immunoelectron-microscopic study on the fine structure of substance-p-containing fibers in the taste...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 227:380-392 (1984)
Immunoelectron-Microscopic Study on the Fine Structure of Substance-P-Containing
Fibers in the Taste Buds of the Rat
HIROSHI YAMASAKI, YOSHIYIJKI KUBOTA, HIROSHI TAKAGI, ANI) MASAYA TOHYAMA
Department of Electron Microscopy, Central Laboratory [H.Y.] and Department of Neuroanatomy, Institute of Higher Nervous Activity [Y.K., M.T.], Osaka University
Medical School, Osaka 530 and 2nd Department of Anatomy, Kinki University School of Medicine [H.T.], Osaka 589, Japan
ABSTRACT The fine structure of substance-P-like immunoreactive [SPI] fibers in
the taste buds of the circumvallate papillae of the rat tongue was investi- gated by means of electron microscopy using the unlabeled antibody-enzyme method.
Outside the epithelium, SPI and non-SPI fibers are surrounded by the cytoplasm of Schwann cells. When the SPI fibers enter the epithelium, they immediately lose this cytoplasmic sheath and begin to traverse the taste buds. Though passing through the taste buds, no profiles suggesting clear synaptic contact between SPI fibers and underlying cells are identified. SPI terminals are filled with small synaptic vesicles and contain a few mitochon- dria. No SPI-positive structures are found in nerve endings t.hat make synaptic contact with type I11 cells, the gustatory receptor cells.
Key words: Substance P, taste bud, immunohistochemistry, fine structures
Substance P [SP] was first recognized by von Euler and Guddam in 1931. Since the chemical nature of SP was elucidated by Leeman and her co-workers [Chang et al., '711, numerous immunohistochemical and radioimmunoas- say studies have shown an uneven distribution of SP in the central and peripheral nervous systems [Hokfelt et al., '75a,b, '77a-d, '78a,b; Brownstein et al., '76, '77; Cuello et al., '77, '78; Ben-Ari et al. '77; Chan-Palay and Palay, '77; Chan-Palay, '78; Ijungdahl et al., '78a,b; Inagaki et al., '81, '82; Sakanaka et al., '821.
However, the exact role of SP remains to be elucidated. Several authors have suggested that SP is involved in pri- mary sensory afferent mechanisms [Lembek, '53; Hokfelt et al., '75a,b, '76, '77a-c; Takahashi and Otsuka, '751. Sub- sequent physiological studies supported this hypothesis [Krivoy et al., '63; Krivoy and Zimmerman, '73; Otsuka and Takahashi, '771.
SP fibers have been demonstrated in the tongue [Hokfelt et al., '77b], particularly in the taste buds [Lundberg et al., '791. Furthermore, we have shown that SP fibers in the foliate and circumvallate papillae [both in the taste buds and epithelium outside the taste buds] are supplied by the glossopharyngeal nerve, those in the taste buds of the fun- giform papillae by the chorda tympani, and those in the
0 1984 ALAN It . LISS, INC.
epithelium of the fungiform papillae outside the taste buds by the third division of the trigeminal nerve by using ex- perimental procedures [Nishimoto et al., '821.
These findings strongly suggest that SP is involved in taste transmission. In order to clarify the exact role of SP in the taste buds, it is important to eludicate the fine structure of SP fibers, including their pre- or postsynaptic structures. Therefore, we investigated the fine structure of SP fibers in the taste buds together with those in the epithelium outside the taste buds of the rat tongue by electron microscopy using the unlabeled antibody-enzyme method [Sternberger et al., '701.
MATERIALS AND METHODS Specificity of the antiserum
The antiserum against SP was obtained after immuniza- tion of a rabbit with SP-bovine serum albumin [BSAl con- jugate. The specificity of this antiserum was tested by radioimmunoassay. The radioimmunoassay system showed less than 0.01% crossreactivity with the structurally simi- lar peptides eledoisin and physalemin, and no crossreactiv-
Accepted March 27, 1984.

SUBSTANCE P IN RAT TASTE BUDS
ity with other peptides such as somatostatin, neurotensin, enkephalins, endorphins, vasoactive intestinal polypeptide, or cholecystokinin-8. The specificity was also examined by an absorption test. The structures stained with SP anti- serum were not seen in the sections stained in the presence of SP I0.08 pg/ml]. Although the structures stained with SP antiserum should be correctly described as SP-like immu- noreactive, the simpler term "SPY was used in this study.
381
Procedure of immunohistochemistry Five albino rats, weighing about 100-150 gm, were used.
The procedures followed the methods developed by Somogyi and Takagi "821. In brief, the animals were perfused via the ascending aorta with 50 ml of ice-cold saline followed by 300 ml of picric acid [0.2%]-paraformaldehyde [4%]-glu- taraldehyde [0.05%] fixative under sodium pentobarbital anesthesia [40 mgkg, i.p.1. Small blocks of the tongue in- cluding the circumvallate papillae were postfixed in this fixative for 1-2 hours at 4°C. The blocks were then washed repeatedly in 0.1 M sodium phosphate buffer, pH 7.4, fol- lowed by the same buffer containing 10% sucrose and then 20% sucrose, repeatedly until the blocks sank. They were then frozen in liquid nitrogen and thawed in 0.1 M phos- phate buffer, pH 7.4. Sections of 70-pm thickness were cut on a Vibratome and immersed in 0.1 M phosphate buffer [pH 7.41 overnight a t 4°C.
