infect. immun....
TRANSCRIPT

– 1 –
1
2
3
4
Differential requirements for NAIP5 in activation of the NLRC4 (IPAF) 5
inflammasome 6
7
Karla L. Lightfield1,5§
, Jenny Persson2,6§
, Norver J. Trinidad2, Sky W. Brubaker
2,7, Eric 8
M. Kofoed2, John-Demian Sauer
2, Eric A. Dunipace
2, Sarah E. Warren
3,4, Edward A. 9
Miao3, Russell E. Vance
2* 10
11
1School of Public Health, 12
2Division of Immunology & Pathogenesis, Department of Molecular & Cell Biology, 13
University of California, Berkeley, CA 94720 14
3Institute for Systems Biology, Seattle, WA 98103 15
4Department of Immunology, University of Washington, Seattle, WA 98195 16
5Current address: Department of Microbiology & Immunology, Stanford University, 17
Stanford, CA 94305 18
6Current address: Department of Microbiology, Tumor and Cell Biology, Karolinska 19
Institute, Stockholm, Sweden 20
7Current address: Children’s Hospital Boston, Boston, MA 02115 21
§These two authors contributed equally 22
*Correspondence: Russell E. Vance, 415 Life Science Addition, Department of 23
Molecular and Cell Biology, University of California, Berkeley, CA 94720. Tel. (510) 24
643-2795. Fax. (510) 642-1386. Email. [email protected] 25
26
Running title: Requirement for NAIP5 in NLRC4 inflammasome activation27
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.01187-10 IAI Accepts, published online ahead of print on 31 January 2011
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 2 –
Abstract 1
Inflammasomes are cytosolic multiprotein complexes that assemble in response to 2
infectious or noxious stimuli and activate the CASPASE-1 protease. The inflammasome 3
containing the nucleotide binding domain-leucine rich repeat (NBD-LRR) protein 4
NLRC4 (IPAF) responds to the cytosolic presence of bacterial proteins such as flagellin 5
or the inner rod component of bacterial type III secretion systems (e.g. Salmonella PrgJ). 6
In some instances, such as infection with Legionella pneumophila, activation of the 7
NLRC4 inflammasome requires the presence of a second NBD-LRR protein, NAIP5. 8
NAIP5 is also required for NLRC4 activation by the minimal C-terminal flagellin peptide 9
that is sufficient to activate NLRC4. However, NLRC4 activation is not always 10
dependent upon NAIP5. In this report, we define the molecular requirements for NAIP5 11
in activation of the NLRC4 inflammasome. We demonstrate that the N-terminus of 12
flagellin can relieve the requirement for NAIP5 during activation of the NLRC4 13
inflammasome. We also demonstrate that NLRC4 responds to the Salmonella protein 14
PrgJ independently of NAIP5. Our results indicate that NAIP5 regulates the apparent 15
specificity of the NLRC4 inflammasome for distinct bacterial ligands.16
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 3 –
Introduction 1
The innate immune system initiates defense against infectious agents by employing 2
germ-line encoded receptors to detect microbial molecules (also called pathogen-3
associated molecular patterns or PAMPs) (10). Examples of PAMPs include 4
lipopolysaccharide, cell wall components, bacterial nucleic acids, and flagellin. The 5
mammalian Toll-like receptors (TLRs) are transmembrane receptors that are capable of 6
detecting microbial products at the cell surface and within intracellular compartments 7
(26). Signaling downstream of TLR stimulation results in the activation of NF-κB and the 8
induction of proinflammatory cytokines, chemokines and other antimicrobial defenses. In 9
addition to TLRs, there are several types of receptors that lack transmembrane domains 10
and function to sense PAMPs within the cytosol (26). For example, the cytosolic 11
presence of RNA is detected by the MDA5/RIG-I family of RNA helicases. It is believed 12
that cytosolic immunosurveillance permits host cells to make specialized or unique 13
responses to intracellular pathogens, and thus to distinguish these pathogens from 14
extracellular microbes that do not access the host cell cytosol (27). 15
The nucleotide binding domain (NBD)-leucine rich repeat (LRR)-containing 16
proteins constitute an important family of cytosolic immunosensors. Certain NBD-LRR 17
proteins, including NAIP5 and NLRC4, are involved in orchestrating the assembly and 18
activation of multiprotein complexes called ‘inflammasomes’ (24). The primary function 19
of inflammasomes is to activate the cysteine protease CASPASE-1 (CASP-1), which is 20
initially produced as an inactive pro-protein and requires recruitment to inflammasomes 21
in order to become activated (16). Once activated, CASP-1 is required for the proteolytic 22
processing and secretion of the proinflammatory cytokines interleukin (IL)-1β and IL-18. 23
In addition, activated CASP-1 can induce a rapid, inflammatory cell death termed 24
‘pyroptosis’ (2). 25
Several distinct NBD-LRR proteins are expressed by host cells, and appear to 26
dictate inflammasome assembly in response to specific stimuli. For example, the NBD-27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 4 –
LRR protein NLRP3 (NALP3, CRYOPYRIN) orchestrates assembly of an 1
inflammasome that responds to a wide variety of stimuli including crystalline substances 2
such as uric acid, asbestos, and alum (24). In the mouse, the related NBD-LRR protein 3
NLRP1B (NALP1B) activates a distinct inflammasome in response to anthrax lethal 4
toxin (3). NBD-LRR proteins are not the only sensor-scaffold proteins responsible for 5
CASP-1 activation. For example, the PYHIN family member AIM2 binds cytosolic 6
DNA and is responsible for CASP-1 activation in response to infection with bacterial 7
pathogens (7, 11, 12, 21, 23, 28). 8
The inflammasome containing the NBD-LRR protein NLRC4 (IPAF) is one of 9
the most well-characterized inflammasomes, and has been shown to activate CASP-1 10
specifically in response to a conserved domain within the bacterial protein flagellin (1, 9, 11
14, 17), as well as to the conserved inner rod component of the type III secretion system, 12
called PrgJ in Salmonella (18). Interestingly, in some instances, activation of the NLRC4 13
inflammasome requires the presence of a second NBD-LRR protein, NAIP5 (formerly 14
BIRC1E) (14, 19, 22, 30). NAIP5 is believed to hetero-oligomerize with NLRC4 (5, 30) 15
but the precise biochemical function of NAIP5 in NLRC4 activation remains enigmatic. 16
