implication of an endogenous muscle-derived signalling system file544 l. p. baker and others...
TRANSCRIPT

Journal of Cell Science 102, 543-555 (1992)Printed in Great Britain © The Company of Biologists Limited 1992
543
Induction of acetylcholine receptor clustering by native polystyrene beads
Implication of an endogenous muscle-derived signalling system
LAUREN P. BAKER, QIMING CHEN and H. BENJAMIN PENG*
Department of Cell Biology and Anatomy and the Curriculum in Neurobiology, University of North Carolina, Chapel Hill, North Carolina27599, USA
*Author for correspondence at Department of Cell Biology and Anatomy, University of North Carolina, CB#7090, 108 Taylor Hall,Chapel Hill, NC 27599, USA
Summary
Aneural muscle cells in culture often form acetylcholinereceptor (AChR) clusters, termed hot spots, which aresimilar to those found at the postsynaptic membraneboth in structure and in molecular composition.Although hot spots form on both dorsal and ventralsurfaces of the cell, the ventral ones are bettercharacterized because of their association with sites ofcell-substratum contact. To understand the stimuli andmechanisms involved in ventral hot spot formation,native, uncoated polystyrene beads were applied tocultured Xenopus myotomal muscle cells to create localmembrane-substratum contacts. These beads were ableto induce a postsynaptic-type development as evidencedby the clustering of AChRs and the development of a setof ultrastructural specializations, including membraneinfoldings and a basement membrane. Whereas thesenative beads were effective in inducing clustering, beadscoated with bovine serum albumin or treated withserum-containing medium were ineffective. Nativebeads were also capable of inducing clusters in serum-free medium, indicating that their effect was mediatedby endogenous molecules that were locally presented bythe beads, rather than by bead adsorption of com-ponents in the medium. Heparan sulfate proteoglycan(HSPG) is a major component of the muscle extracellu-lar matrix and our previous study has shown that basicfibroblast growth factor (bFGF), a member of theheparin-binding growth factor (HBGF) family, and itsreceptor are present in Xenopus myotomal muscleduring the period of synaptogenesis. Therefore, we
tested the involvement of HBGF in bead induction. Theresults of this study show the following: (1) preincu-bation of cultures in heparin, which solubilizes matrix-bound HBGFs, suppressed the bead-induced AChRclustering. (2) Suramin, which interferes with theinteraction between several growth factors and theirreceptors, also inhibited bead-induced clustering.(3) Tyrphostin, which blocks tyrosine kinase activityassociated with a number of growth factor receptors, wasalso inhibitory to the bead effect. (4) The percentage ofbead-induced AChR clusters was significantly enhanced bypretreating the cultures with bFGF prior to beadapplication. This exogenously applied bFGF could belargely removed by treatment of cultures with heparin,suggesting its association with HSPG at the cell surface.(5) An anti-bFGF neutralizing antiserum significantlyreduced the efficacy of the bead stimulation. These datasuggest that uncoated beads, which adhere to the cellsurface and can mimic the cell-substratum interaction,effect a local presentation of HBGFs, such as bFGF,residing with the HSPG to their membrane receptors,thereby locally activating receptor-associated tyrosinekinases. This may be followed by tyrosine phosphorylationof yet unidentified substrate proteins, resulting in cluster-ing of AChRs. We propose that a similar local presentationof HBGF may also underlie the postsynaptic induction atdeveloping neuromuscular junctions.
Key words: ACh receptor, neuromuscular junction,heparin-binding growth factor.
Introduction
Tissue culture of nerve and muscle cells offers aconvenient system for studying synaptogenesis. Whenprocesses from motoneurons come into contact withmuscle cells in culture, functional neuromuscularjunctions (NMJs) develop. These in vitro junctions
acquire a considerable degree of specialization asevidenced by the clustering of acetylcholine receptors(AChRs) in the postsynaptic membrane, the accumu-lation of acetylcholinesterase in the synaptic cleft andthe development of synaptic vesicle clusters in the nerveterminal (Bloch and Pumplin, 1988; Peng, 1987). It isgenerally thought that the signal for postsynaptic

544 L. P. Baker and others
differentiation comes from the motoneuron. In theabsence of innervation, however, AChR clustering stilloccurs (Anderson et al. 1977; Fischbach and Cohen,1973; Sytkowski et al. 1973; Bloch and Pumplin, 1988).Aneural clusters, commonly called hot spots, bear astriking resemblance to the postsynaptic membrane ininnervated cells both in structure and in molecularcomposition. Although hot spots can be found any-where on the muscle cell, the most prominent ones aredisplayed at sites of close cell-substratum contact(Bloch and Pumplin, 1988). They form on substratacoated with proteinaceous molecules, such as collagenand polylysine, as well as on uncoated glass or plastic.Thus, contact with the substratum alone provides thenecessary signal for the induction of AChR clustering.
Previously we showed that AChR clustering can alsobe induced in cultured Xenopus muscle cells bypolycation-coated latex beads (Peng and Cheng, 1982).These studies suggest that the local presentation of astimulus in the form of a mini-substratum can mimic theaction of nerve in inducing postsynaptic differentiation.However, the nature of this stimulation is still un-known. Recently we became interested in the role ofpolypeptide growth factors in NMJ development, sincethis class of molecules is universally used in thesignalling of cell differentiation and proliferation. Ofparticular interest is basic fibroblast growth factor(bFGF), an extracellular matrix (ECM)-associatedgrowth factor (Esch et al. 1985; Gospodarowicz et al.1986). bFGF has a strong affinity for heparin, and forheparan sulfate proteoglycans (HSPG), which provideextracellular storage sites for this growth factor (Bur-gess and Maciag, 1989; Rifkin and Moscatelli, 1989).ECM-associated bFGF can be mobilized by surround-ing cells to exert an autocrine or paracrine regulation oncellular processes (Saksela and Rifkin, 1990; Rifkin andMoscatelli, 1989). Both bFGF and HSPG are found onthe surface of developing skeletal muscle, which alsopossesses receptors for bFGF (Anderson and Fam-brough, 1983; Bayne et al. 1984; Joseph-Silverstein etal. 1989; Seed et al. 1988; Olwin and Hauschka, 1988).Recently we showed that bFGF-coated beads can alsoinduce the formation of AChR clusters (Peng et al.1991). This suggests that bFGF and/or other heparin-binding growth factors (HBGFs) at the cell surface maycontribute to the formation of hot spots.
To test this hypothesis, we examined the effect ofnative uncoated polystyrene beads on the formation ofAChR clusters in cultured Xenopus muscle cells. Herewe report that these uncoated beads also induce AChRclustering, through a mechanism that appears to involvean interaction between HBGFs, their receptors andtyrosine phosphorylation. Since exogenous bFGF,which binds to HSPG at the cell surface, significantlyincreases the efficacy of the bead-induced AChRclustering and a neutralizing bFGF antiserum sup-presses this process, the role of bFGF is implicated.This work was previously presented in abstract form(Chen et al. 1990).
Materials and methods
MaterialsHuman recombinant bFGF was a gift from Synergen(Boulder, CO). The tyrphostins, RG50864 (active) andRG50862 (inactive), were gifts from Rhone-Poulanc RorerCentral Research (Fort Washington, PA). Polystyrene latexmicrospheres were purchased from Polysciences (Warrington,PA). The bFGF monoclonal antibody (mAb; type I) was fromUpstate Biotechnology, Inc. (Lake Placid, NY). Tetramethyl-rhodamine or FITC-conjugated a--bungarotoxin (R-BTX orF1TC-BTX) were purchased from Molecular Probes, Inc.(Eugene, OR). BSA, heparin and chondroitin sulfate A werefrom Sigma Chemical Co. (St. Louis, MO). Suramin waspurchased from FBA Pharmaceuticals (New York, NY).Fluorescent goat anti-mouse secondary antibody was fromOrganon Technika (Durham, NC). Anti-bFGF neutralizingantiserum (77R) was a generous gift from Dr. M. Klagsbrun(The Children's Hospital, Boston, MA). A second polyclonalanti-bFGF antibody from American Diagnostica (Greenwich,CT) was also used for blocking experiments. Sheep anti-rabbit IgG antiserum was also purchased from Organon-Teknika (cat. no. 55603).
Preparation of muscle and nerve culturesMuscle cultures were prepared according to a previouslypublished protocol (Peng and Nakajima, 1978). Stage 20-22Xenopus embryos were freed from the surrounding jelly coatand vitelline membrane, and the dorsal, myotomal portionwas dissected away and dissociated using a calcium- andmagnesium-free Steinberg solution (60 mM NaCl, 0.7 mMKC1, 0.4 mM EDTA, 10 mM HEPES, pH 7.4). Muscle cellswere plated onto glass coverslips and grown in a culturemedium composed of Steinberg solution (60 mM NaCl, 0.7mM KC1, 0.4 mM Ca(NO3)2, 0.8 mM MgSO4, and 10 mMHEPES, pH 7.4) supplemented with 10% L-15 medium, 1%fetal bovine serum, and 0.1 mg/ml gentamycin. Cultures weremaintained at 15°C or at 22°C, and were routinely used within2-7 days after plating.
