in vitro early gametogenesis
TRANSCRIPT

Molecular Human Reproduction, Vol.24, No.5 pp. 233–243, 2018
Advanced Access publication on March 8, 2018 doi:10.1093/molehr/gay011
ORIGINAL ARTICLE
Characterization of migratoryprimordial germ cells in the aorta-gonad-mesonephros of a 4.5-week-oldhuman embryo: a toolbox to evaluatein vitro early gametogenesisMaria Gomes Fernandes1,†, Monika Bialecka1,†, Daniela C.F. Salvatori2,*, and Susana M. Chuva de Sousa Lopes 1,3,*
1Department of Anatomy and Embryology, Leiden University Medical Center, Leiden 2333-ZC, The Netherlands 2Central LaboratoryAnimal Facility, Leiden University Medical Center, Leiden 2333-ZC, The Netherlands 3Department for Reproductive Medicine, GhentUniversity Hospital, Ghent 9000, Belgium
*Correspondence address. Department of Anatomy and Embryology, Leiden University Medical Center, Leiden 2333-ZC, The Netherlands.Tel: +31-71-526-9350; Fax: +31-71-526-8289; E-mail: [email protected] orcid.org/0000-0003-3866-2803
Submitted on July 16, 2017; resubmitted on February 26, 2018; editorial decision on March 5, 2018; accepted on March 7, 2018
STUDY QUESTION: Which set of antibodies can be used to identify migratory and early post-migratory human primordial germ cells(hPGCs)?
STUDY FINDING: We validated the specificity of 33 antibodies for 31 markers, including POU5F1, NANOG, PRDM1 and TFAP2C asspecific markers of hPGCs at 4.5 weeks of development of Carnegie stage (CS12–13), whereas KIT and SOX17 also marked the intra-aortichematopoietic stem cell cluster in the aorta-gonad-mesonephros (AGM).
WHAT IS KNOWN ALREADY: The dynamics of gene expression during germ cell development in mice is well characterized and thisknowledge has proved crucial to allow the development of protocols for the in vitro derivation of functional gametes. Although there is a greatinterest in generating human gametes in vitro, it is still unclear which markers are expressed during the early stages of hPGC development andmany studies use markers described in mouse to benchmark differentiation of human PGC-like cells (hPGCLCs). Early post-implantationdevelopment differs significantly between mice and humans, and so some germ cells markers, including SOX2, SOX17, IFITM3 and ITGA6may not identify mPGCs and hPGCs equally well.
STUDY DESIGN, SIZE, DURATION: This immunofluorescence study investigated the expression of putative hPGC markers in the cau-dal part of a single human embryo at 4.5 weeks of development.
PARTICIPANTS/MATERIALS, SETTING, METHODS: We have investigated by immunofluorescence the expression of a set of 33antibodies for 31 markers, including pluripotency, germ cell, adhesion, migration, surface, mesenchymal and epigenetic markers on paraffinsections of the caudal part, including the AGM region, of a single human embryo (CS12–13). The human material used was anonymouslydonated with informed consent from elective abortions without medical indication.
MAIN RESULTS AND THE ROLE OF CHANCE: We observed germ cell specific expression of NANOG, TFAP2C and PRDM1 inPOU5F1+ hPGCs in the AGM. The epigenetic markers H3K27me3 and 5mC were sufficient to distinguish hPGCs from the surrounding som-atic cells. Some mPGC-markers were not detected in hPGCs, but marked other tissues; whereas other markers, such as ALPL, SOX17, KIT,TUBB3, ITGA6 marked both POU5F1+ hPGCs and other cells in the AGM. We used a combination of multiple markers, immunostaining dif-ferent cellular compartments when feasible, to decrease the chance of misidentifying hPGCs.
LARGE SCALE DATA: Non-applicable.
†These authors contributed equally.
© The Author(s) 2018. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology.This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/), which permitsnon-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact [email protected]
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

LIMITATIONS REASONS FOR CAUTION: Material to study early human development is unique and very rare thus restricting the sam-ple size. We have used a combination of antibodies limited by the number of paraffin sections available.
WIDER IMPLICATIONS OF THE FINDINGS: Most of our knowledge on early gametogenesis has been obtained from model organ-isms such as mice and is extrapolated to humans. However, since there is a dedicated effort to produce human artificial gametes in vitro, it isof great importance to determine the expression and specificity of human-specific germ cell markers. We provide a systematic analysis of theexpression of 31 different markers in paraffin sections of a CS12–13 embryo. Our results will help to set up a toolbox of markers to evaluateprotocols to induce hPGCLCs in vitro.
STUDY FUNDING AND COMPETING INTEREST(S): M.G.F. was funded by Fundação para a Ciência e Tecnologia (FCT) [SFRH/BD/78689/2011] and S.M.C.S.L. was funded by the Interuniversity Attraction Poles (IAP, P7/07) and the European Research CouncilConsolidator (ERC-CoG-725722-OVOGROWTH). The authors declare no conflict of interest.
Key words: human / primordial germ cells / migration / antibodies / expression / pluripotency / epigenetics / surface markers / aorta-gonad-mesonephros
IntroductionThe dynamics of gene expression during specification and furtherdevelopment of primordial germ cells (PGCs) in mouse is well charac-terized (Saitou and Yamaji, 2012; Bertocchini and Chuva de SousaLopes, 2016; Saitou and Miyauchi, 2016; Tang et al., 2016).Consequently, markers (including antibodies) to identify and facilitateFACS-sorting of differentiated PGC-like cells (PGCLCs) from mousepluripotent stem cells (PSCs) in vitro, as well as to evaluate the effi-ciency of in vitro differentiation protocols, are well known and reliablyused. This useful toolbox of antibodies has been crucial for the successof recent protocols using mouse PSCs to recapitulate gametogenesisin vitro (Hayashi et al., 2011, 2012; Hikabe et al., 2016). In humans, thedynamics of gene expression is less well studied and in fact it is stillunclear when PGCs are specified (Bertocchini and Chuva de SousaLopes, 2016; Tang et al., 2016). This lack of knowledge is hamperingthe efficient benchmark of differentiation protocols recapitulating gam-etogenesis in vitro using human PSCs (Clark et al., 2004; Bucay et al.,2009; Kee et al., 2009; Gkountela et al., 2013; Irie et al., 2015; Tilgneret al., 2008; Sasaki et al., 2015; Sugawa et al., 2015).
In mouse, PGC precursors (pPGCs) express PRDM1 (or BLIMP1),TFAP2C (or AP2gamma) and PRDM14 (Bertocchini and Chuva deSousa Lopes, 2016; Tang et al., 2016) to suppress the somatic programand become lineage restricted as PGCs around embryonic day (E)7.2(Tam and Zhou, 1996; de Sousa Lopes et al., 2007). From this stage onand until they undergo meiosis, mPGCs also express key genes asso-ciated with pluripotency such as POU5F1 (or OCT4) (Kehler et al.,2004), NANOG (Chambers et al., 2007), SOX2 (Campolo et al.,2013), DPPA3 (or STELLA) (Payer et al., 2003), SALL4 (Yamaguchiet al., 2015) and ALPL (or TNAP) (MacGregor et al., 1995).
Mouse transgenic PSCs, such as Blimp1::mvenus and Stella::ecfp(Hikabe et al., 2016; Zhou et al., 2016) have proved useful to optimizeprotocols for the differentiation of mPSCs to mPGCLCs. Hence,mPGCLCs FACS-sorted for SSEA1+ and ITGB3+ and subsequentlyco-cultured with either E12.5 female gonads or with newborn testis(a necessary step to induce meiosis), were able to undergo respect-ively oogenesis or spermatogenesis ex vivo, generating functionalgametes (Hikabe et al., 2016; Zhou et al., 2016). Human transgenicPSCs for Blimp1::tdtomato, Tfap2c::egfp (Sasaki et al., 2015) andNanos3::mcherry (Irie et al., 2015) have facilitated differentiation to
human PGCLCs (hPGCLCs), but these do not upregulate late PGCmarkers or undergo meiosis.
