information to users - university of toronto t-space · 2020. 4. 7. · against noradrenaline and...
TRANSCRIPT

INFORMATION TO USERS
This manuscript has been reproduœd from the microfilm master. UMI films the
text diredly from the original or copy submitted. Thus, some thesis and
dissertation copies are in typewriter face, h i l e others may be from any type of
cornputer printer.
The quality of this reproduction is dependent upon the quality of the copy
submitted. Broken or indistinct print, colored or poor quality illustrations and
photographs, pnnt bleedthrough, substandard rnargins, and improper alignment
can adversely affect reproduction. a
In the unlikely event that the author did not send UMI a complete manuscript and
there are missing pages, these will be noted. A h , if unauthorized copyright
material had to be removed, a note will indicate the deletion.
Ovenize ~ t e n a l s (e.g., maps, drawings, charts) are reproduœd by sedioning
the original, beginning at aie upper left-hand corner and continuing from left to
right in equal sections with small overlaps. Each original is also photographed in
one exposure and is induded in reduœd fom at the back of the book.
Photographs induded in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6" x 9' black and white photographie
prints are available for any photographs or illustrations appeafing in this copy for
an additional charge. Contact UMI diredly to order.
Bell & Howell Infornation and Leaming 300 North Zeeb Road, Ann Arbor, MI 481061 346 USA
800-521-0600


CONDUIT VERSUS RESISTANCE BLOOD VESSELS: ADRENOCEPTORS AND NlTRlC
OXIDE.
BY
Habib M. Razavi
A thesis submitted in confomiity with the requirements for the degree of Master of Science in the University of Toronto
01998 Copyright by K.M. Razavi

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services seMces bibliographiques
395 Wellington Street 395, rue Wellington Ottawa ON K1A ON4 Ottawa ON K I A ON4 Canada Cariada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or sel reproduire, prêter, distn'buer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/film, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts kom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

CONDUIT VERSUS RESISTANCE BLOOD VESSELS: ADRENOCEPTORS AND NlTRlC OXlDE
Habib M. Ratavi
Masters of Science, Department of Phamiacology
University of Toronto, 1998
Abstract
Local regulation of hemodynamic control is a complex rnechanism that
employs a number of biochemical processes (including adrenoceptor control and
Nitic Oxide (NO) modulation) depending on the vascular bed and function. The
nature of adrenocepton and nitric oxide modulation were, therefore, investigated
in two distinct types of blwd vessek with different function.
Third generation rat rnesenteric artenes (50 to 200pm diameter) and
descending thoracic aorta were selected as resistance and condul artenes
respectively. These were dissected and mounted in a microvessel chamber or a
standard (10ml) organ bath. The preparaüoons were allowed to equilibrate, and
Thereafter, a series of experiments were performed to examine adrenoceptor
charaderkation and the role played by nitnc oxide in the vascular response.
Specifically the effects of several antagonists were inspected against the agonist
response.
The results showed mat the rat rnesentefic artery and aorta behavd
differently. The a-adrenoceptor mediated contraction in the mesenteric artery
seemed to be of the aiA-adrenoceptor subtype. This conclusion was reached

based on the differences between the ECsoPs (ECI1s were: 29x1 05. 6 . 7 ~ 1 O-=,
8 . 1 ~ 1 0 ~ and 2.2x10-~ (M) for control 10, 30 and 100nM spiperone (are-acting)
against noradrenaline and 8 .5~1 O-', 1 .6~1 O&, 9 .4~1 and 1 . 7 ~ 1 os (M) for
control, 1 O,3O and 1 OOnM BMY 7378 (ai D-acting) against noradrenlaine,
respectively). In contrast, al! of the functionally characterized al-adrenoceptors
appear to be present and play a role in response to a-stimulation of the rat aorta.
This study also illustrated the potential presence of an atypical P-
adrenoceptor, which mediates vascular smooth muscle relaxation. This was
conduded in the aorta but not the mesentetic artery. In the aorta, relaxations
mediated by isoproterenol were not inhibited by propranolol. In the mesenteric
artery relaxations developed to isoproterenol could also not be inhibited by
propranolol. However the ECSo of this latter response was about 50 pM and
therefore not in the usual range of a typical P-rnediated response.
Finally, based on the effects of aminoguanidine and L-NNA, both iNOS
and cNOS sensitive relaxation components appeared to be operative in the
mesentenc artery. However the relaxation in the aorta was exclusively due to the
activation of cNOS.
In conclusion, these studies confirm that different blood vessels Vary
with respect to their mode of cardiovascular regulation, primarily through
differential biochernical pathways and distinct receptor populations (Le. NO
modulatory pathways and differential a and P-adrenoceptors).

I am indebted to the following people for the successful realization of this project:
Dr. Christine Forster, who has been an outstanding supervisor and mentor. Thank you for your guidance, and insight
Dr. Cecil Pace-Açciak, rny advisor. Thank you for monitoring the successful progress of my masters project.
My defense wmrnittee DR. L. Grupp, H. Van Toi and D. Osmond -1 thank you for your special interest in this project.
My close colleagues and friends: Mr. MS. Ng, Mr. A. Mendonca and Mr. H. Hamrahi.
I thank my friend Mr. Vince Mauurco for his skillful illustrations of the research apparatus.
I am forever grateful for the continued and constant support of my parents and my brother Majid.

Table of contents
CONTENTS
LlST OF FIGURES
LlST OF TABLES
LIST OF ABBREVIATIONS
ABSTRACT
1 .O Introduction
vi
viii
ix
1
1.1 Background
1.2 Receptors involved in cardiovascular regulation
1.2.1 Characterization of adrenoceptor subtypes
1.2.2 Classification of al-adrenoceptor subtypes
1 -2.2.1 Functional and classical characterization of
al-adrenocepton 7
1 2 2 . 2 Vascular al-adrenoceptors 9
1.2.3 Classification of cr~adrenoceptor subtypes 15
1.2.3.1 Functional and classical characterization of
aradrenoceptors 15
1 -2.32 Vascular aradrenoceptors 18
1.2.4 Classification of padrenoceptor su bty pes 22
1.2.4.1 Functional and classical characterization of
padrenocepton 22

1.2.4.2 Vascular p-adrenoceptors
1.3 Endothelial modulation of vascular responsiveness
1.3.1 Biotogy of nitric oxide
1.3.2 Synthesis of nitric oxide
1 -3.3 The regulatory role of NO in blood vessels
2.0 RATIONALE, OBJECTIVES AND HYPOTHESES
3.0 METHODOLOGY
3.1 Experimental approach
3.2 Tissue preparation
3.3 Experimental apparatus
3.3.1 Mesentek artery dissection and set up
3.3.1.1 Fine dissection
3.3.1.2 Mesenteric artery set up calibration
3.3.1.3 In vitro mesenteric set up
3.3.2 Aortic fine dissection and set up
3.3.2.1 Fine dissection
3.3.2.2 Aortic organ bath apparatus and calibration
3.3.3 Experimental protocol
3.3.3.1 Delineation of a-adrenoceptors in the srnall
rnesenteric artery versus the aorta
3.3.3.2 Delineation of fbadrenoceptors in the small
rnesenteric artery venus the aorta

3.3.3.3 Endothelial modulation of vascular
responsiveness
3.4 Drugs and solutions
3.5 Data analysis
4.0 RESULTS
4.1 Characteritation of the a-adrenoceptors on the srnall rat
mesentenc arteries versus the aorta
4.1.1 The effect of prazosin on noradrenaline-mediated
contractile response in the mesenteric artery versus
the aorta 67
4.1.2 The effect of prazosin on phenylephrine-mediated
contractile response in the mesentenc artery versus
the aorta 75
4.1.3 The effect of 5 MU on noradrenaline-mediated
contractile response in the mesenteric artery venus
the aorta
4.1.4 The effect of 5 MU on phenylephrine-mediated
contractile response in the mesenteric artery venus
the aorta 83
4.1.5 The effect of spiperone and BMY 7378 on
agonist-rnediated contractile response in the mesenteric
artery versus the aorta 86

4.2 Characterization of the P-adrenoceptors on the small rat
mesenteric arteries versus the aorta
4.3 Angiotensin-mediated contractile response in the rat
4.4 Endothelial modulation of vascular resistance in the small
mesentenc artery versus the aorta of the rat
4.4.1 The effect of L-NNA on the acetylcholine mediated
relaxation in the pre-constricted small mesenteric
artery versus the aorta
4.4.2 The effect of aminoguanidine on the acetylcholine
mediated relaxation in the preconstricted small
mesenteric artery venus the aorta 110
5.0 DISCUSSION 113
5.1 The effect of repeated administration of a-adrenoceptor agonists 1 13
5.2 a-Adrenoceptor heterogeneity in the third generation rat
mesentenc artery 117
5.2.1 The role of aradrenocepton in the third generation
rat mesenteric artery 137
5.2.2 The role of al-adrenoceptors in the third generation
rat mesenteric artery and aorta 117
5.2.3 p-Adrenoceptor responsiveness in rat aorta venus
the mesenteric artery 1 24
5.3 NMc oxide modulation of the vascular contradile responsiveness 126

6.0 FUTURE DIRECTIONS
6.1 Charaderkation of novel p-adrenoceptor-rnediated relaxation
in the rat mesenteric artery versus the aorta 131
6.2 Characteriration of iNOS on vascular location 131
6.3 Role for other endothelial-derived relaxing factors 132
7.0 REFERENCES 133

LIST OF FIGURES
Figure 3.1 Mesentetic artery apparatus
Figure 3.2 The mounting of the mesenteric artery
Figure 3.3 The aortic experimental apparatus
Figure 4.1 The effect of a-adrenoceptor agonists on the small mesenteric
a rtery
Figure 4.2 The effect of consecutive repeated administration in the small
Mesenteric artery and aorta
Figure 4.3 Effect of prazosin on the contractile response of noradrenaline
in the mesenteric artery versus the aoria
Figure 4.4 Effect of prazosin on the contractile response of phenylephrine
in the mesentenc artery versus the aorta
Figure 4.5 Effect of 5 MU on the contractile response of noradrenaline
in the mesenteric artery venus the aorta
Figure 4.6 Effect of 5 MU on the contractile response of phenylephrine
in the mesenteric artery versus the aorta
Figure 4.7 The effect of spiperone and BMY 7378 on contractile responses
to phenylephrine and noradrenaline in the mesenteric artery 87
Figure 4.8 Effect of spiperone on the contractile responses of noradrenaline
and phenylephrîne in the rat aorta 92
Figure 4.9 Effect of BMY 7378 on the contractile responses of noradrenaline
and phenylephtine in the rat aotta 95
Figure 4.10 Efiect of propranolol on the relaxation responses of

Figure 4.1 1
Figure 4.12
Figure 4.1 3
Figure 4.14
P-adrenoceptor agonists in pre-constricted rat rnesenteric artery 98
Effect of repeated administration on the relaxation responses of
p-adrenoceptor agonists in the KCI preconstricted rat aortic
rings 1 O0
Effect of propranolol on the relaxation responses of adrenaline
to the KCI pre-constricted rat sortic rings 1 03
Contrations mediated to noradrenaline and angiotensin II in the
rat mesenteric artery. 105
The effect of L-NNA on the acetylcholine-mediated relaxation to
a-agonist [1 -PM] preconstricted rat mesenteric artery and
aofta
The effect of aminoguanidine on the acetylcholine-mediate
relaxation to a-agonist [1 -PM] pre-constricted rat mesenteric
arterv and aorta 111
Figure 4.15
vii

LIST OF TABLES
Table 1.1
Table 4.1
Table 4.2
Table 4.3
Table 4.4
A summary of a and P-adrenoœptor subtypes 30
ECso values and maximum response for noradrenaline and
phenylephrine for a series of concentration-effect curves in absence
of any antagonist (Time-control study) 61
ECw, values and maximum data for noradrenaline in the presence
of increasing concentrations of a-adrenoceptor antagonists in the
mesenteric artery and aorta 72
ECSo values and maximum data for phenylephrine in the presence
of increasing concentrations of a-adrenoceptor antagonists in the
mesenteric artery and aorta 78
Mesenteric ECK, and maximum data for noradrenaline and
phenylephrine in the presenœ of increasing concentrations
of a-adrenoceptor antagonists BMY 7378 and spiperone 89

LIST OF ABBREVIATIONS
AC
ADP
a
P
CAMP
CEC
cDNA
cNOS
DAG
EDRF
EDHF
IUPHAR
NOS
1P3
L-NAME
LNMMA
LNNA
MDCK
NOS
5 MU
PGE
adenylyl cyclase
adenosine dip hosphate
alpha
beta
cyclic adenosine 3', 5' -monophosphate
chloroethylchlonidine
cloned deoxyri bonucleic acid
constitutive nitric oxide synthase
diacyl glycerol
endothelialdenved relaxing factor
endot helialderived h yperpola rizing factor
International union of Pharmacology
inducible nitric oxide synthase
inositol trisphosphate
La-Nitro-Arginine Methyl Ester
La-Nitro-Monometh yl-Arginine
Lm-N -Nitro-L-Arginine
Madin-Darby canine kidney
nitric oxide synthase
5 Methylurapdil
prostaglandin E

PG l2
PLA2
PLC
PL0
PKC
PKA
TBA
prostag landin l2
phospholipase A2
phospholipase C
Rhpspholipase D
protein kinase C
protein kinase A
tetrabutyl ammonium

1 .O INTRODUCTION
1.1 Background
Phamacological intervention by dnigs that are aimed to correct
cardiovascular rnalfunction, act at the level of the heart, vascular network or both.
A number of drugs (with the exception of those that regulate hemostasis)
act to regulate, stabilize andlor improve cardiac output (CO), which is the product
of the stroke volume (SV) and heart rate (HR). Stroke volume, is the volume of
the blood that is ejected out of the left ventride during systole and depends not
only on the contradile capacity of the heart but also, on the vascular fundion in
the perip hery . According ly , pre-load and after-load are important modulators of
the cardiac function (Ackermann, 1997).
Much cardiovascular research has been devoted to the physiology of
heart and to its phanacological modification. The pharmacology of vascular
networks [whilst having been widely studied (Edwards et al. 1995 and Fulton et
ai. 1995; Quast, 1996 a ~ d van Zwieten, 1996)] has not been poorly examined in
ternis of cardiovascular function and regulation.
The vascular network can be divided into three components, the conduit,
the resistance and the capadtance vessels. The spectnim where the different
parts of the vascular network give rise to other components is based on the
diameter of the vesse1 and the distinct anatomical attributes. The physiology
associated with different parts is also distinct. The conduit vessels, the iarger
artenes, are those that a d to direct large volumes of oxygenated blood from the

heart towards the vital organ systems (Nakamura et al. 1997, Mitchell et al.
1997). The resistance arteries represent a number of vesse1 types that give rise
to the artenoles and regulate blood flow and pressure (Goto et el. 1996, Kam et
al. 1996 and DeFily el al. 1995). Therefore the resistance vessels behave to
compromise the often-oonflicting hemodynamic needs of an organism versus
those of the individual organ systerns (Hall, et el. 1996).
Veins are capacitance vessels. These passively help to return
deoxygenated blood back to the right atrium. Although in times of stress and
exersice sympathetic (adrenergic) tone acts on the large veins to cause
contraction and aid venous return, for the most part these vessels act as a
venous pool.
The biochemical basis of cardiovascular control depends on many factors.
These include a variety of intracellular, message relaying mechanisms, that c m
all be targeted by phannacological tools. Cell to cell communication is achieved
via the interaction of an agonist (neurotransmitter or hormone) with specific
receptors that in tum, interad with guanine regulatory G-proteins with the
subsequent activation of an effedor. The following sections will intmduce the rote
that receptors play in the cardiovascular regulation.
1.2 Receptors involved in cardiovascular regulatiiüon
Regutation of blood Row and pressure is a muttifaceted and complex
mechanism. Therefore, understanding the degree of the local regulation by each
blood vesse1 type requires a thorough understanding of the various chernical,

neurohurnoral and physical modes that act to rnediate these processes. lt is
obvious that the various receptors that sense the physical stimulus of the shear
stress of flow or pressure (Le. stretch receptors) and those that mediate the
signals of the various biological ligands play an important role in the control
hemod ynamics.
Arguably one of the most important reg ulators of cardiovascular
homeostasis is the action of the adrenergic neurotransmitters (adrenaline and
noradrenaline) that stimulate adrenoceptors located on vascular smooth muscle
cells.
1.2.1 Characterization of the adrenoceptor subtypes
When stirnulated, adrenoceptors cause of a host of physiological events in
a wide variety of diHerent species and tissues. In this respect, agents that act on
these recepton c m modify the action of the heart and the vascular network.
Adrenoceptors have been characterized and categorized by their
pharmacological properties (Hieble et al. 1995) and more recently, their
molecular stnidure and signal transduction mechanisms (Hill and Sillenœ,
1 997).
Historîcally, Sir Henry Dale fiist descrÎbed the cc-adrenoœptor mediated
response via some elegantly designed experiments. He was the first to
demonstrate that ergot extracts inhibited the pressor effects of
sympathornimetics (Dale, 1 906). Later Ahlquist put funvard the notion of distinct
adrenoceptors Le. a vs. 8. This classification was based on analysis of

responses to a substance released from stimulation of the heart that was initially
described as adrenaline-like, but was not fuliy identified as noradrenaline until
1942 (VonEuler, 1942 and Ahlquist. 1948). Subsequently, Langer et al. proposed
the subdivision of the a-adrenocepton into distinct subtypes based on the
anatomical location: -postjunctional al and prejunctional aTadrenoceptors
(Langer 1974). The concept that al1 postjunctional a-adrenoceptors are of the al
type was refuted (Schimmel, 1976; Berthelsen and Pettinger 1977) and a 'se
called" functional classification of a-adrenocepton was put forward where
inhibitory responses were mediated by the a2 and excitatov responses were
mediated by the ai-adrenoceptors.
The classification of a family of receptors into subtypes is now based on
the rank order of potency of a number of agonists and antagonist for a given
receptor (Ruffolo, 1994). Hence three main adrenoœptor types namely the al, a2
and p adrenoceptors have been identified and characterizeâ (Bylund, 1988).
Hawevet each of these families are furthet subdivided. At least nine
pharmacologically distinct adrenoceptors have now been cloned and expressed,
the cloned, recombinant receptors having identical pharmacology to their
functional counterparts

1.2.2 Classification of the al-adrenoceptor subtypes
1.2.2.1 Functional and classical characterization of al -adrenoceptors
The al-adrenoceptor subtypes are related proteins, which are
heterogeneous and distinct. These receptors are transmembrane glycoproteins
spanning seven domains (Riek e l al. 1995) that couple G proteins to relay
biological signals to the cell interior (Strader et al. 1994 and Lismaa et al. 1994).
The generation of the second messeng ers diacylglycerol and inositol
trisphosphate are the means by which these receptors produce a cellular effect.
The majority of the data showing the heterogeneous nature of these receptors
has been accumulated fairly recently (Piascik el al. 1996; De Ponti et al. 1996;
Anfossi and Trovati, 1996; Galitzky et al. 1995). Through functional, radioligand
and biochemical studies, 1 was indicated that the various processes rnediated by
these receptors can not passibly be mediated by a single receptor type (Ruffolo
and Hieble, 1994).
Morrow and Cresse, first suggested the subdivision of al-adrenocepton
into the al* and a1~ -adrenoceptors (1 986). This was based on the differential
affinity of these subtypes for a variety of agents including oxymetazoline, WB
41 01 (2-[2,6-dimethoxyoxyphenoxyethyl]-arninomethyl-l, 4-benzodioxane) and
phentolamine. The alKadrenoceptor subtype was recognized as having the
higher affinity for these agents. Further evidence suppoiüng the existence of two
different adrenoceptor subtypes provided by Minneman's group (1 987). They
showed the aie , but not the al*, adrenoceptors can cornpletely be inactivated by
the irreversible alkylating agent, chlorethylclonidine (CEC) (Han et ai. 1987).

Likewise the irreversible inactivation of the al^ but not the alkadrenaceptor
subtype has also been demonstrated by a prazosin analogue azidoprazosin and
SZL-49- [l -(4amino-6, 7dimethoxy-2quinazdnyl-)4-(2-bicycJo [2,2,2] octa-2, 5-
dimethylcarbony1)-piperazinel (Terrnan et al. 1990; Piascik et al. 1989).
The doned a l d -adrenoceptor, which was thought to be a homologue of
the native al*-adrenoceptor is now believed to exist independently. It has
subsequently been demonstmted that the alD-adrenoœptor exists in a vanety of
tissues including vascular smooth muscle (Perez et al. 1991 and Lomasney et al.
1 991).
Many other ligands distinguish the nature of each of these al-
adrenoceptor subtypes. 5-Methylurapidil 15-MU], (+) niguldipine, methoxamine
and benoxathian are compounds that are recognized with high affinity for the a1~-
adrenoceptor binding sites (Graham et al. 1996). Spiperone, a classical D2
dopamine receptor antagonist, shows seledivity towards the UlB- subtype (Ford
et al. 1994). Finally, BMY 7378 [8-(2-[4-(2-methoxypheny1)-1 -piperazinyl] ethyl-û-
azasapirol [4,5] decane-7, 9dione dihydrochloride] and S W 1 05854((E)-a-[2-
butyl-1 - ((4carboxyphenyl) methyl)-1 K i m idazol-5-yl] met h ylene)-2-
thiophenopropoanoate) have been reported to have 50 to 100-fold selecüvity for
the alD-adrenoceptors compareci to the al^ and ai8-adten0~ept0rS (Hieble et al.
1995).
ln addition to the above dassification of the ciladrenoceptors, another
proposed charaderkation of al-adrenoceptors has been suggested. An atypical
a-adrenoceptor was suggested on various blood vessels wtiich demonstrates iow
8

affinity for prazosin (> 1 Nm, Flavahan and Vanhoutte, 1986). . This adrenoceptor
was classified as the aiL-adrenoceptor and belonged to a subfamily put forward
by Muramastu (1 995).
Whi le the functional anaiysis of adrenoceptor action can be directed
on any physiological system, the principal focus of this thesis is concerned
with the rote of adrenoceptors in the vascular network .
1.2.2.2 Vasculai a1 -adrenoceptors
The following sections systematically account for the presence of a1 -
adrenoceptors on three difFerent vesse1 types. Depending on the vascular bed,
the composition of the a1 -adrenoceptors are dflerent. Accordingly the conduit
(aorta), the resistance (3rd generation mesenteric artery) and capacitance (vein)
vessels are discussed.
AORTA
Initial functional studies using non-selective agonists and antagonists
showed that a1 -adrenoœpton seerned to represent an apparent homogeneous
population in human and rabbit aortae (Martinotti et al. 1991). More recently
Fagura and coworkers have confimed the heterogeneous nature of the ai - adrenoceptor on the rat aorta (1997). Using classical organ bath expenments
where the contractile response of phenylephrine was challenged by BMY 7378
and 5-MU revealed the phamacological classifcation of the a10 and a1~-
adrenoceptors as the rnost predominant subtypes in the rat aorta.

Using both functional and radioligand binding studies, Buckner et al.
(1 996) have shown a good correlation between the rank order of potency of
several agonists in mediating contraction in the rat aorta. and their binding profile
on cloned alD-adrenoceptors. These investigators showed that the rank order of
potency for the rat aorta was the same as that obtained for the binding affinity of
the rat cloned airadrenoceptor: noradrenaline > adrenaline s cirazoline >
phenylephrine > oxymetazoline > A-61603 > methoxamine. Correlation
coefficients corn paring rat aortic contraction (pA2) to binding (pK) were 0.09-0.21
for -adrenoceptors, 0.66 for cloned a l b and 0.94 for cloned arc
adrenoceptors (Buckner et al. 1996).
Two studies have shown a good correlation between the affinity estirnates
of the cloned alCadrenoceptor (rat 4broblast) and the pA2 value obtained for
BMY 7378 in the rat aorta (with the correlation coefficient constant r-1). In
contrast, the pA2 values obtained for spiperone and 5-MU had only a modest
correlation with their affinity estimates for the cloned art, and aiA-adrenoceptors
(Kenny et al. 1995 and Testa et al. 1995). Xu and coworken have recently used
solution hybridization to quantii mRNAs for a 1 ~ -, ale -and a10 -adrenocepton in
rat aortae. Although al1 three adrenoceptor subtypes were expressed, the
dominant expression of the ai D -adrenoceptors further support the ment finiding
that al^ -adrenocepton are major functional receptor subtype in the rat aorta
(Xu et al. 1 996).
ûther studies have shown that a le and a~~~adrenuceptor subtypes also
play a role in the contractile response to a-stimulation of rat aorta. In a separate

study Testa et al. (1 995) have shown a significant correlation beONeen the pA2
values of the al-adrenoceptor antagonists and their sensitivity for the cloned a l e
-adrenoceptor subtype, but not for the al D-subtype.
Furthemore, Van der Graaf and colleagues concluded that the contractile
response in rat aortic strips can be mediated by a coexisting population of al1
three al-adrenoceptor subty pes (1 996). This finding has been reiterated by other
techniques. For example, Hussain et al. contend that the contractile response
mediated to phenylephrine is only partly due to activation of alD-adrenoceptors
(1 997). Likewise using in situ hybridization histochemistry, radioligand binding,
and in vitro contractile studies on the rat aorta, Piascik's taboratory has shown
that both the a1A and al^-adrenoceptor subtypes are present. They have
effectively shown that three, possibly four, adrenoceptor subtypes play a role in
the rat aorta vasailar regulation (Piascik el al. 1994 and Vllalobos-Molina 1996).
From these studies, al1 three al- adrenoceptor subtypes mediate a
contractile response with the alD-adrenoceptor being dominant. However,
variation in the al-adrenoceptor population may exist between gender, age,
species and pathological circumstances should not be niled out.
MESENTERIC ARTERY
The resistance vessels are a major locus for hemodynamic regulation.
Some studies have found that resistance artenes are relatively more innervated
when compared with other vessel types (Stassen et al. 1998). This allows the
sympathetic nervous system to have a tighter and more inîtantaneous control of

these vessels. The extent of this regulation is dependent on the location of the
neurotransrnitter receptor (adrenoceptors) which when activated mediates
vascular reactivity (Stassen et ai.).
Evaluating the relationship between the sympathetic innewation and the
presence of al-adrenoceptors in the arterial tree of the rat Stassen et al. have
estimated the total al- and the subcomposition of alA-adrenoceptor in the third
generation rat mesenteric artery (1 998). These investigators have observed that
following chernical sympathactomy both the noradrenaline content of the
sympathetic nerves, and selective alA-adrenoceptor binding and ~ I A -
adrenoceptor mRNA content diminish( Stassen et ai.).
Other evidence which supports the contention, which the major al-
adrenoceptor subtypes in the small mesenteric arteries are of the a1~-
adrenoceptor subtypes was reported by lpsen and coworkers (1 997). These
authors investigated the ened of antipsychotic agent sertindole (adrenergic
antagonist) in rat resistance vessels and in membranes of HEK293 cells
transfected with ut-adrenoceptors (1 997). In mesenteric small arteries, it was
shown that seledive alA-adrenoceptor antagonists (5-MU and WB41 0 1 (2-(2,6-
di methoxyphenoxyet h yl) aminomethyl-1,4-benzodioxane)) inhibited
phen ylephrine responses with high affinity (pA2 9.1 and 9.5, respectively).
Chloroethylclonidine (arB- and aiD-adrenoceptor irrevenible antagonist) and
BMYï378 (8-[2-[4-(2-methoxyphenyl)-i -piperazînyl]ethyl]-8-azaspiro [4,5]
decane-7,Bdione dihydmchloride, -adrenoceptor revenible antagonist) had
leMe efFect. This indicated that the main adrenoœptor subtype in mesenteric

