increased binding at 5-ht1a, 5-ht1b, and 5-ht2a receptors and 5-ht transporters in diet-induced...
TRANSCRIPT

Ž .Brain Research 847 1999 90–97www.elsevier.comrlocaterbres
Research report
Increased binding at 5-HT , 5-HT , and 5-HT receptors and 5-HT1A 1B 2A
transporters in diet-induced obese rats
Soyoung Park ), Joanne A. Harrold, Peter S. Widdowson, Gareth WilliamsDiabetes and Endocrinology Research Group, Department of Medicine, UniÕersity of LiÕerpool, Duncan Building, Daulby Street, LiÕerpool L69 3GA, UK
Accepted 31 August 1999
Abstract
Ž .5-Hydroxytryptamine 5-HT, serotonin , synthesized in midbrain raphe nuclei and released in various hypothalamic sites, decreasesfood intake but the specific 5-HT receptor subtypes involved are controversial. Here, we have studied changes in the regional density ofbinding to 5-HT receptors and transporters and the levels of tryptophan hydroxylase, in rats with obesity induced by feeding a palatablehigh-energy diet for 7 weeks. We mapped binding at 5-HT receptor subtypes and transporters using quantitative autoradiography and
Ž .determined tryptophan hydroxylase protein levels by Western blotting. In diet-induced obese DiO rats, specific binding to 5-HT1AŽw3 x . Ž . Ž .receptors H 8-OH-DPAT was significantly increased in the dorsal and median raphe by 90% P-0.01 and 132% P-0.05 ,
Žw125 x .respectively, compared with chow-fed controls. 5-HT receptor binding sites I cyanopindolol were significantly increased in the1BŽ . Ž . Žw3 x .hypothalamic arcuate nucleus ARC of DiO rats 58%; P-0.05 , as were 5-HT receptor binding sites H ketanserin in both the2A
Ž . Ž . Ž . Žw3 xARC 44%; P-0.05 and lateral hypothalamic area LHA 121%; P-0.05 . However, binding to 5-HT receptors H mesulger-2C.gine in DiO rats was not significantly different from that in controls in any hypothalamic region. Binding to 5-HT transporters
Žw3 x . Ž . Ž .H paroxetine was significantly increased P-0.05 in both dorsal and median raphe, paraventricular nuclei PVN , ventromedialŽ . Ž .nuclei VMH , anterior hypothalamic area AHA and LHA of DiO rats, by 47%–165%. Tryptophan hydroxylase protein levels in the
raphe nuclei were not significantly different between controls and DiO rats. In conclusion, we have demonstrated regionally specificchanges in binding to certain 5-HT receptor subtypes in obesity induced by voluntary overeating of a palatable diet. Overall, thesechanges are consistent with reduced 5-HT release and decreased activity of the 5-HT neurons. Reduction in the hypophagic action of5-HT, possibly acting at 5-HT , 5-HT and 5-HT receptors, may contribute to increased appetite in rats presented with highly1A 1B 2A
palatable diet. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: 5-HT; 5-HT receptor; 5-HT transporter; Hypothalamus; Diet-induced obesity
1. Introduction
Ž .5-Hydroxytryptamine 5-HT, serotonin powerfully in-hibits feeding in rodents when injected into various hy-
w xpothalamic regions 25 , and drugs which increase 5-HTw xavailability in the synaptic cleft, such as fluoxetine 15 ,
w x w xdexfenfluramine 31 and sibutramine 24 , cause weightloss in obese humans.
5-HT neurons are mainly located in the midbrain raphew xnuclei 7,20 , and project to widespread regions in the
central nervous system, including the hypothalamusw x20,40,41 which plays a key role in the regulation of
w xenergy homeostasis 6,48 . In recent years, numerous 5-HT
) Corresponding author. Fax: q44-151-706-5797; e-mail:[email protected]
w xreceptors have been identified 19 and some have beenimplicated in modulating food intake, notably the 5-HT1A
autoreceptor, and the 5-HT , 5-HT and 5-HT sub-1B 2A 2C
types which may mediate 5-HT’s anorexic effectw x3,5,16,28 . Presynaptic 5-HT sites are concentrated in1A
the dorsal and median raphe nuclei, and stimulation ofthese receptors inhibits firing rates of the 5-HT neuronsw x14 . Systemic and central injection of the 5-HT receptor1A
w xagonist, 8-OH-DPAT, induces hyperphagia 3 due to inhi-w xbition of 5-HT release in hypothalamic sites 43 . 5-HT1B
receptors in the hypothalamus are both presynaptic autore-w xceptors and postsynaptic receptors 29 ; the 5-HT ago-1B
nist, CP 93,129, markedly reduces food intake when in-Ž . w xjected into the paraventricular nuclei PVN 28 . A 5-HT2A
agonist injected into the PVN attenuates neuropeptide Y-w xinduced hyperphagia 16 , while injection of the 5-HT2C
w xreceptor antagonist, RS-102221, produces hyperphagia 5 .
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 99 02055-7

( )S. Park et al.rBrain Research 847 1999 90–97 91
The 5-HT system may mediate changes in food intakewhen nutritional status is altered, and its role in obesity isthe focus of this paper. Some studies have demonstratedthat hyperphagic, genetically obese Zucker rats have al-
w xtered hypothalamic 5-HT activity 38,39,45 in both 12-w x w xday-old 39 and adult obese Zucker rats 38 . In diet-in-
Ž .duced obese DiO rats, which mimic human obesity moreclosely than monogenic mutants such as the far fa rat, therole of the 5-HT system is unclear. In rats made obese byfeeding a high-energy diet, 5-HT levels are decreased in
w xthe hippocampus and brainstem 22 , while 5-HT-stimu-Žlated prolactin release an indirect marker of serotonergic
. w xtone is unchanged in cafeteria diet-fed rats 42 .Here, we have investigated the contribution of the 5-HT
system to obesity induced in normal rats by feeding apalatable diet. We measured the density of binding to the
Žmain 5-HT receptor subtypes implicated in feeding 5-.HT , 5-HT , 5-HT , 5-HT and to 5-HT transporters,1A 1B 2A 2C
using quantitative autoradiography of midbrain and hy-pothalamic sections. We speculated that sustained changesin 5-HT neuronal activity would lead to reciprocal alter-ations in binding to the specific 5-HT receptor subtypesinvolved, consistent with observations that increased ordecreased 5-HT release leads to fewer or greater 5-HT
w xbinding sites, respectively 10,21,29 . We also measuredtryptophan hydroxylase levels by Western blotting. Trypto-phan hydroxylase which catalyzes the rate-limiting step of
w x5-HT synthesis 9 , is located throughout serotonergic neu-rons, and along with the pre-synaptic ion-dependent trans-
w xporters 30 , is a potentially important regulatory site for5-HT neuronal activity.