It was previously shown that NAIP5 is required for CASP-1 activation in response to 17
some, but not all, NLRC4-dependent stimuli (14). For example, NAIP5 was required for 18
NLRC4-dependent activation of CASP-1 in response to Legionella pneumophila, 19
whereas Naip5-deficiency had only partial effects on NLRC4-dependent responses to 20
Salmonella and Pseudomonas (14). NAIP5/NLRC4/CASP-1 activation by L. 21
pneumophila is largely dependent on amino acids within the C-terminus of flagellin (14), 22
and accordingly, retroviral-mediated expression of a minimal C-terminal peptide from 23
flagellin activates CASP-1-mediated pyroptotic cell death in a manner completely 24
dependent on NLRC4 and NAIP5 (14). Unexpectedly, however, retroviral-mediated 25
expression of full-length flagellin activated NLRC4- and CASP-1-dependent pyroptosis 26
independently of NAIP5 (14). 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 5 –
The molecular basis for the differential requirements for NAIP5 in NLRC4 1
activation is not well understood. Legionella and Salmonella both activate NLRC4 2
primarily via translocation of flagellin to the cytosol of host cells, but the flagellin 3
molecules translocated exhibit amino acid differences in the critical C-terminal domain. 4
Moreover, while Salmonella translocates flagellin into host cells via its SPI-I type III 5
secretion system (T3SS) (25), Legionella lacks a T3SS and instead utilizes the 6
evolutionarily unrelated Dot/Icm type IV secretion system (T4SS) to translocate flagellin 7
into host cells. T3SSs, but not T4SSs, contain homologs of the bacterial protein PrgJ, 8
another known activator of NLRC4 (18). Thus, differences in flagellin itself, and/or the 9
flagellin-translocating apparatus, may underlie the differential requirements for NAIP5 in 10
NLRC4 activation. 11
In this paper we dissect the molecular features that dictate the requirement for 12
NAIP5 in the activation of the NLRC4 inflammasome. We demonstrate that L. 13
pneumophila engineered to express Salmonella flagellin activates NLRC4 in a strictly 14
NAIP5-dependent manner, thus ruling out polymorphisms within flagellin as an 15
explanation for the differential requirement for NAIP5 in activation of NLRC4 by the 16
two bacterial species. We further demonstrate that the N-terminus of flagellin, while not 17
itself sufficient to activate NLRC4, is nevertheless able to transform the C-terminus of 18
flagellin from a NAIP5-dependent activator of the NLRC4 inflammasome to a NAIP5-19
independent activator. Lastly, we show that the PrgJ protein can activate NLRC4 without 20
a requirement for NAIP5. Together, our results suggest a model in which NAIP5 21
functions to dictate the specificity of NLRC4 for distinct stimuli. 22
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 6 –
Materials and Methods 1
2
Mice. Wildtype C57BL/6J (B6) mice were from Jackson Labs. Nlrc4-/-
mice on a pure 3
B6 background (15) were obtained from S. Mariathasan and V. Dixit (Genentech). 4
Naip5-/-
mice on a pure B6 background mice were described previously (14). 5
6
Bacterial Strains. The Salmonella typhimurium strain LT2 and the flagellin mutant S. 7
typhimurium LT2FliC/FljB- were gifts from A. Van Der Velden and M. Starnbach. The 8
pLIV2-L.p.FlaA and pLIV2-S.t.PrgJ constructs were constructed as IPTG-inducible 9
variants of previously described strains. A construct fusing the amino terminal 300bp of 10
the actA gene with the flaA gene from L.p.FlaA was previously described and used to 11
facilitate ectopic secretion of Legionella pneumophila FlaA (Sauer et al, Submitted). The 12
actA-L.p.FlaA fusion was amplified from the pPL2-L.p.FlaA plasmid with the primers 13
(aaaagCGGCCGgtgggattaaatagatttatgcgtgc) and (aaaaGTCGACcagaaatcgaagtgcagttg) 14
using Pfu Ultra II (Agilent Technologies, Santa Clara, CA). The resulting PCR product 15
and the IPTG inducible variant of pPL2, pLIV2 (8), were double digested with EagI and 16
SalI (New England Biosystems, Waltham, MA) and ligated using the NEB quick ligation 17
kit. The resulting vector pLIV2-L.p.FlaA was subsequently integrated at the tRNAArg
18
locus on the L. monocytogenes 10403s chromosome as previously described (13). 19
pLIV2-S.t.PrgJ was constructed similarly from the previously described actA-prgJ fusion 20
construct (Sauer et al, Submitted). The fusion was amplified from the pPL2-S.t.PrgJ 21
plasmid using the primers (aaaaGCGGCCGCaggagggagtataagtgggattaaatag) and 22
(ttttGTCGACtcatgagcgtaatagcgtttc), digested and ligated into pLIV2, and integrated into 23
the 10403s chromosome as previously described. The Legionella strain LP02 is a 24
streptomycin-resistant thymidine auxotroph derivative of Legionella pneumophila strain 25
LP01. LP02∆flaA contains an unmarked deletion of flaA and was described previously 26
(22). LP02∆flaA::fliC expresses Salmonella fliC from the chromosome under the control 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 7 –
of the endogenous Legionella flaA promoter. To generate this strain, the flaA promoter 1
from LP02 was amplified by PCR using primers 2
pflaAFwd (ATAGTCGACTTAATGCCTCTTTCTCTCCTGTCG) and pflaARev 3
(AATGACTTGTGCCATAATTTTAGTCTCCTCAGACCTGAATCC) while fliC was 4
amplified from S. typhimurium using primers fliCfwd 5
(GAGGAGACTAAAATTATGGCACAAGTCATTAATACAAACAGC) and fliCrev 6
(ATAGGATCCTTAACGCAGTAAAGAGAGGACGTTTTG). These amplicons were 7
mixed and spliced in a second round of PCR using pflaAFwd and fliCrev and cloned into 8
the allelic exchange vector pSR47S. This ‘suicide’ construct was introduced onto the 9
LP02∆flaA chromosome by a single crossover and selection with Kanamycin (50µg/ml). 10
11
Retroviral constructs and transductions. Retroviral constructs were generated and 12
transductions were performed as previously described (14). Briefly, retroviral particles 13
were generated by transient transfection of Phoenix-Eco packaging cells with MSCV2.2-14
based retroviral vectors. 1x106
bone marrow derived cells were cultured 48h in a 6-well 15
plate in media containing MCSF and were then transduced with 1ml of retrovirus-16
containing packaging cell supernatant. Macrophages were analyzed 3-4 days post 17
transduction and analyzed for GFP expression on a Beckman Coulter FC-500 flow 18
cytometer. Greater than 10,000 cells were analyzed per sample. When simultaneously 19