The staining proceeded as follows: Incubation in 20% goat serum for 1 hour at 15"C, wash for 1 hour with phosphate- buffered saline [PBS] at 15"C, incubation in primary anti- serum overnight at 4°C at a dilution of 1:1,000 [diluted with PBS], wash three times with PBS [20 minutes each] at 15"C, incubation in goat antirabbit IgG [DAKO] at a dilution of 150 for 2 hours at 15°C [diluted with PBS], and wash three times with PBS at 15°C [20 minutes each]. The reaction products were visualized by the diaminobenzidine [DAB] reaction.
After immunohistochemistry, the sections were postfixed for 1-2 hours in Os04 in 0.1 M phosphate buffer [pH 7.41 and then dehydrated. To enhance contrast for electron mi- croscopy, the sections were stained with l % uranyl acetate at the 70% alcohol dehydration state. The sections were flat-embedded on slides in Epon 812 according to Somogyi et al. "821.
Immunohistochemically stained structures were selected under light microscopy for subsequent electron microscopic analysis of their fine structure using the correlation method described by Somogyi and Takagi "821 and Somogyi et a1 "821. Serial ultrathin sections were studied with lead stain- ing on an Hitachi HU 12 electron microscope.
RESULTS As shown in Figure 1, a number of SPI fibers were found
in the circumvallate papillae. They were mainly situated in the connective tissue just under the epithelium, where they formed a dense fiber band. The fiber band often sends branches into the epithelium, both into and outside the taste buds. About 40% of the taste buds situated in the circumvallate papilla contain one to five SPI fibers.
SPI fibers were easily identified under electron micro- scope even under low magnification because of the strong electron-dense precipitations [Figs. 2,3]. They were local- ized exclusively in nerve fibers. Immunoreactive end prod- ucts were found diffusely throughout the axoplasm, most of which were associated with the surface of small, electron- lucent vesicles.
Fig. 1. Lightfield photomicrograph showing SPI fibers [arrows] in the taste buds [TB] and epithelium (EP, arrowhead1 outside the taste buds. Note numerous SPI fibers in the lamina propria [LP]. ~ 5 7 0 .
