il-12p40-dependent agonistic effects on the … · il-12p40-dependent agonistic effects on the...
TRANSCRIPT

of September 17, 2018.This information is current as
EnteritidisSalmonellaAdaptive Immunity Against
Development of Protective Innate and IL-12p40-Dependent Agonistic Effects on the
Schröder, Nicole Schütze and Gottfried AlberJörg Lehmann, Steffi Bellmann, Christoph Werner, Regina
http://www.jimmunol.org/content/167/9/5304doi: 10.4049/jimmunol.167.9.5304
2001; 167:5304-5315; ;J Immunol
Referenceshttp://www.jimmunol.org/content/167/9/5304.full#ref-list-1
, 25 of which you can access for free at: cites 42 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

IL-12p40-Dependent Agonistic Effects on the Development ofProtective Innate and Adaptive Immunity Against SalmonellaEnteritidis 1
Jorg Lehmann,* Steffi Bellmann,2† Christoph Werner,* Regina Schroder,* Nicole Schutze,* andGottfried Alber 3*
To study a potential IL-12p40-dependent but IL-12p75-independent agonistic activity regulating the immune response againstSalmonella Enteritidis, the course of infection in IL-12p35-deficient mice (IL-12p35�/�, capable of producing IL-12p40) wascompared with that of IL-12p40�/� mice. Mice lacking IL-12p40 revealed a higher mortality rate and higher bacterial organburden than mice capable of producing IL-12p40. This phenotype was found in both genetically susceptible (BALB/c,Itys) andresistant mice (129Sv/Ev,Ityr) indicating Ity-independent mechanisms. The more effective control of bacteria in the IL-12p35�/�
mice was associated with elevated serum IFN-� and TNF-� levels. In contrast, IL-12p40�/� mice showed reduced IFN-� pro-duction, which was associated with significantly elevated serum IgE levels. Early during infection (days 3 and 4 postinfection), aswell as late (day 20 postinfection), the number of infected phagocytes was strongly increased in the absence of IL-12p40 indicatingimpaired bactericidal activity when IL-12p40 was missing. Liver histopathology revealed a decreased number of mononucleargranulomas in IL-12p40�/� mice. Depletion of CD4� or CD8� T lymphocytes in vivo suggested that both T cell subpopulationscontribute to the IL-12p40-dependent protective functions. Analysis of IL-12p40 vs IL-23p19 mRNA expression revealed anup-regulation of only IL-12p40 mRNA during Salmonella infection. Together these data indicate that IL-12p40 can induce pro-tective mechanisms during both the innate and the adaptive type 1 immune response inSalmonella infection. This novel activityof IL-12p40 complements the well described dominant and essential role of IL-12p75 in protective immunity toSalmonellainfection. The Journal of Immunology, 2001, 167: 5304–5315.
T he genusSalmonella is the causing agent of a variety ofinfections in humans, ranging from acute self-limitinggastroenteritis to systemic typhoid fever (1–3). In ty-
phoid,Salmonella Typhi spread systemically to deeper tissues andvisceral organs (2).Salmonella Typhimurium has proven to be auseful model for studying invasive typhoid-like disease, as somestrains are able to cause systemic salmonellosis following oral in-fection in mice (1). Recently, we established an infection modelbased on i.p. infection withSalmonella Enteritidis, another serovarof Salmonella Enterica, which became recently more important forhuman disease. This pathogen induces a typhoid-like infectionsimilar to that caused byS. Typhimurium in susceptible BALB/cmice (Itys), whereas mice with innately resistant background (e.g.,129Sv/Ev,Ityr) control low doses of virulentS. Enteritidis at theearly stage of infection before T cell- or B cell-mediated immunemechanisms develop (Ref. 4; and J. Lehmann, unpublished data).
IL-12 represents a heterodimeric protein consisting of two gly-cosylated subunits (35 and 40 kDa) that are linked covalently by a
disulfide bond (5, 6). The p40 subunit is strongly induced in APCsby bacterial and viral Ags, whereas the p35 subunit is constitu-tively transcribed (7, 8). Within the endoplasmic reticulum, bothsubunits are joined together to form the bioactive IL-12p75 mol-ecule (9). IL-12p75 has previously been shown to be the key factorfor the induction of a Th1 immune response, which is essential tocontrol infections with intracellular pathogens (10–12).
Besides the heterodimeric molecule IL-12p75, the p40 subunitcan also form homodimers (IL-12(p40)2). An excess of IL-12(p40)2 was found to act antagonistically to IL-12p75 in vitro andin vivo, because this molecule is able to compete with IL-12p75for the IL-12R�1 binding site (13–16). But unlike the IL-12p75,IL-12(p40)2 cannot bind to the IL-12R�2 subunit of the IL-12receptor, which is responsible for signal transduction leading toSTAT-4 activation (17). Recently, a new cytokine designatedIL-23 was identified, which consists of the IL-12p40 subunitcovalently linked to a newly discovered 19-kDa polypeptidechain (18). Like the IL-12(p40)2, this new molecule is alsocapable of binding only to IL-12R�1 but not IL-12R�2 (18). Sofar it is not known whether IL-23 can antagonize IL-12p75-induced effects.
Interestingly, an IL-12p40-dependent activity was found to con-tribute to the protective immune response against the fungal patho-gen Cryptococcus neoformans, a result that was previously pub-lished by our group (19). IL-12(p40)2 was also shown to actagonistically with IL-12 in a murine transplantation model via theactivation of CD8� T cells (20). Here we present for the first timedata which show that an IL-12p40-dependent activity is able tocontribute to protective immunity againstS. Enteritidis indepen-dently of IL-12p75.
Institutes of *Immunology and†Pathology, College of Veterinary Medicine, Univer-sity of Leipzig, Leipzig, Germany
Received for publication June 4, 2001. Accepted for publication August 23, 2001.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This project was supported by Grant AL371/3-1 from the DeutscheForschungsgemeinschaft.2 Current address: Landesveterinar- und Lebensmitteluntersuchungsamt Sachsen-Anhalt, Stendal, Germany.3 Address correspondence and reprint requests to Dr. Gottfried Alber, Institute ofImmunology, College of Veterinary Medicine, University of Leipzig, An den Tier-kliniken 11, D-04103 Leipzig, Germany. E-mail address: [email protected]
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Materials and MethodsMice and bacteria
Breeding pairs of BALB/c and 129Sv/Ev mice of three different genotypes(wild-type; IL-12p35�/�, Ref. 21; and IL-12p40�/� or IL-12p35/40�/�,Ref. 22) were provided by Dr. H. Mossmann (Max-Planck-Institute forImmunobiology, Freiburg, Germany). Mutant mice had been backcrossedfive times on a BALB/c background (23) or were on a genetically pure129Sv/Ev background. Animals were housed under specific pathogen-freeconditions in filter-topped cages within a Maxi-Miser Caging system(Thoren Caging Systems, Hazleton, PA) containing sterile bedding. Micewere fed sterile food and water. For most experiments wild-type and mu-tant BALB/c mice were used unless otherwise stated. To study the role ofthe Ity gene Itys BALB/c mice were compared with Ityr 129Sv/Ev mice.Animals were used at 8–12 wk of age.
Mice were infected with an attenuated vaccination strain of S. entericaserovar Enteritidis (S. Enteritidis; ade�, his�; SALMOVAC SE; Impfst-offwerk Dessau-Tornau, Rosslau, Germany) (4, 24) and, in indicated cases,challenged with one of two different virulent wild-type strains of S. En-teritidis (147 Nalr, high virulent or 64/03 Nalr, low virulent; provided byDr. S. Springer, Impfstoffwerk Dessau-Tornau).
Infection model
The animals (3–8/group) were inoculated i.p. with different doses (5 � 101
to 1 � 107 CFU) (4, 25). Different standard inocula were used to study theearly (1 � 106 CFU) or the late (2.5 � 103 CFU) infection stage in bothBALB/c and 129Sv/Ev background. Using an inoculum of 2.5 � 103 CFUin IL-12p40-deficient mice clinical symptoms occurred starting with day16 postinfection (p.i.)4 on the BALB/c or with day 42 p.i. in the 129Sv/Evmice. If not otherwise stated, wild-type and mutant mice on the BALB/cbackground were used in the experiments described.
Preparation of S. Enteritidis Ag
Heat-killed S. enteritidis (hk S.E.) were prepared by incubating bacteria at60°C for 60 min in a water bath. Killed salmonellae were stored at 1 �1010/ml in PBS at �75°C. This preparation was used as Ag for ex vivostimulation of splenocytes.
Bacterial count in spleen and liver
After different time points (2, 3, 4, 20, 42, or 48 days p.i.), three to six miceper group were sacrificed. The thorax and the peritoneum of the animalswere opened under sterile conditions, and they were bled by cardiac punc-ture, followed by the isolation of spleen and liver. After the determinationof the spleen and liver weights, both organs were homogenized. Then thespleen and liver homogenates were diluted 1/3 or 1/10 (w/v), respectively,in PBS. Log10 serial dilutions of the homogenates in PBS were plated ontoselective Xylose-Lysine-Desoxicholate agar (Merck, Darmstadt, Germa-ny). Colonies were counted after 24- and 48-h incubation periods at 37°C.In cases in which only a part of the organ was homogenized, counts werecorrected for the whole organ weights.
Enumeration of peritoneal cells or splenocytes containing viablesalmonellae
To evaluate the number of peritoneal or splenic phagocytes that were pos-itive for viable intracellular salmonellae, cells were freshly isolated fromthe peritoneal cavity by lavage with 5 ml of ice-cold PBS or from thespleen by passage through a 100-�m mesh cell strainer (Falcon; BD Bio-sciences, Heidelberg, Germany). Before plating, cells were washed threetimes in PBS, then incubated for 1 h in RPMI 1640 containing 100 �g/mlgentamicin (Life Technologies, Karlsruhe, Germany) to completely elim-inate extracellular bacteria. After two additional washing steps in PBS, thecell number was adjusted to 1 � 105/ml or 1 � 106/ml, and 1 ml of theperitoneal cell or splenocyte suspension was plated onto Xylose-Lysine-Desoxicholate agar (Merck) and air-dried under sterile conditions. Thedensity of the plated cells was checked microscopically to verify that singlenonconfluent bacterial colonies could develop from the seeded cells. Grow-ing colonies were counted after 24- and 48-h incubation periods at 37°C.Each single colony represented one Salmonella-positive phagocyte.
