studentsrepo.um.edu.mystudentsrepo.um.edu.my/10359/4/omar.pdfiii. abstract . malaria, especially ....
TRANSCRIPT

MOLECULAR EPIDEMIOLOGY OF MALARIA AND
DETECTION OF ANTI-MALARIAL DRUG RESISTANCE-
ASSOCIATED MARKERS (PFCRT, PFMDR-1, PFDHFR
AND PFDHPS) IN HADHRAMOUT GOVERNORATE,
YEMEN
OMAR ABDULLAH ALI BAMAGA
FACULTY OF MEDICINE
UNIVERSITY OF MALAYA
KUALA LUMPUR
2017 Univers
ity of
Mala
ya

MOLECULAR EPIDEMIOLOGY OF MALARIA AND
DETECTION OF ANTI-MALARIAL DRUG RESISTANCE-
ASSOCIATED MARKERS (PFCRT, PFMDR-1, PFDHFR
AND PFDHPS) IN HADHRAMOUT GOVERNORATE,
YEMEN
OMAR ABDULLAH ALI BAMAGA
THESIS SUBMITTED IN FULFILMENT OF THE
REQUIREMENTS FOR THE DEGREE OF DOCTOR OF
PHILOSOPHY
FACULTY OF MEDICINE
UNIVERSITY OF MALAYA
KUALA LUMPUR
2017
Univers
ity of
Mala
ya

UNIVERSITI MALAYA
ORIGINAL LITERARY WORK DECLARATION
Name of Candidate: Omar Abdullah Ali Bamaga
Registration / Matric No: MHA100059
Name of Degree: DOCTOR OF PHILOSOPHY
Title of Project Paper/Research Report/Dissertation/Thesis (“this Work”):
Molecular epidemiology of malaria and detection of anti-malarial drug resistance-
associated markers (Pfcrt, pfmdr-1, pfdhfr and pfdhps) in Hadhramout
governorate, Yemen
Field of Study: Parasitology
I do solemnly and sincerely declare that:
(1) I am the sole author/writer of this Work;
(2) This Work is original;
(3) Any use of any work in which copyright exists was done by way of fair dealing and
for permitted purposes and any excerpt or extract from,or reference to or
reproduction of any copyright work has been disclosed expressly and sufficiently
and the title of the work and its authorship have been acknowledged in this Work;
(4) I do not have any actual knowledge nor do I ought reasonably to know that the
making of this work constitutes an infringement of any copyright work;
(5) I hereby assign all and every rights in the copyright to this work to the University of
Malaya (“UM”), who henceforth shall be owner of the copyright in this work and
that any reproduction or use in any form or by any means whatsoever is prohibited
without the written consent of UM having been first had and obtained;
(6) I am fully aware that if in the course of making this work I have infringed any
copyright whether intentionally or otherwise, I may be subject to legal action or any
other action as may be determined by UM.
Candidate’s Signature Date
Subscribed and solemnly declared before,
Witness’s Signature Date
Name: Dr. Yvonne Lim Ai Lian
Designation: Professor
Department of Parasitology, Faculty of Medicine, University of Malaya
Univers
ity of
Mala
ya

iii
ABSTRACT
Malaria, especially Plasmodium falciparum malaria is one of the main causes of
mortality and morbidity worldwide. Yemen is an Eastern Mediterranean country where
68% of its population is at risk of malaria. In 2013, it was estimated that there were
150,000 cases recorded in Yemen with 55 malarial deaths, compared to 900,000 cases
in 2000. The anti-malarial treatment policy in Yemen was changed from chloroquine
(CQ) to artemisinin combination therapy (ACT) in 2005.The present study is the first in
Hadhramout, Yemen which aimed to assess the epidemiology of malaria parasites and
to determine the frequency of mutant alleles and genotypes associated with antimalarial
drug resistance in Plasmodium falciparum isolates. Blood specimens were collected
from seven villages in two different districts of the Hadhramout governorate by house-
to-house visits from July 2011 to May 2012. A total of 735 individuals aged 1 to 75
years with a median of 16 years and 22 interquartile range participated in the study. A
pre-tested questionnaire was used to gather demographic, socioeconomic and
environmental data. Plasmodium species were first identified by microscopy
examination and subsequently genomic DNA was extracted from dried archive blood
spots of P. falciparum isolates and analyzed using nested PCR. Mutation-specific nested
polymerase chain reaction (MS-PCR) and restriction fragment length polymorphism
(PCR–RFLP) methods were used to investigate the mutations in the Pfmdr1 (codons 86
and 1246) and Pfcrt (codons 76, 271, 326, 356 and 371) genes. DNA was also amplified
using nested PCR and subsequently sequenced for Pfdhfr and Pfdhps genes. Sequences
were analyzed for mutations in Pfdhfr at codons 51, 59, 108, and 164 and in Pfdhps at
codons 436, 437, and 540. Results of the overall prevalence of malaria parasites in
Hadhramout governorate, Yemen via microscopy was 18.8% (138 of 735) with
Plasmodium falciparum being the predominant species (99.3%; 137 of 138), followed
by Plasmodium vivax (0.7%; 1). Nested PCR detected P. falciparum in four samples
Univers
ity of
Mala
ya

iv
that were previously negative using microscopy. The combination of microscopy and
nested PCR detection resulted in three samples being identified as mixed infections of
P. falciparum and P. vivax. The infection rate was higher in Al-Raydah-Qusyer district
(21.8%) compared to Hajer district (11.8%). Fifty two percent of those positive for
Plasmodium were asymptomatic with low parasite density. The adults had a higher
infection rate as compared to children. Univariate analysis identified those whose
household’s heads are fishermen (OR = 11.3, 95% CI: 3.13–40.5) and farmers (OR =
4.84, 95% CI: 1.73–13.6) as high-risk groups. A higher number of positive rates were
observed in people living in houses with uncemented brick walls (OR = 2.1, 95% CI:
1.32–3.30), without access to toilets (OR = 1.6, 95% CI: 1.05–2.32), without a fridge
(OR = 1. 6, 95% CI: 1.05–2.30), or without TV (OR = 1. 6, (95% CI: 1.05–2.30).
People living in houses with water collection points located less than 200 meters away
were also at higher risk of acquiring malaria (OR = 1.6, 95% CI:1.05–2.30). Knowledge
about the importance of using insecticide-treated mosquito nets (ITNs) and indoor
residual spraying (IRS) for prevention of malaria was 7% and 2%, respectively. The
prevalence of Pfcrt mutations at codons 76, 271, 326 and 371 were 50.4%, 58.7%,
54.3% and 44.9%, respectively. All isolates had wild-type Pfcrt 356 allele. The majority
of Pfmdr1 86 alleles (83.3%) and all Pfmdr1 1246 (100%) alleles were also wild type.
There was no association between Pfcrt mutations and symptomatology, gender and age
groups. For Pfdhfr/Pfdhps mutations, each Pfdhfr mutant allele (I51 and N108) in P.
falciparum isolate had a frequency of 84%. Pfdhfr R59 mutant allele was detected in one
isolate. Pfdhps at codon G437 mutant allele was detected in 44.7% of Plasmodium
falciparum malaria isolates. Frequencies of Pfdhfr double mutant genotype
(I51C59N108I164) and Pfdhfr/Pfdhps triple mutant genotype (I51C59N108I164-S436G437K540)
were 82.8% and 40.6%, respectively. It is important to note that there was one isolate
each which harbored Pfdhfr triple mutant genotype (I51, R59, N108, I164) and
Univers
ity of
Mala
ya

v
Pfdhfr/Pfdhps quadruple mutant genotype (I51R59N108I164-S436G437K540). In conclusion,
several environmental, socioeconomic and behavioral issues were discovered to be the
contributing factors to the high prevalence of malaria in this southeast Yemen
governorate. High frequencies of point mutations in codons 76, 271, 326 and 371 of P.
falciparum, suggested a sustained high CQ resistance even after 6 years of shifting to
ACTs. High frequencies of Pfdhfr and Pfdhps mutant alleles and genotypes in P.
falciparum isolates from Hadhramout, Yemen, highlight the risk of decreasing efficacy
of sulfadoxine pyrimethamine antimalarial drugs. Novel strategies adapted to local
situations need to be established in order to improve the effectiveness of malaria
control. The current study findings necessitate continuous monitoring of the efficacy of
malaria treatment.
Univers
ity of
Mala
ya

vi
ABSTRAK
Malaria, terutamanya malaria Plasmodium falciparum merupakan salah satu faktor
utama mortaliti dan morbiditi di seluruh dunia. Yemen adalah sebuah negara yang
terletak di timur Mediterranean di mana 68% daripada penduduknya adalah berisiko
tinggi untuk dijangkiti malaria. Pada tahun 2013, dianggarkan bahawa 150,000 kes
malaria direkodkan di Yemen dengan 55 kematian berbanding dengan 900,000 kes pada
tahun 2000. Polisi rawatan anti-malaria di Yemen telah ditukar daripada chloroquine
(CQ) kepada terapi gabungan artemisinin (ACT) pada tahun 2005. Kajian ini merupakan
yang pertama di Hadhramout, Yemen untuk mengkaji epidemiologi parasit malaria serta
penentuan kekerapan alel mutan dan genotip yang dikaitkan dengan rintangan ubat anti-
malaria dalam pencilan. Plasmodium falciparum. Spesimen darah telah diambil dari
tujuh kampung di dua daerah yang berlainan di wilayah Hadhramout dari rumah ke
rumah dari Julai 2011 hingga Mei 2012. Sejumlah 735 individu berusia di antara 1
hingga 75 tahun dengan median umur 16 tahun and julat interquartile 22 telah terlibat
dalam kajian ini. Borang pra-soalselidik berdasarkan isi rumah telah digunakan untuk
mengumpul demografi, sosio-ekonomi dan data alam sekitar. Pertama sekali, spesies
Plasmodium dikenalpasti melalui pemeriksaan mikroskop. DNA genomik kemudian
diekstrak daripada tompok darah arkib pencilan P. falciparum untuk analisis tindak
balas berantai polimerase (PCR). Kaedah mutase-spesifik reaksi bersarang rantaian
polimerase (MS-PCR) dan sekatan panjang serpihan polymorphism (PCR-RFLP) telah
digunakan untuk mengenalpasti mutasi pada gen Pfmdr1 (kodon 86 dan 1246) dan Pfcrt
(kodon 76, 271, 326, 356 dan 371). DNA juga telah diamplifikasi menggunakan kaedah
PCR bersarang dan kemudiannya gen Pfdhfr dan Pfdhps dijujuk. Jujukan DNA
dianalisis untuk mutasi dalam gen Pfdhfr pada kodon 51, 59, 108, dan 164 dan di gen
Pfdhps pada kodon 436, 437, dan 540. Keputusan keseluruhan prevalen malaria di
Hadhramout, Yemen melalui kaedah pemeriksaan mikroskop adalah 18.8% (138 dari
Univers
ity of
Mala
ya

vii
735) dengan spesies P. falciparum sebagai spesies utama (99.3%; 137 dari 138) diikuti
dengan Plasmodium vivax (0.7%; 1). PCR bersarang mengesan P. falciparum dalam
empat sampel yang sebelum ini negatif menggunakan mikroskop. Gabungan
penggunaan mikroskop dan pengesanan PCR bersarang menemui tiga sampel yang
dikenalpasti sebagai jangkitan campuran P. falciparum dan P. vivax. Kadar jangkitan
adalah lebih tinggi di daerah Al-Raydah-Qusyer (21.8%) berbanding dengan daerah
Hajer (11.8%). Lima puluh dua peratus daripada individu yang positif untuk
Plasmodium adalah asimptomatik dengan kepadatan parasit yang rendah. Orang dewasa
mempunyai peratusan jangkitan yang lebih tinggi berbanding kanak-kanak. Analisis
univariat mengenalpasti individu di mana ketua rumahnya adalah nelayan (OR = 11.3,
95% CI: 3.13-40.5) dan petani (OR = 4.84, 95% CI: 1.73-13.6) sebagai kumpulan
berisiko tinggi. Peratusan positif didapati lebih tinggi bagi orang yang tinggal di rumah
yang berdinding tidak bersimen (OR = 2.1, 95% CI: 1.32-3.30), tidak mempunyai
tandas (OR = 1.6, 95% CI: 1.05-2.32), tidak mempunyai peti sejuk (OR = 1. 6, 95% CI:
1.05-2.30), atau tidak mempunyai televisyen (OR = 1. 6, 95% CI:. 1.05-2.30). Individu
yang tinggal di rumah di mana sumber pengumpulan airnya terletak kurang daripada
200 meter juga berisiko tinggi untuk dijangkiti malaria (OR = 1.6, 95% CI: 1.05-2.30)
Pengetahuan tentang kepentingan menggunakan insecticide-treated mosquito nets
(ITNs) dan indoor residual spraying (IRS) untuk pencegahan malaria adalah 7% dan
2%, masing-masing. Prevalen mutasi Pfcrt pada kodon 76, 271, 326 dan 371 adalah
50.4%, 58.7%, 54.3% dan 44.9%, masing-masing. Semua pencilan mempunyai alel
Pfcrt 356 jenis liar. Majoriti Pfmdr1 86 alel (83.3%) dan semua alel Pfmdr1 1246
(100%) juga adalah jenis liar. Tidak ada kaitan antara mutasi Pfcrt dengan
simptomatologi, jantina dan kumpulan umur. Untuk mutasi Pfdhfr / Pfdhps, setiap alel
mutan Pfdhfr (I51 and N108) dalam pencilan P. falciparum mempunyai kekerapan 84%.
Alel mutan Pfdhfr R59 ditemui hanya dalam satu pencilan. Alel mutan kodon G437
Univers
ity of
Mala
ya