In the lamina propria just beneath the epithelium, SPI fibers were surrounded by the cytoplasm of Schwann cells together with non-SPI fibers [Fig. 4A,B]. The SPI fibers identified in these areas were unmyelinated and their di- ameters varied widely [from 0.15 to 1.0 pm]. About 30% of the fibers observed were SPI fibers. The fiber bundles pen- etrated the basal lamina and entered the taste buds.
SPI fibers lost the cytoplasmic sheath of Schwann cells [Fig. 5 , arrow] when they entered the taste buds. SPI fibers ascended in the taste buds as intergemmal fibers and ap- proached the outer taste pores [Figs. 6, 7D]. The fibers in the taste buds were filled with small vesicles and often contained mitochondria [Figs. 4-10]. As shown in Figure 6, SPI fibers tended to become larger as they approached the outer taste pores. About 30% of nerve fibers identified in the taste buds were SPI.
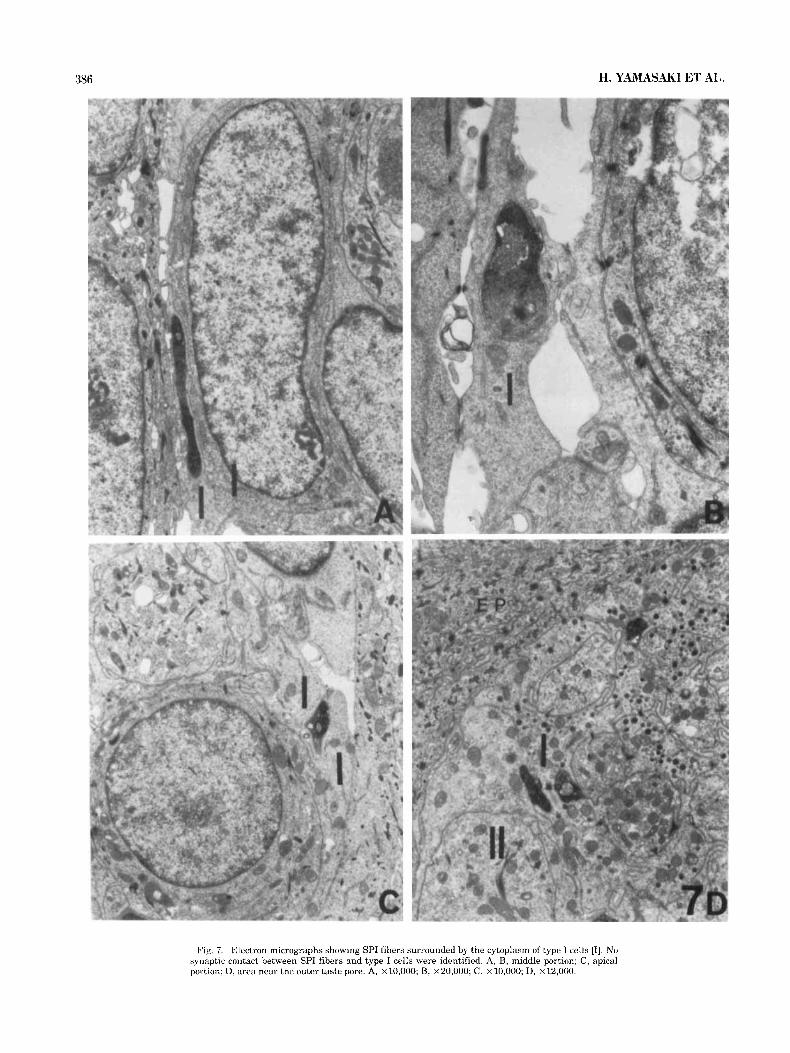
Figure 7 shows SPI fibers surrounded by the cytoplasm of the type I cells. Figure 8 shows SPI fibers surrounded partly by the cytoplasm of type I cells and partly by type I1 cells. No clear profiles that suggest special contact with type I and I1 cells were obtained. Figure 9 shows two nerve end- ings situated close to the cytoplasm of type I11 cells at the midportion of the taste buds; one is labeled by immunoreac- tive products and contains numerous synaptic vesicles [ax- onlike profile, Fig. 9A]; the other is devoid of immunoreactive products and shows dendritelike profiles [Fig. 9B]. Synaptic vesicles in the type I11 cells approach the contact membrane and presynaptic specialization in the cytoplasm of the type I11 cell which is in contact with the nerve ending showing dendritelike profiles is evident. No
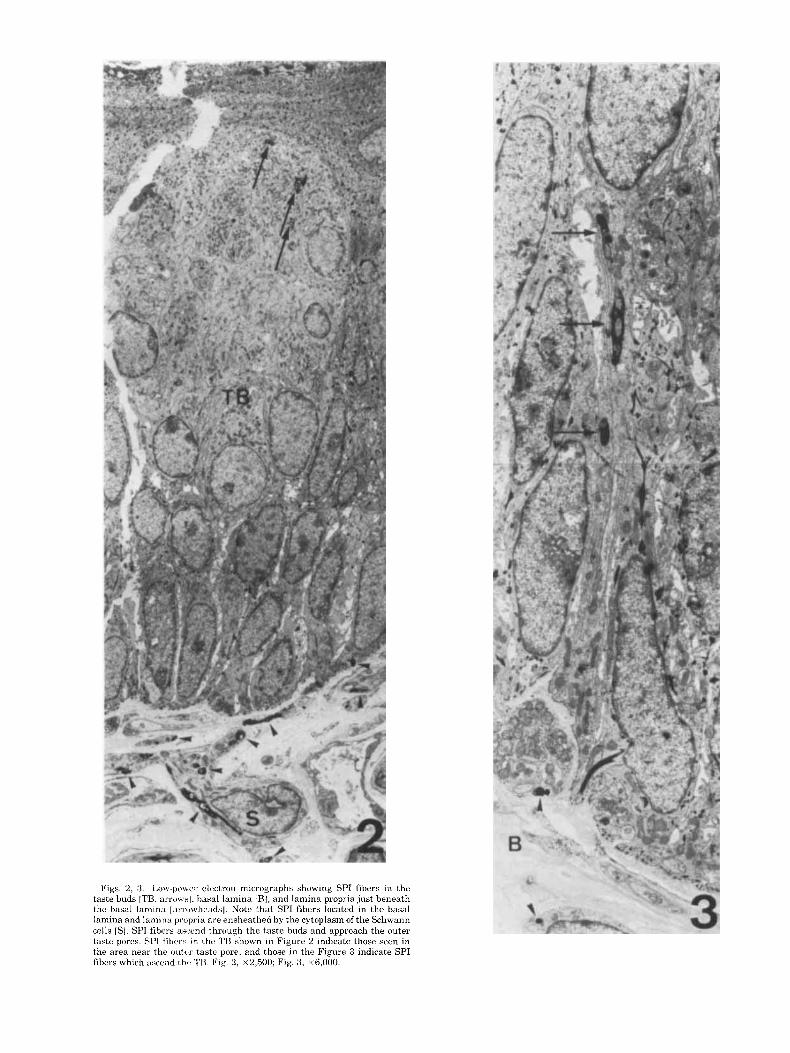
Figs. 2, 3. L o w p o ~ e r electron micrographs showing SPI fibers in the taste buds [TB. arro~.<l , hasal lamina [HI, and lamina propria just beneath the basal lamina [armwheadsl. Note that SPI fibers located in the basal lamina and lamina propria are ensheathed by the cytoplasm of the Schwann cells [S] . SPI fibers ascend through the taste buds and approach the outer taste pores. SPI fibers in the TB shown in Figure 2 indicate those seen in the area near the (1utc.r taste pore, and those in the Figure 3 indicate SPI fibers which ascend tho TB. Fig. 2, ~ 2 , 5 0 0 ; Fig. 3 , ~6,000.