Detection of intracellular Salmonella Ag-positive cells by flowcytometry
Intracellular Salmonella Ag (comprising viable and killed/processed intra-cellular salmonellae) was indirectly fluorochrome-stained using a poly-clonal rabbit anti-S. Enteritidis antiserum as primary Ab and a FITC-la-beled goat anti-rabbit Ig (Jackson Immunoresearch Laboratories, WestGrove, PA) as secondary Ab. The antiserum was obtained after s.c. im-munization, and two intramuscular booster applications of an Ag suspen-sion containing heat-killed and freeze-thawed S. enteritidis bacteria com-bined with Al(OH)3 (Eurobio, Raunheim, Germany). Phagocytes wereidentified by staining with the PE-labeled anti-CD11b mAb M1/70.15(Caltag Laboratories, Hamburg, Germany) simultaneously to the secondarystaining of the Salmonella Ag.
Determination of total and Ag-specific serum IgE
Total IgE serum levels were measured with a sandwich ELISA accordingto a standard protocol by using mAbs and a murine IgE standard from BDPharMingen (Heidelberg, Germany). Briefly, 96-well microtiter plates(Maxisorp; Nunc, Roskilde, Denmark) were coated overnight at 4°C withthe anti-IgE mAb R35-118 in 0.1 M carbonate buffer (pH 9.6). Plates wereblocked and incubated with the 1/5 diluted serum samples and the mouseIgE standard. Then plate-bound IgE was detected by incubation with thebiotinylated anti-IgE mAb R35-72 for 1 h followed by a 30-min incubationat room temperature with streptavidin-peroxidase (Southern BiotechnologyAssociates, Birmingham, AL) and the ABTS substrate reaction (Sigma,Deisenhofen, Germany). The detection limit was 8 ng/ml.
For the detection of Ag-specific IgE microtiter plates (Polysorp; Nunc)were coated overnight at 4°C with whole-cell extracts prepared by repeatedfreeze-thaw cycles of a fresh bulk culture of S. Enteritidis. Plates wereblocked and then incubated with undiluted and 1/5 diluted mouse sera for2 h at room temperature, followed by detection of Ag-bound specific IgEserum Abs using the biotinylated anti-IgE mAb R35-72 and streptavidin-peroxidase as described above.
Isolation of splenocytes and ex vivo stimulation
Single cell suspensions of the spleens were prepared by mincing and pass-ing the tissue through a 100-�m mesh cell strainer (Falcon). The erythro-cytes were lysed by using ammonium chloride buffer (0.15 M NH4Cl, 0.1mM EDTA disodium salt dihydrate, 10 mM NaHCO3; Merck). The spleno-cytes were washed three times in cold PBS and resuspended in RPMI 1640culture medium (Life Technologies) supplemented with 10% heat-inacti-vated FCS, 2 mM L-glutamine (Biochrom, Berlin, Germany), 10 mMHEPES buffer, 100 U/ml penicillin, 100 �g/ml streptomycin, and 50 �M2-ME (Sigma). Following adjustment of the cell number to 5 � 106/ml, 1ml of splenocyte suspension was dispensed into 24-well plates and stim-ulated with either mitogen (5 �g/ml Con A; Sigma) or Ag (106-108 hk S.E.)or left unstimulated. After 48 h of culture at 37°C in a humidified atmo-sphere containing 5% CO2, cell-free culture supernatants were harvestedand stored in aliquots at �75°C until cytokine measurements.
Cytokine determination in sera and splenocyte supernatants
Cytokines (IFN-�, IL-12p40, TNF-�, IL-4, IL-6, IL-10) in sera and cellculture supernatants were measured by sandwich ELISA according to stan-dard protocols by using matching pairs of mAbs. Briefly, IFN-� was as-sessed by using the mAb AN-18 (5 �g/ml; American Type Culture Col-lection (ATCC), Manassas, VA) as capture and the peroxidase-conjugatedmAb XMG1.2 (ATCC) as detection Ab (both mAbs were provided by Dr.H. Gallati, Hoffmann-LaRoche, Basel, Switzerland), with recombinantmouse IFN-� (provided by Dr. G. R. Adolf, Ernst Boehringer Institute,Vienna, Austria) as the standard (detection limit: 16 pg/ml). IL-12p40 wasmeasured by using the mAb 5C3 (5 �g/ml; ATCC) as capture and biotin-ylated polyclonal goat anti-mouse IL-12 as detection Ab (both Abs wereprovided by Dr. H. Gallati, F. Hoffmann-LaRoche) combined with strepta-vidin-peroxidase (Southern Biotechnology Associates), with recombinantmouse IL-12 (provided by Dr. M. Gately, Hoffmann-LaRoche, Nutley, NJ)as the standard (detection limit: 156 pg/ml). IL-6 was measured using themAb MP5-20F3 (5 �g/ml) as capture and the biotinylated mAb MP5-32C11 (5 �g/ml) as detection Ab (both obtained from BD PharMingen)combined with streptavidin-peroxidase (Southern Biotechnology Associ-ates), with recombinant mouse IL-6 (BD PharMingen) as standard (detec-tion limit: 32 pg/ml).
IL-4, IL-10, and TNF-� were measured by using the respective DuoSetELISA development systems (R&D Systems, Wiesbaden, Germany) fol-lowing the manufacturers instructions.
4 Abbreviations used in this paper: p.i., postinfection; hk S.E., heat-killed S.Enteritidis.
5305The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In vivo neutralization of IL-12p40
In indicated experiments IL-12p40 was neutralized in IL-12p35�/� miceby i.p. injection of 100 �g of anti-mouse IL-12p40 mAb 10F6 (ATCC,provided by Dr. H. Gallati) per mouse at days �1 and 1 p.i.
In vivo depletion of CD4� and CD8� cells
To deplete CD4� or CD8� T cells, groups of five animals of each genotype(wild-type, IL-12p35�/�, IL-12p40�/�) were treated with anti-mouse CD4(anti-L3T4, clone GK 1.5, rat IgG2b; a gift from Dr. S. Ehlers, ResearchCenter Borstel, Borstel, Germany) or with anti-mouse CD8 (anti-Lyt2.2,clone 2.43, rat IgG2b; a gift from Dr. S. Ehlers; in combination with anti-Lyt2.1/Lyt2.2, clone 53-6.7, rat IgG2a; BD PharMingen). Both CD4 andCD8 Abs were administered i.p. at total doses of 0.5 mg/mouse on threeconsecutive days (days �6, �5, and �4). Six days after the first mAbinjection the degree of depletion (reaching 99.9% for CD4� and 99.6% forCD8�) was analyzed by FACS analysis (26). Then the depleted animalswere infected with 2.5 � 103 CFU S. enteritidis.
Histopathological examination
Spleen and liver tissue samples from sacrificed mice were immediatelyfixed in 5% phosphate-buffered formalin and embedded in paraffin. Tissuesections (5 �m) were stained with H&E and analyzed for the tissue gran-ulomatous response using light microscopy. Randomly selected (n � 8)high power fields of tissue sections from both organs were examined fromeach animal.
Quantification of IL-12p40, IL-23p19, and �-actin mRNA levelsby real-time PCR
Total RNA was extracted from 10 mg of homogenized mouse spleen usingthe RNeasy minikit (Qiagen, Hilden, Germany) according to the manufac-turer’s instructions combined with a DNA digestion step (DNase, RocheDiagnostics, Mannheim, Germany). Reverse transcription was performedwith 500 ng of total RNA, peqGOLD dNTP-Mix (Peqlab, Erlangen, Ger-many), oligo(dT) primer, and Expand Reverse Transcriptase (both obtainedfrom Roche Diagnostics) according to the manufacturer’s instructions us-ing a conventional thermocycler (GeneAmp PCR System 2400; AppliedBiosystems, Weiterstadt, Germany) in a total volume of 20 �l.
For relative quantification of IL-12p40 and IL-23p19 mRNA, 5 �l ofeach sample cDNA was used for real-time PCR, which was conducted onthe LightCycler Instrument (Roche Diagnostics). The amplification ofcDNA was monitored with the fluorescent DNA binding dye SYBR Greenusing the LightCycler-FastStart DNA Master SYBR Green I kit (RocheDiagnostics), according to the manufacturer’s instructions. Specific prim-ers were designed using Primer Express software (Applied Biosystems).The following primer pairs were used for �-actin (forward primer, 5�-AGAGGG AAA TCG TGC GTG AC-3�; reverse primer, 5�-CAA TAG TGATGA CCT GGC CGT-3�; product size 148 bp), IL-12p40 (forward primer,5�-GGA AGC ACG GCA GCA GAA TA-3�; reverse primer, 5�-AAC TTGAGG GAG AAG TAG GAA TGG-3�; product size 180 bp), IL-23p19(forward primer, 5�-TGC TGG ATT GCA GAG CAG TAA-3�; reverseprimer, 5�-GCA TGC AGA GAT TCC GAG AGA-3�; product size 121bp). The cytokine mRNA levels of infected animals were relatively quan-titated in comparison to the respective level of naive animals by using thecomparative CT (threshold cycle) method (27). Briefly, the amount of tar-get, normalized to an endogenous reference (�-actin) and relative to acalibrator (cytokine mRNA expression in naive animals), is given by:2���CT, where �CT � CT,X � CT,R is the difference in threshold cycles fortarget and reference and ��CT � �CT,q � �CT,cb is the difference in thethreshold cycles for any sample q and the calibrator cb. Before using the��CT method for relative quantitation, a validation experiment was per-formed to demonstrate that efficiencies of target amplification and refer-ence amplification are approximately equal. For validation of IL-12p40 andIL-23p19 mRNA expression total cDNAs prepared from murine primarybone marrow macrophages stimulated with IFN-� (100 U/ml) � LPS (100ng/ml) or RAW264.7 macrophages (ATCC) stimulated with hk S.E. (107/ml) were used as model systems to generate standard curves for IL-12p40or IL-23p19, respectively. The crossing point for each reaction was deter-mined using the Second Derivative Maximum algorithm and the arithmeticbaseline adjustment using LightCycler software (Roche Diagnostics).