small arteries was of the al^ subtype. Furthemore, sertindole inhibited the
phenylephrine response of mesentetic srnall artenes (pA2 9.0), but had little
effect on the response of aorta to phenylephrine hence concluding it to be an
alA-antagonist (Ipsen et al. 1 997).
In contrast to these studies, others have shown that a le and the al^ -
adrenoceptors have a regulatory role in the rat small mesenteric arteries ( Han et
al. 1990 and Hussain et al. 1997). Based on functional studies in the small
rnesenteric artery it was shown that antagonist (SMU, WB 4101 and BMY7378)
affinity correlated highly with their previously published pA2 values in rat aorta
(alD) and less well with those for a1~- and alB-adrenoceptors mediating
contraction of the rat epididymal vas deferens and rat spleen, respectiveiy
(Hussain et al. 1997). They concluded that the contraction to phenylephrine of
mesenteric artery waç in part, via the ale- adrenocaptors.
Using chloroethylclonidine and nifedipine (ca2' channel inhibitor) Han and
coworken estimated the relative abundance of the al- adrenocepton in the small
mesenteric artery (1 990). They mncluded that both al^ and ale- adrenoceptors
were present in the srnall mesenteric arteries. This was based on the finding that
in these artenes there was a chloroethylclonidine and nifedipine sensitive
cornponent in contractions to phenylephrine.
Another study rules out the involvement of the a l~ , aie and ID-
adrenocepton in the mesenteric contractile response altogether (Van der Graaf
et ai. 1997). In ths study, the e W s of a series of al-adrenoceptor antagonists
on the phenyiephrîne-mediated contractions of rabbit isolated prostate, urethra,

trigone and mesenteric artery were investigated. They demonstrate a low
potency displayed by prazosin and HV723 (alpha-ethyl-3,4,5-tflmethoxy-alpha-
(3-((2-(2-rnethoxyphenoxy)eth yl)amino ) propy 1) benzene-acetonitfile fuma rate)
and suggest that the functional recepton in al1 four tissues belong to the ai'-
adrenoceptor class (1 997).
As evidenced from al1 of these studies, the rat mesenteric al-
adrenocpeton are probably heterogeneous. The intrajunctional receptors are
mainly the alA-adrenooeptor subtypes and these most likely, exert the greatest
degree of cardiovascular control.
MESENTERIC VEIN
Vascular control is attributed to the sympathetic nervous system, which
innervates the veins. For example Govyrin et al. have shown that the rat
mesenteflc reactivity is controlled by both humoral and neurogenic factors in the
mesenteric artery but only by neurogenic factors in the rnesenteric vein (1994).
In order to characterire the different ai-adrenoceptors in canine veins, a
study by Shi et al. looked at aie [3H] prazosin and [3H] rauwolscine binding in
plasmaiemma-enriched microsomal fractions isolated from the saphenous and
mesenteric veins (1989). They demonstrated that the microsomal membranes of
dog saphenous and mesenteric vein contained about a fourfold higher density of
the high affinity [3H] rauwolscine binding sites than those for [3H] prazosin
binding. Therefore canine veins seem to contain more a-drenoceptor subtypes
compared to al-adrenocptor subtypes. In a similar study ltoh et al. have

characterized the postsy naptic a-adrenaceptors in isolatecl canine mesenteric
arterial and venous preparations (1987). These investigaton showed that
phenylephrine was a more potent agonist in the mesentenc artery than in the
mesenteric vein and that UK-14,304 (~~adrenoceptor agonist) exhibited the
opposite profile of activity. They also showed that rauwolscine (aradrenoceptor
antagonist) was more potent than prazosin in the mesenteric vein demonstrating
that the more prevalent adrenoceptor was the ~~adrenoceptor.
In a related study based on functional studies of phenylephrine-induced
contractions and binding interactions of [3H]-prazosin wit h prazosin, WB 4 1 0 1, 5
rnethylurapidil, BMY 7378, SK&F 105854 and chloroethylclonidine, Daniel and
coworkers conclude that a~-adrenoceptors in the dog rnesenteric vein resembled
the al D-adrenoceptor su btype (1 997).
Therefore, accorùing to these studies, the dominant a-adrenoceptor is not
the ai-adrenoceptor subtype but is the aradrenoceptor subtype. However the
al-adrenoceptor does exist in the mesenteric vein and is most probably the a1~-
adrenoce ptor subtype.
1 .2.3 Classification of the aradrenoceptor subtypes
1 2.3.1 Functional and classical characterization of a2 -adrenoceptors
The phamacology of the aradrenocepton has wtablished that they are a
group of heterogeneous but related receptor proteins (Bylund, et al. 1988 and
Lomasney et al. 1091). Evidence perîaining to the heterogeneous nature of the
~~adrenoceptors was put forward by radioligand studies of a number of agonists

and antagonists. These results were further corroborated by functional studies in
cell lines and more recently by transfections of the ~~adrenoceptor genes in a
number of eukaryotic recombinant systems (Bylund, et ai. 1988 and 1994;
Lomasney et al. 1991).
The work of Bylund and associates has been quite instrumental, in
determining the nature of the ~~adrenoceptors (1995). In a series of
experiments, Bylund et al., investigated the ability of prazosin to inhibit [ 3 ~ 1-
yohimbine binding in rat and human brains. They revealed two discrete regions
of heterogeneity. One with a low prazosin binding affinity, which had a
pharmacological idently similar to the receptor found in human platelets. The
other binding site displayed a higher affinity to prazosin ( p k 7.5) which was
similar to a binding site identified in the neonatal rat lung. The aracirenoceptors
subtype that is found on the human platelet and human colonic adenocarcinoma
cells (HT29 cells) that displays a low affinity for prazosin and ARC-239 ([-2-
(2,4[0-methoxyphenyll-piperazine)-1-yl] and spiroxatrine) with a high affinity for
yohimbine is believed to be the azpiadrenooaptor subtype (Rufollo el al. 1994;
Bylund, et al. 1988). In wntrast, the adrenoceptor that is found on the rat kidney
and the hybrîd neuroblstorna-glioma cells (NG108-15) is the aZ8-adranoceptor
subtype (Rufollo et al. 1994 and Michel et al. 1990; Bylund et al 1985; Bylund et
al. 1987). In addition, oxyrnetazoline, benoxathian and antagonists WB 4101 and
BRL 44408 [2-(2H-(1 -methyl-1 , 3-dihydroisoindoie)rnethyi)-4, Sdihydro-
imidazoie] selectively interad with the au-adrenoceptor, whereas the a m

adrenoceptor subtype shows selectivity for, chlorpromazine, imiloxan , ARC-239.
(Rufollo, et el. 1994).
A third aradrenoceptors subtype has been identifieci in the opossum
kidney cells, and in the human neuroblastorna cell line Y 79 (Rufollo et al. 1994;
Bylund et al. 1995). This receptor, similar to the azB-adrenoceptor subtype, has a
high affinity for prazosin. The ratio of the affinities for prazosin and yohimbine for
this receptor however, is intemediate to the aa and ~2~adrenoceptors. This
receptor subtype was consequently temed the azc-adrenoceptor (Murphy, et al.
1988; Rufollo, 1994 and Bylund et a1.1995). A fourth aradrenoceptor subtype,
namely the a2D-adrenoceptor, has k e n described in the bovine pineal, rat
subrnaxillary glands and a cell line derived from a rat pancreatic islet tumor ( RIN
m5F). This receptor like the am subtype has a low affinity for prazosin and even
a lower affinity for [ 3 ~ 1-rauwolscine, when cornpared to the other subtypes.
In addition studies aimed to distinguish the pre- venus. post-junctional
~~adrenoceptors have been useful in that they strengthen and give credence to
the fadual existence of this subtype. Therefore a receptor that was resistant to
SK&F104078 ((E)-a-([2-butyl-l-((4-~arboxyphenyl)methyl~l H-imidazolb-
ylmethelyne)-2-thiphenepropanoate and SWFI 04856 [9-[(3-mehtyl-Zbutenyl-
) 0 ~ ] 6 c h l o r o - 3 m e t h y I - 2 , 3 , 4 , 5 , t e t m h y d r ~ was identifid and was
presumed to be the putative a2D-adrenoceptor subtype (Ruffolo, et al. 1987;
Murphy and Bylund 1988). SK8rF104078 and SKBrF104856 will biock the ar
adrenoœptor mediated contracüons of the postjundional canine, hurnan and

rabbit saphenous vein but not the prejunctional neuroinhibitory actions ar
adrenoceptor in atrial or vascular preparations (Hieble et al. 1986; Hieble el al.
1988). Therefore some suggested that the prejunctional aradrenoceptors are
probably exclusively the ~~~-adrenoceptor subtype. However, it was
subsequently demonstrated that the prejunctional a2-adrenoceptor in the rat vas
deferens was inhibited by SKtkF104078, and that the classification of the pre vs.
post-junctional adrenoceptors basad on selectivity for these antagonists was
therefore inadequate (Connaughton et ai. 1989).
Recentty, there has been interest in explaining the centrally activated
cardiovascular versus the analgesic effects of clonidine. Historically, clonidine
mediated al1 of its e W s via aradrenoceptors. However, numerous studies
aimed to delineate the centrally mediated hypdensive venus analgesic effects of
clonidine, pointed towards binding of clonidine to two distinct binding sites
(Head, 1995; Codd et el. 1995). In the medula oblongata, analgesia is facilitated
by binding of clonidine to a putative non-adrenergic irnidazoline receptor (Codd et
al. 1995). These recaptors were tenned irnidazoline 'prefemng 'site and are
found in the brain region which was typically devoid of traditional a2-
adrenoceptors (Bosquet et al. 1984; French, 1995).
1.2.3.2 Vascular a2- adrenoceptor
The major mode of the ar adrenoceptor mediated cardiovascular
regulation is via their regulation of the nucleus tractus solitarius and rostrai
ventrolateral medulla that a d to modulate barorefiexes hence adjusting

hemod ynamic variables (El-Mas et al. 1 997). Postjunctional pen pheral a2-
adrenoceptors, do exist and have various functions depending on the vessel type
and location on the vessel (endothelial œll versus the vascular smooth muscle
cell).
AORTA
At least two separate ar adrenoceptor sensitive cornponents have been
characterizad in the rat aorta. Matsuda and colleagues have shown that the
constitutive nitflc oxide synthase inhibitor L-NNA potentiated the contraction
induced by noradrenaline in the rat aorta depicting a noradrenaline mediated
relaxant component via the endothelium (1995). These investigaton also show
that in the presence of L-NNA. a& adrenocepton-agonists, donidine and UK-
14304 induced a dose-dependent contraction that was inhibited by yohimbine.
They concluded that in the rat aorta the ar adrenoceptors on the vascular
smooth musde and the endothelium are different in that one is coupled to
smooth muscle contraction and the other to EDRF production foilowed by
vascular relaxation.
In a similar study, on the rat aorta, Kaneko et ai. have investigated the
involvement of endothelium-derived nitnc oxide in the depressant action of the
endothelium on noradrenaline-induced contractions. They also charaderized the
recepton involved in the release of nitrÎc oxide (1993). Here they showed that in
the rat aorta the noradrenaline-induced contradion was significantly potentiated
by endothelium removal and in the presence of NO-nitro-Larginine (L-NNA) or

NO-monomethyl-L-arginine (L-NMMA), likewise demonstrating an a-
adrenoceptor mediated component in the endothelial production of the nittic
oxide and vascular relaxation. Furthemore they have also demonstrated that
clonidine wuld induce contraction only in endotheliumdenuded preparations or
in the presence of L-NNA hence proving that the involvement of an ar
adrenoceptor in the vascular smooth muscle contraction. In this study however, it
was also shown that the potentiating action of L-NNA on noradrenaline-induced
contractions could be observed in the presence of yohimbine or rauwolscine
conceivably demonstrating an involvement of the al-adrenoceptors in the
production of endothelial derived nitric oxide and vascular relaxation (Kaneko et
al.)
Frorn these studies it can be seen that in the rat aorta, the ar
adrenoceptors exist both on the endotheliurn and the vascular smooth muscle
cells where they mediate opposite and rnasking efFects. Therefore the stimulation
of the endothelial form produces the formation of nitric oxide followed by vascular
relaxation and the stimulation of the smooth muscle forrn causes direct
contraction.
MESENTER1C ARTERY AND VElN
Several studies have investigated the mle of postjunctional ar
adrenoceptors in the mesentenc vascular bed. For example Kong et al. have
examined the nature of aradrenoceptors in the rat isolated, pemised mesenteric
vascular beâs. They observed that under physiological preqsures of 60 mmHg

the infusion of the aradrenoceptor agonist donidine (3-100nM) has no effect on
the perfusion pressure (1 991). However, they showed that in the presence of an
elevated pressure caused by constant infusions of noradrenaline (6-20 PM) and
bolus injections of clonKline (O. 1-10 nM) caused a dose-related decrease in
perfusion pressure. The mechanism for this was thought to be due to the
presence of aradrenocepton loated on the endothelium. However, destruction
of the endothelium by methylene blue or reactive oxygen radicals did not
attenuate the depressor action of clonidine.
In a similar study, Kannan et. al. (1 986) have looked at the mechanical
responses to penvascular nerve stimulation in rat supenor mesenteric artefies.
They observed that frequencydependent contractions to noradrenaline were
unaffected by the arselective adrenoceptor antagonist yohimbine, but were
rnarkedly attenuated by donidine, the arselective adfenoceptor agonist. The
effect of clonidine was attributed to endothelial production of nRric oxide. Similarly
Neilsen and coworken have looked at mesenteric resistanœ arteries (intemal
diameter 174-337 microns) h m rats, rabbits, pigs, and humans (1991). Using
the selective aradrenoceptor agonist BHT933 they demonstrated that functional
postjunctional a~drenocepton appear to be present in porcine and human
vessels (where they mediate contraction), but not the rat and rabbit vessels.
However in a prior investigation 1 was shown in the rat mesenteric artery,
that nanornola r concentrations of U K44,304 (aradrenoceptor selective
antagonist) could displace [3H] yohimbine implying the presence of ar
adrenocepton (Agrawal et al. 1985).

More research is required to delineate aie ~~adrenoceptor subtypes that
mediate vascular reactivrty in distinct blood vessels and the role played by a2-
adrenoceptors located on the endothelium and the vascular smooth muscle cells.
1.2.4 Classification of p-adrenoceptor subtypes
1.2.4.1 Functional and ciassical characterkation of p -adrenoceptors
The existence of vascular P-adrenoceptors was presented in 1957 when
the partial-agonist dichloroisoproterenot, antagonized P but not a-mediated
responses in the rat systemic circulation. (Powell and Slater, 1957; Moran and
Perkins, 1958). Sub-classification of the Padrenoceptors was suggeçted by
Lands and associates (1967) by comparing the rank order of potency of a
number agonists, they concluded that these reœpton could be subdivided into
two subtypes namely Pl and Pradrenocepton. The Pl subtype as the dominant
subtype in the heart and adipose tissue while the Pradrenoceptor subtype
produced relaxation of the vascular, utenne and airway smooth muscle. In
addition, the & adrenoceptor subtype was shown to be much less sensitive to
noradrenaline vis-&vis adrenaline (which displays up to a 100-fold seledivity for
the pradrenoceptor subtype; Hieble and Ruffolo, 1995).
Further ment evidence, suggested the existence of an additional p-
adrenoceptor which was insensitive to inhibition by classical Pi and Pr
adrenoceptor antagonists (Arch, et a1 1984; Wilson, et al. 1984). This receptor
was temed the 'atypicar P-adrenoceptor. The recombinant characterization of
this reœptor has now been daracterized and selective agonists have been

pmduced. This receptor is now known as the p3-adrenoceptor (Arch et al. 1984;
Wilson et al. 1 984).
Bot h adrenaline and noradrenaline activate the three p-adrenoceptor
subtypes (Alexander 1997). In some tissues and systems noradrenaline seems
to be more potent at the P3-adrenoceptor when compared with adrenaline
(Giacobino, 1995 and Emorine 1994). lsoproterenol activates al1 of the three
subtypes without any consistent selectivity of one over the other (Ruffolo and
Hieble 1 995).
There now exkt a number of agents, which specifically show selecüvity for
one P-adrenoceptor subtype over another. In this manner the agonist procaterol
selectively activates the PTadrenoceptor subtype (Pittman and Molinoff, 1983;
Pittman, et al. 1984 ; Wadlek et al. 1986; Hdberg and Mattssson, 1981). In a
similar manner, several agonists show specificity at the pl-adrenoceptor over the
other subtypes. Some of these rnost selective agents are R0363[ (-)-1-(3,4-
dimethoxy-phenethylamin0-)-3-(3,4dihydro~heno~)-2-pmpanol) oxalate]
(Lakovidis, et al. 1980), denopamine (Naito, et al. 1985) and xamoterol (Nuttall
and Snow, 1982).
In addition, several agents have been identified that inhibit each of the P
adrenoceptors individually. The most common agents used are the compounds
(-) biso prolol and CGP207 1 2A [2-h yd roxy-b (2-[(Z-hydroxy-3- (4-[ l meth y-
4triRurornethyl2imidazolyl] phenoxy) propyl) amino] ethoxy) benzamide], which
have k ' s in the nanomolar range at the pi-adrenocepton. The plipz -selactivity
ratio for these two antagonists are 100 to 1 (Kaumann and Lemoine, 1985) in the

case of the former and 10000 to 1 for the latter (Dooley, et ai. 1986; Schliep and
Harting, 19û4). In a similar manner ICI1 18, 551 ((2)-1-(2,3[dihydro-7-methyl-1 H -
inden-rl-yl] oxy)-3-([l -methyleth yl]-amino)-2-butanol] is a potent Pradrenoceptor
antagonist with a Kg l e s than 1 nM and a pup1 ratio of over 100-fold (Bilski et a/.
1983, Lemoine et al. 1985).
The P3-adrenoœptor mediated lipolysis has been characterized with high
selectivity by novel agon ists such as, BRL 35 1 35 [(R*, Re)- (+/-)-methyl 4-[2-[2-
hydroxy-2 (Sthlrophenyl) ethylamino] propyl] phenoxacetatehydrobrornide), BRL
37344 [sodium 4-(2-[Z-hydro (3chlorophenyl) ethylaminno] propyl)
phenoxyacetate] and carazolol (Wilson et al. 1984 Hollenga and Zaagsma,
1989). While this marked the first characteriration of this novel receptor this
receptor has also been identified in other tissues. Manara et al. (1995) for
example have demonstrated the abolition of the relaxation response of the rat
proximal colon isoproterenol by, SR 588994A (N- (2dimethylaminoethyI)-N- (3-
pyridinylmethyl)[4-(2,4,6-tnisopropylpheyl) thiazol4yl] amine) bupranolol and
SR 59230A [3-(-2ethylphenoxy)-19[1 SI-1 ,2,3,4-tetrahydronapth-l -ylarnino)Qs-
propanol oxalate]. Furthemiore in this study; by cornparhg the lCso values of SR
58894A and the non- selective p blocker propranolol, one obsewed a 77-fold
selectivity for inhibition by SR58894A.
In the heart. the BI and PTadrenocepton not only increase the force of
contraction during systole (ionotropy) but also increase the force of cardiac
muscie relaxation [Lucitropy] (Kaumann, 1997). These events oaxir by coupling
these adrenoceptors to the adenylyl cyclase via Gs. P-drenoceptor -selective

agonists, shorten the cardiac action potential and muse cardiodepression,
suggesting direct coupling of this receptor to a K' channel (Kaurnann 1997 and
Gauthier, 1995 and 1996). Most recently, cardiostirnulant effects of non-
conventional partial agonists (which are related to pindolol, alprenolol and
oxprenolol; Kaumann 1973 and 1989, Kaumann and Blinks 1980b) has been
attributed to a putative cardiac p4-adrenoceptor subtype. Although this distinct
adrenoceptor has yet to be doned it appears to be coupleâ positively to a cyclic
AM?-dependent cascade and seems to undergo soma desensitization (Kaumann
et al. 1997).
1.2.4.2 Vascular B-adrenoceptors
Endogenous adrenaline and noradrenaline act on vascular P-
adrenocepton to produce vasorelaxation. p-Adrenoœpton stimulate the G.
family of G proteins that in tum stimulate adenylyl cyclase and elevate W P
concentrations. This second messenget then activates the CAMP dependent
protein kinase followed by phophorylation of intracellular proteins and relaxation
in blood vessels. The following sections will examine the role of vascular P-
adrenocepton
AORTA
It has been shown in both endothelium denuded and intact phenylephrine
pre-constricted rat aorta that P-adrenergic agonists terbutaline and dobutamine
cause a conœntration dependent vasorelaxation (Guney et al. 1998). In a similar

study, Zheng et al. have shown that NO-nitro-~oarginine meth ylester (L-NAME) ,
an inhibitor of NO synthase, partially inhibited endotheliumdependent relaxations
that were evokeû in phenylephrine-prcxonstnded fings by isoproterenol. They
also showed that L-NAME abolished relaxations mediated by forskolin,
suggesting that p-adrenocepton or any agent, which raises CAMP also, elevates
nitric oxide release from endothelial cells causing relaxation (1995). In an almost
identical study that investigated the importance of the endothelium in the
relaxation of isolated rat aorta caused by a P-adrenoceptor agonist, it was shown
that mechanical removal of the endothelium attenuated the relaxation induced by
isoproterenol. Endothelial denudation however, did not affect the relaxation
produceâ by either forskolin or sodium nitroprusside (Kamata et al. 1989).
Furthemore, mechanical removal of the endotheliurn or treatment with
methylene blue enhanced the maximal contraction induced by isoproterenol a
response that was antagonized phentolamine. The authors concluded that
isoproterenol-induced relaxation of endothelial intact aortic strips is mediated by
f3-adrenocepton located on both the endotheliurn and the smooth muscle, and
that high concentrations of isoproterenol produces an increase in the resting
tension through a-adrenoceptors (Kamata et al.).
In a study exploring the mechanism of relaxation by forskolin in rabbit
aortic rings, relaxant effectç of isoproterenol are also desctibed and are attributed
to the Bradmnoceptor stimulation (Satake et a!. 1997). In this study, fonkolin
potentiated the relaxing response to isoproterenol. Furthemore, the potentiating
e W of forskolin was shown to be inhibited by propranolol but not by methylene

blue. Also the relaxing response to terbutaline, a Pfldrenoceptor agonist, but not
lower concentrations of dobutsrnine, a Pl-adrenoœptor agonist, was also
potentiated by fonkolin. The authors concluded that in rabbit aortic rings forskolin
causes the apparent potentiation of isoproterenol-induced relaxation (as a resuit
of forskolin-induced increase in the b e l of CAMP) via modulation of the Pr
adrenoceptor activity.
More research in the rat aorta has shown that the putative p-
adrenoceptors involved in the relaxant response are not only coupled to
endothelial production of nitric oxide, but that this process may also involve the
cytochrome P-450 system (Iranami et al. 1996 and Satake et al. 1997). For
example Satake et al. showed that in the aorta pretreated with ICI-1 18, 551(a
selective &adrenoceptor antagonist) or atenolol (Pl-antagonist), the residual
relaxing respunse to isoproterenol was inhibited by cytochrome P450
monoxygenase in hibiton, 2-methyl-l , 2-di-3-pyridyl-1 -propanone (metyrapone),
alpha-naphthoflavone or 8-methoxypsoralen (1 997).
These studies thusfar disclose the involvernent of Pl - and the Pr
adrenoceptors in the rat aortic relaxant response. In a study by Oriowo, it has
been shown that nanomolar concentrations of isoproterenol, CGP 121 77 and
BRL 37344 (both P3-adrenoceptor agonists) al1 relax phenylephrine pre-
contracted rat thoracic aorta (1 995). As the rank order of potency in the thoracic
aorta was isoproterenol = CGP 12177 > BRL 37344 Oriowo concluded that there
was probably more than one p-adrenocepton one of which was atypical (1 995).

It is evident that al1 three p-adrenoceptor subtypes exist on the rat aorta
and that they mediate vasorelxation. This response is coupled with the
production of nitric oxide on the endothelium. However, the precise elucidation of
the mechanisrn of the p-adrenoceptor mediated vasorelxation on the vascular
smooth muscle awaits further research.
MESENTERIC ARTERY AND VElN
The rat mesenteric atiery acts much the same way as the aorta where the
endothelial p-adrenocepton cause the production of nitric oxide and
vasorelaxation (Huang et al. 1998). Therefore it was shown by Huang et al. that
isoproterenol and fenoterol concentration-dependently relaxed the
phenylephnne-preçonstncted endothelium-intact mesenteric arteries and this
response was abolished upon the removal of the endothelium (1998).
Furthemiore thesr investigators showed that the activation of
tetrapentylamrnonium-sensitive K* channels contributes toward the relaxations
mediated through P- and pTadrenoceptor stimulation in rat mesenteric arteries.
This was because the relaxant responses to the P-adrenoceptor agonists were
abolished by 5pM (f channel blocker) tetmpentylammonium (Huang el al.
1 998).
Randall and associates (1995) have asseseci the involvement of ATP-
sensitive potassium channels in the vasorelaxant responses to P-adrenoceptor
agonists. The result of their investigation showed that the vasorelaxant potencies
of the non-selective p-adrenoceptor agonist, isoproterenol, the Pl-adrenoceptor

agon ist, dobutamine and the Pradrenoceptor agonist, terbutaline were al1
significantly (P < 0.05) reduced in the presence of the AT? sensitive potassium-
channel blocker, glibenclamide. Based on these results they wncluded that in
the rat mesenteric artery vasorelxation mediated by both the pl-and Br
adrenoœpton is coupled by a hyperpolanzation of the vascular smooth muscle
via the ATP sensitive potassium channels.
Looking at the venous cornponent of the rat mesentetic bed, Martinet-
Cuesta concluded that a PTadrenoceptor mediated vasorelaxation is present and
this response is attenuated in the portal-hypertensive rats (1 996).
In contrast some other studies in the dog mesenteric veins have shown
that the prejunctional (and not postjunctional) p-adrenoceptors may be invotved
in modulation of noradrenaline release. For example their stimulation c m lead to
increased venous contraction via the postjunctional venous a-adrenoceptors
(Seki et ai. 1989 and Seki et al. 1990).
80th Pi- and Pradrenocepton seem to exist on the mesenteric artery and
mediate a relaxation response. However more research is required to elucidate
the nature of padrenoceptors on the mesenterk veins.
Table 1.1 summarizes adrenoceptor su btypes, seledive agonists,
antagonists and their second messenger products.