SUBSTANCE P IN RAT TASTE BUDS
Fig. 4. Electron micrographs showing SPI fibers located in the lamina propria just beneath the basal lamina [B]. Numerous SPI fibers were ensheathed by the cytoplasm of the Schwann cell LS] together with nonimmunoreactive nerve fibers. A, x 12,000; B, ~20 ,000 .
383
384 H. YAMASAKI ET AL.
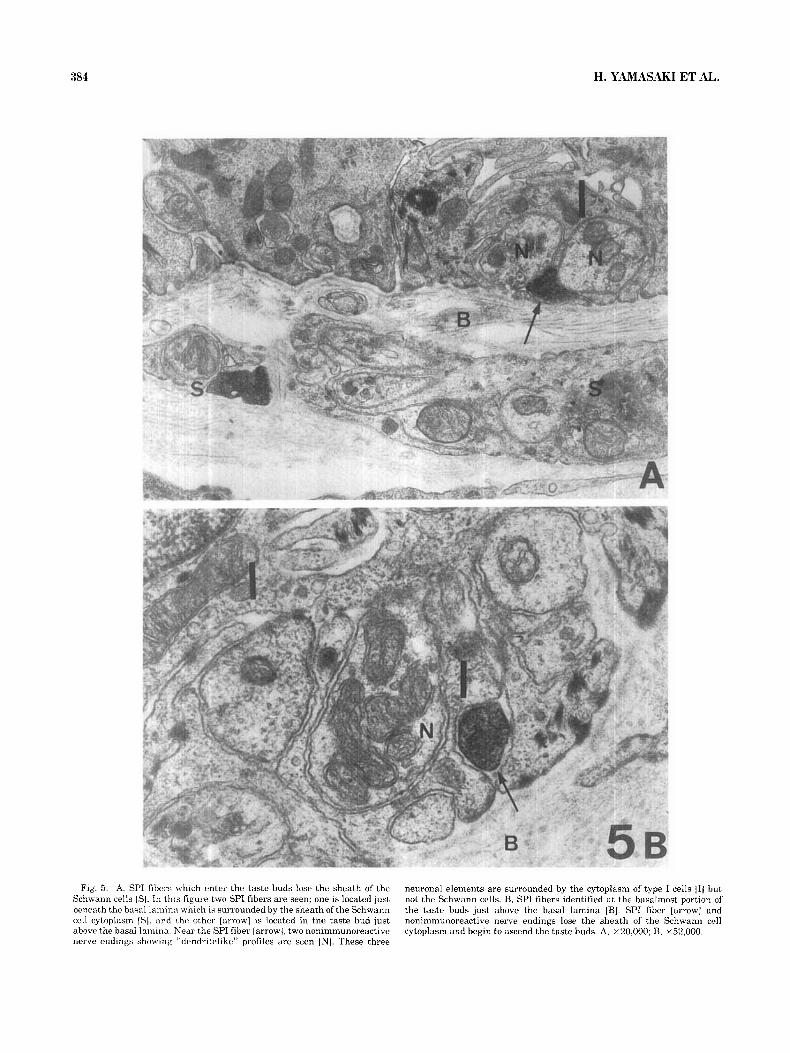
Fig. 5 . A. SPI fihers which enter the taste buds lose the sheath of the Schwann cells IS]. In this figure two SPI fibers are seen; one is located just beneath the basal lamin:] which IS surrounded by the sheath of the Schwann cell cytoplasm ISI, and rht, other [arrow] is located in the taste bud just above the basal lamina Near the SPI fiber [arrow], two nonimmunoreactive nerve endings showing “dendritelike” profiles are seen [NJ. These three
neuronal elements are surrounded by the cytoplasm of type I cells 111 but not the Schwann cells. B. SPI fibers identified at the basalmost portion of the taste buds just above the basal lamina [BI. SPI fiber [arrow] and nonimmunoreactive nerve endings lose the sheath of the Schwanri cell cytoplasm and begin to ascend the taste buds. A, ~20,000; B, ~52,000.

SUBSTANCE P IN RAT TASTE BUDS 385
Fig. 6. Electron micrograph showing SPI fibers identified near the outer taste pore region. The fibers are moderately larger than those seen in other regions of the taste buds (TBI. EP, epithelium outside the TB. ~ 6 , 6 0 0
specialization is seen in the cytoplasm of type I11 cell lo- cated adjacent to SPI fibers. Although 12 synaptic contacts were observed between type I11 cell and nerve ending, the postsynaptic elements with which type I11 formed synapses are dendritelike profile and devoid of immunoreactive products.
These findings indicate that nerve endings [dendritelike profiles] which receive gustatory information and convey it to the solitary nucleus do not contain SP. In addition, the taste buds contained nonlabeled nerve fibers that showed axonlike profiles [Fig. 101. These fibers rarely made contact with SPI fibers. However, no aggregation of the vesicles was found in the contact membrane; these contacts seemed to belong to the zonula adherens.