transducing cells with multiple constructs (Figure 2) equal volumes of each retroviral 20
supernatant were used. These macrophages were analyzed on a BD Influx Cell Sorter 21
for GFP and mCherry expression. The retroviral constructs used in this paper were 22
generated as follows: GFP-C65 by amplifying the C-terminal 65 amino acids of flagellin 23
using the primers pC65fwd (GACGAGCTGTACAAGTTTGAATCAACGATAG) and 24
pFlaA65rev (TTGCGGCCGCCTATCGACCTAACAAAGATAATACAGATTGCG) 25
while GFP was amplified using the primers pGFPfwd 26
(ATAAGATCTCCACCATGGTGAGCAAGGGCGAGGA) and pGFP65rev 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 8 –
(CTATCGTTGATTCAAACTTGTACAGCTCGTC). These amplicons were mixed and 1
spliced in a second round of PCR using the primers pGFPfwd and pFlaArev and cloned 2
into the retroviral expression vector pMSCV2.2 using the enzymes BglII and NotI. 3
Similarly, the construct N65-GFP-C65 was generated by amplifying GFP-C65 with the 4
primers pN65GFPfwd (GGATGAACCAAGCCGTTATGGTGAGCAAGGGC) and 5
pFlaA65rev while the N-terminal 65 amino acids of flagellin were amplified with the 6
primers pN65fwd 7
(ATAAGATCTCCACCATGGCTCAAGTAATCAACACTAATGTGG) and 8
pN65GFPrev (GCCCTTGCTCACCATAACGGCTTGGTTCATCC). These amplicons 9
were spliced in a second round of PCR using the primers pN65fwd and pFlaA65rev. 10
N65-GFP was generated by amplifying the N-terminal 65 amino acids of flagellin with 11
the primers pN65fwd and pN65GFPrev while amplifying GFP with the primers 12
pN65GFPfwd and pGFPrev (TTGCGGCCGCTTACTTGTACAGCTCGTC). These 13
amplicons were mixed and spliced in a second round of PCR using the primers pN65fwd 14
and pGFPrev. The constructs N65-GFP-C65 L12L32::II and N65-GFP-C65 15
I5V9L12:AAA were generated using site directed mutagenesis on the parental N65-GFP-16
C65 construct, pL12fwd 17
(CACTAATGTGGCGTCGatcACAGCCCAACGTAATTTGGG), pL12rev 18
(CCCAAATTACGTTGGGCTGTgatCGACGCCACATTAGTG), pL32fwd 19
(CATCGATCCAGCGTatcTCATCGGGATTAAGG) and pL32rev 20
(CCTTAATCCCGATGAgatACGCTGGATCGATG) for L12L32II and pL12Afwd 21
(CACTAATGCAGCGTCGGCAACAGCCCAACG), pL12Arev (CGTTGGGCTGTT 22
GCCGACGCTGCATTAGTG), pI5V9AAfwd(GGCTCAAGTAGCAAACACTAA 23
TGCAGCGTCGCTCAC and pI5V9AArev(GTGAGCGACGCTGCATTAGTGTTT 24
GCTACTTGAGCC). mCherry-C65 was generated by amplifying mCherry using 25
pmCherryfwd (ATAAGATCTCCACCATGGTGAGCAAGGGCGAGG) and 26
pmCherry65rev (GGCTATCGTTGATTCAAACTTGTACAGCTCGTCC), while the C-27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 9 –
terminal 65 amino acids of flagellin were amplified using the primers pmCherry65fwd 1
(GGACGAGCTGTACAAGTTTGAATCAACGATGCC) and pFlaA65rev. These 2
amplicons were mixed and spliced during a second round of PCR with the primers 3
pmCherryfwd and pFlaA65rev and cloned into pMSCV2.2 with BglII and NotI. 4
5
Cytotoxicity Assays. Cytotoxicity was measured as described previously (6) by 6
determining the activity of lactate dehyrogenase (LDH) released by cells. Bone marrow-7
derived macrophages (5x104 to 1x10
5) were seeded onto tissue culture-treated 96 well 8
plates and infected with bacteria at the indicated multiplicity of infection. Plates were 9
centrifuged at 400xg to ensure equivalent infectivity of WT and flagellin-mutant strains. 10
At 30 min-1hr post infection, media was removed and replaced with media containing 11
10-100µg/mL Gentamicin to kill extracellular bacteria. Six hours after infection, plates 12
were centrifuged at 400xg, supernatant was collected and assayed for LDH release. 13
Legionella strains used for infection were grown overnight to an OD>3.8. Salmonella 14
cultures were grown overnight with shaking at 37°C and were diluted 3h prior to 15
infection at 1:40 in 1ml of LB and re-grown with shaking until reaching late-exponential 16
phase. Listeria strains were grown as standing cultures at 37°C in BHI media. In the 17
experiments with Listeria, infected cells were treated with Isopropyl β-D-1-18
thiogalactopyranoside (IPTG) to induce expression of flaA or prgJ. Percent specific lysis 19
of infected cells was calculated as the amount of LDH released (minus background 20
release by uninfected cells) divided by the amount of LDH released by detergent lysed 21
cells (minus background) x 100%. 22
23
Growth Curves. Bacterial growth was determined as previously described (4). 24
Macrophages were plated at a density of 5x105 cells per mL in 24 well plates, allowed to 25
adhere overnight and infected with stationary phase Legionella (OD>3.8) at an MOI of 26
0.01. Colony forming units (CFUs) were determined by lysing washed macrophage 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 10 –
monolayers with sterile distilled water and plating lysates on buffered charcoal yeast 1
extract plates. 2
3
Protein Transfection. Salmonella FliC, PrgJ and SsaI proteins were purified as described 4
previously (18). Macrophages were plated 5x104 per well in a 96 well plate, allowed to 5
adhere overnight and primed with tripalmitoyl cysteinyl seryl tetralysine lipopeptide 6
(Pam3CSK4, 0.5µg/ml) for 3.5 hours prior to protein transfection to induce expression of 7
pro-interleukin(IL)-1β. 0.2µg of protein per well was transfected using the Profect 8
P1(Targeting Systems) transfection reagent according to manufacturer’s instructions. 9
Supernatants were collected for analysis 3h post transfection and analyzed by ELISA for 10
IL-1β (R&D Systems). 11
12
Statistics. Statistical significance was assessed by the t-test or 1-way ANOVA with 13
Bonferroni posttests. Graph Pad Prism 5 software was for statistical analyses. 14
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 11 –
Results 1
The N-terminus of flagellin can relieve the requirement for NAIP5 in flagellin-sensing. 2
In order to better understand the molecular basis for NAIP5/NLRC4 activation, we first 3
sought to determine what regions of flagellin enable full-length flagellin to be toxic to 4
Naip5-/-
macrophages in the retroviral lethality assay (14). In this assay, flagellin is 5
expressed downstream of a mammalian promoter in macrophages using a retrovirus-6
based construct. Transduction of macrophages with a construct expressing a stimulatory 7
flagellin molecule results in rapid pyroptotic cell death of the macrophages and thereby 8
prevents expression of a co-expressed green fluorescent protein (GFP) reporter (14). 9