viii
dikesan pada 44.7% daripada pencilan malaria Plasmodium falciparum. Kekerapan
Pfdhfr genotip dua mutan (I51C59N108I164) dan Pfdhfr / Pfdhps genotip tiga mutan
(I51C59N108I164-S436G437K540) adalah 82.8% dan 40.6%, masing-masing. Terdapat satu
pencilan setiap satu untuk Pfdhfr genotip tiga mutan (I51, R59, N108, I164) dan Pfdhfr /
Pfdhps genotip empat mutan (I51R59N108I164-S436G437K540). Kesimpulannya, beberapa
isu alam sekitar, sosio-ekonomi dan tingkah laku telah ditemui sebagai faktor yang
menyumbang kepada kes malaria yang tinggi di wilayah tenggara Yemen. Frekuensi
tinggi mutasi titik dalam kodon 76, 271, 326 dan 371 Pfcrt dan Pfdhfr / Pfdhps alel
mutan P. falciparum, menunjukkan rintangan tinggi terhadap CQ adalah berterusan
walaupun telah 6 tahun beralih kepada ACT. Frekuensi alel mutan dan genotip Pfdhfr
and Pfdhps yang tinggi dalam pencilan P. falciparum di Hadhramout, Yemen,
menunjukkan risiko penurunan effikasi ubat anti-malaria sulfadoxine pyrimethamine.
Strategi dan langkah pengawalan yang baru yang bersesuaian dengan keadaan tempatan
perlu diwujudkan dalam usaha untuk meningkatkan keberkesanan kawalan malaria.
Hasil daripada kajian ini memerlukan pemantauan berterusan terhadap tahap
keberkesanan rawatan malaria.
Univers
ity of
Mala
ya

ix
ACKNOWLEDGEMENTS
First and foremost, I would like to thank Allah for all the wonderful blessings and
giving me the courage, guidance, strength and perseverance throughout the duration of
my whole life.
I would like to express my deepest appreciation and extend my profound gratitude to
my supervisors, Professor Dr. Yvonne Lim Ai Lian and Associate Professor Dr.
Mohammed Mahdy for their support, assistance and guidance during the course of this
study. Their advice, boundless ideas, skills, expertise, comments, criticism,
encouragement and challenges were very much appreciated. You will forever remain an
indelible part of my life as mentors.
Special thanks to the head and staff of Department of Parasitology, Faculty of
Medicine, for their support. I thank the University of Malaya for supporting the research
under the University of Malaya Research Grant (UMRG/RG503-13HTM) and the
University of Malaya High Impact Research Grant UM-MOHE UM.C /62 /1/ HIR /
MOHE/MED/18 from the Ministry of Higher Education Malaysia. I would like to thank
all my lecturers, colleagues, and friends for their unending encouragements.
I also would like to thank all the technical staff in the field study and laboratory
expert groups for their assistance in the molecular laboratory of Parasitology
Department, the National Malaria Control Program in Hadhramout governorate Yemen,
Ministry of Health and communities in Hadhramout for their cooperation during this
study.
Univers
ity of
Mala
ya

x
Most importantly, I would like to thank my Mother; my Father, Abdullah Ali; my
wife, my brothers, my sisters for their undivided support, encouragement, assistance and
their prayers and many others who are too numerous to mention here.
I am also thankful to the Hadhramout University of Science and Technology of my
country for the support given to me in terms of scholarship to study in Malaysia.
Finally, I would like to thank everyone for their assistance and prayers. I hope that
this dissertation has provided meaningful ideas and significant contributions that will be
beneficial to the field of malaria and I hope that everyone who reads this dissertation
finds it useful.
Univers
ity of
Mala
ya

xi
TABLE OF CONTENTS
ABSTRACT iii
ABSTRAK vi
ACKNOWLEDGEMENTS ix
TABLE OF CONTENTS xi
LIST OF FIGURES xvi
LIST OF TABLES xviii
LIST OF SYMPOLS AND ABBREVIATIONS xxi
LIST OF APPENDICES xxiv
CHAPTER 1: INTRODUCTION 1
1.1 BACKGROUND OF STUDY 1
1.2 PROBLEM STATEMENTS 3
1.3 RESEARCH HYPOTHESES 5
1.4 OBJECTIVES 6
1.4.1 General objective 6
1.4.2 Specific objectives 6
1.5 SIGNIFICANCE OF STUDY 7
CHAPTER 2: LITERATURE REVIEW 8
2.1 MALARIA 8
2.1.1 History 8
2.1.2 Biology, etiology and life cycle 8
2.1.3 Clinical manifestations 12
2.1.4 Epidemiology 13
2.1.4.1 Malaria burden and geographical distribution of Plasmodium
species
13
Univers
ity of
Mala
ya

xii
2.1.4.2 Factors associated with malaria 17
2.1.5 Diagnosis of malaria 22
2.1.6 Treatment 23
2.1.6.1 Anti-malarial drugs 23
2.1.6.2 Methods of anti-malarial drug resistance surveillance 26
A) In vivo methods 26
B) In vitro method 31
C) Molecular markers 32
2.2 MALARIA IN EASTERN MEDITERRANEAN REGION 35
2.2.1 Current status 35
2.2.2 Anti-malarial drug resistance 38
2.3 MALARIA IN YEMEN 43
2.3.1 Plasmodium species and types of vectors 43
2.3.2 Trend of confirmed malaria cases in the last 23 years 46
2.3.3 Risk factors 48
2.3.4 Malaria distribution and intensity of transmission 52
2.3.5 Prevention and control 53
2.3.6 Malaria diagnosis 53
2.3.7 Malaria treatment in Yemen 54
2.3.7.1 The old strategy (from 1999) 54
2.3.7.2 The new strategy (from 2005) 54
2.3.7.3 Monitoring anti-malarial drug resistance 55
A) In vivo studies 55
B) Molecular markers based studies 55
Univers
ity of
Mala
ya

xiii
CHAPTER 3: METHODOLOGY 57
3.1 OVERVIEW OF STUDY METHODS 57
3.6.2 Ethical clearance 59
3.3 STUDY AREA AND STUDY POPULATIONS 59
3.3 DESIGN OF STUDY 62
3.4 SAMPLE SIZE 62
3.5 DESCRIPTION OF VARIABLES 63
3.6 DATA AND SAMPLE COLLECTION 63
3.6.1 Strategy of field work 63
3.6.2 Questionnaire 63
3.6.3 Blood sampling 64
3.7 DETECTION OF MALARIA PARASITE BY MICROSCOPY METHOD 65
3.7.1 Staining blood smears 65
3.7.2 Microscopy examination 65
3.8 MOLECULAR IDENTIFICATION AND GENOTYPING OF MALARIA
SPECIES
66
3.8.1 DNA extraction 66
3.8.2 Molecular identification of malaria species 66
3.8.3 Molecular detection of mutation in Pfcrt gene at codon K76T 70
3.8.4 Molecular detection of point mutations in Pfcrt gene (positions Q271E,
N326S, I356T, R371I) and Pfmdr1 gene (positions N86Y and
D1246Y)
72
3.8.5 Molecular detection of point mutations in Pfdhfr gene at different
codons
72
3.8.6 Molecular detection of point mutations in Pfdhps gene at different 73
Univers
ity of
Mala
ya

xiv
codons
3.9 STATISTICAL ANALYSIS 76
CHAPTER 4: RESULTS 77
4.1 EPIDEMIOLOGICAL RESULTS OF MALARIA IN THE HADHRAMOUT
GOVERNORATE, YEMEN
77
4.1.1 Characteristic of study population 77
4.1.2 Prevalence of malaria and identify the risk factors associated with
malaria in the Hadhramout governorate, Yemen
80
4.1.3 Assessment of knowledge, attitude and practices (KAP) towards
malaria in the Hadhramout governorate, Yemen
84
4.1.4 Clinical manifestations of individuals positive with malaria 86
4.2 MOLECULAR CHARACTERIZATION OF MALARIA IN THE
HADHRAMOUT GOVERNORATE, YEMEN
89
4.2.1 Malaria parasite identification using nested PCR based on 18SSU
rRNA gene
89
4.2.2 Prevalence and distribution of mutations in Pfcrt gene at 76, 271, 326,
356 and 371 and Pfmdr1 gene at 86 and 1246 as molecular markers
of CQ resistance of Plasmodium falciparum isolates in Hadhramout
governorate, Yemen
91
4.2.3 Prevalence of mutations in Pfdhfr and Pfdhps genes at different codons
as molecular markers of SP resistance of Plasmodium falciparum
isolates in Hadhramout governorate, Yemen
94
CHAPTER 5: DISCUSSION 98
5.1 THE EPIDEMIOLOGICAL OF MALARIA IN THE HADHRAMOUT
GOVERNORATE, YEMEN
98
5.1.1 Prevalence of malaria and identify the risk factors associated with 98
Univers
ity of
Mala
ya

xv
malaria in the Hadhramout governorate, Yemen
5.1.2 Assessment of knowledge, attitude and practices towards malaria in the
Hadhramout governorate, Yemen
100
5.2 MOLECULAR CHARACTERIZATION RESULTS OF MALARIA IN THE
HADHRAMOUT GOVERNORATE, YEMEN
102
5.2.1 Point mutations in Pfcrt gene at 76, 271, 326, 356 and 371 and Pfmdr1
gene at 86 and 1246 as molecular markers of CQ resistance of
Plasmodium falciparum isolates in Hadhramout governorate
102
5.2.2 Point mutations in Pfdhfr and Pfdhps genes at different codons as
molecular markers of sulfadoxine-pyrimethamine resistance of
Plasmodium falciparum isolates in Hadhramout governorate
105
5.3 LIMITATIONS OF STUDY 110
CHAPTER 6: CONCLUSIONS AND RECOMMENDATIONS 111
6.1 CONCLUSIONS 111
6.2 RECOMMENDATIONS 113
References 115
Appendices 148
List of publications and paper presented 182
Univers
ity of
Mala
ya

xvi
LIST OF FIGURES
Figure 2.1: Life cycle of Plasmodium spp. 11
Figure 2.2: The malaria control stage and the countries contributing to the
global death
15
Figure 2.3: The intensity of malaria transmission worldwide 16
Figure 2.4: Modified map of distribution of Anopheles mosquito in
governorates, Yemen
45
Figure 2.5: Malaria trend in Yemen from 1990 till 2014 47
Figure 3.1: Schematic diagram of samples and data collection and molecular
marker detections
58
Figure 3.2: Map of study area highlighted in the Hadhramout governorate,
Yemen
61
Figure 4.1: Malaria prevalence in endemic areas of the two districts (i.e, Al
Raydah-Qusyer and Hajer) of Hadhramout governorate, Yemen
78
Figure 4.2: Prevalence of parasitemia among populations infected with
malaria in Hadhramout governorate, Yemen
88
Figure 4.3: Agarose gel electrophoresis for identification of Plasmodium
species (Secondary nested PCR-genus specific).
178
Figure 4.4: Agarose gel electrophoresis for identification of Plasmodium
falciparum (Secondary nested PCR-species specific)
178
Figure 4.5: Agarose gel electrophoresis for identification of Plasmodium
vivax (Secondary nested PCR-species specific)
178
Figure 4.6: Secondary mutant specific nested PCR of Pfcrt 76 using
restriction enzymes
179
Figure 4.7: Digestion of secondary nested PCR for Pfcrt 271 using 179
Univers
ity of
Mala
ya

xvii
restriction enzymes
Figure 4.8: Digestion of secondary nested PCR for Pfcrt 326 using
restriction enzymes
179
Figure 4.9: Digestion of secondary nested PCR for Pfcrt 356 using
restriction enzymes
179
Figure 4.10: Digestion of secondary nested PCR for Pfcrt 371 using
restriction enzymes
180
Figure 4.11: Digestion of secondary nested PCR for Pfmdr1 86 using
restriction enzymes
180
Figure 4.12: Digestion of secondary nested PCR for Pfmdr1 1246 using
restriction enzymes
180
Figure 4.13: Secondary nested PCR for Pfdhfr 181
Figure 4.14: Secondary nested PCR for Pfdhfr 181
Univers
ity of
Mala
ya

xviii
LIST OF TABLES
Table 2.1: Socioeconomic factors, behavior factors and environmental
factors of malaria
20
Table 2.2: Classification of antimalarial drug and brief outline of mechanism
of action
25
Table 2.3: Definitions of parasitological response to drug in in vivo
therapeutic efficacy studies
29
Table 2.4: Definitions of parasitological and clinical response to drug in in
vivo therapeutic efficacy studies
30
Table 2.5: Most commonly used antimalarial drugs along with their
molecular markers to determine their drug susceptibility /
resistance
34
Table 2.6: Malaria cases in countries with high transmission areas at the
Eastern Mediterranean region in 2013
37
Table 2.7: Summary of some previous studies on mutations of crt, mdr1,
dhfr and dhps genes in Plasmodium isolates and their role in
antimalarial drugs resistance in Mediterranean countries
39
Table 2.8 Socio-economic, behavioral and environmental risk factors
associated with acquiring malaria in four governorates in Yemen
50
Table 3.1: Protocol for the detection of Plasmodium malarial species based
on 18SSU rRNA gene
69
Table 3.2: Detection of point mutations in Pfcrt , Pfmdr1, Pfdhfr, and Pfdhps
genes at different codons
71
Table 3.3: Forward and reverse primers sequences for Pfcrt , Pfmdr1,
Pfdhfr, and Pfdhps genes at different codons
75
Table 4.1: Demographic characteristics of study populations in Hadhramout 79
Univers
ity of
Mala
ya