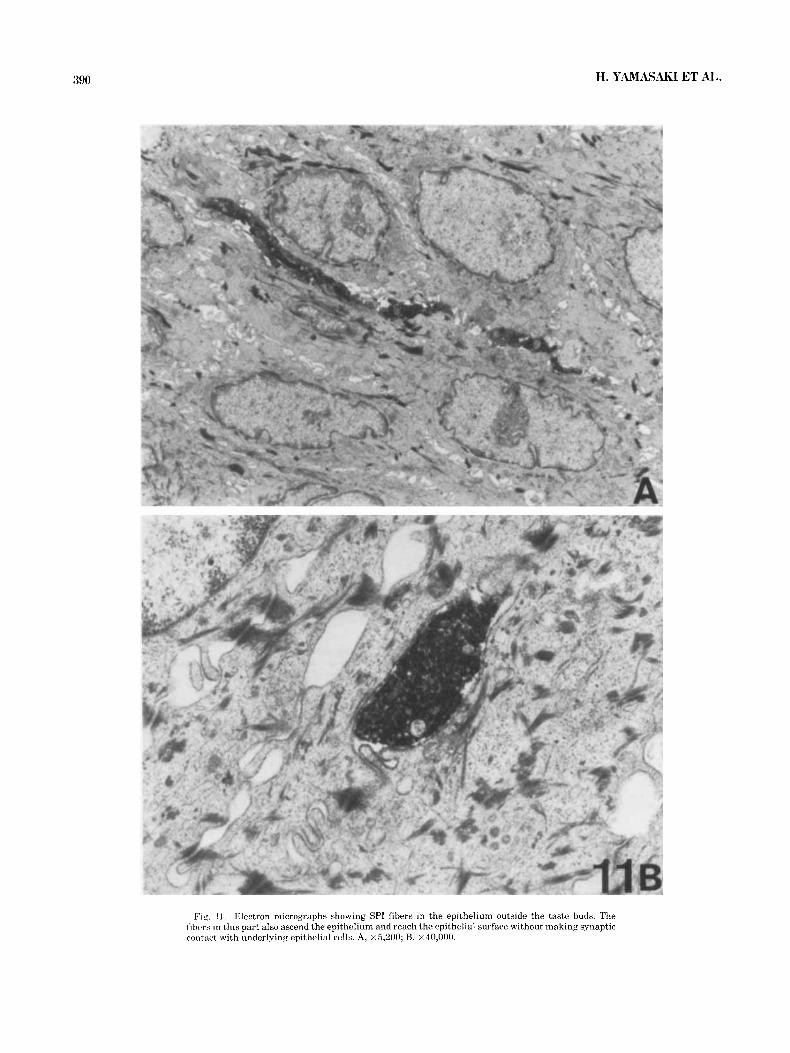
In the epithelium outside the taste buds, the general features of SPI fibers were similar to those seen in the taste buds [Fig. 111. The fibers from the bundles located in the lamina propria just beneath the epithelium entered the epithelium and ascended as intergemmal fibers to reach the epithelium [Fig. llA1. The SPI fibers contained numer- ous synaptic vesicles and a few mitochondria. They passed through the epithelium [Fig. 11A] without making clear synaptic contact with the underlying epithelial cells [Fig. 11B] .
A summary of the findings is schematically presented in Figure 12.
DISCUSSION The existence of SPI fibers in the taste buds was first
reported in the cat by Lundberg et al. "791. We have also
confirmed their presence in the taste buds of the rat and further eludicated their origins by using experimental im- munohistochemical procedures [Nishimoto et al., '821. In our previous study, SPI located in the taste buds and epi- thelium outside the taste buds in the circumvallate papillae were found to be supplied by the glossopharyngeal nerve, since unilateral neurotomy of the peripheral branch of this nerve caused a significant decrease of SPI fibers, and bilat- eral neurotomy resulted in a disappearance of SPI fibers in this papilla. On the other hand, Nagy et al. "€321 failed to demonstrate the presence of SPI fibers in the taste buds of the rat circumvallate and fungiform papillae. The discrep- ancies between their findings and ours are attributable to the differences in the antisera used.
Several investigators have suggested that nerve endings which convey primarily gustatory information from taste bud to the medulla oblongata make synaptic contact with type I11 cells, since these cells have been classified as gus- tatory receptors and are involved in the transmission of taste stimuli [Murray, '73; Murray et al., '69; Olivieri- Sangiacomo, '70; Fujimoto and Murray, '69; Takeda, '76; Takeda and Hoshimo, '751. This concept is based upon the following findings: [ l] type III cells are the only ones that make synaptic contact with the nerve endings, [2] nerve endings in contact with type I11 cells show dendritelike profiles, and [3] synaptic vesicles near the contact mem- brane and presynaptic specialization were identified in the cytoplasm of the type I11 cells. If SPI fibers located in the taste buds relate primarily to conveying the gustatory in- formation from taste buds to the medulla oblongata, SPI
386 H. YAMASAKI ET AL.
Fig. 7. Electron micrographs showing SPI fibers surrounded by the cytoplasm of type 1 cells [IJ. No synaptic contact between SPI fibers and type I cells were identified. A, B, middle portion; C, apical portion: D, area near the outer taste pore. A, ~10,000; B, ~20,000; C , ~10,000; D, ~12,000.