Table 1.1 A summary of a and Badrenoceptor subtypes
The non-selective agonists and the more selective agonist are tabulated for each adrenoceptor subtype. The antagonists that are listed are al1 selecüve agents that specifically inhibit, the respective adrenoceptor subtype. Azidoprazosin and bchloroethylclondine are alkylating agents that irreversibly inhibit the crin and the a1~- adrenoceptor subtype respective1 y. The signal transduction machinery along with the relevant G-proteins for each adrenoœptor is also listed. Abbreviations not previously mentioned in the text : A61603: N - ( w , 5-dihydro- I H-imidazole-2-y]-2-hydmxy-5,6,7,8- tetahydmaphthalen- 1 -yl)methanesul phoamîde hydmbnnide A H 7 1 f iOA: l -(Biphenyl-2-yloxy)-4-imino-4-pipendin- 1-y/-butan-2-O/ RS i 7053: n N- [2-(2-cycIopmpy~methuxyphenoxy) ethy&-5chloro- a, adimethyl- I Hindole-3-ethanamide SNA P 5272: 5canboxamide-2, ôdiethyl- 1, edihydro-3-[-K( 3-[4- hydm~y-4-phenylpipe~din-y~pmpyI)] ca&oxamid@(#-nifrophenyI)

Table 1.1 ADRENOCEPOTR
FüNCflONAL CLONED
Noradrenaline Adrenaline Dopami ne Phenylephrine Methoxamine A61 6603 (Seiective) Noradrenaline Adrenaline Dopamine Phenylephrine Adrenaline Noradrenaline Phenylephrine Noradrenaline
Adrenaline Clonidine BHT920
(+) Niguidipine 5-Methylurapidil SNAP5272 RS7 7053 Aadoprazosin '
IPdDAG and GqIl1 ca2' iflux
Oxyrnetazoline (Selective)
Noradrenaline Prazosin(7.5) Adrenaline ARC239 (8.0) Clonidine BHT920
Adrenaline Prazosin(7.5) Noradrenaline ARC239 (8.0)
Noradrenaline Betaxolol Adrenaline Atenolol lsoproterenol CGP20712A cl-Dobutamine Xamoterol (Selective) Noradrenaline ici1 18551 Adrenaline Sai butamol Procaterd (Seledive)
Noradrenaline Bupranolol Adrenaline SR59230A BRL 37344(Selective)
Gilo

1.3 Endothelial modulation of vascular responsiveness
At least three endothelium derived relaxing agents have been implicated
in affecüng the biology of underiying vascular smooth muscle. These are EDRF
(endot helium derived relaxi ng factor) or n i tk oxide, prostacyclin (Pei2) and
EDHF (endothelium derived hyperpolariring factor). Al1 three produce vascular
relaxation in preanstricted vessels albeit via different mechanisms (Vanhoutte
et al. 1997). Endothelium detived hyperpolarizing factor hyperpolarizes the
vascular srnooth muscle cells by directly stimulating the tetra butyl ammonium
(TBA) sensitive K' channels. Prostacydin activates the vascular smooth muscle
adenylyl cylcase causing the production of CAMP leading activation of the
glibendamide sensitive K' channels and hyperpolarization (Parkington et al.
1 996).
Although nitric oxide also has a putative hyperpolarking role in vascular
relaxation, it is more importantly a direct vasodilator (Vanhoutte et al. 1997).
1.3.1 Biology nitric oxide
In 1980 Furchgot and Zawadzki describecl that the relaxant response of
isolated blood vessels by acetylcholine was dependent on an intact endothelium
(Furchgot et al. 1980). Nitrîc oxide was later identified as one of the finai, if not
the most important, mediators of the 'so-clalled' endothelial derîved relaxing ,
pathway (Palmer et al. 1987).
Although NO possesses an unquestionable chernical sirnplicity, the range
and complexity of its biological actions are only now emerging. Involvement of

NO in a range of physiological and pathophysiological systems has been
desctibed. These indude a major regulatory role in the cardiovascular system
(Alastair, et al. 1 994).
1.3.2 Synthesis of NO
Nitric oxide is pruduced from the amino acid L-arginine via nitric oxide
synthase (NOS). This results from the cleavage of one of the two terminal
guanidino nitmgen atorns, and the incorporation of an oxygen atom from
molecular oxygen to produce the enzyrnatic by-products NO and Lcitnilline
(Moncada et al. 1991; Leone et al. 1991). Nitric oxide is very short-lived with a
half-life of -6 seconds and hence, its actions are highly localised (Alastair, et ai.
1 994).
The production of NO is mediated via hivo isoforms of a dioxygenase
flavoprotein namely constitutive nitric oxide synthase (cNOS) and cytokine
inducible nlric oxide synthase (iNOS), in a vanety of cell types (Forstennann, et
al. 1 991 ; White and Marietta 1 992).
Constitutive NOS is mainly expressed in endothelial cells, neurons and
platelets (Sessa et al. 1993) and is ~a~'/calmodulin dependent for its short-lived
activity (8redt et al. 1990). lnducible NOS is the prominent NOS subtype in the
cells of the immune system. For example it exists both in neutrophils and
macrophages, is ca2'-independent for its actions which are associated with long
term production of NO. This leads to cytotoxiclcytostatic mechanisms against
foreign andor turnour cells (Hibbs et al. 1988). Moreover, the pathology

associated with a host of disorders is also mediated through the induction of
iNOS and prolonged synthesis NO in an array of organ systems (Corbett et al.
1992 and Radomski et el. 1990).
The two NOS isoforms are differentially inhibited. Accordingly Larginine
analogues such as Na-monomethyl-Larginine (L-NMMA) and Nm-nitro-L-arginine
methyl ester (L-NAME) and Na -Nlro-L-arginine (L-NNA) preferentially inhibit the
cNOS isofom( Kilbourn et el. 1990; Sakuma et al. 1988). Conversely
aminoguanidine is about 40-fold more potent at inhibiting the iNOS compared
with cNOS (Gnffiths et al. 1993).
1.3.3 The regulatory role of NO in the blood vessets
Nitric oxide is highly lipophilic and readily diffises between adjacent cells
and thus can be produced in one cell and have its biological effects in the next
d l . It exerts its physiological role as a result of activating soluble guanylyl
cyclase (Radomski et al. 1987 and Schmitt et. a1.1992). In this respect NO
u tirnately increase intracellular cGMP.
In the vascu lar system, activation of muscarinic recepton, serotonerg ic
(5HT1) receptors, aradrenoceptors and shear stress (Flavahan et. al. 1992;
Wang et. al. 1997 and Kichuk et al. 1996) lead to endothelium-dependent
relaxation which can be inhibited by cornpetitive NOS inhibiton such as L-
NMMA,
The diffusion of NO to the vascular smooth muscle cells result in
increasing cGMP levels which activate protein kinase G(s) phosphorylating target

pmteins involved in ca2' deposition. This results in accelerated ca2+ rernoval
from the smooth muscle ceIl leading to vasorelaxation.
Nodependent vasodilator tone is implicated in the physiology of all
vascular beds to some extent. Most importantly, however, are the tesistance
beds where tissue metabolic events, hormonal stimuli and flow stress locally
regulate these adaptive responses in the cardiovascular system.
AORTA
The progression of vascular research has witnessed the advent of new
and exciting ways for studying endothelium derived relaxing factor (NO). For
instance, diaminofluoresœins (fluorescence indicators for NO) have been used
to detect the release of NO from bovine aortic endothelial cells. This directly
shwed the generation of NO (via the constitutive nitric oxide synthase) from
bovine aortic endothelial cells (Nakatsubo et el. 1998). In a related study, a nitric
oxide-selective electrode has been used to directty measure NO release frwn
isolated rat aortic endothelium and cultureci rat aortic endothelial cells (Guo et el.
1996). These investigators showed that basal release of NO was detectable in
isolated rat aortic rings with intact endothelium and the response was abolished
upon administration of (1 mM) N'-nitro-~marginine methyl ester (L-NAME).
A number of endogenous compounds have been implicated in stimulation
of the vascular nlric oxide synthase. For example in the rat aorta, interieukinl-P
(Ikeda et al. 1995), adenosine (Ikeda et al. 199?), insulin like growth factor
l(Muniyappa et al. 1997), angiotensin II (Pueyo et al. 1998) and oxygen

(Whorton et al. 1997) al1 a d to alter or modify the cardiovaswlar regulation
pnmanly by stimulating the release of NO.
The tonic release of NO is set by the physical property of shear stress
followed by flow-induced production of NO. For example in a study to determine
mechanistic regulation of endothelial constitutive NO synthase (ecNOS), Corson
and associates have measuted NO production by bovine aorüc endothelial cells
exposed to shear stress in a larninar flow chamber (1996). In this study it was
shown that concomitant with the augmented production of NO,
immunoprecipitation of ecNOS showed a 210% increase in phosphorylaüon after
1 minute of flow initiation. The investigaton concluded that shear stress can
increase NO production for which phosphorylatian of ecNOS may importantly
modulate its activity during the imposition of increased shear stress (Corson et
al.). A sirnilar study has demonstratecl that exposure of cultured rat aortic
endothelial cells to well-defined laminar fluid flow results in a biphasic elevated
rate of NO production (Kuchan et ai. 1994). An initial rapid production of NO
consequent to the onset of Row followed by a less rapid, sustained production. It
was also noted that only the sustained production of nitric oxide required the
continua1 presence of flow stress and this response was insensitive to either ca2+
or calmodulin inhibitors. The rapid production of NO was shown not to be
dependent on shear stress within a physiological range (6-25 dynlcm2) but was
dependent on the rate of change in shear stress. This component was shown to
be sensitive to of ~ a * and Calrnodulin inhibiton (Kuchan et al.).

In the aorta, production of endothelial NO probably plays a major mle in
hemodynamic regulation. However, more research is needed to elucidate the
relative importance of nitnc oxide in relation to regulatory roles of prostacyclin
and endathelial derived hyperpolarizing factor (EDHF).
MESENTERIC ARERY AND VElN
Direct measurement of NO release due to increase in blood flow has been
shown in canine mesenteric resistance arteries (Hyre et al. 1998). In this in vivo
study, NO concentration was measured with NO-specific electrodes (200-micro-
tip diameter) on the outer wall of the mesenteric arteries. It was shown that by
doubling blood flow an increase in NO concentration of at least 100 nM at the
outer arterial wall occurred (Hyre et al.). Recently, an in vitro study investigating
the relative significance of NO mediated relaxation in different sized blood
vessels, showed that acetylcholine elicited an endothelium-dependent NO
mediated relaxation in phenylephrinecontracteâ superior mesenteric arteries but
not in the resistance mesenteric arteries (Hwa et al. 1994). This conclusion was
reached in view of the fact that in the superior mesenteric artery the NO inhibitor
N=-monomethyl-~earginine abolished the relaxant response. The acetylcholine-
mediated relaxation in the resistance mesenteric arteries was attributed to a
hyperpolarization where this response was attenuated by pre-administration of
(0.1 PM) ~a*'-activated K' channel blocker charybdotoxin (Hwa et aL).
Similar to aortic tissue, in the mesenteric artery, apart f m flow-mediated
NO production, a number of endogenous agents sümulate the production and

release of NO. For example in a study by Champion et al. it has been shown that
in the rat mesenteric artery, the stimulation of the HI and Hz but not the H3
histaminergic recepton caused a signifiant vasorelaxation (1 998). In a related
study on the rat mesenteric artery, Chataigneau and associates compared
acetylcholine mediated release of NO followed by direct vasorelaxation with
hyperpolarization and vasorelaxation mediated by anandamide [a
cannabiniod](1998).
Other studies in the mesenteric artery point to a neurogenic role of NO.
For instance, Okamura et al. have observed neurally induced venous relaxations
via NO release from perivascular nerves in dog and rnonkey mesenteric arteries.
They concluded that nitroxidergic and sympathetic nerves innervate mesenteric
vessels and act opposingly to balance the vasodilation (1 995).
In a similar study, Ahiuwalia and coworken show that capsaiciri, a
selective C-fiber activator, relaxes small rat mesenteric veins in an endothelium-
dependent manner, demonstrating that the venous side of the mesenterk
microcirculation can also respond directly to sensory stimulation (1 997). Although
the production and release of NO is a major regulator pathway in the control of
mesenteric hemodynamics, some studies also point out to the relative
importance of prostanoids (Peredo et. al. 1997). For instance it has been shown
that both acetylcholine and bradykinin (albeit via d-ïfferent mechanisms) cause
the release of prostacyclin in the rat mesenteric vein (Peredo et al).
As it is evident from the studies mentioned, the vascular mesentek bed
utilizes a number of mechanisms to locally regulate blood flow and pressure.

Aithough the production and release of endothelial NO is not the only adaptive
mechanism used by these blood vesse1 1 is probably one of the moût signifiant
processes.

2.0 RATIONALE, OBJECTIVES AND HYPOTHESES
The intention of this study, is to address the following pharmacological
h y potheses:
1. That a and P- adrenoceptor subtypes differ according to vascular
bed and function.
2. There is a differential response to NO production in conduit and
resistance artenes which is due to activation of distinct NOS
isoforms.
Therefore, in order to test these hypotheseç the following experirnents are
proposed:
1. Functional delineation of a and 0- adrenoceptor subtypes on
conduit and resistance vessels. This will be done utilizing a number
of selective and non-selective agonists and antagonists.
Experiments will be designed to look at contraction and relaxation
responsiveness using organ bath and microvessel chambers.
2. Examination of the role that NO may play in control of vascular tone
in large conduit and resistance artenes. The use of selective NOS
inhibitors will shed light on the nature this control by suggesting that
different NOS isoforms may play distinct mies.
The result of these studies should add to the present knowledge ( with respect to
the differential ability of the cardiovascular components that regulate
hemodynamics) but also help to elucidate the distinct roles of resistance versus
the conduit vascular teactivity in cardiovascular regulation.

3.0 METHODOLOGY
3.1 Experimental approach
In these studies in-vitro experiments testing the responsiveness of rat
blood vessels were conducted. Therefore, contractions and relaxation caused by
a variety of agents were recorded in the rat aorta and the small (100 to 250
microns in diameter) branches of the mesenteric artery. Wrth the help of selective
agonists and antagonists various recepton involved in physiological
hemoâynamic wntrol were characterized.
3.2 Tissue preparation
Ail animal experiments were approved by the Animal Care Cornmittee of
the University of Toronto in accordance with the Animals of Research Act and
Canadian Guidelines on Animal Care. Male SpragueDawley rats (n=59)
weighing 300-4509 (Charles River, Quebec, Canada) were killed, by elher
decapitation or cervical dislocation. Immediatsly following death, a large incision
was made to open the pleural and the abdominal cavities. The rib cage, the
abdominal wall, esophagus, trachea and the lungs were sequentially reroved to
locate the underlying aorta. The pericardium and the thoracic aorta were blunt
disseded and excised beginning from the aortic arch and descending to the
border of the thoracic and abdominal aorta at the level of the diaphragm.
For extraction of the mesenteric artenal tree, the smell intestines, and its
mesentery were cnidely removed to be fine dissected at a later stage.

The freshly excised aorta and mesentery were then placed in a beaker
filled with ice cold ( 4 O C) Kreb's-Henseleit (Kreb's; see the drugs section for the
solution composition) solution and immediately subjected to a fine dissection
protocol. (See below)
3.3 Experimental apparatus
3.3.1 Mesenteric artery dissection and set up
3.3.1.1 Fine dissection
The intestine and the rnesentery were placed in a petri dish coated with a
transparent elastomer (# 184 Sylgard, Paisley Products of Canada Inc..
Scarborough, Ontario, Canada) and filled with ice cdd (4") Kreb's. The
mesenteric bed was subsequently spread out by pinning and securing the
intestines. The third generation mesenteric arteries (diameter range: 75 to 250
pm) were then careful ty disseded with the aid of a dissecting microscope (Nikon
SM- IB , Melville, NY, USA) at 12-50 x magnification. The arteries were
distinguished from the parallel ninning veins by striated demarcations that nin
dong the length of the vessel. In addition, the arteries appeared more rigid due
to their elastic properties as opposed to the placid and inelastic nature of the
corresponding veins.
The microvessel was then rneticulously cleaned of any surrounding fat
and adherent tissue using a pair of Dumont #5 Carbon Forceps (FST 11251-10,
Fine Science Tools Inc. Vancouver, B.C. Canada) and mini-Vanas scissors

(FST-15000-10 Fine Science Tools Inc.) Care was taken not to darnage the
vascular endothelium.
3.3.1 -2 Mesenbric ertery set up calibration
Diagrammatic representation of the rnesenteric artery set up is shown in
figure 3.1. A Video Dimension Analyzer (Living Systems, Burlington Vermont,
USA) was used to provide a visible and accurate measurement of the luminal
diameter directly in microns (pm). Prior to each experirnent, the Video Dimension
Analyzer was calibrated using a micrometer, which was placed on the stage of
an inverted microscope (Nikon, TMSnMS-F X500 total magnification). The
transluminated micrometer was then visualired on the television monitor so that
its precise 5p divisions were dearly visible. The white highlighted scan line was
subsequently positioned so that it would intersect the divisions of the stage
micrometer using the 'SCAN LINE' knob on the apparatus. This subsequently,
would provide readings of these divisions by the apparatus. Any discrepant
reading from the Video Dimension Analyzer was subsequently corrected using
the 'ZERO and SCALE' knobs. This manual adjustment continued until the Video
Dimension Analyzer correctly measured the micrometer divisions.
In al1 experiments that were conducted, the Video Dimension Analyzer
was calibrated so that its reading emr for the micrometer was no more than one
pm (i.e. at worst for a 50pm division of the micrometer the reading was 49pm or
S I pm).

Fiaure 3.1 Mesenteric artery apparatus
A. The plexiglass microvessel set up is placed on the stage of the inverted microscope. The camera wntinually filmed the change in vesse1 diameter and displayed it on the monitor.
B. The television monitor is depicted. Any diameter changes were continually recorded via the Video Dimension Analyzer and were graphically shown on the chart recorder.
Cm The close up of the mesenteric microvessel chamber is shown.


The 'PRESSURE MONITOR' of the pressure control unit of the rnicrovessel set-
up was calibrated by attaching a manometer to the pressure transducer. In
addition, following connedion of the manorneter, the 'PRESSURE ADJUST' knob
was dialed to zero. However if the meter did not read zero, the dial marked as
'ZERO' was tumed using a screwdriver until zero was achieved. Various
pressures (5-60 mmHg) were applied by the manometer and calibrated by the
'PRESSURE ADJUST' knob, accordingly, to regulate the digital reading to the
correct pressure.
Following the calibration of the Vdeo Dimension Analyzer and the
pressure unit, the polygraph recorder (Fisher Recirdall Series 5000) was
calibrated. This was accomplished by tuming on the recorder and moving the left
and right windows to adjust the subsequent diameter readings. The pen on the
recorder was placed at zero using the 'PEN' knob. The sensitivity of the
polygraph was set at 10 volt in the recording channel. Finally the chart paper was
allowed to run at a speed of 0.25 cmlrninute throughout ail of the experiments.
3.3.1.3 In vitro mesenteric artery set up
The dissected rnesenteric microvessel was transfenad into a special
plexiglass (8 ml volume) microvessel chamber containing Kreb's solution (pH
7.4) constantly being bubbled with a 95% O2 and 5% CO2 (Figure 3.2). Previous
studies have indicated that continuos bubbling of oxygen does not change the
physiological H' concentration and therefore the pH (Larosa, 1 997). lnitially

Finure 3.2 The mounting of the mesenteric artery
The fine dissected third generation mesenteric artery was mounted in the Kreb's filled (warmed and oxygenated) plexiglass chamber. The residual blood was flushed out of the vessel. Subsequently the vessel was secured at both ends with the aid of ethicon surgical sutures. The mounted vessel was then inflated (20 mmHg) and equilibrated for an hour prior to any experiments.


any residual blood in the vessel was flushed out by fastening one end of the
preparation to one of the two glas cannulae in the chamber via the peristaltic
pump (Living Systems Instrumentation, Burlington, Vermont, USA) which was
wnnected to the cannula. Following this, the dissected arterial segment was
secured with 10-0 Ethicon nylon monofilament ligatures (Johnson and Johnson
Medical Products, Peterborough, Ontario, Canada) and mounted at both ends of
the chamber between the two glass canulae (50-100 pm in diameter). More
elaborately, in the chamber, one of the cannulae was blind-ended in a non-fiow
mode whiie the other was attached (via Tygon tubing, diamete~2mm) to a
pressure transducer that constantly sampled and monitored the vessel's
intraluminal pressure. A second pump was connected (via Tygon tubing
diameter=5 mm) to a heated water jacket (that was heated by a Haake D2
circulator) which maintained the temperature at 37°C where it wnstantly
replenished the Kreb's solution in the chamber during equilibration from a
resewoi r.
After mounting the vessel, the pressure servo adjuster knob was used to
apply a pressure of 20 mmHg in order to Mate the mounted vessel in a 'no flow'
state. The central portion of the vessel that was suspended between the two
cannulae was not wnnulated so that endothelial integriiy was maintained.
The mounted microvessel was then placad ont0 the microscope stage.
Translumination of the plexiglass chamber allowed the filming of the apparatus
by an inverted carnera that was attached to the microscope (Nikon, TMSKMS-F
X 500 total magnification j. In the same manner, as previously described for the

micrometer, the central portion of the vesse1 could subsequently be visualizeâ by
the television monitor (Figure 3.1). Furthemore the calibrated Video Dimension
Analyzer could accurately measure changes both in the luminal diameter as well
as the wall thickness of the vessels following the administration of the vanous
agonists either in the absence or presence of antagonists.
Following drug administration, continuous diameter measurements were
made using a v i d a tracking device (Living systems, Burlington Vermont, USA)
as previously described by Halpern et al. (1984). The tracking device (a scanline
running horizontally across the television monitor that allowed the measurement
of diameter changes based on the principles of optical density changes) hence
allowed the recording of diameter changes on a polygraph trace (in
micrometers). This was due to changes in the signal curent across the television
monitor which were produced by changes in the dismeter. In this respect the
changes in distention and dilations of the vessel could be recorded.
In all experirnents prior to any intervention, the mounted pressurized artery
was allowed to equilibrate for at least one hour in Kreb's solution bubbled with
95% O2 and 5% COz. The viability of the vessel was rneasured as a greater than
20% distention of the vessel as a result of 20 mM KCI administration (Myen et
al. 1989). The optimal, intraluminal distending pressure was determined in pilot
studies where pressure vs. contraction curves were constnicted. Accordingly it
was established that the vesse1 conveys the greatest degree of contraction under
a pressure of 20 mmHg.

3.3.2 Aortic fine dissection and set-up
3.3.2.1 Fine Dissection
As previously described approximately 5 to 6 cm of aorta was excised
frorn the rat and placed in ice cold Kreb's solution. In order to further fine dissect
the aorta, it was placed on a Kreb's-moistened filter paper. It has k e n described
previously (Larosa, 1997 and Rubanyi et. al., 1985) that dissection of vascular
segments over Kreb's -moistenecl filter papers does not diminish the viability of
the tissue. In light of this. the aorta was scnipulously cleaned of as much
ad herent tissue as possible.
The aortic tissue was then cut into six rings of equal length in length (5
mm). In some experiments the vascular endothelium were removed, by placing
the tip of fine forceps through the lumen and gently rolling the preparation back
and forth over the moistened filter paper (Larosa 1997).
3.3.2.2 Aortic organ bath apparatus and calibmtîon
Prior to commencement of each experiment, the force transducer was
calibrated using standard 200mg and IOOOmg weights that were pfaced on the
force transducer to measure the accuracy of its measurement at vadous voltage
channel amplifications. Any discrepancy was subsequently manually correcteci
and the procedure was repeated until the polygraph reading corresponded to the
known value of the weig ht by precise defiection of the Pen.
Following calibration, aortic rings were mounted in separate lOmL glass
charnbers (Ealing Scientifc Ltd. St. Laurent. Quebec) that contained Kreb's (pH
7.4) which was continously bubbled with a gas wntaining 95% O2 and 5% C a .

A constant temperature of 37°C was maintained using a cirwlating water pump
( ~ a a k e ~ D1). Subsequently every ring that was dissected was carefully
suspendeci between two L -shaped stainless steel hooks where one of the hooks
was attached to a metal tissue holder in the organ bath's interior and the other
was connected to the force transducer (Model FTOBC, Grass Instrument Co.,
Quincy, Mass. USA). This latter connection was made possible by means of a
surgical suture (~ th icon~, 4.0, silk). Subsequently the development of changes in
the isometric tension as the result of dnig administration could be amplified and
recorded at a speed of 2.5 mmlmin using a polygraph [Grass mode1 7D] (Figure,
3.3).
Prior to any expenment, al1 rings were allowed to equilibrate for a period of
at least one hour dunng which time the rings were constantly washed with Krebs
solution. In this periad the aorüc rings were progressively stretched to mach an
optimum resting tension of 2 grams. Once this resting tension was stable over
several washings the experimental protocol was cornmenced.
3.3.3 Experimental protocol
Following equilibraüon, viability was established by a 20mM challenge of
KCI. Likewise endotheliurn integrii and function was assessed in KCI pre-
constrided endothelium intact and denuded preparations using cumulative
acetylcholine administration (1U8 M-~o-'M) with 100 ML log unit concentration
increments.

Fioure 3.3 The aortic experimental appaatus
A. The aortic ring was secured between two L-shaped hooks, one of which was attached to the tissue holder and the other to the transducer via a silk thread. The mounted ring was subsequently subjected to constant stretching until it stabilized under two grams tension. Meanwhile the ring was continually washed and bubbled with oxygen.
B. During the course of the expetiment, the development of any response (either a constriction or a relaxation) was graphically represented via aie Grass polygraph.


Hence responses developed to the effect of each acetylcholine concentration
were allowed to plateau before addition of a subsequent concentration. Vîability
of the endothelium was established by successive incremental relaxation of the
preconstricted vesse1 after administration of each concentration. Successful
removal of the endothelium was evaluated by the lack of this relaxation
response.
3.3.3.1 Delineation of a-adrenoceptors in the small mesenteiic artery versus the aorta
A control (absence of antagonist) cumulative concentrationeffect curve
was constnicted where the most dilute concentration of the agonist (1 O" M) was
added to the tissue bath chamber. When the recorded response plateaued the
next concentration was added (in half log units). When this response plateaued
the procedure was repeated until the highest concentration was added or until
the contractile response maxirnized fully.
After the highest concentration of agonist was added in the absence of
any antagonist (Le. the control concentrationsffect curve), the preparaüon was
wntinously washed wlh Kreb's solution until the baseline diameter was
reestablished. At this point in the expriment the preparation was incubated with
the lowest concentration of antagonist for at least fifteen minutes. Subsequently,
concentration effect curves were constnicted but in presence of pre-incubated
antagonist. This procedure was repeated using at least Wo higher concentrations
of antagonist Only one antagonist was assessed on a single preparation.

3.3.3.2 Delineation of P-adrenoceptors in the small mesenteric artery versus the aorta
In an analogous manner, the effects of salbutamol and isoproterenol (1 0%
104) were examined either on the KCI (20 mM) preconstricted mesenteric artery
or phenylephrine (108 M) pre-ainstricted aortic rings. In addition, the nature of
the p-adrenoceptors were identified via antagonisrn of the P-mediated responses
by the non-selective p-blocker propranolol (1 o4 to 1 0-7 M )
3.3.3.3 Endothelial modulation of vascular responsiveness
In these experiments specific NOS inhibiton (namely Lm-NNA and
arninoguanidine [10m5 to 104 MI) were utilized to assess the nature of NO
production in conduit versus resistanœ vessels. Therefore, relaxations to
acetylcholine either in KCI pre-conçtricted mesenteric artery, or phenylephrine
pre-constricted aortic rings, were subjected to the pra-incubation with NOS
inhi bitors.
In these experiments, Gare was taken to maintain endothdial integrity, as
this vascular component was the underlying modulator of contractiky via the
actions of NOS and NO production.
3.4 Drugs and solutions
The KrebtîHenselel solution of the following mM composition: d-glucose.
10; NaHC03, 25; KCI, 5.6; NaCl 120; MgS04 1.2; NaH2P04, 1.2; and CaCI*, 2.5.
The solution was prepared fresh each day of the study and was bubbled with a
gas mixture of 95% O2 and 5% COz for at hast an hour.