Much of what is known regarding human early gametogenesis, in par-ticular PGC specification, relies heavily on extrapolation from studies inmouse (Bertocchini and Chuva de Sousa Lopes, 2016; Tang et al.,2016), monkey (Sasaki et al., 2016) and pig (du Puy et al., 2011;Kobayashi et al., 2017). Although several studies have tested antibody-markers of pluripotency and germ cells in histological sections of humanfoetal gonads (Gaskell et al., 2004; Pauls et al., 2006; Anderson et al.,2007; Gkountela et al., 2013; Heeren et al., 2015, 2016; Kerr et al.,2008a, b; Rajpert-De Meyts et al., 2004), only few report the analysis ofmigratory hPGCs (Mollgard et al., 2010; Mamsen et al., 2012).Moreover, studies on hPGCs have highlighted differences in markerexpression and hence gametogenesis between mice and humans.
Despite recent advances in hPGC single-cell transcriptomics (Guoet al., 2015; Li et al., 2017) and differentiation protocols from hPSCs tohPGCLCs, many of the markers that are currently used to access differ-entiation and to sort pure populations of hPGCs/hPGCLCs are notuniquely expressed in hPGCs and their expression has not been valid-ated in earlier stages of human development. In this study, we evaluatedand validated for the expression of several pluripotency- and PGC-associated markers/antibodies in migratory and early colonizing hPGCsin one single human embryo of Carnegie stage 12–13 (CS12–13). Ourresults showed the specificity of a panel of 31 markers to distinguishhPGCs, crucial to evaluate hPGCLC differentiation in vitro.
Materials andMethods
Ethical approval for use of human foetaltissueAll procedures conformed to the Declaration of Helsinki for MedicalResearch involving Human Subjects and were approved by the MedicalEthical Committee of the Leiden University Medical Center (P08.087).The embryo was donated for research with informed consent from elect-ive abortions without medical indication.
Collection and sex genotyping of humanfoetal materialThe developmental age of the embryo was determined by ultrasonog-raphy. The embryo was isolated in cold 0.9% NaCl (Fresenius Kabi, Zeist,
234 Gomes Fernandes et al.
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

the Netherlands) and fixed in 4% paraformaldehyde (PFA) (Merck,Darmstadt, Germany) overnight (o/n) at 4°C, washed 3× in phosphate-buffered saline without Ca++ and Mg++ (PBS0) and stored in 70% etha-nol at 4°C.
The sex was determined by genomic PCR for Amelogenin (AMELX/AMELY), that distinguishes the X and Y chromosomes by amplicon size(977 bp and 790 bp, respectively) as described (Heeren et al., 2015). Theprimers used were: FW 5′-CTG ATG GTT GGC CTC AAG CCT GTG-3′ and RV 5′-TAA AGA GAT TCA TTA ACT TGA CTG-3′; the PCR pro-gramme used was 5 min 95°C, 34× (1 min 95°C, 30 s 60°C, 2 min 72°C),10 min 72°C, and the PCR products were run on a 1.5% agarose gel.
Immunofluorescence in paraffin sectionsTheW4.5 embryo was embedded in paraffin using a Shandon Excelsior tis-sue processor (Thermo Scientific, Altrincham, UK) and sectioned (5 μm)using a RM2065 microtome (Leica Instruments GmbH, Wetzlar,Germany) onto StarFrost slides (Waldemar Knittel, Braunschweig,Germany). Human foetal material (W8–9 gonad, W9 mesonephros,W16–18 kidney, W19 adrenal, W19 placenta and W15 colon) were iso-lated, embedded in paraffin and processed for immunofluorescence.Immunofluorescence was performed as described (Heeren et al., 2015).Briefly, paraffin sections were deparaffinised in xylene, rehydrated throughan ethanol series and finally water, followed by antigen retrieval in 0.01 Mcitric buffer (pH 6.0) for 12 min at 98°C on a microwave (TissueWave 2,Thermo Scientific) and allowed to cool down. After being rinsed in PBS0,sections were treated for 1 h at room temperature (RT) with blockingsolution (1% bovine serum albumin (BSA, Sigma-Aldrich, St. Louis, USA),0.05% Tween-20 (Merck-Schuchardt, Hohenbrunn, Germany) in PBS0).Thereafter, sections were incubated with primary antibodies diluted inblocking solution overnight at 4°C, washed three times for 20 min at RTwith PBS0 and incubated with the respective secondary antibodies for 2 hat RT. Primary antibodies and dilutions used, as well as used matching iso-types used as negative controls, are listed in Supplementary Table S1.Secondary antibodies and dilutions used are listed in SupplementaryTable S2. Results from the isotype controls (negative controls) are pre-sented in Supplementary Fig. S1.
Immunofluorescence of whole mountsHuman W8–9 gonads were cut transversally in several smaller pieces(12–15 pieces) using a scalpel (Swann Morton, Sheffield, England). Thesesmall pieces were permeabilized in 0.2% Triton-X100 (Merck, Darmstadt,Germany) in PBS0 for 20 min at RT and blocked in a solution of 1% BSA(Life Technologies, Carlsbad, USA) and 10% foetal calf serum (LifeTechnologies, Carlsbad, USA) in PBS0 for 1 h at RT. The gonadal pieceswere then incubated with primary antibodies (Supplementary Table S1)diluted in 1% BSA in PBS-T (0.1% Tween-20 (Merck, Darmstadt,Germany) in PBS0) overnight at 4°C, washed twice with PBS-T, incubatedwith secondary antibodies (Supplementary Table S2) diluted in 1% BSA/PBS-T o/n at 4°C, washed twice with PBS-T and once with MilliQ water,and counterstained with DAPI (Life Technologies, Carlsbad, USA).Samples were mounted on StarFrost slides using ProlonGold.
Teratoma assayParaffin sections of teratomas were a gift from D. Salvatori. The formationof teratoma was ethically approved by the Animal Ethical Committee ofthe Leiden University Medical Center (DEC 13 165) and previouslydescribed (Bouma et al., 2017). Briefly, adult male mice (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ, Charles River) were subcutaneously injectedwith 2102Ep cells (1 × 106 cells per injection) in the flank region. Thetumour growth was monitored periodically and, when reaching a volume
of 2 cm3, was isolated, embedded in paraffin blocks and used for immuno-fluorescence and as described above. Antibodies (and dilutions) used arelisted in Supplementary Tables S1 and S2.
ImagingBright field images of the embryo were made using a Tablet-PC PETW1010 I0NL (Peaq, Oberursel, Germany). Fluorescence images weremade on an inverted Leica TCS SP5 confocal microscope (LeicaMicrosystems, Wetzlar, Germany) using the Leica Application SuiteAdvanced Fluorescence software (LAS AF, Leica). Different channels wereacquired sequentially and the merged imaged was generated afterwards.Colour settings were performed in Fiji (Schindelin et al., 2012) andfigures were assembled in Adobe Photoshop CC (Adobe Systems, SanJose, CA, USA) and Adobe Illustrator CC (Adobe Systems, San Jose, CA,USA).
Results
Morphological characteristic of the humanembryo analysedIn 1948, Emil Witschi performed a detailed histological analysis of 23serially sectioned embryos (ranging from 3.5 to 8 mm) from theCarnegie collection, and generated a graphical reconstruction of themigratory trajectory of the hPGCs based on morphology (Witschi,1948). The quantification of the hPGCs in a 4.2 mm embryo revealedthat, at that stage, most hPGCs had left the gut endoderm and 71%were migrating thought the mesentery and rounding the coelomicangle heading for either the left or right gonadal primordium (Witschi,1948).
We have analysed a rare (and almost intact) embryo of 4 weeks and5 days gestation (Fig. 1A), corresponding to CS12–13 (Hill, 2017). Wecounted 30 somites, but the most posterior part of the embryo wasmissing and therefore, the somite number was likely higher. Sectionsthrough the caudal part of the embryo, containing the aorta-gonad-mesonephros (AGM) region, revealed the gut, dorsal mesenterium,mesonephros, dorsal aorta, somites, notochord, neural tube and sur-face ectoderm (Fig. 1B). These are important landmarks to evaluatethe specificity of the antibodies tested. The thickening of the gonadalprimordia had not formed yet (Fig. 1B), suggesting that most hPGCswere still actively migrating.