Statistical analysis
Statistical analyses were performed with the Mann-Whitney rank sum testor Student’s t test if the samples passed the normality and the equal vari-ance analyses by using SigmaStat for Windows software (Jandel, Erkrath,Germany).
ResultsElevated survival rate and longer survival time in S. Enteritidis-infected IL-12p35�/� vs IL-12p40�/� mice
To characterize the course of S. Enteritidis infection in mice in theabsence of either IL-12p75 or IL-12p40, the survival rates as wellas the survival times of IL-12p35�/�, IL-12p40�/�, and wild-typemice were determined. Wild-type mice that were able to produceIL-12p75 as well as IL-12p40 survived inocula of up to 107 CFUof S. Enteritidis (LD50 � 3 � 107) and showed clinical symptomsfollowing inoculation with �106 CFU. In contrast, both IL-12p35-and IL-12p40-deficient mice were more sensitive to Salmonellainfection. Mice of both strains died when infected with �5 � 103
CFU. However, at doses lower than 5 � 103 CFU the survivalrates and the survival times differed between IL-12p35- and IL-12p40-deficient mice in a dose-dependent fashion (Fig. 1). At aninoculum of 50 CFU IL-12p35�/� mice completely survived up today 50 p.i., whereas 50% of the IL-12p40�/� mice died starting at
FIGURE 1. Following i.p. infection with live attenuated S. Enteritidisbacteria IL-12p35�/� mice showed higher survival rates or longer survivaltimes than IL-12p40�/� mice depending on the infection dose. At the low-est inoculum of 5 � 101 CFU (top) all IL-12p35�/� stayed alive, whereas50% of the IL-12p40�/� mice died of the infection (n � 6). At a 10-foldhigher inoculum of 5 � 102 CFU (middle) only 50% of the IL-12p35�/�
but none of the IL-12p40�/� survived (n � 6), and at a 100-fold higherinoculum of 5 � 103 CFU (bottom), neither IL-12p40�/� nor IL-12p35�/�
mice survived the infection (n � 6). However, at this infection dose themean survival time of IL-12p35�/� mice was significantly higher than thatof IL-12p40�/� mice (26.7 vs 21.5 days, p � 0.032; Student’s t test).
5306 IL-12p40-DEPENDENT AGONISTIC EFFECTS IN MURINE SALMONELLOSIS
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
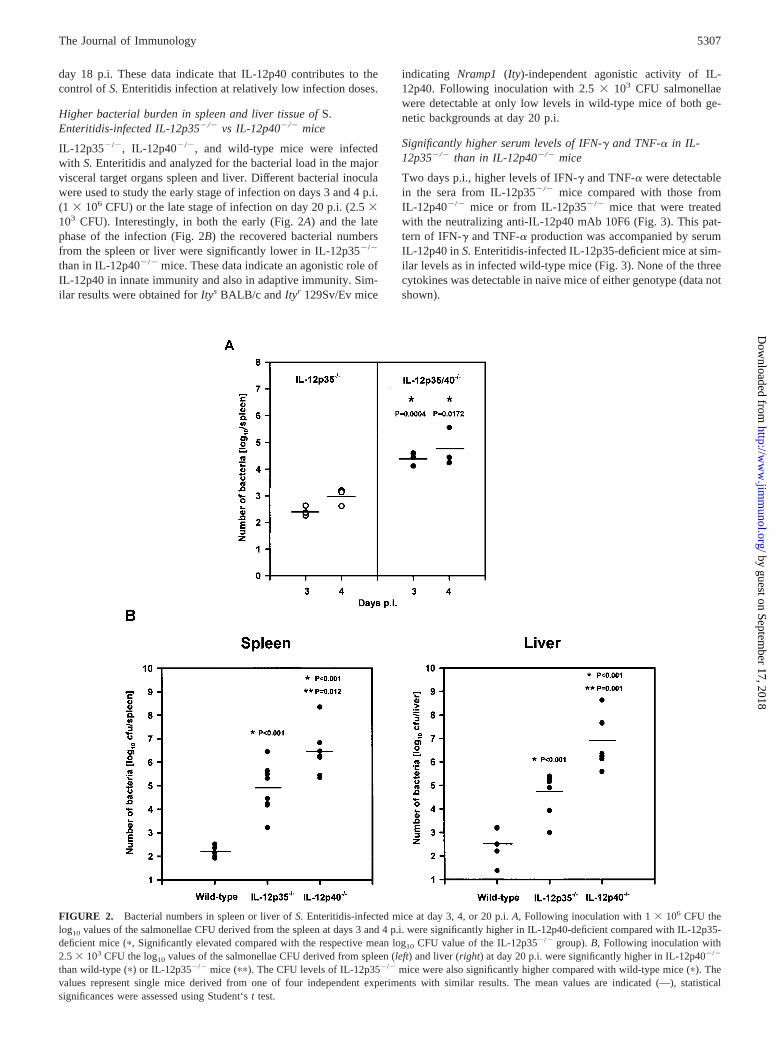
day 18 p.i. These data indicate that IL-12p40 contributes to thecontrol of S. Enteritidis infection at relatively low infection doses.
Higher bacterial burden in spleen and liver tissue of S.Enteritidis-infected IL-12p35�/� vs IL-12p40�/� mice
IL-12p35�/�, IL-12p40�/�, and wild-type mice were infectedwith S. Enteritidis and analyzed for the bacterial load in the majorvisceral target organs spleen and liver. Different bacterial inoculawere used to study the early stage of infection on days 3 and 4 p.i.(1 � 106 CFU) or the late stage of infection on day 20 p.i. (2.5 �103 CFU). Interestingly, in both the early (Fig. 2A) and the latephase of the infection (Fig. 2B) the recovered bacterial numbersfrom the spleen or liver were significantly lower in IL-12p35�/�
than in IL-12p40�/� mice. These data indicate an agonistic role ofIL-12p40 in innate immunity and also in adaptive immunity. Sim-ilar results were obtained for Itys BALB/c and Ityr 129Sv/Ev mice
indicating Nramp1 (Ity)-independent agonistic activity of IL-12p40. Following inoculation with 2.5 � 103 CFU salmonellaewere detectable at only low levels in wild-type mice of both ge-netic backgrounds at day 20 p.i.
Significantly higher serum levels of IFN-� and TNF-� in IL-12p35�/� than in IL-12p40�/� mice
Two days p.i., higher levels of IFN-� and TNF-� were detectablein the sera from IL-12p35�/� mice compared with those fromIL-12p40�/� mice or from IL-12p35�/� mice that were treatedwith the neutralizing anti-IL-12p40 mAb 10F6 (Fig. 3). This pat-tern of IFN-� and TNF-� production was accompanied by serumIL-12p40 in S. Enteritidis-infected IL-12p35-deficient mice at sim-ilar levels as in infected wild-type mice (Fig. 3). None of the threecytokines was detectable in naive mice of either genotype (data notshown).
FIGURE 2. Bacterial numbers in spleen or liver of S. Enteritidis-infected mice at day 3, 4, or 20 p.i. A, Following inoculation with 1 � 106 CFU thelog10 values of the salmonellae CFU derived from the spleen at days 3 and 4 p.i. were significantly higher in IL-12p40-deficient compared with IL-12p35-deficient mice (�, Significantly elevated compared with the respective mean log 10 CFU value of the IL-12p35�/� group). B, Following inoculation with2.5 � 103 CFU the log10 values of the salmonellae CFU derived from spleen (left) and liver (right) at day 20 p.i. were significantly higher in IL-12p40�/�
than wild-type (�) or IL-12p35�/� mice (��). The CFU levels of IL-12p35�/� mice were also significantly higher compared with wild-type mice (�). Thevalues represent single mice derived from one of four independent experiments with similar results. The mean values are indicated (—), statisticalsignificances were assessed using Student‘s t test.
5307The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Phagocyte cytotoxicity against intracellular S. Enteritidis isstrongly impaired in IL-12p40-deficient mice
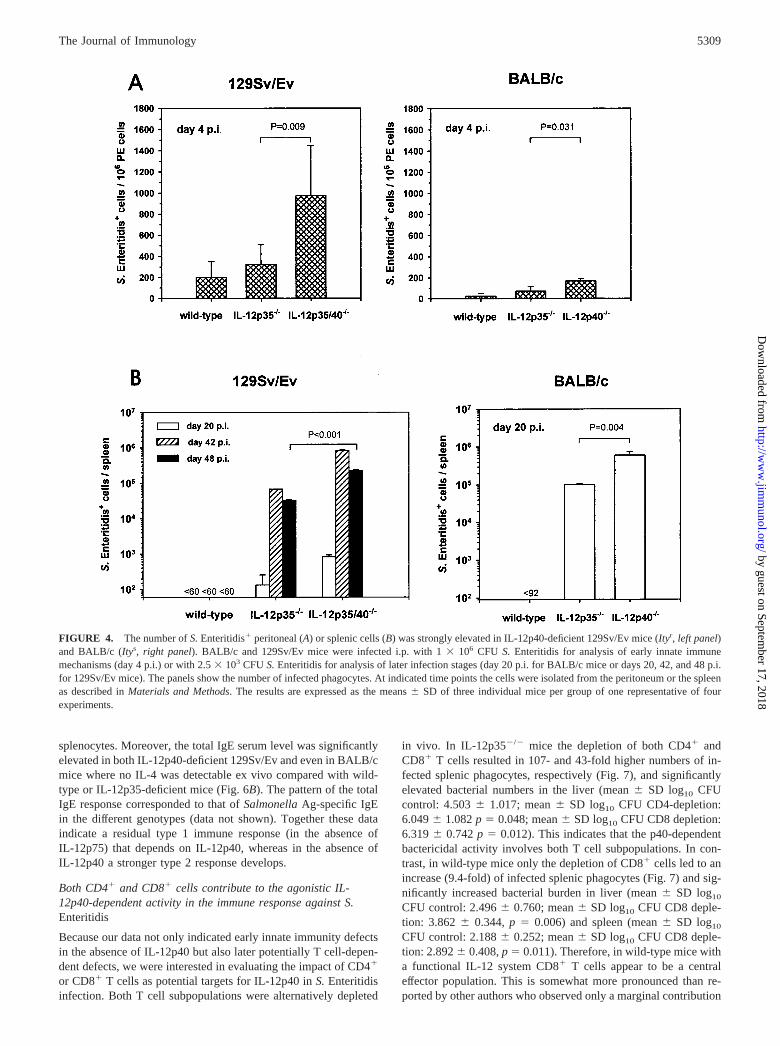
To study the underlying IL-12p40-dependent protective mecha-nism(s) in S. Enteritidis-infected wild-type, IL-12p35-, and IL-12p40-deficient mice, the bactericidal capacity of splenic phago-cytes was determined by plating and subsequently countingSalmonella-positive splenocytes. Using this assay it was possibleto detect single intracellularly infected phagocytes. As shown inFig. 4 the number of S. Enteritidis-infected phagocytes was sig-nificantly elevated in IL-12p40�/� compared with IL-12p35�/�
mice as early as day 4 p.i. in the peritoneal cavity (Fig. 4A) as wellas later on (i.e., days 20, 42, and 48 p.i.) in the spleen (Fig. 4B).