Preparation and application of polystyrene beadsPolystyrene latex beads (5 to 10 /im in diameter) were washedfor 30 min in 95% ethanol, and rinsed three times in distilledwater. They were then immediately applied to cultures orstored in PBS at 4°C. These beads were not exposed to anyproteinaceous molecules (uncoated). The effect of maskingthe bead surface by coating them with BSA or serum proteinswas also examined in this study. To coat beads with bovineserum albumin (BSA), they were treated with PBS containingBSA at various concentrations for 0.5-1 h after the ethanolwash and then rinsed three times with PBS. To coat beadswith serum proteins, the ethanol-washed beads were treatedwith culture medium that contained 1% fetal bovine serum for0.5-1 h.
Beads were applied to muscle cells via micropipette. Toprevent uncoated beads from being coated by serum, themedium level over the cells was kept at a minimum during thebead application. The beads were allowed to associate withthe cells for a brief period (approximately 10-50 s), and thenexcess, nonadhered beads were rinsed away with culturemedium. Bead-seeded cultures were incubated for 1-30 h at22°C, and then were labeled with 0.3 fM R-BTX for 30 min.Cells were fixed for 3 min in 95% ethanol at -20°C andvisualized with fluorescence microscopy. The effect of bead-induced AChR clustering was quantified by scoring the

ACh receptor clustering induced by polymer beads 545
percentage of bead-muscle contacts associated with fluor-escent BTX labeling. This was calculated for each cell andthen the mean and standard error of the percentages from allcells in a culture were calculated. These values are presentedin the figures. The cells were chosen at random.
Visualization of bFGF by immunofluorescencemicroscopyImmunofluorescence studies were conducted to visualizebFGF bound to the cell surface in untreated and in bFGF-treated cultures. First, the cultures were treated with a mAbagainst bFGF for 30 min at a concentration of 50 nM, followedby extensive washing, and labeled with rhodamine or FITC-conjugated goat anti-mouse IgG for 30 min. The cultures werethen fixed with 95% ethanol at —20°C for 3 min and examinedunder a fluorescence microscope. In some experiments,cultures were double-labeled with FJTC-BTX and the bFGFantibody followed by rhodamine-conjugated secondary anti-body.
Confocal microscopy, using a Biorad MRC-600 laserscanner and a Nikon inverted microscope, was also conductedon these specimens. The cells were optically sectioned at 0.5-1,um increments and stereo images were constructed with theBiorad software.
Electron microscopyMuscle cells were cultured in Lux Permanox tissue culturedishes (Nunc, Naperville, IL). After incubation with beadsfor 24 h, they were fixed with 1% glutaraldehyde in 0.1 Mphosphate buffer (pH 7.2), postfixed with 1% OsO4, en blocstained with 1% uranyl acetate in 0.1 M acetate buffer (pH5.5), dehydrated through an ethanol series, and embedded inepoxy resin. Bead-bearing cells were scored with a Leitzdiamond marker on a microscope and sawed off the block.Ultrathin sections were cut on a microtome with a diamondknife. They were poststained with uranyl acetate and leadcitrate and examined with a JEOL 200CX transmissionelectron microscope.
Drug applicationSuramin was dissolved in Steinberg solution to make a 10 mMstock solution. The stock solution was filter-sterilized andfinal concentrations were prepared in Steinberg culturemedium by serial dilution. Muscle cell cultures were incu-bated in medium containing suramin for the duration of theexperiment.
The tyrphostins RG50864 (active) and RG50862 (inactive)were dissolved in dimethylsulfoxide (DMSO) to make a 40mM stock solution. This solution was divided into aliquotsand stored at —80°C. Before each experiment, workingsolutions were prepared in Steinberg medium by serialdilution. Cultures were preincubated in tyrphostins for 7 h.After the addition of beads, the cultures were maintained inthe presence of the tyrphostins for 12 h, labeled with R-BTXand fixed. In addition to comparing the effect of the active andthe inactive tyrphostins, we also examined the effect of 0.2%DMSO (corresponding to the highest concentration used inthe experimental groups) on bead-induced AChR clusteringand found that DMSO had no effect at this concentration.
Results
Induction of AChR clustering by uncoated beadsClean polystyrene latex beads attached readily tocultured Xenopus cells. After an incubation period of
24 h, clusters of AChRs, as shown by bright fluorescentstaining of R-BTX, were observed at the bead-musclecontacts (Fig. la-b). These clusters developed discre-tely at bead-muscle contacts and their size wasproportional to the size of the beads applied. Althoughmost muscle cells already had hot spots on their surfacebefore the addition of beads, the bead-associatedclusters arise as a result of de novo assembly and notdue to attachment of beads to preexisting hot spots forthe following reasons: (1) beads seldom landed on pre-existing clusters as shown by the lack of R-BTX stainingat contact sites immediately after the addition of beads.(2) Beads came into contact only with the top surface ofthe cell whereas most hot spots were present on thebottom surface. Similar to results of a previous study(Peng, 1986), we also found that hot spots disappearwith time on cells exhibiting bead-induced clusters.
On average, 70% of bead-muscle contacts developedAChR clusters. When the bead-muscle contacts wereexamined by electron microscopy, a highly specializedmembrane area was revealed. First, beads came intoclose contact with the muscle cell membrane, leaving acleft distance of less than 30 nm at the narrowest pointof contact (Fig. lc). The membrane apposing the beadshowed a thickening of the plasmalemma and itsassociated cortical cytoplasm and was overlain withpatches of basal lamina. In addition, the membrane wasdeeply invaginated (Fig. lc). These features are allcharacteristic of the postsynaptic membrane at theNMJ.
AChR clusters could be detected by R-BTX labelingwithin the first hour of bead application, and thenumber of clusters reached a plateau after 7-8 h ofincubation. Afterward, there was an increase influorescence intensity without an increase in numberduring the first 24 h. Thus, the mechanism for clusteringAChRs is set up within the first hour of bead-musclecontact and the recruitment of receptors to the contactsite continues at least through the first 24 h.
Proteins adsorb readily to clean polystyrene surfaces.To examine whether this property was responsible forthe induction of AChR clustering, the free bead surfacewas masked with culture medium containing 1% fetalbovine serum, or with BSA. After beads were incu-bated for 30 min in serum-containing medium andapplied to cells, they were found to be ineffective ininducing AChR clustering. The same effect was ob-served after the beads were incubated with BSA (Fig.2). Beads coated with more than 100 ^g/ml BSA wereunable to induce clustering. BSA-coated beads wereless adherent to the cells than uncoated beads as shownby a decrease in the mean number of beads remainingattached to cells after the cultures were rinsed immedi-ately after bead application. However, remaining beadsstill became firmly anchored to the cell surface withtime. Electron micrographs of these ineffective bead-muscle contacts revealed that the cleft distance was stillnarrow, ranging from 30 to 50 nm. However, noultrastructural specializations were detected at thesebead-muscle contacts (data not shown; cf. Fig. 6 ofPeng et al. 1991). These studies suggest that the AChR

546 L. P. Baker and others
Fig. 1. AChR clusters inducedby uncoated polystyrene beads.9 )jm beads were used in thisstudy, (a) Fluorescencemicrographs showing R-BTX-labeled AChR clusters that arelocated at bead-musclecontacts as shown in thephase-contrast micrograph in(b). The numbers point to thecorrespondence in thepositions of the AChR clustersand the beads, (c) Electronmicrograph of ultrastructuralspecializations induced by anuncoated bead. Blackarrowheads point to basallamina accumulation. Openwhite arrowheads point tomembrane infoldings. Filledwhite arrows point tomembrane densities indicativeof sites of AChRconcentration. B, bead.
cluster-inducing capability of uncoated beads is notsimply due to a mechanical deformation of themembrane at the bead-muscle contacts. We alsoconducted experiments in Steinberg solution alone,without added serum or L-15 medium. Under thiscondition, beads were still effective in inducing AChRclustering (data not shown). Thus, this effect is notmediated by factors in the culture medium that bind tothe beads. Rather, factors that are localized at the cellsurface may be responsible for cluster formation. Thesefactors may pre-exist on the cell surface, secreted by thecells themselves during bead-muscle contact, or derivedfrom the culture medium.
We also experimented with beads coated with othermolecules. As reported previously (Peng et al. 1991),beads coated with polycations or with bFGF are veryeffective in inducing AChR clustering. However, beadscoated with other basic molecules, such as cytochrome cor the active (fi) subunit of nerve growth factor, wereineffective. Beads coated with nonspecific immuno-globulins, laminin or tenascin were similarly ineffective.All of these coated beads adhered tightly to the cellsurface, suggesting that the adhesion alone is notsufficient to trigger the clustering process.
Inhibition of AChR clustering by heparin and suraminPrevious studies have shown the existence of HSPG atthe surface of Xenopus muscle cells (Anderson and
101 10J 10*
BSA concentration (|ig/ml)
Fig. 2. Inhibition of uncoated bead-induced AChRclustering by masking the bare bead surface with BSA.Beads were treated with BSA solution for 30 min beforeaddition to the cells. The percentage of beads associatedwith AChR clusters showed a decline when the BSAconcentration reached 0.1 ^g/ml. AChR clustering at thebeads was abolished when the BSA concentration reached100 /ig/ml. 20 cells, each with 1-20 bead-muscle contacts,were scored for each data point. Each point represents themean±s.e.m.