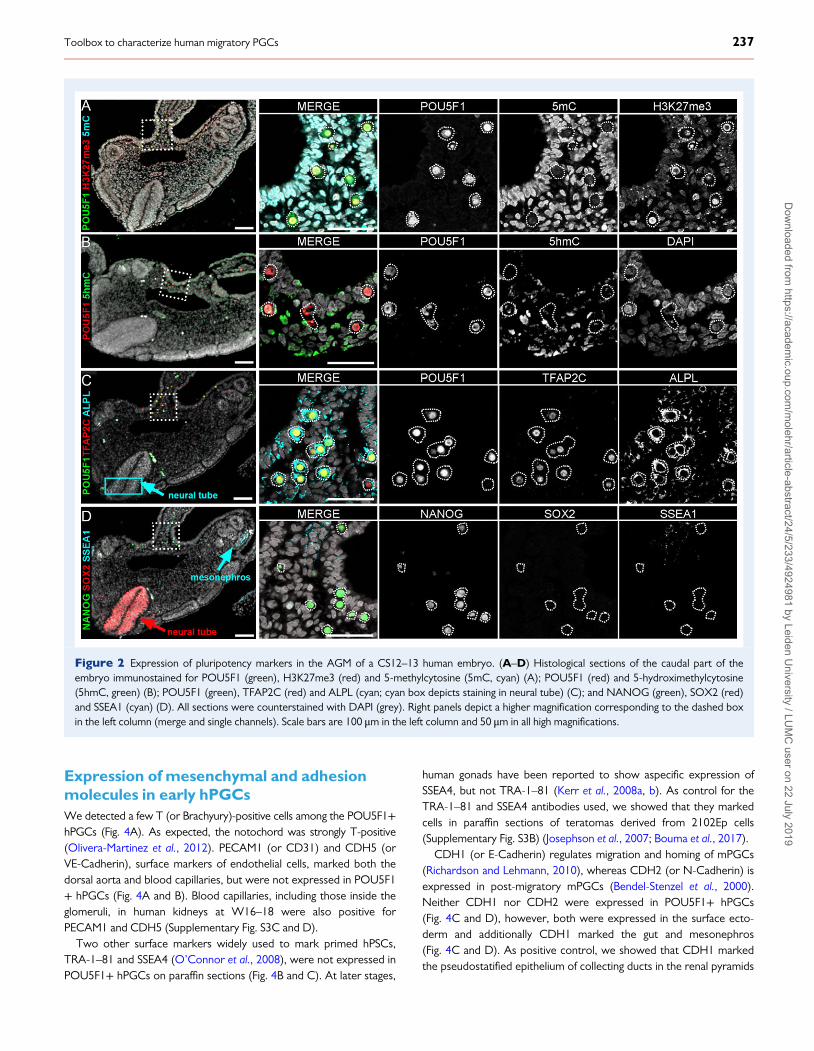
Early hPGCs showed a distinct epigeneticstate from the somatic compartmentWe used POU5F1 (or OCT4) to mark hPGCs unambiguously andobserved hPGCs migrating through the dorsal mesentery and roundingthe coelomic angle to reach the gonadal primordia (Fig. 2A). POU5F1showed strong nuclear localization but was also visible in thecytoplasm.
The sex of the embryo (XX) was confirmed by immunostaining forhistone 3 lysine 27 trimethylation (H3K27me3), as the characteristicperinuclear accumulation of H3K27me3, corresponding to the silentchromosome X in somatic cells, was visible (Geens and Chuva DeSousa Lopes, 2017). By contrast, in hPGCs H3K27me3 coated theentire nuclear envelope and this was in fact sufficient to distinguishhPGCs from the surrounding somatic cells (Fig. 2A, SupplementaryFig. S2A). As described in late hPGCs (Gkountela et al., 2013), early
235Toolbox to characterize human migratory PGCs
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

hPGCs showed much lower levels of global DNA methylation, markedby anti 5-methylcytosine (5mC), than the surrounding somatic cells(Fig. 2A, Supplementary Fig. S2A), another striking feature distinguishingearly hPGCs and somatic cells. The levels of 5-hydroxymethylcytosine(5hmC), generated by oxidation of 5mC (Ficz et al., 2011; Hackett et al.,2013) were also evaluated. Both early hPGCs and neighbouring somaticcells exhibited perinuclear foci of 5hmC (Fig. 2B, SupplementaryFig. S2B).
POU5F1, NANOG and TFAP2Cmarkmigratory and early colonizing hPGCsMost migratory and early colonizing POU5F1+ hPGCs were also posi-tive for other pluripotency markers, including NANOG, ALPL (orTNAP) and TFAP2C (or AP2γ) (Fig. 2C and D). ALPL was also pre-sent, albeit at lower level, in the neural tube (Fig. 2C). Interestingly,this is also observed in mouse embryos of comparable developmentalstage (Kwong and Tam, 1984).
SSEA1 has been one of the markers (together with ITGB3) used toFACS-sort differentiated mPGCLCs from mPSCs (Hikabe et al., 2016;Zhou et al., 2016), therefore it was important to test its specificity inhPGCs. In agreement with Liu and colleagues (Liu et al., 2004), wewere unable to detect SSEA1 in paraffin sections of early hPGCs, butobserved expression in parts of the mesonephros (Fig. 2D,Supplementary Fig. S3A). This contrasted with studies that showedSSEA1 in paraffin sections of human gonads from later developmentalstages (Kerr et al., 2008a, b; Park et al., 2009).
We observed that the pluripotency marker SOX2 was absent fromearly hPGCs (Fig. 2D), as described for later stage hPGCs (Perrettet al., 2008). However, prominent SOX2 staining marked the neuraltube (Fig. 2D), confirming previous observations in human CS12 andCS16 (Olivera-Martinez et al., 2012). Moreover, abundant SOX2 wasobserved in paraffin sections of teratomas derived from the embryonalcarcinoma line 2102Ep (Supplementary Fig. S3B).
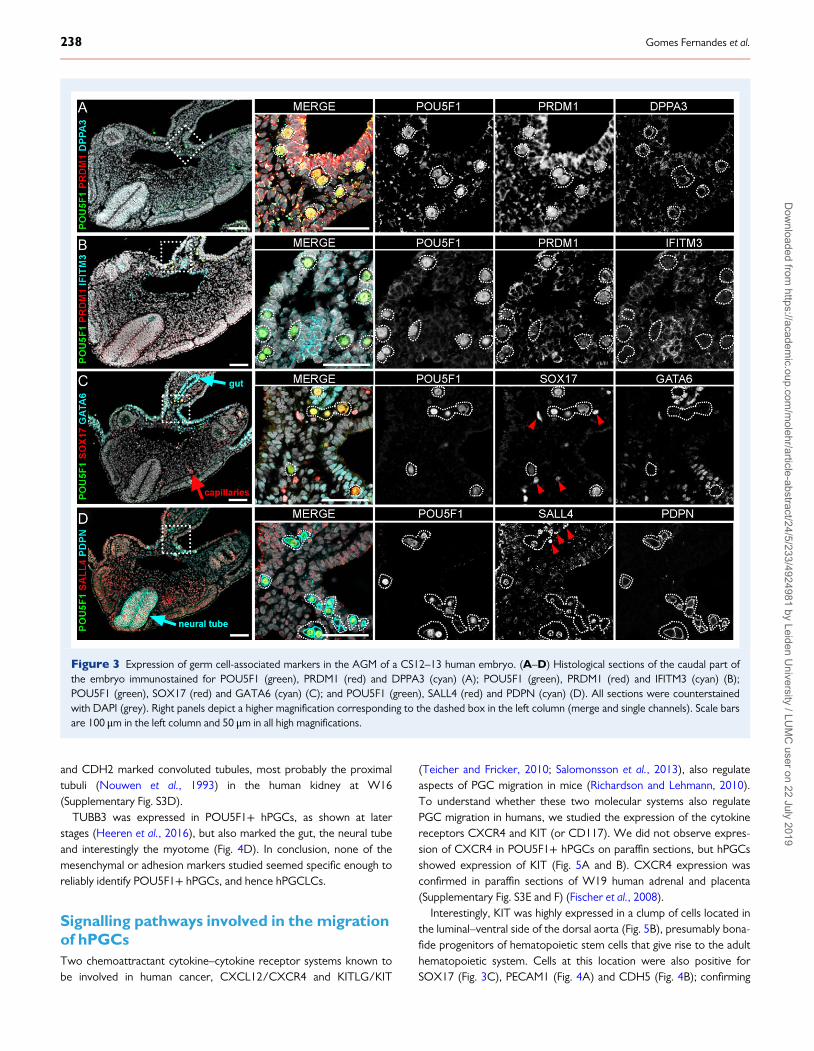
Expression of mPGC-markers PRDM1,DPPA3 and IFITM3 in early hPGCsNext, we investigated the expression of genes known to mark earlymPGCs in mice (Saitou et al., 2002; Ohinata et al., 2005) and regularlyused to access in vitro differentiation to hPGCLCs (Clark et al., 2004;Bucay et al., 2009; Kee et al., 2009; Gkountela et al., 2013; Irie et al.,2015; Tilgner et al., 2008; Sasaki et al., 2015; Sugawa et al., 2015).Specific nuclear PRDM1 was sufficient to identify POUF51+ hPGCs(Fig. 3A and B). The antibody used against DPPA3 showed higherexpression in POUF51+ hPGCs, but the expression was restricted tothe cytoplasm instead of being nuclear (Fig. 3A). IFITM3 showed lowlevels of expression overall in the embryo and did not mark hPGCs spe-cifically (Fig. 3B). We further analysed the expression of DPPA3 andIFITM3 in OCT4+ hPGCs in older human embryos [week (W)8–9 ofdevelopment] in both paraffin sections and whole mount and confirmedthe cytoplasmic staining of DPPA3 in hPGCs and the aspecific staining ofIFITM3 (Supplementary Fig. S4). Thus, we suggest caution when usingDPPA3 and IFITM3 antibodies to identify hPGCLCs.