Interestingly, the frequency of infected phagocytes was elevatedin BALB/c, and even more pronounced in 129Sv/Ev mice, indi-cating that the IL-12p40-dependent effect on the phagocyte bacte-ricidal capacity was independent of the Nramp1(Ity) genotype.Wild-type mice of both genetic backgrounds had lower amounts ofSalmonella-positive peritoneal cells than IL-12p35�/� mice be-tween days 1 and 4 p.i. (these differences were not statisticallysignificant; Fig. 4A). At the later time points (i.e., days 20, 42, and48 p.i.) viable Salmonella-positive cells were no more detectablein the spleens from wild-type mice, at detection limits of 92 and 60cells/spleen (Fig. 4B).
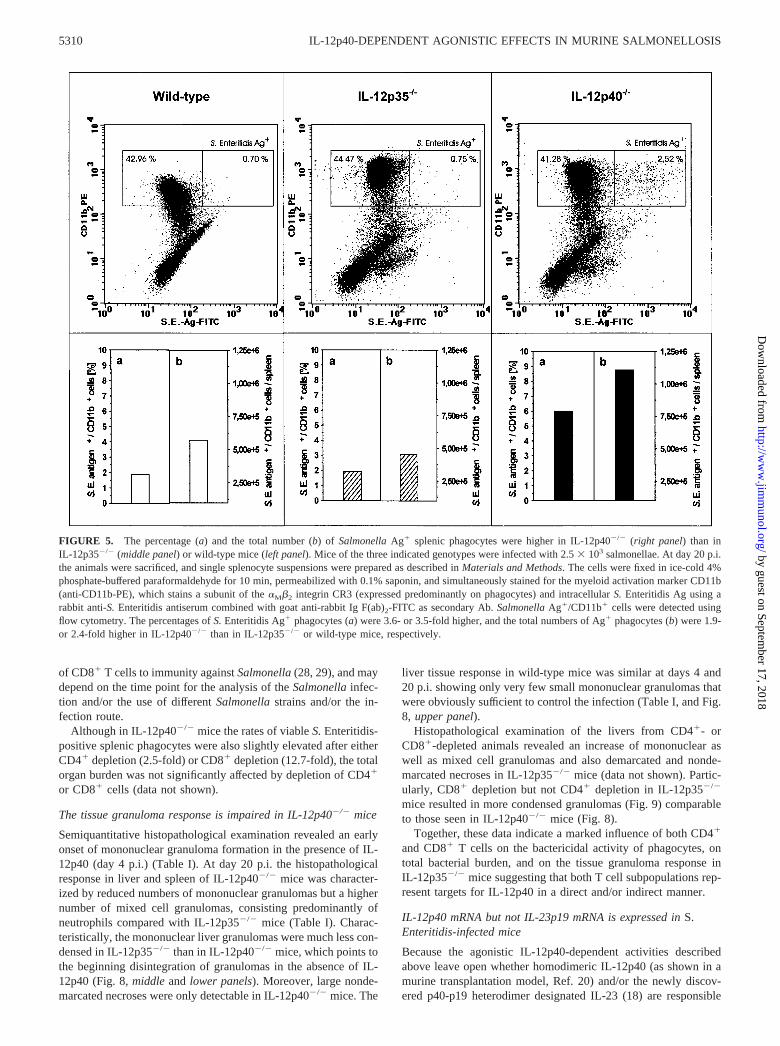
The data obtained for viable Salmonella-positive splenic phago-cytes were confirmed by intracellular staining of Salmonella Ag insplenic macrophages derived from wild-type, IL-12p35�/�, andIL-12p40�/� mice (day 20 p.i.) and analysis by flow cytometry. InIL-12p40�/� mice the percentages of S. Enteritidis Ag-positivemacrophages were 3.6-fold higher than in wild-type mice, and 3.5-fold higher than in IL-12p35�/� mice. In addition, the totalnumber of Ag-positive splenic macrophages was 1.9-fold highercompared with wild-type mice, and 2.4-fold higher compared withIL-12p35�/� mice; this shows that the total cell number was lowerin wild-type than in IL-12p35�/� mice (Fig. 5). These data indi-cate a correlation between the high levels of intracellular Ag andthe number of live intracellular salmonellae shown in Fig. 4 inIL-12p40-deficient mice. Interestingly, this analysis demonstratedalso the presence of residual Salmonella Ag in wild-type splenicmacrophages at a time point where no viable bacteria could befound anymore in wild-type mice.
IL-12p35�/� mice mount a partial Th1 response against S.Enteritidis, whereas the Th1 response is completely absent inIL-12p40�/� mice
To differentiate the immune responses induced by S. Enteritidis inwild-type, IL-12p35-, and IL-12p40-deficient mice into Th1- orTh2-biased responses, IFN-� and TNF-� vs IL-4 levels in mito-gen- or Ag-induced splenocyte culture supernatants, and serumIgE levels from S. Enteritidis-infected animals were analyzed after20 (BALB/c) or 48 (129Sv/Ev) days of infection. As mentioned inMaterials and Methods the symptomatic stages of the S. Enteritidisinfection occurred between days 16 and 20 p.i. in BALB/c miceand between days 42 and 48 p.i. in 129Sv/Ev mice when infectedwith 2.5 � 103 CFU. Therefore, we analyzed T cell responses atday 20 p.i. in BALB/c and day 48 p.i. in 129Sv/Ev mice.
As shown in Fig. 6A, the mitogen-induced IFN-� concentrationsin splenic culture supernatants from IL-12p35�/� 129Sv/Ev andfrom IL-12p35�/� BALB/c mice were 2.5- and 9.8-fold higherthan from IL-12p40-deficient mice, respectively. Similar data wereobtained for Ag-induced IFN-� (data not shown). In IL-12p40-deficient 129Sv/Ev mice, lower IFN-� amounts corresponded withelevated IL-4 amounts. However, IL-4 was almost undetectable inculture supernatants derived from IL-12p40-deficient BALB/c
FIGURE 3. Serum IFN-� (upper) and TNF-� levels (middle) are sig-nificantly decreased in S. Enteritidis-infected IL-12p40-deficient mice atday 2 p.i. As shown in the lower graph IL-12p40 was induced in vivo byS. Enteritidis infection in wild-type as well as in IL-12p35-deficient mice.The mice were inoculated with 1 � 107 CFU S. Enteritidis, and thesera were taken at day 2 p.i. To verify the specific importance ofIL-12p40 during S. Enteritidis infection in mice with a functional p40gene, IL-12p40 was neutralized in IL-12p35�/� mice by i.p. injection ofthe anti-IL-12p40 mAb 10F6 (100 �g/application at days �1 and 1 p.i.).The values represent single mice. The mean or median values are indicated(—), statistical significances were assessed using Student‘s t test or theMann-Whitney rank sum test, respectively (� vs IL-12p35�/��anti-IL-12p40; �� vs IL-12p40�/�). None of the three cytokines was detectable innaive animals of either genotype (data not shown).
5308 IL-12p40-DEPENDENT AGONISTIC EFFECTS IN MURINE SALMONELLOSIS
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
splenocytes. Moreover, the total IgE serum level was significantlyelevated in both IL-12p40-deficient 129Sv/Ev and even in BALB/cmice where no IL-4 was detectable ex vivo compared with wild-type or IL-12p35-deficient mice (Fig. 6B). The pattern of the totalIgE response corresponded to that of Salmonella Ag-specific IgEin the different genotypes (data not shown). Together these dataindicate a residual type 1 immune response (in the absence ofIL-12p75) that depends on IL-12p40, whereas in the absence ofIL-12p40 a stronger type 2 response develops.
Both CD4� and CD8� cells contribute to the agonistic IL-12p40-dependent activity in the immune response against S.Enteritidis
Because our data not only indicated early innate immunity defectsin the absence of IL-12p40 but also later potentially T cell-depen-dent defects, we were interested in evaluating the impact of CD4�
or CD8� T cells as potential targets for IL-12p40 in S. Enteritidisinfection. Both T cell subpopulations were alternatively depleted
in vivo. In IL-12p35�/� mice the depletion of both CD4� andCD8� T cells resulted in 107- and 43-fold higher numbers of in-fected splenic phagocytes, respectively (Fig. 7), and significantlyelevated bacterial numbers in the liver (mean � SD log10 CFUcontrol: 4.503 � 1.017; mean � SD log10 CFU CD4-depletion:6.049 � 1.082 p � 0.048; mean � SD log10 CFU CD8 depletion:6.319 � 0.742 p � 0.012). This indicates that the p40-dependentbactericidal activity involves both T cell subpopulations. In con-trast, in wild-type mice only the depletion of CD8� cells led to anincrease (9.4-fold) of infected splenic phagocytes (Fig. 7) and sig-nificantly increased bacterial burden in liver (mean � SD log10
CFU control: 2.496 � 0.760; mean � SD log10 CFU CD8 deple-tion: 3.862 � 0.344, p � 0.006) and spleen (mean � SD log10
CFU control: 2.188 � 0.252; mean � SD log10 CFU CD8 deple-tion: 2.892 � 0.408, p � 0.011). Therefore, in wild-type mice witha functional IL-12 system CD8� T cells appear to be a centraleffector population. This is somewhat more pronounced than re-ported by other authors who observed only a marginal contribution
FIGURE 4. The number of S. Enteritidis� peritoneal (A) or splenic cells (B) was strongly elevated in IL-12p40-deficient 129Sv/Ev mice (Ityr, left panel)and BALB/c (Itys, right panel). BALB/c and 129Sv/Ev mice were infected i.p. with 1 � 106 CFU S. Enteritidis for analysis of early innate immunemechanisms (day 4 p.i.) or with 2.5 � 103 CFU S. Enteritidis for analysis of later infection stages (day 20 p.i. for BALB/c mice or days 20, 42, and 48 p.i.for 129Sv/Ev mice). The panels show the number of infected phagocytes. At indicated time points the cells were isolated from the peritoneum or the spleenas described in Materials and Methods. The results are expressed as the means � SD of three individual mice per group of one representative of fourexperiments.