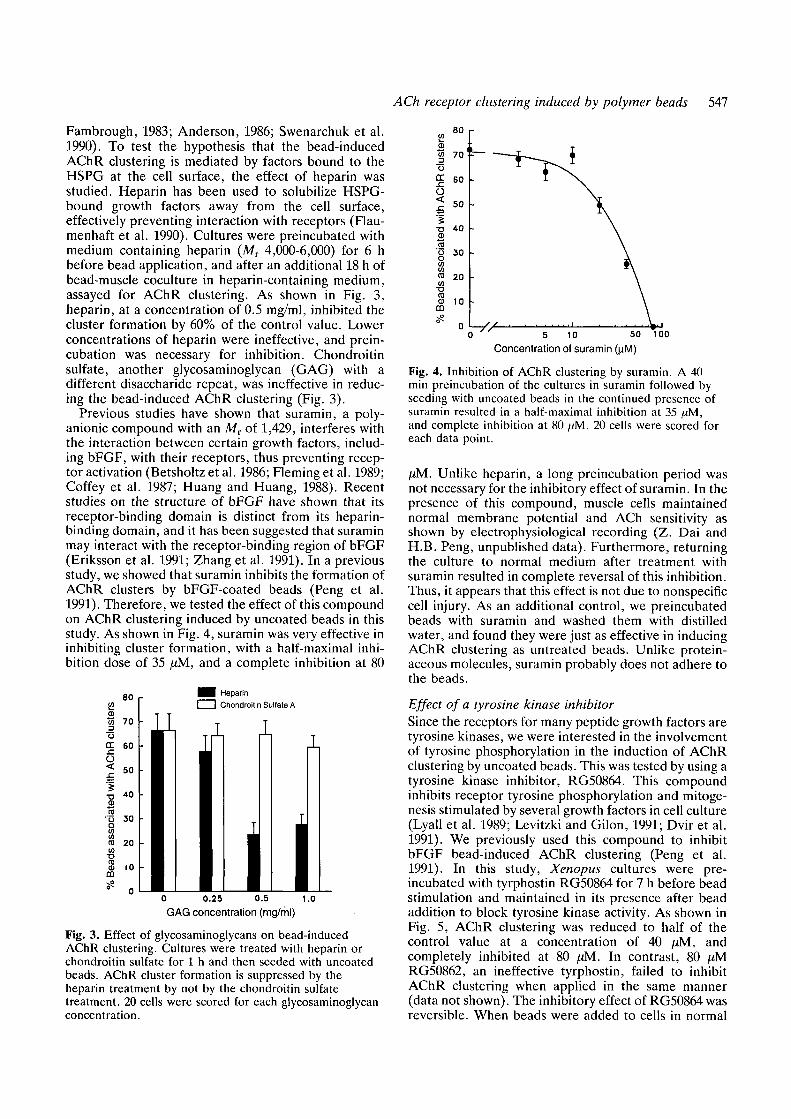
ACh receptor clustering induced by polymer beads 547
Fambrough, 1983; Anderson, 1986; Swenarchuk et al.1990). To test the hypothesis that the bead-inducedAChR clustering is mediated by factors bound to theHSPG at the cell surface, the effect of heparin wasstudied. Heparin has been used to solubilize HSPG-bound growth factors away from the cell surface,effectively preventing interaction with receptors (Flau-menhaft et al. 1990). Cultures were preincubated withmedium containing heparin (MT 4,000-6,000) for 6 hbefore bead application, and after an additional 18 h ofbead-muscle coculture in heparin-containing medium,assayed for AChR clustering. As shown in Fig. 3,heparin, at a concentration of 0.5 mg/ml, inhibited thecluster formation by 60% of the control value. Lowerconcentrations of heparin were ineffective, and prein-cubation was necessary for inhibition. Chondroitinsulfate, another glycosaminoglycan (GAG) with adifferent disaccharide repeat, was ineffective in reduc-ing the bead-induced AChR clustering (Fig. 3).
Previous studies have shown that suramin, a poly-anionic compound with an MT of 1,429, interferes withthe interaction between certain growth factors, includ-ing bFGF, with their receptors, thus preventing recep-tor activation (Betsholtz et al. 1986; Fleming et al. 1989;Coffey et al. 1987; Huang and Huang, 1988). Recentstudies on the structure of bFGF have shown that itsreceptor-binding domain is distinct from its heparin-binding domain, and it has been suggested that suraminmay interact with the receptor-binding region of bFGF(Eriksson et al. 1991; Zhang et al. 1991). In a previousstudy, we showed that suramin inhibits the formation ofAChR clusters by bFGF-coated beads (Peng et al.1991). Therefore, we tested the effect of this compoundon AChR clustering induced by uncoated beads in thisstudy. As shown in Fig. 4, suramin was very effective ininhibiting cluster formation, with a half-maximal inhi-bition dose of 35 fxM, and a complete inhibition at 80
80COCDw 70O
cr 60x:O< 50
I•5 toS(0
i 3 0to 20to
•o
ID 10CD
^ n
Heparin
1 1 Chondroitin Sulfate AT T
" I T T Ti-U T•in•i•- •••I• ••- •
in•I••••1•••
_
0 0.25 0.5 1.0
GAG concentration (mg/ml)
Fig. 3. Effect of glycosaminoglycans on bead-inducedAChR clustering. Cultures were treated with heparin orchondroitin sulfate for 1 h and then seeded with uncoatedbeads. AChR cluster formation is suppressed by theheparin treatment by not by the chondroitin sulfatetreatment. 20 cells were scored for each glycosaminoglycanconcentration.
5 10 50 100
Concentration of suramin (uM)
Fig. 4. Inhibition of AChR clustering by suramin. A 40min preincubation of the cultures in suramin followed byseeding with uncoated beads in the continued presence ofsuramin resulted in a half-maximal inhibition at 35 ,aM,and complete inhibition at 80 [tM. 20 cells were scored foreach data point.
fiM. Unlike heparin, a long preincubation period wasnot necessary for the inhibitory effect of suramin. In thepresence of this compound, muscle cells maintainednormal membrane potential and ACh sensitivity asshown by electrophysiological recording (Z. Dai andH.B. Peng, unpublished data). Furthermore, returningthe culture to normal medium after treatment withsuramin resulted in complete reversal of this inhibition.Thus, it appears that this effect is not due to nonspecificcell injury. As an additional control, we preincubatedbeads with suramin and washed them with distilledwater, and found they were just as effective in inducingAChR clustering as untreated beads. Unlike protein-aceous molecules, suramin probably does not adhere tothe beads.
Effect of a tyrosine kinase inhibitorSince the receptors for many peptide growth factors aretyrosine kinases, we were interested in the involvementof tyrosine phosphorylation in the induction of AChRclustering by uncoated beads. This was tested by using atyrosine kinase inhibitor, RG50864. This compoundinhibits receptor tyrosine phosphorylation and mitoge-nesis stimulated by several growth factors in cell culture(Lyall et al. 1989; Levitzki and Gilon, 1991; Dvir et al.1991). We previously used this compound to inhibitbFGF bead-induced AChR clustering (Peng et al.1991). In this study, Xenopus cultures were pre-incubated with tyrphostin RG50864 for 7 h before beadstimulation and maintained in its presence after beadaddition to block tyrosine kinase activity. As shown inFig. 5, AChR clustering was reduced to half of thecontrol value at a concentration of 40 /JM, andcompletely inhibited at 80 (iM. In contrast, 80 fiMRG50862, an ineffective tyrphostin, failed to inhibitAChR clustering when applied in the same manner(data not shown). The inhibitory effect of RG50864 wasreversible. When beads were added to cells in normal

548 L. P. Baker and others
20 40 SO
Tyrphostin concentration (uM)BO
Fig. 5. Inhibition of AChR clustering by a tyrphostin,RG50864. Preincubation of cultures in tyrphostin for 7 h,followed by addition of uncoated beads in the continuedpresence of the drug resulted in half-maximal inhibition atapproximately 40 /xM. Complete inhibition of AChRclustering was observed at 80 JJM. 20 cells were scored foreach data point.
medium after a 7 h tyrphostin pre-incubation, AChRclustering returned to 90% of the control value.Although cells showed the formation of vacuolarstructures at higher concentrations, they were stillviable after prolonged incubation (24 h) as shown bytrypan blue exclusion (Peng et al. 1991).
Immunocytochemical localization of bFGF oncultured muscle cellsThe presence of bFGF and its receptor has beendemonstrated in the muscle tissue of Xenopus embryosat stages during which synaptogenesis occurs byWestern blot analysis in previous studies (Peng et al.
1991; Friesel and Dawid, 1991). In order to visualizebFGF associated with the muscle cell surface, indirectimmunofluorescence studies were carried out with amonoclonal antibody against bFGF. As shown in Fig.6a-b, a non-uniform, punctate staining pattern ofendogenous bFGF on the cell surface was revealed bythis procedure. It was not, however, concentrated atAChR clusters. The labeling was specific, as it wasabolished by omission of the primary anti-bFGFantibody.