The solution was prepared fresh each day of the study and was bubbled with a
gas mixture of 95% O2 and 5% COz for at least an hour.
All dnig solutions were prepared in double distilled water. Noradrenaline
and isoproterenol stock concentrations (1 o-*M) were prepared in 0.2 % ascorbic
acid solution. Spiperone was prepared in 1% lactic acid solution. Ascorbic acid
inhibited the oxidization of noradrenaline and isoproterenol while lactic acid
pmmoted the solubility of the spi perme. BHT 933 [6-allyl-2-amino-5, 6,ï1 BI-
tetrahydro-4H-thiazoI01[4,5-~azepine ] is light sensitive ( Larosa , 1997) and
therefore care was taken not to expose it to sunlight or nonal fluorescent
labofatory lights. For this reason the tubes containing this solution were
stopcocked and wrapped in atuminum tin foi1 during the course of experiment.
Stock solution of both prazosin and Na-nlro-L-arginine (L-NNA) as well as
dopamine were sonicated for better dissolution.
All stock dnig solutions were prepared fresh as (IO-~M) and serially diluted
accordingly on the day of the experiment. Subsequently, just before every
experiment, serial dilutions were made with distilled water. Al\ of the drug
saluiions were placed on ice during the course of experiments with the exception
of KCI stock solution, which was stored at r o m temperature.
Ail experirnents analyzing the effects of aadrenoceptors were conductecl
in 5pM propranolol Kreb's-Hensleit solution to antagonize the effects of p-
adrenoceptors (Forster et ai. 1983; Cum and Greenberg 1983). Likewise when
the effects of P-adrenooepton were assessed, phentolamine (5pM) was added to

the Kreb's solution to inhibit a-adrenoœptor mediateâ responses (Grassby and
Broadley 1986).
Dopamine and lactic acid were purchased from the Aldrich Chemical
Company. Rqitine maleate (Phentolamine) was kindly donated by Ciba Geigy
(Mississauga, Ontario, Canada). The following agents were obtained frorn Fisher
Scientific (Neapean, Ontario, Canada): NaHC03, KCI, MgSO4, CaC121 NaH2PO4,
Prazosin hydrochloride was a gift from Pfizer Canada (Kirkland, Quebec,
Canada). S-(-)-propranolol was purchased from ICN Biochemical tnc. (Auroia,
OH, USA). D-glucose and NaCl were acquired from BDH lnc. (Toronto, Ontario,
Canada) BHT 933 was a gift from Boehringer lngelheim (Ridgefield, CT, USA).
Acetylcholine iodide, Na-nitro-L-arginine (L-NNA), aminoguanidine bicarbonate.
spiperone, 5-MU , chlorethylclonidine (CEC), BMY7378. (1)-phenylephrine, (-
)noradrenaline, (-) adrenaline, salbutamol and isoproterenol were al1 purchased
from the Sigma Chemical Company (St. Louis , MO, USA).
3.5 Dataanalysis
The data was graphically represented as raw data or was norrnalized
using a cuwfit program (Parker and Waud. 1971) to obtain maximum responses
and ECso values. In either case the graphs that were produced were plotted as a
concentration response relationship. The reliability of the normalized data has
previously been addressed in Dr. Forsteh laboratory (Larosa, 1997). In addition
a sample of fitted data is presented in appendix A. The concentration-effect
curves that are produced using the cunht program, foilow the general sigrnoidal

relation, that utilizes the logistic equation Y= {[ady(l +[xlclb)}+d, where Y is the
response, X is the arithmetic dose, a is the response when X=O, d is the
response for an infinite dose, c is the €Cm, and b is the dope factor(larosa,
1997, Parker and Waud 1979). Using this data processing program, estimates of
the individual maximum responses as well as the ECS0 values can be obtained.
In this manner, in aortic rings, the individual parameters described above
were expressed as % relaxation in the case of aœtylcholine dose and grarn.rnmm2
in al! other experimental treatments. Although dilatations in the mesenteric artery
in some cases were also given as % relaxation, the contractions in the
mesenteric artery were calculateci as the subtraction of the individual diameter
changes developed at a certain agonist concentration from the basal diameter (in
microns) of each experiment, averaged for the number of experiments.
To quantify antagonism, the ECw, as well as the maximum tension
developed to the agonist in the presence of different antagonist concentrations
were compared with the appropriate controls. The median €Cs& were wmpared
using the Kruskal-Wallis ANOVA on ranks test. Foltowing this test, the ECW of
each individual antagonist-treated curve was separately compared to the control
value, using the non-paramettic Mann-Whitney rank sum test. In this manner
ECsis that were most significantly different from the control values were
determined. In a similar manner the degree of variance in the antagonist-treated
maxima from the control was tested with the one way ANOVA test. Also each
individual antagonist treated curve was also separately compared to its
respective control value using the student t- test with a bonferroni correction.

In al1 of the tests a p value c 0.05 was considered statistically significant.
If as detemined by the appropriate ANOVA tests, the ECso and not the maximum
values for the experimental treatments were statisticall y different from the control
(NO ANTAGONIST), antagonism was deemed competitive. In this instance, a
Schild regression was constructed to quantify competitive antagonism at a
particular receptor (Aninlakshana and Schild, 1959). If on the other hand both the
ECso and the maximum values for the experirnental treatments were significantly
different fmm the control values then the antagonism was deemed non-
competitive and an lCso regression was conducted to appraise the non-
corn petitive antag onism.
The Schild regression is the plot of the LOG (DR-1) versus the negative
LOG [ANT] where DR is the dose ratio of the treatment group over the control
(for example, ECso amgon* ~ ( e g l IE& A dope of unity in this
relationship, designates cornpetitive antagonism. The pA2 (x-intercept, which is
the negative log concentration of the antagonist that causes the twofold shift to
the right in the concentrationefbct curves for the agonist) value on the other
hand, determines the measure of antagonistic potency and can be used to
detenine receptor identity.
The lCso regression was plotted as the percentage reduction in the
maximum response developed to agonist versus the incremental concentration of
antagonist. The ICW value was subsequently detennined as the concentration of
the antagonist which reduces the maximum tension (control tension) by 50
percent (Jenkinson et al. 1995). In the regression plots, coefficients of correlation

are depicted as (r) and represent the measure of the correlation of the
independent variables (namety LOG [DR-1) and %Reduction in contraction)
versus the dependent variables i.e. the concentration of the antagonist. When (r)
equals 0, the values of the independent variable do not allow any prediction of
the dependent variables. On the other hand when (r) equals 1, one can perfèctly
predict the dependent variables from the independent variable.

RESULTS
4.1 Characterizalion of the aadrenoceptors on the small rat mesenteric arteries vemus the aorta.
Initial experiments demonstrated that the rat mesenteric artery was
sensitive to the e W s of noradrenaline and phenylephnne Mth BHT 920
producing only a weak response (Figure 4.1). The noradrenaline concentration-
e W cuwe was steep and generated a maximum constriction of 150 p. Although
the maximum response generated to phenylephrine was not significantly d'flerent
from that of noradrenaline, the concentrationefkt curve was shallower and
required higher concentrations of phenylephrine to attain effects of similar
magnitude as those seen with noradrenaline. The efkt of BHT 920, on the
other hand, produced negligible diameter changes of only 30 to 35 micrometers
(Figure 4.1 ).
In the absence of antagonists, the effect of repeateâ administration of
noradrenaline on the third generation mesenteric arteries is shown in figure 4.2A.
It was observed that upon repeated administration, the contractile effect of
noradrenaline became sensitized in the 1 0"-1 O-?M noradrenaline concentration
range. However, statistical analysis of the ECW and the maximum values
demonstrated that the differences between the individual curves were not
statistically significant (p0.05, Figure 424). The ECso values for the four
consecutive concentration- response curves to noradrenaline were

Fiaure 4.1 The effect of a-adrenoceptor agonists on the small mesenteric artery.
The effect of noradrenaline, phenylephrine and BHT 920 is shown. Each point is the mean I SEM. The data is expressed as the diameter constriction in p for at least 5 (n) mesenteric preparations from 5 different animals (hi).

Figure 4.1
Phenylephrine (N=b,n=5) Noradrenaline (5,s) T T T T T
BHT 920 (5,s)
-9 -8 -7 4
LOG [Agonist] (M)

Finure 4.2 The effect of consecutive repeated administration in the small mesenteric artery and aorta.
In the mesenteric artery (A), four consecutive concentration- effect curves were constructed to noradtenaline in the absence of antagonist. Each point represents the mean I SEM. The data are expressed as mean diameter constriction in p developed for increasing noradrenaline concentrations. Repetition of the agonist administrations enhances the contractile re ponse in the 1-1 00nM C concentration range for noradrenaline. An represents significant enhancement of the contractions upon repeated administration (~~0.05, one way ANOVA). The EC501s and the maxima were not significantly different between the groups as determined by the ANOVA on ranks and one way ANOVA respectively. 20 (n) preparations were extracted from 15 (N) anirnals.
in rat aortic rings, four consecutive concentration-effect curves were constructed to noradrenaline (6) and phenylehrine (C) in the absence of antagonist. Each point represents the mean I SEM. The data are expressed as the grams tension developed per millimeter squared of dry aortic tissue. Repetition of noradrenaline and phenylephrine administration significantly lowers the maximum contractile response. A or represent concentrations of noradrenaline and phenylephnne where the contraction is significantly lowered between the four curves ( ~ 4 . 0 5 , one way ANOVA). The ECSO's were not significantly different between the curves as a group (~~0.05, ANOVA on ranks). 15 rings from 10 rat were used.


8.7xl0", 4 . 5 ~ 1 06, 1.8~1 O-' and 4 .9~1 o5 molar, respectivel y. Sirnilady the
maximum responses attained for noradrenaline were 175, 178, 173 and 143
microns, respectivel y.
Figure 4.2 panels B and C demonstrate the effect of repeated
administration of noradrenaline and phenylephrine in the rat aorta respectively.
As shown, repeated administration of noradrenaline and phenylephrine in the
absence of any antagonist progressively lowered the maximum tensions (pc0.05)
without changing the ECS0 values (p>0.05).
Table 4.1 demonstrates the degree of reduction in the maximum response
upon each subsequent administration of noradrenaline and phenylephrine. It was
obsenred that after four consecutive concentration-effect curves, the maximum
tension is reduced 50.3 and 4 1.9 percent respectively (p<O.OS).
4.l.l The effect of prazosin on noradrenaline-mediated contractile response in the mesenteric artery versus the aorta.
In the mesenteric artery, in the absence af antagonist noradrenaline
produced a contractile response that created a maximum effect of 173 n m n s
(Figure 4.3 Al.). In the presence of increasing concentrations of prazosin,
noradrenaline produced concentration-effect curves that appeared parallel and
displaced to the nght of the control. Tables 4.2 and 4.3 are central tables where
the ECS0 and maximum values [in the absence and presence al -antagonists] to
noradrenaline (Table 4.2) and phenylephnne (Table 4.3) are sumrnarized both for
the mesenteric artery and the aorta.

Table 4.1 ECa values and maximum response for noradrenaline and phenylephrine for a series of concentration-effect curves in absence of any antagonist. (Time 4ontrol study.)
The mean ECsa (M) and the maximum tension (gram. mm-2) are presented. The agonist response is repeated in absence of antagonist. The percent reduction in the maximum tension attained by repeated administration of agonist is also shown. 15 rings from 10 rats are used. An + shows significant reduction in the contraction (pe0.05, one way ANOVA).

Table 4.1
NORADRENALINE PHENYLEPHRINE
1 2 3 4 1 2 3 4
Ecs0 3 . 9 ~ 9 O* 1 2x1 u7 1.6~1 o4 9 .9~1 od 9.4~1 od 2.3~1 2x1 O-? 2.8~1 od * Maximum 167.8 k14.7 123.72 10.9 99% 12 83.42 10.5 141.2 * 13.9 110.4~ 13 70.9 r 11.2 82I16.7 *
% Reduction O 26.8 40.7 50.3 O 21.8 44 41.9

mure 4.3 Effect of prarosin on the contractile response of noradrenaline in the mesenteric artery versus the aorta.
Concentration-effect curves (CEC) were constructed to noradrenaline in the absence or presence of IO", 3x10" and low7 M prazosin in the rat rnesenteric artery (Al) and aorta (BI). Each point represents the mean & SEM. (N) represents the number of rats used and (n) the num ber of preparations extracted.
In the rat mesenteric artery (Al) prazosin shifts the noradrenaline CEC to the right where the EC501~ significantly increase (PcO.05, ANOVA on ranks). The projected maximum contractile responses are not significantly different between the four curves as a group (P>0.05 one way ANOVA). A Schild analysis was plotted (A2) with the pA2 equaling 8.6 and slope 1.7.
In the rat aorta (BI) both the ECm and the projected maxima were significantly different between the four CEC's (p*0.05, ANOVA on ranks and one way ANOVA respectively). Therefore the measure of antagonisrn was assessed using an lCSo regression (82). The [Cu, was calculated and equaled 2.4~1 o-~M. A represents significant reduction in the contraction (pe0.05, one way ANOVA).

ngm 4 3 Al.
-a -7
LOG (Prazodn] (M)

Table 4.2 ECso and maximum data for noradrenaline in the presence of increasing concentrations of a- adrenoceptor antagonists in the mesenteric artery and aorta.
A The mean EC& were detemined using the curvfit analysis. These were calculated for noradrenaline and both in absence and increasing concentrations of antagonist. In the mesenteric artery, the EC501s in prazosin and 5MU treated groups were significantly different between the four curves as detemined by ANOVA on ranks (pe0.05). In the aorta the ECSols of the entire antagonist treated curves were significantly different ( ~ ~ 0 . 0 5 ) .
B. In the mesenteric artery, the maximum tension (measured directly in p's) developed to noradrenaline is listed. The maxima between the control and the antagonist treated groups are not significantly different as determined by one way ANOVA (p>0.05). In the rat aorta, the treatment with the antagonists significantly attenuated the maximum contraction developed with al1 of the antagonists (p<O.OS).

Table 4.2
Control [ I O 30 1001 [nM] VARIANCE -
MESENTERK ARTERY Prazosin 4x1 c7 1. f x i o4 7.6~1 ou 4.5~1 o5 Significant (pc0.05) 5 MU - 54x1 o;T 1.3~1 3.3~1 O' 2.1 XI o4 Significant (~~0.05)
Pmzosin 1.3x10-~ 2.9~10~ 4 . 1 ~ 1 0 ~ 35x10' Significant ( ~ ~ 0 . 0 5 ) 5 MU - i . i x lod 7 . 2 ~ 1 0 ~ I.~XIO-~ 6 .2~10 ‘~ Signifiant ( ~ ~ 0 . 0 5 ) S~ i~erones .7~10~ s.aiOd 1.4~105 3 . 5 ~ 1 ~ ~ Significant (peo. 05) BMY7378 3.1 x l od 7 .5~1 O* 2.2~10" 1.5~10" Signifiant (p<0.05!
MAXIMUM
Controt Il O 30 IO0 ] [nM] VARIANCE MESENTERIC ARTERY ~ a t a ~ i n 172.5 + 72.6 170 119.3 164 M 169 114.3 Insignificant (pMI.05)
5 MU - 200 11 3.6 210 11 0.9 230 +6.3 196 21 5.1 lnsignificant ( ~ ~ 0 . 0 5 )

As shown in table 4.2 al1 prazosin concentrations tested resulted in significant
increases in EC& compared to cantrol (p < 0. 05).
The projedeci maximum values as denved from the curvfit program are
also shown in Table 4.2. There were no significant differences in the projected
maxima (for noradrenaline) in the presence of prazosin comparecl with control (p
> 0.05). Prazosin displaced the noradrenaline concentration-effect curve to the
right, yet attained the same maximum, demonstrating possible cornpetitive
antagonism. Hence a Schild analysis was conducted and plotted (Figure 4.3
A2). The siope of the regression line was equal to 1.7 with a pA2 value of 8. 63.
In the rat aorta, noradrenaline produced a concentration-dependent
contracu'le response with the maximum effect equaling 160 gram.mmS2 (Figure
4.3 BI). In the presence of prazosin, the ECs0 (Table 4.2) was significantly
increased compared to control (pc0.05). Prazosin concentrations caused a
significant decrease in the maximum response to noradrenaline in the absence of
antagonist ( ~ ~ 0 . 0 5 ) . When mmpared to the respective timecontrol curves
(Figure 4.28) it c m be seen that prazosin causes a further reduction with the
respective time-control curve.
In light of the fact that the maximum tensions developed in presence of
prazosin concentrations were lower than the respective time-controls (Figure
4.28), the ICHi was deterrnined taking into account the reported change in the
time-control study. lCSo regression analysis was used to measure the degree of
antagonisrn The ICs4 value was detennined to be 2.4 x 1 0 " ~ (Figure 4.3 82).

4.1.2 The effect of pratosin on phenylephrine-mediabd contractile response in the rat mesenteric artery versus the aorta.
In the mesenteric artery, phenylephrine generated concentration
dependent contractions with a maximum response equal to 128 microns (Figure
4.4 Al).
Addition of each antagonist concentration displaced the concentration-
effect curves to the right of the control. This response, however, was
concomitant with an apparent depression of the maximum efFect (Table 4.1).
Cornparison of the maximum effect attained in the absence of prazosin with that
in the presence prarosin, resulted in a statistically significant dedine of the
maximum values with 30 and 100nM concentrations of prazosin (p < 0.05).
Observation that the antagonist treatment did not achieve the same maximum
effect as the wntrol led to the measurement of antagonism by measuring the
ICso. The ICw value was therefore calculated to be 8.5 x 208 M (Figure 4.4 A2).
in the rat aorta, phenylephrine produced concentrationdependent
contractions. which generated a maximum response of 130 gram. mm". In figure
4.3 BI , prazosin caused a concentrationdependent antagonism of
phenylephrine. According to the aiwf~t analysis, the maximum response (see
Table 4.3) was significantl y (p<0.05), reduced. T hese reductions amounted to a
much greater dectine (particularly with 30 and 100 nM prazosin, peO.05 one way
AOVA) than that seen with the perspective time~ontrols (Table 4.1).
Cornpanson of the ECso values for phenylephrine in the presence of

Fiaure 4.4 Effect of prazosin on the contractile response of phenylephrine in the mesenteric artery versus the aorta.
Concentration-effect curves (CEC) were constnicted to phenylephrine in the absence or presenœ of IO", 3 x 1 0 ~ and 'IO-' M prazosin in the rat rnesenteric artery (Al) and aorta (BI). Each point represents the mean t SEM. (N) represents the number of rats used and (n) the number of preparations extracted.
In the rat mesenteric artery (Al) prazosin shifts the phenylephrine CEC to the right where the ECsols significantly increases (Pe0.05, ANOVA on ranks). The maximum contractile responses are also significantly different between the four curves as a group (~~0.05 one way ANOVA). ICm regression anal sis is shown 1 in panel (A2). The lCso value was determined at 8.5X10- .
In the rat aorta, prazosin antagonized the contractions mediated to phenylephrine (Bl). Both the and the maxima were significantly different between the four CEC's ( ~ ~ 0 . 0 5 , ANOVA on ranks and one way ANOVA respedively). Therefore the measure of antagonism was assessed by ICxi regression (BZ). The ICm value was determined as 1. W 1 o-~M. A + represents significant reduction in the contraction (pc0.05, one way ANOVA).

A l . Figure 4.4

Table 4.3 ECSo and maximum data for phenylephrine in the presence of increasing concentrations of a- adrenoceptor antagonists in the mesenteric artery and aorta.
A. The mean EC& were determined using the curvfit analysis. These were calculated for phenylephrine both in absence and increasing concentrations of antagonist. In the mesenteric artery, the EC& in pratosin and 5MU treated groups were significantly different between the four curves as determined by ANOVA on ranks (pe0.05). In the aorta the ECsols of the entire antagonist treated curves were significantly different (~~0.05).
B. In the mesenteric artery, the maximum tension (measured directly in pas) developed to phenylephrine is listed. The projected maximum between the control and the antagonist treated groups are significantly different as determined by one way ANOVA (pe0.05). In the rat aorta, the treatrnent of the antagonists also significantly attenuated the maximum contractions developed with al1 of the antagonists (~~0 .05 ) .

Table 4.3
Conbol [ I O 30 1001 [nM] VARIANCE - --
MESENTERIC ARTERY Prazosin 6.6~1 o4 1 .9~1 O~ I . ~ X I O= 5. I x i o4 Significant ( ~ C O . 05)
Prazosin 2 . 2 ~ ~ ~ 2 . 7 ~ 1 0 ~ 1 . 8 ~ 1 0 ~ 6.6x106 Significant (pc0.05) 5 MU - 2 . 5 ~ 1 0 ~ 9.9~1 o4 4.0~1 O-' 2.08~1 o4 Significant (PO. 0s)
Sd~emne 8 . 2 ~ 1 0 ~ 1.7~10" 5 . 5 ~ 1 ~ ~ 2.3x10d Signifiant (pc0.05) BMY7378 2.03~10" 6.98~10" 2.3~1 o6 1.8~1 o4 Significant (pg0.05)
MAXIMUM
Control [1 O 30 100 ] [nM] VARIANCE MESENTERIC ARTERY ~ a z o s i n 128 i I 5.8 i 50 I I 6.5 86 *I 3.4 16 -.O Significant (pc0.05)
5 MU - 233 112.1 21821 1.2 60 21 8.2 53 f 13.6 Signifiant (p0.05)
Pmzosin 128.a 33.4 t27 .s 20.7 56 I 9 4&t0 Signifiant (pc0.05) 5 MU - 112~ 14.8 6% 12.2 61r 14 68k16.6 Significant (~~0.05) Sni~erone i a9.n i 1 14% 10.9 1 1411 1 -4 91 k12.8 Significant (pc0.05)
BMY7378 24% 35.5 23th 20.7 2 1 W 126I10.3 Signifcant (pc0.05)

increasing concentrations of prazosin venus the control, demonstrated that
prazosin significantly increased the ECso values (p c 0. 05, Table 4.3).
From these data, ICW1s were determined (Figure 4.108). The ICs0 value
was calculated to be 1 2x1 o-~M.
4.1.3 The effect of 5 MU on noradrenalinemediated contractile msponse in the rat mesenteric artery versus the aorta.
In the mesenteric preparation, noradrenaline produced a maximum
contraction of 200 ps (Figure 4.5 Al). In the presence of increasing
concentrations of 5 MU, noradrenaline produced concentration-effect curves that
were displaced to the right of the control curve in an apparent parallel manner.
Table 4.2 shows ECW data for each curve and demonstrated that increasing 5
MU concentrations significantly augrnented the EC& in a concentration-
dependent manner (p < 0.05).
The projected maximum values denved from the wtvfit program are
show in Table 4.2. Cornparison of the maximum values for noradrenaline in the
presence of 5MU showed no significant differenœs (p, O. 05). This suggested
cornpetitive antagonisrn. A Schild analysis was conducted and plotted (Figure 4.5
A2) and the dope of the regresçion line was 1.9 wlh a pA2 value of 7.9.
In the rat aorta, 5 MU caused a parallel, rightward displaœment of the
concentration effect curves generated to noradrenaline (Figure 4.5 BI).
Although the ECw, for noradrenaline signficantly increased in the
presence of 5 MU, (pe0.05) the differences in maximum response developed

Fiaure 4.5 Effect of 5MU on the contractile response of noradrenaline in the mesenteric artery versus the aorta.
Concentration-effect curves (CEC) were constructed to noradrefialine in the absence or presence of 10" , 3 x 1 0 ~ and 1 oD7 M 5MU in the rat mesenteric artery (Al) and aorta (Bi). Each point represents the mean 2 SEM. (N) represents the number of rats used and (n) the number of preparations extracted.
In the rat mesentetic artery (Al) 5MU shifts the noradrenaline CEC to the right and EC5& significantly increase (P<0.05, ANOVA on ranks). However the maximum contractile responses were not significantly different between the four curves as a group (P~0.05 one way ANOVA). A Schild analysis was plotted (A2) with the pA2 equaling 7.9 and dope 1.9.
In the rat aorta (61) both the ECa and the projected maxima were significantly different between the four CEC's (~~0.05, ANOVA on ranks and one way ANOVA respectively). The decrease in the maximum response was not more than that seen in the time control. Therefore the measure of antagonism was also assessed by a Schild plot (82). The siope of the Schild regression was 0.8 and the pA2 was equal to 8.9. A represents significant reduction in the contraction (~~0.05, one way ANOVA).

Al . Figure 4.5
. 0 8 -7 d 4 4
LOG [NA] (M)
a 4
LOG[S MU] (M)

(Table 4.2) to noradrenaline were not statistically signifcant (p>0.05). In addition
the maximum responses for the four curves did not significantly differ from their
respective ürnecontrois (Table 4.1). The antagonisrn appeared cornpetitive and
the Schild analysis yieided a pA2 value of 8. 9 with a siope equal to 0.8 (Figure
4.5 82).
4.1.4 The effect of 5 MU on phenylephrine-mediated contractile response in the rat mesenteric artery versus the aorta.
In the mesenteric artery, the concentrationeffect curves generated to
phenylephrine in the presence of 5 MU were displaced in a downward fashion
(Figure 4.6 Al). The ECso values for curves in the presence of 5 MU (30 and
100nM) were significantly difierent from cuntrol (Table 4.3, p 4.05). Likewise, on
the projeded maximum response devaloped to phenylephrine in the absence
and presence of 5 MU, a concentration-dependent decline was seen (Table 4.3,
p< 0. 05). Quantification of antagonism was measured by the lC50 (Figure 4.6 B).
The IC 50 value was calculated to be 9. 3 x IO~M.
in the rat aorta, 5 MU displaced the concentrationefiect curve generated
to phenyiephrine in a paraltel and rigMward manner (Figure 4.6 BI). In contrast
to noradrenaline, there was a significant decrease in the maximum response
(see Table 4. 3) with al1 concentrations of 5 MU (p< 0.05). However this
decrease was not concentrationdependent. In addition, when compared to the
timeantrol data, 5 MU did not have a significant effect on the maximum
response generated to phenyiephnne. Moreover, ECso values were significantly

Fiaure 4.6 Effect of 5MU on the contractile response of phenylephrine in the mesenteric artery versus the aorta.
Concentration-effect curves (CEC) were constructed to phenylephrine in the absence or presence of 1 O* M, 3x1 0-*M and 1 0-7 M 5MU in the rat mesenteric artery (A l ) and aorta (BI). Each point represents the mean I SEM. (N) represents the number of rats used and (n) the number of preparations extracted.
In the rat mesentenc artery (Al) 5MU shifts the phenylephrine CEC to the right where EC5& significaniiy increase (PcG.05, ANOVA on ranks). The projected maximum contractile responses are also significantly different between the four curves as a group (pe0.05 one way ANOVA). lCSo regression analysis (A2) determined the lCso value at 9.3~10~ M.
In the rat aorta, 5MU antagonized the contractions mediated to phenylephrine (BI). Both the ECK, and the projected maxima were significantly different between the four CEC's (~~0.05, ANOVA on ranks and one way ANOVA respectively). The decline in the maximum response was not more than that seen due to time alone (see the appropriate time -control in Figure 4.2). Therefore the measure of antagonism was assessed by a Schild regression (62) and the pA2 value was 8.6 with the dope 1.2. A represents significant reduction in the contraction (pe0.05, one way ANOVA).

Bi.

(pg0.05) increased in the presence of 5 MU in a concentrationdependent fashion
(see Table 4.3). The antagonism appeared competlive in nature and a Schild
analysis gave a p& of 8.6 with a slope equal to 1.2 (Figure 4.126).
4.1.5 Tne effects of spiperone and BNlY 7378 on agonist- mediated contractile response in the rat mesenteric artery versus the aorta.
Figure 4.7 depicts the concentration-effect c w e s to noradrenaline and
phenylephrine in the absence and presenœ of increasing concentrations of
spiperone ana BMY 7378 in the mesenteric preparations.
As indicated in panel A, spiperone did not inhibit the response to
noradrenaiine but surprisingly potentiated the response at al1 concentrations
tested. This efFed was also seen with 30nM and 1OOnM spiperone against
phenylephrine (Figure 4.7 B). Statistical cornparison of the ECso and the
maximum vaiues of the control venus aie antagonist treated groups, conveyed
that they were not, however, statistically significant (Table 4.4, p> 0. 05).
Pre-incubation of the small mesenteric artery with BMY 7378 displaceci
the noradrenaline concentration-effid curve to the right with no diange in the
maximum response (Figure 4.7C, p>O.OS). However, 10 nM BMY 7378 appeared
to potentiate the efkcts of low concentrations of noradrenaline (< IO-? Ml.
Nevertheless, the overall displacement of the antagonist pre-treated curves to
the right of control were not concenûaüon dependent as the ECso values, were
not significantly different (p >O. 05, Table 4.4). Similady the effect of BMY 7378
prpincubation on phenylephrine mediated contractions is dernonstrated in figure
4.7D. Neither the ECm nor the maxima of the curves were statistiwily

Fiaure 4.7 The effect of spiperone and BlllM378 on contractile responses to phenylephrine and noradrenaline in the mesenteric aftery.
Concentration-effect curves constnicted to A. noradrenaline in the presence of spiperone B. phenylephrine in the presence of spiperone C. noradrenaline in the presence of BMY7378 and D. phenylephrine in the presence of BMY 7378. In each experiment the mean* SEM of diameter changes developed to agonist in the absence presence of IO-$ M, 3~10% and 10'~ M antagonist is presented. Neither antagonist has any effect on the ECS0 or the maximum contraction (p>0.05, one way ANOVA and ANOVA on ranks respectively). (N) represents the nurnber of animals that were used and (n) the nurnber of preparations extracted.


Table 4.4 Mesenteric ECso and maximum data for noradrenaline and phenylephrine, in the presence of increasing concentrations of a-adrenoceptor antagonists BMY 7378 and spiperone.
A. The mean ECSo (M) values calculated for noradrenaline and phenylephrine. The EC5& were determined using the curvfit analysis. They were calculated for noradrenaline and phenylephrine both in the absence and increasing concentrations of antagonist.
B. The maximum tension developed in p's, to noradrenaline and phenylephrine in the absence and presence of a-adrenoœptor antagonists.