SUBSTANCE P IN RAT TASTE BUDS 387
Fig. 8. Electron micrographs showing SPI fibers which are partly surrounded by the cytoplasm of type I cells [I] and partly by type I1 cells [III. No synaptic contact between the fibers and these cells are identified. A, ~26,000; B, ~23,000.
must be localized in the nerve endings that make synaptic contact with type I11 cells. However, as shown in this study [see Fig. 9B], the nerve endings making synaptic contact with type I11 cells were not labeled by SP. In addition, although these nerve endings showed dendritelike profiles, SPI fibers showed axon-like profiles. These findings strongly suggest that ganglion cells which conduct primarily gusta- tory information from the taste bud to the medulla ob- longata do not contain SP, and even if they do, their SP is preferentially conveyed centrally to the medulla oblongata, not peripherally. In any case, these findings suggest that SP fibers in the taste buds are not fibers of the ganglion cells that conduct primarily gustatory information to the central nervous system.
The present study demonstrates that SPI fibers in the taste buds as well as those in the epithelium outside the taste buds are filled with synaptic vesicles. Previously, we showed that SP in the epithelium of the tongue is trans- ported from sensory ganglia [Nishimoto et al., '821, for neurotomy of the peripheral branches resulted in disap- pearance of SPI from the tongue. Thus, SP may influence the taste buds or epithelium outside the taste buds.
Little is known of the function of SPI in the taste buds. The present study suggests that SPI fibers in the taste buds
as well as those in the epithelium do not form synaptic contact with the underlying cells, although it is possible that a few populations of SPI fibers contain synaptic spe- cialization. The SPI which wat3 released from SPI fibers may diffusely affect underlying cells. Accordingly, if SPI influences type I11 cells or nonimmunoreactive fibers which make synaptic contact with type I11 cells, it is probable that SPI also has some effect on gustatory transmission, even though this effect is not primary. Similarly, if SPI fibers located in the epithelium outside the taste buds affect the nerve endings which receive sensory stimulation, SPI fibers located in the epithelium outside the taste buds also second- arily are related to the transmission of pain.
On the other hand, it has been reported that neuronal elements are necessary for the maintenance of the normal function of the taste buds, because taste buds disappear after neurotomy and reappear when the neuronal elements regenerate [Farbman, '69; Fujimoto and Murray, '70; Guth, '58; Olmsted, '20; State and Bowden, '74; Whiteside, '271. Toray "271 speculated that nerve endings in the taste buds secrete substances essential for maintaining the taste buds or promoting cell differentiation. As shown in this study, SP terminals which are filled with synaptic vesicles pass through the taste buds and reach the outer pore. Thus, SP

388 H. YAMASAKI ET AL.
Fig. 9. A. Electron niirrokTaph showing SPI fibers which show close apposition to type 111 llIl1 and type I1 cells. No special contact is identified between the fiber and type 111 or type I1 cells. B. Electron micrograph showing nonimmunoreattivc, nerve ending “1 which makes synaptic con- tact with type I11 cells [III]. This nerve ending exhibits a “dendritelike”
profile, while in the SPI fibers shown in Fig. 9A numerous synaptic vesicles are concentrated laxonlike profile]. Since synaptic vesicles in the cytoplasm are located near the contact membrane [arrows] and presynaptic special- izations are seen, this nonimmunoreactive nerve ending receives gustatory informatinn. A, ~20 ,000; B, x16,600.

SUBSTANCE P IN RAT TASTE BUDS
Fig. 10. In addition to SPI fibers in the taste buds, nerve endings which show axonlike profiles [devoid of immunoreaction] are observed [N]. No clear features that suggest axoaxonic contact between SPI terminals and nonimmunoreactive nerve endings are identified. Arrow indicates the zonu- la adherens between SPI fibers and nonimmunoreactive axonlike nerve ending "1, because no clear aggregation of synaptic vesicles is seen near the contact membrane. ~20,000.
389
390 H. Y.4MBSA4KI ET AL.
Fig. 11. Electron micrographs showing SPI fibers in the epithelium outside the taste buds. The fihcrs in this part also ascend the epithelium and reach the epithelial surface without making synaptic contact with underlying epithelial cells. A, ~ 5 . 2 0 0 ; B, X40,OOO.

SUBSTANCE P IN RAT TASTE BUDS 391
T B
I- . -_-
Fig. 12. Schematic representation summarizing the findings. Nerve end- ings that conduct primarily gustatory information and form synaptic con- tact with a type 111 cell [111] do not contain SP. SP fibers that enter the taste buds [TB] or epithelium [EP] outside the taste buds traverse these regions
released from the terminals may be involved in the main- tenance of the taste buds as a trophic factor, though direct evidence in support of this hypothesis is lacking.
Furthermore, we demonstrated the presence of nerve end- ings that show axonlike profiles but do not contain SPI. These findings suggest the presence of nerve fibers which exert an action in the taste bud in addition to SPI fibers; this indicates very complicated interactions among the nerve fibers that control the general level of sensitivity of the taste buds.