Conversely, transduction of a construct encoding a non-stimulatory flagellin mutant 10
results in GFP+ cells (14). Previously, the retroviral lethality assay was used to establish 11
that NLRC4 and NAIP5 are both required for the response to the C-terminal domain of 12
flagellin, whereas pyroptotic cell death in response to full-length flagellin required 13
NLCR4 but not NAIP5 (14). Thus, in order to determine what regions in full-length 14
flagellin relieve the requirement for NAIP5, we began by making a series of retroviral 15
constructs expressing N-terminally deleted flagellin molecules (Figure 1A). These 16
constructs were transduced into primary macrophages. As shown previously (14), 17
retroviral expression constructs encoding full-length flagellin (FlaA) did not stably 18
transduce C57BL/6 (WT) or isogenic Naip5-/-
macrophages, as evidenced by a lack of 19
GFP+ cells 3 days after transduction (Figure 1B). However, we found that a retroviral 20
construct that expresses a flagellin molecule lacking the N-terminal 65 amino acids from 21
flagellin (FlaAN∆65), could be transduced into Naip5-/-
macrophages (Figure 1B). 22
Consistent with our previous observation that the C-terminus of flagellin is sufficient to 23
activate NLRC4, FlaAN∆65 (which retains the normal flagellin C-terminus) could not be 24
transduced into WT macrophages. Further N-terminal deletions of FlaA (∆N85, ∆N100, 25
∆N125) resembled ∆N65 and could be transduced into Naip5-/-
macrophages (Figure 26
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 12 –
1B). These results suggest that the N-terminus of flagellin is important for the ability of 1
full-length flagellin to activate NLRC4 independently of NAIP5. 2
3
Molecular basis by which the N-terminus affects the requirement for Naip5. 4
Next, we sought to address whether the N-terminus of flagellin is sufficient to convert the 5
C-terminus of flagellin into a NAIP5-independent activator of NLRC4. Consistent with 6
our previous studies (14), we found that the C-terminal 65 amino acids of flagellin, fused 7
to GFP, is a NAIP5-dependent activator of NLRC4 (Figure 1C). In contrast, full length 8
flagellin (whether or not it is fused to GFP) is a NAIP5-independent activator of NLRC4 9
(Figure 1B, 1C). To determine the role of the N-terminus in NAIP5/NLRC4 activation, 10
we created an expression construct that consisted of GFP flanked by the N-terminal and 11
C-terminal 65 amino acids of FlaA (see Figure 1A). The resulting N65-GFP-C65 12
construct behaved just like full-length flagellin; namely, it was cytotoxic to macrophages 13
in a manner requiring NLRC4, but independent of the presence or absence of NAIP5 14
(Figure 1C). The N-terminus of flagellin fused to GFP was itself noncytotoxic and could 15
be transduced into WT (Nlrc4+) macrophages (Figure 1C). Thus, the N-terminus of 16
flagellin is not sufficient to activate NLRC4. However, the N-terminus of flagellin 17
appears to enhance or alter NLRC4-dependent responses to the C-terminus of flagellin in 18
such a way that NAIP5 is no longer required. 19
Although the C-terminal region of flagellin is thought to be unstructured when 20
flagellin is in its monomeric form, the structure of flagellin within the flagellin filament 21
(29) suggests that the N and C-termini of flagellin may interact weakly via a coiled-coil 22
interaction. Moreover, the crystal structure of GFP reveals that its N- and C-termini are 23
near to each other (20), and it is therefore possible that the appended N- and C-termini of 24
flagellin could interact with each other in the N65-GFP-C65 construct. Thus, we 25
hypothesized that perhaps a weak coiled-coil interaction with the N-terminus of flagellin 26
resulted in a stabilized or structurally altered C-terminal region that no longer required 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 13 –
NAIP5 for stimulation of NLRC4. To test this idea, we mutated the N-terminus of 1
flagellin within the N65-GFP-C65 construct in order to disrupt the putative interaction 2
between the N- and C-terminus. Two leucines (L12, L32) in the N-terminus of flagellin, 3
were mutated to more bulky isoleucines, which would not be easily accommodated 4
within the core of a coiled-coil and should thus result in a disruption of putative coiled-5
coil interactions. In addition, a series of mutations predicted to be even more disruptive 6
to the putative coiled-coil interaction were made, namely, isoleucine 5, valine 9 and 7
leucine12 were all mutated to alanines (I5V9L12:AAA). Neither series of mutations 8
were sufficient to transform the parental N65-GFP-C65 stimulus from a Naip5-9
independent stimulus (N65-GFP-C65) into a NAIP5-dependent one (Figure 1D, 1E). 10
Thus, we were unable to find evidence that a coiled-coil interaction between the N- and 11
C-termini of flagellin is important for the ability of full-length flagellin to activate 12
NLRC4 independent of NAIP5. These results do not rule out the possibility that an 13
interaction between the N- and C-termini of flagellin (that we did not disrupt with the 14
mutations we made) is important in determining the requirement for NAIP5 in NLRC4 15
activation. However, a lack of a role for a coiled-coil interaction would not be surprising 16
given that the interaction between the N- and C- termini of flagellin is likely very weak 17
(29) and probably not significant when flagellin is in its monomeric form, which is the 18
form that appears to be recognized by NLRC4 (17). 19
20
The N- and C-termini of flagellin can act in trans to activate NLRC4. 21
Because coiled-coil interactions did not appear to be required for the NAIP5-independent 22
activation of NLRC4 by flagellin, we questioned whether the N- and C-termini of 23
flagellin needed to be expressed on the same molecule, or whether they could cooperate 24
in trans to activate NLRC4 without NAIP5. In order to address this question, a construct 25
encoding the N-terminal 65 amino acids of flagellin fused to GFP (N65-GFP), and a 26
separate construct encoding the C-terminal 65 amino acids of flagellin fused to mCherry 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 14 –
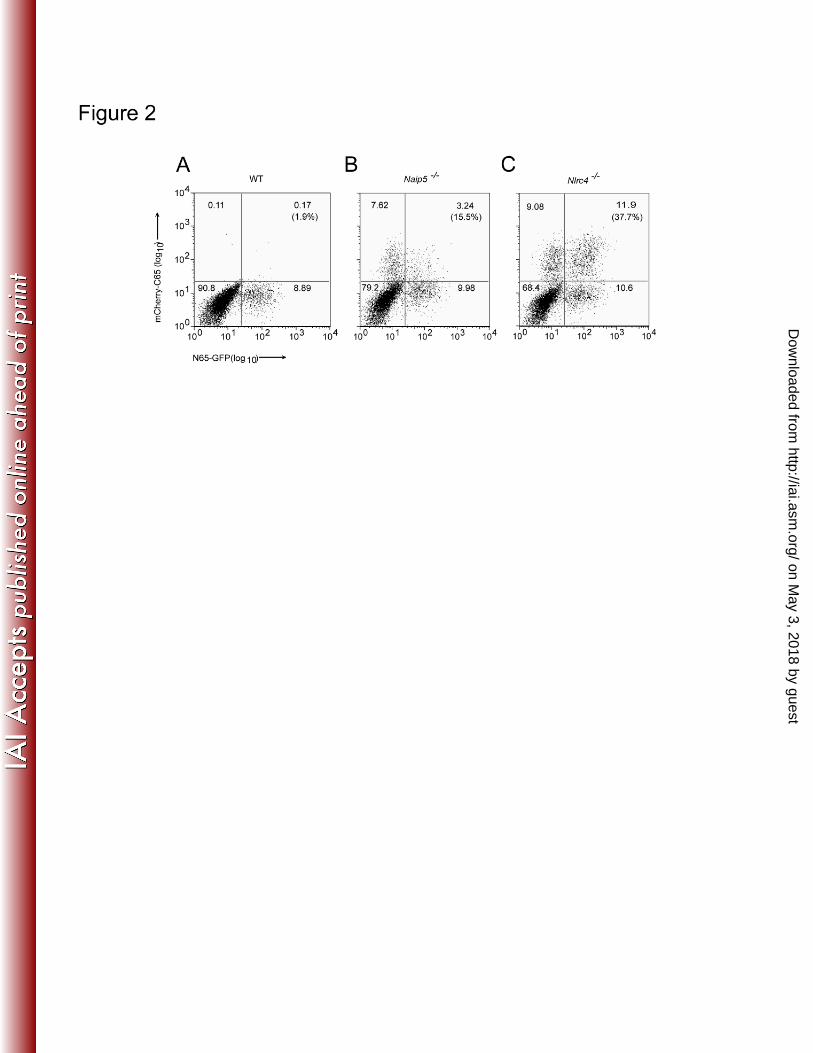
(mCherry-C65), were expressed simultaneously in wild-type, Naip5-deficient, and Nlrc4-1
deficient macrophages via retroviral transduction. As expected, mCherry-C65 was 2