Table 4.4 E G O
NORADRENALINE PHENYLEPHRINE
ControI [ IO 30 1001 [nM] Control 110 30 100][nM]
Spiperone 23x1 o4 6 .7~1 od a. 1 o4 2.2~1 o6 2.9~1 od 4 .5~1 oJ 1 . 7 ~ 1 O* 2.2~1 O&
BMY 7378 8.5~1 o - ~ 1.6~1 O= 9 . 4 ~ 1 0 ~ 1.7~1 o6 3.9~10' 2 . 0 5 ~ 1 0 ~ 6 . 9 ~ 1 0 ~ 1 . 7 ~ 1 0 ~
B. MAXIMUM
NORADRENALINE PHENYLEPHRINE
Control 110 30 f 00 1 [nM] Control 11 O 30 1 OOIfn Ml Spiperone 198 t 12.3 193.2 26.1 178 ki8.2 204 114.7 121.2 2 14.2 123 t10.5 121 i 9.8 121.7 11 1.9

diffkent form control ( ~ ~ 0 . 0 5 , Table 4.4 A and B, respedively).
In contrast to the rat mesenteric artery, in the rat aorta spiperone and BMY
7378 antagonized responses to both noradrenaline and phenylephnne.
lncreasing concentrations of spiperone depressed the maximum contractile
response to noradrenaline in a concentration-dependent manner (Figure 4.8 A l
and Table 4.3). Spiperone also significantly increased the ECso's of
noradrenaline (Table 4.3) wbich was also concentration dependent (pc0.05).
In addition, although the maximum responses developed to noradrenaline
were significantly different from the control (pç 0.05) the percent reduction in the
maximum response was similar to the respective time-controls (Table 4.3 and
Figure 4.28). Therefore the antagonism was cornpetitive and a Schild analysis
yielded a pA2 value of 8.9 with the slope k ing 1 .O (Figure 4.8 A2).
Spiperone was found to significantly reduœ the maximum response to
phenylephrine and displace the concentration-effect curves to the right in a
concentration-dependent manner (Figure 4.8 BI). As seen in Table 4.3, addition
of spiperone, statistically increaseâ the ECw, values in a concentrationdependent
manner (pe0.05). The doifferences in the maximum contractions attained in the
spiperone treated curves were also significant (Table 4AB, p< 0.05 ). However,
these effects were not significantly different from their respective tirne-controls
(Figure 4.2 C) thus indicating cornpetitive antagonisrn and Schild analysis gave
a pA2 value of 9.5, and slope of 1 .O (Figure 4.8 82).
In the rat aorta, increasing concentraüons of BMY 7378, displaced the
noradrenaline concentration-effect curve to the right, and significantly (pc 0.05)

Fiaure 4.8 Effect of spiperone on the contractile response of noradrenaline and phenylephrine in the rat aorta.
Concentration-effect Cumes (CEC) were constructed to noradrenaline A l and phen lephrine (BI) in the absence or 6 ) Y presence of 10 ,3x10a and 1 O M spiperone. Each point represents the mean t SEM. (N) represents the number of rats used and (n) the number of preparations extracted.
Wth noradrenaline,(Al) spiperone shifts the CEC to the right with an increase in ECds (Pc0.05, ANOVA on ranks). The maximum responses were significantly different ( ~ ~ 0 . 0 5 one way ANOVA). The decline of the maximum response was not more than that seen with the appropriate üme control (see Figure 4.2). Schild regression (A2) yielded a pA2 value of 8.9 with the dope equal to 1.
With phenylephrine (BI) both the ECSo and the maxima were also significantly different between the four CEC's ( ~ ~ 0 . 0 5 , ANOVA on ranks and one way ANOVA respectively). The decline in the maximum response was not more than that seen due to time alone (see the appropriate time - control in Figure 4.2). Schild regression (B2)gaveapA2valueof9.5andaslopeof 1 . A represents significant reduction in the contraction (pe0.05, one way ANOVA).

Figure 4.8 Al.

decreased the maximum response (Figure 4.9 A1 and Table 4.4). The €Cu,
(Table 4.3) values for noradrenaline in the presence of BMY 7378 were
significantly different Rom the control (pc0.05). Furthemore, cornparison of the
decline in the maximum response in the presence of increasing concentrations of
BMY 7378 with the appropriate tirnecontrols (Figure 4.2 8) revealed that they
were not significantly different. Schild regression (Figure 4.9 A2) gave a pAz
value of 9.9 and a slope of 0.4.
Figure 4.9 61 depids the effect of BMY 7378 on phenylephnne induced
contractions in the rat aorta. BMY 7378 disptaced the concentration-effect
curves to the right of the wntrol with an apparent decrease of the maximum
response. However, cornparison of the mean maximum response data (Table
4.3) in the presence and absence of BMY 7378 as derÎved h m the curvfit
analysis showed that there were no signifiant differences in the derived maxima
( ~ ~ 0 . 0 5 ) . Thersfore 8MY 7378 had no effect per se. on the maximum response
developed to phenylephrine. Analysis of the ECSots (Table 4.3), demonstrated
that the values were significantly increased with each BMY 7378 concentration
(pe0.05). Schild analysis gave a pA2 value of 9.2 and a dope of 0.4.

Fiaure 4.9 Effect of BMY7378 on the contractile response of noradrenaline and phenylephrine in the rat aorta.
Concentrationeffect curves (CEC) were constructed to noradrenaline A l and phen lephrine (BI) in the absence or 4 ' 7 presenœ of 10 ,3x1o4 and 10- M BMY 7378. Each point represents the mean k SEM. (N) represents the number of rats used and (n) the number of preparations extracted.
With noradrenaline, (A l ) BMY 7378 shifts the CEC tu the right with an increase in ECSo's (Pe0.05, ANOVA on ranks). The projected maximum responses were significantl y (~~0.05 one wa y ANOVA). The decrease of the maximum response was not more than that seen with the appropriate time-control (see Figure 4.2). Schild regression (A2) yielded a pA2 value of 9.9 with the dope equal to 0.4.
Wth phenylephrine (BI) both the ECso and the maxima were also significantly different between the four CEC's (pe0.05, ANOVA on ranks and one way ANOVA respectively). The decline in the maximum response was not more than that seen due to time alone (see the appropriate time - control in Figure 4.2). Schild regression (B2) gave a pA2 value of 9.2 and a slope of 0.4. A represents signifiant reduction in the contraction (~~0.05, one way ANOVA).

Flgure 4.9 Al.
340- -e conml 0 4 BMY?S78 (1OnMJ (6. =-- B M m 7 8 p O n ~ ( r
200-
150 -

4.2 Characterization of the Psdrenoceptors on the smal mesenteric a-ry venrus aorta in the rat
For both types of preparation, the effects of P-adrenoceptor agonists were
tested in pre-constricted vessels using 20 mM KCl. The responsiveness of these
preparations to P-adrenoceptor stimulation was then assessed in the presence of
increasing concentrations of propranolol.
In the mesenteric artery, isoproterenol produced a concentration-
dependent relaxation (Figure 4.10 A). The addition of propranolol in the
concentrations tested failed to antagonize the e%ds of isoproterenol (Figure
4.1 0 A). The ECso value for the control airve was 5 . 9 ~ 1 o5 M which was sirnilar to
€Cm values seen in presence of 1 O*, 3x1 0' and 1 O-' M propranolol (2.9~1 M,
4 . 0 ~ 1 0 ' ~ M and 4. MM. respectively),.
The effect of salbutamoi on the mesenteric artery is shown in figure
(Figure 4.10 C). it can be seen that salbutamol produced a negligible effect
exœpt at IO+ M, where it produced a mean relaxation of 30 &IO%. Propranolol
also had a negligible effect against salbutamol.
Propranolol also did not have any efkt on the relaxation mediated by
either isoproterenol or salbutamol in the aortic preparations (Figures 4.1 0 6 and
D respectively). The apparent potentiating e M of propranolol is identical to the
enhancement of the isoproterenol and salbutarnol relaxation due to repeated
administration alone (Figure 4.1 f A and 6).

Fiaure 4.10 Effect of propranolol on the relaxation responses of Padrenoceptor agonists in pre-constricted rat mesenbric artery.
Concentration-effect cunre constructed to the effects of isoproterenol in the rnesenteric (A) and aorüc (B) pre-constricted preparations [KCI (20mM)l. The response is recorded in the absence and presence of 1 x 1 0 ~ and 3x10" M [in aortic preparations] propranolol.
Conœntrationeffect curves constructed to the salbutamol mediated relaxation of the KCI pre-constricted rat mesenteric artery (C) and aorta (D). The CEC's were made in the absence and presence of 10" , 3 x I o4 and 1 O'~M propranolol. The data is presented as meanSEM. of contractions developed as a percentage of KCI mediated preconstricüon of the mesenteric artery and as the grams of tension deveioped per millimetre squared of dry aortic tissue for the aomc preparation. (N) represents the number of
and (n) the number of preparations extracted. A anima# represents significant enhancernent in the relaxation
response (p<0.05, one WaY ANOVA).

LOG OS01 (M)
- -. -10 8 4 -7 4 5 4
LOG [SALI (M)
140 - D*
120 -
lm-
da-
m-
40 -
20-

Fiaure 4.1 1 Effect of repeated administration on the relaxation responses of Padrenoceptor agonists in the KCI pre-constncted rat aortic rings.
Concentration-effect CU rve constructed to the effects of isoproterenol (A) and salbutamol (B) in KCI (20mM) pre-constricted rat aorta. Concentration-effect curves were constructed in the absence of propranolol at three consecutive times. The data is presented as mean+SEM. of contractions developed as a percentage of KCI mediated pre-constriction of the mesenteric artery and as the grams of tension developed per miilimetre squared of dry aortic tissue for the aortic preparation. (N) represents the number of animals and (n) the number of preparations extracted. A represents significant enhancement in the relaxation response (pe0.05, one way ANOVA).

Tension developed Gram. mmœ2 Tension developed Gram . h m

This observation was based on the comparison of series of üme controls in which
a time-dependent effect was apparent during a senes of successive
concentration-effect curves (Figures 4.1 1 A and 0 respectively).
Testing the effects of propranolol against adrenaline (NB. in this case a-
adrenocsptors were blocked throughout with phentoiamine [see methods]), figure
4.12 shows these data (A) along with the tirnecontrd comparison (B).
Adrenaline slightly relaxed the preconstricted aortic preparations, however in
concentrations above 1 O-' M produced concentration-dependent contractions.
Propriinolol failed to antagonize the responses generated to adrenaline when
compared to the tirne-control (Figures 3.12 A and B, respectively).
4.3 Angiotensin-mediated contractile effects in the rat
Figure 4. 13 compares the effect of angiotensin II with noradrenaline-
induced contraction in the rat rnesenteric artery. The maximum contraction
created by noradrenaline was 173 microns. At the highest concentration of
angiotensin II, a very small contraction was observeâ. No response was seen
with angiotensin I andior angiotensin I-VI1 (data not shown).
Because of a lack of response in the mesenteric artery, the effect of
angiotensin rnediated contradions in the aorta were not tested.

Fiaurr, 4.12 Effect of propranolol on the relaxation responses of adrenaline to the KCI pre-constricted rat aortic rings.
Concentration-effect curve constructed to the effects of adrenaline (A)in 20mM KCI pre-constricted rat aorta. The response is recorded in the absenœ and presenœ of Io9 M and 3x10"~ propranolol.
Time-controls constructed (at hourly intervals) (B) to observe the effect of repeated adrenaline administration on pre-constricted rat aortic rings. Concentrationeffect curves were constructed in the absence of propranolol three consecutive times. The data is presented as meanSEM. of contractions developed as a percentage of KCI mediated pre-constriction of the mesenteric artery and as the grams of tension developed per millimetre squared of dry aortic tissue for the aortic preparation. (hi) represents the number of animals and (n) the number of preparations extracteci. A * represents significant enhancement in the relaxation response (pe0.05, one way ANOVA).
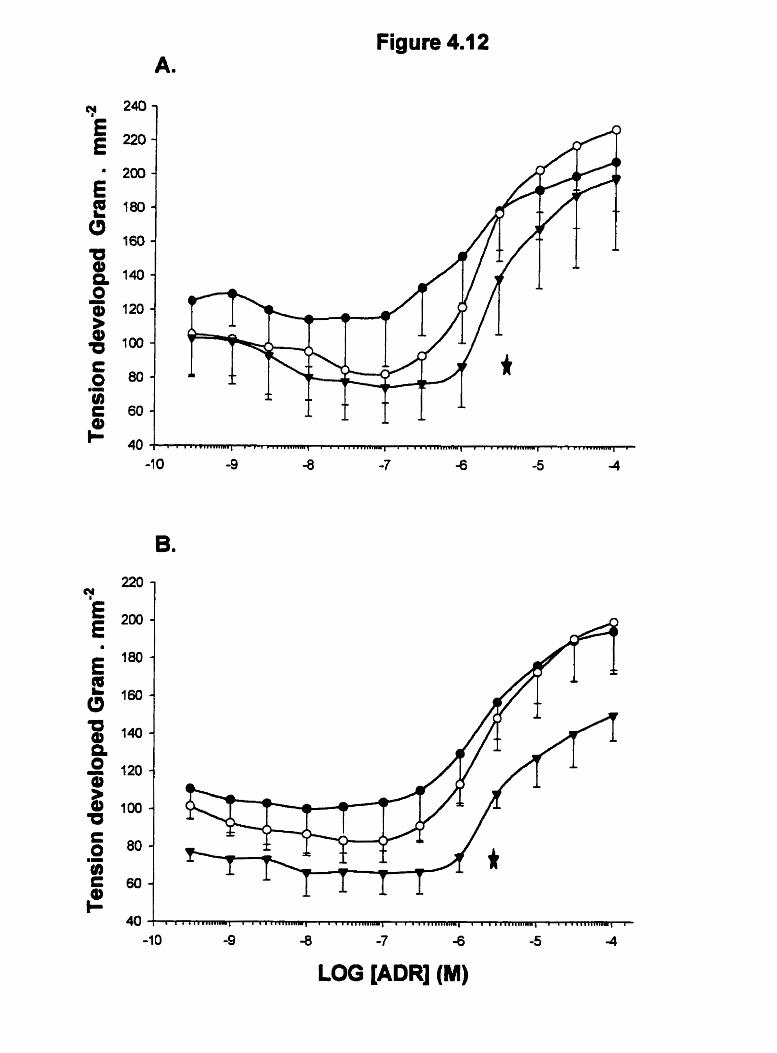
Figure 4.12
LOG [ADR] (M)

Fiaure 4.13 Contractions mediabd to noradrenaline and angiotensin II in the rat mesenteric artery
Concentration-effect curves constructed to the effect of noradrenaline and angiotensin II. The data is presented as rneanSEM. of contractions developed as a percentage of KCI rnediated pre-constriction of the mesenteric artery and as the grams of tension developed per rnillimetre squared of dry aortic tissue for the aortic preparation. (N) represents the number of animals and (n) the number of preparations ext racted.

1 + Angiotensin II * Noradrenaline
Figure 4.13

4.4 Endothelfal modulation of vascular resistance in the small mesenteric artery versus aorta of the rat
In order to assess endothelial dependent relaxation in the preconstncted
mesenteric artery venus the aorta. inhibitors of NOS were tested.
4.4.1 The effect of 1-NNA on the acetylcholine mediated relaxation in the pre-constricted small mesenteric artery vemus aorta.
The effect of L-NNA on the relaxation mediated by acetylcholine in pre-
constricted (noradrenaline and phenylephrine [l PM]) small mesenteric artery is
shown in figures 4.14 A and B respectively. In a concentration-ùependent
manner, 1-N NA diminished the acety lcholine-mediated response in
noradrenaline (A) and the phenylephrine (B) preconstricted small mesenteric
artery. In both cases, L-NNA [: O ~ M ] totally abolished the relaxant response.
The degree of dedine in relaxation by L-NNA was concentration-
dependent in the noradrenaline preconstricted mesenteric artery but for the
phen ylephrine preconstricted rnesenteric artery the curves were almost
superimposable for the L-NNA at 30 and 100 PM.
Similarly, L-NNA attenuated the relaxation developed to acetylcholine in the
aortic preparations (Figure 4.14 C). This reduction in relaxation was
concentration-dependent relaxation was concentrationdependent and 1 O ~ M L-
NNA almost totally abolished the relaxation response (Figure 4.14 C).

Fiaure 4.14 The effect of L-NNA on the acetylcholinemediated relaxation to aagonist [1 PM] pre-constrictsd rat mesenteric artery and aorta.
Concentrationeffect curves constructed to aœtylcholine in noradrenaline (A) [l FM] pre-constrkted rat mesenteric artery. The response is recorded in the absence and presence of 1 oo5, 3x1 o ' ~ and 1 0' M L-NNA.
Concentrationeffect curve constructed to acetylcholine in phenylephrine (B) [l PM] pre-wnsincted rat mesenteric artery. The response is recorded in absence and presence of presence of IO-=, 3x100~ and lo4 M L-NNA.
Concentrationeffect curve constructed to acetylcholine in phenylephrine (1pM) pre-constricted rat aorta (C). The response is recorded in the absence and presence of presence of lu5, 3x10-~ and 104 M L-NNA. Each point represents mean~SEM relaxation developed to acetylcholine administration on the pre-constricted vessel. (N) represents the number of animals used and (n) the number of preparations extracted. A )) represents significant inhibition of the relaxation response (pe0.05, one way ANOVA).

Figure 4.14

relaxation was concentration-dependent where the highest concentration of L-
NNA alrnost abolished the relaxation response (Figure 4.14 C) .
4.4.2 The effect of aminoguanidine on the acetylcholine relaxation iïi Gie piu+uii&&t~G sniaii inesenteric artery versus aorta.
Similar to L-NNA, arninoguanidine, decreased the relaxant response to
a-tÿichuiiria iiï a ancentration-dependent rnanner in noradrenaline and
phenylephrine [bothl PM] preanstricted srnaIl mesentenc artery (Figures 4.15 A
and C respectively). Wdh Io4 M aminoguanidine the relaxant responses to
acetylcholine were almost totally abolished.
il-I mtîifast to the effect ol L-NNA, aminoguanidine did not show any
significant effects on the relaxant responses mediated &y at;etyichoiine in ihe rai
aorta (Figures 4.15 B and D). In both the noradrenaline and phenylephrhe pre-
constncted aortic rings, pre-incubation with aminoguanidine seemed to potentiate
the relaxant effect of higher concentrations of acetylcholine [Io4 to Io4 Ml.
However, these differences were not statistically significant (p0.05).

Fiaure 4.15 The effect of aminoguanidine on the acetylcholine- mediated relaxation to aagonist [l p m pre-constricted rat mesenteric artery and aorta.
Concentration-effect curves were constructed to acetylcholine in noradrenaline (1 PM) pre-constricted rat mesenteric artery A . The 5 ' response was recorded in the absence and presence of 10- , 3x1U5 and lo4 M aminoguanidine.
Concentration-effect curves were constructed to acetylcholine in phenylephrine (1 FM) pre-wnstricted mesenteric artery (6)). The response was also recorded in the absence and presence of IO", 3 x 1 0 ~ and 104 M aminoguanidine.
Also concentrationeffect curves were constructed to acetylcholine in noradrenaline (1pM) pe-constriad rat aorta (C). The response was recorded in the absence and presence of 1 O-', 3x1 o - ~ and 1 o4 M aminoguanidine.
Finally, conœntrationeffect curves were constructed to acetylcholine in phenylephrine (1 PM) pre-constricted rat aorta (D). The response was also recorded in absence and presence of presence of 1 O-', 3x1 oo5 and 1 o4 M aminoguanidine Each point represents meankSEM relaxation developed to aœtylcholine administration on the pre-constricted vessel. (N) represents the number of animals used and (n) the number of preparations extracted . A represents significant inhibition of the relaxation response ( peO.05, one way ANOVA).

Figure 4.15 A.
LOG [ACW (M)

5. O DISCUSSION
The key findings of this thesis were:
That the response to repeated successive noradrenaline administration
produced eithet a sensitizing effect (in the case of third generation mesenteric
arteties) or a desensitizing effect (aorta). In the third generation mesenteric
artery, only orlA-adrenoceptors were present. In contrast al1 three al-
adrenoceptor subtypes were observed in the rat aorta. The lack of response by
BHT 920 indicated th& aradrenoceptors play a minimal role in the contractile
response of the mesenteric artery. The vasoreactive response of the third
generation mesenteric artery and aorta to padrenoceptor stimulation revealed an
efFect that could not be antagonized by propranolol. Nanomolar and micromolar
concentrations of isoproterenol were needed to relax the preconstrided rat '
mesenteric artery and aorta respectively. This implied that at least in the rat aorta
an atypical P-adrenocaptor might be present. And finally, endothelial modulation
by inhibiting NOS revealeâ that in the third generation mesentenc artery, there
was both an aminoguanidine selecüve component and an L-NNA selective
component. In wntrast no aminoguanidine selective component was seen in the
rat aorta.
5.1 The effect repeated administration of a-adrenoceptor agonists
Successive concentration-effect wrves generated to noradrenaline
produced sensitization to low concentrations (1-100 nM) in the third generation
mesenteric artery. This was particularly evident after the third and fourth

concentration-effect curves. This is possibly Iinked to phosphorylation as it has
been shown that sensitization occurs with phosphorylation of the mysoin light
chains in the vascular smooth muscle from the skinned rabbit aorta and the
mesenteric artery (Satoh et el. 1994 and Fujiwara et al. 1989). For instance, in
the rabbit thoracic aorta noradrenaline pmduceû phosphorylation of myosin light
chains (by increasing the ca2+) sensitivity of the contractile elements via the a 1 ~
and the U ~ B ~drenocepton. It was also demonstrated that ca2+ sensitization
produced by a,* -adrenoceptor was rnediated by G-protein activation of protein
kinase C (Satoh et al.). Similarly in the rabbit mesentenc artery it was shown that
GTP and its analogue GTPyS enhanced the ca2'-induced contraction in skinneâ
vascular smooth muscle due to increase in the ca2' senslivity of contractile
proteins (Fujiwara et al.). Based on these studies it is possible that upon third
and fourth administration of agonist on the rat mesenteric arteries, the contractile
proteins becorne sensitized to ca2*. This response could simply be mediated via
the second messenger rnechanisms associatecl with the a-adrenoceptor
activation leading to phosphorylation of key proteins.
Aitematively, build up of constrictor prostaglandins such as PGFh (Le. an
increase in the synthesis or desensiüzaüon of prostaglandin degradation
machinery) in the third or the fourth concentration curves could result in an
exaggerated contractile response. This explanation is likely since indomethacine
was not used in these experiments.
Anather possibility is that the atadrenoceptors becorne desenslized.
These are pflrnarily located on the endothelium and would lead to diminished

pmduction of nitnc oxide and indirectly inhibiüng any vasodilator tone.
Denudation of the third generation mesentek artery can determine if the
endothelial production of nitric oxide has any effect on the noradrenaline
sensitization.
Repeated administration of noradrenaline and phenylephrine in the rat
aortic preparations produced a significant aecrease in the maximum response
with no change in the ECso and hence no evidence of sensitization. The
hydrolysis of membrane phospholipids is believed to be the underlying cause for
the development of desensitization of al-adrenoceptor mediated contractions in
vascular smooth muscle (Rufollo and Hieble, 1994). It has been shown that
desensitization of al-admnoceptor-mediated contractions of rabbit aorta by the
continuous exposure of adrenaline (10%l for 7 houn) was not associated with a
loss of post-junctional vascular ai-adt~ocepto~. With the use of [1251] BE2254
(Bbeta (4-hydroxyphenyl)-ethybminomethyl)-tetrao) a seleetive al -
adrenoceptor antagonist, it was shown that the loss in sensitivrty ?O adrenaline
was not mediated by the down-regulation of affinw of the al-adrenocepton for
adrenaline. This was instead attributed to a marked blunting of the adrenaline-
induced increase in phosphotidylinositol turnover (Lurie et al. 1985). In contrast
desenslizaüon of al-adrenocepton by longer exposure period (24 hours) to
noradrenaline was characterized by a decrease in receptor number with no
change in affinity as well as a reducüon in phosphotidylinositol tumover (Bobik et
al. 1984). In a retated study, Seasholtz and associates have show that in vivo
infusion of noradrenaline in the rat causes heterologous desensitiration of the

contractile raçponse and inositol phosphate accumulation in the rat aorta where
these changes are mediated by an uncoupling of receptors from their G proteins
(1997). Similarly to having a doser look at the role of G proteins in a
desensitiation response Johnson et al. conclude vascular desensitization
produced by noradrenaline infusion involves seledive reducüons in the ability of
al-adrenoceptors to acüvate Gç and Gi ( A 991). In another study the endothelium
in the rat aorta has been irnplied in having a role in a desensitization response.
Therefore it has been shown that erther mechanical removal of the endotheliurn
or chernical treatment wiar hemoglobin restores sensitivity of the contractile
response in a desensitked vesse1 (Hiremath et el. 1 991 )
Based on these therefore the desensitization of vasailar al-adrenoceptor
response can result, at least in part, from an uncoupling between al-
adrenocepton and membrane phospholipid hydrolysis. A possible mechanism
for this uncoupling may involve activation protein kinase C by diacylglycerol.
Activation of protein kinase C by phorbol esters has been shown to markedly
reduce the turnover of inositol phospholipids produced by asdrenoceptor
activation in cuttured smooth muscle cells from the rabbit aorta and the hamster
vas deferens (Leeb-Lundberg et a1.1985,). Thus, the cornplex metabolism of
membrane inositol phospholipids may be responsible for the transduction of al-
adrenoœptor activation into a final mediator for a-adrenoœptor stimulated
desensitiration.