5309The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
of CD8� T cells to immunity against Salmonella (28, 29), and maydepend on the time point for the analysis of the Salmonella infec-tion and/or the use of different Salmonella strains and/or the in-fection route.
Although in IL-12p40�/� mice the rates of viable S. Enteritidis-positive splenic phagocytes were also slightly elevated after eitherCD4� depletion (2.5-fold) or CD8� depletion (12.7-fold), the totalorgan burden was not significantly affected by depletion of CD4�
or CD8� cells (data not shown).
The tissue granuloma response is impaired in IL-12p40�/� mice
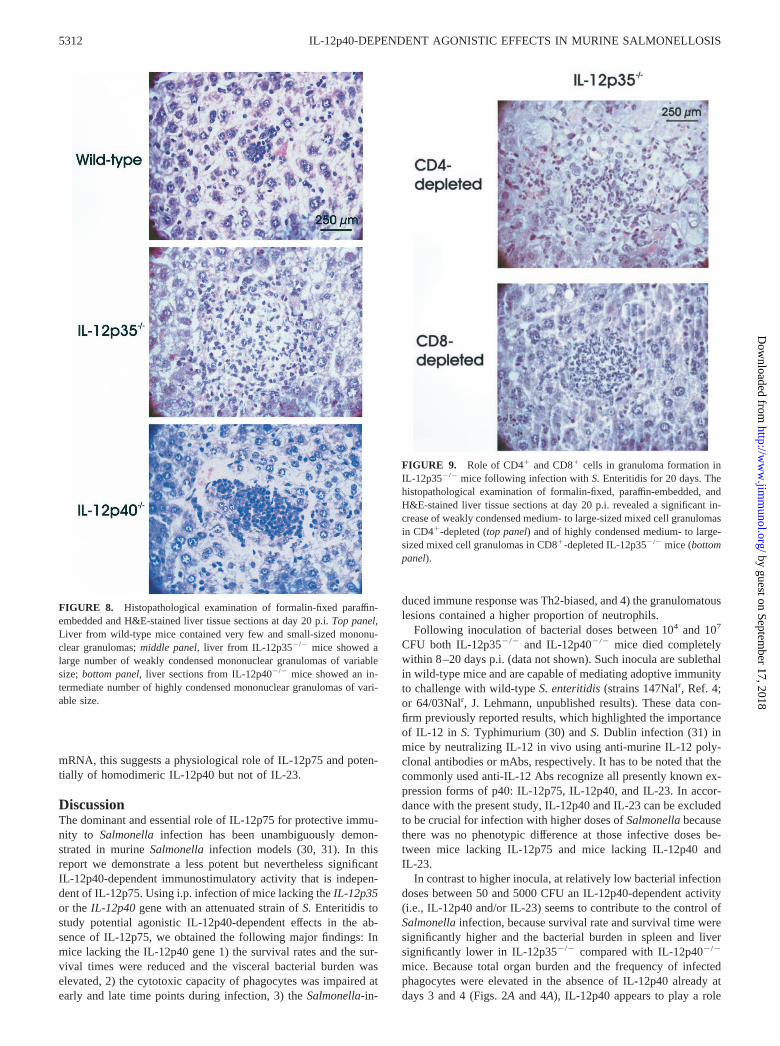
Semiquantitative histopathological examination revealed an earlyonset of mononuclear granuloma formation in the presence of IL-12p40 (day 4 p.i.) (Table I). At day 20 p.i. the histopathologicalresponse in liver and spleen of IL-12p40�/� mice was character-ized by reduced numbers of mononuclear granulomas but a highernumber of mixed cell granulomas, consisting predominantly ofneutrophils compared with IL-12p35�/� mice (Table I). Charac-teristically, the mononuclear liver granulomas were much less con-densed in IL-12p35�/� than in IL-12p40�/� mice, which points tothe beginning disintegration of granulomas in the absence of IL-12p40 (Fig. 8, middle and lower panels). Moreover, large nonde-marcated necroses were only detectable in IL-12p40�/� mice. The
liver tissue response in wild-type mice was similar at days 4 and20 p.i. showing only very few small mononuclear granulomas thatwere obviously sufficient to control the infection (Table I, and Fig.8, upper panel).
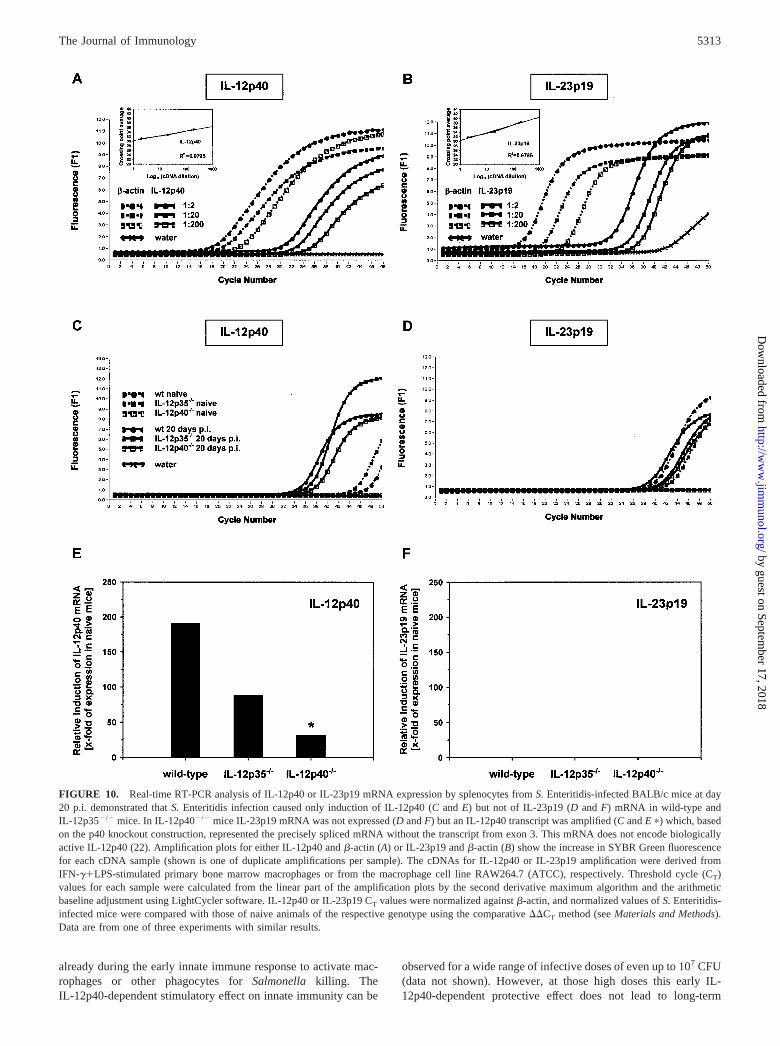
Histopathological examination of the livers from CD4�- orCD8�-depleted animals revealed an increase of mononuclear aswell as mixed cell granulomas and also demarcated and nonde-marcated necroses in IL-12p35�/� mice (data not shown). Partic-ularly, CD8� depletion but not CD4� depletion in IL-12p35�/�
mice resulted in more condensed granulomas (Fig. 9) comparableto those seen in IL-12p40�/� mice (Fig. 8).
Together, these data indicate a marked influence of both CD4�
and CD8� T cells on the bactericidal activity of phagocytes, ontotal bacterial burden, and on the tissue granuloma response inIL-12p35�/� mice suggesting that both T cell subpopulations rep-resent targets for IL-12p40 in a direct and/or indirect manner.
IL-12p40 mRNA but not IL-23p19 mRNA is expressed in S.Enteritidis-infected mice
Because the agonistic IL-12p40-dependent activities describedabove leave open whether homodimeric IL-12p40 (as shown in amurine transplantation model, Ref. 20) and/or the newly discov-ered p40-p19 heterodimer designated IL-23 (18) are responsible
FIGURE 5. The percentage (a) and the total number (b) of Salmonella Ag� splenic phagocytes were higher in IL-12p40�/� (right panel) than inIL-12p35�/� (middle panel) or wild-type mice (left panel). Mice of the three indicated genotypes were infected with 2.5 � 103 salmonellae. At day 20 p.i.the animals were sacrificed, and single splenocyte suspensions were prepared as described in Materials and Methods. The cells were fixed in ice-cold 4%phosphate-buffered paraformaldehyde for 10 min, permeabilized with 0.1% saponin, and simultaneously stained for the myeloid activation marker CD11b(anti-CD11b-PE), which stains a subunit of the �M�2 integrin CR3 (expressed predominantly on phagocytes) and intracellular S. Enteritidis Ag using arabbit anti-S. Enteritidis antiserum combined with goat anti-rabbit Ig F(ab)2-FITC as secondary Ab. Salmonella Ag�/CD11b� cells were detected usingflow cytometry. The percentages of S. Enteritidis Ag� phagocytes (a) were 3.6- or 3.5-fold higher, and the total numbers of Ag� phagocytes (b) were 1.9-or 2.4-fold higher in IL-12p40�/� than in IL-12p35�/� or wild-type mice, respectively.
