in order to neutralize the contaminating conca- navalin A.
Induction and assay of cytotoxic T lymphocytes. Our procedures for the induction and assay of CTL in microcultures have previously been described in detail ( 10, 11) and were based upon procedures of Teh et al. (12) and Shearer (13), with minor modifications. The microcultures contained usually 2 X lo4 or 5 X lo4 lymph node cells together with 3 X lo5 irradiated (1500 rad) stimulator cells and the indicated amount of CSCS in 0.2 ml RPMI-1640 (GIBCO) supplemented with glutamine, streptomycin, penicillin, HEPES, 10% fetal calf serum, and 3 X 10e5 M 2-mercap- toethanol ( 10, 11). Each group was tested in four parallel microcultures with a separate control of four wells without responder cells. The cultures were incubated for 5 days at 37°C under 5% COZ and then tested for cytotoxic activity in a 4 hr 5’Cr-release assay ( lo- 13) using 2 X 1 O4 target cells per well.
Our macrocultures contained usually 10 X lo6 responder cells and 10 X lo6 ir- radiated ( 1500 rad) stimulator cells in 4.5 ml culture medium if not indicated oth- erwise. The cytotoxic activities were tested 5 days later on 2 X lo4 target cells at effector:target cells ratios of 25:l and 5:l.
The target cells were prepared by culturing spleen cells together with 2.2 pg/ml concanavalin A (Pharmacia, Uppsala, Sweden) for 2 days (11). Stimulator and target cells were haptenated with 10 mM 2,4,6,-trinitrobenzene sulfonic acid (Sigma) for 10 min at 37°C (12). The data from cytotoxic assays were indicated as specific 5’Cr- release or as cytotoxic activities (Nat) which were computed according to the equa- tion ( 12) Nat = log, [ 1 - (100/P)] where P = percentage specific 5’Cr-release.
The application of indomethacin and prostaglandin Ez. Indomethacin is a non- steroidal anti-inflammatory drug and a potent irreversible inhibitor of prostaglandin synthesis (14-16). A stock solution of indomethacin (Sigma) was prepared by dis- solving 10 mg/ml in ethanol. The cell cultures contained usually 0.5 pg indomethacin per ml, i.e., a 1:20,000 dilution of the stock solution in culture medium.
Prostaglandin E2 was a gift from Dr. John Pike (Upjohn, Kalamazoo, Mich.) and also kindly provided to us by Dr. D. Gemsa. Our stock solution contained lo-* M PGE2 in ethanol (3.5 mg/ml) and was stored at -20°C. It was applied to cultures at concentration of 10-6-10-9 M.
Irradiation. y-Irradiation was performed with a 6oCo source usually at a dose of 1500 rad.
RESULTS
The Efect of Indomethacin on Cytotoxic Responses against TNP-Haptenated Syn- geneic Cells in Macrocultures and against Allogeneic Cells in Microcultures
Indomethacin is an irreversible inhibitor of the prostaglandin biosynthesis (14- 16) and was found to enhance cytotoxic responses against ailogeneic cells in macro-

288 WOLF AND DROEGE
LJQ lndomethacm added per culture
day 0 day 1 day 2 day 3
OOOO1
1 1 0 0
2 2 0 0
1 0 0 2
2 0 0 2
1 1 1 0
2 2 2 0
1 1 1 1
2 2 2 2
effector-target ratio 25 1 m 51 0
I I
1
I I I 01 02 03
CytOIOxlc actwity (Nat )
I I 1 I I I 0 4 6 12 16 20 24
‘0 Weclfic 5’0.release
FIG. 1. Effect of indomethacin in macrocultures at different time intervals. DBA/Z spleen cells ( 10 X 106) were cultured together with 10 X IO6 TNP-haptenated irradiated (1500 tad) syngeneic spleen cells in 4.5 ml macrocultures and the cytotoxic activity was tested on TNP-haptenated syngeneic cells 5 days later. The indicated amounts of indomethacin were added at various days after initiation of the culture.