5.2. a-Admnoceptors heterogeneity in the third generation rat mesenteric artery venus the aorta
52.1 The role of a2-adrenoceptors in the third generation rat mesenteric artery
It appean that most of the a-adrenoceptor-mediated responses of both
the rat rnesenteric artery and the aorta wem due to activation of al-
adrenoceptors. This was baseà on the lack of effect of BHT 920, which is a
relatively potent a2.adrenoceptor agonist (Alexander 1997). This meant that the
aradrenocepton play a negligible rde in the third generation mesenteric artery.
However it should be mentioned that a lack of response to BHT 920 could
potentially have been the result of a counter-balancing effect of the a*mediated
contractile response of vascular smooth muscle with that of a;! -mediated nitric
oxide production via the vascular endothelium. In order to examine this potential
interaction, administration of BHT 920 to endothelium denuded venus intact
preparations would shed light on the true nature of aradrenoceptor mechanisms
in the vasculaf contraction of the rat mesenteric bed.
5.2.2 The role of ai-adrenoceptors in the third generation rat mesenteric artery and aorta
In the rat mesenteric artery the antagonism of the noradrenaline-mediated
contractions by both prazosin and 5-MU were similar and competitive in nature.
As a result the pA2 values were found to be 8.6 and 7.8 respectively. The pA2 for
prazosin was similar to that reported in the literature, however there was little
confidence that this was the true pA2 value because of poor correlation i.e. low (r)

value (Alexander et el.). The p& value for ,MU was somewhat lower than the
reported values (Alexander et al.). The higher (r) value on the other hand,
demonstrated good confidence in the accuracy of this value. It is possible that
the cornplex interaction of noradrenaline of the al-subtypes might be masking the
true effect of 5-MU on the orrA-adrenoceptor subtypes. This is unlikely however,
since spiperone and BMY 7378 failed to antagonize noradrenaline. 5-MU has
b e n reported to inhibit other receptors (5HT receptors, Kenakin, 1987) which
was not the subject of this thesis and therefore was not characterized.
The slopes of the linear regression for these antagonists were greater
than un*Ry. A slope which is greater than one may represent a non-equilibrium
steady-state where inhibitory effects of the lower concentrations of prazosin and
5-MU (namely 10 nM) are underestimated (Kenakin, 1987). Therefore,
antagonist-receptor interaction and not diffusion is the rate-limiting step. This
would mean that the interaction of lower concentrations of the antagonists may
require a longer equilibration tirne as compared with that for higher
concentrations. In retrospect, al1 concentrations of the antagonist were incubated
for a total of 15 minutes. Thus the effect of the lower concentrations of prarosin
and 5-MU may have been mis-represented as compared to the higher
concentrations, and would result in a steepening of the Schild regreçsion.
In contrast to the antagonism of noradrenaline-mediated contractions in
the third generation mesenteric artery, prazosin and 5-MU produced a non-
cornpetitive inhibition of the phenylephrine-rnediated response. In this cese, lCso
regression analyses showed that nanornolar concentrations of both of these

antagonists resulteâ in the maximum response generated to phenylephrine to be
reduced by 50%. The correlation coefficients in the C s o regressions were 0.86
and 0.76 show-ng good confidence in the regression.
The nonampetitive nature of the antagonism by both prazosin and 5-MU
rnay be due to the fact that phenylephrine may engage in tighter interactions with
the receptor. Furthermore, it is not subjected to re-uptake and metabolic
processes. In these experiments, phenylephrine may have had a desensitizing
role on a-adrenoceptors and subsequent administration of phenylephrine would
interact with fewer fundional adrenocepton teading to a decline of the maximum
response. This possibility could be addressed by repeated , successive
concentration-effect curves to phenylephnne to detemine whether an inherent
desensitization process takes place with tirne.
The failure of spiperone and BMY 7378 to produce any antagonism
whatsoever against both noradrenaline and phenylephrine mediated contractions
indicated that the a ie and alD-adrenoceptors do not play a role in the al-
rnediated contraction of the third generation rat mesentenc artery.
In the rat aorta, prazosin antagonized the contractions to both
noradrenaline and phenylephrine in an apparent insurmountable manner. The
decline in the maximum response in presence of increasing concentrations of
pratosin was greater than the inherent desensitization seen in the respedive
timeantrols. Nanomolar concentrations (240 and 120 nM) of prazosin were
required to produce a 50% inhibition of the maximum contractile response
developed to both noradrenaline and phenylephrine, respecüvely. There was Little

confidence in the validity of the IC50 regression as the r value was only 0.22. This
showed that based on the prazosin concentrations one can not reliably predict
the dependent variable, namely the degree of redudion in contraction. The
nature of prazosin's antagonism was unusual since it has b e n dassically
described as a cornpetitive antagonist (Lip and Femer, 1995; Ogbonna et al.
1997) of ai-adrenoceptors. Several explanations for the lack of competitive
antagonism might be operative in these experiments. It could be that the
washout period between successive concentration-effect curves might have
b e n inadequate. Consecutive administration of drug with intermittent washouts
of equal length may result in the build of dnig, in a rnanner where it ocaipies the
receptor but has no fundional response as in the case of a partial agonist. Hence
repeated administration of these drugs should be accompanied with longer
washout pends to ovemme this e W . In addition, Dr. Forster's laboratory has
reported variable effects of prazosin dernonstrating non-competitive antagonism
and potentiation of noradrenalineçontraction in coronary arteries of different
sizes (Larosa 1997). Prazosin has been show to antagonite the ar
adrenoceptors (namely the a2e and azc-adrenoceptors) which could further add to
the complexity of these results. Aitematively more experiments might reveal the
tnie nature of prazosin is antagonisrn with greater confidence.
In contrast, 5-MU produced competiüve antagonism of both noradrenaline
and phenylephrine responses. In this regard the pA2 value wen 8.9 and 8.6
respedively which are similar to that reported in the literature (Alexander et al.).
The correlation coefficients in the Schild analyses were 0.66 and 0.81 for

noradrenaline and phenylephrine respecüvely which generally dernonstmted
good confidence in the results. Further supporting the competitive nature of this
antagonism was that the slope of the noradrenaline Schild plot was -0.8, which
was not considerably less than one. A slope less than unity represents a sluation
where, re-uptake mechanisms may deplete the agonist from the receptor
cornpartment and therefore overestimate the antagonist potency, effectively
decreasing the dope of the Schild regression (Kenakin , 1987). However given
that the p h values were similar for both noradrenaline and phenylephrine
(against 5-MU) and to that reported in the literature, increasing the number of
experiments could possibly annul the discrepancy of this slope with unity.
Contrary to the mesenteric artery, in aortic preparations both spiperone
and BMY 7378 acted as antagonists and caused significant inhibition of the
contractile responses mediated by both noradrenaline and phenylephrine.
Taking into account the inherent desensitization of response as shown in time-
wntrol studies both of these antagonists producd cornpetitive antagonism of
noradrenaline and phenylephrine.
The pA2 values obtained from Schild regression analyses for spiperone
against noradrenaline and phenylephrine were 8.9 and 9.5 respedively, which
was similar to the consensus reporting the p h value of 8.9 for this antagonist in
the literature (Alexander et al.). Although these pA2 values were fairly similar to
the consensus reporting of the p 4 values for the ale-adrenocepton, their validity
was questionable since the (r) values from the Schild regression analyses of
spiperone against noradrenaline and phenylephtîne were both 0.44. The s lop of

the Schild plot for both noradrenaline and phenylephrine, equaled to 0. 95
(rounded up to 1 .0 in the results section) which was not significantly different
form unity. This clearly would favo~ cornpetitive antagonism.
The pA2 values obtained for BMY 7378 against noradrenaline and
phenylephrine were 9.9 and 9.2 respectively. This was somewhat of an
overestimate according to the values reported in the literature (8.4; Alexander et
ai.). There was even less confidence with these results as the r-values for these
Schild regfessions were 0.39 and 0.24 for noradrenaline and phenylephrine
respectively. Aiso as evidenced from the slope of the Schild plot, for BMY 7378
against noradrenaline and phenylephrine were significantly less than unity
suggesting an involvement of a re-uptake process which would considerably
exaggerate the antagonist potency of BMY 7378. However uptake for
phenylephrine is negligible so this explanation is inadeqwte to clarify the effect
of BMY 7378 versus phenylephrine. Another possibility would be that apart from
the arD-adrenoceptor, phenylephrine is ading on other al-adrenoceptor
subtypes, Mich would lead to a dope of < 1 .O.
The overall results of these experiments show that sub-populations of al-
adrenoœptors exist in blood vessels. However these are distributed to different
extents depending on the vascular bed and function. In this respect, the al-
adrenoceptor subtype that exists on the mesenteric artery is of the a1~-
adrenoceptor subtype whereas al1 three aadrenoceptor subtypes, namely the
aj~, a l g and the alDadrenoceptors are present in the rat aorta. However 1 should
be mentioned that based on these correlation analyses of the regression plots, it

is safe to Say that the prominent aortic and rnesenteric al-adrenoceptors are the
ai*-subtypes. Other al-adtenoceptors may exist on these vessels (for example
the ai^, a l N and the ail-adrenoceptor, Muramastu 1995). However these have
not been characterized using the recombinant techniques available. Also
selective tools that unequivocally would distinguish between these receptors
were not yet available. WiVi the advent of selective agents in the future, it is likely
that the nature of these adrenoceptors can be characterized on different blood
vessels.
These results differ from what has been reported in the literature where
the dominant aorlic and rnesenteric ai- adrenoceptors have k e n suggested to
be the alo- (aorta) and alD and a~~subtypes (mesentenc artery) respedively (Xu
et al. 1996, Piascik et al. 1994 and Piascik et al. 1994). For example in a
thorough examination of in vitro functional contractile analysis,
immunohistochemistry and quantification of mRNA, Piascik and coworkerç have
descri bed overwhelrning evidence for a al D-adrenoceptor subtype-rnediated role
O in the contractile response (to noradrenaline) in the rat aorta and the rnesenteric
artery (1 994).
The difTerenœ in our fndings and those in the literature can conceivably
be explained in the following ways. The antagonists used in our functional
experiments are not as selective (for their respective adrenaceptors) as they
have been led to believe. Therefore these antagonists inhibit the contractile
response via antagoniring the activation of a sole adrenoceptor (namely the a10-
adrenoceptor subtype). It is likely that the difierences in the antagonistic profile of

this study with those reported in the litsrature are due to an age difFerenœ in the
subject animals. For instance, Gurdal et al. have shown that al-adrenoceptor-
mediated contractile responses change with age in the rat aorta (1995). In
radioligand binding studies with rat aortic membranes, they demonstrated a high-
affinity constant for WB41 01 (1.4 nM) in 1 -month-old aorta which was consistent
wlh that for al^ -adrenoceptor subtype, whereas the high-affinity constants (0.03
nM) in 6 and 24-monthold aortas were consistent with those for the al^-
adrenoceptor subtype (1 995).
5.2.3 fbAdrenoceptor responsiveness in rat aorta vemus the mesenteric artery
The result of these experiments demonstrated that isoproterenol elicited
relaxation of the preanstricted third generation mesenteric arteries. Salbutamol
produced a small relaxation at high concentrations. Thus the lack of response
with salbutamol most likely indicates B-mediated relaxation is not due to
activation of a pradrenoceptor. However, isoproterenol (a non-seledive P-
adrenoceptor agonist) produced relaxation responses that could not be
antagonized by the non-selective P-blocker propranolol. The €Cs0 fOr this
response however was almost 2 X 104 M and therefore the relaxant response
may have been due to activation of receptors other than the fl-adrenoceptor.
Nevertheless given that the response could not inhibited by propranolol indicates
that an atypical P-adrenoceptor subtype might be involved. The atypical Ch-
adrenoceptor subtype is not inhibited by propranolol (Gemack et el. 1997). This

potentially interesting observation is the first to irnply a possible involvement of
atypical P-adrenocepton in the physiology of the rat mesenteric vascular bed.
In contrast to the third generation mesenteric artery, salbutamol resulted in
relaxation of the pre-constricted aortic preparations. In addlion adrenaline (in
presence of phentolamine) also produced a relaxation respanse. These
responses of the aorta to salbutamol and adrenaline, would indicate a potential
involvernent of the PTadrenoceptor which is the p-adrenoceptor that is
traditionally expressed in the vascular smooth muscle cells. The ECso 's for each
of the agonists were in the concentration range (accepted in the literature,
Alexander et al.) were 1X IO-*, 2.4 X104 and 1.2 X104 M for isoproterenol,
salbutamol and adrenaline respectively.
Similar to the mesenteric artery, none of these agonist responses were
sensitive to the e W s of propranolol. Although propranolol seemed to have an
effect (see results) this was not significant when the propranolol-treated vessels
were compared to the timeantrol concentrationeffect curves. In at least one
study atypical p-adrenocepton have been implied in mediating the relxation
effect by isoproterenol (Oriowo 1995). ln this study isoprenaline and the selective
Pragonists CGP 121 77 and BRL 37344 concentrationdependently relaxed the
pre-constricted rat aortic rings. The rank order of potency of these agents were
as follows: isoproterenol = CGP 12177 > BRL 37344 demonstrating a high
affinity in the rat aorta for at least one of the two p3-agonists (Oriowo. 1995).
Based on these resuîts the investigators concluded that there were at least two
atypical adrenocepton in the rat aortic rings that mediated a relaxant response.

The results in this thesis also indicate that some form of p-adrenoceptor
exists on the third generation rat mesenteric artery and aorta. These B-
adrenoceptors appear distinct from either the Pl or the Eadrenoceptor subtypes
as they are not inhibited by propranolol. To date, some blood vessels have been
found to possess a lirnited number of PTadrenoceptors (Steinkraus et al. 1996;
Feuerstein and Ruffolo, 1996). In contrast, the general consensus is that
coronary arteries most likely possess an abundance of the pl-adrenoceptors
(Ghaleh et al. 1995). Unusual vascular f3-adrenoceptors (other than in the aorta)
have not yet been identified. Therefore the results from this thesis clearly warrant
further investigation. In addition there is a difference in agonist potency between
the mesenteric artery and the aorta. Therefore it is interesting to speculate that
two distinct subpopulations of the p-adrenoceptors exist; one novel P-
adrenoœptor on the third generation mesenter~c artery and a second novel B-
adrenoceptor on the aorta. To support this, not only have P3-adrenoceptor k e n
identified recently in several different tissues (De Ponti 1997; Gerrnack, et
ai. 1997; Oriowo. 1997) but another newly reported P4-adrenoceptor has also been
suggested to exist. Of major interest to cardiovascular physiology and
phamacology the 'so callad" Pradrenoceptor has been located in cardiac tissue
(Kaurnann et al. 1 997).
5.3 Nitnc oxide modulation of the vascular contractile respome
When mesenteric arteries were preconstricted to a similar level of
contraction with either noradrenaline or phenylephrine, acetylchdine produced

concentration-dependent relaxation. These relaxation responses could be
inhibited by L-NNA in a concentration-dependent manner, independent of
whether vessels were pre-constricted with either noradrenaline or phenylephnne.
This would imply that the acetylcholine produces relaxation indirectly via
activation of cNOS. In this manner it has been shown that cNOS inhibitors such
as L-NNA block both the conversion of L-arginine to N "-hydroxy-L-arginine (a
readive intemediate in the L arginine: NO pathway) as well as the conversion of
this intenediate to nitric oxide and citrulline with similar affinities(Stuehr, et al.
1991).
Surprisingly, however, relaxation mediated to acetylcholine could also be
inhibited by aminoguanidine in a concentration-dependent manner. Once again
these effects were independent of the preconstriction agonist. This would
indicate a potential involvement of NOS.
Although iNOS is thought to be present in endotheliurn ( Gross et al.
1991) and vascular smooth muscle( Thomae et al. 1993), its activity is not
dependent on the intracellular calcium levels as is the case with cNOS. Under
conditions of long tenn stimulation (such as the fong-terni presence of
imrnunologic stimuli), iNOS produces nitric oxide over long periods of time (hours
to days) [Moncada et al 1991; Larosa 19971. Furthemore, although NO
production via the iNOS does not require calcium, it is calmodulin dependent. In
fact it is thought that calmodulin tightly binds to iNOS to sustain enzymatic
activity even at resting levels of intracellular calcium (Cho et al 1992; McCall et

al. 1991) ensuring that 1 is tonically active in the presence of oxygen, L-arginine
and NADPH (Abu and Stuehr, 1993; Cho et el. 1992; Larosa, 1997).
It can be condudeci that in the rat mesenteric artery, aminoguanidine and
L-NNA inhibit the relaxation responses to acetylcholine facilitated by NOS. Given
that L-NNA inhibits the relaxation, one can conclude that the relaxation is due to
activation of cNOS. However the effects of aminoguanidine which is believed to
be some 40 times more seledve for NOS would imply a significant activation of
iNOS in response to acetylcholine as well as cNOS. To date, NOS has only
been found to be advatad in long temi stimulation (see above). To our
knowledge if this is an iNOS activation , the data presented in this thesis would
be the first to demonstrate an acute activation of the enzyme in response to
pharmacolog ic stimulation. Whether this iNOS is the classicall y descri bed
isoforrn or whether it is a novel isofom requires further investigation . Likewise
the location of these isoforms c m not be determined from these experiments. it
is interesüng to predict that a NOS isoforrn might exist on the vascular smooth
muscle thus modulating the overall vascular responsiveness of the small
mesenteric arieries.
Muscarinic stimulation of the phenylephrine preconstricted aorüc
preparations, also resulted in concentration-dependent relaxation. lncreasing L-
NNA concentrations decreaseâ the acetylcholine dependent relaxation in a
concentration-dependent manner. In contrast to the mesenteric artery however,
aminoguanidine failed to have any effect on the relaxation response to

acetylcholine. This would cleariy indicate that only the cNOS isozyme is present
in this tissue.
As indicated in this study, in the mesenteric artery both an L-NNA and an
aminoguanidine sensitive components exist. In contrast, In the aortic
preparations, only a L-NNA sensitive component was seen. It can be concluded
that different NOS isozymes are therefore activated in distinct blood vessels.
Whether the arninoguanidine sensitive component is due to a classical iNOS
awaits further study.
These novel findings should have significant impact upon the
understanding of endcthelial regulation of vascular readivity, in the resistance
vessels versus the conduit vessels.
CONCLUSIONS
In summary the experiments conducted in this study have shown that the
rat rnesenteric artery and aorta are phamiacologically distinct apart from the
obvious anatomical difierences. It was shown that the dominant adrenoceptors in
the mesenteric artery were of the alA-adrenoceptor subtype , as ale-and a3~-
adrenoceptor selective antagonists had no effect. In addition an atypical P-
adrenoceptor component was indicated to possibly exist in this tissue sinœ
isoprenaline-mediated relaxations were not antagonized by propranolol. Finally,
in the mesenteric artery both an L-NNA and an aminoguanidine sensitive
component were observed against the relaxant actions of acetylcholine signaling
the presence of cNOS and iNOS isozymes.

In the rat aorta, al1 of the t h m a-edrenocepton seemed to be operative.
In this tissue the afKadrenoceptor also seemed to be the dominant subtype. In
the preconstricted rat aorta a significant Padrenoceptor mediated
vasorelaxation was present which also proved to be atypical. Finally the last
major differenœ between the two vessels was the fact that only the cNOS
seemed to be operative in the rat aorta.

6.0 FUTURE DIRECTIONS
6.1 Characterization of novel p-adrenoceptor-mediated relaxation in the rat mesenteric attery versus the aorta
As indicated in this study the p-rnediated relaxant responses to
isoproterenol and salbutamol were, neither meùiated through the Pl- nor the p2.
adrenoceptor subtypes. By exarnining the effect of selective Pl-agonists and
antagonists ( for eg . BRL37344 and bupranolol, respective1 y; Alexander et al.), it
will be shown whether the p-adrenoceptor mediated relaxation in these blood
vessels are mediated via the p3-adrenoceptor subtype. A lack of relaxation
responses with p3-adrenoceptor selective agents would indicate the existence of
p-adrenoœpton that are as yet not fully characterized .
6.2 Characterization of iNOS on vascular location
There are several methods that cm be utilized for characterizing and
delineating the nature of NOS on the V ~ ~ ~ O U S components of the blood vessels.
Some suggestions are as follows:
1. Utilkation of more potent iNOS inhibiton could potentially show that iNOS
exists in some vascular beds and not in others. Alternatively modulation of the
Krebs-Hensleit could demonstrate the same effect. In this respect depletion of
ca2' and the resultant abolition of a previously described acetyfcholine-
mediated relaxation, would unequivocally depict existence of cNOS in the
vascular bed .

Denudation of the vessels followed by observation of an acetylcholine-
mediated relaxation response would pinpoint NOS activity in the vascular
smooth muscle.
Several molecular techniques are now available to monitor the existence of
the various proteins. In this respect monitoring the mRNA levels of the
different NOS isoforms is readily accessible in different species and tissues.
Therefore northern analysis andlor the more sensitive RNAse protection
assay could potentially be ernployed to compare and contrast the NOS
isoform mRNA in the resistance versus the conduit vessels.
6.3 Role for other endothelialderived relaxing factors
The current knowledge regarding the status of endotheliumdependent
hyperpolarization allows the conclusion that endothelial cells do indeed release
factors which effect the membrane potential of the underiying vascular smooth
muscle cells (Feletou and Vanhoutte 1996; Beny and Chabaud, 1996; Parkington
et al. 1996). However, the relative importance of the EDHF pathways as
wmpared to the EDRF-mediated relaxation remains l e s clear. This is potentially
due to the fact that the chernical entity underiying this response has yet to be
identified and has been the subject of a controvenial debate amongst various
groups. Severai candidates have been suggested to be responsible, but no dear-
cut identification has been put forward. Such candidates include endogenous K'
channel openen and canabiniods (Petersson et al. 1997).

7.0 REFERENCES
Abu-Soud, H.M. and Stuehr, D.J.: Nitric oxide synthases reveal a role for calmodulin in controlling electron transfer. Proc. Nat. Acad. Sci. USA. Vol. 90(22),pp. 10769-10772, 1993.
Ackemann. U.: Cardiovascular system oveMew and organization, Principles of medical phanacology, sixth edition. Oxford univenity press. Toronto, 1 998.
Agrawal, D. K. and Daniel, E. E.: Agonists interaction with radiolabeled alpha- adrenoceptor antagonists binding sites in rat mesenteric artery. J. Cardiovasc Pharmacol Vo1.7 Suppl6 pp. S66-S75,1985.
Alastair, G.S., Phan, L. and Girgoriadis, G.: Physiological and pathophysiological roles of nitric oxide. Microsurgery, Vol. 1 5, pp. 693-702, 1 994.
Alexander, S.P.H., Peter, J.A. and Mead, A.: 1997 Receptor and ion channel nomenclature supplement. Trends in Pharmaculogical Sciences. Suppl: 1 -84, 1997.
Ahluwalia, A and Vallance, P.: Evidence for functional responses to sensory nerve stimulation of rat small mesenteric veins. J Phamacol Exp Ther Vol. 281 No.1 pp. 9-14, 1997
Ahlquist, R. P.: A study of adrenergic receptors. Am. J. Physiol. Vol. 153, pp.586-600, 1948.
Anfossi G. and Trovati M.: Role of catecholamines in platelet function: pathophysiological and clinical significance. Eur. J. of Clinic. Invest. V01.26(5),~~.353-370, 1 996.
Ami, M., Alpert, N.R., Maclennan, D.H., Barton, P. and Periasamy, M.: Alteraüon in sarwplasmic reticulum gene expression in human heart cells to beta 2 adrenergic stimulation. Circulation. Vol. 92, pp. 161 2-1 61 8, 1993.
Arch, J.R., Ainsworth, A.T., Cawthome, M.A, Piercy, V., Sennitt, M.V., Thod, V.E., Wilson, C. and Wilson, S.: Atypical beta adrenooeptors on brown adipocytes as target for anti-obesity dnigs. Nature , Vol. 309, pp. 163-1 65, 19û4.
Arch, J.R.: Subclassification of beta adrenoceptors : The phanacology if beta 3 adrenoceptors in tissues. Pharmacol. Cornmu. Vol. 6, Nos. 1-3, pp. 223-228, 1995.
Arunlakshana, 0. and Schild, H.O.: Some quntitative uses of dnig antagonists. British J. Phamacol. Vol. 14,pp. 48-58, 1959.

Baker, E.N. and Sipkema, P.: Components of acetylcholne-indu dilation in isolated rat artenoles. Am. J. Physiology. Vol. 273(4 Pt 2),pp.H1848-1853, 1997.
Beny , J. L. and Chabaud, F .: Kinins and endotheliumdependent hyperpolarization in porcine coronary arteries. Endotheliumderived hyperpolarizing factor. Harwood academic publishers. France 1996.
Berthelsen, S. and Pettinger, W. A.: A fundional basis of classification of alpha- adrenergic receptors. Life Sci. Vol., 21, pp. 595-606, 1977. Bilski, AJ., Halliday, S.E., Fitzgerald, J.D. and Wale, J.L.: The phamacology of a beta 2 seledive adrenoceptor antagonist (ICI 1 18551) . J. Cardiovas. Pharmacol. , Vol. 5 pp. 430-437, 1983.
Blank, J.L., Brattain, KA. and Exton, J.H.: Activation of the cytosolic phosphoinositide phospholipase C by G-protein Beta-Gama subunits. J. Biol. Chem., Vol. 267, pp. 2308923075, 1992.
Bobik A., Campbell J.H. and Little P.J.: Desensitization of the alpha 1 adrenoceptor system in vascular srnooth muscle. Biodiern.1 Phamacol.Vol. 33(7),~~.1143-1145, 1984.
Bousquet, P., Feldman, J. and Schwartz, J.: Central cardiovascular effects of alpha-adrenergic drugs: differences between catecholamines and imidazolines . J. Pharmacol. Exp. Ther. Vol. 230, pp. 232-236, 1984.
Boyer, J.L., Cardenas, C.. Posadas, C., and Garcia-Saint, LA.: Pertussis toxin induces tachycardia and impairs the increase in blood pressure produœd by alpha2-adrenergic agonists. Life Sci. Vol. 33, pp. 2627-2633, 1 983.
Bredt, D.S., and Snyder, S.H.: Isolation of nitnc oxide synthase, a calmodulin - requirîng enzyme. Proc. Natl. Acad. Sci. USA. Vo1.87, pp. 682-685, 1990. Bredt, D.S. and Snyder, S.H.: No, a novel, neuronal messenger. Neuron, Vol. 8, pp. 3-1 1, 1992.
Bridge, J.H. B., Smolley, J. R. and Spitzer, K W.: The relationship between charge rnovements associated with I calcium and I sodiumîalcium in cardiac myocytes. Science Vol. 248, pp. 378-378, 1990.
Buckner SA. Oheim W. Morse PA. Knepper SM. Hancock M.: Alpha 1- adrenoceptor-induœd contractility in rat aorta is mediated by the alpha I D subtype. European Journal of Pharmawlogy. Vo1.297 N0.3 pp.241-8, 1996.
Bylund, 0. B. : Subtypes of the alpha2adrenocptors: Phamacological and molecular biologieal evidence converge. Trends Pharmac. Sci. Vol. 9 pp. 351- 356,1988.

Bylund, D. B.: P harmacolog ical characteristics of alpha2 adrenerg ic receptor subtypes. Annals of the New York Acaderny of Sciences. Vol. 763,pp. 1-7, 1995.
Bylund, D.B., Eikenberg, D.C., Heible, J.P., Langer, S.Z., Lefkowitz, R.J., Minneman, K.P., Molinoff, PB., Ruffolo, R.R., Jr. and Trendelenburg, U. : International Union of Pharrnacdogy Nomenclature of Adrenoceptors. Phamacol. Rev. Vo1.46, pp. 121 -1 36, 1994.
Bylund, D. B. : Heterogeneity of alpha-2 adrenergic receptors. Phannac. Biochern. Behav., Vol. 22, pp.835-843, 1985.
Bylund, D.B., Ray,-Prenger, C. and Murphy, T.J.: Alpha2A and alpha26 adrenergic receptor subtypes: antagonist binding in the tissues and cell lines containing only one receptor subtype, J. Phanacol. Exp. Ther., Vol. 245, pp. 600-607, 7988.
Bylund D.B., Regan J.W., Faber J.E., Hieble J.P., Triggle C.R. and Ruffolo R.R. Jr.: Vascular alpha-adrenocepton: from the gene to the human. Can. J. of Physiol. & Pharmacol. Vol. 73(5), pp. 533-543, 1995.
Bylund, D.B., Hass, N.A., Ceriutis, D. R., and Blaaxall, H.S.: Characterization of alpha2 adrenergic receptor su btypes. Pharmacol. Commu., Vol., 6, Nos. 1 3 , pp.87-90, 1995.
Champion, H.C. and Kadowitz, P.J.: R-(-)-alpha-methyl-histamine has nitric oxide-mediated vasodilator activity in the mesenteric vascular bed of the cat. Eur J Pharmacol Feb Vol. 19;343 Nos.2-3 pp. 209-216, 1998.
Chataigneau, Tm, Feletou, M., Thollon, C., Villeneuve, N., Vilaine, J.P., Duhault, J. and Vanhoutte, F.M.: Cannabinoid CB1 receptor and endothelium-dependent hyperpoiarization in guinea-pig carotid, rat mesenteric and porcine coronary arteries. Br J Pharmacol Vol. 123 N0.5 pp. 968-974, 1998
Cho, H.J., Xie, Q.W., Calaycay, J., Mumford, R.A., Swiderek, KM,. Lee, T.D., and Nathan, C.: Calrnodulin is a subunit of nitric oxide synthase from macrophages. J. Experimental Medicine.Vo1. 176(Z),ppS9%6O4, 1992.
Codd, E.E., Press, J.B. and Raffa, R: Minireview AIpha2-adrenocepton versus imidazoline receptors: Implications for alpha 2 mediated analgesia and other non-cardiovaswlar therapeutic uses. Lik Sa. Vol. 56, No.2, pp.63-74, 1995.
Corson, M.A., James, N.L., Latta, S.E., Nerem, R.M., Berk, B.C. and Harrison, D.G.: Phosphorylation of endothelial nitric oxide synthase in response to fluid shear stress. Circ Res Vo1.79 No. 5 pp. 984-991.1996.

