in situ transformation of striatal glia into cerebellar-like glia after brain transplantation
TRANSCRIPT

Neuroscience Letters, 136 (1992) 181-184 181 © 1992 Elsevier Scientific Publishers Ireland Ltd. All rights reserved 0304-3940/92/$ 05.00
NSL 08497
In situ transformation of striatal glia into cerebellar-like glia after brain transplantation
Claude Jacque a, J ean -L60n Tchel inger ian a, Pe ter Collins b and Isabelle Suard"
alNSERM U-134, H6pital de la Salpdtrikre, Paris (France) and b Ludwig Institut for Cancer Research, Stockholm (Sweden)
(Received 14 October 1991; Revised version received 30 December 1991; Accepted 2 January 1992)
Key words: Astrocyte; Cerebellum; Glial plasticity; Xenograft; Cell migration
Transplants of striatum from rabbit embryo were implanted into the colliculus posterior of newborn mice. After 4 weeks, astroglial cells derived from the transplant had migrated into the cerebellum of the host. Wherever they had settled in the cerebellum they presented forms similar to local glia. Some migrated glial cells were found to transform into forms of glia, such as radial-like glia, which are not present in the striatum. This observation confirms that glial precursor cells are highly plastic. It is an in vivo demonstration that local conditions alone define the morphology of glial cells. After grafting in an heterotopic location they take on forms that they were not destined to express in the region of origin.
Astroglial cells have characteristics of local specificity which may be functional or morphological. In the cerebellum, radial glia in the molecular layer and intersti- tial astrocytes in the granular layer, present a morphol- ogy which is encountered nowhere else in the adult central nervous system. The astroglia in the striatum is composed of two different populations with the ap- pearance of either fibrous or protoplasmic astrocytes. We have used the capacity of glial cells to migrate in the host parenchyma after brain transplantation to allow striatal glial cells, implanted in posterior brain, the op- portunity to enter the cerebellum. We then carefully ob- served these cells in order to see if they had taken the morphological forms of the cerebellar astroglia or, al- ternately, had kept the morphology of striatal glia. We report here that striatal astroglia, once in the cerebellum, present the morphological appearances ofcerebellar glia: they participate in all sub-classes of astroglia normally present in the cerebellum, including sub-classes which do not normally develop in the striatum such as glia with long processes radially oriented.
According to their localization in the brain the astrocytes express different receptors for neuromediators or neuropeptides [3-5, 10]. Moreover, in vitro experi- ments have shown that striatal astrocytes do not exert the same influence on mesencephalic neurons, in terms of neuritic development, as do mesencephalic astrocytes [1,
Correspondence." C. Jacque, INSERM U-134, H6pital de la Salp6tri~re, 47 bid de l'H6pital, 75651 Paris c6dex 13, France.
7]. These are examples of functional specificity. The glial cells are not only region specific, but they are also mobile. They have the capacity to move in a tissue, espe- cially when they are not fully differentiated and the tissue is not completely mature. Examples of such migration are found in brain transplantation experiments in which immature brain tissue fragments are implanted into xe- nogenic newborn brains [11-13, 16, 17] or into isogenic adult brain [9, 14] or spinal cord [8]. During the first few days after grafting the glial precursors escape the graft mass, enter the host parenchyma and migrate, sometimes over long distances [16]. We don't know when the glial precursors start to express, in the course of their matura- tion, the topographic specificity that differentiated astrocytes exhibit. Do the precursors become specifically committed due to in situ influences once they have reached their final location or do they reach a specific location because they are genetically programmed to do so? Are striatal glial precursors committed to become striatal glia, only composed of fibrous and protoplasmic astrocytes, or do they have the potential to develop into other forms of glial cells which are not normally present in this region of the brain? In order to answer this ques- tion we have taken advantage of the capacity of im- planted glial cells to migrate following transplantation in the colliculus of recipient newborn mice. From this loca- tion they would likely follow the flux of autologous glial cells that populate the cerebellum [15]. This protocol offers the potent advantage of determining the fate of striatal astrocytes in non-damaged cerebellum, avoiding

!