2. Materials and methods
2.1. Animals
ŽForty-two male Wistar rats approximately 200 g;.breeding stock, Liverpool University, UK were random-
ized into two groups under environmentally controlledŽ .conditions 22"28C, humidity 30%–40% and a 12:12-h
Ž .lightrdark cycle lights on at 0700 h . They were allowedtap water ad libitum throughout. Half of the rats were fed
Žstandard laboratory chow CRM, Labsure, Poole, Dorset,.UK , providing 65% energy as carbohydrate, 24% as
protein and 9% as fat. The other half were fed a highlypalatable diet for 7 weeks, consisting of 33% chow, 33%Nestle condensed milk and 7% sucrose by weight, with theremainder being water. This diet provided 68% energy ascarbohydrate, 17% as protein and 13% as fat. As the twodiets were of a similar nutrient composition, their blood
Ž .tryptophan–large neutral amino acid TrprLNAA ratioswere also comparable, ensuring that the test diet did notdirectly introduce changes in 5-HT levels within thesynapse. In addition, it was verified that no single compo-nent of the test diet contained levels of tryptophan suffi-
cient to alter the TrprLNAA ratio. Therefore, any ob-served changes reflect alterations in the 5-HT system andits receptors.
Rats were killed by carbon dioxide inhalation and bloodremoved from the heart for measurement of plasma leptinand insulin concentrations using radioimmunoassay kits
Ž .from Linco Biogenesis Dorset, UK and Pharmacia Up-Ž .john Buck, UK , respectively, and of plasma glucose
using a glucose oxidase-based kit from BoehringerŽ .Mannheim Lewes, Sussex, UK . Epididymal and perirenal
white adipose tissues and gastrocnemius muscles wereŽ .removed, weighed and their ratio fatrlean calculated as
an index of body composition.For autoradiography, brains were immediately removed
Ž .and frozen in cold isopentane y308C to y408C , thenstored at y408C. For Western blotting, rear portion ofbrains were removed by cutting between the pineal glandand cerebellum and then two 1 mm3 blocks were dissectedout using a dissection microscope, each containing thedorsal and median raphe nuclei. The blocks were placed in1.5-ml microcentrifuge tubes containing 250 ml of 30 mM
Ž .Tris–HCl pH 7.8 for homogenization.
2.2. Autoradiography
Using a microtome, 20-mm sections were cut from ratŽ .brain blocks at the levels of the arcuate nucleus ARC ,
PVN and raphe nuclei, and the sections thaw-mounted ongelatin-coated microscope slides. Sections were then driedat 48C overnight and stored at y408C before being briefly
Ž .thawed at room temperature RT prior to incubation. Eachof two slides bearing adjacent pairs of sections were
w3 xprocessed for total and nonspecific binding. H 8-OH-Ž . w3 x Ž .DPAT 127 Cirmmol , H ketanserin 66 Cirmmol ,
w3 x Ž . w125 xH paroxetine 15 Cirmmol and I cyanopindololŽ .2200 Cirmmol were purchased from NEN DuPontŽ . w3 x ŽStevenage, Herts, UK , and H mesulergine 83 Cir
. Žmmol was purchased from Amersham Little Chalfont,.Bucks, UK .
2.2.1. 5-HT receptors1 Aw3 xH 8-OH-DPAT binding was performed in sections cut
at the level of the raphe nuclei. These were preincubatedfor 30 min in 170 mM Tris–HCl supplemented with 4 mM
Ž .CaCl 2H O and 0.1% ascorbate pH 7.4 at 258C . The2 2
sections were subsequently transferred to the same bufferw3 xcontaining 2 nM H 8-OH-DPAT in the presence or ab-
sence of unlabeled 1 mM 5-HT for measurement of non-specific binding. After incubating for 60 min at RT, sec-tions were washed twice in ice-cold buffer for 5 min,rapidly dipped in ice-cold distilled water to remove buffersalts, and quickly dried with a stream of warm air. Slides
3 Žwere apposed to H-sensitive film Hyperfilme, Amer-. 3sham in X-ray cassettes with H autoradiography stan-Ž .dards Microscalese, Amersham for 4–6 weeks. The
autoradiographs were developed and fixed with Kodak

( )S. Park et al.rBrain Research 847 1999 90–9792
GBX, and the receptor density in the dorsal and medianŽraphe nuclei measured using computer densitometry AIS
system, Imaging Technology, Brock University, St..Catharine’s, Ontario, Canada . Adjacent sections were
stained with Cresyl violet and the anatomical regionsconfirmed using the Paxinos and Watson rat brain stereo-
w xtactic atlas 33 .