The level of endogenous bFGF as shown by thismethod was relatively low when compared with theapparent abundance of HSPG on the surface of thesemuscle cells, as shown previously by immunofluor-escence microscopy (Anderson, 1986; Swenarchuk etal. 1990). This suggested that HSPGs on the cell surfacecould accommodate a much higher amount of bFGF.To study this possibility, human recombinant bFGF, ata concentration of 1 jig/ml, was bath-applied to theculture. After an incubation period of 1 h, the culturewas washed extensively and labeled with the anti-bFGFantibody. This resulted in a dramatic increase in thebFGF labeling at the surface of muscle cells. Fig. 7shows a composite confocal stereo image through amuscle cell treated according to this procedure. Theapplication of exogenous bFGF resulted in brightstaining of the cell surface. It showed that this staining isentirely restricted to the cell surface and no intracellularstaining was observed (Fig. 7). Again, the staining wasabolished if the primary antibody was omitted.Interestingly, not all cell types in the culture exhibitedthis intensified labeling after bFGF application. Forexample, melanocytes showed an absence of bFGFlabeling and fibroblasts showed a much reducedstaining as compared with muscle cells (results notshown). Thus, muscle cells appeared to be highlyenriched in surface binding sites for bFGF.
Fig. 6. Immunolocalization ofbFGF on cultured muscle cells,(a-b) Phase-contrast andfluorescence micrographs showingendogenous bFGF labeling.Arrowheads point to punctatesites of bFGF localizationrevealed by labeling with an anti-bFGF mAb and a rhodamine-conjugated secondary antibody,(c, d) The culture was treatedwith 1 jig/ml recombinant humanbFGF for 1 h and then washedwith medium containing 0.5mg/ml heparin for 1 h. It wasthen labeled with FITC-BTX (c)and anti-bFGF mAb followed byrhodamine-conjugated secondaryantibody (d). The residual bFGFwas especially concentrated atAChR clusters (white arrows).Away from AChR clusters, bFGFwas still present in a punctatemanner (arrowheads).
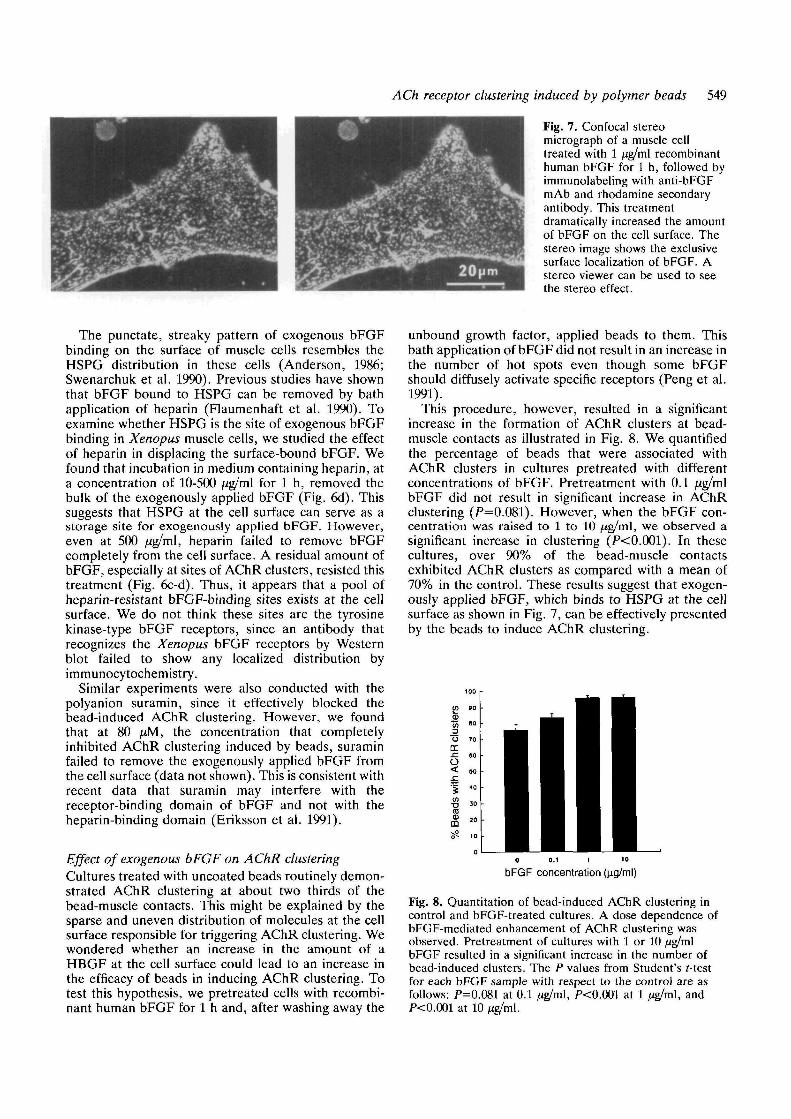
ACh receptor clustering induced by polymer beads 549
Fig. 7. Confocal stereomicrograph of a muscle celltreated with 1 /ig/ml recombinanthuman bFGF for 1 h, followed byimmunolabeling with anti-bFGFmAb and rhodamine secondaryantibody. This treatmentdramatically increased the amountof bFGF on the cell surface. Thestereo image shows the exclusivesurface localization of bFGF. Astereo viewer can be used to seethe stereo effect.
The punctate, streaky pattern of exogenous bFGFbinding on the surface of muscle cells resembles theHSPG distribution in these cells (Anderson, 1986;Swenarchuk et al. 1990). Previous studies have shownthat bFGF bound to HSPG can be removed by bathapplication of heparin (Flaumenhaft et al. 1990). Toexamine whether HSPG is the site of exogenous bFGFbinding in Xenopus muscle cells, we studied the effectof heparin in displacing the surface-bound bFGF. Wefound that incubation in medium containing heparin, ata concentration of 10-500 /ig/ml for 1 h, removed thebulk of the exogenously applied bFGF (Fig. 6d). Thissuggests that HSPG at the cell surface can serve as astorage site for exogenously applied bFGF. However,even at 500 /ig/ml, heparin failed to remove bFGFcompletely from the cell surface. A residual amount ofbFGF, especially at sites of AChR clusters, resisted thistreatment (Fig. 6c-d). Thus, it appears that a pool ofheparin-resistant bFGF-binding sites exists at the cellsurface. We do not think these sites are the tyrosinekinase-type bFGF receptors, since an antibody thatrecognizes the Xenopus bFGF receptors by Westernblot failed to show any localized distribution byimmunocytochemistry.
Similar experiments were also conducted with thepolyanion suramin, since it effectively blocked thebead-induced AChR clustering. However, we foundthat at 80 JJM, the concentration that completelyinhibited AChR clustering induced by beads, suraminfailed to remove the exogenously applied bFGF fromthe cell surface (data not shown). This is consistent withrecent data that suramin may interfere with thereceptor-binding domain of bFGF and not with theheparin-binding domain (Eriksson et al. 1991).
Effect of exogenous bFGF on AChR clusteringCultures treated with uncoated beads routinely demon-strated AChR clustering at about two thirds of thebead-muscle contacts. This might be explained by thesparse and uneven distribution of molecules at the cellsurface responsible for triggering AChR clustering. Wewondered whether an increase in the amount of aHBGF at the cell surface could lead to an increase inthe efficacy of beads in inducing AChR clustering. Totest this hypothesis, we pretreated cells with recombi-nant human bFGF for 1 h and, after washing away the
unbound growth factor, applied beads to them. Thisbath application of bFGF did not result in an increase inthe number of hot spots even though some bFGFshould diffusely activate specific receptors (Peng et al.1991).
This procedure, however, resulted in a significantincrease in the formation of AChR clusters at bead-muscle contacts as illustrated in Fig. 8. We quantifiedthe percentage of beads that were associated withAChR clusters in cultures pretreated with differentconcentrations of bFGF. Pretreatment with 0.1 fJ.g/m\bFGF did not result in significant increase in AChRclustering (P=0.081). However, when the bFGF con-centration was raised to 1 to 10 /ig/ml, we observed asignificant increase in clustering (P<0.001). In thesecultures, over 90% of the bead-muscle contactsexhibited AChR clusters as compared with a mean of70% in the control. These results suggest that exogen-ously applied bFGF, which binds to HSPG at the cellsurface as shown in Fig. 7, can be effectively presentedby the beads to induce AChR clustering.
« »°DC
6
3
bFGF concentration (ug/ml)
Fig. 8. Quantitation of bead-induced AChR clustering incontrol and bFGF-treated cultures. A dose dependence ofbFGF-mediated enhancement of AChR clustering wasobserved. Pretreatment of cultures with 1 or 10 /ig/mlbFGF resulted in a significant increase in the number ofbead-induced clusters. The P values from Student's f-testfor each bFGF sample with respect to the control are asfollows: P=0.081 at 0.1 /ig/ml, P<0.001 at 1 /ig/ml, andP<0.001 at 10 /ig/ml.