Chruscinski, A.J., Link, R E , Duant, D.A., Barsh, G-S., and Kobilka, B.K:Cloning and the expression of the mouse homologue of the human alpha2C-adrenergic receptor. Biochem. Biophys. Res. Comm. , Vol. 15, pp. 49-57, 1992.
Cohen, J.A., Baggott, LA., Romano, C., Ami, M., Southerling , T E , Young, L. H., Kozak, C.A., Moniloff, P.B. and Greene, M.I.: Characteriration of a mouse beta 1 adrenergic receptor genomic clone. DNA and Cell Biol. Vol. 12, pp. 537- 547,1993.
Connaughton, S. and Docherty, J. R.: No evidence for differences between pre- and postjunctional alpha2-adrenoceptors in the perip hery. Br. J. P harmacol. Vol. 99, pp. 97-102, 1989.
Corbett, J.A, and Macdaniel, M.L. : Does nitnc oxide mediate autoimmune destruction of beta cells? Diabetes. Vol. 41, pp. 897-903, 1992.
Cotecchia, S., Schwinn, D A , Lefkowitz, R.J., Randall, R.R., Caron, M.J. and Kobilka, B.K.: Molecular doning and expression of the cDNA for the hamster alpha 1 adrenergic receptor, Proc. Natl. Acad. Sci. USA. Vol. 85, pp.7 1 59-7 163, 1988.
Curro F.A. and Greenberg S.: Characteristics of pst-synaptic alpha 1 and alpha 2 adrenergic receptors in canine vascular smooth muscle. Can. J. Physiol. & Pharmacol. Vol.61(8),pp. 893-904, 1983.
Dale, H.H.: On some biological actions of ergot. J. Physiol. Vo1.34, pp.163-206, 1906.
Daniel E.E., Low A.M., Lu-Chao H., Gaspar V. and Kwan C.Y.: Characterization of alpha-adrenoceptors in canine mesentenc vein. J Cardiovasc Pharmacol Vol. 30(5) pp. 591 -598,199.
DeFily D.V. and Chilian W.M.: Coronary microcirculation: autoregulation and metabolic control. Basic Research in Cardiology. Vo1.90(2),pp. 1 1 2-8, 1995.
Defeudis, F.: Excess, EDRFNO , a potentially deleterious condition that may be involved in accelerated atherogenesis and other chronic disease States. J. Immunology, Vol. 2614, pp. 667-679, 1995.
De Ponti F., Giaroni C., Cosentino M., Lecchini S. and Frigo G.: Adrenergic mechanisms in the control of gastrointestinal motility: from basic science to clinical applications. Pharmacology & Therapeutics. Vd.69(1), pp.59-78, 1996.
De Ponti, F.: Phamacotogical critena for the detection of beta 3-adrenoceptors. Trends in Phamacological Sciences. Vol. 18(2),pp.52-53, 1 997.

Dixon, R A F . , Kobilka, B.K, Strader, D.J., Benovic, J.L., Dohlrnan, H.G., Fnelle, T., Bolanowski, M.A., Bennett, G.D., Rnads, e. Diehl, RE., Mumford, R.A., Slater, E.E., Sigal, I.S. , Caran, MG., Lefkowitz, R.J., and Strader, C.D.: Cloning of the gene and cDNA for mammalian beta adrenergic receptor and homology with hoctopsin, Nature, Vol. 321, pp. 75-79, 1986.
Dooley, D.J., Bittiger, H., and Reyman, N.C.: CGP20712A: A useful tool for quantitating beta 1 and beta 2 adrenoceptors, Eur. J. Phamacol. Vol. 130,pp. 137-139, 1986.
Edwards G. and Weston AH.: Pharrnacology of the potassium channel openers. Cardiovascular Drugs 8 Therapy. Vol. 9 Suppl 2, pp., 185-93, 1995.
El-Mas M.M. and Abdel-Rahman AA: Aortic barodenervation up-regulates alpha2-adrenoceptors in the nucleus tractus solitarius and rostra l ventrolateral medulla: an autoradiographic study. Neuroscience Vol. 79(2) pp. 581-590, 1997.
Emorine, L.J., Marullo, S., BriendSutren, M.M.,Patey, G., Tate, K., Delavier- Klutchko, C., and stroberg , A.D.: Molecular characterization of the human Beta 3 adrenergic receptor. Science, Vol. 245, pp. 1 18-1 12 1, 1989.
EmorÎne, L.: The human Psadrenoœptor: the search for a physiological function. Trends Phannacol. Sci.. Vol. 15, pp. 3-7, 1994.
Fabiato, A. and Fabiato, F. : Calcium and cardiac excitation *ntraction coupling, Ann. Rev. Physiol. , Vol. 41, pp. 473484, 1979.
Fagura MS. Lydford SJ. Dougall IG.: Pharrnacological classification of alpha 1- adrenoceptors mediating contractions of rabbit isolated ear artery: cornparison with rat isolated thoracic aorta. British Journal of Pharmacology. Vol. 120 No.2 pp.247-58, 1997.
Fain, J.N., and GarciaSainz, J.A.: Role of phosphotidylinosÎtol turnover in alpha 1 -and of adenyîyl cyclase inhibition in alpha2-effects of catecholamines. Life Sci. ,Vol. 26 pp. 1 183-1 194, 1980.
Feuerstein,G.Z. and Ruffolo, R.R.Jr.: Carvedilol, a novel vasodilating beta- blocker with the potentbl for cardiovascular organ protection. European Heart Journal. Vol. 1 7 Suppl BI pp. 24-29, 1996.
Flavahan, N.A.: Atherosderosis or lipoprotein induceâ endothelial dysfundion: Potential mechanisrns underlying reductîon in EDRFINO actMty. Circulation, Vol. 8515, pp. 1927-1938, 1992.
Flavahan, N A and Vanhoutte, P.M.: alpha1 adrenoceptor subclassification is vascular smooth muscle. Trends in phannacul. Sci. Vol. 7, pp. 347-349, 1986.

Ford, A.P.D.W., Williams, T.J., Blue, DR., and Clarke, D E : Alpha1 adrenoceptor classificaüon: Sharpening Occam's razor. Trends. Pharmacol. Sci. Vol. 1 5 , ~ ~ . 167-1 70, 1994.
Forster C., Drew G.M., Hilditch A. and Whalley ET.: Dopamine receptors in human basilar artenes. Eur. J. Pharmacol. Vol. 87(2-3),pp. 22735, 1983.
Forster C. and Armstrong P.W.: Pacing-inducad heart Mure in the dog: evaluation of peripheral vascular alpha-adrenoceptor subtypes. J.1 Cardiovascular Pharmacul.Vo1. 16(5),pp.708-718, 1 990.
Forster C., Naik G. and Armstrong P.W.: Noradrenaline biosynthesis and metabolism during development and recovery from pacing-induced heart failure in the dog. Can. J. Physiol. & Pharmacol. .Vol. 72(1),pp.45-599, 1 994.
Forsterman, U., Schmidt, H.H.H.W., Pollack, J.S., Sheng, H-, Mitchell, J.A., Wamer, T.D., Nakane, M. and Murad, F.: Isofoms of nitric oxide synthase. Biochern. Pharmacol. Vol. 42, pp. 1849-1 857, 1991.
French, N.: Alpha2 adrenocepton and 12 sites in the marnmalian central nervous system. Pharmacol. Ther. Vol. 68, No.2. pp. 175-208, 1995.
Frielle, T., Collins, S., Daniel, K.W., Caron, MG.. Lefkowitz. R.J. and Kobilka. B.K.: Cloning of the cDNA for the human beta 1 adrenergic receptor. Proc. Natl. Acad. Sci. USA, Vol. 84, pp. 7920-7924, 1987.
Fur~hg~t, R.F. and Zawadski, P.M.: The obligatory role of the endothelial cells in the relaxation of arterial smooth muscle by acetylcholine, Nature, Vol. 288, pp. 373-376, 1 980.
Fulton B., Wagstaff AJ. and Sorkin E.M.: Doxazosin. An update of its dinical pharmacology and therapeutic applications in hypertension and benign prostatic hyperplasia. Dnigs. Vol. 49(2), pp.,295-320, 1995.
Galitrky J., Carpene C.. Bousquet-Melou A.. Berlan M. and Lafontan M.: Differential activation of beta 1-, beta 2- and beta 3-adrenoceptors by catecholamines in white and brown adipocytes. Fundamental & Clinical Pharmacol. Vol.9(4), pp.324-331, 1995.
Garcia-Sainaz, J.A.: Deueased sensitivity to al p haîadrenerg ic amines , adenosin and prostaglnsind in white fat cells from hamster treated with pertussis vaccine. , FEBS, Lett. Vol. 1 26, pp. 306-308, 1 98 1.

Garthwaite, J.: NWc oxide synthesis linkeâ to activation of excitatory neurotransmitter receptors in the brain. In: Nitric Oxide from L-arginine : a bioregulatory systern , 4 s . Moncada, S. and Higgs E.A. , pp. 1 15-1 37, Elsevier, Amsterdam, 1990.
Gauthier, C., Charpentier,F., Laurent, K. and Trochu, J-E.: Pharmacolagical evidence for the presence of psadrenoceptors in human ventricular cells: ( Abstract 3065), Circulation Vol. 92. pp. 639, 1995.
Gauthier, C., Travemier, G., Charpentier, F., Langin, O. and Le Marec, H.: Funcüonal P3-adrenergic receptor in human heart. J. Clinc. Invest. Vol. 98, pp. !%6-562., 1 996.
Germack, R., Starzec, AB., Vassy, R. and Perret, G.Y.: Beta-adrenoceptor subtype expression and funcüon in rat white adipocytes. British J. Pharmacol.. Vol. 120(2),pp.201-10, 1997.
Germack, R., Starrec, A.B., Vassy. R., and Perret, G.Y.: Beta-adrenoceptor subtype expression and function in rat white adipocytes. British J. Phanacol. Vol. 120(2),pp.201-21 O, 1997.
Ghaleh, B., Bea, M L , Dubois-Rande. J.L., Giudicelli, J.F ., Hittinger, L. and Berdeaux, A.: Endothelial modulation of beta-adrenergic dilation of large coronary artenes in conscious dogs. Circulation.Vol. 92(9),pp.2627-2635, 1995.
Giacobino. 1-P. : P3-adrenocepton. An Update, Eur. J. Endocrinol. Vol. 132, pp.377-385,1995.
Goto M., VanBavel E., Giezeman M.J. and Spaan J.A: Vasodilatory effect of pulsatile pressure on coronary resistance vessels. Circ. Research. Vol. 79(5),pp. 1039-45, 1996.
Govyrin V.A. and Leont'eva G.R.: r h e reactivity of the mesenteric arteries and vein to exposure to endogenous noradrenaline in normal rats and rats with DOCA-salt hypertension]. [Russian] Fiziologicheskii Zhumal lmeni I . M. Sechenova. Vol. 80 No. 1 1 pp. 14,1994.
Graham, R.M., Perez, D-M., Hwa, J. and Piascik M.T.: Aiphai-adrenergic receptor subtypes , Molecular structure, fundion and signalling. Circ. Res.Vo1.78, NO. 5, pp.737-749, 1996.
Grant J.A. and Scnitton M.C.: Interaction of selectÎve alpha-adrenoceptor agonists and antagonists with human and rabbit blood platelets. British J. of Phannacol. Vol. 71 (?)$p. 121 -1 34, 1980.

Grassby P.F. and Broadley KJ.: Responses mediated via beta-1 adrenoceptors but not beta-2 adrenoceptors exhibit supersensitivity after chronic reserpine pretreatment. J. Phanacology & Experimental Therapeutics. Vol. 237(3),pp. 950-958, 1986.
GrifTiths, M.J.D., Messent, M.. MacAllister, R.J. and Evans, T.W.: Aminoguanidine selectively inhibas iNOS. Br. J. Pharmacol. Vol. 110, pp. 963- 968, 1993.
Gross, S.S., Jaffe, E.A,. Levi, R. and Kilboum, R.G.: Cytokine-activated endothelial cells express an isotype of nittic oxide synthase which is tetrahydrobiopterin-dependent, calmodulin-independent and inhibited by arginine analogs with a rank-order of potency characteristic of activated macrophages. Biochemical & Biophysical Research Communications. Vol. 1 78(3),pp. 823-629, I W I .
Guarino, RD., Perez, D.M. and Piascik, M.T.: Recent advances in the molecular phamacology of the alphal ~adrenergic receptors, Cell Signal, Vol.8, pp. 323-333, 1996.
GUO, J.P., Murohara, T., Buerke, M., Scalia, R. and Lefer, AM.: Direct measurement of nitric oxide release h m vascular endothelial cells. J Appl Physiol Vo1.81 Na2 pp. 774-779, 1996.
Guney, H.Z., Hodoglugil, U., Uluoglu, C., Gorgun, C.Z., Ercan, Z.S., Abacioglu, N. and Zengil, H.: In vitro susceptibility rhythrns. II. Biological-time-dependent differences in effect of beta 1- and beta 2-adrenergic agonists of rat aorta and influence of endothelium. Chronobiol Int Mar Vol. 15 No. 2 pp. 159-1 72, 1998.
Hadjilam bris, O.W., Skwish, S. and Dickinson, K E. J. : DifFerent Pharmacological characteristics of CHO cells transfected wlh mouse and human beta3 adrenergic receptors. FASEB J. Vol. 8, pp. A354 1994.
Hall, LE., Guyton, A.C. and Brands, M.W.: Pressure-volume regulation in hypertension. [Review]. Kidney International - Su pplement. 55, pp.S3541, 1 996.
Halpern, W., Osol, G. and Coy, G.: Mechanical behavior of pressurized in vitro prearteriolar vessels determined with a video system. Ann. Biomed. Eng. Vol. 1 2, pp. 463479,1984.
Haslam, R.J.: Role of cyclic nucieotides in platelet funcüon. ClBA Found. Symp. Vol. 25, pp. 121 -1 SI, 1975.
Han, C., Abel, P.W., and Minneman,, ICP. : Heterogeneity of alphal-adrenergic recepton revealed by chlorethylclonidine. Molec. Pharmac., Vol. 32, pp. 505-510 1987-

Han C., Li J. and Minneman K.P.: Subtypes of alpha 1-adrenoceptors in rat blood vessels. Eur J Phannacol Vol. 6 Nos. 190(1-2) pp. 97-1 04, 1990
Harrison, J.K., D'Angelo, D.D., Zeng, D., and Lynch, KR.: Phamacological characterization of rat alpah2adrenergic receptors. Mol. Phanacol. , Vo1.40, pp. 407-4 12,199 1.
Head G A : Central monoamine systems and new antihypertensive agents. Clinical8 Experimental Hypertension. Vol. 1 7(1-2), pp. 141 -1 52, 1 995.
Hedberg, A. and Matisson, H.: Beta adrenoceptor interaction of full and partial agonists in the cat heart and the soleus muscle. J. Pharmacol. Exp. Ther. Vol. 21 9, pp. 798-808, 1981.
Hibbs, J.B.. Taintor, R.R., Vavrin, 2. and Rachlin, E.M.: Nitric oxide: a cytotoxic activated macrophage effector molecule. , Biochem. Biophys. Res. Corn m. Vol. 1 5 7 , ~ ~ . 87-94, 1988.
Hibbs, J. B., Vavrin, 2. and Taintor, R. R. : L-argininelN0 pathway expression of the activated macrophages effedor mechanism can cause metabolic in hibition in the target cells. J. Immunology, Vol. 138, pp. 550-565, 1990.
Hibbs, J.B., Corbett, J.A., Sweetland, M.A. and Macllaniel, M.L.: 11-18 induœd formation of EPR- detectable iron-nitmsyl complexes in islets of Iangerhans. J. Biol. Chemistry, Vol. 266, pp. 21 351 -21 354, 1991.
Hieble,J.P., Sulpizio, A C , Nichols, A.J., Dernarinis, R.M., Pfeiffer,F.R., Lavanchy, P.J., and Ruffolo, R.R.,Jr. Phamacological differentiation of pre and postjunctional alpha2-adrenoœptors., J. Hypertens., V0l.4,Suppl.G~ pp. SI 89- S192, 1986.
Hieble, J.F., Bylund, D.B., Clarke, D.E., Eikenburg, D.C., Langer, S.Z., Lefkowitz, R.J., Minneman, K.P., and Ruffolo, R.R. Jr.: International Union of Phamacology, X: recommendation for nomendature of alphaladrenocepton: consensus update. Pharmacol. Rev. Vol. 47, pp. 267-270, 1995.
Hieble, J.P., Ruffolo, R.R. Jr., Sulpizio, AC., Naselsky, D.P., Conway, T.M., Ellis, C., swift. AM., Ganguly, S. and Bergsma, D.: Functional subclassification of alpha2-adrenoceptors. Pharmacol. Commun. Vol.6, Nos. 1-3, pp.91-97, 1995.
Hieble, J. P and Ruffolo, R. R. Jr.: Subclassification of the beta-adrenoceptors. Pharmacol. Commun. Vol. 6 , Nos. 1-3, pp. 183-193, 1995.

Hieble J.P., Bondinell W.E. and Rufblo R.R. Jr.: Alpha- and beta-adrenoceptors: from the gene to the clinic. 1. Molecular biology and adrenoceptor subclassification. J. of Med. Chem. Vol. 38(18),pp.3415-3444, 1995.
Hill R.A. and Sillenœ M.N.: lmproved membrane isolation in the purification of beta 2-adrenoceptors from transgenic Escherichia coli. Protein Expression & Purification. Vol. 10(1),pp. 162-1 67, 1997.
Hollenga, C. and Zaagsma, J.: Direct evidence for the atypical nature of fundional beta adrenoceptors in the rat adipocytes. Br. J. Phamacol., Vol. 68, pp. 1420-1 424, 1989.
Huang, Y. and Chan, N.W.: Involvement of endothelium in relaxant action of glibenclamide on the rat mesenteric artery. Eur J Pharmacol Vol. 5; 343 No.1 pp. 27-33, 1998.
Hussain M.B. and Manhall 1.: Charaderization of alphel-adrenoceptor subtypes mediating contractions to phenylephrine in rat thoracic aorta, mesenteric artery and pulmonary artery. Br. J. Phanacol. Vol. 122 No. 5 pp.849-858 1997.
Hwa, J.J., Ghibaudi, L., Williams, P. and Chattejee, M.: Cornparison of acetylchoiinedependent relaxation in large and small artenes of rat rnesenteric vascular bed. Am J Physiol Vol. 266 Nos. 3 Pt 2 pp. H952-H958, 1994.
Hyre, C.E., Unthank, J.L. and Daising, M.C.: Direct in vivo measurement of fiow- dependent nittic oxide production in mesenteric resistance arteries. J Vasc Surg Vol. 27 No.4 pp. 726-732, 1998.
lakovidis, D., Malta, E., Mcpherson, G.A. and Raper, C.: In Vitro activity of RO 363, e betal adrenoceptor seledive agonist. Br. J. Pharmacol. Vol. 68, pp. 677- 685, 1980.
lismaa, T. Biden, T.J., and Shine, J.: G proteincoupled recepton. New York, CRC Press. 1994.
Ikeda, U., Furuhashi, K, Kanbe, T. and Shirnada, K.: Ouabain enhances nitric oxide synthesis in rat vascular smooth muscle cells induced by interleukin-1 M a . Eur J Phamacol Vol. 15;288 No.3 pp. 379-383 1995.
Ikeda, U., Kurosaki, K., Ohya, K. and Shimada, K.: Adenosine stimulates nitric oxide synthesis in vasarlar smooth musde cells. Cardiovasc Res Vol. 35 No. 1 pp. 168-1 74 ,1997.
lpsen M., Zhang Y., Dragsted N.. Han C. and Mulvany M.J.: The antipsychotic drug sertindole is a specific inhibitor of alphalA-adrenocepton in rat mesenteric small arteries. Eur J Phamacol Vol. 336 No. 1 pp. 29-35 1997.

Iranami, H., Hatano, Y., Tsukiyarna. Y., Maeda, H. and Mizumoto, K.: A beta- adrenoceptor agonist evokes a nitric oxide-djMP relaxation rnechanism modulated by adenylyl cydase in rat aorta. Halothane does not inhibit this mechanism. Anesthesiology Vol. 85 No. 5 pp. 1 1 29-1 1 38, 1 996.
ltoh H., Kohli J.D. and Rajfer S.1.: Phanacdogical characteriration of the postsynaptic alpha-adrenoceptors in isolated canine mesenteric arteries and veins. Naunyn Schmiedebargs Arch Pharmacol Vol. 335 No. 1 pp. 44-49, 1987.
Jenkinson D.H., Barnard E.A., Hoyer D., Humphrey P.P., Leff P. and Shankley N.P.: International Union of P harmacology Cornmittee on Receptor Nomenclature and Drug Classification. IX. Recornmendations on ternis and symbols in quantitative phamacology. Pharrnacological Reviews. Vo1.47 (2),pp. 255-266, 1 995.
Kaneko K. and Sunano S.: lnvolvement of alpha-adtenocepton in the endothelium-dependent depression of noradrenaline-induced contraction in rat aorta. Eur J Pharmacol Vo1.24240 Nos.2-3 pp. 195-200, 1993.
Kannan M.S. and Seip A.E.: Neurogenic dilatation and constdction of rat superior mesentefic artery in vitro: mechanisms and meâiators. Can J Physiol Phannacol Vol. 64 NO. 6 pp. 729-736, 1986.
Kam K.L., Pfaffendorf M. and van Zwieten P.A.: Pharmacudynamic behavior of isolated resistance vessels obtained from hypertensivediabetic rats. Fundamental & Clinical Pharmawlogy. Vol. IO(4),pp.329-36, 1 996.
Kamata, K., Miyata, N. and Kasuya, Y.: lnvolvement of endothdial cells in relaxation and contadion responses of the aorta to isoproterenol in naive and streptozotocin-induced diabetic rats. J Pharmacol Exp Ther Vol. 249 No. 3 pp. 890-894, 1989
Katada, T. and Ui , M.: Slow interaction of islet acüvating protein with pancreatic islets during prirnary culture to cause reversal of alpha -adrenergic inhibition of insulin secretion. J. Biol. Chem. , Vol. 255. pp. 950-958, 1980.
Kaumann, A. J. : Adrenergic receptors in heart muscle. Two different mechanisms of P-blockers as partial agonists. International Union of Biochemsitry, Symposium 52, Acta Physiol. LatAm vol. 23, pp., 235-236, 1973.
Kaumann, A.J.: 1s there a third heart p-adrenoceptor? Trends Pharmacol. Sci. Vol. 1 O, pp., 31 6-320, 1 989.

Kaumann, A.J., Blinks, J.R.: p-Adrenoceptor blocking agents as partial agonists. Dissociation of stimulation and blockade. Naunyn-Schmeide berg's Arch. Pharmacol. Vd., 31 1, pp., 237-248, 1 980b.
Kaumann, A.J. and Lemoine, H.: Direct labeling of rnyomrdial beta one adrenoceptors: Cornparison of binding affinity of 3~-(-)-bisoprolol with its blocking potency. Naunyn-Schmiedeberg's Ardi. Pharmacol.Vol. 331, pp. 27-39, 1985.
Kaumann, A.J. and Molenaar, P.: Modulation of human cardiac function through four beta adrmoceptor populations, Naunyn-Schmiedeberg's &ch. Pharmacoi.Vol.355, pp. 667-681, 1997.
Kenny, B.A.. C halrners, D.H., Philpott, P.C. and Naylor, A.M.: Characterization of an alpha ID-adrenoceptor mediating the contractile response of rat aorta to noradrenaline. British Journal of Pharmacology. Vol. 1 7 5 No.6 pp.98l-6, 1995.
Kichuk, M.R., Seyedi, N., Zhang, X., Marobe, C.C., Michler, RE., Kaley, G., Nasjletti, A. and Hintze T.H.: Regulation of NO production in human wronary microvesseles and the contribution of local kinin formation. Circulation, Vol. 94(1), pp., 44-51, 1996.
Kilboum, R.G., Gross, S.S., Jurban. A., Adams, J., Griffith, O.W., Levi, R. and Lodato, R.F.: N6-methyl-L-arginine inhibits Tnf induced hypotension : implications for the involvement of nitric oxide . P m . Natl. Acad. Sci. USA, Vol. 87, pp. 362903632, 1990.
Kilbourn, R. G., Klostergaard, J. and Lopez-Berestein, J. : Activated macrophages secrete a soluble factor that inhibits mitochondrial respiration of turnor cells. J. Immunology, Vol. 133 , pp. 2577-2581, 1984.
Kong J.Q., Taylor D.A. and Fleming W.W.: Antagonism of nompinephrine by donidine in the isolated rat mesenteric vascular bed. J Pharmacol Exp Ther Vol. 259 N0.2 pp. 653-658, 1991.
Kuchan, M.J., JO, H. and Frangos, J.A.: Role of G proteins in shear stress- mediated nitric oxide production by endothelial cells. Am J Physiol Vo1.267 Nos3 Pt 1 pp. C753-C758,1994.
Lands, AM., Arnold, A, McAullif, J.P., Luduena, F.P. and Brown, T.J.: Differentiation of receptor systems by sympathornemetic amines. Nature Vol. 214, pp. 597-598, 1967.
Langer. S. 2.: Presynaptic regulation of catecholamine retease. Biochem. Pharmacol. Vol. 23, pp. 1793-4 8OO1I99O.

Lanier, S.M. : Regulaüon of alpha2-adrenergic receptor signaling, Phancol. Comm. Vol. 6, Nos.1-3, pp. 133-137, 1995.
Larosa,G.: Altered regulation of coronary vascular tone in exparimental heart failure: Modulation by nitnc oxide , nitrovasodilators, and alpha adrenoceptors, Ph.D. Thesis, Department of phamacology, University of Toronto, 1997.
Laz, T.M., Forray, C., Smith, K., Bard, J.A, Vaysee, P.J.J., Branchek, T.A., and Weinshank, R.L.: The rat homologue of the bovine alpha1 C-adrenergic receptor shows the pharmacolagical properties of the classical alphalA subtype. Mol. Pharmacol. Vol. 46, pp. 414-422, 1994.
Leeb-Lundberg, L.M.F., Dickinson, K.€. Jel Heald, S.L., Wickberg, J.E.S., Hagen, P.D., Debemardis, J.F., Wnn, M., Andersen, D L , Lefkowitz, R.J. and Caron, M.G. : Photoafinity labeling of mamrnalian alpha1 adrenergic receptors. J. Biol. Chem. Vol. 259, pp. 2579-2587, 1984.
Leeb-Lundberg, L.M., Cotecchia, S., Hagen, P.O., Lefkowitz, R. J. and Caron MG.: Phorbol ester effects nn alpha 1-adrenoceptor binding and phosphotidylinositol metabolism in cultured vascular smooth muscle œlls. Life Sciences.Vo1.37(25),pp.2389-2398, 1985.
Leone, AM., Palmer, R.M.J., Knowlen, R.G., Francis, P.L., Ashton, D.S., and Moncada, S.: Constitutive and inducible nitnc oxide synthases are 1-arginine N6- C6- dioxygenase. J. Biol. Chemistry, Vol. 1 3, pp. 424-430, 1 991 .
Lemoine, H., Ehle, B. and Kaumann, A.J.: Direct labeling of beta 2 adrenoceptor: Cornparison of the binding potency of 3 ~ - ( - ) - 1 ~ ~ 1 15551 and blocking potency of ICI 1 7 8,551. Naunyn-Schmiedeberg's Arch. Pharmacol.Vol.331, pp.40-51,1985.
Limbird, LE., Macmillan, L.B., and Keffer J. R.: Specificity in alpha2- andrenoceptor signal transduction: receptor subtypes: Coupling to distinct signal transduction pathways and tocalization to discrete subdomains in target cells. Pharmacol. Commu. Vol6 , Nos 1-3, pp. 139-145, 1995.
Link, R., Duant, B., Barsh, J., Chruscinski, AJ. and Kobilka, B.: Clonning of two mouse genes encoding alpha2-adrenergic receptor subtypes and identification of a single amino acid in the mouse alpha2C-homologue responsible for interspecies variation in antagonist binding. Mol. Pharmacol. Vo1.42, pp. 16- 27,1992.
Lip, G.Y. and Femer, RE.: Poisoning with antihypertensive dnigs: alpha adrenoceptor antagonists. J. Human Hypertension.Vol. 9(7), pp.523-6, 1995.