cultures (3). We confirmed this enhancing effect for the cytotoxic response of DBA/ 2 cells against TNP-haptenated DBA/2 stimulator cells, and we found in this case that a single dose of indomethacin at the onset of the culture was clearly less effective than repeated applications (Fig. 1). This indicated that the late phase of the cytotoxic response was still sensitive against the inhibitory effect of prostaglandin(s). We also confirmed in this system that the cytotoxic response was equally well enhanced by the drug Ro20-5720 (from Roche, Nutley, N.J., kindly provided by Dr. Gemsa) which inhibits as a competitive and reversible inhibitor a different step in the bio- synthesis of prostaglandin (5, 16) (data not shown).
The subsequent experiment (Fig. 2) showed that the response of DBA/2 cells against TNP-haptenated DBA/2 stimulator cells was also enhanced by the addition of concanavalin A-activated spleen cell supematant (CSCS), and that the combined application of indomethacin and CSCS resulted in no further enhancement of the response. Essentially the same effect was observed in the microculture reaction of C3H responder cells against allogeneic stimulator cells (Table 1). C3H mice and other strains with the H-2k haplotype generate exceptionally strong CTL responses against TNP-haptenated syngeneic cells (see (10)); and C3H spleen and lymph node cells produce in macrocultures substantial CTL responses even in the absence of indomethacin and CSCS (data not shown). The response of C3H lymph node cells in microcultures in the absence of CSCS and indomethacin, however, was minute and significantly enhanced by the addition of 0.1 mg indomethacin per culture. Stronger responses were obtained in the presence of CSCS, and the cytotoxic re- sponses were in this case not further enhanced by the combined application of CSCS and indomethacin. The results in these two very different experimental systems were
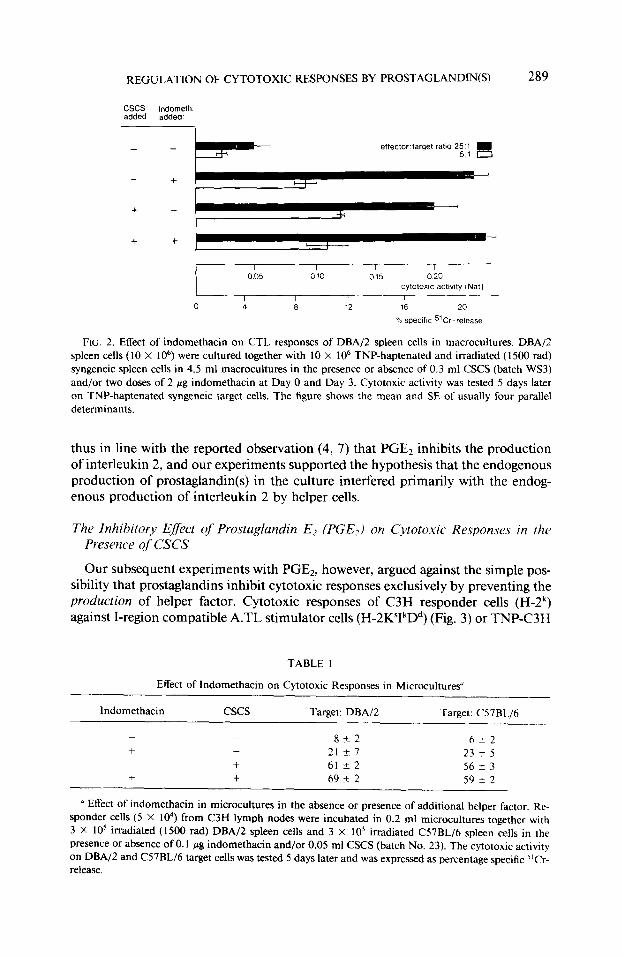
REGULATION OF CYTOTOXIC RESPONSES BY PROSTAGLANDIN(S) 289
- -I-- effector target ratio 25 1 I 51 n
+
+ -
+ f
0.b5 I I
010 015 020 cytotoxlc actwty I Nat 1
I I I / I 0 4 8 12
%‘:,ec,fI, 5k.rL
FIG. 2. Effect of indomethacin on CTL responses of DBA/Z spleen cells in macrocultures. DBA/Z spleen cells (10 X 1 06) were cultured together with 10 X lo6 TNP-haptenated and irradiated (1500 rad) syngeneic spleen cells in 4.5 ml macrocultures in the presence or absence of 0.3 ml CSCS (batch WS3) and/or two doses of 2 pg indomethacin at Day 0 and Day 3. Cytotoxic activity was tested 5 days later on TNP-haptenated syngeneic target cells. The figure shows the mean and SE of usually four parallel determinants.