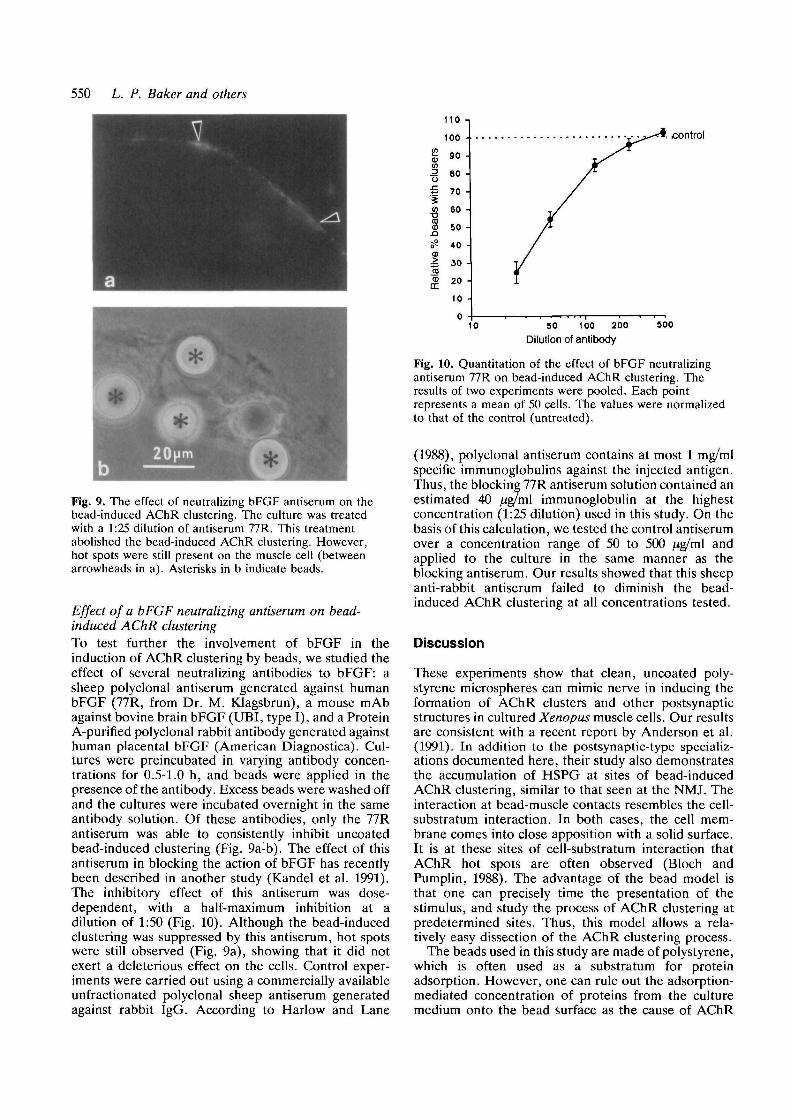
550 L. P. Baker and others
Fig. 9. The effect of neutralizing bFGF antiserum on thebead-induced AChR clustering. The culture was treatedwith a 1:25 dilution of antiserum 77R. This treatmentabolished the bead-induced AChR clustering. However,hot spots were still present on the muscle cell (betweenarrowheads in a). Asterisks in b indicate beads.
Effect of a bFGF neutralizing antiserum on bead-induced AChR clusteringTo test further the involvement of bFGF in theinduction of AChR clustering by beads, we studied theeffect of several neutralizing antibodies to bFGF: asheep polyclonal antiserum generated against humanbFGF (77R, from Dr. M. Klagsbrun), a mouse mAbagainst bovine brain bFGF (UBI, type I), and a ProteinA-purified polyclonal rabbit antibody generated againsthuman placental bFGF (American Diagnostica). Cul-tures were preincubated in varying antibody concen-trations for 0.5-1.0 h, and beads were applied in thepresence of the antibody. Excess beads were washed offand the cultures were incubated overnight in the sameantibody solution. Of these antibodies, only the 77Rantiserum was able to consistently inhibit uncoatedbead-induced clustering (Fig. 9a-b). The effect of thisantiserum in blocking the action of bFGF has recentlybeen described in another study (Kandel et al. 1991).The inhibitory effect of this antiserum was dose-dependent, with a half-maximum inhibition at adilution of 1:50 (Fig. 10). Although the bead-inducedclustering was suppressed by this antiserum, hot spotswere still observed (Fig. 9a), showing that it did notexert a deleterious effect on the cells. Control exper-iments were carried out using a commercially availableunfractionated polyclonal sheep antiserum generatedagainst rabbit IgG. According to Harlow and Lane
110 -i
100 -
90 -
80 •
70 -
60 -
50 -
40 •
30 •
20 -
10
0
control
10 SO 100 200
Dilution of antibody500
Fig. 10. Quantitation of the effect of bFGF neutralizingantiserum 77R on bead-induced AChR clustering. Theresults of two experiments were pooled. Each pointrepresents a mean of 50 cells. The values were normalizedto that of the control (untreated).
(1988), polyclonal antiserum contains at most 1 mg/mlspecific immunoglobulins against the injected antigen.Thus, the blocking 77R antiserum solution contained anestimated 40 ^g/ml immunoglobulin at the highestconcentration (1:25 dilution) used in this study. On thebasis of this calculation, we tested the control antiserumover a concentration range of 50 to 500 /ig/ml andapplied to the culture in the same manner as theblocking antiserum. Our results showed that this sheepanti-rabbit antiserum failed to diminish the bead-induced AChR clustering at all concentrations tested.
Discussion
These experiments show that clean, uncoated poly-styrene microspheres can mimic nerve in inducing theformation of AChR clusters and other postsynapticstructures in cultured Xenopus muscle cells. Our resultsare consistent with a recent report by Anderson et al.(1991). In addition to the postsynaptic-type specializ-ations documented here, their study also demonstratesthe accumulation of HSPG at sites of bead-inducedAChR clustering, similar to that seen at the NMJ. Theinteraction at bead-muscle contacts resembles the cell-substratum interaction. In both cases, the cell mem-brane comes into close apposition with a solid surface.It is at these sites of cell-substratum interaction thatAChR hot spots are often observed (Bloch andPumplin, 1988). The advantage of the bead model isthat one can precisely time the presentation of thestimulus, and study the process of AChR clustering atpredetermined sites. Thus, this model allows a rela-tively easy dissection of the AChR clustering process.
The beads used in this study are made of polystyrene,which is often used as a substratum for proteinadsorption. However, one can rule out the adsorption-mediated concentration of proteins from the culturemedium onto the bead surface as the cause of AChR

ACh receptor clustering induced by polymer beads 551
clustering, since the same effect was observed in a saltsolution without any exogenously added proteins orserum. Furthermore, if the bead surface is masked byinert molecules, such as BSA or serum, the beads losetheir inductive action, even though they are stillattached to the cell surface. This suggests that a simplemechanical perturbation of the membrane does notactivate the clustering process. A remaining possibilityis that the beads somehow utilize components at the cellsurface to stimulate the cells. These components maybe intrinsic to the cells in culture, or derived fromculture medium. In addition, endogenous factors maypre-exist at the cell surface or be secreted during thetime of bead contact, and since the process of AChRclustering is triggered soon after the beads come intocontact with the cell surface, we favor the formerpossibility.
The possible role of a HBGFIn this study, we have concentrated on the role ofHBGF(s) at the surface of muscle cells in postsynapticinduction for two reasons.
(1) HSPG is a prominent component of the musclesurface (Anderson and Fambrough, 1983; Bayne et al.1984; Anderson et al. 1984; Anderson et al. 1991).Certain forms of this proteoglycan are present in theECM, whereas others exist as integral membraneproteins, e.g. syndecan (Ruoslahti and Yamaguchi,1991). It is not known which forms are present onXenopus muscle cells. However, ultrastructural studieshave shown that there is very little ECM on thesemuscle cells in culture, except at AChR clusters(Weldon and Cohen, 1979; Luther and Peng, 1985).Thus, a major portion of the HSPG in the area awayfrom AChR clusters may be in membrane-bound form,whereas both forms may be present at clusters. Recentstudies have shown that an important function of eitherform of HSPG is to provide binding sites ('low-affinityreceptors') for HBGFs (Rifkin and Moscatelli, 1989;Ruoslahti and Yamaguchi, 1991).
(2) bFGF is a member of the HBGF family. Amongmembers of this family, bFGF binds heparin with thehighest affinity (Burgess and Maciag, 1989). Our recentstudy has shown that beads coated with bFGF caninduce the formation of AChR clusters, and byimmunoblot analysis, both bFGF and its receptor arepresent in Xenopus myotomal muscle in vivo at stagesduring which synaptogenesis occurs (Peng et al. 1991).Previous studies have also shown that bFGF is presentin skeletal muscle in vivo in other species (Joseph-Silverstein et al. 1989; Gonzalez et al. 1990; DiMario etal. 1989). Thus, bFGF is a possible candidate for aHSPG-bound factor available for synaptic induction.