xix
governorate, Yemen
Table 4.2: Prevalence and distribution of malaria among population in
Hadhramout governorate, Yemen according to age, gender and
areas
82
Table 4.3: Risk factors associated with malaria in Hadhramout governorate,
Yemen
83
Table 4.4: KAPs of malaria in the rural areas of Hadhramout governorate,
Yemen
85
Table 4.5: Clinical manifestations among humans infected with malaria in
Hadhramout governorate, Yemen
87
Table 4.6: Detection of Plasmodium species using nested PCR among
populations infected with malaria in Hadhramout governorate,
Yemen
90
Table 4.7: Frequency and distribution of Pfcrt and Pfmdr1 alleles in P.
falciparum isolates from populations in Hadhramout governorate,
Yemen
92
Table 4.8: Frequency and distribution of Pfcrt and Pfmdr1 allels according
to symptomatology in P. falciparum isolates from populations in
Hadhramout governorate, Yemen
93
Table 4.9: Prevalence of mutant alleles of Pfdhfr and Pfdhps genes in P.
falciparum isolates from populations in Hadhramout governorate,
Yemen
96
Table 4.10: Prevalence of genotypes of Pfdhfr, Pfdhps and combined Pfdhfr–
Pfdhps genes in P. falciparum isolates from populations in
Hadhramout governorate, Yemen
97
Univers
ity of
Mala
ya

xx
Table 6.1: Morphological characteristics of the Plasmodium species
infecting human
173
Univers
ity of
Mala
ya

xxi
LIST OF SYMPOLS AND ABBREVIATIONS
% Percentage
< Less than
> More than
µg/l Microgram per liter
µg/ml Microgram per milliliter
µL Microliter
ACPR Adequate clinical and parasitological response
ACT: Artemisinin-based combination therapy
Ala (A): Alanine
AL: Artemether lumefantrine
Arg (R): Arginine
Asn (N): Asparagine
Asp (D): Aspartate
MS-PCR Mutant-specific nested polymerase chain reaction
AS Artesunate
bp Base pair
C° Degree Celsius
CDC: Centers for Disease Control and Prevention
CI: Confidence interval
Cys: Cysteine
Cyt-b The mitochondrial cytochrome b gene
Dhfr-ts Dihydrofolate reductase-thymidylate synthase
Endo. Dig: Endonuclease digestion
ETF Early treatment failure
g Gram
g/dl Gram per deciliter
Gln (Q): Glutamine
Glu (E): Glutamate
Gly (G): Glycine
Hb: Haemoglobin
IL: Interleukin
Ile (I): Isoleucine
IRS: Indoor Residual Spraying
Univers
ity of
Mala
ya

xxii
ITNs: Insecticide-treated Nets
KAP: Knowledge, Attitude and Practices
kb: Kilobase
LCF Late clinical failure
LLINs: Long Lasting Insecticide-treated Nets
LPF: Late parasitological failure
Lys (K): Lysine
mg/dl Milligram per deciliter
min Minute
ml Milliliter
mM Millimolar
MQ: Mefloquine
n Sample size
nM Nano mole
NMCP: National Malaria Control Programme
OR: Odds-ratio
P: Level of significance
PCR: Polymerase Chain Reaction
Pfcrt :
Plasmodium falciparum chloroquine resistance
transporter
Pfdhfr: Plasmodium falciparum dihydrofolate reductase
Pfdhps: Plasmodium falciparum dihydropteroate synthase
Pfmdr-1: Plasmodium falciparum multidrug resistance gene-1
Pfmrp Plasmodium falciparum multidrug resistance-associated
protein
pfnhe Plasmodium falciparum Sodium Hydrogen Exchanger
Phe (F): Phenylalanine
Post-Dig: Post Digestion
RBC: Red blood cell
RE: Restriction enzyme
RFLP: Restriction fragment length polymorphism
SD: Standard deviation
sec Seconds
Ser (S): Serine
SERCA: Sarco/endoplasmic reticulum Ca2+-ATPase
Univers
ity of
Mala
ya

xxiii
SNP: Single nucleotide polymorphism
SP: Sulfadoxine-pyrimethamine
SPSS: Statistical Package for Social Science
SSU-rRNA: Small subunit ribosomal RNA
TAE: Tris- acetate EDTA
Thr (T): Threonine
TNF: Tumor necrosis factor
Tyr (Y): Tyrosine
U Unit
V Voltage
WHO: World Health Organization
χ2: Chi-square
Univers
ity of
Mala
ya

xxiv
LIST OF APPENDICES
Appendix A: Ethical clearance form (English) 148
Appendix A: Ethical clearance form (Arabic) 151
Appendix B: Consent form (English) 153
Appendix B: Consent form (Arabic) 154
Appendix C: Photography of field study and specimens collection 155
Appendix D: Defination of variables 160
Appendix E: Questionnaire (English) 162
Appendix E: Questionnaire (Arabic) 167
Appendix F: Wright-giemsa stain 172
Appendix G: Key morphological differences between the blood stages of
human Plasmodium species.
173
Appendix G: Photograph of diagnostic stages 174
Appendix H: DNA extraction protocol 175
Appendix I: Gel electrophoresis of PCR products 178
Appendix J: The SNPs alignment of dhfr gene sequences of Plasmodium
falciparum isolates in Hadhramout governorate, Yemen
182
Appendix K: The SNPs alignment of dhps gene sequences of Plasmodium
falciparum isolates in Hadhramout governorate, Yemen
183
Appendix L: List of Publications and Presenting Papers 184
Univers
ity of
Mala
ya

1
CHAPTER 1: INTRODUCTION
1.1 Background of study
Malaria, especially Plasmodium falciparum malaria is one of the main causes of
mortality and morbidity worldwide where 3.3 billion people are at risk of malaria
transmission and 1.2 billion individuals are at high risk of being infected with malaria
(Dyer et al., 2007; Joubert et al., 2009; WHO, 2011). Globally, the transmission of
malaria mostly occurs in tropical and subtropical countries, particularly, in sub-Saharan
Africa and South Asia, affecting 124 to 283 million people and resulting in an estimated
584,000 deaths due to complications, mostly among children less than five years of age
in the African region (Waitumbi et al., 2000; WHO, 2014b). Almost all deaths were
caused by Plasmodium falciparum (Färnert et al., 2001; Snow et al., 2013). In endemic
countries, the people at higher risk of infection with malaria and those severely affected
reside in the poorest communities, with limited or without proper access to effective
prevention, diagnosis and treatment. Thus, combating and elimination of malaria are
related to strengthening of health system, development of infrastructure and reduction of
poverty (WHO, 2014b).
In the WHO Eastern Mediterranean region, which consists of 12 countries,
approximately 280 million people in eight countries including Yemen are at risk of
malaria transmission. Of these, 104 million people are at high risk of malaria
transmission. Six countries (i.e., Sudan, Pakistan, Yemen, Afghanistan, Somalia and
Djibouti) have areas with high incidence of malaria with an estimated 1,027 deaths,
occurred mostly in Sudan and Pakistan (WHO, 2011, 2014b).
Univers
ity of
Mala
ya

2
Yemen is a Mediterranean country where 62% of its population (about 24 million) are at
risk of malaria. In 2013, there were more than 25% of the population at high risk of
acquiring the infection, with 149,451 confirmed cases (WHO, 2014b). Most cases of
malaria in Yemen belongs to the afro-tropical type with the predominance of P.
falciparum which is accountable for nearly 99% of malaria cases with only minimal
cases caused by Plasmodium vivax and with Anopheles arabiensis as the predominant
vector (WHO, 2014b). However, the malaria parasite vector in Socotra Island and the
eastern governorate of Al-Maharah belongs to the oriental type with Anopheles
culicifacies as the predominant vector (NMCP, 2011). The National Malaria Control
Program (NMCP) in Yemen is proactive in controlling malaria through prompt
diagnosis and proper treatment, distribution of insecticide-treated mosquito nets (ITN),
indoor residual spraying (IRS), and active case surveillance (WHO, 2012). However,
Yemen is placed in the control phase and was not on track to achieve the Global Malaria
Action Plan (GMAP)’s objective which was to reduce malaria cases by 75% by the end
of 2015 (WHO, 2012, 2014b). By contrast, Saudi Arabia, the northern neighbouring
country of Yemen, showed more than 75% reduction in malaria case incidence placing
it in the elimination phase, and Oman, the eastern neighbouring country of Yemen is
now in the prevention of re-introduction phase (WHO, 2012).
Socio-economic, environmental and human behavioral factors might contribute to
the slow progress of malaria control in Yemen. Previous malaria indicator survey,
conducted in Yemen between 2008 and 2009, reported that only 4.2% of people and 7%
of children under 5 years slept under long lasting insecticide-treated net (LLINs) (Noor,
2009). Several factors have been identified to increase the malaria transmission in
different countries including house condition, education level, occupation, usage of bed
net, spraying of insecticide inside the house, agriculture, knowledge, beliefs and
Univers
ity of
Mala
ya

3
practices toward malaria disease (Worrall et al., 2003; Yé et al., 2006; Tipmontree et
al., 2009; Ayele et al., 2012; Liu et al., 2014).
Following the emergence of chloroquine (CQ) resistance, the antimalarial
treatment policy in Yemen has shifted to artemisinin-based combination therapy (ACT)
in 2005, where artesunate plus sulphadoxine-pyrimethamine (SP) has been used as the
first line, and artemether lumefantrine (AL) as the second line therapy for
uncomplicated malaria (NMCP, 2006). However, CQ and SP are still being prescribed
as monotherapy by clinicians in both public and private health facilities because they
have limited knowledge of the newer treatment policy (Bashrahil et al., 2010; Bin
Ghouth, 2013). Despite a highly efficacious current antimalarial dug policy in Yemen
(Adeel et al., 2015), several related dihydrofolate reductase (dhfr) gene mutations which
is a molecular marker for SP failure, the partner drug of AS have been reported recently
among Plasmodium falciparum isolates from different governorates in Yemen,
suggesting that the emergence and spread of SP resistance will expose the parasite to
AS monotherapy, which has the potential to contribute to the emergence of ACT
resistance in Yemen. These mutations include double mutant genotype of Pfdhfr
(I51/N108) in Taiz, Dhamar, and Hodeidah governorates in western Yemen (Al-
Hamidhi et al., 2013) and single mutant genotype of Pfdhfr (N108) in Hodeidah
governorate (Abdul-Ghani et al., 2014).
1.2 PROBLEM STATEMENTS
Based on WHO reports, malaria is still a public health threat in Yemen and this
infection contributes a high proportion to the total cases of malaria reported from WHO
Mediterranean region. Besides, it is a challenge to achieve a reduction of malaria cases
Univers
ity of
Mala
ya

4
by 75% by the end of 2015 as stated in the Global Malaria Action Plan (GMAP) (WHO,
2014b). Barriers and factors that are challenging the success of malaria control in
Yemen should be identified in order to develop an effective control strategy. Firstly,
there is a scarcity of data on malaria predictors in this country. Thus, the present study
aimed to determine the prevalence and risk factors of malaria in the southeastern part of
Yemen, and to explore the residents’ knowledge, attitude and practices (KAP) toward
malaria.
In Yemen, malaria treatment is also another challenge. The national antimalarial
drug policy in Yemen was formulated in 1999, included CQ as first-line and SP as a
second line monotherapies for treating uncomplicated falciparum malaria (NMCP,
2006). As mentioned above, CQ is still being prescribed as monotherapy by clinicians
in both public and private health facilities because they have limited knowledge of the
newer treatment policy (Bashrahil et al., 2010; Bin Ghouth, 2013). Continued use of CQ
sustains the selection of CQ resistant mutations leading to persistence of mutant
parasite. The complete withdrawal of CQ use may enhance the emergence of CQ
sensitive parasite over time and make CQ possible to be re-introduced for malaria
treatment (Kublin et al., 2003; Laufer et al., 2006). However, the persistence of CQ
resistance will be prolonged if the shift to ACT and the simultaneous withdrawal of CQ
are not rigorously implemented. The aim of the current survey is to also determine the
prevalence of CQ resistant mutations, since they will be important for future monitoring
and assessment of antimalarial drug policy in Yemen.
In 2005, due to the increased CQ resistance, antimalarial drug policy shifted to a
combination of AS and SP as the first-line therapy and AL as a second-line treatment
for uncomplicated falciparum malaria (WHO, 2011). Various factors such as continued
Univers
ity of
Mala
ya

5
use of SP in the new policy, availability of this drug in the private sector, and poor
knowledge of the national policy among physicians (Bin Ghouth, 2013) may have
increased the usage of monotherapy of SP against P. falciparum, which is likely to
compromise drug efficacy. However, the efficacy of AS + SP as first-line treatment for
uncomplicated falciparum malaria was rated at 97% ACPR in a recent clinical drug
efficacy trail carried out in 2013 (Adeel et al., 2015). It is noteworthy that currently
used routine clinical efficacy trail is the gold standard for the assessment of the
efficiency of the combined antimalarial drugs, although it does not differentiate between
the efficacy of AS and its partner drug. The evolution of SP resistant parasite will
expose the malaria to AS monotherapy and speed the emergence of resistance to
artemisinins. Molecular markers are practical for tracking the resistance toward
antimalarial drugs. Quintuple mutant of combined dhfr and dhps genes (Pfdhfr I51, R59,
T108 plus Pfdhps G437, E540) is significantly associated with in vivo resistance to SP, as
reported in a systematic review in 2009 (Picot et al., 2009). Therefore, the current study
will investigate mutations in dhfr and dhps genes associated with SP resistance and
findings from this study will be used to predict and monitoring the development of SP
resistance in Hadhramout governorate, Yemen.
1.3 RESEARCH HYPOTHESES
1- There are foci with high prevalence of malaria in the Hadhramout governorate,
Yemen.
2- There is a significant association between socio-economic, human behavioural and
environmental factors and malaria in the Hadhramout governorate, Yemen.
3- There is a high prevalence of Pfcrt 76 mutant allele in the Hadhramout governorate,
Yemen.
Univers
ity of
Mala
ya