Specific expression of SOX17, SALL4 andPDPN in early hPGCsRecently, SOX17 has been shown to be expressed in hPGCs and to be acritical determinant during in vitro differentiation to both hPGCLCs fate(Irie et al., 2015), endodermal fate (Wang et al., 2011) and endothelial fate(Zhang et al., 2017). We showed that SOX17 was expressed almostexclusively in POU5F1+ hPGCs and endothelial cells including the dorsalaorta, but unexpectedly not in the endoderm-derived GATA6+ gut(Fig. 3C). In addition, we investigated the expression pattern of SALL4, anovel determinant of mPGCs (Yamaguchi et al., 2015) and showed thatSALL4 was expressed almost exclusively in POU5F1+ hPGCs, (Fig. 3D).Furthermore, we report the specific expression of the surface markerPDPN in POU5F1+ hPGCs, but also in the neural tube (Fig. 3D).
Figure 1 Morphological characteristics of a CS12–13 human embryo. (A) Bright field image of a human embryo with 4 weeks and 5 days of develop-ment, corresponding to Carnegie stage (CS)12–13. The caudal part of the embryo, containing the aorta-gonad-mesonephros (AGM) region was sec-tioned (black square shows the orientation). (B) Histological section of the embryo with several anatomic landmarks identified. Nuclei are stained withDAPI (grey). Scale bars are 1 mm in (A) and 50 μm in (B).
236 Gomes Fernandes et al.
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019
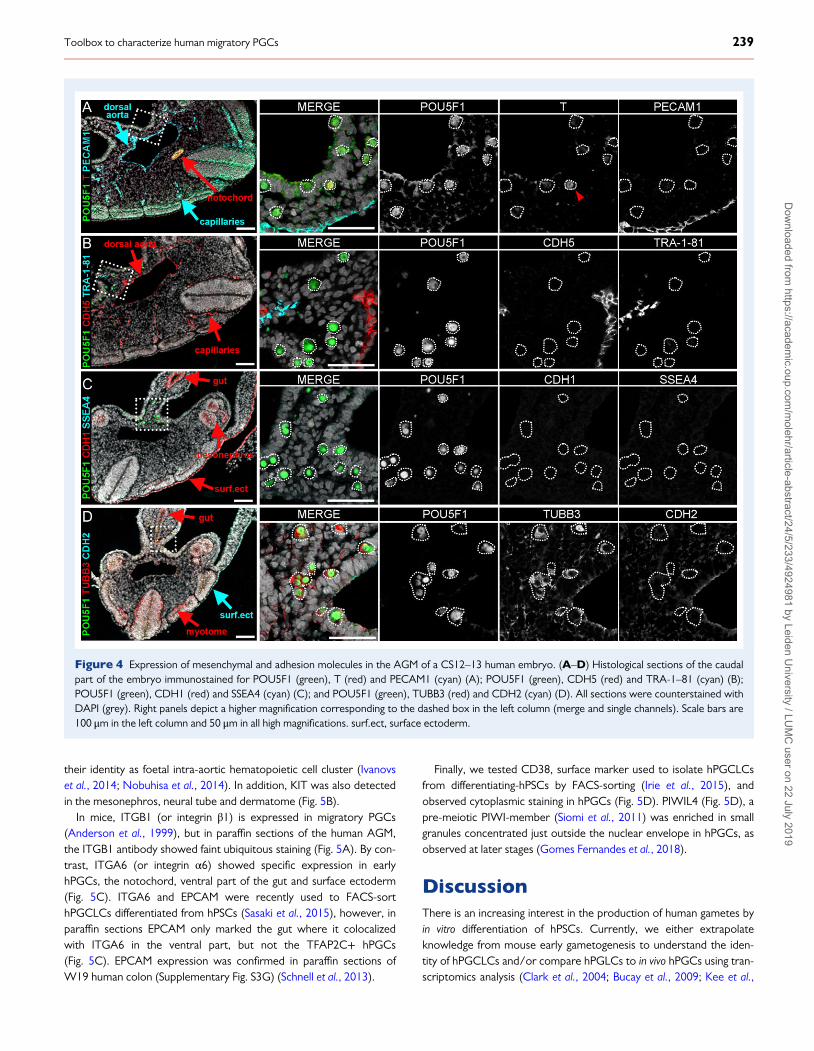
Expression of mesenchymal and adhesionmolecules in early hPGCsWe detected a few T (or Brachyury)-positive cells among the POU5F1+hPGCs (Fig. 4A). As expected, the notochord was strongly T-positive(Olivera-Martinez et al., 2012). PECAM1 (or CD31) and CDH5 (orVE-Cadherin), surface markers of endothelial cells, marked both thedorsal aorta and blood capillaries, but were not expressed in POU5F1+ hPGCs (Fig. 4A and B). Blood capillaries, including those inside theglomeruli, in human kidneys at W16–18 were also positive forPECAM1 and CDH5 (Supplementary Fig. S3C and D).
Two other surface markers widely used to mark primed hPSCs,TRA-1–81 and SSEA4 (O’Connor et al., 2008), were not expressed inPOU5F1+ hPGCs on paraffin sections (Fig. 4B and C). At later stages,
human gonads have been reported to show aspecific expression ofSSEA4, but not TRA-1–81 (Kerr et al., 2008a, b). As control for theTRA-1–81 and SSEA4 antibodies used, we showed that they markedcells in paraffin sections of teratomas derived from 2102Ep cells(Supplementary Fig. S3B) (Josephson et al., 2007; Bouma et al., 2017).
CDH1 (or E-Cadherin) regulates migration and homing of mPGCs(Richardson and Lehmann, 2010), whereas CDH2 (or N-Cadherin) isexpressed in post-migratory mPGCs (Bendel-Stenzel et al., 2000).Neither CDH1 nor CDH2 were expressed in POU5F1+ hPGCs(Fig. 4C and D), however, both were expressed in the surface ecto-derm and additionally CDH1 marked the gut and mesonephros(Fig. 4C and D). As positive control, we showed that CDH1 markedthe pseudostatified epithelium of collecting ducts in the renal pyramids
Figure 2 Expression of pluripotency markers in the AGM of a CS12–13 human embryo. (A–D) Histological sections of the caudal part of theembryo immunostained for POU5F1 (green), H3K27me3 (red) and 5-methylcytosine (5mC, cyan) (A); POU5F1 (red) and 5-hydroximethylcytosine(5hmC, green) (B); POU5F1 (green), TFAP2C (red) and ALPL (cyan; cyan box depicts staining in neural tube) (C); and NANOG (green), SOX2 (red)and SSEA1 (cyan) (D). All sections were counterstained with DAPI (grey). Right panels depict a higher magnification corresponding to the dashed boxin the left column (merge and single channels). Scale bars are 100 μm in the left column and 50 μm in all high magnifications.
237Toolbox to characterize human migratory PGCs
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019
and CDH2 marked convoluted tubules, most probably the proximaltubuli (Nouwen et al., 1993) in the human kidney at W16(Supplementary Fig. S3D).
TUBB3 was expressed in POU5F1+ hPGCs, as shown at laterstages (Heeren et al., 2016), but also marked the gut, the neural tubeand interestingly the myotome (Fig. 4D). In conclusion, none of themesenchymal or adhesion markers studied seemed specific enough toreliably identify POU5F1+ hPGCs, and hence hPGCLCs.