A series of experiments were conducted in this studyto test the involvement of HBGF, in general, andbFGF, in particular, in uncoated bead-induced AChRclustering. First, heparin was found to be inhibitory tothe effect of beads. Recent studies have shown thatbFGF bound to an artificial matrix or to a cellmonolayer can be solubilized by heparin (Flaumenhaftet al. 1990). Thus, this inhibitory effect on AChR
clustering may be due to the a removal of bFGFnormally bound to HSPG at the cell surface. This is alsosupported by our finding that heparin treatmentremoves exogenously applied bFGF. However, thisremoval is incomplete even at the highest concentration(0.5 mg/ml) used in this study. This suggests that somebFGF may be bound to a pool of cell surface bindingsites with much higher affinity. These sites are particu-larly concentrated at AChR hot spots (Fig. 6c-d), wherethe ECM is organized into a basement membrane(Weldon et al. 1981; Luther and Peng, 1985), suggestingthat basement membrane-bound bFGF is resistant toheparin. The incomplete removal of exogenous bFGFmay explain the partial suppression of bead-inducedAChR clustering by heparin (Fig. 3). Previously,Hirano and Kidokoro (1989) showed that heparin orheparan sulfate, at concentrations similar to those usedhere, causes a 50% inhibition in nerve-induced cluster-ing in Xenopus cultures. Thus, heparin essentiallyexerts the same inhibitory effect on AChR clusteringinduced by beads and by nerve.
In contrast to heparin, another polyanionic com-pound, suramin, inhibits AChR clustering completely.With regard to bFGF, the action of suramin appears tobe directed to the receptor-binding domain, rather thanthe heparin-binding domain (Eriksson et al. 1991).Consistent with this notion is our finding that it does notremove exogenous bFGF from the cell surface. Theaction of suramin is not specific to bFGF, as it alsointerferes with interactions of other growth factors withtheir receptors (Betsholtz et al. 1986; Fleming et al.1989; Coffey et al. 1987; Huang and Huang, 1988), aswell as with signalling pathways involved in embryonicinduction mediated by peptide growth factors (Gerhartet al. 1989; Smith, 1989).
A second piece of evidence supporting the role of aHBGF is the finding that exogenous application ofbFGF that results in a large increase in the amount ofbound bFGF to the surface of muscle cells enhances theefficacy of the bead-induced AChR clustering. Thissuggests that the failure of some beads to elicit AChRclustering in control cultures may be due to the fact thatthe endogenous HBGF level at the cell surface isrelatively low and unevenly distributed, as our immuno-fluorescence study has suggested.
Previous studies have shown that the number ofbFGF binding sites in the form of HSPG greatlyexceeds that of the bFGF receptors and most of theexogenously applied bFGF binds to these sites (Mosca-telli, 1988). In fact, bath application of bFGF toXenopus muscle cells by itself has no effect on AChRclustering (Peng et al. 1991), in contrast to the highdegree of cluster induction when beads are applied tobFGF-treated cells. This suggests that the HSPG-boundbFGF at the cell surface may be more effectivelypresented to the bFGF receptor by beads. Recentstudies have suggested that complexing with heparin orHSPG may be essential for bFGF to activate itsreceptors (Rapraeger et al. 1991; Ruoslahti and Yama-guchi, 1991; Yayon et al. 1991). Given that the beadscan elicit AChR clustering at any site on the cell

552 L. P. Baker and others
surface, the distribution of the endogenous moleculesthat can be presented by the beads to trigger thisprocess are likely to exist in a diffuse manner.Consistent with this notion is our finding that endogen-ous bFGF exists in a diffuse pattern and is not visiblyconcentrated at specific sites on the cell surface, e.g. athot spots.
The suppression of bead-induced AChR clustering byan anti-bFGF antiserum (77R, generated in sheep)offers support for the involvement of bFGF in uncoatedbead effects. This antiserum apparently recognizes thereceptor-binding domain of bFGF as opposed to theheparin-binding domain (Dr. M. Klagsbrun, personalcommunication). The inhibitory effect does not appearto be due to serum-coating of the beads, as highconcentrations (up to 500 jUg/ml) of a non-specific sheepantiserum were not inhibitory. The reason why theother two antibodies used in this study were notinhibitory is not known. Both of these antibodies werepartially purified, but the epitope that they recognize isnot known. The inhibitory antiserum (77R), on theother hand, is not purified, and the titer was relativelylow according to our dot blot analysis. However, theinhibitory effects were clear cut. Species differences, ordifferences in antibody affinity for specific epitopes mayexplain these results.
These studies suggest that the stimulation presentedby uncoated beads is mediated by the presentation ofmolecules bound to HSPG at the cell surface. Since thebare polystyrene surface is sticky, one can imagine thatthe beads might adsorb HSPG and its associated growthfactor molecules and at the same time make a tightcontact with the cell surface. By this action, the HBGFis forced into contact with specific membrane receptorsand exerts a local activation. This hypothesis is shownschematically in Fig. 11. Subsequent intracellularamplification may eliminate the necessity for anaccumulation of extracellular signalling molecules atthe site of synaptic differentiation. Consistent with thisnotion is our observation of a relatively diffuse, lowlevel of endogenous bFGF labelling. If the sticky beadsurface is masked by inert molecules, it can no longeradsorb HBGF to present to receptors. This may explainthe lack of effect of beads coated with BSA, or othersmall basic proteins such as cytochrome c or NGF, eventhough these beads still adhere to the cell surface.
Recently, Anderson et al. (1991) showed that native,uncoated polystyrene beads can desorb fluorescentlyconjugated ECM molecules, such as laminin andfibronectin, that are adsorbed on a solid substratum.On the basis of this result, they proposed that thesynaptogenic activity of these beads is due to a focalremoval of certain weakly bound peripheral cell surfacecomponents. While there is yet no experimentalevidence indicating that such a desorptive process perse can serve as a stimulatory signal on inductiveprocesses, it is conceivable that molecules removed bybeads from the ECM can be focally presented to the cellas a positive signal. This interpretation is entirelyconsistent with the data presented in our current work.
Beads coated with polycations or bFGF, on the other
Fig. 11. A model depicting the role of bFGF (or someother HBGF) in mediating the formation of synapticspecializations induced by beads. The adhesive surface ofthe bead can adsorb bFGF bound to HSPG at the cellsurface (A) and present it to cell surface receptors (B).This local signal activates a mechanism to cluster AChRs atthe bead-muscle contact (C).
hand, can directly present effective stimuli for AChRclustering. We think that these coated beads maydirectly activate membrane receptors to signal theinitiation of the clustering process. bFGF-coated beadspresumably activate bFGF receptors. The mechanismof activation by polycation-coated beads is unknown.However, recent studies have suggested that polyca-tions such as polylysine can nonspecifically activatereceptors for several growth factors, including thebFGF receptor (Dauchel et al. 1989; Kuo et al. 1990).Thus, the stimulation by bFGF and polycation-coated,and by uncoated, beads may utilize a common pathway.
The ability of exogenously applied bFGF to increasesignificantly bead-induced AChR clusters suggests thatbFGF is able to synergize with the endogenous stimuluspresented to the cell by the beads. The exact identity ofthis endogenous signal is unknown; however, given theresults of this study, it appears that a HBGF, perhapsbFGF itself, may be involved. The possibility remainsthat other factors secreted by the cells in culture anddeposited on the cell surface, such as agrin (Nitkin et al.1987; Fallon and Gelfman, 1989; Godfrey et al. 1988),are similarly capable of being presented to specific cell-surface receptors by these beads. Whatever the identityof the endogenous molecule, the ability of its localizedpresentation to initiate clustering may shed light on theintracellular mechanisms involved in this process.
The contact and adhesion model of NMJ formationPreviously, Bloch and Pumplin proposed the 'contactand adhesion' model to explain the formation of AChRclusters (Bloch and Pumplin, 1988). This hypothesisdepicts that the muscle cell's contact with the sub-stratum or between synaptic partners initiates a se-quence of events, including the organization of an actin-

ACh receptor clustering induced by polymer beads 553
based cytoskeleton, which leads to the clustering ofAChRs. The occurrence of AChR clusters at close cell-substratum contacts and at bead-muscle contacts isconsistent with this model. Our study has provided amolecular basis for understanding why this adhesiveinteraction can lead to synaptic induction. In theformation of hot spots, a simple substratum-mediatedpresentation of HBGF, similar to the scheme proposedabove for the bead-muscle interaction, may be suf-ficient to trigger the clustering process. However, amore elaborate process has to be invoked for the nerve-muscle interaction, since the cell-cell contact may befundamentally different. The finding that nerve-induced AChR clustering can be partially inhibited byheparin (Hirano and Kidokoro, 1989) suggests thatHBGFs may also play an important role in NMJdevelopment. Recent studies have shown that therelease of HBGF, in particular bFGF, from the HSPGmay involve a proteolytic step (Saksela and Rifkin,1990). Proteases secreted by nerve growth cones, suchas plasminogen activator (Pittman et al. 1989; McGuireand Seeds, 1990), may cleave HSPG at the developingNMJ and release HBGF-HSPG complexes locally toeffect AChR clustering. Indeed, a breakdown of HSPGduring early stages of synaptogenesis in Xenopus hasbeen suggested by a previous study (Anderson, 1986).