6
4- There is an existence of Pfmdr1 mutant allele at different loci (86 and 1246) in the
Hadhramout governorate, Yemen.
5- There is an existence of mutants in Pfdhfr and Pfdhps genes in the Hadhramout
governorate, Yemen.
1.4 OBJECTIVES
1.4.1 General objective
The general objective of this study is to determine the epidemiology of malaria and to
detect the frequency of alleles and genotypes of genes associated with antimalarial drug
resistance (Pfcrt, Pfmdr1, Pfhdfr, and Pfdhps) in the Hadhramout governorate, Yemen.
1.4.2 Specific objectives
1. To determine the prevalence of malaria in the Hadhramout governorate, Yemen.
2. To identify the risk factors associated with malaria in the Hadhramout
governorate, Yemen.
3. To assess the knowledge, attitude and practices towards malaria in the
Hadhramout governorate, Yemen.
4. To detect the point mutations of Pfcrt gene at 76, 271, 326, 356 and 371 positions
as molecular markers of CQ resistance of P. falciparum in the Hadhramout
governorate, Yemen
5. To detect the point mutations of Pfmdr1 gene at 86 and 1246 positions as
molecular markers of antimalarial drug resistance of P. falciparum in Hadhramout
governorate, Yemen.
Univers
ity of
Mala
ya

7
6. To ascertain the point mutations of Pfdhfr and Pfdhps genes at different positions
as molecular markers of SP resistance of P. falciparum in Hadhramout
governorate, Yemen.
1.5 SIGNIFICANCE OF STUDY
The present study is the first in Hadhramout governorate, Yemen which will apply new
molecular technologies based on DNA to determine the frequency of mutant alleles and
genotypes associated with antimalarial drug resistance in P. falciparum. The study also
determine the prevalence of malaria and identify environmental, socioeconomic and
behavioural factors associated with the high prevalence of malaria in Hadhramout
governorate, Yemen. Information from this study would help public health local
authorities to develop an effective malaria control strategy based on better
understanding of malaria epidemiology. The study highlight the importance of the
continuous surveying of P. falciparum population for molecular markers as an early
alarming tool for the emergence of antimalarial drug resistance.
Univers
ity of
Mala
ya

8
CHAPTER 2: LITERATURE REVIEW
2.1 Malaria
2.1.1 History
Malaria is an infectious disease and is caused by protozoan parasites of the genus
Plasmodium (Barillas‐Mury & Kumar, 2005). The name malaria is derived from the
Italian ‘mal’ aria,’ which means bad air, derived from the belief that the illness occurs
due to inhalation of bad air around marshy area. Plasmodium was discovered by Charles
Louis Alphonse Laveran at the end of the 19th century. He noticed the parasites in the
blood film of a patient suffering from malaria and for this discovery, he was awarded
the Nobel Prize in 1907 (Launiala & Kulmala, 2006). Later, Dr. Ronald Ross, a British
medical officer in the Indian Medical Service, was the first to discover that mosquitoes
transmit malarial parasites to human and then an Italian professor Giovanni Battista
Grassi identified that only female Anopheline mosquitoes are able to transmit malarial
parasites (Launiala & Kulmala, 2006).
2.1.2 Biology, etiology and life cycle
Out of two hundred species of Plasmodium, five species can cause human malaria and
these include Plasmodium falciparum, Plasmodium vivax, Plasmodium malariae,
Plasmodium ovale. and Plasmodium knowlesi (Singh et al., 1999; Singh et al., 2004a;
Chavatte et al., 2007; Cox-Singh et al., 2008; White, 2008a; Piekarski, 2012).
Although malaria transmission is anthroponotic from human to human via the bites of
infected female Anopheles mosquito, monkeys have been implicated as a source of P.
knowlesi which had been considered as a monkey malaria (Singh et al., 2004a; Cox-
Univers
ity of
Mala
ya

9
Singh et al., 2008; Lee et al., 2011). About 465 Anopheles mosquitoes have been
identified (Harbach, 2011; Raghavendra et al., 2011) Of these, 70 species are able to
transmit malaria parasite to human (Warrell & Gilles, 2002) with 41 species being
reported as dominant natural malaria vectors (Joy et al., 2008; Coutinho-Abreu et al.,
2010; Hay et al., 2010; Dhandapani et al., 2011; Cholewiński et al., 2015).
Transmission of malaria is affected by the mosquito behaviour such as night or day
feeding behaviour, indoor or outdoor, and human or animal preference. The presence of
these species depends on the geographical region and the environmental conditions
(Subbarao & Sharma, 1997).
Plasmodium of mammalian hosts has a complex life cycle, sexual (in Anopheles
as definitive host) and asexual (in human as the intermediate host) life cycles. The
typical life cycle of Plasmodium malaria is demonstrated in Figure 2.1. The asexual life
cycle in human starts with a single bite of infected female Anopheles mosquito
containing 20-30 sporozoites which are able to initiate the malaria disease (Satoskar et
al., 2009). The sporozoites travel though the blood stream to liver cells in 30 minutes
where asexual multiplications occur inside the hepatocytes to form mature schizont
which contains 2000 - 40,000 merozoites. This phase is called exo-erythrocytic
schizogony which usually takes 7-16 days based on the Plasmodium species (Satoskar
et al., 2009). The mature schizonts rupture releasing numerous merozoites that enter
blood stream and intiate erythrocytic schizogony phase. Some sporozoites of P. vivax
and P. ovale may remain inside liver as dormant form called hypnozoite for months or
years prior to the development into mature schizonts that then cause malaria relapse. In
the erythrocytic phase, merozoites invade red blood cells (erythrocytes) to form early
trophozoites (ring form) which have cytoplasm, nucleus and vacuole. The mature
trophozoites develop into erythrocytic schizonts which rupture releasing merozoites.
Univers
ity of
Mala
ya

10
The erythrocytic cycle takes 24 -72 hours based on the species of Plasmodium; one day
for P. knowlesi (Bronner et al., 2009), three days for P. malariae, and two days for P.
vivax, P. ovale and P. falciparum (Satoskar et al., 2009). The released merozoites
invade new erythrocytes and start a new erythrocytic cycle. The gametogenesis starts
after several asexual erythrocytic cycles which produces male (microgametocytes) and
female gametocytes (macroga metocytes) that are infective for female anopheline
mosquito (Lacroix et al., 2005). The circulating gametocytes can be observed for
months or years in the absence of treatment (Bousema et al., 2004). The sexual life
cycle in mosquito starts when the mosquito picks up blood meal with infective
transmission stages (gametocytes) of malaria. The male microgamete and female
macrogamete fuse in the mosquito midgut to form a zygote which undergoes maturation
forming a motile stage called ookinete which will be able to penetrate the gut wall and
further develop into oocysts that contain numerous spindle shapes sporozoites. The
oocysts undergo maturation and asexual multiplications of sporozoites leading to
oocysts rupture and the release of large numbers of sporozoites which travel to the
salivary glands of the mosquito. This process of parasite development is called
sporogony which takes 10-18 days depending on the species of Plasmodium and the
infected Anopheles mosquito which may remain infectious for 1-2 months (Day et al.,
1998; Barry, 2005; Liljander, 2010).
Univers
ity of
Mala
ya

11
Figure 2.1: Life cycle of Plasmodium spp.
Source: Centers of Disease Control and Prevention (CDC, 2014)
www.cdc.gov/malaria/about/biology/index.html
Sporogonic cycle
Exo-erythrocytic cycle
Erythrocytic cycle
Univers
ity of
Mala
ya

12
2.1.3 Clinical manifestations
The first appearance of the symptoms which varies depending on species of
Plasmodium; 9-14 days for P. falciparum, 12-16 days for P. vivax, 16-18 for P. ovale,
18-40 or more for P. malariae and 10-12 days for P. knowlesi (Warrell & Gilles, 2002).
The longest incubation periods reported previously are 4 years for P. falciparum and 30
years for P. vivax (Moody & Chiodini, 2000; Moody, 2002; Trampuz et al., 2003;
Tangpukdee et al., 2009). The long incubation period could be due to the level of
immunity acquired via previous infection, chemoprophylaxis of malaria and prior
partial treatment (Ohrt et al., 2008). Paroxysms of malaria fever occurs as a result of
schizonts rupture within two or three days according to the type of Plasmodium species
(tertian or quartan fever). The ruptured erythrocytic schizont releases into blood stream
Plasmodium pigments, toxins, antigens, and a series of pathological factors such as
cytokines, interleukin 1 (IL-1), interleukin 6 (IL-6), and tumor necrosis factor-alpha
(TNF-α), which lead to the common symptoms of malaria; high fever, profuse sweating,
chills, headache, fatigue, vomiting, nausea, diarrhea and anaemia (Miller et al., 1994;
Miller et al., 2002). Severe malaria may cause serious complications including cerebral
malaria, severe anaemia, hepatosplenomegaly, pulmonary oedema, jaundice,
haemoglobinuria, acute kidney injury, acute respiratory distress syndrome,
hypoglycemia, acidosis, hypotension and brain inflammation that may lead to coma
(Mackintosh et al., 2004; Fritsche & Selvarangan, 2011). The high risk groups for
severe malaria are young children, pregnant women and travellers to malaria endemic
areas (Bejon et al., 2009; Phillips et al., 2009; Mali et al., 2010). Malaria is classified as
severe based on the following criteria; hyperparasitemia (>100,000 parasites/µl in
hypoendemic areas and >200,000 parasites/µl in hyperendemic areas), impaired
consciousness, respiratory distress, or severe anaemia (White, 1996; WHO, 2003;
Univers
ity of
Mala
ya

13
Mackintosh et al., 2004; Crawley et al., 2010; White et al., 2014; WHO, 2015a).
Partially protective immunity is developed in individuals living in a malaria endemic
area after frequent exposure to malaria infections. The individuals with the partial
immunity may be asymptomatic carriers or develop mild signs and symptoms (Bousema
et al., 2014; WHO, 2014b).
2.1.4 Epidemiology
2.1.4.1 Malaria burden and geographical distribution of Plasmodium species
Malaria is a major health problem worldwide with 3.2 billion individuals (representing
about half of the world’s population) at risk of being infected with malaria, and 1.2
billion people are at high risk (WHO, 2014b). In 2013, 198 million cases of malaria
were reported globally with an estimated 584 000 deaths, of them 90% occurred among
children less than five years old in the African region (Dyer et al., 2007; Joubert et al.,
2009; WHO, 2011, 2014b). Approximately 90% of total malaria deaths are due to
falciparum malaria (WHO, 2014b). The countries that have contributed to malaria-
related death are illustrated in Figure 2.2. As a result of the scale up of malaria control
in the period from 2000 to 2012, malaria incidence rate dropped by 31% and mortality
rate by 49% in the WHO African Region (WHO, 2013). Several factors have challenged
the achievement of the goals designed by the Roll Back Malaria (RBM) partnership and
the World Health Organization to decrease the cases of malaria and death recorded in
2000 by 50% and 70% by the end of 2010 and 2015, respectively, in the poor countries.
These factors included climate changes, emergence and spread of antimalarial drugs and
insecticide resistance, lack of infrastructure, international travels to endemic areas,
political instability, civil war, poverty, low-income, weak or unavailability of public
Univers
ity of
Mala
ya

14
health services, and outdoor and indoor biting habits of mosquitoes (Tatem et al., 2010;
WHO, 2012; Cotter et al., 2013).
WHO has classified the malaria endemic areas into six WHO regions including 97
countries; three countries in the Europe region, 10 in the Southeast Asian region, 10 in
the Western pacific region, 8 in the Eastern Mediterranean region, 21 in the America
region and 45 countries in the African region. Majority of malaria cases (82%)
occurred in the WHO African region, followed by the WHO Southeast Asian region
(12%) and the WHO Eastern Mediterranean region (5%) (WHO, 2014b). The
transmission of malaria can be either stable with continuous seasonal or non-seasonal
transmission for many years or unstable malaria transmission with fluctuation variations
(Kiszewski & Teklehaimanot, 2004). In high transmission areas, infants and young
children are the most infected groups, while in low transmission areas, most malaria
cases occur in older children and adults (Carneiro et al., 2010). The intensity of malaria
transmission is indicated in Figure 2.3.
The geographical distribution of malaria species in the world is distinct with most
overlapping in certain geographical areas; P. vivax occurs in many parts of the world
and is predominant in the Asia region, P. falciparum is widespread in tropical and
subtropical regions and highly prevalent in Africa, P. ovale is found in limited parts of
Africa particularly in the western coast, P. malariae is much less frequent in South
America, Asia, and Africa, and P. knowlesi is most commonly reported from Southeast
Asia (Baird, 2007; Mueller et al., 2007; Gupta et al., 2009; Snow et al., 2013).
Univers
ity of
Mala
ya

15
Figure 2.2: The malaria control stage and the countries contributing to the
global death (http://www.rbm.who.int/) (Alonso & Tanner, 2013)
Univers
ity of
Mala
ya

16
(http://www.rbm.who.int/) (WHO, 2014b)
Figure 2.3: The intensity of malaria transmission worldwide
Univers
ity of
Mala
ya

17
2.1.4.2 Factors associated with malaria
The identification of predictors of malaria in an endemic area is a key factor for an
effective control strategy in the area. Multiple risk factors of malaria have been
identified including socioeconomic, environmental and behavioral factors (Table 2.1).
Climate change has a great effect on malaria. Rainfall, temperature and humidity
have influence on vector multiplication or differentiation rate (Longstreth & Wiseman,
1989; Caminade et al., 2014), therefore, high density of malaria vectors increases the
vectors biting rate and consequently increases the prevalence of malaria. The increase in
the environmental temperature may shorten the duration of the sporogonic cycle in the
malaria vector and affect the dynamic of human-vector contact and vector longevity
(Craig et al., 1999; Teklehaimanot et al., 2004). Climate changes, in particular rainfall,
temperature and humidity affect malaria transmission and consequently influence vector
multiplication or differentiation rate (Kiszewski & Teklehaimanot, 2004; Paaijmans et
al., 2009; Blanford et al., 2013; Caminade et al., 2014).
Travelling to malaria endemic areas is a significant risk factor of malaria
(Prothero, 1965; Singhanetra-Renard, 1993; Martens & Hall, 2000). In the same vein,
migration of people due to wars or disasters can caused malaria outbreaks. About
30,000 malarial cases were documented in Tajikistan as a result of civil war in southern
Azerbaijan (Sabitinelli, 2002). Furthermore, as a result of the Soviet Union breakup in
central Europe, about 60,000 malaria epidemic cases were reported (Sabitinelli, 2002).
Occupation may represent a significant predictor of malaria by exposing human to
mosquito bites. Higher prevalence of malaria was reported among soldiers and gem-
Univers
ity of
Mala
ya