Signalling pathways involved in the migrationof hPGCsTwo chemoattractant cytokine–cytokine receptor systems known tobe involved in human cancer, CXCL12/CXCR4 and KITLG/KIT
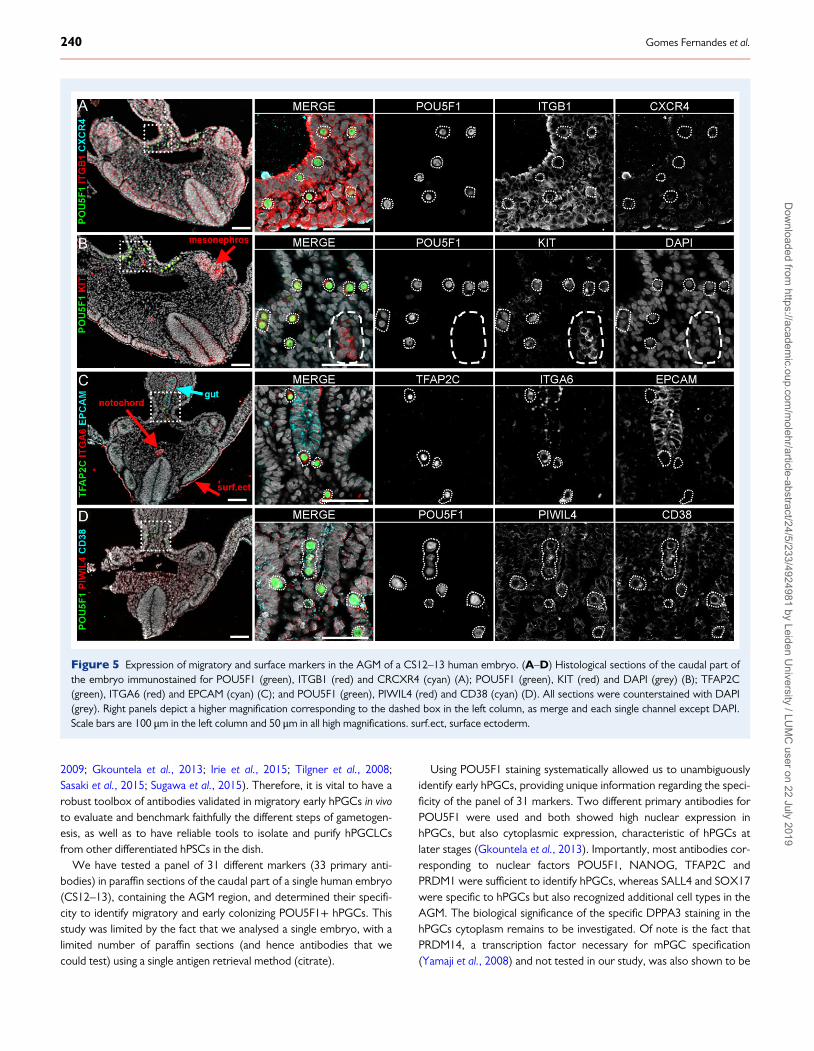
(Teicher and Fricker, 2010; Salomonsson et al., 2013), also regulateaspects of PGC migration in mice (Richardson and Lehmann, 2010).To understand whether these two molecular systems also regulatePGC migration in humans, we studied the expression of the cytokinereceptors CXCR4 and KIT (or CD117). We did not observe expres-sion of CXCR4 in POU5F1+ hPGCs on paraffin sections, but hPGCsshowed expression of KIT (Fig. 5A and B). CXCR4 expression wasconfirmed in paraffin sections of W19 human adrenal and placenta(Supplementary Fig. S3E and F) (Fischer et al., 2008).
Interestingly, KIT was highly expressed in a clump of cells located inthe luminal–ventral side of the dorsal aorta (Fig. 5B), presumably bona-fide progenitors of hematopoietic stem cells that give rise to the adulthematopoietic system. Cells at this location were also positive forSOX17 (Fig. 3C), PECAM1 (Fig. 4A) and CDH5 (Fig. 4B); confirming
Figure 3 Expression of germ cell-associated markers in the AGM of a CS12–13 human embryo. (A–D) Histological sections of the caudal part ofthe embryo immunostained for POU5F1 (green), PRDM1 (red) and DPPA3 (cyan) (A); POU5F1 (green), PRDM1 (red) and IFITM3 (cyan) (B);POU5F1 (green), SOX17 (red) and GATA6 (cyan) (C); and POU5F1 (green), SALL4 (red) and PDPN (cyan) (D). All sections were counterstainedwith DAPI (grey). Right panels depict a higher magnification corresponding to the dashed box in the left column (merge and single channels). Scale barsare 100 μm in the left column and 50 μm in all high magnifications.
238 Gomes Fernandes et al.
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019
their identity as foetal intra-aortic hematopoietic cell cluster (Ivanovset al., 2014; Nobuhisa et al., 2014). In addition, KIT was also detectedin the mesonephros, neural tube and dermatome (Fig. 5B).
In mice, ITGB1 (or integrin β1) is expressed in migratory PGCs(Anderson et al., 1999), but in paraffin sections of the human AGM,the ITGB1 antibody showed faint ubiquitous staining (Fig. 5A). By con-trast, ITGA6 (or integrin α6) showed specific expression in earlyhPGCs, the notochord, ventral part of the gut and surface ectoderm(Fig. 5C). ITGA6 and EPCAM were recently used to FACS-sorthPGCLCs differentiated from hPSCs (Sasaki et al., 2015), however, inparaffin sections EPCAM only marked the gut where it colocalizedwith ITGA6 in the ventral part, but not the TFAP2C+ hPGCs(Fig. 5C). EPCAM expression was confirmed in paraffin sections ofW19 human colon (Supplementary Fig. S3G) (Schnell et al., 2013).
Finally, we tested CD38, surface marker used to isolate hPGCLCsfrom differentiating-hPSCs by FACS-sorting (Irie et al., 2015), andobserved cytoplasmic staining in hPGCs (Fig. 5D). PIWIL4 (Fig. 5D), apre-meiotic PIWI-member (Siomi et al., 2011) was enriched in smallgranules concentrated just outside the nuclear envelope in hPGCs, asobserved at later stages (Gomes Fernandes et al., 2018).
DiscussionThere is an increasing interest in the production of human gametes byin vitro differentiation of hPSCs. Currently, we either extrapolateknowledge from mouse early gametogenesis to understand the iden-tity of hPGCLCs and/or compare hPGLCs to in vivo hPGCs using tran-scriptomics analysis (Clark et al., 2004; Bucay et al., 2009; Kee et al.,
Figure 4 Expression of mesenchymal and adhesion molecules in the AGM of a CS12–13 human embryo. (A–D) Histological sections of the caudalpart of the embryo immunostained for POU5F1 (green), T (red) and PECAM1 (cyan) (A); POU5F1 (green), CDH5 (red) and TRA-1–81 (cyan) (B);POU5F1 (green), CDH1 (red) and SSEA4 (cyan) (C); and POU5F1 (green), TUBB3 (red) and CDH2 (cyan) (D). All sections were counterstained withDAPI (grey). Right panels depict a higher magnification corresponding to the dashed box in the left column (merge and single channels). Scale bars are100 μm in the left column and 50 μm in all high magnifications. surf.ect, surface ectoderm.
239Toolbox to characterize human migratory PGCs
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019
2009; Gkountela et al., 2013; Irie et al., 2015; Tilgner et al., 2008;Sasaki et al., 2015; Sugawa et al., 2015). Therefore, it is vital to have arobust toolbox of antibodies validated in migratory early hPGCs in vivoto evaluate and benchmark faithfully the different steps of gametogen-esis, as well as to have reliable tools to isolate and purify hPGCLCsfrom other differentiated hPSCs in the dish.
We have tested a panel of 31 different markers (33 primary anti-bodies) in paraffin sections of the caudal part of a single human embryo(CS12–13), containing the AGM region, and determined their specifi-city to identify migratory and early colonizing POU5F1+ hPGCs. Thisstudy was limited by the fact that we analysed a single embryo, with alimited number of paraffin sections (and hence antibodies that wecould test) using a single antigen retrieval method (citrate).