5310 IL-12p40-DEPENDENT AGONISTIC EFFECTS IN MURINE SALMONELLOSIS
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
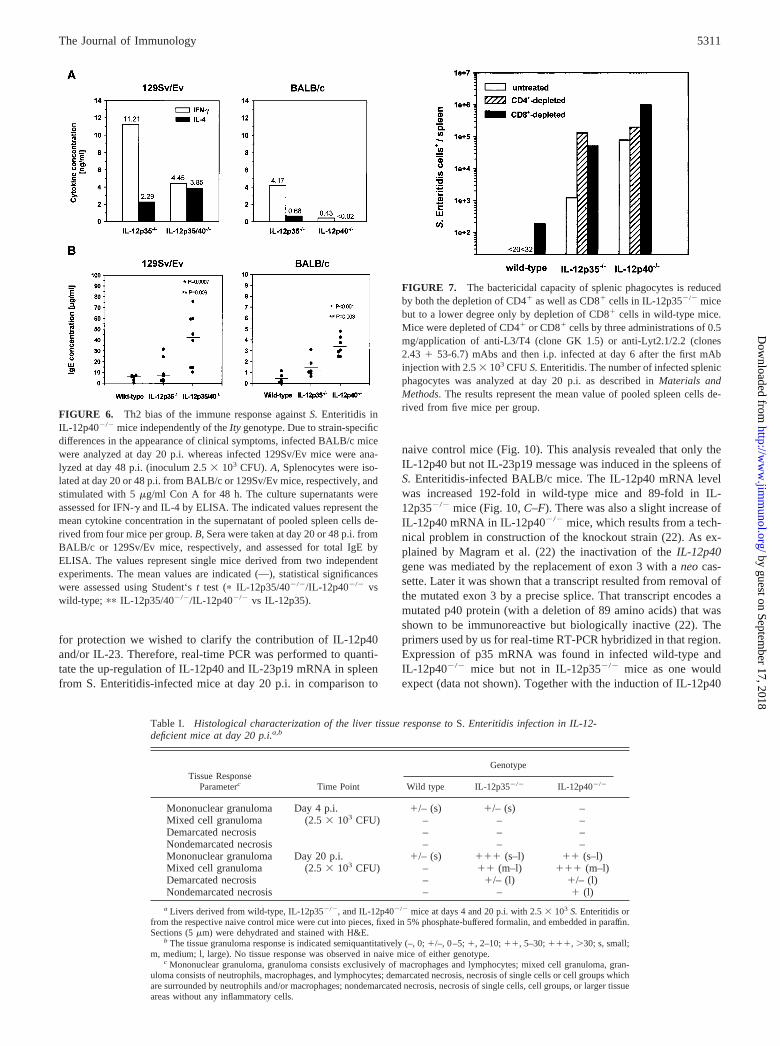
for protection we wished to clarify the contribution of IL-12p40and/or IL-23. Therefore, real-time PCR was performed to quanti-tate the up-regulation of IL-12p40 and IL-23p19 mRNA in spleenfrom S. Enteritidis-infected mice at day 20 p.i. in comparison to
naive control mice (Fig. 10). This analysis revealed that only theIL-12p40 but not IL-23p19 message was induced in the spleens ofS. Enteritidis-infected BALB/c mice. The IL-12p40 mRNA levelwas increased 192-fold in wild-type mice and 89-fold in IL-12p35�/� mice (Fig. 10, C–F). There was also a slight increase ofIL-12p40 mRNA in IL-12p40�/� mice, which results from a tech-nical problem in construction of the knockout strain (22). As ex-plained by Magram et al. (22) the inactivation of the IL-12p40gene was mediated by the replacement of exon 3 with a neo cas-sette. Later it was shown that a transcript resulted from removal ofthe mutated exon 3 by a precise splice. That transcript encodes amutated p40 protein (with a deletion of 89 amino acids) that wasshown to be immunoreactive but biologically inactive (22). Theprimers used by us for real-time RT-PCR hybridized in that region.Expression of p35 mRNA was found in infected wild-type andIL-12p40�/� mice but not in IL-12p35�/� mice as one wouldexpect (data not shown). Together with the induction of IL-12p40
Table I. Histological characterization of the liver tissue response to S. Enteritidis infection in IL-12-deficient mice at day 20 p.i.a,b
Tissue ResponseParameterc Time Point
Genotype
Wild type IL-12p35�/� IL-12p40�/�
Mononuclear granuloma Day 4 p.i. �/– (s) �/– (s) –Mixed cell granuloma (2.5 � 103 CFU) – – –Demarcated necrosis – – –Nondemarcated necrosis – – –Mononuclear granuloma Day 20 p.i. �/– (s) ��� (s–l) �� (s–l)Mixed cell granuloma (2.5 � 103 CFU) – �� (m–l) ��� (m–l)Demarcated necrosis – �/– (l) �/– (l)Nondemarcated necrosis – – � (l)
a Livers derived from wild-type, IL-12p35�/�, and IL-12p40�/� mice at days 4 and 20 p.i. with 2.5 � 103 S. Enteritidis orfrom the respective naive control mice were cut into pieces, fixed in 5% phosphate-buffered formalin, and embedded in paraffin.Sections (5 �m) were dehydrated and stained with H&E.
b The tissue granuloma response is indicated semiquantitatively (–, 0; �/–, 0–5; �, 2–10; ��, 5–30; ���, �30; s, small;m, medium; l, large). No tissue response was observed in naive mice of either genotype.
c Mononuclear granuloma, granuloma consists exclusively of macrophages and lymphocytes; mixed cell granuloma, gran-uloma consists of neutrophils, macrophages, and lymphocytes; demarcated necrosis, necrosis of single cells or cell groups whichare surrounded by neutrophils and/or macrophages; nondemarcated necrosis, necrosis of single cells, cell groups, or larger tissueareas without any inflammatory cells.
FIGURE 6. Th2 bias of the immune response against S. Enteritidis inIL-12p40�/� mice independently of the Ity genotype. Due to strain-specificdifferences in the appearance of clinical symptoms, infected BALB/c micewere analyzed at day 20 p.i. whereas infected 129Sv/Ev mice were ana-lyzed at day 48 p.i. (inoculum 2.5 � 103 CFU). A, Splenocytes were iso-lated at day 20 or 48 p.i. from BALB/c or 129Sv/Ev mice, respectively, andstimulated with 5 �g/ml Con A for 48 h. The culture supernatants wereassessed for IFN-� and IL-4 by ELISA. The indicated values represent themean cytokine concentration in the supernatant of pooled spleen cells de-rived from four mice per group. B, Sera were taken at day 20 or 48 p.i. fromBALB/c or 129Sv/Ev mice, respectively, and assessed for total IgE byELISA. The values represent single mice derived from two independentexperiments. The mean values are indicated (—), statistical significanceswere assessed using Student‘s t test (� IL-12p35/40�/�/IL-12p40�/� vswild-type; �� IL-12p35/40�/�/IL-12p40�/� vs IL-12p35).
FIGURE 7. The bactericidal capacity of splenic phagocytes is reducedby both the depletion of CD4� as well as CD8� cells in IL-12p35�/� micebut to a lower degree only by depletion of CD8� cells in wild-type mice.Mice were depleted of CD4� or CD8� cells by three administrations of 0.5mg/application of anti-L3/T4 (clone GK 1.5) or anti-Lyt2.1/2.2 (clones2.43 � 53-6.7) mAbs and then i.p. infected at day 6 after the first mAbinjection with 2.5 � 103 CFU S. Enteritidis. The number of infected splenicphagocytes was analyzed at day 20 p.i. as described in Materials andMethods. The results represent the mean value of pooled spleen cells de-rived from five mice per group.
5311The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
mRNA, this suggests a physiological role of IL-12p75 and poten-tially of homodimeric IL-12p40 but not of IL-23.
DiscussionThe dominant and essential role of IL-12p75 for protective immu-nity to Salmonella infection has been unambiguously demon-strated in murine Salmonella infection models (30, 31). In thisreport we demonstrate a less potent but nevertheless significantIL-12p40-dependent immunostimulatory activity that is indepen-dent of IL-12p75. Using i.p. infection of mice lacking the IL-12p35or the IL-12p40 gene with an attenuated strain of S. Enteritidis tostudy potential agonistic IL-12p40-dependent effects in the ab-sence of IL-12p75, we obtained the following major findings: Inmice lacking the IL-12p40 gene 1) the survival rates and the sur-vival times were reduced and the visceral bacterial burden waselevated, 2) the cytotoxic capacity of phagocytes was impaired atearly and late time points during infection, 3) the Salmonella-in-
duced immune response was Th2-biased, and 4) the granulomatouslesions contained a higher proportion of neutrophils.
Following inoculation of bacterial doses between 104 and 107
CFU both IL-12p35�/� and IL-12p40�/� mice died completelywithin 8–20 days p.i. (data not shown). Such inocula are sublethalin wild-type mice and are capable of mediating adoptive immunityto challenge with wild-type S. enteritidis (strains 147Nalr, Ref. 4;or 64/03Nalr, J. Lehmann, unpublished results). These data con-firm previously reported results, which highlighted the importanceof IL-12 in S. Typhimurium (30) and S. Dublin infection (31) inmice by neutralizing IL-12 in vivo using anti-murine IL-12 poly-clonal antibodies or mAbs, respectively. It has to be noted that thecommonly used anti-IL-12 Abs recognize all presently known ex-pression forms of p40: IL-12p75, IL-12p40, and IL-23. In accor-dance with the present study, IL-12p40 and IL-23 can be excludedto be crucial for infection with higher doses of Salmonella becausethere was no phenotypic difference at those infective doses be-tween mice lacking IL-12p75 and mice lacking IL-12p40 andIL-23.
In contrast to higher inocula, at relatively low bacterial infectiondoses between 50 and 5000 CFU an IL-12p40-dependent activity(i.e., IL-12p40 and/or IL-23) seems to contribute to the control ofSalmonella infection, because survival rate and survival time weresignificantly higher and the bacterial burden in spleen and liversignificantly lower in IL-12p35�/� compared with IL-12p40�/�
mice. Because total organ burden and the frequency of infectedphagocytes were elevated in the absence of IL-12p40 already atdays 3 and 4 (Figs. 2A and 4A), IL-12p40 appears to play a role
FIGURE 8. Histopathological examination of formalin-fixed paraffin-embedded and H&E-stained liver tissue sections at day 20 p.i. Top panel,Liver from wild-type mice contained very few and small-sized mononu-clear granulomas; middle panel, liver from IL-12p35�/� mice showed alarge number of weakly condensed mononuclear granulomas of variablesize; bottom panel, liver sections from IL-12p40�/� mice showed an in-termediate number of highly condensed mononuclear granulomas of vari-able size.