18
mining workers in the Thai-Cambodian border. Working in the rural areas was also
identified as a significant factor of getting malaria in Colombia (Luxemburger et al.,
1996; Luxemburger et al., 1997; Mendez et al., 2000; Kitvatanachai et al., 2003).
Additional working activities were identified as risk factors of malaria in endemic areas
such as agricultural, logging, ore-digging and harvesting activities in the farm or forest
(Butraporn et al., 1986; Marques, 1987; Martens & Hall, 2000; Singh et al., 2004b).
Poor housing has been associated with the increased occurrence of malaria in
Gambia, Ethiopia, western of Kenya, Tanzania, Malaysia, Eritrea and Thailand
(Pichainarong & Chaveepojnkamjorn, 2004; Atieli et al., 2009; Peterson et al., 2009;
Ahmad et al., 2014) which increases the indoor density of mosquitoes and subsequently
increases the rate of mosquito bites. Incorrect knowledge about transmission of malaria,
etiology and prevention (Fungladda et al., 1987; Arasu, 1991) and not using insecticide-
treated bed nets have been associated with the high prevalence of malaria (Butraporn et
al., 1986; Fungladda & Sornmani, 1986; Singhanetra-Renard, 1986).
In addition, many malaria risk factors have been identified such as sleeping
outdoors, stagnant water nearby the house and wearing insufficient protective clothes
(Arasu, 1991; Ghebreyesus et al., 2000; Sintasath et al., 2005; Graves et al., 2009;
Alemu et al., 2011), age and gender (Mendez et al., 2000; Van Der Hoek et al., 2003;
Incardona et al., 2007; Winskill et al., 2011). Vector ecology is another factor which
may affect the malaria occurrence. Malaria vector in Asia are described as zoophilic.
Thus, the presence of alternative hosts such as cattles, goat and sheep may reduce the
human exposure to mosquito bites and decreases the incidence of malaria (Habtewold et
al., 2004; Do Manh et al., 2010). Moreover, delayed treatment of malaria cases has been
identified as predictor of developing severe malaria which often occur in rural endemic
Univers
ity of
Mala
ya

19
areas, among people staying far away from health centers, or among people who treat
themselves with traditional drugs (Fungladda & Sornmani, 1986; Arasu, 1991;
Oemijati, 1992; Alemu et al., 2011).
Univers
ity of
Mala
ya

20
Table 2.1: Socioeconomic factors, behavior factors and environmental factors of
malaria
Factors Countries References
Socio-economic factors:-
Family size (more than 5)
Low education
Low income
Age of individuals
(20-39 years/rural area)
(below 17 years)
Pregnant women
Occupation
Poorly constructed houses
Mud walls
Material of the roof
Open of house windows
Some bedrooms without ceiling
Household own cattle
Traveling to another endemic area
Thialand
Sudan
French Guiana
Thailand
Burkina Faso
Thailand
Burkina Faso
Colombia
Srilanka
Thialand
China
Sudan
Brazil
Malawi
India
Thialand
Kenya
Sri lanka
Sri Lanka
Gambia
Eritrea
Gambia
Sri Lanka
Ethiopia
Gambia
Ethiopia
Ethiopia
Burkina Faso
(Butraporn et al., 1986)
(El Samani et al., 1987)
(Hustache et al., 2007)
(Butraporn et al., 1986)
(Baragatti et al., 2009)
(Butraporn et al., 1986)
(Baragatti et al., 2009)
(Mendez et al., 2000)
(van der Hoek et al., 1998)
(Nosten et al., 1991)
(Moore et al., 2008)
(Adam et al., 2005)
(Martínez-Espinosa et al., 2004)
(Brabin et al., 1993)
(Sharma et al., 2015a)
(Inchana et al., 2013)
(Mutero et al., 2000)
(Yapabandara et al., 2001)
(Campaign, 1991)
(Koram et al., 1995)
(Sintasath et al., 2005)
(Adiamah et al., 1993)
(Konradsen et al., 2003)
(Ghebreyesus et al., 2000)
(Adiamah et al., 1993)
(Ghebreyesus et al., 2000)
(Ghebreyesus et al., 2000)
(Baragatti et al., 2009)
Univers
ity of
Mala
ya

Continued
21
Behavioral factors:
Sleeping or staying outdoor
Non sleeping under bed nets
Some bedrooms without ceiling
Non residual spraying of the
walls in a house
Non or Partial coverage of the
body with clothing
Malaysia
Indonesia
China
Thialand
Burkina Faso
Sri Lanka
Gambia
Ethiopia
Thialand
Sri Lanka
Gambia
India
(Arasu, 1991)
(Oemijati, 1992)
(Moore et al., 2008)
(Butraporn et al., 1986)
(Baragatti et al., 2009)
(van der Hoek et al., 1998)
(Adiamah et al., 1993)
(Ghebreyesus et al., 2000)
(Honrado & Fungladda, 1994)
(van der Hoek et al., 1998)
(Koram et al., 1995)
(Lwin et al., 2014)
Environmental factors:
Distance from mosquitoes
breeding sites
Water collections nearby
Presence of stream
Swamp existence
Man-made water tank
Rainy season
Agricultural and irrigation area
Workers outdoor or forest
Colombia
Thailand
Gambia
Kenya
Uganda
Sri Lanka
Sudan
Pakistan
Sri lanka
Thailand
Uganda
Guiana
Ethiopia
Sudan
Eritrea
Burkina Faso
Ghana
Burkina Faso
Ethiopia
Vietnam
Bangladesh
(Mendez et al., 2000)
(Butraporn et al., 1986)
(Clarke et al., 2002)
(Minakawa et al., 2002)
(Staedke et al., 2003)
(Konradsen et al., 2003)
(El Samani et al., 1987)
(Klinkenberg et al., 2004)
(Van Der Hoek et al., 2003)
(Butraporn et al., 1986)
(Staedke et al., 2003)
(Hustache et al., 2007)
(Yewhalaw et al., 2009)
(Ranson & Lissenden, 2016;
Rayah et al., 2016)
(Sintasath et al., 2005)
(Baragatti et al., 2009)
(Klinkenberg et al., 2008)
(Baragatti et al., 2009)
(Ghebreyesus et al., 2000)
(Erhart et al., 2005)
(Haque et al., 2011)
Univers
ity of
Mala
ya

22
2.1.5 Diagnosis of malaria
Accurate and rapid diagnosis of malaria is a key control measure in the strategy of
malaria control which depends on detecting malaria parasite or its antigens or DNA in
the blood of the patient (Fritsche & Selvarangan, 2011). Microscopy is the most
common technique used for detecting and identifying the blood stages of malaria and
has remained the gold standard for malaria diagnosis. Besides, microscopy is the
suitable technique for estimating the parasite density and the assessment of anti-malaria
drug efficacy. However, it is laborious and time consuming, needs expertise and has low
sensitivity in low parasitaemia cases (Kawamoto, 1991; Milne et al., 1994; Coleman et
al., 2006; Ohrt et al., 2008; Hassan et al., 2010). Rapid diagnostic tests (RDTs) were
developed to overcome the drawbacks of microscopy. It is rapid, easy to use, storable at
room temperature and has shown similar or superior sensitivity compared to microscopy
(Azikiwe et al., 2012). Therefore, it has been introduced as alternative tool for malaria
diagnosis in areas where good microscopy cannot be maintained. In contrast, the low
specificity of RDTs has been reported (Bell & Peeling, 2006; Cunningham & Gatton,
2014). Several malaria RDTs of different manufacturers are commercially available
with a variation in their reliability which therefore necessitate selection criteria and in
sometimes field evaluation for decision on procurement and implementation
(Cunningham & Gatton, 2014). Although molecular approaches have shown high
sensitivity and specificity for detecting malaria and identifying Plasmodium species,
they are sophisticated techniques, limited to reference laboratories and therefore not
practical in low income countries (Coleman et al., 2002; Bates et al., 2004; Mitiku et
al., 2004; Azikiwe et al., 2012; Adams et al., 2015)
Univers
ity of
Mala
ya

23
2.1.6 Treatment
2.1.6.1 Anti-malarial drugs
Several antimalarial drugs have been developed and the prescription depends on the
Plasmodium species and the severity of disease. The anti-malarial drug policy is
different from one country to another according to the emergence and spread of anti-
malarial drug resistance. Guidelines for malaria treatment are published and available
from the World Health Organization (WHO, 2015a). The most widely used antimalarial
drugs are classified into classes; 4-aminoquinolines and 8-aminoquinoline (quinolones),
diaminopyrimidine and aminosulfonamide (antifolate drugs), sesquiterpene lactone
endoperoxides (Artemisinin’s drugs) and antibiotics, depending on the chemical
structure and/or mechanism of action (Table 2.2).
Quinine was the first and the oldest antimalarial drug which has been introduced
as pure form for treating malaria since 1820 (Sullivan & Krishna, 2006), but the
resistance of P. falciparum to quinine has been documented in many endemic areas
(Cowman & Foote, 1990; Pukrittayakamee et al., 1994; Jelinek et al., 1995; Segurado et
al., 1997). Chloroquine was the drug of choice for the treatment of uncomplicated
malaria for more than 40 years, and sulfadoxine-pyrimethamine was the second line for
the treatment of uncomplicated malaria in CQ resistance endemic areas. Up to date, P.
falciparum, P. vivax, and P. malariae have shown resistance to antimalarial drugs
(Reyburn, 2010). P. falciparum has evolved resistance to many antimalarial drugs such
as chloroquine (WHO, 2010; Klein, 2013), sulfadoxine-pyrimethamine (Clyde & Shute,
1957; Nair et al., 2003; Roper et al., 2003) and recently artemisinin derivatives in
Southeast-Asia and Western Cambodia (Ashley et al., 2014; WHO, 2014a). The
Univers
ity of
Mala
ya

24
evolution of P. vivax strains resistant to chloroquine and sulfadoxine-pyrimethamine has
been reported in many endemic areas (Reyburn, 2010; WHO, 2015a). As a result, WHO
has recommended the shifting from monotherapy of malaria to combination drugs;
artemisinins combination therapy.
Univers
ity of
Mala
ya

25
Table 2.2: Classification of antimalarial drug and a brief outline of the mechanism of
action
Class/chemical
family
Name of drug Mechanism of action References
4-aminoquinolines
(Quinoline
derivatives)
Chloroquine,
Hydroxychloro
-quine,
Amodiaquine,
Piperaquine
Inhibiting
detoxification of Haem
(Sullivan et al.,
1996; Pagola et al.,
2000; Pandey et al.,
2001; Lvova et al.,
2016)
4-
methanolquinolines
(Quinoline
derivatives)
Mefloquine,
Lumefantrine,
Halofantrine,
Quinine,
Quinidine
Inhibiting
detoxification of Haem
(Kumar &
Bandyopadhyay,
2005; Kumar et al.,
2007; Lvova et al.,
2016)
Diaminopyrimidine
(Antifolates
derivatives)
Pyrimethamine,
Cycloquanil,
Proguanil
(Chloroguanide)
Inhibiting plasmodial
dihydrofolate reductase
(DHFR)
(Zhang & Meshnick,
1991; Cunha‐Rodrigues et al.,
2006)
Aminosulfonamide
(Antifolate
derivatives)
Sulfadoxine,
Dapsone,
Sulfametho-
pyrazine
Inhibiting plasmodial
dihydropteroate
synthase (DHPS)
(Ferone et al., 1969;
Olliaro, 2001)
Sesquiterpine
lactones
(Artemisinin
derivatives)
Artemether,
Arteether,
Artesunate,
Arterolane,
Artemisinin
Free-radical induced
damage or Inhibition of
Sarcoplasmic reticulum
Calcium-dependent
ATPase 6 (SERCA)
(Terkuile et al.,
1993; White, 2008b;
Fidock, 2010)
8-aminoquinoline
(Quinoline
derivatives)
Primaquine,
Tafenoquine,
Bulaquine
Inhibits electron
transport chain in
Plasmodium
(Butterworth et al.,
2013)
Naphthoquinone
(Quinoline
derivatives)
Atovaquone
Inhibiting
mitochondrial electron
transport chain in
Plasmodium and
mimicking ubiquinone
(Fry & Beesley,
1991; Fry & Pudney,
1992; Hudson, 1993;
Srivastava et al.,
1997)
Antibiotics Tetracycline
Doxycycline
Clindamycin
Azithromycin
Inhibiting protein
synthesis in apicoplast
(Cunha‐Rodrigues et
al., 2006; van Eijk &
Terlouw, 2011)
Univers
ity of
Mala
ya