2.2.2. 5-HT receptors1Bw125 xTo measure I cyanopindolol binding, sections at
ARC and PVN levels were preincubated for 20 min in 50mM Tris supplemented with 154 mM NaCl and 10 mM
Ž .pargyline pH 7.7 at 258C . Sections were then transferredw125 xto the same buffer containing 150 pM I cyanopindolol,
Ž .30 nM NAN-190 a 5-HT blocker and 30 mM isoprote-1AŽ .renol a general b-adrenergic blocker in the presence or
absence of unlabeled 10 mM 5-HT to determine non-specific binding. After incubating for 90 min at RT, sec-tions were washed twice for 5 min in ice-cold buffer,rapidly dipped in ice-cold distilled water to remove buffersalts, and quickly dried with a stream of warm air. Slides
3 Žwere apposed to H-sensitive film Hyperfilme, Amer-. 125sham in X-ray cassettes with I autoradiography stan-Ž .dards Microscalese, Amersham for 24 h. Sections were
processed as for 5-HT .1A
2.2.3. 5-HT receptors2 Aw3 xTo measure H ketanserin binding, sections at ARC
and PVN levels were preincubated for 15 min in 170 mMŽ .Tris pH 7.4 at 258C and then transferred to the same
w3 xbuffer containing 2 nM H ketanserin in the presence orabsence of unlabeled 1 mM cinanserine for non-specificbinding. After incubating for 30 min at RT, sections werewashed twice for 10 min in ice-cold buffer, rapidly dippedin ice-cold distilled water to remove buffer salts, andquickly dried as above. Sections were processed as for5-HT .1A
2.2.4. 5-HT receptors2Cw3 xTo measure H mesulergine binding, sections contain-
ing ARC and PVN nuclei were preincubated for 15 min inŽ .170 mM Tris pH 7.4 at 258C and then transferred to the
w3 xsame buffer containing 5 nM H mesulergine in the pres-ence or absence of unlabeled 10 mM mianserine for non-specific labelling. After incubating for 120 min at RT,sections were processed as for 5-HT .2A
2.2.5. 5-HT transportersw3 xTo measure H paroxetine binding, sections containing
the raphe, ARC and PVN nuclei were preincubated for 15min in 50 mM Tris supplemented with 120 mM NaCl and
Ž .5 mM KCl pH 7.4 at 258C . Sections were subsequentlytransferred to the same buffer containing 0.5 nMw3 xH paroxetine in the presence or absence of unlabeled 10mM fluoxetine for non-specific binding. After incubatingfor 60 min at RT, sections were processed as for 5-HT .2A
2.3. Western blotting for tryptophan hydroxylase
Western blotting was essentially carried out accordingw xto the method of Laemmli 23 . Briefly, midbrain dorsal
Žand median raphe blocks were sonicated Soniprep 150,. Ž .MSE, UK in 250 ml of 30 mM Tris–HCl pH 7.8 for 10
s and centrifuged for 4–5 min at 14,000 rpm. SupernatantŽ .50 ml was diluted with an equal volume of sample bufferŽ150 mM Tris–Cl, 25% glycerol, 2.5% b-mercaptoethanol,
.2.4% SDS, 0.25% bromophenol blue, pH 6.8 and heatedat 958C for 3 min. Protein concentration was determined
Žusing a commercially available kit BCA, Pierce, Rock-.ford, IL, USA .
Ž .Samples 100 mgrlane were separated by SDS–PAGEwith 10% acrylamide gel and then electrophoretically
Žtransferred to nitrocellulose membranes Hybonde-Cpure,. w xAmersham , according to the method of Towbin et al. 47 ,
at a constant current of 3 mArcm2 for 2–3 h at RT inŽtransfer buffer 25 mM Tris, 200 mM glycine and 20%
.methanol . Pre-stained SDS–PAGE molecular weightŽ .markers Rainbow markers; Amersham were electro-
phoresed in parallel lanes. After non-specific binding siteswere blocked for 30 min at RT with 3% fat-free milk inPBS, the membranes were washed briefly in PBS contain-ing 0.1% Tween-20e and then incubated overnight at 48Cwith a commercially available monoclonal tryptophan hy-
Ž .droxylase antibody Sigma, Dorset, UK . This well charac-terised antibody was diluted to the recommended concen-
Ž .tration for Western blotting 1:1000 in PBS containing3% milk. Membranes were washed with PBS for 30 minand then incubated with secondary anti-mouse peroxidase
Ž .labeled antibody Sigma , diluted 1:10,000 in PBS, for 1 hat RT. Following 3=5 min washes in PBS and 1=5 minwash in PBS containing 0.1% Tween-20e, membraneswere detected with Amersham’s ECL kit and exposed to
Ž .Hyperfilme-ECL Amersham for 2–10 min, ensuring thatthe signal obtained was in the middle of the linear rangeand the film was not saturated. Control for non-specificstaining by the secondary antibody was monitored andshown to be negative. Optical density readings for the
Table 1Body weight, fat and muscle mass, and plasma glucose, insulin and leptinlevels in control and 7 weeks DiO rats. Values expressed as mean"S.E.M.
Žfor groups of eight rats per group. Fatrlean: perirenal fat massq.epididymal fat mass rgastrocnemius muscle mass
Controls DiOUŽ .Final body weight g 450"5 510"7
UUŽ .Perirenal fat mass g 5.7"0.71 10.0"1.15UUŽ .Epididymal fat mass g 5.5"0.51 10.2"1.05
Ž .Muscle mass g 2.5"0.05 2.6"0.06UUFatrlean 4.6"0.5 7.5"0.6
Ž .Plasma glucose mmolrl 11.9"0.63 13.5"1.01UŽ .Plasma insulin mUrl 22.5"1.37 30.1"3.14UUUŽ .Plasma leptin ngrl 5.3"0.25 7.3"0.39
UP -0.05,
UUP -0.01,
UUUP -0.001 vs. controls.