182
Fig. 1. Frozen sections from the brain of a non-implanted 5-week-old mouse. Staining with rabbit polyclonal antibodies to the glial fibrillary acidic protein (GFAP) (Dako, Glostrup, Denmark). A: fronto-dorsal portion of the striatum with part of the corpus callosum (cc) visible on the top, B: central portion of the striatum. On both A and B proto- plasmic (arrows) and pericapillar glia (arrowheads) are the most fre-
quent aspects of the astroglia in the striatum. Magnification: × 150.
the possible influence of trauma or inflammation on the morphological appearance of neighbouring astrocytes. The morphological question is: will striatal astrocytes become cerebellar glia or remain striatal glia in the cerebellum? One may object that fibrous astrocytes in the white matter as well as protoplasmic astrocytes in the grey matter have similar appearances whatever their lo- calization, in the striatum or in the cerebellum. However, the cerebellum has glial cell populations which, in the adult, are unique: radial glia and interstitial glia of the granular layer. These types of glial cells are not present in the striatum. Therefore, striatal precursors, if prede- termined to become only mature striatal glia, would be unable to develop into these specific forms of cerebellar glia.
The model we used to trace astrocytes after brain transplantation is based on the capacity of a monoclonal antibody raised against glial fibrillary acidic protein (GFAP) to specifically recognize rabbit [11], but not mouse [6] GFAP under appropriate fixation conditions [2]. This identification method imposes the use of xeno- transplantation.
Charcoal-mixed fragments of striatum taken from E25 rabbit embryos were implanted into the colliculus post- erior of newborn C3H/SW mice. After survival times ranging from one to 8 weeks the grafted mice were perfused and the brains removed and treated for immu- nohistochemistry on sagittal frozen sections. The exact location of the site of implantation was determined by means of the charcoal bits and we checked that the cerebellum had not been damaged by the operation. Transplant-derived astroglia were sought on serial sec-
tions of whole brain including the cerebellum. Immu- nostaining with polyclonal anti-GFAP antibodies was used to obtain an overview of the aspect of astroglia in the host cerebellum, whereas monoclonal Tp-GFAP 1 antibodies were used to identify those glial cells which came from the rabbit striatal implant. The indirect im- munofluorescence techniques have been previously de- scribed [16].
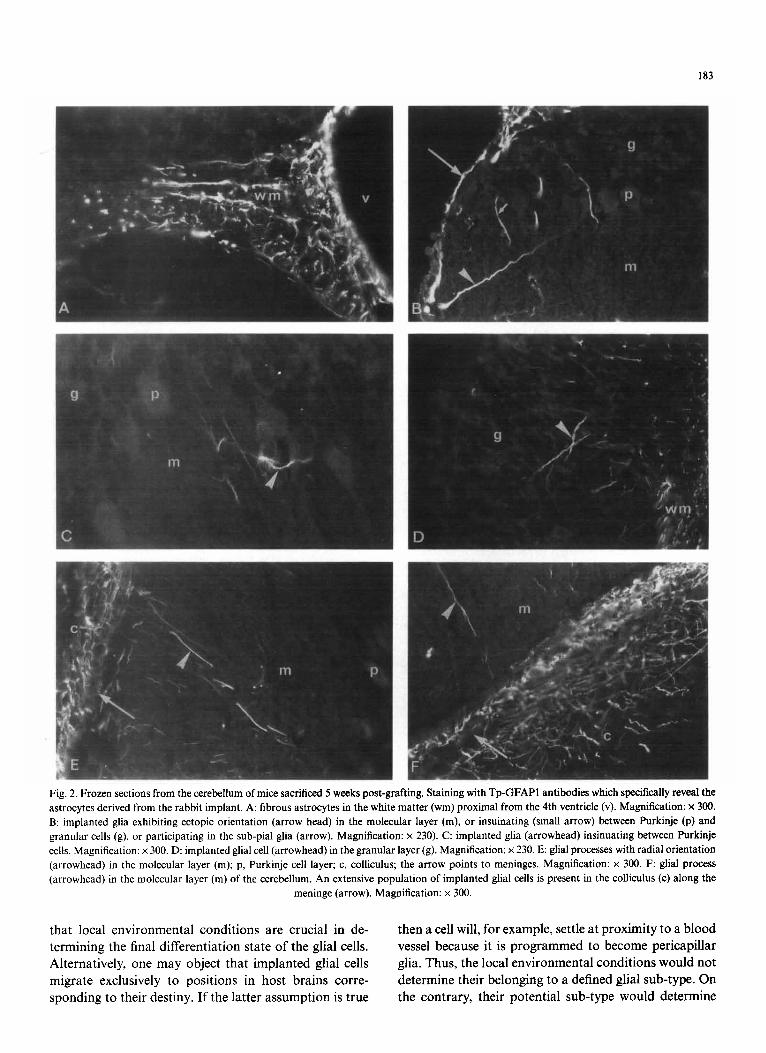
The classical aspect of striatal astroglia is shown in Fig. 1. Protoplasmic and pericapillar glia in the grey matter and fibrous glia in the white matter constitute most of the astroglia in the striatum of a normal adult brain. In brain transplantation experiments, glial cells from the implants were found in different regions of the host brains, including the cerebellum. As shown in Fig. 2, rabbit astrocytes were found to assume the different forms of host glia, including the cerebellar-specific glia. They were observed in the white matter with the form of fibrous astrocytes (Fig. 2A); in the granular layer in- sinuating between neurons (Fig. 2B,D) and in the molec- ular layer presenting the form of radial processes (Fig. 2E,F). The two latter forms are not found in the striatum. Implanted glial cells could also be found in ec- topic positions (Fig. 2B).