26
2.1.6.2 Methods of anti-malarial drug resistance surveillance
The emerging resistance of malaria parasites has resulted in reduced efficacy of
antimalarial drugs and drug combinations in some areas. Surveillance of antimalarial
drug efficacy is required for effective malaria management of cases and early detection
of resistance to antimalarial drugs. Three main approaches have been used to evaluate
the effectiveness of several antimalarial drugs: in vivo drug efficacy testing, in vitro
assay susceptibility testing, and molecular methods (WHO, 2003). These methods have
advantages and disadvantages. It is essential to distinguish the resistance of malaria
parasites from treatment failure, which is unable to clear malarial parasitaemia and/or
resolve clinical symptoms of the disease after treatment. The susceptibility of malarial
parasites is just one factor that determines the outcome of the antimalarial drug
treatment. Other factors that contribute to treatment failure include incorrect dose of
treatment, poor treatment persistence and compliance, poor quality of drug, inadequate
drug absorption and/or interaction with other drugs (Laufer, 2009).
A) In vivo methods
This method is based on the WHO standardized observation period of 7, 14, or 28 days
and subsequent follow-up of parasitological outcomes (S/RI/RII/RIII levels of
resistance) (Table 2.3) or parasitological and clinical signs and symptoms (adequate
clinical response, early or late treatment failure) (Table 2.4) and treatment of a
symptomatic patient with a standard dose of an antimalarial drug (WHO, 2003;
Stepniewska et al., 2004). The in vivo therapeutic efficacy method remains the gold
standard for monitoring antimalarial drug efficacy and guiding drug policy. It is a
straightforward method, provides indicator results of the efficacy of an antimalarial drug
Univers
ity of
Mala
ya

27
and requires minimal training (except microscopy), equipment and supplies in contrast
to the in vitro method (WHO, 2003). The in vivo therapeutic efficacy method was
introduced as a result of CQ resistance in 1965 and the protocols were developed,
revised and standardized for the assessment and monitoring antimalarial drug efficacy
in children and infants in high transmission areas of malaria (WHO, 1996). Recently,
the methodology of in vivo test has been developed and has undergone many
modifications to render it suitable to apply in areas with low to moderate transmission
of malaria (WHO, 2002, 2003). However, this method has many disadvantages such as
long periods of monitoring leading to possible high patient loss, does not necessarily
reflect the level of true antimalarial drug resistance due to many factors such as
treatment outcome interference with patient immunity, previous drug intake,
metabolism and drug absorption variations, also misclassification of reinfection as
recrudescence.
Following the WHO recommendation, genotyping to detect populations of
malaria parasite can be a useful tool for examining a number of infecting parasite clones
and diversity of infection due to host immunity and transmission intensity. In
antimalarial drug clinical trials, the PCR based parasite genotyping can be used as a
correction method to differentiate between new infections and recrudescence in
Plasmodium malaria (WHO, 2008b). Furthermore, the characterisation of length
polymorphism of the merozoite surface proteins (MSP-1, MSP-2 ) and the glutamate
rich protein (GLURP) genes in samples collected at day zero and on the day
reappearance of parasitemia was present. Recrudescence infection was defined when at
least one identical length polymorphism for each genotype markers (MspI,Msp2, and
Glurp) was present between samples collected on the same period. Wherase, a new
infection was occured when, the length polymorphisms were different for one or more
Univers
ity of
Mala
ya

28
genotype markers between the sample collected on the same period (Mugittu et
al.,2006; WHO 2008b).
Univers
ity of
Mala
ya

29
Table 2.3: Definitions of parasitological response to drug in in vivo therapeutic efficacy
studies
Parasitological response outcomes (WHO, 1996) a
Sensitive Reduce of asexual parasitemia to 25% within 48 hours after
initiation of treatment and complete clearance of parasitemia on
day 7, without subsequent recrudescence up to day 28.
RI Reduce of asexual parasitemia to < 25% within 48 hours after
initiation of treatment, but reappears between day 7 and day 28.
RII Reduce of asexual parasitemia to > 25% but < 75% within 48
hours after initiation of treatment, without complete clearance on
day 7.
RIII Absence or reduce level of parasitemia to < 25% or an increase
in parasitaemia after 48 hours from initiation of treatment.
a Outcomes for extended test protocol (i.e., 14 Day or 28 Day follow-up); R, resistance.
Univers
ity of
Mala
ya

30
Table 2.4: Definitions of parasitological and clinical response to drug in in vivo
therapeutic efficacy studies (WHO, 2003)a
WHO in vivo test follow up protocol
ETF Signs of complicated severe malaria on day 1, 2, or 3, or level of parasitaemia
on day 2 higher than on day 0, or level of parasitaemia on day 3 ≥ 25% of day
0, or parasitaemia with axillary temperature ≥ 37.5°C on day 3.
LCF Signs of complicated severe malaria after day 3, or parasitaemia with axillary
temperature ≥37.5°C from day 4 to day 28 without meeting any of the criteria
for ETF.
LPF Parasitaemia without axillary temperature ≥37.5°C from day 7 to day 28
without meeting any of the criteria for ETF or LCF.
ACPR Absence of parasitaemia on day 28, without meeting any of the criteria for
ETF, LCF or LPF.
a 14 day follow-up protocol for high transmission areas and 28 day follow-up for low to
moderate transmission areas.
ETF; Early treatment failure, LCF; Late clinical failure, LPF; Late parasitological
failure, ACPR; Adequate clinical and parasitological response.
Univers
ity of
Mala
ya

31
B) In vitro method
In vitro assays measure the intrinsic sensitivity of malaria parasites to a range of
antimalarial drug concentrations. There are several in vitro assay methods, which differ
in respect to the measured effect and period of exposure to the test compound. These
include the WHO mark III test (assessment of schizont maturation or replication by
using microscopic examination of blood films), the radioisotopic test (Incorporation of
radiolabelled nucleotide precursors), the enzyme-linked immunosorbent assay (ELISA)
test together with antibodies against Plasmodium lactate dehydrogenase or histidine-rich
protein 2 (Noedl et al., 2002; Olliaro, 2005) and fluorometric assays with DNA binding
fluorescent dyes (Noedl et al., 2003; Corbett et al., 2004). The in vitro method has many
advantages, such as multiple analyses can be carried out with the same isolated sample
and numerous drug sensitivities can be evaluated simultaneously, including drugs that
are undergoing experiments. It also removes host-confounding factors which influence
in vivo tests as a result of isolated parasites from the host and placed into the controlled
environment. It can accurately detect true drug resistance and provides quantitative
results. The in vitro and molecular markers monitoring could help as complementary
methods for in vivo studies and provide early alarm tools for antimalarial drug
resistance (Shah et al., 2011). However, the disadvantages of in vitro method include
being costly and time-consuming, require expensive equipment and supplies, proper
training and advanced skills for parasite cultures (Basco & Ringwald, 2000).
Furthermore, there is a lack of standardized in vitro protocols, long-term in vitro
adaptation may lose parasites (LeRoux et al., 2009), threshold values for resistance are
not validated, and the correlation with therapeutic response studies is not yet fully
reliable and understood. Therefore, they are not readily amenable to large scale
epidemiological mapping, especially in poor countries, and are recommended to be used
Univers
ity of
Mala
ya

32
with in vivo efficacy studies (Miller et al., 2005). Also, once it has been identified that
the antimalarial drug resistance is associated with genetic changes, the assessment of
drug resistance needs to be confirmed using molecular techniques.
C) Molecular markers
Table 2.5 indicates the commonly used antimalarial drugs and their corresponding
molecular markers that predict the evolution of resistant parasite. Screening the parasite
population for mutations at these genetic loci has been considred essential tool for
antimalarial drug resistance surveillance in malaria endemic areas (Plowe et al., 2007).
This molecular approach has many advantages, such as multiple tests can be performed
on one isolated sample on filter paper and numerous drugs can be evaluated
simultaneously. In addition, numerous samples can be easily collected, transported,
stored, assessed, and examined within a short period. This approach is also not affected
by host-confounding factors that are usually controlled in the routine clinical efficacy
trails (WHO, 2003; Ekland & Fidock, 2008). Furthormore, screening for mutations in
molecular markers corresponding to combined drugs in a combined based antimlarial
drug policy may provide early indications for evloution of a resistance to the partner
drug in the combination therapy, avoiding the expose of the malaria parasite to
monotherapy which can not be achieved using the routine therapeutic efficacy methods
(Plowe et al., 1995; Su et al., 1997; Kublin et al., 2003; Picot et al., 2009). However,
the presence of the molecular marker is not necessary correlated with the failure of the
corresponding treatment (Sidhu et al., 2002). Beside the evolution of mutations
associated with developing antimalarial drug resistance, the possibility of treatment
failure depends on host-confounding factors such as the immune response of the host,
drug dose, drug absorption variation and metabolism (Hastings, 2007).
Univers
ity of
Mala
ya

33
Many methods are available for monitoring and assessment of antimalarial drug
resistance genes which include PCR-RFLP (polymerase chain reaction-restriction
fragment length polymorphism) (Djimdé et al., 2001a; Djimdé et al., 2001b), a nested
mutation-specific PCR (Djimdé et al., 2001a; Lopes et al., 2002; Ranford-Cartwright et
al., 2002; Sangster et al., 2002). These methods have disadvantages, which include
SNPs analysis for a limited number of samples usually limited by high cost. Other
techniques such as pyrosequencing (Nair et al., 2002), real-time PCR (de Monbrison et
al., 2003; Alker et al., 2004), molecular beacons (Durand et al., 2000), or clamped-
probe PCR (Senescau et al., 2005), are also complex and expensive. Most of these
technologies have disadvantages such as a lack of standardised protocols for specimen
collection, processing and DNA extraction. Also, these technologies involve expensive
infrastructure, equipment, and supplies, especially in the poorest countries.
Univers
ity of
Mala
ya

34
Table 2.5: Most commonly used antimalarial drugs along with their molecular
markers to determine their drug susceptibility/resistance
Anti-
malarial
derivatives
Class/
Chemical
family
Name of
drug
Presence
of
resistance
Corres-
ponding
genetic
markers
References
Quinoline
derivatives
4-amino-
quinolines
Chloroquine,
Amodiaquine
Yes Point
mutations
in Pfcrt , Pfmdr1,
Pfmrp1 and
copy
number variation
in Pfmdr1
Djimdé et al., 2001b;
Sidhu et al., 2002; Mu
et al., 2003; Duraisingh, & Cowman 2005;
Humphreys et al., 2007;
Nawaz et al., 2009; Ecker et al., 2012;
Gupta et al., 2014.
4-methanol-
quinolines
(Amino
alcohol)
Mefloquine,
Lumefantrin,
Halofantrine,
Quinine
Yes Point
mutations
in Pfcrt ,
Pfmdr1, Pfmrp,
Pfnhe-1
and copy number
variation in
Pfmdr 1
Duraisingh et al., 2000;
Mu et al., 2003; Ferdig
et al.,2004; Price et al.,
2004; Sidhu et al., 2006; Humphreys et al.,
2007; Preechapornkul
et al., 2009; Koenderink et al., 2010.
8-amino-
quinoline
Primaquine Yes Not known Murphy et al., 1993;
Baird & Hoffman 2004;
Thomas et al., 2016
Naphtho-
quinone
Atovaquone
Yes Point
mutation in
Cyt-b gene
Korsinczky et al., 2000;
Fivelman et al., 2002.
Antifolate
derivatives
Diamino-
pyrimidine
Pyrimethami
n,
Proguanil
Yes Point
mutation in
Pfdhfr
Plowe et al., 1997;
Gregson et al., 2005;
Gama et al., 2009; Alifrangis et al., 2014;
Sharma et al., 2015b.
Amino-
sulfonamide
Sulfadoxine,
sulfene
Yes Point
mutation in Pfdhps
Yuvaniyama et al.,
2003; Pearce et al., 2003; Gama et al.,
2009; Lumb et al., 2011;
Alifrangis et al., 2014; Sharma et al., 2015b
Artemisinin
derivatives
Sesquiterpine
lactones
Artemether,
Artesunate,
Artemisinin
Yes Polymorphi
sm in
Kelch 13 protein
Noedl et al., 2008;
Dondorp et al., 2009;
Ariey et al., 2014; Ashley et al.,2014;
Takala-Harrison et al.,
2015.
Univers
ity of
Mala
ya

35
2.2 Malaria in Eastern Mediterranean region
2.2.1 Current status
In 2013, approximately 280 million people were at risk of malaria in eight countries in
the Eastern Mediterranean. P. falciparum is responsible for the majority of malaria
cases in all countries except in Iran, Afghanistan, and Pakistan, where P. vivax is the
most common species responsible for malaria cases. The majority of malaria cases were
reported from Sudan, Pakistan, Afghanistan, Yemen, Somalia and Djibouti with more
than 55% of the cases occurring in Sudan. Most malaria attributed death cases occurred
in Sudan (67%) and Pakistan (24%) (Table 2.6) (WHO, 2014).
All endemic countries in the Eastern Mediterranean have reported a decline in the
number of confirmed malaria cases and deaths due to the strengthening of malaria
control programmes which include the use of effective artemisinin-based combination
therapies (WHO, 2014b), and vector control such as long-lasting insecticidal nets and
indoor residual spraying. Improved health services especially in urban areas have also
contributed to progress and improved outcomes. The number of malaria cases dropped
from two million to one million during the period from 2000 to 2013. The incidence of
malaria in six countries (Saudi Arabia, Oman, Syria, Afghanistan, and Iran) decreased
more than 75% during the same period. Only 519 and 34 indigenous cases were
reported from Iran and Saudi Arabia in 2013, respectively, and zero indigenous cases
were reported from Iraq since 2009 (WHO, 2014b).
Seven countries are still in control phase including Yemen, Sudan, South Sudan,
Pakistan, Somalia, Afghanistan and Djibouti, while two countries have limited
transmission of malaria which are the Kingdom of Saudi Arabia and Iran and have been
Univers
ity of
Mala
ya

36
classified in the elimination phase. Four countries are in the prevention of reintroduction
phase. They are Egypt, Oman, the Syrian Arab Republic, Iraq, since 1998, 2004; 2005,
and 2009, respectively, however, few local malaria cases may be recorded due to
imported cases, such as in Aswan governorate, Egypt has reported 22 local malaria
cases in 2014 (WHO, 2014b). in addition, the countries that have been certified free of
malaria including Morocco (2010) and the United Arab Emirates (2007) and Tunisia
(2012) (WHO, 2012, 2013). The remaining countries belong to Eastern Mediterranean
region that have eliminated local transmission of malaria long time ago or have a few
imported cases include Qatar (1970), Bahrain (1979), Kuwait (1979), Libya (1973),
Lebanon (1963), Jordan (1970), Palestine (1965) and Cyprus (1953), but they have not
request certification from WHO (WHO, 2001; Malaria, 2002; Atta & Zamani, 2008).
Univers
ity of
Mala
ya