Using POU5F1 staining systematically allowed us to unambiguouslyidentify early hPGCs, providing unique information regarding the speci-ficity of the panel of 31 markers. Two different primary antibodies forPOU5F1 were used and both showed high nuclear expression inhPGCs, but also cytoplasmic expression, characteristic of hPGCs atlater stages (Gkountela et al., 2013). Importantly, most antibodies cor-responding to nuclear factors POU5F1, NANOG, TFAP2C andPRDM1 were sufficient to identify hPGCs, whereas SALL4 and SOX17were specific to hPGCs but also recognized additional cell types in theAGM. The biological significance of the specific DPPA3 staining in thehPGCs cytoplasm remains to be investigated. Of note is the fact thatPRDM14, a transcription factor necessary for mPGC specification(Yamaji et al., 2008) and not tested in our study, was also shown to be
Figure 5 Expression of migratory and surface markers in the AGM of a CS12–13 human embryo. (A–D) Histological sections of the caudal part ofthe embryo immunostained for POU5F1 (green), ITGB1 (red) and CRCXR4 (cyan) (A); POU5F1 (green), KIT (red) and DAPI (grey) (B); TFAP2C(green), ITGA6 (red) and EPCAM (cyan) (C); and POU5F1 (green), PIWIL4 (red) and CD38 (cyan) (D). All sections were counterstained with DAPI(grey). Right panels depict a higher magnification corresponding to the dashed box in the left column, as merge and each single channel except DAPI.Scale bars are 100 μm in the left column and 50 μm in all high magnifications. surf.ect, surface ectoderm.
240 Gomes Fernandes et al.
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

cytoplasmic in gonadal hPGCs (Irie et al., 2015), suggesting that theDPPA3 staining pattern here observed may be of relevance.
The antibodies for the epigenetic marks H3K27me3 and 5mC (glo-bal DNA methylation) were sufficient to distinguish POU5F1+ hPGCsfrom the surrounding somatic cells. These two marks will be importantto show whether differentiating hPGCLCs are undergoing correctreprogramming (von Meyenn et al., 2016). Interestingly, the localiza-tion of H3K27me3 to the nuclear lamina of migratory hPGCs is similarto that in post-migratory gonadal (E11.5–E13.5) mPGCs (Prokopuket al., 2017) and different from that in migratory (E7.5–E9.5) mPGCs(Chuva de Sousa Lopes et al., 2008). This species-specific difference isin agreement with the different dynamics regarding epigenetic remod-elling observed in mice and humans (Gkountela et al., 2015; Guo et al.,2015; Tang et al., 2016; von Meyenn et al., 2016).
We were unable to detect IFITM3, SSEA1, SSEA4, TRA-1–81,CXCR4, CDH1, CDH2, CHD5, ITGB1 and EPCAM specifically inhPGCs in paraffin sections Nevertheless, SSEA1, CDH1, CDH2,CDH5, ITGB1 and EPCAM were not only expressed in other specificregions of the same paraffin section, but we also showed positive con-trols in paraffin-sections of human different tissues (teratoma, mesone-phros, kidney, placenta, adrenal and colon). This suggested thatmPGCs and hPGCs may respond to difference cues to migrate andhence express different surface markers, highlighting the need for func-tional studies and the validation of in vitro discoveries in the human.
The surface antibodies to detect ALPL, KIT and ITGA6 have beenused successfully to isolate hPGCs (Gkountela et al., 2013; Guo et al.,2015) and/or hPGCLCs (Gkountela et al., 2013; Irie et al., 2015;Sasaki et al., 2015; Sugawa et al., 2015) by FACS. The surface markerPDPN may also be a suitable marker to include when identifyinghPGCs and or hPGCLCs. However, we show here that those surfacemarkers identified POU5F1+/TPAP2C+ hPGCs, but recognizedother cell types in the caudal/AGM region. In the same line, cytoplas-mic TUBB3 marked POU5F1+ hPGCs, but is also expressed in neuralcrest derivatives (Locher et al., 2014; Heeren et al., 2016) and otherprogenitor cell types such as the myotome. Therefore, we stronglysuggest a combinatorial use of markers to unambiguously identifyhPGCs or hPGCLCs.
We observed that several markers, such as KIT (Ivanovs et al.,2014) and SOX17 (Zhang et al., 2017), were expressed by both earlyPOU5F1+ hPGCs and the intra-aortic hematopoietic stem cell cluster(luminal–ventral part of the dorsal aorta). Therefore, using these twomarkers alone may lead to the misidentification (or bulk isolation) ofthese two cell types. Interestingly, although IFITM3 showed no specifi-city for POU5F1+ hPGCs, we did notice an enrichment in the intra-aortic hematopoietic stem cell cluster (Fig. 3B). In mouse, IFITM3 hasnot been described as marker of intra-aortic hematopoietic stem cellclusters, but has been detected in (Runx1+) yolk sac hematopoieticcells (Mikedis and Downs, 2013).
We provide a unique insight in the specificity of a panel of 31 differ-ent markers, including pluripotency, surface and epigenetic markers, toidentify and distinguish early hPGCs (and hPGCLCs) from the sur-rounding somatic cells. We report several striking differences betweenmPGCs and hPGCs and show that (surface) markers tend to reactwith several cell types in the embryo, including the intra-aortic hem-atopoietic stem cells present in the AGM. Transcription factors areusually not solely involved in the specification of a single lineage andcells can share the expression of many markers, hence, a careful and
thoughtful choice of markers is crucial when studying in vitro differenti-ation. Our results provide a toolbox of markers to better evaluateprotocols to induce the formation of hPGCLCs in vitro.
Supplementary dataSupplementary data are available at Molecular Human Reproductiononline
AcknowledgementsWe would like to thank the staff of CASA den Haag and CASALeiden, for all the efforts and availability to collect and provide the foe-tal material, Liesbeth van Iperen for isolating the material and NannanHe for discussions.
Authors’ rolesM.G.F., M.B., D.C.F.S. and S.M.C.d.S.L. designed the study, conductedexperiments, analysed data and wrote the article. All authorsapproved the last version of the article.
FundingFundação para a Ciência e Tecnologia (FCT) [SFRH/BD/78689/2011] to M.G.F., and the Interuniversity Attraction Poles (IAP, P7/07)and the European Research Council Consolidator (ERC-CoG-725722-OVOGROWTH) to S.M.C.S.L.
Conflict of interestThe authors declare no conflict of interest.
ReferencesAnderson R, Fassler R, Georges-Labouesse E, Hynes RO, Bader BL,Kreidberg JA, Schaible K, Heasman J, Wylie C. Mouse primordial germcells lacking beta1 integrins enter the germline but fail to migrate nor-mally to the gonads. Development 1999;126:1655–1664.
Anderson RA, Fulton N, Cowan G, Coutts S, Saunders PT. Conservedand divergent patterns of expression of DAZL, VASA and OCT4 in thegerm cells of the human fetal ovary and testis. BMC Dev Biol 2007;7:136.
Bendel-Stenzel MR, Gomperts M, Anderson R, Heasman J, Wylie C. Therole of cadherins during primordial germ cell migration and early gonadformation in the mouse.Mech Dev 2000;91:143–152.
Bertocchini F, Chuva de Sousa Lopes SM. Germline development inamniotes: a paradigm shift in primordial germ cell specification. BioEssays2016;38:791–800.
Bouma MJ, van Iterson M, Janssen B, Mummery CL, Salvatori DCF, FreundC. Differentiation-defective human induced pluripotent stem cells revealstrengths and limitations of the teratoma assay and in vitro pluripotencyassays. Stem Cell Rep 2017;8:1340–1353.
Bucay N, Yebra M, Cirulli V, Afrikanova I, Kaido T, Hayek A, MontgomeryAM. A novel approach for the derivation of putative primordial germcells and sertoli cells from human embryonic stem cells. Stem Cells 2009;27:68–77.
Campolo F, Gori M, Favaro R, Nicolis S, Pellegrini M, Botti F, Rossi P,Jannini EA, Dolci S. Essential role of Sox2 for the establishment andmaintenance of the germ cell line. Stem Cells 2013;31:1408–1421.
241Toolbox to characterize human migratory PGCs
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

Chambers I, Silva J, Colby D, Nichols J, Nijmeijer B, Robertson M, Vrana J,Jones K, Grotewold L, Smith A. Nanog safeguards pluripotency andmediates germline development. Nature 2007;450:1230–1234.