cytotoxic to wild-type macrophages, but not Naip5-/-
or Nlrc4-/-
macrophages (Figure 3
2A-C). While a robust population of brightly double positive macrophages was detected 4
in transduced Nlrc4-/-
macrophages, Naip5-/-
macrophages harbored only a 5
proportionally smaller, less bright population of double positive macrophages (Figure 2B, 6
2C). These results indicate that, at the relatively high expression levels achieved by 7
retroviral transduction, the N-terminus of flagellin can act in trans to convert the C-8
terminus of flagellin into a NAIP5-independent stimuli. We were unable to detect a 9
physical interaction between mCherry-C65 and N65-GFP by immunoprecipitation (data 10
not shown). Taken together, our data therefore raise the possibility that the N- and C-11
termini cooperate to activate NLRC4 without physically interacting with each other, 12
though a role for a transient or weak physical interaction cannot be rule out. 13
14
NAIP5-independent detection of Salmonella 15
We previously reported that while activation of the inflammasome by Legionella was 16
strictly flagellin-and NAIP5-dependent, activation of the inflammasome by Salmonella 17
was partially NAIP5-independent. It is difficult to compare infections across two 18
different bacterial species when there may be differences in infectivity, translocation 19
levels of flagellin, or flagellin molecules themselves. Thus, to begin to determine if there 20
is in fact a qualitative difference between Salmonella and Legionella with respect to 21
activation of NLRC4, we assayed macrophage cell death at a wide range of MOIs for 22
both bacterial species (Figure 3). We centrifuged bacteria onto the macrophages to 23
minimize differences in infectivity between motile and non-motile strains and carefully 24
monitored the infectivity of the various strains by determining gentamicin-resistant cell-25
associated colony forming units 1h after infection. We found that infection of 26
macrophages with Legionella causes macrophage cell death in a manner dependent upon 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 15 –
both host expression of NAIP5 and NLRC4 and bacterial expression of flagellin across a 1
broad range of MOIs, from 0.3 to 10 (Figure 3A). In contrast, we found that Salmonella 2
causes a partially flagellin-independent cell death at higher bacterial loads (e.g., MOI>2) 3
(Figure 3A), consistent with previous results (18). As previously described (14), we also 4
found that at these higher MOIs, Salmonella also induces a partially NAIP5-independent 5
(but NLCR4-dependent) cell death. Some of the NAIP5-independent death appeared to 6
be flagellin-dependent, but some appeared to be flagellin-independent. The flagellin-7
dependent activation of NLRC4 could not be explained by a reduced infectivity of the 8
∆fliC Salmonella mutant since the cfu per cell were quantified and not found to be 9
significantly different. Thus, the activation of NLRC4 by Salmonella seems to be only 10
partially NAIP5-dependent and flagellin-dependent. This contrasts with the activation of 11
NLRC4 by Legionella, which was more uniformly NAIP5- and flagellin-dependent. 12
To further assess the underlying differences in NLRC4 activation between 13
Salmonella and Legionella, we asked whether the NAIP5-independent death initiated by 14
Salmonella is due to an intrinsic difference between Legionella and Salmonella flagellin. 15
Thus we generated a strain of Legionella that expresses Salmonella flagellin (fliC) in 16
place of its own flagellin. This strain (LP02 ∆flaA::fliC) was capable of activating the 17
NLRC4 inflammasome almost as efficiently as wildtype Legionella (22), and 18
interestingly, did so in a fully NAIP5-dependent manner (Figure 3B). In addition, 19
complementing flagellin-deficient Legionella with Salmonella fliC restored the Naip5-20
dependent IL-1β secretion in response to infection with Legionella (Figure 3C), and the 21
growth restriction of Legionella within macrophages (Figure 3D). Thus, when expressed 22
from Legionella, Salmonella flagellin activates NLRC4 in an entirely NAIP5-dependent 23
manner, similar to Legionella’s own flagellin. For reasons that remain unclear, we were 24
unable to generate a strain of Salmonella that was capable of translocating Legionella 25
flagellin into host cells. Nevertheless, our data strongly suggest that there are not 26
intrinsic differences between the flagellins that underlie the differing abilities of 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 16 –
Legionella and Salmonella to activate the NLRC4 inflammasome independently of 1
NAIP5. These results are consistent with previous experiments that demonstrated that 2
the C-termini of Legionella and Salmonella flagellin behaved identically in the retroviral 3
lethality assay (14). 4
5
PrgJ activates Nlrc4 in a Naip5-independent manner. 6
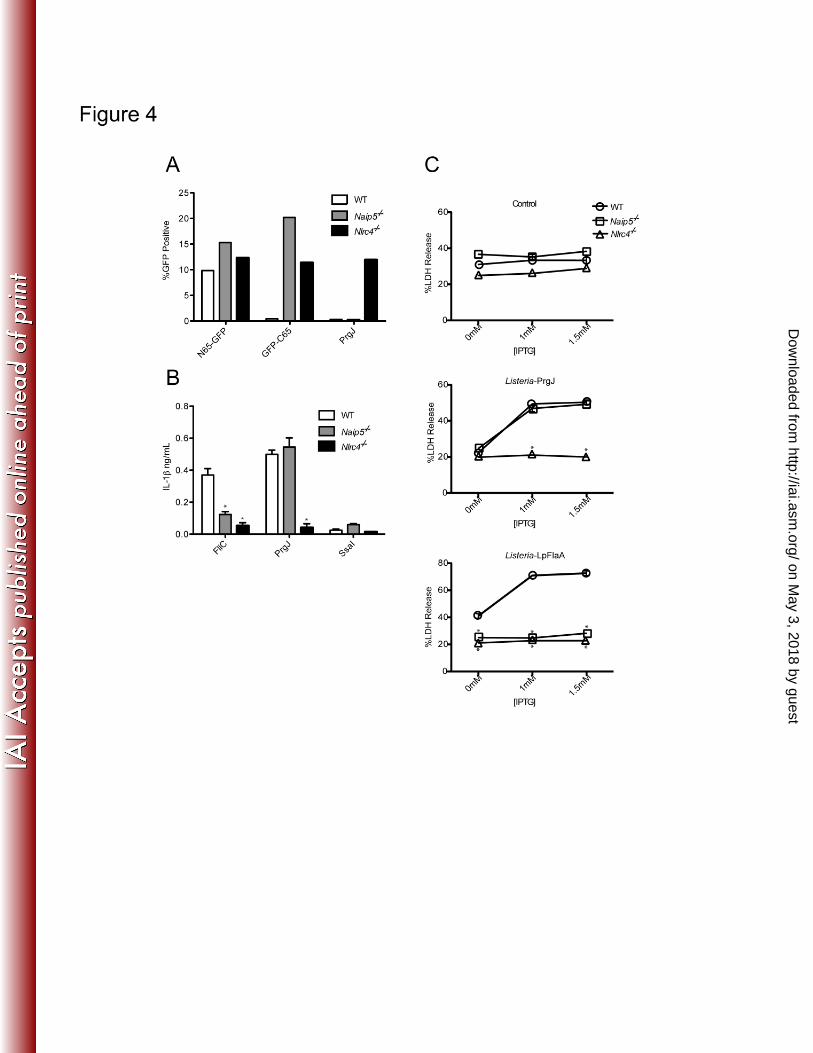
Recently, it was reported that Salmonella PrgJ activates NLRC4 (18). PrgJ is an essential 7
inner rod component of the SPI-1 type III secretion system. Legionella utilizes a type IV 8
secretion system to translocate flagellin into host cells and lacks both a type III secretion 9
system and a PrgJ homolog. Thus, one hypothesis to explain the ability of Salmonella to 10
activate NLRC4 independently of NAIP5 is that Salmonella PrgJ can activate NLRC4 11
independently of NAIP5. We therefore utilized the retroviral lethality assay to test 12
whether PrgJ activated NLRC4 independently of NAIP5 (Figure 4A). We found that in 13
contrast to the control retrovirus expressing the Naip5-dependent GFP-C65 stimulus, the 14
PrgJ-expressing retrovirus was not efficiently transduced into Naip5-/-
macrophages. 15
This result is consistent with PrgJ being a NAIP5-independent activator of NLRC4; 16
however, interpretation of the result is complicated by the fact that full-length flagellin 17
itself behaves as a NAIP5-independent activator of NLRC4 in the retroviral lethality 18
assay (Figure 1B; (14)). Thus, the lack of NAIP5-dependence for PrgJ in the retroviral 19
lethality assay may in part be due to a peculiarity of the assay (for example, the very high 20
expression levels obtained from the retroviral promoter). Therefore, to test whether PrgJ 21