()
S.Park
etal.r
Brain
Research
8471999
90–
9793
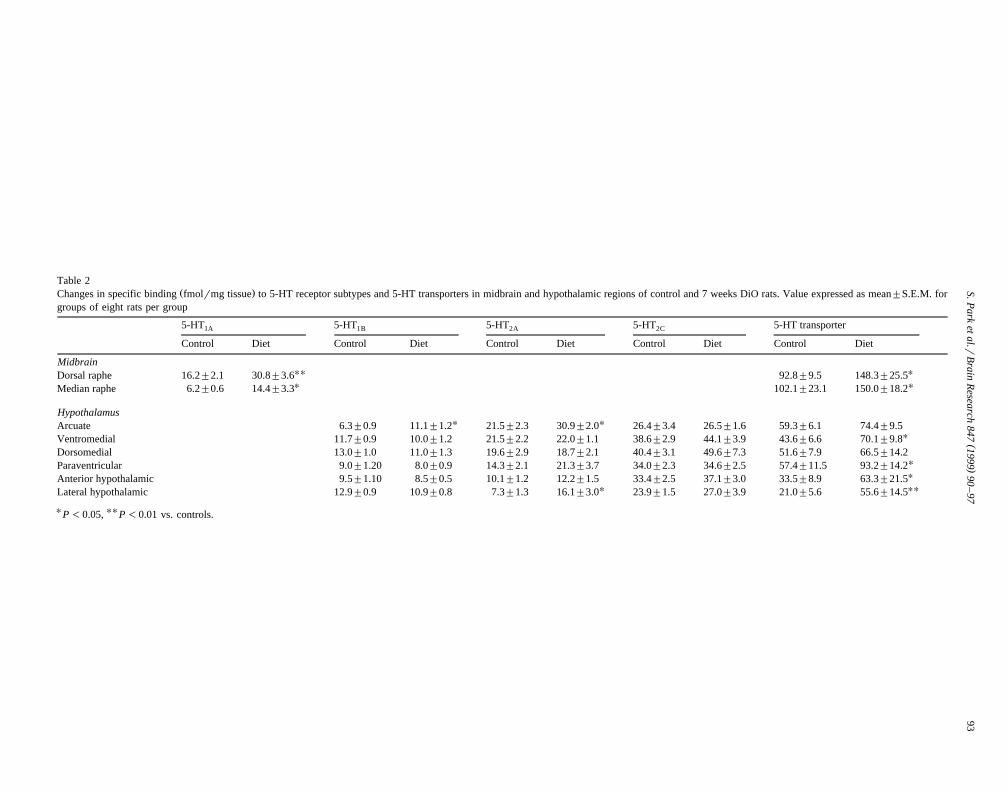
Table 2Ž .Changes in specific binding fmolrmg tissue to 5-HT receptor subtypes and 5-HT transporters in midbrain and hypothalamic regions of control and 7 weeks DiO rats. Value expressed as mean"S.E.M. for
groups of eight rats per group
5-HT 5-HT 5-HT 5-HT 5-HT transporter1A 1B 2A 2C
Control Diet Control Diet Control Diet Control Diet Control Diet
MidbrainUU UDorsal raphe 16.2"2.1 30.8"3.6 92.8"9.5 148.3"25.5U UMedian raphe 6.2"0.6 14.4"3.3 102.1"23.1 150.0"18.2
HypothalamusU UArcuate 6.3"0.9 11.1"1.2 21.5"2.3 30.9"2.0 26.4"3.4 26.5"1.6 59.3"6.1 74.4"9.5
UVentromedial 11.7"0.9 10.0"1.2 21.5"2.2 22.0"1.1 38.6"2.9 44.1"3.9 43.6"6.6 70.1"9.8Dorsomedial 13.0"1.0 11.0"1.3 19.6"2.9 18.7"2.1 40.4"3.1 49.6"7.3 51.6"7.9 66.5"14.2
UParaventricular 9.0"1.20 8.0"0.9 14.3"2.1 21.3"3.7 34.0"2.3 34.6"2.5 57.4"11.5 93.2"14.2UAnterior hypothalamic 9.5"1.10 8.5"0.5 10.1"1.2 12.2"1.5 33.4"2.5 37.1"3.0 33.5"8.9 63.3"21.5
U UULateral hypothalamic 12.9"0.9 10.9"0.8 7.3"1.3 16.1"3.0 23.9"1.5 27.0"3.9 21.0"5.6 55.6"14.5
UP -0.05,
UUP -0.01 vs. controls.
( )S. Park et al.rBrain Research 847 1999 90–9794
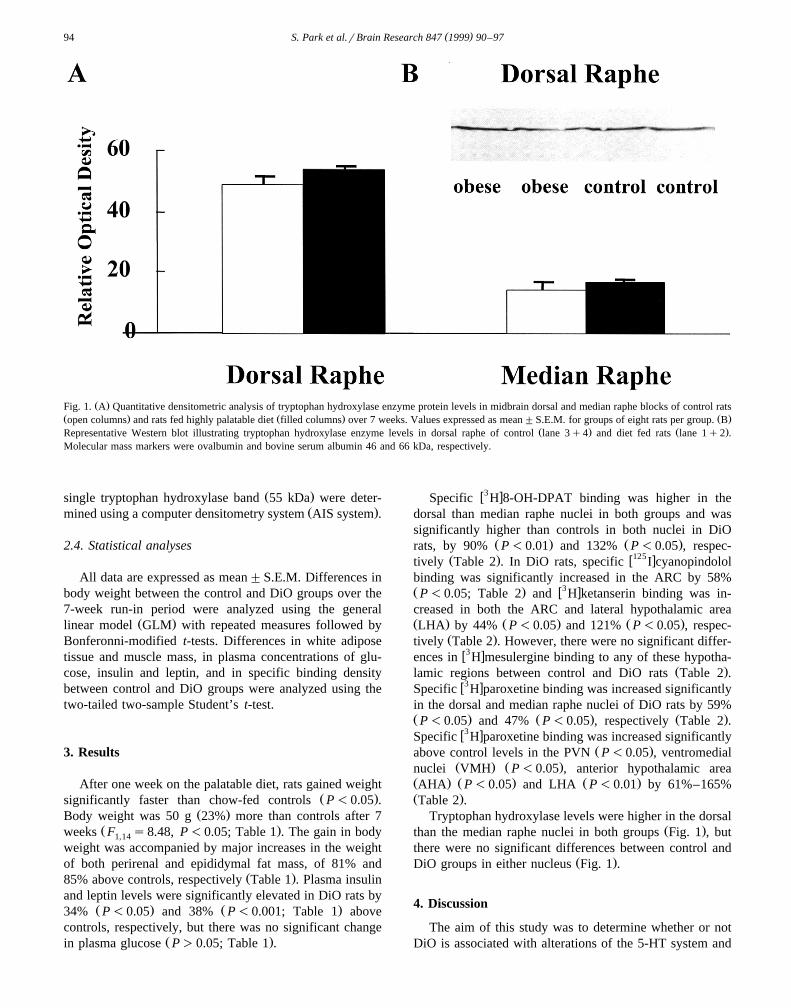
Ž .Fig. 1. A Quantitative densitometric analysis of tryptophan hydroxylase enzyme protein levels in midbrain dorsal and median raphe blocks of control ratsŽ . Ž . Ž .open columns and rats fed highly palatable diet filled columns over 7 weeks. Values expressed as mean"S.E.M. for groups of eight rats per group. B
Ž . Ž .Representative Western blot illustrating tryptophan hydroxylase enzyme levels in dorsal raphe of control lane 3q4 and diet fed rats lane 1q2 .Molecular mass markers were ovalbumin and bovine serum albumin 46 and 66 kDa, respectively.