thus in line with the reported observation (4, 7) that PGE2 inhibits the production of interleukin 2, and our experiments supported the hypothesis that the endogenous production of prostaglandin(s) in the culture interfered primarily with the endog- enous production of interleukin 2 by helper cells.
The Inhibitory Efect qf Prostaglandin E2 (PGEJ on Cvtotoxic Responses in the Presence of CSCS
Our subsequent experiments with PGE2, however, argued against the simple pos- sibility that prostaglandins inhibit cytotoxic responses exclusively by preventing the production of helper factor. Cytotoxic responses of C3H responder cells (H-2k) against I-region compatible A.TL stimulator cells (H-2KSIkDd) (Fig. 3) or TNP-C3H
TABLE I
Effect of Indomethacin on Cytotoxic Responses in Microcultures”
Indomethacin cscs Target: DBA/Z Target: CSlBL/6
- 842 6i-2 + - 2127 23 k 5
+ 61 -t2 56 -+ 3 + + 69 -t 2 59 k 2
(i Effect of indomethacin in microcultures in the absence or presence of additional helper factor. Re- sponder cells (5 X 104) from C3H lymph nodes were incubated in 0.2 ml microcultures together with 3 X IO’ irradiated (1500 rad) DBA/2 spleen cells and 3 X 10’ irradiated C57BL/6 spleen cells in the presence or absence of 0.1 pg indomethacin and/or 0.05 ml CSCS (batch No. 23). The cytotoxic activity on DBA/Z and C57BL/6 target cells was tested 5 days later and was expressed as percentage specific 5’Cr- release.

290 WOLF AND DROEGE
10
0
20
10
0
stim: 3 x lo5 A.TL + 2 x 105 C3H 0-0 3 X lo5 A.TH + 2 x 3 X lo5 A.TH
105 C3H o-o A-A
I
I I I I
i
C)ZX~O~LN
\
- cscs
Is
I
i+H p-8 1 I I 1 I I I I I 0 10-B 10-7 10-6 0 10-8 10-7 10-G
B)Sx104LN + cscs
Prostaglandin E2 (Mall I )
FIG. 3. The effect of prostaglandin Ez on the CTL response in the presence or absence of additional helper factor (CSCS). (A and C) 2 X lo4 or (B and D) 5 X lo4 C3H lymph node cells were incubated in 0.2 ml microcultures together with 0.1 sg indomethacin and the indicated irradiated stimulator spleen cells from A.TL (H-2KsIkD“), A.TH (K’I”D“), and C3H (KkIkDK) mice and the indicated concentrations of prostaglandin E2 in the presence (A and B) or absence (C and D) of 0.05 ml CSCS (batch No. 23). The cytotoxic activity on A.TL target cells (KSkDd) was tested 5 days later. The figure shows the mean and SE of usually four parallel determinants. Note that the cytotoxic responses were strongly suppressed by PGE2 even in the presence of CSCS (top), unless the cultures contained two sets of I-region incompatible irradiated spleen cells (A.TH + C3H). But also these cultures of I-region incompatible spleen cells were suppressed by PGEz if tested in the absence of CSCS (bottom).
stimulator cells (Fig. 4) were strongly suppressed by low concentrations of PGE2 even in the presence of CSCS. That this suppression was not the result of a general toxic effect of PGE2 on the cultures was indicated by the fact that the cultures were essentially insensitive to this suppressive effect when they contained a combination of irradiated I-region incompatible spleen cells such as C3H (H-2k) plus A.TH cells (H-2KSISDd) (Figs. 3 and 4, panels A and D). This was observed irrespective of whether the stimulator cell under test was I-region incompatible (Fig. 3) or com- patible (Fig. 4) to the responder cell. On the other hand, cultures with irradiated I- region incompatible cells were also suppressed by PGEI, when CSCS was omitted from the cultures (Figs. 3 and 4, panels C and D). Cultures without I-region incom-

REGULATION OF CYTOTOXIC RESPONSES BY PROSTAGLANDIN(S) 291
50-
40-
30
20.