could stimulate NLRC4 independently of NAIP5, it was necessary to utilize an assay in 22
which full-length flagellin behaved as a NAIP5-dependent stimulus. Two such assays 23
were employed. In the first, full-length recombinant FliC and PrgJ proteins were 24
transfected into the macrophage cytosol using protein transfection, as described 25
previously (17, 18). Activation of NLRC4 was followed by measuring the 26
NLRC4/CASP-1-dependent release of IL-1β into the supernatant (Figure 4B). Although 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 17 –
IL-1β release in response to transfected FliC protein required NAIP5, IL-1β release in 1
response to PrgJ was nearly entirely NAIP5-independent (Figure 4B). 2
In the second assay to assess whether PrgJ is a NAIP5-independent activator of 3
NLRC4, we utilized flagellin- and PrgJ-expressing strains of the gram-positive bacterium 4
Listeria to activate NLRC4. Listeria strains engineered to express Legionella flagellin 5
(FlaA) strongly activate NLRC4-dependent cell death (J.-D. Sauer et al, submitted). 6
Although the cell death induced by Listeria-flaA was NAIP5-dependent, cell death 7
induced by Listeria-prgJ was entirely NAIP5-independent (Fig. 4C). Thus we conclude 8
that PrgJ can activate NLRC4 independently of NAIP5. 9
10
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 18 –
Discussion 1
Our results demonstrate that NAIP5 is required for activation of the NLRC4 2
inflammasome only in response to certain stimuli. These stimuli include infection with 3
Legionella (expressing Legionella or Salmonella flagellin), or transduction with a 4
retrovirus expressing the C-terminus of flagellin fused to GFP. Other stimuli, such as 5
constructs expressing both the N- and C-termini of flagellin, or PrgJ, appear to activate 6
NLRC4 without a requirement for NAIP5. Lastly, infection with Salmonella activates 7
the NLRC4 inflammasome in a partially NAIP5-dependent, partially NAIP5-independent 8
manner. 9
The NLRC4 inflammasome has yet to be reconstituted or purified, and thus the 10
biochemical role of NAIP5 in the activation of NLRC4 remains poorly understood. 11
Nevertheless, our results appear to indicate that NAIP5 affects the ability of the NLRC4 12
inflammasome to respond to specific stimuli. There are several possible models that 13
might account for the ability of NAIP5 to regulate the apparent specificity of NLRC4. 14
One model (the ‘potentiation’ model) is that NAIP5 functions as a potentiator of NLRC4 15
activation. In this model, the NLRC4 stimuli that exhibit a requirement for NAIP5 are 16
intrinsically ‘weak’ activators of NLRC4 and thus require NAIP5 to activate NLRC4. 17
For example, the C-terminus of flagellin might be poorly structured in the absence of the 18
N-terminus of flagellin and may therefore be a relatively poor activator of NLRC4. The 19
presence of NAIP5 could stabilize the C-terminus of flagellin, and thus improve its 20
ability to stimulate NLRC4. NAIP5 might also potentiate NLRC4 responses by acting 21
downstream of flagellin recognition, and act to amplify NLRC4-dependent activation of 22
CASP-1 in response to a weak stimulus. We found that the N-terminus of flagellin can 23
relieve the requirement for NAIP5 in NLRC4 activation. In the potentiation model, the 24
N-terminus might function to stabilize the C-terminus, thereby improving its signaling 25
capacity, and thus relieving the requirement for NAIP5. A direct biochemical interaction 26
between flagellin and NLRC4 or NAIP5 has yet to be demonstrated; thus, these models 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 19 –
are difficult to distinguish at present. It is also not clear whether Legionella is in fact a 1
‘weaker’ activator of NLRC4 than is Salmonella, as would be implied under the 2
potentiation model. In fact, at similar MOI, Legionella and Salmonella appear to induce 3
comparable levels of NLRC4-dependent cell death (Figure 3). It is also not clear whether 4
PrgJ, a NAIP5-independent activator of NLRC4, is a substantially stronger activator of 5
NLRC4 than is flagellin. In fact, transfection of wildtype macrophages with equivalent 6
amounts of each protein activates NLRC4 similarly, but PrgJ appears to be a much better 7
activator of NLRC4 specifically in Naip5-/-
macrophages. Thus, it is not clear whether 8
our results can be readily explained by a model in which NAIP5 functions primarily to 9
potentiate NLRC4 signaling. 10
Although it is widely assumed that NLRC4 is the proximal sensor of the ligands 11
that leads to its activation, this has never been formally demonstrated. In fact, it is 12
possible that NAIP5, or another host protein, is the true direct cytosolic sensor (or 13
‘receptor’) of flagellin. Thus, another model to explain our data could be that NAIP5 14
functions not as a general potentiator of NLRC4 activation/signaling, but as a specific 15
sensor of certain bacterial ligands. We call this latter model the ‘specificity’ model. For 16
example, NAIP5 may specifically recognize the C-terminus of flagellin, whereas another 17
host protein may specifically recognize full-length flagellin or PrgJ. This model could 18
explain the differential NAIP5-dependence for NLRC4 activation by Legionella versus 19
Salmonella, since Salmonella expresses PrgJ, a NAIP5-independent stimulus, whereas 20
Legionella does not. The specificity model is difficult to address until direct evidence is 21
obtained that NAIP5, NLRC4, or another host protein is the true receptor for flagellin 22
and/or PrgJ. 23
It was interesting that full-length flagellin delivered by protein transfection or 24
Listeria activated NLRC4 in a NAIP5-dependent manner (Figure 4B, 4C), whereas full-25
length flagellin expressed from a retroviral construct activated NLRC4 in a NAIP5-26
independent manner (Figure 1B; (14)). Although there are many possible explanations 27
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 20 –
for this result, one plausible idea is that the very high levels of flagellin expression 1
attained via retroviral transduction are responsible for relieving the requirement for 2
NAIP5. Thus, both the expression levels of the stimulus, as well as its molecular 3
characteristics, may influence the requirement for NAIP5. 4
Regardless of the underlying molecular mechanism, our data illustrate the ability 5