Ž .single tryptophan hydroxylase band 55 kDa were deter-Ž .mined using a computer densitometry system AIS system .
2.4. Statistical analyses
All data are expressed as mean"S.E.M. Differences inbody weight between the control and DiO groups over the7-week run-in period were analyzed using the general
Ž .linear model GLM with repeated measures followed byBonferonni-modified t-tests. Differences in white adiposetissue and muscle mass, in plasma concentrations of glu-cose, insulin and leptin, and in specific binding densitybetween control and DiO groups were analyzed using thetwo-tailed two-sample Student’s t-test.
3. Results
After one week on the palatable diet, rats gained weightŽ .significantly faster than chow-fed controls P-0.05 .
Ž .Body weight was 50 g 23% more than controls after 7Ž .weeks F s8.48, P-0.05; Table 1 . The gain in body1,14
weight was accompanied by major increases in the weightof both perirenal and epididymal fat mass, of 81% and
Ž .85% above controls, respectively Table 1 . Plasma insulinand leptin levels were significantly elevated in DiO rats by
Ž . Ž .34% P-0.05 and 38% P-0.001; Table 1 abovecontrols, respectively, but there was no significant change
Ž .in plasma glucose P)0.05; Table 1 .
w3 xSpecific H 8-OH-DPAT binding was higher in thedorsal than median raphe nuclei in both groups and wassignificantly higher than controls in both nuclei in DiO
Ž . Ž .rats, by 90% P-0.01 and 132% P-0.05 , respec-Ž . w125 xtively Table 2 . In DiO rats, specific I cyanopindolol
binding was significantly increased in the ARC by 58%Ž . w3 xP-0.05; Table 2 and H ketanserin binding was in-creased in both the ARC and lateral hypothalamic areaŽ . Ž . Ž .LHA by 44% P-0.05 and 121% P-0.05 , respec-
Ž .tively Table 2 . However, there were no significant differ-w3 xences in H mesulergine binding to any of these hypotha-
Ž .lamic regions between control and DiO rats Table 2 .w3 xSpecific H paroxetine binding was increased significantly
in the dorsal and median raphe nuclei of DiO rats by 59%Ž . Ž . Ž .P-0.05 and 47% P-0.05 , respectively Table 2 .
w3 xSpecific H paroxetine binding was increased significantlyŽ .above control levels in the PVN P-0.05 , ventromedial
Ž . Ž .nuclei VMH P-0.05 , anterior hypothalamic areaŽ . Ž . Ž .AHA P-0.05 and LHA P-0.01 by 61%–165%Ž .Table 2 .
Tryptophan hydroxylase levels were higher in the dorsalŽ .than the median raphe nuclei in both groups Fig. 1 , but
there were no significant differences between control andŽ .DiO groups in either nucleus Fig. 1 .
4. Discussion
The aim of this study was to determine whether or notDiO is associated with alterations of the 5-HT system and

( )S. Park et al.rBrain Research 847 1999 90–97 95
its receptors. In rats fed a highly palatable diet for 7weeks, we found increased 5-HT receptor binding in the1A
midbrain dorsal and median raphe nuclei, increased 5-HT1B
binding in the ARC, and increased 5-HT in the ARC and2A
LHA. By contrast, no significant changes in binding to5-HT receptors were observed in any of these hypothala-2C
mic regions.5-HT neurons situated in both the dorsal and median
w xraphe nuclei project to the hypothalamus 20 . 5-HT1A
autoreceptors influence the activation not only of 5-HT1A
receptors themselves, but of all other types of 5-HT recep-tors as well, by inhibiting the firing rates of these neuronsw x10,14 . A single injection of the 5-HT agonist, 8-OH-1A
w xDPAT, produces functional desensitization 2 , while 2weeks of treatment with another 5-HT agonist, alne-1A
Ž .spirone S-20499 , significantly reduces 8-OH-DPAT bind-ing which in turn attenuates the ability of alnespirone to
w xreduce extracellular 5-HT release 10 . Seven days oftreatment with fluoxetine, which increases 5-HT availabil-ity in the raphe nuclei, results in functional desensitization
w xof 5-HT receptors 35 . Therefore, our reported increases1A
in the 5-HT autoreceptors in the dorsal and median raphe1A
of DiO rats, possibly reflecting decreased 5-HT availabilityin those regions, is expected to reduce 5-HT transmission.
Decreased synaptic availability of 5-HT may also leadto an increase in 5-HT and 5-HT binding sites in the1B 2A
w xhypothalamus and cortex 21,29 . Selective denervation ofŽ5-HT neurons by intracerebroventricular injection of 5,7
.dihydroxytryptamine causes increased binding at 5-HT1B
receptors in the entorhinal cortex and the dorsomedial andw xsuprachiasmatic nuclei of the hypothalamus 29 . More-
over, reduced 5-HT release induced by the 5-HT1Ar1B
receptor agonist, RU24969, increases 5-HT receptor2Aw xbinding 21 . We demonstrated regional increases in bind-
ing to 5-HT and 5-HT receptors in the hypothalamic1B 2A
ARC and LHA, both of which are important regions forw xfeeding behavior and energy metabolism 6 and also have
bi-directional connections with 5-HT neurons in the mid-w xbrain raphe 20 . Overall, our data suggest that the 5-HT
system in the ARC and LHA of the hypothalamus mightplay an important role in DiO.
Interestingly, 5-HT and 5-HT receptor knock-out1A 1B
mice show no changes in 5-HT levels in the brain, in foodw xintake or body weight 27,36 . This does not exclude these
receptors from playing a role in the regulation of feeding,as it is possible that other neurotransmitters and receptorsinvolved in appetite control may have undergone changesto compensate for their absence in the knock-out mice.