Therefore, if committed to become striatal glia exclu- sively, the glial precursors present in striatal transplant should be unable to participate in interstitial or radial glia. Since they do so, it means that they are not pre- programmed. Thus, their final differentiation keeps open. They are flexible for late differentiation according to local determinants. Although the majority of the im- planted glial cells are morphologically well integrated in host parenchyma, a few cells are found in ectopic posi- tions. Their abnormal orientation might result from local space constraints which prevent them from extending their processes properly.
This study shows clearly that glial precursors present in the striatum of E25 rabbit embryos have the potential to become cerebellar glia, including those forms of cerebellar glia which are not found in the striatum. We do not know how long into ontogenetic development that striatal glial precursors retain this flexibility. In- tegration of heterotopic implanted glia with local host glia had already been shown by previous investigations [13, 16]. Those studies showed that wherever they settle, migrating glial cells adopt morphological appearances similar to that of the neighboring host astroglia. This is true whatever their region of origin. When settling at proximity to pia mater they participate in sub-pial glia, at proximity to a blood vessel they ensheath it and be- come pericapillar glia, in white matter they present a fibrillar morphology, in grey matter they present a proto- plasmic appearance. In terms of plasticity it may mean
183
J
i
Fig. 2. Frozen sections from the cerebellum of mice sacrificed 5 weeks post-grafting. Staining with Tp-GFAP1 antibodies which specifically reveal the astrocytes derived from the rabbit implant. A: fibrous astrocytes in the white matter (wm) proximal from the 4th ventricle (v). Magnification: x 300. B: implanted glia exhibiting ectopic orientation (arrow head) in the molecular layer (m), or insuinating (small arrow) between Purkinje (p) and granular cells (g), or participating in the sub-pial glia (arrow). Magnification: x 230). C: implanted glia (arrowhead) insinuating between Purkinje cells. Magnification: × 300. D: implanted glial cell (arrowhead) in the granular layer (g). Magnification: × 230. E: glial processes with radial orientation (arrowhead) in the molecular layer (m); p, Purkinje cell layer; c, colliculus; the arrow points to meninges. Magnification: x 300. F: glial process (arrowhead) in the molecular layer (m) of the cerebellum. An extensive population of implanted glial cells is present in the colliculus (c) along the
meninge (arrow). Magnification: x 300.
that local environmental condit ions are crucial in de- termining the final differentiation state o f the glial cells. Alternatively, one may object that implanted glial cells migrate exclusively to positions in host brains corre- sponding to their destiny. I f the latter assumpt ion is true
then a cell will, for example, settle at proximity to a b lood vessel because it is p rog rammed to become pericapillar glia. Thus, the local environmental condit ions would not determine their belonging to a defined glial sub-type. On the contrary, their potential sub-type would determine

184
their topographic position after migration in the host pa- renchyma. In our previous studies [13, 16], the final dif- ferentiation states of the implanted glia were the diverse forms of glia which are present in all brain regions. Therefore, these studies could not support one of these hypothesis.
The important observation in the present study is that developmental flexibility allows cells, which, if main- tained in the striatum, would not have become interstitial or persistent radial glia to do so after migration into the cerebellum. It strongly suggests that the local envi- ronmental conditions play a crucial role in determining the morphological appearance of glial cells and there- fore, to a certain extent, their belonging to a defined sub- type.