of NLRC4 to respond to distinct bacterial ligands. In future studies, it will therefore be 6
important to determine the specific molecular role that NAIP5 plays in NLRC4 7
activation. 8
9
Acknowledgements 10
This work was supported by Investigator Awards to R.E.V. from the Burroughs 11
Wellcome Fund and the Cancer Research Institute as well as by NIH grants (AI075039 12
and AI080749). J.P. was supported by Stiftelsen Olle Engkvist Byggmästare through the 13
Swedish Research Council. JD.S. was supported by the American Cancer Society PF-07-14
066-01-LIB. We also thank Tom Alber for help with the analysis of the putative coiled 15
coil in the D0 region of flagellin. 16
17
Figure 1. Role for the N-terminus of flagellin in NLRC4 activation. 18
(A) Diagram of all retroviral constructs used in transduction experiments. Note that all 19
constructs expressed GFP, either as a direct fusion to L. pneumophila flagellin (FlaA, as 20
shown), or downstream of an internal ribosome entry site (IRES) (not shown). C65, C-21
terminal 65 amino acids of FlaA. C20, C-terminal 20 amino acids of FlaA. N65, N-22
terminal 65 amino acids of FlaA. FlaA∆N constructs lack the indicated number of amino 23
acids from the N-terminus. (B-C) Wild-type (C57BL/6), or isogenic Naip5-/-
or Nlrc4-/-
24
macrophages were transduced with the indicated constructs and the percentage of GFP+ 25
cells was enumerated by flow cytometry 3-4 days after transduction. (D-E) Wild-type, 26
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 21 –
Naip5-deficient or Naip5-/-
Nlrc4-/-
doubly-deficient macrophages were transduced with the 1
indicated constructs and the percentage of GFP+ cells was enumerated by flow cytometry 2
3-4 days after transduction. >10,000 cells were analyzed for each experiment. For each 3
panel, data shown are from a single representative experiment of at least three that 4
produced similar results. 5
6
Figure 2. The N-terminus of flagellin can relieve requirement for NAIP5 in flagellin 7
sensing even when expressed on a distinct molecule from the C-terminus. 8
A retroviral construct expressing the N-terminal 65 amino acids of flagellin fused to GFP, 9
and a separate retroviral construct expressing the C-terminal 65 amino acids of flagellin 10
fused to mCherry, were transduced simultaneously into (A) wild-type, (B) Naip5-/-
, or (C) 11
Nlrc4-/-
macrophages, and the percentage of cells expressing GFP and/or mCherry was 12
enumerated by flow cytometry 4 days post transduction. The percentage of double-13
positive cells among total transduced cells is indicated in parentheses. The results are 14
representative of two representative experiments. 15
16
Figure 3. Differential requirement for NAIP5 in sensing Legionella and Salmonella 17
is dictated by the bacterium and not the flagellin itself. 18
(A) Cell death was assessed by measurement of lactate dehydrogenase (LDH) release 19
from wild-type (B6), isogenic Naip5-/-
, or isogenic Naip5-/-
Nlrc4-/-
doubly-deficient 20
macrophages infected with either wild-type (WT) Legionella (LP02), isogenic flagellin-21
deficient Legionella (∆flaA), wild-type Salmonella (LT2), or isogenic flagellin-deficient 22
Salmonella (fliC/fljB-). Gentamicin (100ug/mL) was added 30 min after infection to kill 23
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 22 –
extracellular bacteria. (B) Cell death was assessed by LDH release from B6, Naip5-/-
, or 1
Nlrc4-/-
macrophages infected with WT Legionella (LP02), ∆flaA, or Legionella 2
expressing Salmonella fliC from the Legionella chromosome in place of its native flaA 3
gene (∆flaA::fliC). Gentamicin (100ug/mL) was added 30 min after infection to kill 4
extracellular bacteria. (C) IL-1β release assayed from macrophages infected with the 5
indicated strains, as in panel B. Macrophages were pretreated with tripalmitoyl cysteinyl 6
seryl tetralysine lipopeptide (Pam3CSK4, 0.5µg/ml) for 3.5 hours to induce expression of 7
pro-IL-1β prior to infection. (D) Growth of ∆flaA or ∆flaA::fliC Legionella in B6 wild-8
type (WT), isogenic Naip5-/-
or isogenic Nlrc4-/-
macrophages was assayed by 9
determining colony forming units at the time points indicated. *P<.001. Two-Way Anova 10
and Bonferroni’s test were used to determine the statistical significance (P) of differences 11
in IL-1β and LDH release (3B-3C) against infection with ∆flaA Legionella. For each 12
panel, data shown are from a single representative experiment of at least two that 13
produced similar results. 14
15
16
Figure 4. Sensing of Salmonella PrgJ is independent of NAIP5. 17
(A) Flow cytometry of wild-type (WT; C57BL/6), isogenic Naip5-/-
or Nlrc4-/-
18
macrophages transduced with retroviruses expressing N65-GFP, GFP-C65 or Salmonella 19
PrgJ. The PrgJ construct expressed GFP downstream of an internal ribosome entry site. 20
(B) IL-1β release by wild-type, Naip5-/-
or Nlrc4-/-
macrophages transfected with purified 21
Salmonella flagellin (FliC), PrgJ or SsaI proteins. (C) Cell death assessed by LDH 22
released from wild-type, Naip5-/-
or Nlrc4-/-
macrophages infected with wild-type 23
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 23 –
Listeria, Listeria expressing PrgJ, or Listeria expressing Legionella FlaA. Expression of 1
PrgJ or FlaA was induced by addition of the indicated concentrations of IPTG. *P<.001. 2
Two-Way Anova and Bonferroni’s test were used to determine the statistical significance 3
(P) of differences in IL-1β and LDH release (4B-4C). In 4B-4C, significance tests were 4
done versus WT macrophages or versus both Naip5-/-
and WT macrophages when 5
transfecting with PrgJ (4B) or infecting with Listeria expressing PrgJ (4C). For each 6
panel, data shown are from a single representative experiment of at least two that 7
produced similar results. 8
9
10
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 24 –
References 1
1. Amer, A., L. Franchi, T. D. Kanneganti, M. Body-Malapel, N. Ozoren, G. 2
Brady, S. Meshinchi, R. Jagirdar, A. Gewirtz, S. Akira, and G. Nunez. 2006. 3
Regulation of Legionella phagosome maturation and infection through flagellin 4
and host IPAF. J Biol Chem. 5
2. Bergsbaken, T., S. L. Fink, and B. T. Cookson. 2009. Pyroptosis: host cell 6
death and inflammation. Nat Rev Microbiol 7:99-109. 7
3. Boyden, E. D., and W. F. Dietrich. 2006. Nalp1b controls mouse macrophage 8