% lo-
: 2 6 o-
L=l +z
% 50 s
40
30
20
10
0
stim.: TNP-C3H + A.TH 01 TNP-CM A-
A) 2 x lo4 LN + cscs
C)2 x 104LN -cscs
\ W-P--$
I, ! *I I I
0 10-B 10-7
B)2x104LN + cscs
Prostaglandin Ep (Mall I )
FIG. 4. The effect of prostaglandin E2 on the CTL response against haptenated syngeneic cells in the presence or absence of additional helper factor (CSCS). (A and C) 2 X lo4 or (B and D) 5 X lo4 C3H lymph node cells were incubated in 0.2 ml microcultures together with 3 X 10’ irradiated and TNP- haptenated C3H spleen cells either alone or in combination with 3 X IO5 irradiated A.TH spleen cells and in the presence (A and B) or absence (C and D) of 0.05 ml CSCS (batch No. 23). The cultures contained also 0. I pg indomethacin and the indicated concentrations of prostaglandin El. The cytotoxic activity on TNP-haptenated C3H target cells was tested 5 days later. The figure shows the mean and SE of usually four parallel determinants.
patible cells did not respond at all in the absence of CSCS. The helper activity of this CSCS batch had been titrated prior to these experiments, and the concentration of 0.05 ml/O.2 ml microculture was well within the plateau range of cytotoxic ac- tivity. This CSCS batch mediated substantial cytotoxic responses also in MHC com- patible systems, indicating that it contained sufficient amounts of interleukin 2 and of the late acting helper factor that has been described recently (17).

292 WOLF AND DROEGE
DISCUSSION
Our experiments provide direct evidence for a strong suppressive effect of PGE2 on cytotoxic responses. Such a suppressive effect on cytotoxic responses was sug- gested indirectly by several previous experimental observations (l-7). Previous ex- periments (4, 7) indicated in particular that PGE2 suppresses the pro&&on of interleukin 2, a helper factor which is usually produced by helper T cells and is required as a second signal for the activation of CTL (for review see (8, 9)).
The simple possibility that the inhibition of the cytotoxic responses was exclusively mediated by suppression of IL-2 production in the culture, however, was rendered unlikely. PGEr also showed a strong dose-dependent suppression in microculture systems which contained I-region compatible stimulator cells and concanavalin A- activated spleen cell supernatant (CSCS) as an external source of helper factor. These microculture systems generated no detectable cytotoxicity in the absence of external helper components, but produced plateau levels of cytotoxic activity with CSCS concentrations in the range of 0.025-o. 1 ml per microculture. It should be empha- sized that the effector phase of the cytotoxic T lymphocyte response is not affected by PGE, in concentrations up to 1 X lo-’ M (18) or higher ( 19), and the effector phase is reportedly also not enhanced by indomethacin (3).
Microcultures which contained CSCS and a combination of irradiated I-region incompatible spleen cells were largely resistent to the suppressive effect of PGE2, indicating that this suppression was not mediated by a general toxicity of the drug. The mechanism of this resistance is not known. Cultures with I-region incompatible cells produced cytotoxic responses in the absence of CSCS indicating that these cultures generated helper factor including IL-2 endogenously. But these cytotoxic responses in the absence of CX’S were again sensitive to the inhibitory effect of PGE*. One explanation for the resistence against suppression by PGE2 in the pres- ence of CSCS and I-region incompatible spleen cells is, therefore, that the mere quantity of endogenously produced plus exogenously added factor antagonizes the suppressive effect of PGEz. The antagonizing agent may be IL-2 or yet another factor which is produced after stimulation with allogeneic I-region determinants. There is clearly the possibility that the effect of IL-2 and PGE2 neutralizes one another on a quantitative basis. This conclusion is also supported by the observation of Baker et al. (4) that PGE2 reduced the proliferation and viability of a CSCS-dependent cytolytic T cell line; a PGE2 concentration of lop6 M was reported to reduce the thymidine incorporation and viability by about 50% and this effect was compensated by a fourfold increase of the TCGF concentration in the culture (4).