FIGURE 9. Role of CD4� and CD8� cells in granuloma formation inIL-12p35�/� mice following infection with S. Enteritidis for 20 days. Thehistopathological examination of formalin-fixed, paraffin-embedded, andH&E-stained liver tissue sections at day 20 p.i. revealed a significant in-crease of weakly condensed medium- to large-sized mixed cell granulomasin CD4�-depleted (top panel) and of highly condensed medium- to large-sized mixed cell granulomas in CD8�-depleted IL-12p35�/� mice (bottompanel).
5312 IL-12p40-DEPENDENT AGONISTIC EFFECTS IN MURINE SALMONELLOSIS
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
already during the early innate immune response to activate mac-rophages or other phagocytes for Salmonella killing. TheIL-12p40-dependent stimulatory effect on innate immunity can be
observed for a wide range of infective doses of even up to 107 CFU(data not shown). However, at those high doses this early IL-12p40-dependent protective effect does not lead to long-term
FIGURE 10. Real-time RT-PCR analysis of IL-12p40 or IL-23p19 mRNA expression by splenocytes from S. Enteritidis-infected BALB/c mice at day20 p.i. demonstrated that S. Enteritidis infection caused only induction of IL-12p40 (C and E) but not of IL-23p19 (D and F) mRNA in wild-type andIL-12p35�/� mice. In IL-12p40�/� mice IL-23p19 mRNA was not expressed (D and F) but an IL-12p40 transcript was amplified (C and E �) which, basedon the p40 knockout construction, represented the precisely spliced mRNA without the transcript from exon 3. This mRNA does not encode biologicallyactive IL-12p40 (22). Amplification plots for either IL-12p40 and �-actin (A) or IL-23p19 and �-actin (B) show the increase in SYBR Green fluorescencefor each cDNA sample (shown is one of duplicate amplifications per sample). The cDNAs for IL-12p40 or IL-23p19 amplification were derived fromIFN-��LPS-stimulated primary bone marrow macrophages or from the macrophage cell line RAW264.7 (ATCC), respectively. Threshold cycle (CT)values for each sample were calculated from the linear part of the amplification plots by the second derivative maximum algorithm and the arithmeticbaseline adjustment using LightCycler software. IL-12p40 or IL-23p19 CT values were normalized against �-actin, and normalized values of S. Enteritidis-infected mice were compared with those of naive animals of the respective genotype using the comparative ��CT method (see Materials and Methods).Data are from one of three experiments with similar results.
5313The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

survival (see Fig. 1) suggesting that at those high infective dosesIL-12p75 plays a major role for long-term protection. It has still tobe investigated whether IL-12p40 is able to stimulate the phago-cytes directly or indirectly for secretion of other proinflammatorycytokines such as TNF-� or IFN-�. Of particular interest withinthis frame appears the role of macrophage-derived IFN-�, whichwas recently described in other systems (32, 33). Our data supportthe hypothesis that the IL-12p40-dependent activity is biologicallyrelevant in immune responses toward low-dose infections (e.g.,critical in persistent Salmonella infection) and that high-dose in-fections also require IL-12p75.
The IL-12p40-dependent promoting activity on innate cellularimmune mechanisms is not related to the Nramp1 genotype. Miceof both the resistant and the susceptible genetic backgrounddeveloped a Th2-biased immune response against S. Enteritidis inthe absence of IL-12p40. This is in agreement with previouslypublished data showing similar cytokine response profiles inItyr and Itys congenic mouse strains following infection withS. Dublin (34).
IL-12p75 has previously been shown to be the key factor forTh1 cell differentiation (10–12). Intracellular pathogens such asSalmonella induce predominantly type 1 immune responses, whichinclude differentiation and activation of pathogen-specific Th1and/or Tc1 cells (3). Therefore, type 1 response-inducing cyto-kines (e.g., IL-12p75) or effector cytokines (e.g., IFN-�, TNF-�)are often crucial in intracellular infections. This was confirmed forIL-12 (30, 31), IFN-� (29, 35–38), and TNF-� (29, 37, 39–41) inmurine salmonellosis. To study a potential promoting effect of IL-12p40 on the development of the S. Enteritidis-induced type 1response we have analyzed the concentrations of IFN-� andTNF-� vs IL-4 or IgE in splenocyte supernatants and sera. Theresults demonstrate a significant Th2 shift in S. Enteritidis-infectedIL-12p40-deficient BALB/c and even more pronounced in129Sv/Ev mice, characterized by higher in vivo IgE but lower invivo and ex vivo IFN-� levels. Thus, not only IL-12p75 contrib-utes to Th1 induction during salmonellosis but also IL-12p40. Thisis in agreement with results from a murine CMV model showingthat endogenous IL-12p40 in the absence of IL-12p75 appears tostimulate early IFN-� and TNF-� responses (42).
Histopathological analysis also provided evidence for early andlate effects mediated by IL-12p40. At day 4 p.i. no mononucleargranulomas were formed in the absence of IL-12p40. This is rem-iniscent of our previously published finding in IL-12p40-deficientmice infected with C. neoformans (19). In infection with this fun-gal pathogen granuloma formation was found to depend on IL-12p40 (19). Moreover, homodimeric IL-12p40 was recently shownto exert chemotactic activity on macrophages in vitro (43). In S.Enteritidis infection later neutrophils appear to compensate for thedefect in formation of mononuclear granulomas. In addition, T celldepletion in infected IL-12p35�/� mice, especially the depletionof CD8� T cells, leads to loss of structured mononuclear granu-lomas, which is followed by the appearance of condensed necroticfoci (see Fig. 9) very similar to IL-12p40�/� mice (see Fig. 8).This suggests that T cells are important sources for cytokines (e.g.,TNF-�) required for the recruitment of mononuclear cells andgranuloma formation.
There are three derivatives of the “p40 cytokine family” de-scribed so far: 1) IL-12p75 (f p40 � p35) (5, 6), 2) IL-12(p40)2
(13–15, 19), and 3) IL-23 (f p40 � p19) (18). It was previouslyshown in vitro that mouse IL-12(p40)2 is 25- to 50-fold more po-tent as an IL-12p75 antagonist than mouse IL-12p40 monomer(14). Mice lacking the p35 gene are deficient in IL-12p75 produc-tion but are capable of producing (p40)2 and IL-23. In contrast,mice lacking the p40 gene are unable to produce IL-12p75, IL-
12(p40)2, and IL-23. Thus, the results from this study do not allowa differentiation between IL-12p40- (monomer or homodimer) andIL-23-mediated functions. However, real-time RT-PCR measure-ment showed that neither in wild-type nor in IL-12p35�/� miceIL-23p19 is induced at the mRNA level, whereas IL-12p40 mRNAis strongly up-regulated in both wild-type and IL-12p35�/� miceat day 20 p.i. This points to IL-12p40 as the critical immuno-stimulatory cytokine and does not support an involvement of IL-23. Data from a murine transplantation model (20) and a murineparamyxoviral bronchitis model (44) also point to homodimericIL-12p40 as the active compound. Considering these studies andthe higher affinity of IL-12(p40)2 vs IL-12p40, it is likely thathomodimeric IL-12p40 but not monomeric IL-12p40 is the activemolecule that mediates the protective effects in Salmonella-in-fected IL-12p35�/� mice. To define the active p40-dependentcompound experiments are planned to treat infected IL-12p35/40�/� mice with rIL-12(p40)2 and, for comparison, with rIL-23.
The absence of detectable p19 mRNA expression as describedabove is not sufficient for final exclusion of a potential role ofIL-23 in immunity to Salmonella infection. In further investiga-tions we have to neutralize p19 or specifically IL-23 or use p19�/�
mice to address the question more directly, whether the IL-12p40-dependent protective mechanism(s) on immunity against S. Enter-itidis are mediated by IL-12(p40)2 or IL-23.
AcknowledgmentsWe gratefully acknowledge R. Haack for excellent technical assistance.Furthermore, we thank Drs. H. Gallati (Hoffmann-LaRoche, Basel, Swit-zerland) and G. R. Adolf (Ernst Boehringer Institute, Vienna, Austria) forproviding reagents, Dr. H. Mossmann (Max-Planck-Institute for Immuno-biology, Freiburg, Germany) for providing the animal breeding pairs, andDrs. H.-J. Selbitz, S. Springer, and T. Lindner (Impfstoffwerk Dessau-Tornau, Rosslau, Germany) for providing the S. Enteritidis strains. Wethank Dr. R. K. Straubinger for critically reading the manuscript.
References1. Collins, F. M., G. B. Mackaness, and R. V. Blanden. 1966. Infection-immunity
in experimental salmonellosis. J. Exp. Med. 124:601.2. Hornick, R. B., S. E. Greisman, T. E. Woodward, H. L. DuPont, A. T. Dawkins,
and M. J. Snyder. 1970. Typhoid fever: pathogenesis and immunologic control.N. Engl. J. Med. 283:686.
3. Kaufmann, S. H. E., and G. Kaplan. 1996. Immunity to intracellular bacteria. Res.Immunol. 147:487.
4. Springer, S., J. Lehmann, T. Lindner, J. Thielebein, G. Alber, and H.-J. Selbitz.2000. A new live Salmonella enteritidis vaccine for chickens: experimental ev-idence of its safety and efficacy. Berl. Munch. Tierarztl. Wochenschr. 113:246.
5. Kobayashi, M., L. Fitz, M. Ryan, R. M. Hewick, S. C. Clark, S. Chan, R. Loudon,F. Sherman, B. Perussia, and G. Trinchieri. 1989. Identification and purificationof natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologiceffects on human lymphocytes. J. Exp. Med. 170:827.
6. Stern, A. S., F. J. Podlaski, J. D. Hulmes, Y. C. Pan, P. M. Quinn, A. G. Wolitzky,P. C. Familletti, D. L. Stremlo, T. Truitt, and R. Chizzonite. 1990. Purification tohomogeneity and partial characterization of cytotoxic lymphocyte maturation fac-tor from human B-lymphoblastoid cells. Proc. Natl. Acad. Sci. USA 87:6808.