LITERATURE CITED Ben-Ari, Y., G. Le Gal Salle, and I. Kanazawa [1977] Regional distribution
of substance P within the amygdaloid complex and bed nucleus of the stria terminalis. Neurosci. Lett. 4:299-302
Brownstein, M.J., E.A. Mroz, J.S. Kizer, M. Palkovits, and S.E. Leeman [1976] Regional distribution of substance P i n the brain of the rat. Brain Res. 116:299-305.
Brownstein, M.J., E.A. Mroz, M.L. Tappaz, and S. E. Leeman [1977] On the origins of substance P and glutamic acid decarboxylase [GAD] in the substantia nigra. Brain Res. 135~315-323.
Chan-Palay, V. [1978] The parabigeminal nucleus. 11. Identification and interactions of catecholamine, indoleamine axons, and substance P im- munoreactive cells in the neurophil. J. Neurocytol. 7t419-442.
Chan-Palay, V., and S.L. Palay [1977] Ultrastructural identification of sub- stance P cells and their processes in rat sensory ganglia and their terminals in the spinal cord by immunocytochemistry. Proc. Natl. Acad. Sci. USA 74:4050-4054.
Chang, M.M., S.E. Leeman, and H.D. Niall [I9711 Amino acid sequence of substance P. Nature New Biol. 232%-87.
Cuello, A.C., T. Jessel, I. Kanazawa, and L.L. Iversen [1977] Substance P Localization in synaptic vesicles in rat central nervous system. J. Neu- rochem. 29t747-751.
. -- .
--_ 12 to reach the surface without making synaptic contact with the underlying taste buds cells such as type I [I], type I1 [II], and type I11 [111] or epithelial cells.
Cuello, A.C., and I. Kanazawa 119781 The distribution of substance P im- munoreactive fibers in the rat central nervous system. J. Comp. Neurol. 178~129-156.
Farbman, A.I. [1969] Fine structure of degenerating taste buds after dener- vation. J. Embryol. Exp. Morphol. 22t55-68.
Fujimoto, S., and R.G. Murray [1970J Fine structure of degeneration and regeneration in denervated rabbit vallate taste buds. Anat. Rec. 168~393- 414.
Guth, L. [I9581 Taste buds on the cat's circumvallate papilla after reinner- vation by glossopharyngeal vagal and hypoglossal nerves. Anat. Rec. 13Ot25-37.
Hokfelt, T., J.-0. Kellerth, G. Nilsson, and B. Pernow [1975a] Substance P localization in the central nervous system and in some primary sensory neurons. Science 190~889-890.
Hokfelt, T., J.-0. Kellerth, G. Nilsson, and B. Pernow [1975b] Experimental immunohistochemical studies on the localization and distribution of substance P in cat primary sensory neurons. Brain Res. 100:235-252.
Hokfelt, T., R.P. Elde, 0. Johansson, R. Luft, G. Nilsson, and A. Arimura 119761 Immunohistochemical evidence for separate populations of so- matostatin-containing and substance P-containing primary afferent neurons. Neuroscience 1~131-136.
Hokfelt, T., R.P. Elde, 0. Johansson, L O . Kellerth, A. Ljungdahl, G. Nils- son, B. Pernow, and L. Terenius (1977al Substance P and enkephalin: Distribution in the nervous system as revealed with immmunohisto- chemistry. In C. Ransouco-Thomas Led]: Proc. Collegium Internationale Neuropharmacologia, pp. 477-487.
Hokfelt, T., 0. Johansson, J.-0. Kellerth, A. Ljungdahl, G. Nilsson, A. Nygards, and B. Pernow [1977b] Immunohistochemical distribution of substance P. In U.S. von Euler and B. Pernow [eds]: Substance P. New York: Raven Press, Nobel Symposium 37, pp. 117-145.
Hokfelt, T., J.-0. Kellerth, A. Ljungdahl, G. Nilsson, A. Nygrds, and B. Pernow 11977~1 Immunohistochemical localization of substance P in the central and peripheral nervous system. In J. Barchas, E. Costa, and E. Usdin [eds]: Neuroregulators and Hypotheses of Psychiatric Disorders. Oxford: University Press, pp. 290-310.

392 H. YAMASAKI ET AL.
Hakfelt, T., A. Ljungdahl, I,. Terenius, R.P. Elde, and G. Nilsson [1977d] lmmunohistochemical analysis of peptide pathways possibly related to
Enkephalin and substance P. Proc. Natl. Acad. Sci. USA 74:3081-3085.
Inagaki, S., M. Sakanaka, S. Shiosaka, E. Senba, H. Takagi, K. Takatsuki, Y. Kawai, T. Matsuzaki, H. Iida, Y. Hara, and M. Tohyama 119821 Experimental and immunohistochemical studies on the cerebellar sub- stance P of the rat: Localization, postnatal ontogeny and ways of entry to the cerebellum. Neuroscience 7251-277.
Inagaki, S., M. Sakanakzi, S. Shiosaka. E. Senba, K. Takastuki, H. Takagi, Y. Kawai, H. Minagawa. and M. Tohyama 119811 Ontogeny of substance P-containing neuron system of the rat: Immunohistochemical analysis. I. Forebrain and upper brain stem. Neuroscience 7:639-645.