37
Table 2.6: Malaria cases in countries with high transmission areas at the Eastern
Mediterranean region in 2013
Countries Confirmed
Cases
Deaths cases Total estimated
cases
Pakistan 3,472,727 244 7,752,797
Sudan 989,946 685 2,197,653
Afghanistan 319,742 24 787,624
Yemen 149,451 55 924,821
Somalia 60,199 - 119,752
Djibouti 1,684 17 7,934
Univers
ity of
Mala
ya

38
2.2.2 Anti-malarial drug resistance
There have been various drug-resistant molecular markers identified for P. falciparum
in Mediterranean countries and these are summarised in Table 2.7. All studies have
shown a predominance of Pfcrt 76T allele suggesting the spread of CQ resistance (Al-
Mekhlafi et al., 2011b; Afsharpad et al., 2012; Al-Farsi et al., 2012; Dajem et al., 2012;
Khatoon et al., 2013). For mutations in Pfdhfr and Pfdhps genes, the single mutant
allele R59 has been reported from Saudi Arabia (Al Harthi, 2007; Dajem & Al-Qahtani,
2010; Zakai et al., 2013), Iran (Zakeri et al., 2003; Heidari et al., 2007; Zakeri et al.,
2007; Afsharpad et al., 2012; Rouhani et al., 2015) and Yemen (Mubjer et al., 2011; Al-
Hamidhi et al., 2013). The double mutant 51I/108N was reported from Saudi Arabia
(Al-Farsi et al., 2012; Dajem et al., 2012), Pakistan (Ghanchi et al., 2011), Iran (Zakeri
et al., 2003; Zakeri et al., 2007; Afsharpad et al., 2012) and Yemen (Al-Hamidhi et al.,
2013). The triple mutant 51I/59R/108N was not seen in Saudi Arabia (Al-Farsi et al.,
2012) but was reported in Sudan (A-Elbasit et al., 2008). The single mutant allele
Pfdhps 437G was reported from Saudi Arabia and Iran (Afsharpad et al., 2012; Al-Farsi
et al., 2012; Dajem et al., 2012; Rouhani et al., 2015). In addition, 20% isolates
harboring a single mutant 540E of the Pfdhps gene were recorded in 1999 in Iran
(Eskandarian et al., 2002). It is noteworthy that the absence of the quintuple mutant
genotype (Pfdhfr I51R59N108 and Pfdhps G437 E540) which has a significant association
with SP failure (Picot et al., 2009) shows that SP is still justified as AS-partner drug in
the ACTs drug policy in the Mediterranean region. However, the emergence of the
triple mutant 51I/59R/108N genotype in the region necessitate the continued monitoring
of the efficacy of ACTs and the AS partner drugs.
Univers
ity of
Mala
ya

39
Table 2.7: Summary of some previous studies on mutations of Pfcrt, Pfmdr1, Pfdhfr and Pfdhps genes in Plasmodium isolates and their role in
antimalarial drugs resistance in Mediterranean countries.
Country Mutations in gene markers Notes Reference
Pfcrt Pfmdr1 Pfdhfr Pfdhps
Saudi
Arabia
76T
76T
76T
76T
76T
86Y,184F
86Y
86Y
86Y,184F
59R
51I,108N
59R
59R
51I,108N
437G
437G
-Low level of mutations in Pfdhfr against SP drug
indicates that SP is still an effective treatment.
-No significant difference in distribution of drug
resistance genes was observed between Saudis and
expatriates.
-Showed high prevalence of K76T and N86Y
mutations in the Pfcrt and Pfmdr1 genes by
pyrosequencing.
-Association of CQR with Pfcrt 76T and Pfmdr1 86Y
mutations.
-Showed initial phase of SP resistance in Jazan district.
-Showed prevalence of mutations was associated with
CQR and evolution and spread of SP resistance.
(Zakai et al., 2013)
(Dajem et al., 2012)
(Dajem et al., 2011)
(Dajem & Al-
Qahtani, 2010)
(Al Harthi, 2007)
(Al-Farsi et al.,
2012)
Iran
76T
76T
86Y
86Y
59R, 108N
15I, 59R, 108N
15I, 59R, 108N
59R, 108N,164L
437G
437G
437G
436A,437G
- No association between the clinical outcome of SP
treatment and presence of single or double mutations.
- No correlation between in vivo resistance and the
presence of mutations.
-Association between in vivo SP resistance and
mutations in Pfdhfr and Pfdhps genes before adoption
of SP + AS as the first-line, fixed level of Pfcrt 76T
was observed.
-No correlation between in vivo resistance and the
quintuple mutant, but A437G showed correlation.
(Rouhani et al.,
2015)
(Afsharpad et al.,
2012)
(Zakeri et al., 2003;
Zakeri et al., 2007)
(Heidari et al., 2007)
Univers
ity of
Mala
ya
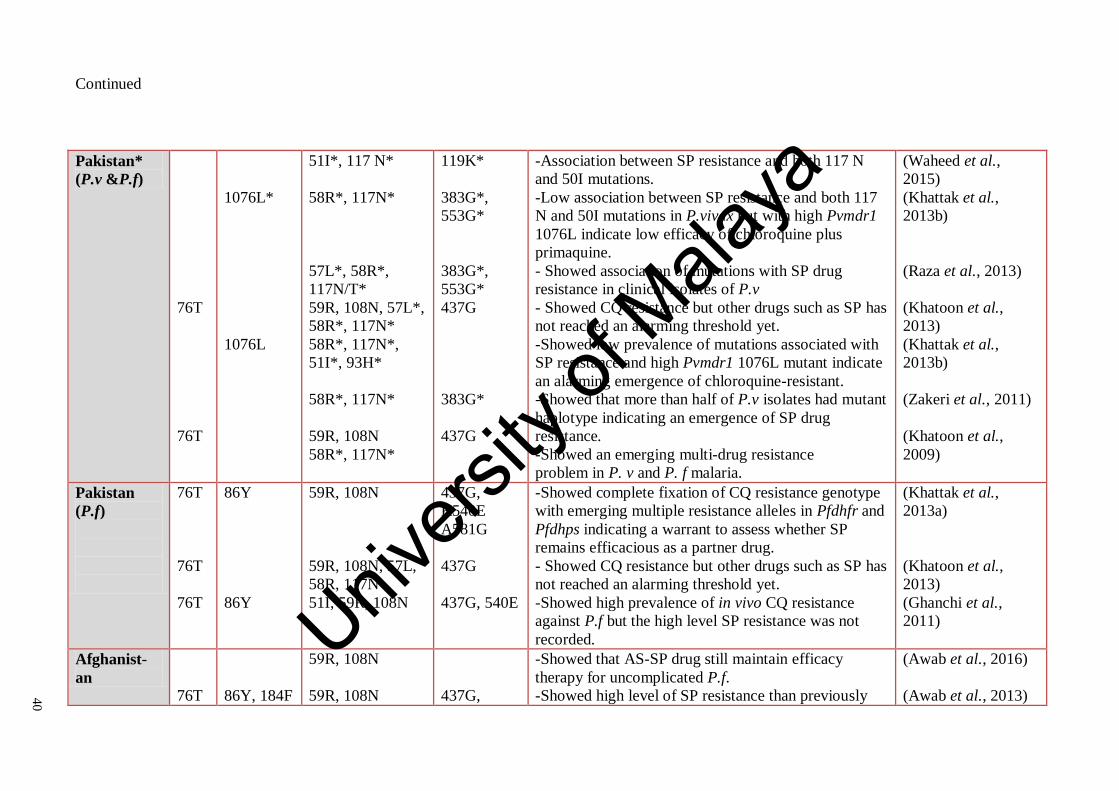
Continued
40
Pakistan*
(P.v &P.f)
76T
76T
1076L*
1076L
51I*, 117 N*
58R*, 117N*
57L*, 58R*,
117N/T*
59R, 108N, 57L*,
58R*, 117N*
58R*, 117N*,
51I*, 93H*
58R*, 117N*
59R, 108N
58R*, 117N*
119K*
383G*,
553G*
383G*,
553G*
437G
383G*
437G
-Association between SP resistance and both 117 N
and 50I mutations.
-Low association between SP resistance and both 117
N and 50I mutations in P.vivax but with high Pvmdr1
1076L indicate low efficacy of chloroquine plus
primaquine.
- Showed association of mutations with SP drug
resistance in clinical isolates of P.v
- Showed CQ resistance but other drugs such as SP has
not reached an alarming threshold yet.
-Showed low prevalence of mutations associated with
SP resistance and high Pvmdr1 1076L mutant indicate
an alarming emergence of chloroquine-resistant.
-Showed that more than half of P.v isolates had mutant
haplotype indicating an emergence of SP drug
resistance.
-Showed an emerging multi-drug resistance
problem in P. v and P. f malaria.
(Waheed et al.,
2015)
(Khattak et al.,
2013b)
(Raza et al., 2013)
(Khatoon et al.,
2013)
(Khattak et al.,
2013b)
(Zakeri et al., 2011)
(Khatoon et al.,
2009)
Pakistan
(P.f)
76T
76T
76T
86Y
86Y
59R, 108N
59R, 108N, 57L,
58R, 117N
51I, 59R, 108N
437G,
K540E
A581G
437G
437G, 540E
-Showed complete fixation of CQ resistance genotype
with emerging multiple resistance alleles in Pfdhfr and
Pfdhps indicating a warrant to assess whether SP
remains efficacious as a partner drug.
- Showed CQ resistance but other drugs such as SP has
not reached an alarming threshold yet.
-Showed high prevalence of in vivo CQ resistance
against P.f but the high level SP resistance was not
recorded.
(Khattak et al.,
2013a)
(Khatoon et al.,
2013)
(Ghanchi et al.,
2011)
Afghanist-
an
76T
86Y, 184F
59R, 108N
59R, 108N
437G,
-Showed that AS-SP drug still maintain efficacy
therapy for uncomplicated P.f.
-Showed high level of SP resistance than previously
(Awab et al., 2016)
(Awab et al., 2013)
Univers
ity of
Mala
ya
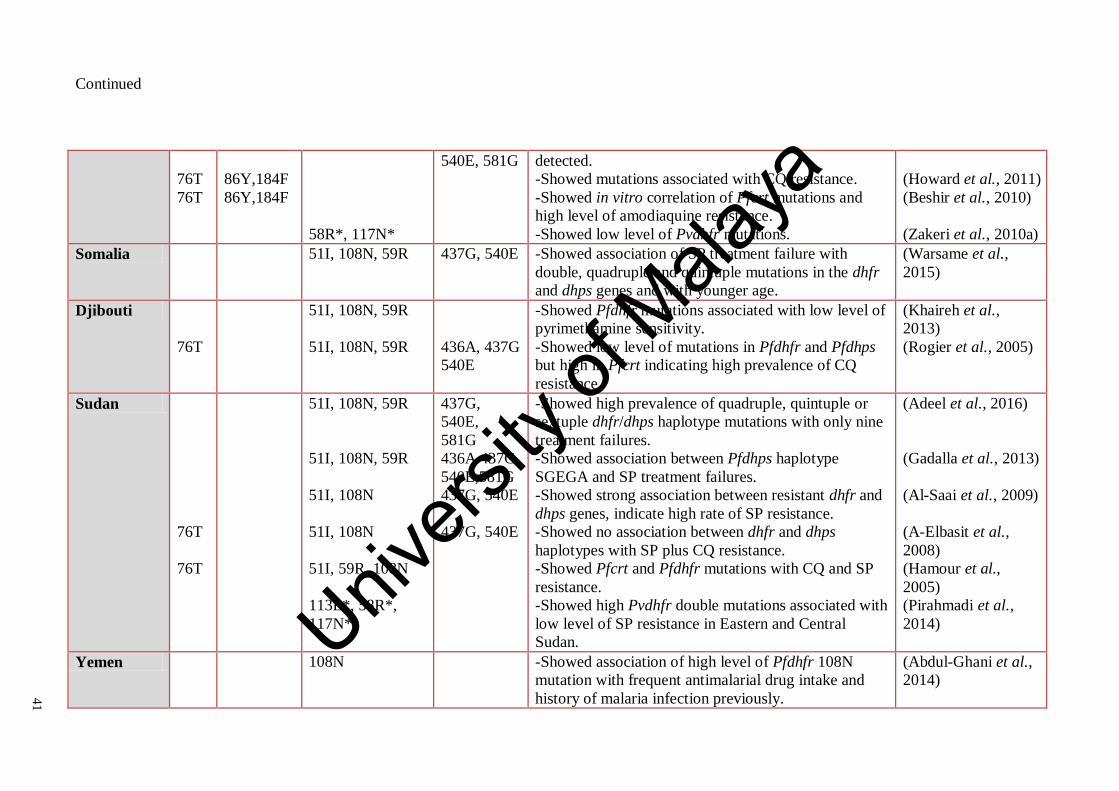
Continued
41
76T
76T
86Y,184F
86Y,184F
58R*, 117N*
540E, 581G
detected.
-Showed mutations associated with CQ resistance.
-Showed in vitro correlation of Pfcrt mutations and
high level of amodiaquine resistance.
-Showed low level of Pvdhfr mutations.
(Howard et al., 2011)
(Beshir et al., 2010)
(Zakeri et al., 2010a)
Somalia 51I, 108N, 59R 437G, 540E -Showed association of SP treatment failure with
double, quadruple and quintuple mutations in the dhfr
and dhps genes and with younger age.
(Warsame et al.,
2015)
Djibouti
76T
51I, 108N, 59R
51I, 108N, 59R
436A, 437G
540E
-Showed Pfdhfr mutations associated with low level of
pyrimethamine sensitivity.
-Showed low level of mutations in Pfdhfr and Pfdhps
but high in Pfcrt indicating high prevalence of CQ
resistance.
(Khaireh et al.,
2013)
(Rogier et al., 2005)
Sudan
76T
76T
51I, 108N, 59R
51I, 108N, 59R
51I, 108N
51I, 108N
51I, 59R, 108N
113L*, 58R*,
117N*
437G,
540E,
581G
436A 437G,
540E,581G
437G, 540E
437G, 540E
-Showed high prevalence of quadruple, quintuple or
sextuple dhfr/dhps haplotype mutations with only nine
treatment failures.
-Showed association between Pfdhps haplotype
SGEGA and SP treatment failures.
-Showed strong association between resistant dhfr and
dhps genes, indicate high rate of SP resistance.
-Showed no association between dhfr and dhps
haplotypes with SP plus CQ resistance.
-Showed Pfcrt and Pfdhfr mutations with CQ and SP
resistance.
-Showed high Pvdhfr double mutations associated with
low level of SP resistance in Eastern and Central
Sudan.
(Adeel et al., 2016)
(Gadalla et al., 2013)
(Al-Saai et al., 2009)
(A-Elbasit et al.,
2008)
(Hamour et al.,
2005)
(Pirahmadi et al.,
2014)
Yemen
108N
-Showed association of high level of Pfdhfr 108N
mutation with frequent antimalarial drug intake and
history of malaria infection previously.
(Abdul-Ghani et al.,
2014)
Univers
ity of
Mala
ya