The role of tyrosine phosphorylationThe cytoplasmic domain of the bFGF receptor containsa tyrosine kinase sequence (Pasquale and Singer, 1989;Johnson et al. 1990; Musci et al. 1990). The binding ofbFGF to its receptor activates this kinase, whichundergoes autophosphorylation and phosphorylatesother substrate proteins on tyrosine residues. Pre-viously we showed that a tyrosine kinase inhibitorabolishes the formation of AChR clusters induced bybFGF-coated beads (Peng et al. 1991). This resultsuggests the essential role of tyrosine phosphorylationin AChR clustering. The current study has shown thatthe same inhibitor also blocks the clustering effect ofuncoated beads. The substrates for tyrosine phos-phorylation responsible for AChR clustering are notknown. Qu et al. (1990) showed that fi, yand <5subunitsof the AChR undergo tyrosine phosphorylation duringsynaptogenesis. However, this is seen after the AChRsare already clustered. Recently, it has been shown thatagrin increases tyrosine phosphorylation of the/3-subunit with a time course similar to AChR clusteringwhen applied to chick myotubes (Wallace et al. 1991).Thus, tyrosine phosphorylation of both the AChR andother proteins may be involved in cluster formation. Inaddition to the autophosphorylation of the growthfactor receptor itself, phospholipase C-y is an earlysubstrate for a number of growth factor receptor-associated tyrosine kinases, including the bFGF recep-tor (Ullrich and Schlessinger, 1990; Burgess et al. 1990).A recent study has shown that a consequence oftyrosine phosphorylation of phospholipase C-y is arelease of the actin-binding protein profilin from itsphosphoinositide anchor in the plasma membrane(Goldschmidt-Clermont et al. 1991). In addition, there
is mounting evidence for tyrosine phosphorylation ofother cytoskeletal proteins, e.g. spectrin and protein4.1, by activated growth factor receptors (Subrahma-nyam et al. 1991; Akiyama et al. 1986). Thus, tyrosinekinase activity resulting from the activation of growthfactor receptors may lead to a reorganization of thecytoskeleton. In fact, the assembly of an actin-basedcytoskeleton is one of the earliest events that can bedetected at sites of presumptive AChR clusteringinduced by beads (Peng and Phelan, 1984; Rochlin et al.1989). Our recent experiments have shown that anaccumulation of phosphotyrosine-containing proteinsat sites of bead-muscle contact can be detected within15 min of the bead addition and these become foci ofAChR clustering during later stages (Baker and Peng,1991). Thus, localized tyrosine phosphorylation stimu-lated by a synaptogenic signal may play a pivotal role inthe formation of the postsynaptic specialization. Thelocal presentation and subsequent intracellular amplifi-cation of this signal may be involved in the activation ofpostsynaptic development.
We thank Ms. Victoria J. Madden (Department ofPathology, UNC) for her help with electron microscopy. Weare grateful to Synergen, Rhone-Poulanc Rorer and Dr. M.Klasgbrun for their generous gifts of reagents. This investi-gation was supported by NIH grant NS23583 and theMuscular Dystrophy Association.
References
Akiyama, I., Kadowaki, T., Nishida, E., Kadooka, T., Ogawara, H.,Fukami, Y., Sakai, H., Takaku, F. and Kasuga, M. (1986).Substrate specificities of tyrosine-specific protein kinases towardcytoskeletal proteins in vitro. J. Biol. Chem. 261, 14797-14803.
Anderson, M. J. (1986). Nerve-induced remodeling of muscle basallamina during synaptogenesis. J. Cell Biol. 102, 863-877.
Anderson, M. J., Champaneria, S. and Swenarchuk, L. E. (1991).Synaptic differentiation can be evoked by polymer microbeads thatmimic localized pericellular proteolysis by removing proteins fromadjacent surfaces. Develop. Biol. 147, 464-479.
Anderson, M. J., Cohen, M. W. and Zorychta, E. (1977). Effects ofinnervation on the distribution of acetylcholine receptors oncultured muscle cells. J. Physiol. (Lond.) 268, 731-756.
Anderson, M. J. and Fambrough, D. M. (1983). Aggregates ofacetylcholine receptors are associated with plaques of a basallamina heparan sulfate proteoglycan on the surface of skeletalmuscle fibers. J. Cell Biol. 97, 1396-1411.
Anderson, M. J., Klier, F. G. and Tanguay, K. E. (1984).Acetylcholine receptor aggregation parallels the deposition of abasal lamina proteoglycan. J. Cell Biol. 99, 1769-1784.
Baker, L. P. and Peng, H. B. (1991). Tyrosine phosphorylation atacetylcholine receptor clusters induced by a variety of stimuli incultured Xenopus muscle cells. Soc. Neurosci. Abslr. 17, 219.
Bayne, E. K., Anderson, M. J. and Fambrough, D. M. (1984).Extracellular matrix organization in developing muscle: correlationwith acetylcholine receptor aggregates. J. Cell Biol. 99, 1486-1501.
Betsholtz, C , Johnsson, A., Heldin, C.-H. and Westermark, B.(1986). Efficient reversion of simian sarcoma virus-transformationand inhibition of growth factor-induced mitogenesis by suramin.Proc. Nat. Acad. Sci. USA 83, 6440-6444.
Bloch, R. J. and Pumplin, D. W. (1988). Molecular events insynaptogenesis: nerve-muscle adhesion and postsynapticdifferentiation. Am. J. Physiol. 254, C345-C364.
Burgess, W. H., Dionne, C. A., Kaplow, J., Mudd, R., Friesel, R.,Zilberstein, A., Schlessinger, J. and Jaye, M. (1990).Characterization and cDNA cloning of phospholipase C-y, a majorsubstrate for heparin-binding growth factor 1 (acidic fibroblast

554 L. P. Baker and others
growth factor)-activated tyrosine kinase. Mol. Cell. Biol. 10, 4770-4777.
Burgess, W. H. and Maciag, T. (1989). The heparin-binding(fibroblast) growth factor family of proteins. Annu. Rev. Biochem.58, 575-606.
Chen, Q., Baker, L. P. and Peng, H. B. (1990). Mechanism of AChreceptor hot spot formation in cultured muscle cells: role of basicfibroblast growth factor. /. Cell Biol. I l l , 493a.
CofTey, R. J., Leof, E. B., Shipley, G. D. and Moses, H. L. (1987).Suramin binding of growth factor receptor and mitogenicity inAKR-2B cells. J. Cell. Physiol. 132, 143-148.
Dauchel, M. C , Courty, J., Mereau, A. and Barritault, D. (1989).Modulation of mitogenic activity and cellular binding of basicfibroblast growth factor by basic proteins. J. Cell. Biochem. 39,411-420.
DiMario, J., Buffinger, N., Yamada, S. and Strohman, R. C. (1989).Fibroblast growth factor in the extracellular matrix of dystrophic(mdx) mouse muscle. Science 244, 688-690.
Dvir, A., Milner, Y., Chomsky, O., Gilon, C , Gazit, A. and Levitzki,A. (1991). The inhibition of EGF-dependent proliferation ofkeratinocytes by tyrphostin tyrosine kinase blockers. / . Cell Biol.113, 857-865.
Eriksson, A. E., Cousens, L. S., Weaver, L. H. and Matthews, B. W.(1991). Three-dimensional structure of human basic fibroblastgrowth factor. Proc. Nat. Acad. Sci. USA 88, 3441-3445.
Esch, F., Baird, A., Ling, N., Ueno, N., Hill, F., Denoroy, L.,Klepper, R., Gospodarowicz, D., Bohlen, P. and Guillemin, R.(1985). Primary structure of bovine pituitary basic fibroblast growthfactor and comparison with the amino-terminal sequence of bovinebrain acidic FGF. Proc. Nat. Acad. Sci. USA 82, 6507-6511.
Fallon, J. R. and Gelfman, C. E. (1989). Agrin-related molecules areconcentrated at acetylcholine receptor clusters in normal andaneural developing muscle. J. Cell Biol. 108, 1527-1535.
Fischbach, G. D. and Cohen, S. A. (1973). The distribution ofacetylcholine sensitivity over uninnervated and innervated musclefibers grown in cell culture. Develop. Biol. 31, 147-162.
Flaumenhaft, R., Moscatelli, D. and Rifkin, D. B. (1990). Heparinand heparan sulfate increase the radius of diffusion and action ofbasic fibroblast growth factor. J. Cell Biol. I l l , 1651-1659.
Fleming, T. P., Matsui, T., Molloy, C. J., Robbins, K. C. andAaronson, S. A. (1989). Autocrine mechanism for v-sistransformation requires cell surface localization of internallyactivated growth factor receptors. Proc. Nat. Acad. Sci. USA 86,8063-8067.
Friesel, R. and Dawid, I. B. (1991). cDNA cloning and developmentalexpression of fibroblast growth factor receptors from Xenopuslaevis. Mol. Cell. Biol. 11, 2481- 2488.
Gerhart, J., Danilchik, M., Doniach, T., Roberts, S., Rowning, B.and Stewart, R. (1989). Cortical rotation of the Xenopus egg:consequences for the anteroposterior pattern of embryonic dorsaldevelopment. Development 1989 Supplement 37-51.
Godfrey, E. W., Siebenlist, R. E., Wallskog, P. A., Walters, L. M.,Bolender, D. L. and Yorde, D. E. (1988). Basal lamina componentsare concentrated in premuscle masses and at early acetylcholinereceptor clusters in chick embryo hindlimb muscles. Develop. Biol.13, 471-486.