Chuva de Sousa Lopes SM, Hayashi K, Shovlin TC, Mifsud W, Surani MA,McLaren A. X chromosome activity in mouse XX primordial germ cells.PLoS Genet 2008;4:e30.
Clark AT, Bodnar MS, Fox M, Rodriquez RT, Abeyta MJ, Firpo MT, PeraRA. Spontaneous differentiation of germ cells from human embryonicstem cells in vitro. Hum Mol Genet 2004;13:727–739.
de Sousa Lopes SMC, Hayashi K, Surani MA. Proximal visceral endodermand extraembryonic ectoderm regulate the formation of primordialgerm cell precursors. BMC Dev Biol 2007;7:140.
du Puy L, Lopes SM, Haagsman HP, Roelen BA. Analysis of co-expressionof OCT4, NANOG and SOX2 in pluripotent cells of the porcineembryo, in vivo and in vitro. Theriogenology 2011;75:513–526.
Ficz G, Branco MR, Seisenberger S, Santos F, Krueger F, Hore TA,Marques CJ, Andrews S, Reik W. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation.Nature 2011;473:398–402.
Fischer T, Nagel F, Jacobs S, Stumm R, Schulz S. Reassessment of CXCR4 che-mokine receptor expression in human normal and neoplastic tissues usingthe novel rabbit monoclonal antibody UMB-2. PLoS One 2008;3:e4069.
Gaskell TL, Esnal A, Robinson LL, Anderson RA, Saunders PT.Immunohistochemical profiling of germ cells within the human fetal tes-tis: identification of three subpopulations. Biol Reprod 2004;71:2012–2021.
Geens M, Chuva De Sousa Lopes SM. X chromosome inactivation inhuman pluripotent stem cells as a model for human development: backto the drawing board? Hum Reprod Update 2017;23:520–532.
Gkountela S, Li Z, Vincent JJ, Zhang KX, Chen A, Pellegrini M, Clark AT.The ontogeny of cKIT+ human primordial germ cells proves to be aresource for human germ line reprogramming, imprint erasure andin vitro differentiation. Nat Cell Biol 2013;15:113–122.
Gkountela S, Zhang KX, Shafiq TA, Liao WW, Hargan-Calvopina J, ChenPY, Clark AT. DNA demethylation dynamics in the human prenatalgermline. Cell 2015;161:1425–1436.
Gomes Fernandes M, He N, Wang F, Van Iperen L, Eguizabal C, MatorrasR, Roelen BAJ, Chuva De Sousa Lopes SM. Human-specific subcellularcompartmentalization of P-element induced wimpy testis-like (PIWIL)granules during germ cell development and spermatogenesis. HumReprod 2018;33:258–269.
Guo F, Yan L, Guo H, Li L, Hu B, Zhao Y, Yong J, Hu Y, Wang X, Wei Yet al. The transcriptome and DNA methylome landscapes of humanprimordial germ cells. Cell 2015;161:1437–1452.
Hackett JA, Sengupta R, Zylicz JJ, Murakami K, Lee C, Down TA, SuraniMA. Germline DNA demethylation dynamics and imprint erasurethrough 5-hydroxymethylcytosine. Science 2013;339:448–452.
Hayashi K, Ogushi S, Kurimoto K, Shimamoto S, Ohta H, Saitou M.Offspring from oocytes derived from in vitro primordial germ cell-likecells in mice. Science 2012;338:971–975.
Hayashi K, Ohta H, Kurimoto K, Aramaki S, Saitou M. Reconstitution ofthe mouse germ cell specification pathway in culture by pluripotentstem cells. Cell 2011;146:519–532.
Heeren AM, He N, de Souza AF, Goercharn-Ramlal A, van Iperen L,Roost MS, Gomes Fernandes MM, van der Westerlaken LA, Chuva deSousa Lopes SM. On the development of extragonadal and gonadalhuman germ cells. Biol Open 2016;5:185–194.
Heeren AM, van Iperen L, Klootwijk DB, de Melo Bernardo A, Roost MS,Gomes Fernandes MM, Louwe LA, Hilders CG, Helmerhorst FM, vander Westerlaken LA et al. Development of the follicular basement mem-brane during human gametogenesis and early folliculogenesis. BMC DevBiol 2015;15:4.
Hikabe O, Hamazaki N, Nagamatsu G, Obata Y, Hirao Y, Hamada N,Shimamoto S, Imamura T, Nakashima K, Saitou M et al. Reconstitutionin vitro of the entire cycle of the mouse female germ line. Nature 2016;539:299–303.
Hill MA. Embryology Embryonic Development. 2017, pp. Retrieved July 15,2017, https://embryology.med.unsw.edu.au/embryology/index.php/Embryonic_Development.
Irie N, Weinberger L, Tang WW, Kobayashi T, Viukov S, Manor YS,Dietmann S, Hanna JH, Surani MA. SOX17 is a critical specifier of humanprimordial germ cell fate. Cell 2015;160:253–268.
Ivanovs A, Rybtsov S, Anderson RA, Turner ML, Medvinsky A.Identification of the niche and phenotype of the first human hematopoi-etic stem cells. Stem Cell Rep 2014;2:449–456.
Josephson R, Ording CJ, Liu Y, Shin S, Lakshmipathy U, Toumadje A, LoveB, Chesnut JD, Andrews PW, Rao MS et al. Qualification of embryonalcarcinoma 2102Ep as a reference for human embryonic stem cellresearch. Stem Cells 2007;25:437–446.
Kee K, Angeles VT, Flores M, Nguyen HN, Reijo Pera RA. Human DAZL,DAZ and BOULE genes modulate primordial germ-cell and haploidgamete formation. Nature 2009;462:222–225.
Kehler J, Tolkunova E, Koschorz B, Pesce M, Gentile L, Boiani M, LomeliH, Nagy A, McLaughlin KJ, Scholer HR et al. Oct4 is required for primor-dial germ cell survival. EMBO Rep 2004;5:1078–1083.
Kerr CL, Hill CM, Blumenthal PD, Gearhart JD. Expression of pluripotentstem cell markers in the human fetal ovary. Hum Reprod 2008a;23:589–599.
Kerr CL, Hill CM, Blumenthal PD, Gearhart JD. Expression of pluripotentstem cell markers in the human fetal testis. Stem Cells 2008b;26:412–421.
Kobayashi T, Zhang H, Tang WWC, Irie N, Withey S, Klisch D, Sybirna A,Dietmann S, Contreras DA, Webb R et al. Principles of early humandevelopment and germ cell program from conserved model systems.Nature 2017;546:416–420.
Kwong WH, Tam PP. The pattern of alkaline phosphatase activity in thedeveloping mouse spinal cord. J Embryol Exp Morphol 1984;82:241–251.
Li L, Dong J, Yan L, Yong J, Liu X, Hu Y, Fan X, Wu X, Guo H, Wang Xet al. Single-cell RNA-Seq analysis maps development of human germlinecells and gonadal niche interactions. Cell Stem Cell 2017;20:858–873e854.
Liu S, Liu H, Tang S, Pan Y, Ji K, Ning H, Wang S, Qi Z, Li L.Characterization of stage-specific embryonic antigen-1 expression dur-ing early stages of human embryogenesis. Oncol Rep 2004;12:1251–1256.
Locher H, Frijns JH, Huisman MA, Chuva de Sousa Lopes SM. TUBB3:neuronal marker or melanocyte mimic? Cell Transplant 2014;23:1471–1473.
MacGregor GR, Zambrowicz BP, Soriano P. Tissue non-specific alkalinephosphatase is expressed in both embryonic and extraembryoniclineages during mouse embryogenesis but is not required for migrationof primordial germ cells. Development 1995;121:1487–1496.
Mamsen LS, Brochner CB, Byskov AG, Mollgard K. The migration and lossof human primordial germ stem cells from the hind gut epitheliumtowards the gonadal ridge. Int J Dev Biol 2012;56:771–778.
Mikedis MM, Downs KM. Widespread but tissue-specific patterns ofinterferon-induced transmembrane protein 3 (IFITM3, FRAGILIS, MIL-1) in the mouse gastrula. Gene Expression Patterns 2013;13:225–239.
Mollgard K, Jespersen A, Lutterodt MC, Yding Andersen C, Hoyer PE,Byskov AG. Human primordial germ cells migrate along nerve fibers andSchwann cells from the dorsal hind gut mesentery to the gonadal ridge.Mol Hum Reprod 2010;16:621–631.