Our experiments did not allow us to distinguish whether the inhibitory effect was mediated by prostaglandin(s) directly or via the activation of inhibitory T cells as suggested by others (2, 20).
Our experiments with indomethacin confumed and extended those of Darrow and Tomar who reported that indomethacin enchanced cytotoxic responses against allogeneic stimulator cells in macrocultures (3). We found that the cytotoxic response of DBA/2 spleen cells against TNP-haptenated syngeneic stimulator cells in macro- cultures was minute if tested in the absence of indomethacin and CSCS; but this response was equally well enhanced by the addition of indomethacin or CSCS, and was not further enhanced by the combined addition of indomethacin plus CSCS. Similarly, the microculture response of C3H cells against allogeneic stimulator cells

REGULATION OF CYTOTOXIC RESPONSES BY PROSTAGLANDIN(S) 293
was significantly enhanced by indomethacin if tested in the absence but not if tested in the presence of CSCS. This group of experiments indicated, therefore, that the cytotoxic responses are usually regulated by endogenously produced prostaglan- din(s), and the endogenously produced amount of prostaglandins appeared to inhibit essentially only the endogenous production of helper factor or its effect.
ACKNOWLEDGMENTS
We are grateful to Mrs. S. Panknin and Mrs. H. Schmidt for expert technical assistance and to Mrs. A. Riedl for her assistance in the preparation of the manuscript.
REFERENCES
1. Goodwin, J. S., Bankhurst, A. D., and Messner, R. P., J. Exp. Med. 146, 17 19, 1977. 2. Webb, D. R., and Nowowiejski, M. D., Ceil Immunoi. 41, 72, 1978. 3. Darrow, T. L., and Tomar, R. H., Ceil. Immunoi. 56, 172, 1980. 4. Baker, P. E., Fahey, J. V., and Munck, A., Ceil. Immunoi. 61, 52, 1981. 5. Gemsa, D., Lymphokines 4, 335, 1981. 6. Bray, M. A., Immunoi. Today 80, 65, 1980. 7. Rappaport, R. S., and Dodge, G. R., J. Exp. Med. 155, 943, 1982. 8. Moller, G., Immunoi. Rev. 51, 1980. 9. Moller, G., Immunoi. Rev. 63, 1982.
10. Drijge, W., Reddehase, M., and Wolf, M., Ceil. Immunoi. 64, 53, 1981. 1 I. Galli, H. P., and Droge, W., Eur. J. Immunol. 10, 87, 1980. 12. Teh, H.-S., Harley, E., Phillips. R. A., and Miller, R. G., J. Immunol. 118, 1049, 1977. 13. Shearer, G. M., Eur. 1. Immunoi. 4, 527, 1974. 14. Vane, J. R., Nature New. Bioi. 231, 232, 1971. 15. Thomas, D. R., Philpott, G. W., and Jaffe, B. M., Exp. Ceil Res. 84, 40, 1974. 16. Gaut, Z. N., Baruth, H., Randall, L. O., Ashley, C., and Paulsrud, J. R., Prostaglandins 10, 59, 1975. 17. Reddehase, M., Suessmuth, W., Meyers, C., Falk, W., and Droege, W., J. Immunoi. 128, 61, 1982. 18. Henney, C. S., Boume, H. R., and Lichtenstein, L. M., J. Immunoi. 108, 1526, 1972. 19. Plaut, M., J. Immunoi. 123, 692, 1979. 20. Fischer, A., Durandy, A., and Griscelli, C., J. Immunol. 126, 1452, 1981.