Krivoy, W.A., M. Lane, and D.C. Kroeger [1963j The action of certain polypeptides of the synaptic transmission. In E.G. Erdos Led]: Structure and Function of Biologically Active Peptides Bradykinin, Kallidin and Cogeners. Ann. N Y. Acad. Sci., Val. 104. New York: The Academy, pp. 312-329.
Krjvoy, W., and E. Zimmerman 119731 A possible role of polypeptides in synaptic transmission. In H.C. Sabelli [edl: Chemical Modulation of Brain Function Ntw York: Raven Press, pp. 111-121.
Leeman, S.E.. and E.A. Mroz 119741 Substance P. Life Sci. 15.9033-2044. Lembeck, F. 11953) Ztir Frange der zentralen Ubertragung afferenter Im-
pulse. 111. Das Vorkoinmen und die Bedeuting der Substance P in den dorsalen Wurzeln des Ruckenmarks. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 219: 197-213.
Ljungdahl. A., T. Hokfdt. and G. Nilsson [1978a] Distribution of substance P-like imrnunoreactivity on the central nervous system of the rat. 11. Light microscopic localization in relation to catecholamine-containing neurons. Neuroscience 3.945-976.
Lundberg, J.M., T. Hokfelt, A. Anggrd, B. Pernow, and P. Emson 119791 Immunohistochemical evidence for the substance P immunoreactive fibers in the task, huds of the cat. Acta Physiol. Scand. 107:389-391.
Murray, R.G. 119731 The ultrastructure of taste buds. In I. Friedmann led]: The Ultrastructure of Sensory Organs. North Holland: American Elsev- ier, pp. 1-81.
Murray, R.G., A. Murrxy, and S. Fujimoto 119691 Fine structure of gustatory cells in rabbit tastc huds. J. Ultrastruct. Res. 27:444-461.
Nagy, J.I., M Goedcrt. S.P. Hunt, and A. Bond [1982] The nature of the substance P-containing nerve fibers in taste papillae of the rat tongue. Neuroscience 7:3137-3151
Nishimoto, T.. M. Akai. S. Inagaki. S. Shiosaka, Y. Shimizu, K. Yamanoto, E. Senba, M. Sakanaka. K. Takatsuki, Y. Hara, H. Takagi, T. Matsu-
zaki, Y. Kawai, and M. Tohyama (19821 On the distribution and origins of substance P in the papillae of the rat tongue: An experimental and immunohistochemical study. J. Camp. Neurol. 207:85-92.
Olivieri-Sangiacomo, C. 119701 Neurosecretory cell types in normal taste buds. Experientia 26.289-290.
Olmsted, J.M.D. [1920] The results of cutting the seventh cranial nerve in Amerius nebulosus. J. Exp. Zool. 31:369-403.
Sakanaka, M., S. Inagaki, S. Shiosaka, E. Senba, H. Takagi, K. Takatsuki, Y. Kawai, H. Iida, Y. Hara, and M. Tohyama [1982] Ontogeny of sub- stance P-containing neuron system of the rat: Immunohistochemical analysis. 11. Lower brain stem. Neuroscience 7:1097-1126.
Somogyi, P., J.V. Priestley, A.C. Cuello, A.D. Smith, and J.P. Bolam 119821 Synaptic connections of substance P immunoreactive terminals in the substantia nigra of the rat: A correlated light and electron microscopic study. Cell Tissue Res. 223:469-486.
Somogyi, P. and H. Takagi 119821 A note on the use of picric acid-paraform- aldehyde-glutaraldehyde fixative for correlated light and electron mi- croscopic immunocytochemistry. Neuroscience 7: 1779-1784.
State, F.A., and R.E.M. Bowden 119741 The effect of transection of the glossopharyngeal nerve upon the structure, cholinesterase activity and innervation of taste buds in rabbits. J. Anat. 118:77-100.
Sternberger, L.A., P.H. Hardy, J.J. Curculis, and H.G. Meyer [1970] The unlabeled antibody-enzyme method of immunocytochemistry. Prepara- tion and properties of soluble antigen-antibody complex [horseradish peroxidase-antihorseradish peroxidase] and its use in identification of spirochetes. J. Histochem. Cytochem. 18:315-333.
Takahashi, T., and M. Otsuka [1975] Regional distribution of substance P in the spinal cord and nerve roots of the cat and the effect of dorsal root section. Brain Res. 87:l-11.
Takeda, M. 119761 An electron microscopic study on the innervation in the taste buds of the mouse circumvallate papillae. Arch. Histol. Jpn. 39:257- 269.
Takeda, M., and T. Hoshimo (19751 Fine structure of taste buds in the rat. Arch. Histol. Jpn. 37:395-413.
Torry, T.W. [19271 The regeneration of the gustatory apparatus in the rat. J. Camp. Neurol. 40:33-45.
Von Euler, US. , and J.H. Guddam [1931] An identified depressor substance in certain tissue extracts. J. Physiol. [Lond.l 72:74-87.
Whiteside, B. (19271 The regeneration of the gustatory apparatus in the rat. J. Camp. Neurol. 40:33-45.