Nobuhisa I, Osawa M, Uemura M, Kishikawa Y, Anani M, Harada K,Takagi H, Saito K, Kanai-Azuma M, Kanai Y et al. Sox17-mediated
242 Gomes Fernandes et al.
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019

maintenance of fetal intra-aortic hematopoietic cell clusters.Mol Cell Biol2014;34:1976–1990.
Nouwen EJ, Dauwe S, van der Biest I, De Broe ME. Stage- and segment-specific expression of cell-adhesion molecules N-CAM, A-CAM, and L-CAM in the kidney. Kidney Int 1993;44:147–158.
Ohinata Y, Payer B, O’Carroll D, Ancelin K, Ono Y, Sano M, Barton SC,Obukhanych T, Nussenzweig M, Tarakhovsky A et al. Blimp1 is a criticaldeterminant of the germ cell lineage in mice. Nature 2005;436:207–213.
Olivera-Martinez I, Harada H, Halley PA, Storey KG. Loss of FGF-dependent mesoderm identity and rise of endogenous retinoid signallingdetermine cessation of body axis elongation. PLoS Biol 2012;10:e1001415.
O’Connor MD, Kardel MD, Iosfina I, Youssef D, Lu M, Li MM,Vercauteren S, Nagy A, Eaves CJ. Alkaline phosphatase-positive colonyformation is a sensitive, specific, and quantitative indicator of undifferen-tiated human embryonic stem cells. Stem Cells 2008;26:1109–1116.
Park TS, Galic Z, Conway AE, Lindgren A, van Handel BJ, Magnusson M,Richter L, Teitell MA, Mikkola HK, Lowry WE et al. Derivation of prim-ordial germ cells from human embryonic and induced pluripotent stemcells is significantly improved by coculture with human fetal gonadal cells.Stem Cells 2009;27:783–795.
Pauls K, Schorle H, Jeske W, Brehm R, Steger K, Wernert N, Buttner R,Zhou H. Spatial expression of germ cell markers during maturation ofhuman fetal male gonads: an immunohistochemical study. Hum Reprod2006;21:397–404.
Payer B, Saitou M, Barton SC, Thresher R, Dixon JP, Zahn D, ColledgeWH, Carlton MB, Nakano T, Surani MA. Stella is a maternal effect generequired for normal early development in mice. Curr Biol 2003;13:2110–2117.
Perrett RM, Turnpenny L, Eckert JJ, O’Shea M, Sonne SB, Cameron IT,Wilson DI, Rajpert-De Meyts E, Hanley NA. The early human germ celllineage does not express SOX2 during in vivo development or uponin vitro culture. Biol Reprod 2008;78:852–858.
Prokopuk L, Stringer JM, Hogg K, Elgass KD, Western PS. PRC2 is requiredfor extensive reorganization of H3K27me3 during epigenetic reprogram-ming in mouse fetal germ cells. Epigenetics Chromatin 2017;10:7.
Rajpert-De Meyts E, Hanstein R, Jorgensen N, Graem N, Vogt PH,Skakkebaek NE. Developmental expression of POU5F1 (OCT-3/4) innormal and dysgenetic human gonads. Hum Reprod 2004;19:1338–1344.
Richardson BE, Lehmann R. Mechanisms guiding primordial germ cellmigration: strategies from different organisms. Nat Rev Mol Cell Biol2010;11:37–49.
Saitou M, Barton SC, Surani MA. A molecular programme for the specifica-tion of germ cell fate in mice. Nature 2002;418:293–300.
Saitou M, Miyauchi H. Gametogenesis from pluripotent stem cells. CellStem Cell 2016;18:721–735.
Saitou M, Yamaji M. Primordial germ cells in mice. Cold Spring HarbPerspect Biol 2012;4:a008375.
Salomonsson A, Jonsson M, Isaksson S, Karlsson A, Jonsson P, Gaber A,Bendahl PO, Johansson L, Brunnstrom H, Jirstrom K et al. Histologicalspecificity of alterations and expression of KIT and KITLG in non-smallcell lung carcinoma. Genes Chromosomes Cancer 2013;52:1088–1096.
Sasaki K, Nakamura T, Okamoto I, Yabuta Y, Iwatani C, Tsuchiya H, SeitaY, Nakamura S, Shiraki N, Takakuwa T et al. The germ cell fate of cyno-molgus monkeys is specified in the nascent amnion. Dev Cell 2016;39:169–185.
Sasaki K, Yokobayashi S, Nakamura T, Okamoto I, Yabuta Y, Kurimoto K,Ohta H, Moritoki Y, Iwatani C, Tsuchiya H et al. Robust in vitro induc-tion of human germ cell fate from pluripotent stem cells. Cell Stem Cell2015;17:178–194.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T,Preibisch S, Rueden C, Saalfeld S, Schmid B et al. Fiji: an open-sourceplatform for biological-image analysis. Nat Methods 2012;9:676–682.
Schnell U, Cirulli V, Giepmans BN. EpCAM: structure and function inhealth and disease. Biochim Biophys Acta 2013;1828:1989–2001.
Siomi MC, Sato K, Pezic D, Aravin AA. PIWI-interacting small RNAs: thevanguard of genome defence. Nat Rev Mol Cell Biol 2011;12:246–258.
Sugawa F, Arauzo-Bravo MJ, Yoon J, Kim KP, Aramaki S, Wu G, StehlingM, Psathaki OE, Hubner K, Scholer HR. Human primordial germ cellcommitment in vitro associates with a unique PRDM14 expression pro-file. EMBO J 2015;34:1009–1024.
Tam PP, Zhou SX. The allocation of epiblast cells to ectodermal and germ-line lineages is influenced by the position of the cells in the gastrulatingmouse embryo. Dev Biol 1996;178:124–132.
Tang WWC, Kobayashi T, Irie N, Dietmann S, Surani MA. Specificationand epigenetic programming of the human germ line. Nat Rev Genet2016;17:585–600.
Teicher BA, Fricker SP. CXCL12 (SDF-1)/CXCR4 pathway in cancer. ClinCancer Res 2010;16:2927–2931.
Tilgner K, Atkinson SP, Golebiewska A, Stojkovic M, Lako M, Armstrong L.Isolation of primordial germ cells from differentiating human embryonicstem cells. Stem Cells 2008;26:3075–3085.
von Meyenn F, Berrens RV, Andrews S, Santos F, Collier AJ, Krueger F,Osorno R, Dean W, Rugg-Gunn PJ, Reik W. Comparative principles ofDNA methylation reprogramming during human and mouse in vitroprimordial germ cell specification. Dev Cell 2016;39:104–115.
Wang P, Rodriguez RT, Wang J, Ghodasara A, Kim SK. Targeting SOX17in human embryonic stem cells creates unique strategies for isolatingand analyzing developing endoderm. Cell Stem Cell 2011;8:335–346.
Witschi E. Migration of germ cells of human embryos from the yolk sac tothe primitive gonadal folds. Contrib Embryol Carnegie Inst 1948;209:67–98.
Yamaguchi YL, Tanaka SS, Kumagai M, Fujimoto Y, Terabayashi T, MatsuiY, Nishinakamura R. Sall4 is essential for mouse primordial germ cellspecification by suppressing somatic cell program genes. Stem Cells2015;33:289–300.
Yamaji M, Seki Y, Kurimoto K, Yabuta Y, Yuasa M, Shigeta M, Yamanaka K,Ohinata Y, Saitou M. Critical function of Prdm14 for the establishmentof the germ cell lineage in mice. Nat Genet 2008;40:1016–1022.
Zhang L, Jambusaria A, Hong Z, Marsboom G, Toth PT, Herbert BS, MalikAB, Rehman J. SOX17 regulates conversion of human fibroblasts intoendothelial cells and erythroblasts by dedifferentiation into CD34(+)progenitor cells. Circulation 2017;135:2505–2523.
Zhou Q, Wang M, Yuan Y, Wang X, Fu R, Wan H, Xie M, Liu M, Guo X,Zheng Y et al. Complete meiosis from embryonic stem cell-derivedgerm cells in vitro. Cell Stem Cell 2016;18:330–340.
243Toolbox to characterize human migratory PGCs
Dow
nloaded from https://academ
ic.oup.com/m
olehr/article-abstract/24/5/233/4924981 by Leiden University / LU
MC
user on 22 July 2019