Lipton, S.A., Choi, Y.B., Pna, Z.H., Lei, S.Z., Chen, H.S.. Sucher, NA., Loscaizo, J., Signel, D.J. and Stamler, J.S.: A redox based mechanism, for the neuroprotedive and neurodestructive effects of NO and related nitroso compounds . Nature, Vol. 364, pp. 626-631, 1993.
Lomasney, J .W., Cottechia, S., Lefkowitz, R. J . , and Caron, M. J.: Molecular biology of the alpha-adrenergic receptors : Implications for receptor classification and for structure-function relationships. Biochem. Biophys. Acta. Vol 1095, pp. 127-139, 1991.
Lomasney, J.W., Cottechia, S., Lorenz, W.,Leung, W.Y., Schwinn, D.A., Yang- Feng, T.A., Brownstein, M., Lefkowitz, R.J. and Caron MG.: Molecular cloning and expression of the cDNA for the alpha? A adrenergic receptors, J. Biol. Chem. V01.266,pp.6365-6369, 1991.
Lurie KG., Tsujimoto G. and Hofhan B.B.: ûesensitization of alpha4 adrenergic receptor-mediated vascular smooth muscle contraction. J. Pharmacol. & Experimental Therapeutics. Vol. 234(1),pp. 147-1 52, 1985.
Macdonald, P.S., Read, M.A., and Dusting, G.J.: Synergistic inhibition of platelet aggregation by endothelium derived relaxing factor and prostacyciin : Thrombosis Res. Vol. 49. pp. 437-449, 1988.
Machida, C.A., Bunzow, J.R., Searies, R.P., Van Tol. H., Tester, B., Neve, K.A. Teal, P., Nipper, V., and Civelli, O.: Molecular cloning and expression of the rat beta 1 adrenergic raceptor gene, J. Biol. Chem, Vol. 265,pp. I296û-lZ96S1 1990.
Manara, L., Badone, O., Baroni, M., Boccardi, G., Croci, T., Giudice, A., Guni, U. and Le Fur, G. : AryloxypropanolaminotetraIins are the first selective antagonists for atypical beta3 adrenoceptors., Phannacol. Commu. Vol. 6, Nos. 1-3, pp. 253- 258, 1995.
Maramatsu, I . , Ohmura, T., Hashimoto, S. and Oshita, M.: Functionai subclassification of vascular alpha 1 adrenoceptors, Pharmacol. Commu. Vol. 6, Nos. 1-3, pp. 23-28, 1995.
Martinotti E. Breschi MC. Omini C. Pani M. Ciucci MA. Nieri P.:Cumparative study of pwtsynaptic alpha-adrenoceptors in aorta obtained from human and other rnammalian species, Pharmacdogical Research. Vol. 23 NO.lpp.57-69, 1993.
Martinez-Cuesta, M.A., Moreno, L., Pique. J.M., Bosch, J., Rodrigo, J. and Esplugues, J.V.: Nitnc oxîde-mediated beta 2-adrenoceptor relaxation is impaired in mesenteric veins from portal-h ypertensive rats. Gastroenterology Vol. 1 1 1 No.3 pp. 727-735, 1996

Matsuda K., Sekiguchi F., Tojo M., Shirnamura K. and Sunano S.: Effects of NG- nitro-1-arginine on alpha-agonists-induced contraction of aortae from Wistar Kyoto rats and stroke-prone spontaneously hypertensive rats. J Smooth Muscle Res Vo1.31 No.2 pp. 51-60, 1995.
McCall, T.B., Feelisch, M., Palmer, R.M. and Moncada, S.: Identification of N- iminoethyl-L-ornlhine as an irrevenible inhibitor of nitric oxide synthase in phagocytic cells. British J. Phamacol. Vol. 1 02(1), pp.234-238, 1991.
Meissner, G.: Ryanodine receptors I calcium release channels and their regulation by endogenous eff&tors. Ann. Rev. Physiol. , Vol. 485-508, 1994.
Merdier, J.J., Lompre, A.M., Duc, P., Boheler, K.R., Fraysse, J.B., Wknewsky, C., Allen, P.D., Komajda, M. and Schwartz, K.: Altered sarcoplasmic reticulurn calcium ATPase gene expression in the human ventride during end stage heart failure. J. Clin. Invest. , Vol, 85, pp. 305-309, 1990.
Mills, D.C.B.: Initial biochemical responses of platelets to stimulation. ClBA found. Symp. Vd. ,35, pp. 1 53-1 67,1975.
Minneman, K.P., and Esbenshade, T.A.: Alphal-adrenergic receptor subtypes. Annu. Rev. Pharmacol. Toxicol., Vol. 34, pp. 1 17-1 33, 1994.
Minneman, K. P. : Alphal ~drenergic receptor subtypes , inositol phosphates and sources of ceIl calcium. Pharmacol. Rev. Vol. 40, pp. 87-1 19, 1988.
Moran, N.C. and Perkins, M.E.: Adrenergic blockade of the mammalian heart by a dichloro analogue of isoprenaline. J. Phannacol. Exp. Ther. Vol. 124, pp. 223- 237, 1958.
Mitchell G.F., Pfeffer J.M. and Pfeffer MA.: The heart and conduit vessels in hypertension. Medical Clinics of North America. Vol., 81 (6), pp. 1247-7 1, 1997.
Mo~~ow, A.L. and Creese, 1. : Characterization of alphalsdrenocaptor subtypes in rat brain: a reevaluation of [ 3 ~ ] ~ ~ 4 1 0 4 and [ 3 ~ ] p m s i n binding. Mol. Pharmacol. Vo1.29, pp. 321-330, 1986.
Moncada, S., Palmer, R.M.J. and Higgs, E.A.: Nitric Oxide : Physiology , pathophysiology and pharmacology. Phamacol. Rev. Vol. 43, pp. 109134, 1991.
Muniyappa, R-, Walsh, M.F., Rangi, J.S., Zayas, R.M., Standley, P.R., Ram, J.L. and Sowers, J.R.: lnsulin like growth factor 1 increases vascular smooth muscle nitric oxide production. Life Sci Vol.61 No. 9 pp. 925-931 1997.

Muraki, Tel Nakaki, T., and Kato, R.: Predominance of alphaBadrenacsptors in porcine thyriod: biochemical and pharmacological correlations. Endocrinology, Vol. 114, pp. l645-I6Sl1 1984.
Murphy, T.J. and Bylund, D.B.: Characteriration of alpha2-adrenergic receptors in the OK cell, an oposum kidney cell 1ine.J. Phannacol. Exp. Ther.Vo1.244, pp.57 1-578,1988.
Muuin, P., Revelli, J.P., Kuhne, F., Gocayne, J.D., McCombie, W.R., Venter, J-C., Giocobino, J.P. and Fraser, C.M.: An adipose tissue specific beta adrenergic receptor. J. Bol. Chem. Vol. 266 , pp. 24053-24058, 1991 .
Myers, P.R., Banitt, R.F., Guerra, R.J., Bates, N.J., and Harrison , D.G.: Characteristics of canine wronary resistance arteries: Importance of endothelium. Am. J. Physiol. Vol. 257, (Heart Cire. Physiol. 26): H603-H610, 1989.
Nakaoka, H. Perez, D.M., Baek, K-J., Das, T., Husain, A., Misono, K., lm, M.J. and Graham, R.M.: A GTP-binding protein with transglutaminase activity and receptor signaling function. Science Vol. 264, pp. 1593-1 596, 1994.
Nakamura T., Saito Y., Kato T., Sumino H., Hoshino J., Ono Z., Sakamaki T. and Nagai R.: Flow-mediated vasodilation of a condul artery in relation to downstream peripheral tissue blwd flow during reactive hyperemia in humans. Japanese Circulation J. Vol. 61 (9), pp.772-80, 1 997.
Nakatsubo, N., Kojima, H., Kikuchi, K., Nagoshi, Hel Hirata, Y., Maeda, D., Imai, Y., Mmura, T. and Nagano, T.: Direct evidenœ of nitric oxide produdion from bovine aortic endothelial cells using new fluorescence indicators: diaminofluoresceins. FEBS Lett Vol. 427 No.2 pp. 263-266 1998.
Natio, K., Nagao, T., Otsuka, M., Harigays, S. and Nakajima, H.: Studies on the affinity and selectivity of denopamine (TA-064) , a new cardiotonic agent, for beta adrenergic receptor. Japan J. Phannacol. Vol. 38, pp. 235-241, 1985.
Nebigil, C. and Malik, KU.: Cornparison of signal transduction mechanisms of alpha2C adrenergic receptor-stimulated prostaglandin synthesis. J Pharmacol. Exp. Ther. Vol. 263, pp. 987-996,1992.
Nielsen H., Pilegaard H.K, Hasenkam J.M., Mortensen F.V. and Mulvany M.J.: Heterogeneity of postjundional alpha-adrenacepton in isolated rnesentenc resistance arteries from rats, rabbits, pigs, and humans. J Cardiovasc Pharmacol Vol. 18 NO. 1 pp. 4-1 0, 199.1

Nichols, AJ., Motley, E.D. and Ruffolo, R.R. Jr.: Differential effect of pertussis toxin on pre- and postjunctional alpha2 adrenoceptors in the cardiovascular system of the pithed rat. Eur. J. Pharmacol. Vol. 145, pp. 345-349, 1988.
Nichols, A.J., Motley, E.D. and Ruffolo, R.R. Jr.: Effect of pertussis toxin on treatment on postjunctianal alpha1 and apha2 adrenacsptor function in the cardiovascular system of the pithed rat. J. Phanacol. Exp. Ther. Vol. 249, pp. 203-209, 1 989.
Nuttall, A., and Snow, H.M.: The cardiovascular effects of ICI 118, 587: A Betal adrenoceptor partial agonist. Br. J. Pharmacol., Vol. 77, pp. 381 -388, 1982.
Ogbonna, B.C., Okeahialam, B. N. and Ramyil, V.M.: alpha-receptor blockade for benign prostatic hyperplasia. British J. Urology. VoI.79(1), pp.32-5, 1 997.
Okamura, T., Uchiyama, M., An, J. and Toda, N.: Nitnc oxidenediated neurogenic relaxation in rnonkey mesenteric veins. Jpn J Pharmacol Vo1.68 No. 4 pp. 405-41 1,1995.
Oriowo, M.A.: Beta(3)-adrenoœptorç mediate smooth muscle relaxation in the rat lower oesophageal sphincter. J. Autonomie Pharmacology. Vol. 17(3),pp.175- 182, 1997.
Oriowo, M.A.: Different atypical Ma-adrenocepton mediate isoprenaline- induced relaxation in vascular and non-vascular smooth muscles. Life Sciences. Vol. 56 NO. 15 pp. PL269-75, 1995.
Palmer, R.M.J., Femdge, A.G. and Moncada, S.: Nitric oxide release accounts for the biological activity of endothelium derived relaxing factor. Nature, Vol. 327, pp. 524-526, 1987.
Parker, RB. and Waud, D.R.: Pharmacological estimation o f drug-receptor dissociation constants. Statistical evaluation. J. Pharmacol. Exp. Ther. Vol. 177, pp. 1-12, 1971.
Parkington, H.C., Tare, M., Hamma~strom, A.K.M., Tonta, M.A. and Coleman, H.A.: The role of endothelium-derivecl proctacyclin in regulating tone in the vascular smooth muscle. Endotheliumderived hyperpolaning factor, Harwwd academic publishen, The Netherlands, pp.57-64,1996.
Peredo, HA, Feleder, E.C. and Adler-Graxhinsky, E.: Differential effects of acetylcholine and bradykinin on prostanoid release from the rat mesentek bed: role of endothelium and of nitric oxide. Proçtaglandins Leukot Essent Fatty Acids Vol. 56 No.4 pp. 253-258, 1997.

Perez, D.M., Piascik, M.Y., Malik, N., Gaivin, R., and Graham, R.M.: Cloning , expression and tissue distribution of the rat homolog of the bovine alphalc- adrenergic receptor provide evidence for its classification as the alphalA- subtype, Mol. Pharmacol. Vol. 46, pp.823-831, 1994.
Perez, D-M., Piascik, M.T. and Graham, R.M.: Solution-phase library screening for the identification of rare clones: isolation of an alpha-adrenergic receptor cDNA. Mol. Pharmacol. Vol. 40, pp. 876-883, 1991.
Petenson, J., Zygmunt, P.M. and Hogestatt, E. D. : Characterization of the potassium channels involved in EDHF-mediated relaxation in cerebral artenes. British J. Phamacol. Vol. 1 20(7),pp. 1 344-1 350, 1997.
Piascik M.T., Hrometz S.I., Edelmann S.E., Guarino R.D., Hadley R.W.and Brown R. D.: lmmunocytochemiml localization of the alpha-1 b adrenergic receptor and the contribution of this and the other subtypes to vascular smooth muscle contraction: analysis with selective ligands and antisense oligon ucleotides. J. Phamacol. Exp. Ther. Vo1.283 No.2 pp.854-868 1 997.
Piascik, M.T., Butler, B.T., Kusiak, J.W., Pitha, J. and Holtman, J.R. : Effect of an alkylating analog of prazosin on alphal-adrenoceptor subtypes and arterial blood pressure. J. Phanacol. Exp. Ther. Vol. 251 ,pp. 878-883, 1989.
Piascik M.T., Soltis E.E., Piascik M. M. and Macmillan L.B.: Aipha-adrenoceptors and vascular regulation: molecular, pharmacologie and clinical correlates. Pharmacology & Therapeuücs. Vol. 72(3), pp. 21 5-241, 1996.
Pietri-Rouxel, , F., Dnimare, M.F. and Strosberg, A. D.: Pharmacological characterization of beta3 adrenergic receptor. Phannacol. Commu. Vo16, Nos. 1 - 3, pp. 229-236, 1995.
Pittman, R.N., Reynolds, E.E., and Monilof, P.B.: Relationship between intrinsic adivities of agonists in normal and desensitized tissue and agonist induced loss of beta adrenergic receptors. J. Pharmacol. Exp. Ther. Vo1.230, pp. 614-61 8, 1984.
Phan, L., Hickey, M., Niazi, Z., and Alastair, S.G.: Nitric oxide synthase inhibitor, Nitro-iminoethyl-lornithine , reduces ischemia- reperfusion injury in rabbit skeletal muscle, Microsurgery, Vol. 1 5, pp. 703-707, 1994.
Powell, C.E. and Slater, I.H.: Blocking of inhibitory adrenergic receptors by a dichloro analogue of isoprenaline. J. Phannacol. Exp. Ther. . Vol. 122, pp. 480- 488,1957.

Pueyo, M.E., Amal J.F., Rami, J. and Michel, J.B.: Angiotensin II stimulates the production of NO and peroxynitrite in endothelial cells. Am J Physiol Vol. 274 NOS. 1 Pt 1 pp. C214-C220 1998.
Quast U.: ATP-sensitive K+ channels in the kidney. Naunyn-Schmiedebergs Arch. of Phannacol, Vol. 354(3), pp.,213-25, 1996.
Radomski, M.W., Palmer, R.M.J. and Moncada, S.: Charaderization of the L- arginine MO pathway in hurnanplatelets. Br. J. Pharmacol. Vol. 101, pp. 325- 328, 1990.
Radomski, M.W., Palmer, R.M.J. and Moncada, S.: The role of NO and cGMP in platelet adhesion to vascular endothelium. iancet, Vol. 2, pp. 1057-1 058, 1987.
Radomski, M.W., Palmer, R.M.J. and Moncada, S.: The anti-aggregation properties of the vascular endothelium : Interactions between prostacyclin and NO. Br. J. Pharmacol. Vol. 92, pp. 639-646, 1987.
Randall, M.D. and McCulloch A.I.: The involvement of ATP-sensitive potassium channels in beta-adrenoœptor-mediated vasorelaxation in the rat isolated mesenteric arterial bed. Br J Pharmacol Vol. 1 15 No.4 pp. 607-61 2 1995.
Ramaro, CS., Kincade, J.M., Perez, D.M., Garvin, R.J., Riek, R.P. and Graham, R.M.: Genomic organization and expression of the human alpha1 8 adrenergic receptor. J. Biol. Chem. Vol. 267, pp. 21 936-2 1 945, 1 992.
Riek, R.P., Handschumacher, M.K., Sung, S.S., Tan, M., Glynias. M.J., Lomansney, J.W., Cotecchia, S., Lorenz, W., Leung. W.Y., Schwin, DA., Yang- Feng, T.L., Braunstein M., Lefkowitz, R.J., and Caron M.J. : Molecular cloning and expression of the cDNA for the alphalA-adrenergic receptor. J. Biol. Chem. Vo1.266, pp.6365-6369, 1991.
Rubanyi, G.M., Lorenz, R.R. and Vanhoutte, P.M.: Bioassay of the endothelium- denved relaxing factor(s): Inactivation by catecholarnines. Am. J. Physiol. Vol. 21 5, pp. H l O77-HIO80, 1985.
Ruffdo, R. R. and Hieble J.P.: Alpha-adrenoceptors, Pharmac. Ther. Vol. 61, pp. 1-64, 1994.
Sabol, S. L., and Nirenberg, M.: Regulation of the adenylyl cyclase of neuroblasotmaXGlioma hybrid cells by alpha-adrenergic recepton. 1. Inhibition of adenylyl cydase mediated by alpha2 -receptors, J. Biol. Chem. Vol. 254, pp. 191 3-1 920, 1979.
Sakuma, I., Stuehr, D.J., Gross, S.S., Nathan, C. and Levi, R.: Identification of arginine as a precursor of endothelium derived relaxing factor (EDRF) . Proc. Natl. Acad. Sci. USA Vol. 85, pp. 8664-8667, 1988.

Satake, N. And Shibata, S.: Potentiation of the relaxing adion of isoproterenol by forskolin in rabbit aortic rings: The involvement of beta 2-adrenoceptors. Gen Phannacol Vol. 28 No3 pp. 753-756, 1997.
Seki, N., Zhang, G. and Suzuki, H.: Transmitter release modulated by isoprenaline in the dog isolatad mesenteric vein. Comp Biochern Physiol Vol. 96 NO. 1 pp. 14 1-1 46, I W O .
Sessa, W.C., Hamson, J.K., Luthin, D.R., Pollock, J. S. and Lynch, KR.: Genomic analysis and expression patterns reveal distinct genes for endothelial and brain nitric oxide synthase. Hypertension, Vol. 21, pp. 934-938, 1993.
Schliep, H.J. and Harting, J.: Betal selectivity of bisoprolol , a new beta adrenoceptor antagonist in anaesthetized dogs and guinea pigs. J. Cardiovas. Pharmacol. , Vol. 6, pp. 1 156-1 160, 1984.
Schmit, H.H.H.W.: No, Co, OH: Endogenous, soluble guanylate cyclase activating factors, FEBS Lett, Vol. 307,pp. 102-1 07, 1 992.
Schmit, H.H.H.W., Pollock , J.S., Nakane, M., Gonky, L.D., Forstennann, U. and Murad, F.: Purification of a soulble isoform of guanylyl cyclase -activating - factor synthase. P m . Nat!. Acad. Sci. USA. Vol. 88 , pp. 365369. 1991.
Schulacter,M.D., Novoteny, J. and Graham, R.M.: Evolutionary conservation of both the hydrophylic and the hydrophobic nature of aie transrnernbrane residues. 3. Theoretical Biol. 1995.
Schuman, €.M. and Madison, D.V.: A requirement for the intracellular messenger NO in long term potentiation. Science , Vol. 254, pp. 15034 506, 1991.
Schwin, D A , Lomansney, J.A., Loreng, W., Szklut, P.J., Fremeau, R.T., Yang- Feng, T.L., Caron, MG., Lefkowitz, R.J. and Cottechia, S.: Molecular cloning and expression of the cDNA for a novel alpha 1-adrenoceptonubtype. J. Biol. Chem. Vol. 265, pp.8183-8109,1990.
Schwinger, HG., Bohrn, M., Schrnit, U., Kraczewski, P., Bavendick, U., Flesch, M.. Krause, E.G. and Erdmann, E.: Unchanged protein levels of S e m II and Phopholarnban but reduced Calcium re-uptake and Calcium-ATPase activity of cardiac sarcoplasmic reticulurn from dilated cardiomyopathy patients compared with patients with non-failing hearts. Circulation, Vol. 92, pp. 3220-3228, 1995.
Shi AG., Kwan C.Y. and Daniel E.E.: Relation between density (maximum binding) of alpha adrenoceptor binding sites and contractile response in four canine vaçcular tissues. J Pharmacd Exp Ther Vol. 250 No.3 pp. 1 19-1 124, 1989.

Vanketecom H-, Matth ys P., and Denef C.: Inducible nlric oxide synthase in the antenor pituitary gland: induction by interfermgamma in a subpopulation of folliculostellate cells and in an unidentifiable population of non-hormone- secreting cells. J. of Histochem. & Cytochern. Vo1.45(6), pp.847-57, 1997.
Vanhoutte P. M. and Feletou, M.: EDHF: Conclusion: Existence of multiple endothelial deflved hyperpolarizing factors? Hawood academic publishers GmbH , the netherlands, pp.303-305, 1996.
Van Zwieten P.A.: Clinical pharmacology of calcium antagonists as antihypertensive and anti-anginal dnigs. J. of Hypertension [ Supplement]. Vol. l4(3), pp. ,S3-9, 1996.
Vllalobos-Molina R. Ibarra M.: Alpha 1-adrenoceptors mediating contraction in arteries of norrnotensive and spontaneously hypertensive rats are of the alpha I D or alpha 1A subtypes [published enatum appears in Eur J Pharmacol 1996 Dec 30;318(2-3):511]. European Journal of Phannacology Vol. 298 No.3 pp.257- 63, 1996.
Waldeck , B., Jeppson, A. B., and Widmark, E.: Partial agonisrn and selectivity : A study on beta adrenoceptor mediated effects in tracheal , cardiac and skeletal muscle . Acta. Pharmacol. Toxicol. , Vol. 58, pp. 209-218, 1986.
Wang, J.X., Ikomi, F. and Ohhashi, T.: 5-HT-induced endotheliurn dependent and independent relaxation in isolated dog anterior spinal small arteries. Can. J. Physiol. Pharmacol. Vol. 75(5), pp. 357-362, 1 997.
Weinberg. 0-H., Trivedi,P.. Tan, C.P., Mitra, S., Perkins-Borrow, A., Barkowski, D., Stradet, CD., and Bayne, M.: Clonning expression and characteriration of alphal-adrenergic receptors, alpha1 A, alpha1 8 and alpha1 C. Biochem. Bioph ys. Res. Commun. Vol. 201, pp. 1296-1 304, 1994.
White, KA. and Marletta, M.A.: Nlric oxide synthase is a cytochrome p450 type, hemoprotein. Biochernistry, Vol. 31, pp. 6627-6631, 1 992.
Whorton, A.R., Simonds, D.B. and Piantadosi, C.A.: Regulation of nlnc oxide synthesis by oxygen in vascular endothelial cells. Am J Physiol Vol. 272 Nos. 6 Pt 1 pp. L I 16141 166, 1997.
Wilson, C., Wilson, S., Piercy, VI Sennitt, M.V., and Archl J.R.S.: The rat lipolytic beta-adrenoceptor : studies using novel beta adrenoceptor agonists. Euro. J. Pharmacol. Vol. 100, pp. 309-31 9, 1984.
Woodcock, E.A. and Johnston, C.I.: Seledive inhibition by epinephnne by paralhynod hormone stimulated adenylyl cyclase in rat renal cortex. Am. J. Physiol. Vol. 242, pp. F721-i726 ,1982.

Shimomura, H. and Terada, A: Primary structure of the rat beta 1 adrenergic receptor gene. Nucleic Acids Res. Vol. 18, pp. 4591, 1990.
Stadler, J-, Billiar, T.R., Cunan, R.D., Stuehr, D. J., Ochao, J.B. and Simmojns, R.L.: Effects of exogenous and endogenous NO on mitochondrial respiration of rat hepatocytes. Am. J. Physiol. Vol. 260, pp. Cg1 06916, 1 991.
Stassen F.R., Maas R.G., Schiffen F.M., Janssen G.M. and De Mey J.G.: A positive and reversible relationship between adrenergic nerves and alpha-IA adrenoceptors in rat artenes. Journal of Phamacology 8 Experimental Therapeutics. Vol. 284 No. 1 pp. 399-405, 1998.
Steinkraus, V., Mak, J.C., Pichlmeier, U., Mensing, H., Ring, J., Bames and P.J.: Autoradiographic mapping of beta-adrenocepton in human skin. Archives of Dermatolog ical Research. Vol.288(9), pp. 549-553, 1 996.
Strader, C.D. Fong, T. M., Tota, M. R., and Underwood, D.: Structure and function of the G-protein-coupled recepton. Annu. Rev. Biochem. , Vol. 63, pp. 101-1 32, 1994.
Strosberg, A.D. and Pietri-Rouxel, F.: Function and regulation of the ps adrenocepton . Trends Pharmacol. Sci. Vol. 17,pp.373-381, 1996.
Stuehr, D.J., Kwon, N.S., Nathan, C.F., Griffith, O.W,. Feldman, P.L. and Wiseman, J.: N omega-hydroxy-L-arginine is an intermediate in the biosynthesis of nitric oxide from 1-arginine. J. Biological Chemistry. VoI.266(1 O), pp.6259- 6263, 1991.
Terman, B.L., Riek, R.P., Grodski, A., Hess, H.G. and Graham R.M.: Identification and characterization of alphal-adreergic receptor subtypes. Mol. Phanacol. Vol. 37, pp. 526-534. 1990.
Testa R. Destefani C. Guarneri L. Poggesi E. Simonaui 1. Taddei C. Leonardi A.:The alpha 1 d-adrenocaptor subtype is involved in the noradrenaline- induced contradons of rat aorta. Life Sciences. Vo1.57 No. 13 pp.PL159-63, 1995.
Thomae, KR., Gelter, D.A., Billiar, T.R., Davies, P., Pitt, BR. and Simmons, R.L.: Antisense oligodeoxynucleotide to inducible nitric oxide synthase inhibits nitric oxide synthesis in rat pulmonary artery smooth musde cells in culture. Surgery. Vol. 1 14(2),pp.272-277, 1993.
Van der Graaf PH. Deplanne V. Duquenne C. Angel 1.: Analysis of alphal- adrenoceptors in rabbit lower urinary tract and mesenteric artery. European Journal of Phamecology. Vol. 327 No. 1 pp. 25-32, 1997.

Wu, O. Katz, A., Lee, C.H., Simon, M.I.: Activation of phospholipase C by alphal - adenergic reœptors is medieted by the alpha subunits of Gq family. J. Biol Chem. Vol. 267, pp. 231 89-25802, 1992.
XU K, HAN C.: Quantification of mRNAs for three alpha ladrenoceptor subtypes in rat aorta by solution hybridization. Liie Sci Vol. 59 No. 21 pp. P1343- P1347, 1996.
Yamakazi, S., Katada, T. and Ui, M.: Alpha2-adrenergic inhibition of insulin secretion via interference with cAMP generated in rat pancreatic islets. Mol. Parmacol. , Vol.. 21, pp. 648453. 1982.
Yarden, Y., Rodriguez, H., Wong, S.K.F., Brandt, D.R., May, D.C., Bumier, J., Harkins, R.M., Chen, E.Y., Ramachandaran, J., Ulrich, P., and Ross, E.M.: The avian beta adrenergic receptor: Primary structure and membrane topology . Proc. N d . AC&. Sd. USA. Vol. 83, pp. 6795-6799, 1986.
Zeng, Dal Harrison, J.K., D1AngeloI D.D., Barber, C.M., Tucker, AL., and Lynch, KR.: Molecular characterization of a rat alpha26-adrenergic receptor. Proc. Natl. Acad. Sci., Vol., 87, pp. 3102-3106. 1990.
Zheng, X.F., Kwan, C.Y. and Daniel, E. E.: Beta-Adrenoceptor activates endotheliumdependent release of nitric oxide in rat aorta. Chung Kuo Yao Li Hsueh Pao Vol. 16 No. 5 pp. 385390, 1995.