5-HT receptors have been strongly implicated in in-2C
hibiting feeding. For example, the selective 5-HT antag-2C
onist, RS-102221, increases food intake and body weightw xwhen injected intraperitoneally 5 and 5-HT receptor2C
w xknock-out mice are hyperphagic and obese 46 . Our in-ability to detect changes in the 5-HT receptor density2C
does not exclude these receptors from the regulation offood intake in the DiO rats, since there may be a large
receptor reserve, which could preclude measurable changesin receptor density. Alternatively, the observed difference
Žin receptor subtypes of the 5-HT family 5-HT and2 2A.5-HT could reflect their distinct regulation. Previous2C
studies suggest that this may be the case. For example,5-HT receptors have a much lower intrinsic ability to2A
spontaneously adopt or maintain the active receptor con-w xformation than 5-HT receptors 17 . The receptor sub-2C
types also respond differently to 24 h pre-treatment withw xthe serotonergic antagonist mianserin 32 .
We have suggested that the observed increase in bind-ing at 5-HT , 5-HT and 5-HT receptors in the mid-1A 1B 2A
brain and hypothalamus represents reduced activity of5-HT neurons. This is consistent with previous studies. Forexample, in Osborne–Mendel rats, which are sensitive todietary-fat induced obesity, D-fenfluramine, which stimu-lates 5-HT release, completely abolished hyperphagia andweight gain on a high-fat diet compared with S 5BrP1
w xrats, which are obesity-resistant 13 . It has also beenshown that CSF levels of the 5-HT metabolite, 5-HIAA,
w xare decreased in obese women 4 , and a reduced TrprLNAA ratio, which leads to decreased 5-HT synthesis in
w xbrain 40 , has been reported in the plasma of obese humanw xsubjects 1 . Additionally, it was found that supplementa-
Ž .tion with 5-hydroxytryptophan a precursor of 5-HT for 2weeks decreased daily energy intake and reduced bodyweight in obese non-insulin-dependent diabetic subjectsw x8 . However, from the present study, the regulatory factorscausing the proposed reduction in 5-HT activity in DiO arenot clear. It is possible that central or peripheral factorsaltered in obese subject may be responsible for this change.
We reported that binding to 5-HT transporters washigher in the raphe than in the hypothalamus of chow-fedrats, which is consistent with reported local levels of
w xrelease and synthesis 26,34 . After feeding a palatable dietfor 7 weeks, binding at 5-HT transporters in both midbrainand hypothalamus was increased compared with chowfedcontrols. Few studies have examined for changes in bind-ing density of 5-HT transporters in vivo under differentphysiological conditions, and so the regulatory factorsgoverning these phenomena are only partly understood. Ithas been shown that 5-HT depletion by p-chlorophenyl-alanine injection decreases 5-HT transporter mRNA levels
w3 xand expression of the gene product, measured by H cita-w xlopram binding 37 . This data is consistent with a regula-
tion of 5-HT transporters by 5-HT concentrations. How-ever, additional studies provide contradictory evidence asto the mechanism of transporter regulation. For example,whilst observations based on streptozotocin-induced dia-betic rats are consistent with this regulatory mechanism, in
w xthis case a reduction in 5-HT 44 is associated withw xincreased 5-HT transporter mRNA levels 34 . Further still,
w xstress, known to increase 5-HT release 11 , also produceschanges in the density of 5-HT transporter binding sites inrodents, but here the results are less conclusive. For exam-ple, shock stress increases the binding to 5-HT transporters

( )S. Park et al.rBrain Research 847 1999 90–9796
Ž . w xin the hippocampus of Sprague–Dawley SD rats 12 ,while restraint stress has no effect on 5-HT transporters inany of the area examined including the hippocampus in SD
w xrats 49 . Therefore, it is inferred from these data that theregulation of 5-HT transporters may not simply be gov-erned by the availability of 5-HT in the synaptic cleft, butby more complicated regulatory mechanisms. The possibleregulatory factors resulting in increased binding at 5-HTtransporters in DiO rats are not clear from the presentstudy.
The enzyme tryptophan hydroxylase is not saturatedunder normal physiological conditions, and thus is therate-limiting enzyme in 5-HT synthesis, making it animportant regulator of 5-HT activity. However, we foundno change in its levels in the raphe of DiO rats, despitereceptor alterations being observed. This does not excludethe enzyme from playing a role in the control of foodintake. It is possible that localized changes in enzymelevels within specific hypothalamic nuclei occur. Therecould also be alterations in the intrinsic activity of the
w xenzyme 18 . Measurement of the levels of enzyme sub-strate, tryptophan, would also help to clarify this issue, as5-HT synthesis not only depends on the amount of enzymeprotein but also its degree of saturation. Further workshould elucidate the role of tryptophan hydroxylase inregulating the reduced 5-HT activity proposed in DiO rats.
In conclusion, we deduce from the observed changes inbinding to 5-HT receptors that DiO in rats is associatedwith decreased 5-HT activity. Increased 5-HT transporternumbers may also play a role in the reduction of 5-HTavailability in synaptic cleft. However, we found no changein binding to the 5-HT receptor, which has been postu-2C
lated to mediate the appetite-suppressing effect of 5-HTw x46 .
Acknowledgements
This work was supported by the Korea Science andŽ .Engineering Foundation KOSEF and by the Liverpool
Diabetes Research Action Fund. We are grateful to RobinBuckingham for comments and to Peter King and LucyPickavance for discussion. We are also grateful to LeighThompson for his conscientious care of the animals.
References
w x1 D.V. Ashley, M.O. Fleury, A. Golay, E. Maeder, P.D. Leathwood,Evidence for diminished brain 5-hydroxytryptamine biosynthesis inobese diabetic and non-diabetic humans, Am. J. Clin. Nutr. 42Ž .1985 1240–1245.
w x2 M. Beer, G.A. Kennett, G. Curzon, A single dose of 8-OH-DPATw3 xreduces raphe binding of H 8-OH-DPAT and increase the effect of
raphe stimulation on 5-HT metabolism, Eur. J. Pharmacol. 178Ž .1990 179–187.
w x Ž .3 C. Bendotti, R. Samanin, 8-Hydroxy-2- di-n-propylamino tetralinŽ .8-OH-DPAT elicits eating in free-feeding rats by acting on central
Ž .serotonin neurons, Eur. J. Pharmacol. 121 1986 147–150.w x4 P. Bjorntorp, Neuroendocrine abnormalities in human obesity,
Ž .Metabolism 44 1995 238–241.w x5 D.W. Bonhaus, K.K. Weinhardt, M. Taylor, A. DeSouza, P.M.