This work was supported by an INSERM grant. We are grateful to R. M. Alvarado-Mallart for helpful dis- cussions and to J. Booss for reviewing this manuscript.
1 Barbin, G., Katz, D., Chamak, B., Glowinski, J. and Prochiantz, A., Brain astrocytes express region-specific surface glycoproteins in culture, Glia, 1 (1988) 96--103.
2 Bell, P., Runquist, I., Svensson, I. and Collins, P., Formaldehyde sensitivity of a GFAP epitope, removed by extraction of the cytos- queleton with high salt, J. Histochem. Cytochem., 35 (1987) 1375 1380.
3 Beyer, C., Pilgrim, C., Reisert, I. and Misgeld, U., Cells from em- bryonic rat striatum cocultured with mesencephalic glia express do- paminergic phenotypes, Neurosci. Lett., 128 (1991) 1-3.
4 Chneiweiss, H., Glowinski, J. and Premont, J., Modulation by mo- noamines of somatostatin-sensitive adenylate cyclase on neuronal and glial cells from the mouse brain in primary cultures, J. Neu- rochem., 44 (1985) 1825-1831.
5 Cholewinski, A. and Wilkin, G., Astrocytes from forebrain, cerebel- lum and spinal cord differ in their responses to vasoactive intestinal peptide, J. Neurochem., 51 (1988) 1626-1633.
6 Collins, P. and Moser, R., Monoclonal antibodies to glial fibrill~try acidic protein, Acta Path. Microbiol. Immunol. Scand. A, 91 (1983) 269-279.
7 Denis-Donini, D., Glowinski, J. and Prochiantz, A., Glial hetero- geneity may define the three-dimensional shape of mouse mesence- phalic dopaminergic neurons, Nature, 307 (1984) 641-643.
8 Goldberg, W. and Bernstein, J., Transplant-derived astrocytes migrate into host lumbar and cervical spinal cord after implantation of El4 fetal cerebral cortex into adult thoracic spinal cord, J. Neurosci. Res., 17 (1987) 391~,03.
9 Goldberg, W. and Bernstein, J., Fetal cortical astrocytes migrate from cortical homographs throughout the host brain and over the glia limitans, J. Neurosci. Res., 20 (1988) 3845.
10 Hansson, E. and R6nnb~ick, L., Interaction between catecho- lamines and vasoactive intestinal peptide in cultured astrocytes, Neuropharmacology, 27 (1988) 295-300.
11 Jacque, C., Suard, I., Collins, P. and Raoul, M., Identification of astrocytes after intracerebral transplantation, Dev. Neurosci., 8 (1986) 142-149.
12 Jacque, C., Suard, I., Ignacio, V., Collins, P., Raoul, M. and Baumann, N., Time-course expression of GFAP by implanted astrocytes after intracranial grafting of immature and mature brain tissues, Prog. Brain Res., 78 (1988) 387-393.
13 Jacque, C., Suard, I., Collins, P. and Baumann, N., Migration patterns of donor astrocytes after reciprocal striatum-cerebellum transplantation into newborn hosts, J. Neurosci. Res., 29 (1991) 421~,28.
14 Raisman, G., Lawrence, J., Zhou, C, and Lindsay, R., Some neu- ronal, glial and vascular interactions which occur when developing hippocampal primordia are incorporated into adult host hip- pocampi. In A. Bj6rklund and U. Stenevi (Eds.), Neural grafting in the mammalian CNS, Elsevier, Amsterdam, 1985, pp, 125-150.
15 Reynolds, R. and Wilkin, G., Development ofmacroglial ceils in rat cerebellum. II. An in situ immunohistochemical study of oligoden- droglial lineage from precursor to mature myelinating cell, Devel- opment, 102 (1988) 409~,25.
16 Suard, I., Collins, P., Ignacio, V. and Jacque, C., Implantation of rabbit embryo brain fragments into newborn mice: integration and survival of xenogeneic astrocytes, J. Neurosci. Res., 23 (1989) 172- 179.
17 Zhou, H.F., Lee, L.H.C. and Lund, R., Timing and patterns of astrocyte migration from xenogenic transplants of the cortex and corpus callosum, J. Comp. Neurol., 292 (t990) 320-330.