susceptibility to anthrax lethal toxin. Nat Genet 38:240-244. 9
4. Coers, J., R. E. Vance, M. F. Fontana, and W. F. Dietrich. 2007. Restriction of 10
Legionella pneumophila growth in macrophages requires the concerted action of 11
cytokine and Naip5/Ipaf signalling pathways. Cell Microbiol 9:2344-2357. 12
5. Damiano, J. S., V. Oliveira, K. Welsh, and J. C. Reed. 2004. Heterotypic 13
interactions among NACHT domains: implications for regulation of innate 14
immune responses. Biochem J 381:213-219. 15
6. Decker, T., and M. L. Lohmann-Matthes. 1988. A quick and simple method for 16
the quantitation of lactate dehydrogenase release in measurements of cellular 17
cytotoxicity and tumor necrosis factor (TNF) activity. J Immunol Methods 18
115:61-69. 19
7. Fernandes-Alnemri, T., J. W. Yu, C. Juliana, L. Solorzano, S. Kang, J. Wu, 20
P. Datta, M. McCormick, L. Huang, E. McDermott, L. Eisenlohr, C. P. 21 Landel, and E. S. Alnemri. 2010. The AIM2 inflammasome is critical for innate 22
immunity to Francisella tularensis. Nat Immunol 11:385-393. 23
8. Fischetti, V. A., and American Society for Microbiology. 2006. Gram-positive 24
pathogens, 2nd ed. ASM Press, Washington, D.C. 25
9. Franchi, L., A. Amer, M. Body-Malapel, T. D. Kanneganti, N. Ozoren, R. 26
Jagirdar, N. Inohara, P. Vandenabeele, J. Bertin, A. Coyle, E. P. Grant, and 27 G. Nunez. 2006. Cytosolic flagellin requires Ipaf for activation of caspase-1 and 28
interleukin 1beta in salmonella-infected macrophages. Nat Immunol 7:576-582. 29
10. Janeway, C. A., Jr. 1989. Approaching the asymptote? Evolution and revolution 30
in immunology. Cold Spring Harb Symp Quant Biol 54 Pt 1:1-13. 31
11. Jones, J. W., N. Kayagaki, P. Broz, T. Henry, K. Newton, K. O'Rourke, S. 32
Chan, J. Dong, Y. Qu, M. Roose-Girma, V. M. Dixit, and D. M. Monack. 33 2010. Absent in melanoma 2 is required for innate immune recognition of 34
Francisella tularensis. Proc Natl Acad Sci U S A 107:9771-9776. 35
12. Kim, S., F. Bauernfeind, A. Ablasser, G. Hartmann, K. A. Fitzgerald, E. 36
Latz, and V. Hornung. 2010. Listeria monocytogenes is sensed by the NLRP3 37
and AIM2 inflammasome. Eur J Immunol 40:1545-1551. 38
13. Lauer, P., M. Y. Chow, M. J. Loessner, D. A. Portnoy, and R. Calendar. 39
2002. Construction, characterization, and use of two Listeria monocytogenes site-40
specific phage integration vectors. J Bacteriol 184:4177-4186. 41
14. Lightfield, K. L., J. Persson, S. W. Brubaker, C. E. Witte, J. von Moltke, E. 42
A. Dunipace, T. Henry, Y. H. Sun, D. Cado, W. F. Dietrich, D. M. Monack, 43 R. M. Tsolis, and R. E. Vance. 2008. Critical function for Naip5 in 44
inflammasome activation by a conserved carboxy-terminal domain of flagellin. 45
Nat Immunol 9:1171-1178. 46
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 25 –
15. Mariathasan, S., K. Newton, D. M. Monack, D. Vucic, D. M. French, W. P. 1
Lee, M. Roose-Girma, S. Erickson, and V. M. Dixit. 2004. Differential 2
activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 3
430:213-218. 4
16. Martinon, F., K. Burns, and J. Tschopp. 2002. The inflammasome: a molecular 5
platform triggering activation of inflammatory caspases and processing of proIL-6
beta. Mol Cell 10:417-426. 7
17. Miao, E. A., C. M. Alpuche-Aranda, M. Dors, A. E. Clark, M. W. Bader, S. I. 8
Miller, and A. Aderem. 2006. Cytoplasmic flagellin activates caspase-1 and 9
secretion of interleukin 1beta via Ipaf. Nat Immunol 7:569-575. 10
18. Miao, E. A., D. P. Mao, N. Yudkovsky, R. Bonneau, C. G. Lorang, S. E. 11
Warren, I. A. Leaf, and A. Aderem. 2010. Innate immune detection of the type 12
III secretion apparatus through the NLRC4 inflammasome. Proc Natl Acad Sci U 13
S A 107:3076-3080. 14
19. Molofsky, A. B., B. G. Byrne, N. N. Whitfield, C. A. Madigan, E. T. Fuse, K. 15
Tateda, and M. S. Swanson. 2006. Cytosolic recognition of flagellin by mouse 16
macrophages restricts Legionella pneumophila infection. J Exp Med 203:1093-17
1104. 18
20. Ormo, M., A. B. Cubitt, K. Kallio, L. A. Gross, R. Y. Tsien, and S. J. 19
Remington. 1996. Crystal structure of the Aequorea victoria green fluorescent 20
protein. Science 273:1392-1395. 21
21. Rathinam, V. A., Z. Jiang, S. N. Waggoner, S. Sharma, L. E. Cole, L. 22
Waggoner, S. K. Vanaja, B. G. Monks, S. Ganesan, E. Latz, V. Hornung, S. 23 N. Vogel, E. Szomolanyi-Tsuda, and K. A. Fitzgerald. 2010. The AIM2 24
inflammasome is essential for host defense against cytosolic bacteria and DNA 25
viruses. Nat Immunol 11:395-402. 26
22. Ren, T., D. S. Zamboni, C. R. Roy, W. F. Dietrich, and R. E. Vance. 2006. 27
Flagellin-deficient Legionella mutants evade caspase-1- and Naip5-mediated 28
macrophage immunity. PLoS Pathog 2:e18. 29
23. Sauer, J. D., C. E. Witte, J. Zemansky, B. Hanson, P. Lauer, and D. A. 30
Portnoy. 2010. Listeria monocytogenes triggers AIM2-mediated pyroptosis upon 31
infrequent bacteriolysis in the macrophage cytosol. Cell Host Microbe 7:412-419. 32
24. Schroder, K., R. Zhou, and J. Tschopp. 2010. The NLRP3 inflammasome: a 33
sensor for metabolic danger? Science 327:296-300. 34
25. Sun, Y. H., H. G. Rolan, and R. M. Tsolis. 2007. Injection of flagellin into the 35
host cell cytosol by Salmonella enterica serotype typhimurium. J Biol Chem. 36
26. Takeuchi, O., and S. Akira. 2010. Pattern recognition receptors and 37
inflammation. Cell 140:805-820. 38
27. Vance, R. E., R. R. Isberg, and D. A. Portnoy. 2009. Patterns of pathogenesis: 39
discrimination of pathogenic and nonpathogenic microbes by the innate immune 40
system. Cell Host Microbe 6:10-21. 41
28. Warren, S. E., A. Armstrong, M. K. Hamilton, D. P. Mao, I. A. Leaf, E. A. 42
Miao, and A. Aderem. 2010. Cutting edge: Cytosolic bacterial DNA activates 43
the inflammasome via Aim2. J Immunol 185:818-821. 44
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

– 26 –
29. Yonekura, K., S. Maki-Yonekura, and K. Namba. 2003. Complete atomic 1
model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2
424:643-650. 3
30. Zamboni, D. S., K. S. Kobayashi, T. Kohlsdorf, Y. Ogura, E. M. Long, R. E. 4
Vance, K. Kuida, S. Mariathasan, V. M. Dixit, R. A. Flavell, W. F. Dietrich, 5 and C. R. Roy. 2006. The Birc1e cytosolic pattern-recognition receptor 6
contributes to the detection and control of Legionella pneumophila infection. Nat 7
Immunol 7:318-325. 8
9
10
on May 3, 2018 by guest
http://iai.asm.org/
Dow
nloaded from