McNeeley, K. Szczepanski, D.J. Fontana, J. Trinh, C.L. Rocha,M.W. Dawson, L.A. Flippin, R.M. Eglen, RS-102221: a novel highaffinity and selective, 5-HT receptor antagonist, Neuropharmacol-2C
Ž .ogy 36 1997 621–629.w x6 G.A. Bray, D.A. York, The MONALISA hypothesis in the time of
Ž .leptin, Horm. Res. 53 1998 95–117.w x7 M.J. Brownstein, M. Palkovits, Catecholamines, serotonin, acethy-
choline and g-aminobutyric acid in the rat brain: biochemical stud-ies, Handbook of Chemical Neuroanatomy. Vol. 2: Classical Trans-mitter in the CNS, Part I, Elsevier, Amsterdam, 1984, pp. 23–54.
w x8 C. Cangiano, A. Laviano, M. Del Ben, I. Preziosa, F. Angelico, A.Cascino, F. Rossi-Fanelli, Effects of oral 5-hydroxy-tryptophan onenergy intake and macronutrient selection in non-insulin dependent
Ž .diabetic patients, Int. J. Obes. Relat. Metab. Disord. 22 1998648–654.
w x9 A. Carlsson, M. Lindqvist, Dependence of 5-HT and catecholaminesynthesis on concentrations of precursor amino acids in the rat brain,
Ž .Naunyn-Schmiedeberg’s Arch. Pharmacol. 303 1978 157–164.w x10 J.M. Casanovas, M.T. Vilaro, G. Mengod, F. Artigas, Differential
regulation of somatodendritic serotonin 5-HT receptors by 2-week1AŽ .treatments with the selective agonists alnespirone S-20499 and
Ž .8-hydoxy-2 di-n-propylamino tetralin: microdialysis and autoradio-Ž .graphic studies in rat brain, J. Neurochem. 72 1999 262–272.
w x11 H.W. Clement, M. Kirsch, C. Hasse, C. Opper, W. Wesemann,Effect of repeated immobilization on serotonin metabolism in differ-ent rat brain areas and on serum corticosterone, J. Neural Transm.
Ž .105 1998 1155–1170.w x12 E. Edwards, W. Komrich, P.V. Houtten, F.A. Henn, Presynaptic
serotonin mechanisms in rats subjected to inescapable shock, Neu-Ž .ropsychopharmacology 31 1992 323–330.
w x13 J.S. Fisler, S.J. Underberger, D.A. York, G.A. Bray, D-Fenfluraminein a rat model of dietary fat-induced obesity, Pharmacol. Biochem.
Ž .Behav. 45 1993 487–493.w x14 A.M. Gardier, I. Malagie, A.C. Trillat, C. Jacquot, F. Artigas, Role
of 5-HT autoreceptors in the mechanism of action of serotonergic1A
antidepressant drug: recent findings from in vivo microdialysisŽ .studies, Fundam. Clin. Pharmacol. 10 1996 16–27.
w x15 D.J. Goldstein, A.H. Rampey, G.G. Enas, J.H. Potvin, L.A. Fludzin-ski, L.R. Levine, Fluoxetine: a randomized clinical trial in the
Ž .treatment of obesity, Int. J. Obes. Relat. Metab. Disord. 18 1994129–135.
w x16 G. Grignaschi, F. Sironi, R. Samanin, Stimulation of 5-HT recep-2A
tors in the paraventricular hypothalamus attenuate neuropeptide Y-induced hypophagia through activation of corticotrophin releasing
Ž .factor, Brain Res. 708 1996 173–176.w x17 M.S. Grotowiel, E. Sanders-Bush, Differences in agonist-indepen-
dent activity of 5-Ht2A and 5-Ht2c receptors revealed by heterolo-gous expression, Naunyn-Schmiedeberg’s Arch. Pharmacol. 359Ž .1999 21–27.
w x18 M. Hamon, S. Bourgoin, F. Artaud, S. El Mestikawy, The respectiveroles of tryptophan uptake and tryptophan hydroxylase in the regula-tion of serotonin synthesis in the central nervous system, J. Physiol.Ž . Ž .Paris 77 1981 269–279.
w x19 D. Hoyer, D.E. Clarke, J.R. Fozard, P.R. Hartig, G.R. Martin, E.J.Mylecharane, P.R. Saxena, P.P. Humphrey, International union ofpharmacology classification of receptors for 5-hydroxytryptamineŽ . Ž .serotonin , Pharmacol. Rev. 46 1993 157–203.
w x20 B.L. Jacobs, E.C. Azmitia, Structure and function of the brainŽ .serotonin system, Physiol. Rev. 72 1992 165–229.
w x21 H. Johanning, P. Plenge, E. Mellerup, Serotonin receptors in thebrain of rats treated chronically with imipramine or RU24969:

( )S. Park et al.rBrain Research 847 1999 90–97 97
support for the 5-HT receptor being a 5-HT autoreceptor, Pharma-1BŽ .col. Toxicol. 70 1992 131–134.
w x22 T.D. Kimbrough, L.B. Weekley, The effect of high-fat diet onbrainstem and duodenal serotonin metabolism in Sprague–Dawley
Ž .and Osborne–Mendel rats, Int. J. Obes. 8 1984 305–310.w x23 U.K. Laemmli, Cleavage of structural proteins during the assembly
Ž .of the head of bacteriophage T4, Nature 227 1970 680–685.w x24 M.E. Lean, Sibutramine — a review of clinical efficacy, Int. J.
Ž .Obes. Relat. Metab. Disord. 21 1997 S30–S36.w x25 S.F. Leibowitz, G.F. Weiss, G. Shor-Posner, Hypothalamic sero-
tonin: pharmacological, biochemical and behavioral analysis of itsŽ .feeding-suppressive action, Clin. Neuropharmacol. 11 1988 S51–
S71.w x26 K.P. Lesch, C.S. Aulakh, B.L. Wolozin, T.J. Tolliver, J.L. Hill, D.L.