Goldschmidt-Clermont, P. J., Kim, J. W., Machesky, L. M., Rhee, S.G. and Pollard, T. D. (1991). Regulation of phospholipase C-gammal by profilin and tyrosine phosphorylation. Science 251,1231-1233.
Gonzalez, A.-M., Buscaglia, M., Ong, M. and Baird, A. (1990).Distribution of basic fibroblast growth factor in the 18-day rat fetus:localization in the basement membrane of diverse tissues. J. CellBiol. 110, 753-765.
Gospodarowicz, D., Neufeld, G. and Schweigerer, L. (1986).Fibroblast growth factor. Mol. Cell. Endocrinol. 46, 187-204.
Harlow, E. and Lane, D. (1988). Antibodies: A Laboratory Manual,p. 291. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Hirano, Y. and Kidokoro, Y. (1989). Heparin and heparan sulfatepartially inhibit induction of acetylcholine receptor accumulationby nerve in Xenopus culture. J. Neurosci. 9, 1555-1561.
Huang, S. S. and Huang, J. S. (1988). Rapid turnover of the platelet-derived growth factor receptor in ay-transformed cells and reversalby suramin. J. Biol. Chem. 263, 12,608-12,618.
Johnson, D. E., Lee, P. L., Lu, J. and Williams, L. T. (1990). Diverseforms of a receptor for acidic and basic fibroblast growth factors.Mol. Cell. Biol. 10, 4728-4736.
Joseph-Silverstein, J., Consigli, S. A., Lyser, K. M. and Ver Pault, C.(1989). Basic fibroblast growth factor in the chick embryo:immunolocalization to striated muscle cells and their precursors. J.Cell Biol. 108, 2459-2466.
Kandel, J., Bossy-Wetzel, E., Radvanyi, F., Klagsbrun, M.,Folkman, J. and Hanahan, D. (1991). Neovascularization isassociated with a switch to the export of bFGF in the multistepdevelopment of fibrosarcoma. Cell 66, 1095-1104.
Kuo, M.-D., Huang, S. S. and Huang, J. S. (1990). Acidic fibroblastgrowth factor receptor purified from bovine liver is a novel proteintyrosine kinase. / . Biol. Chem. 265, 16455-16463.
Levitzki, A. and Gilon, C. (1991). Tyrphostins as molecular tools andpotential antiproliferative drugs. Trends Pharmacol. Sci. 12, 171-174.
Luther, P. W. and Peng, H. B. (1985). Membrane-relatedspecializations associated with acetylcholine receptor aggregatesinduced by electric fields. J. Cell Biol. 100, 235-244.
Lyall, R. M., Zilberstein, A., Gazit, A., Gilon, C , Levitzki, A. andSchlessinger, J. (1989). Tyrphostins inhibit epidermal growthfactor(EGF)-receptor tyrosine kinase activity in living cells andEGF-stimulated cell proliferation. J. Biol. Chem. 264,14503-14509.
McGuire, P. G. and Seeds, N. W. (1990). Degradation of underlyingextracellular matrix by sensory neurons during neurite outgrowth.Neuron 4, 633-642.
Moscatelli, D. (1988). Metabolism of receptor-bound and matrix-bound basic fibroblast growth factor by bovine capillary endothelialcells. J. Cell Biol. 107, 753-759.
Musci, T. J., Amaya, E. and Kirschner, M. W. (1990). Regulation ofthe fibroblast growth factor receptor in early Xenopus embryos.Proc. Nat. Acad. Sci. USA 87, 8365-8369.
Nitkin, R. M., Smith, M. A., Magill, C , Fallon, J. R., Yao, Y. M.,Wallace, B. G. and McMahan, U. J. (1987). Identification of agrin,a synaptic organizing protein from Torpedo electric organ. J. CellBiol. 105, 2471-2478.
Olwin, B. B. and Hauschka, S. D. (1988). Cell surface fibroblastgrowth factor and epidermal growth factor receptors arepermanently lost during skeletal muscle terminal differentiation inculture. /. Cell Biol. 107, 761-769.
Pasquale, E. B. and Singer, S. J. (1989). Identification of adevelopmentally regulated protein-tyrosine kinase by using anti-phosphophotyrosine antibodies to screen a cDNA expressionlibrary. Proc. Nat. Acad. Sci. USA 86, 5449-5453.
Peng, H. B. (1986). Elimination of preexistent acetylcholine receptorclusters induced by the formation of new clusters in the absence ofnerve. J. Neurosci. 6, 581-589.
Peng, H. B. (1987). Development of the neuromuscular junction intissue culture. CRC Crit. Rev. Anal. Sci. 1, 91-131.
Peng, H. B., Baker, L. P. and Chen, Q. (1991). Induction of synapticdevelopment in cultured muscle cells by basic fibroblast growthfactor. Neuron 6, 237-246.
Peng, H. B. and Cheng, P.-C. (1982). Formation of postsynapticspecializations induced by latex beads in cultured muscle cells. J.Neurosci. 2, 1760-1774.
Peng, H. B. and Nakajima, Y. (1978). Membrane particle aggregatesin innervated and noninnervated cultures of Xenopus embryonicmuscle cells. Proc. Nat. Acad. Sci. USA 75, 500-504.
Peng, H. B. and Phelan, K. A. (1984). Early cytoplasmicspecialization at the presumptive acetylcholine receptor cluster: ameshwork of thin filaments. J. Cell Biol. 99, 344-349.
Pittman, R. N., Ivins, J. K. and Buettner, H. M. (1989). Neuronalplasminogen activators: cell surface binding sites and involvementin neurite outgrowth. J. Neurosci. 9, 4269-4286.
Qu, Z., Moritz, E. and Huganir, R. L. (1990). Regulation of tyrosinephosphorylation of the nicotinic acetylcholine receptor at the ratneuromuscular junction. Neuron 4, 367-378.
Rapraeger, A. C , Krufka, A. and Olwin, B. B. (1991). Requirementof heparan sulfate for bFGF-mediated fibroblast growth andmyoblast differentiation. Science 252, 1705-1708.
Rifkin, D. B. and Moscatelli, D. (1989). Recent developments in thecell biology of basic fibroblast growth factor. / . Cell Biol. 109,1-6.
Rochlin, M. W., Chen, Q., Tobler, M., Turner, C. E., Burridge, K.

ACh receptor clustering induced by polymer beads 555
and Peng, H. B. (1989). The relationship between talin andacetylcholine receptor clusters in Xenopus muscle cells. / . Cell Sci.92, 461-472.
Ruoslahti, E. and Yamaguchi, Y. (1991). Proteoglycans as modulatorsof growth factor activities. Cell 64, 867-869.
Saksela, O. and Rifkin, D. B. (1990). Release of basic fibroblastgrowth factor-heparan sulfate complexes from endothelial cells byplasminogen activator-mediated proteolytic activity. /. Cell Biol.110, 767-775.
Seed, J., Olwin, B. B. and Hauschka, S. D. (1988). Fibroblast growthfactor levels in the whole embryo and limb bud during chickdevelopment. Develop. Biol. 128, 50-57.
Smith, J. C. (1989). Mesoderm induction and mesoderm-inducingfactors in early amphibian development. Development 105, 665-677.
Subrahmanyam, G., Bertics, P. J. and Anderson, R. A. (1991).Phosphorylation of protein 4.1 on tyrosine-418 modulates itsfunction in vitro. Proc. Nat. Acad. Sci. USA 88, 5222-5226.
Swenarchuk, L. E., Champaneria, S. and Anderson, M. J. (1990).Induction of a specialized muscle basal lamina at chimaericsynapses in culture. Development 110, 51-61.
Sytkowski, A. J., Vogel, Z. and Nirenberg, M. W. (1973).Development of acetylcholine receptor clusters on cultured musclecells. Proc. Natl. Acad. Sci. USA 70, 270-274.
Ullrich, A. and Schlessinger, J. (1990). Signal transduction byreceptors with tyrosine kinase activity. Cell 61, 203-212.
Wallace, B. G., Qu, Z. and Huganir, R. L. (1991). Agrin inducesphosphorylation of the nicotinic acetylcholine receptor. Neuron 6,869-878.
Weldon, P. R. and Cohen, M. W. (1979). Development of synapticultrastructure at neuromuscular contacts in an amphibian cellculture system. J. Neurocytol. 8, 239-259.
Weldon, P. R., Moody-Corbett, F. and Cohen, M. W. (1981).Ultrastructure of sites of cholinesterase activity on amphibianembryonic muscle cells cultured without nerve. Develop. Biol. 84,341-350.
Yayon, A., Klagsbrun, M., Esko, J. D., Leder, P. and Ornitz, D. M.(1991). Cell surface, heparin-like molecules are required forbinding of basic fibroblast growth factor to its high affinity receptor.Cell 64, 841-848.
Zhang, J., Cousens, L. S., Barr, P. J. and Sprang, S. R. (1991).Three-dimensional structure of human basic fibroblast growthfactor, a structural homolog of interleukin 1/3. Proc. Nat. Acad. Sci.USA 88, 3446-3450.
(Received 28 January 1992 - Accepted 26 March 1991)