7. Ma, X., M. Aste-Amezaga, G. Gri, F. Gerosa, and G. Trinchieri. 1997. Immu-nomodulatory functions and molecular regulation of IL-12. Chem. Immunol.68:1.
8. D’Andrea, A., M. Rengaraju, N. M. Valiante, J. Chehimi, M. Kubin, M. Aste,S. H. Chan, M. Kobayashi, D. Young, and E. Nickbarg. 1992. Production ofnatural killer cell stimulatory factor (interleukin-12) by peripheral blood mono-nuclear cells. J. Exp. Med. 176:1387.
9. Murphy, F. J., M. P. Hayes, and P. R. Burd. 2000. Disparate intracellular pro-cessing of human IL-12 preprotein subunits: atypical processing of the p35 signalpeptide. J. Immunol. 164:839.
10. Germann, T., M. K. Gately, D. S. Schoenhaut, M. Lohoff, F. Mattner, S. Fischer,S. C. Jin, E. Schmitt, and E. Rude. 1993. Interleukin-12/T cell stimulating factor,a cytokine with multiple effects on T helper type 1 (Th1) but not on Th2 cells.Eur. J. Immunol. 23:1762.
11. Hsieh, C. S., S. E. Macatonia, C. S. Tripp, S. F. Wolf, A. O’Garra, andK. M. Murphy. 1993. Development of TH1 CD4� T cells through IL-12 pro-duced by Listeria-induced macrophages. Science 260:547.
12. Trinchieri, G. 1993. Interleukin-12 and its role in the generation of TH1 cells.Immunol. Today 14:335.
5314 IL-12p40-DEPENDENT AGONISTIC EFFECTS IN MURINE SALMONELLOSIS
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

13. Mattner, F., S. Fischer, S. Guckes, S. Jin, H. Kaulen, E. Schmitt, E. Rude, andT. Germann. 1993. The interleukin-12 subunit p40 specifically inhibits effects ofthe interleukin-12 heterodimer. Eur. J. Immunol. 23:2202.
14. Gillessen, S., D. Carvajal, P. Ling, F. J. Podlaski, D. L. Stremlo, P. C. Familletti,U. Gubler, D. H. Presky, A. S. Stern, and M. K. Gately. 1995. Mouse interleu-kin-12 (IL-12) p40 homodimer: a potent IL-12 antagonist. Eur. J. Immunol. 25:200.
15. Gately, M. K., D. M. Carvajal, S. E. Connaughton, S. Gillessen, R. R. Warrier,K. D. Kolinsky, V. L. Wilkinson, C. M. Dwyer, G. F. Higgins, F. J. Podlaski, etal. 1996. Interleukin-12 antagonist activity of mouse interleukin-12 p40 ho-modimer in vitro and in vivo. Ann. NY Acad. Sci. 795:1.
16. Mattner, F., L. Ozmen, F. J. Podlaski, V. L. Wilkinson, D. H. Presky,M. K. Gately, and G. Alber. 1997. Treatment with homodimeric interleukin-12(IL-12) p40 protects mice from IL-12-dependent shock but not from tumor ne-crosis factor-�-dependent shock. Infect. Immun. 65:4734.
17. Szabo, S. J., A. S. Dighe, U. Gubler, and K. M. Murphy. 1997. Regulation of theinterleukin (IL)-12R�2 subunit expression in developing T helper 1 (Th1) andTh2 cells. J. Exp. Med. 185:817.
18. Oppmann, B., R. Lesley, B. Blom, J. C. Timans, Y. Xu, B. Hunte, F. Vega,N. Yu, J. Wang, K. Singh, et al. 2000. Novel p19 protein engages IL-12p40 toform a cytokine, IL-23, with biological activities similar as well as distinct fromIL-12. Immunity 13:715.
19. Decken, K., G. Kohler, K. Palmer-Lehmann, A. Wunderlin, F. Mattner,J. Magram, M. K. Gately, and G. Alber. 1998. Interleukin-12 is essential for aprotective Th1 response in mice infected with Cryptococcus neoformans. Infect.Immun. 66:4994.
20. Piccotti, J. R., S. Y. Chan, K. Li, E. J. Eichwald, and D. K. Bishop. 1997.Differential effects of IL-12 receptor blockade with IL-12 p40 homodimer on theinduction of CD4� and CD8� IFN-�-producing cells. J. Immunol. 158:643.
21. Mattner, F., J. Magram, J. Ferrante, P. Launois, K. Di Padova, R. Behin,M. K. Gately, J. A. Louis, and G. Alber. 1996. Genetically resistant mice lackinginterleukin-12 are susceptible to infection with Leishmania major and mount apolarized Th2 cell response. Eur. J. Immunol. 26:1553.
22. Magram, J., S. E. Connaughton, R. R. Warrier, D. M. Carvajal, C. Y. Wu,J. Ferrante, C. Stewart, U. Sarmiento, D. A. Faherty, and M. K. Gately. 1996.IL-12-deficient mice are defective in IFN-� production and type 1 cytokine re-sponses. Immunity 4:471.
23. Piccotti, J. R., K. Li, S. Y. Chan, J. Ferrante, J. Magram, E. J. Eichwald, andD. K. Bishop. 1998. Alloantigen-reactive Th1 development in IL-12-deficientmice. J. Immunol. 160:1132.
24. Martin, G., I. Hanel, R. Helmuth, A. Schroeter, W. Erler, and H. Meyer. 1996.Immunization with potential Salmonella enteritidis mutants. 1. Production and invitro characterization. Berl. Munch. Tierarztl. Wochenschr. 109:325.
25. Martin, G., U. Methner, G. Steinbach, and H. Meyer. 1996. Immunization withpotential Salmonella enteritidis mutants. 2. Investigations on the attenuation andimmunogenicity for mice and young hens. Berl. Munch. Tierarztl. Wochenschr.109:369.
26. Kruisbeek, A. M. 1991. In vivo depletion of CD4- and CD8-specific T cells. InCurrent Protocols in Immunology. J. E. Coligan, A. M. Kruisbeek, D. H. Mar-gulies, E. M. Shevach, and W. Strober, eds. Wiley, New York, pp. 4.1.1–4.1.5.
27. Comparative CT Method (Separate Tubes). 1997. PE Applied Biosystems. UserBulletin 2: ABI PRISM 7700 Sequence Detection System.
28. Nauciel, C. 1990. Role of CD4� T cells and T-independent mechanisms in ac-quired resistance to Salmonella typhimurium infection. J. Immunol. 145:1265.
29. Mastroeni, P., B. Villarreal-Ramos, and C. E. Hormaeche. 1992. Role of T cells,TNF-� and IFN-� in recall of immunity to oral challenge with virulent salmo-nellae in mice vaccinated with live attenuated aro- Salmonella vaccines. Microb.Pathog. 13:477.
30. Mastroeni, P., J. A. Harrison, J. A. Chabalgoity, and C. E. Hormaeche. 1996.Effect of interleukin-12 neutralization on host resistance and � interferon pro-duction in mouse typhoid. Infect. Immun. 64:189.
31. Kincy-Cain, T., J. D. Clements, and K. L. Bost. 1996. Endogenous and exoge-nous interleukin-12 augment the protective immune response in mice orally chal-lenged with Salmonella dublin. Infect. Immun. 64:1437.
32. Munder, M., M. Mallo, K. Eichmann, and M. Modolell. 1998. Murine macro-phages secrete interferon-� upon combined stimulation with interleukin (IL)-12and IL-18: a novel pathway of autocrine macrophage activation. J. Exp. Med.187:2103.
33. Xing, Z., A. Zganiacz, and M. Santosuosso. 2000. Role of IL-12 in macrophageactivation during intracellular infection: IL-12 and mycobacteria synergisticallyrelease TNF-� and nitric oxide from macrophages via IFN-� induction. J. Leu-kocyte Biol. 68:897.
34. Eckmann, L., J. Fierer, and M. F. Kagnoff. 1996. Genetically resistant (Ityr) andsusceptible (Itys) congenic mouse strains show similar cytokine responses fol-lowing infection with Salmonella dublin. J. Immunol. 156:2894.
35. Ramarathinam, L., D. W. Niesel, and G. R. Klimpel. 1993. Salmonella typhi-murium induces IFN-� production in murine splenocytes: role of natural killercells and macrophages. J. Immunol. 150:3973.
36. Muotiala, A., and P. H. Makela. 1993. Role of � interferon in late stages ofmurine salmonellosis. Infect. Immun. 61:4248.
37. Nauciel, C., and F. Espinasse-Maes. 1992. Role of � interferon and tumor ne-crosis factor-� in resistance to Salmonella typhimurium infection. Infect. Immun.60:450.
38. Pie, S., P. Truffa-Bachi, M. Pla, and C. Nauciel. 1997. Th1 response in Salmo-nella typhimurium-infected mice with a high or low rate of bacterial clearance.Infect. Immun. 65:4509.
39. Mastroeni, P., J. N. Skepper, and C. E. Hormaeche. 1995. Effect of anti-tumornecrosis factor-� antibodies on histopathology of primary Salmonella infections.Infect. Immun. 63:3674.
40. Tite, J. P., G. Dougan, and S. N. Chatfield. 1991. The involvement of tumornecrosis factor in immunity to Salmonella infection. J. Immunol. 147:3161.
41. Everest, P., M. Roberts, and G. Dougan. 1998. Susceptibility to Salmonella ty-phimurium infection and effectiveness of vaccination in mice deficient in thetumor necrosis factor-� p55 receptor. Infect. Immun. 66:3355.
42. Carr, J. A., J. A. Rogerson, M. J. Mulqueen, N. A. Roberts, and A. A. Nash. 1999.The role of endogenous interleukin-12 in resistance to murine cytomegalovirus(MCMV) infection and a novel action for endogenous IL-12 p40. J. InterferonCytokine Res. 19:1145.
43. Ha, S. J., C. H. Lee, S. B. Lee, C. M. Kim, K. L. Jang, H. S. Shin, and Y. C. Sung.1999. A novel function of IL-12p40 as a chemotactic molecule for macrophages.J. Immunol. 163:2902.
44. Walter, M., N. Kajiwara, P. Karanja, M. Castro, and M. Holtzman. 2001. Inter-leukin-12 p40 production by barrier epithelial cells during airway inflammation.J. Exp. Med. 193:339.
5315The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from