Murphy, Regional brain expression of serotonin transporter mRNAand its regulation by reuptake inhibiting antidepressants, Brain Res.
Ž .Mol. Brain Res. 17 1993 31–35.w x27 J.J. Lucas, A. Yamamoto, K. Scearce-Levie, F. Saudou, R. Hen,
Absence of fenfluramine-induced anorexia and reduced c-fos induc-tion in the hypothalamus and central amygdala complex of serotonin
Ž .1B receptor Knock-Out mice, J. Neurosci. 18 1998 5537–5544.w x28 J.E. Macor, C.A. Burkhart, J.H. Heym, J.L. Ives, L.A. Lebel, M.E.
Newman, J.A. Nielsen, K. Ryan, D.W. Schulz, L.K. Torgersen, B.K.Ž . Ž .Koe, 3- 1,2,5,6-Tetrahydropyrid-4-yl pyrrol 3.2-b pyrid-5-one: a po-
tent and selective serotonin 1B agonist and rotationally restrictedŽ .phenolic analogue of 5-methoxy-3 1,2,5,6-tetrahydropyrid-4-yl in-
Ž .dols, J. Med. Chem. 33 1990 2087–2093.w x29 C. Manrique, L. Segu, F. Hery, M. Faudon, A.M. Francois-Bellan,
Increase of central 5-HT binding sites following 5,7-dihy-1BŽ .doxytryptamine axotomy in the adult rat, Brain Res. 623 1993
345–348.w x30 J.O. Marcusson, S.B. Ross, Binding of some antidepressants to the
5-HT transport in the brain and platelets, PsychopharmacologyŽ . Ž .Berlin 102 1990 145–155.
w x31 D. McTavish, R.C. Heel, Dexfenfluramine. A review of its phar-macological properties and therapeutic potential in obesity, Drugs 43Ž .1992 713–733.
w x32 R.A. Newton, J.M. Elliott, Mianserin-induced down-regulation ofhuman 5-hydroxytryptamine2A and 5-hydroxytryptamine2C recep-tors stably expressed in the human neuroblastoma cell line SH-SY5Y,
Ž .J. Neurochem. 69 1997 1031–1038.w x33 G. Paxinos, C. Watson, The Rat Brain, 2nd edn., Academic Press,
London, 1986.w x34 M.S. Petrisic, S.J. Augood, R.J. Bicknell, Monoamine transporter
gene expression in the central nervous system in diabetes mellitus, J.Ž .Neurochem. 68 1997 2435–2441.
w x35 E. Le Poul, N. Laaris, M. Hamon, L. Lanfumey, Fluoxetine-induceddesensitization of somatodendritic 5-HT autoreceptors is indepen-1A
Ž . Ž .dent of glucocorticoid s , Synapsis 27 1997 303–312.w x36 S. Ramboz, R. Oosting, D.A. Amara, H.F. Kung, P. Blier, M.
Mendelsohn, J.J. Mann, D. Brunner, R. Hen, Serotonin receptor 1Aknock-out: an animal model of anxiety-related disorder, Proc. Natl.
Ž .Acad. Sci. U. S. A. 95 1998 14476–14481.w x37 M. Rattray, S. Baldessari, M. Gobbi, T. Mennini, R. Samanin, C.
Bendotti, p-Chlorophenylalanine changes serotonin transportermRNA levels and expression of the gene product, J. Neurochem. 67Ž .1996 463–472.
w x38 V.H. Routh, D.M. Murakami, J.S. Stern, C.A. Fuller, B.A. Horwitz,Neuronal activity in hypothalamic nuclei of obese and lean Zucker
Ž .rats, Int. J. Obes. 14 1990 879–891.w x39 V.H. Routh, J.S. Stern, B.A. Horwitz, Serotonergic activity is
depressed in the ventromedial hypothalamic nucleus of 12-day-oldŽ .obese Zucker rats, Am. J. Physol. 267 1994 R712–R719.
w x40 J.M. Saavedra, M. Palkovits, M.J. Brownstein, J. Axelrod, Serotonindistribution in the nuclei of the rat hypothalamus and preoptic
Ž .region, Brain Res. 77 1974 157–164.w x41 P.E. Sawchenko, L.W. Swanson, H.W. Steinbusch, A.A. Verhofstad,
The distribution and cells of origin of serotonergic inputs to paraven-Ž .tricular and supraoptic nuclei of the rat, Brain Res. 277 1983
355–360.w x42 J. De Schepper, X. Zhou, B. Velkeniers, E. Hooghe-Peters, L.
Vanhaelst, 5-Hydroxytryptophan-stimulated prolactin levels in cafe-teria diet fed rats: an in vivo evaluation of the central serotonergic
Ž .tonus, J. Endocrinol. Invest. 17 1994 783–786.w x43 T. Sharp, S. Hjorth, Application of brain microdialysis to study the
pharmacology of the 5-HT autoreceptor, J. Neurosci. Methods 341AŽ .1990 83–90.
w x44 H. Shimizu, Alteration in hypothalamic monoamine metabolism ofŽ .freely moving diabetic rat, Neurosci. Lett. 131 1991 225–227.
w x45 S.E. Specter, J.S. Stern, B.A. Horwitz, Hypothalamic monoaminer-gic activity in obese Zucker rats in response to acute and chronic
Ž .distary stimuli, Am. J. Physiol. 270 1996 E677–E688.w x46 L.H. Tecott, L.M. Sun, S.F. Akana, A.M. Strack, D.H. Lowenstein,
M.F. Dallman, D. Julius, Eating disorder and epilepsy in miceŽ .lacking 5-HT serotonin receptors, Nature 374 1995 542–546.2C
w x47 H. Towbin, T. Staehelin, J. Gordon, Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedure
Ž .and some applications, Proc. Natl. Acad. Sci. U. S. A. 76 19794350–4354.
w x48 U. Ungerstedt, Stereotaxic mapping of the monoamine pathways inŽ .the rat brain, Acta Physiol. Scand. 82 1971 1–48.
w x49 Y. Watanabe, R.R. Sakai, B.S. McEwen, S. Mendelson, Stress andantidepressants effects on hippocampus and cortical 5-HT and1A
5-HT receptors and transporter sites for serotonin, Brain Res. 6152Ž .1993 87–94.