Continued
42
* point mutations are shown for genes in Plasmodium vivax at different codons.
76T
76T
76T
76T
86Y, 184F
51I, 108N, 59R
59R
-Showed high level of mutations in Pfcrt, Pfmdr1 and
Pfdhfr with high diversity of P. falciparum, indicating
a large parasite reservoir.
-Showed association of Pfcrt 76T mutation with
parasitemia and treatment seeking behaviors.
-Showed association of Pfcrt 76T mutation with
moderate/low parasitaemi, the age group > 10 years
and low household income in Hodeidah and Taiz
governorates, Yemen.
-Showed an emerging resistance to SP and haplotype
associated with CQ resistance P. falciparum parasites
from Lahj governorate, Yemen.
(Al-Hamidhi et al.,
2013)
(Abdul-Ghani et al.,
2013)
(Al-Mekhlafi et al.,
2011b)
(Mubjer et al., 2011)
Univers
ity of
Mala
ya

43
2.3 Malaria in Yemen
2.3.1 Plasmodium species and types of vectors
Malaria is still a major public health problem in Yemen. The epidemiology is classified
as an Afrotropical type with 99% of malaria cases being caused by P. falciparum as the
predominant species and the remainder are P. vivax and P. malariae (NMCP, 2002; Al-
Maktari et al., 2003; Azazy & Raja'a, 2003; Bassiouny & Al-Maktari, 2005; Al-Taiar et
al., 2006; Alkadi et al., 2006; Abdulsalam et al., 2010). The epidemiology of
falciparum malaria in the Arabian Peninsula including Yemen can be divided according
to topographical criteria into three eco-epidemiological zones of malaria; Oriental,
Palaearctic and Afrotropical, leading to a wide variation in vectors and transmissions of
malaria parasites and the subsequent increased risk of malaria (Zahar, 1974;
Kouznetsov, 1976; Kravchenko, 1979; Snow et al., 2013).
In Yemen, many species of anopheline mosquitoes have been reported to be
responsible for malaria transmission (Figure 2.4). The most common vectors include
Anopheles arabiensis which has been reported as the main vector within the country,
Anopheles culicifacies which is an important vector in the coastal areas and it is the
predominant vector in Socotra Island and the eastern governorate of Al Maharah,
Anopheles sergenti has been reported to be a vector in the mountainous hinterland and
highland areas, and recently An. algeriensis (Sinka et al., 2010; Snow et al., 2013).
Understanding the time of biting and indoor or outdoor resting behaviours, distribution
of Yemeni anopheline mosquitoes, the susceptibility of vectors to insecticides and their
role are important in the control and transmission of malaria and can assist in planning
for vector control strategies in Yemen. Increase or decrease of malaria incidence
depends on the densities of mosquitoes. Peak transmission of malaria in endemic areas
Univers
ity of
Mala
ya

44
is mostly related to rainy seasons as there are plentiful of breeding sites available for
mosquitoes. Yemen shares many similarities of the ecology of mosquitoes with Saudi
Arabia, Jazan Provine. In addition, high transmission of malaria in Yemen as a result of
illegal immigration of people from the malaria endemic countries in the horn of Africa
such as Eritrea, Somalia and Ethiopia due to conflict, political instability, civil wars and
poverty (Soucy, 2011).
Univers
ity of
Mala
ya

45
Source: (NMCP, 2006)
Figure 2.4: Modified map of distribution of Anopheles mosquito in governorates,
Yemen
Univers
ity of
Mala
ya

46
2.3.2 Trend of confirmed malaria cases in the last 23 years
The estimate of malaria cases in Yemen fluctuates from year to year, making estimation
of malaria cases incidences difficult (WHO, 2013). For example, there were 2.7 million
cases in 1999 and 3.2 million cases in 2001 after which the malaria cases declined
gradually between 800,000 -900,000 cases in 2006 with 1% estimated related deaths
(Figure 2.5). According to a WHO report, malaria cases in Yemen in 2009 were
265,074 cases with 779 related deaths (WHO, 2008a; NMCP, 2011). The weather and
climate of Yemen varies from one region to another due to diverse topography, changes
in climates especially in rainfall, temperature and humidity leading to variation in the
rates of malaria transmission.
Univers
ity of
Mala
ya

47
Figure 2.5: Malaria trend in Yemen from 1990 till 2014
0
150000
300000
450000
600000
750000
900000
1050000
1200000
1350000
1500000
1650000
1800000
1950000
2100000
2250000
2400000
2550000
2700000
2850000
3000000
1990
1991
1992
1993
1994
1995
1996
1997
1999
2000
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
2012
2013
2014
Mal
aria
ca
ses
Years
Univers
ity of
Mala
ya

48
2.3.3 Risk factors
Yemen is the remaining country in the Arabian Peninsula with high transmission rate of
malaria because of unstable political issues and civil wars for many years. The country
undergoes numerous environmental and social stresses such as food and water
insecurity, and severe depletion in water resources, weak institutions and health system,
rapid population growth and climate change (Husain & Chaudhary, 2008).
The research on risk factors for malaria in Yemen is still very scant as there were
only four studies in particular governorates such as Taiz, Hodiedah, Dhamar and
Raymah that have determined some factors that could be associated with the increased
risk of acquiring malaria (Table 2.8). For example, Al‐Taiar et al. (2008) study found
that socioeconomic factors (distance to nearest health center >2 km, driving time to
reach health center > 10 min and house with earth roof), behavior factors (spray
insecticides at home, delay of treatment > 3 days, burning mosquito coils) and
environmental factors (nearby the water pump to house, nearby the man-made water
collection/tank and more than 2 km of the distance from health center) were risk factors
for malaria. In addition, Al-Taiar et al. (2009) have also demonstrated that
socioeconomic factors (house with earth roof with or without opening in the roof, and
traveling to another endemic area in the last 2 month), behavior factors (burning animal
dung) and environmental factors (water collections nearby, presence of water
stream/spring, presence of water pump, swamp existence/marshy land, presence of
latrine outside the house or non and water storage at home:in jerry cans) could be risk
factors that lead to increase the chance of acquiring malaria in the country.
Univers
ity of
Mala
ya

49
A more recent study in 2011 found a significance risk factors for malaria include
children ≤ 12 year, large family size, gender, living in rural area, low income, not
working, not sleeping under bed nets, burning mosquito coils, spray insecticides at
home, wear short clothes and water collections nearby the houses (Al-Mekhlafi et al.,
2011a) (Table 2.8).
Univers
ity of
Mala
ya

50
Table 2.8: Socio-economic, behavioral and environmental risk factors associated with
acquiring malaria in four governorates in Yemen
Factors Governorates References
Socio-economic factors:-
Family size (more than 5)
Gender
Male
Female
Low income
Rural area
Age of individuals
age ≤ 12 year
Occupation (not working)
Distance to nearest health center >2
km
Driving time to reach health center
> 10 min
Poorly constructed houses
Material of the roof (earth)
Presence of opening in the roof
Presence of latrine:
Outside the house
Not at all
Traveling to another endemic area
In the last 2 month
Dhamar,and Raymah
Dhamar,and Raymah
Taiz, Hodiedah
Taiz, Hodiedah,
Dhamar,and Raymah
Dhamar,and Raymah
Taiz, Hodiedah
Taiz, Hodiedah
Taiz
Taiz
Taiz, Hodiedah,
Taiz
Taiz
Taiz
Taiz
Taiz
(Al-Mekhlafi et al.,
2011a)
(Al-Mekhlafi et al.,
2011a)
(Al-Taiar et al., 2006)
(Al-Mekhlafi et al.,
2011a)
(Al-Mekhlafi et al.,
2011a)
(Al-Mekhlafi et al.,
2011a)
(Al-Mekhlafi et al.,
2011a)
(Al‐Taiar et al., 2008)
(Al‐Taiar et al., 2008)
(Al-Taiar et al., 2006)
(Al‐Taiar et al., 2008)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
Behavioral factors:
Not sleeping under bed nets
Burning mosquito coils
Spray insecticides at home
Taiz
Taiz, Hodiedah,
Dhamar,and Raymah
Taiz
Taiz
(Al-Mekhlafi et al.,
2011a)
(Al-Mekhlafi et al.,
2011a)
(Al‐Taiar et al., 2008)
(Al‐Taiar et al., 2008)
Univers
ity of
Mala
ya

Continued
51
Burning animal dung
delay of treatment >3days
Wear short clothes
Dhamar,and Raymah
Taiz
Taiz
Taiz, Hodiedah,
Dhamar,and Raymah
(Al-Mekhlafi et al.,
2011a)
(Al-Taiar et al., 2009)
(Al‐Taiar et al., 2008)
(Al-Mekhlafi et al.,
2011a)
Environmental factors:
Water collections nearby
Presence of water stream/spring
Presence of water pump
Swamp existence/marshy land
Man-made water collection/tank
Water sources for house:
Well
Stream/spring
Water truck
Water storage at home:
In jerry cans
Distance from health center>2km
Taiz, Hodiedah,
Taiz
Taiz, Hodiedah,
Dhamar,and Raymah
Taiz
Taiz
Taiz
Taiz, Hodiedah,
Dhamar,and Raymah
Taiz
Taiz
Taiz
Taiz
Taiz
Taiz
Taiz
(Al-Mekhlafi et al.,
2011a)
(Al-Taiar et al., 2009)
(Al-Mekhlafi et al.,
2011a)
(Al-Taiar et al., 2009)
(Al‐Taiar et al., 2008)
(Al-Taiar et al., 2009)
(Al-Mekhlafi et al.,
2011a)
(Al-Taiar et al., 2009)
(Al‐Taiar et al., 2008)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al-Taiar et al., 2009)
(Al‐Taiar et al., 2008)
Univers
ity of
Mala
ya

52
2.3.4 Malaria distribution and intensity of transmission
In Yemen, malaria is seasonal and unstable, the malaria endemicity ranges from
mesoendemic in the south to hyperendemic in the north, especially the coastal plains,
including Tehama areas, foothill regions and coastal districts in the Hadhramout
governorate which is characterised by extensive wadis and seasonal rainfall (Yahia,
2005; Mohanna et al., 2007). From the past to present, the prevalence of malaria in
Yemen fluctuates and is a major health problem with prevalence ranging from 12.8% to
18.6% (Alkadi et al., 2006; Mohanna et al., 2007; Abdulsalam et al., 2010; Othman et
al., 2015).
Yemen has four major epidemiological stratification of malaria, and the
classification was based on the attitude, rainfall, and topography (Table 2.8) (NMCP,
2011; Adeel et al., 2015). The areas of the first stratum has an attitude of 0-600 meters
with an average of 4-6 months of rain. The transmission season mainly occur in winter
from November to April and is characterised by occurrence of malaria infection in
wadis along the coastal areas whereas the desert areas are malaria free zones (e.g
Hadhramout). The second stratum consists of an attitude of 601-1000 meters above the
sea level, and malaria transmission occurs in the winter season from November to April.
In addition, there is partial transmission in summer from May to September and is
characterised by occurrence of malaria infection in the Wadis (valleys), and in the
foothills (e.g. Tihama region). The third stratum has an attitude above 1001-2000
meters, malaria transmission occurs in the summer season, especially in the foothills
and wadis of the central highlands. The four stratum is the areas above 2000 meters and
the desert areas which are usually free from malaria. Hadhramout governorate is divided
into three zones which are, coastal plain, mountains and foothill areas (Yahia, 2005).
Univers
ity of
Mala
ya

53
The area is classified by the NMCP as belonging to stratum one and peak malaria
transmission occurs in winter between October and April.
2.3.5 Prevention and control
The National Control Malaria Program (NCMP) in Yemen, is proactive in combating
malaria through the implementation of several interventions that include distribution of
insecticide-treated mosquito nets (ITNs), indoor residual spraying (IRS), proper
diagnosis, proper treatment, and reactive and proactive case surveillance.
2.3.6 Malaria diagnosis
In Yemen, the diagnosis of malaria depends on clinical examination and confirmation
by microscopic detection of malaria parasites in blood smear. The microscopic
examination is usually conducted in the main hospitals and health centers where trained
technicians are present. However, this technique may not be available in the rural areas
due to the lack of required facilities and qualified health workers. Furthermore, WHO
reported that the standard of microscopy examination in Yemen is poor due to the lack
of effective national standards; poor quality of blood films, poor quality stains and
staining techniques, generally unsatisfactory laboratory equipment; and the absence of
an effective quality assurance program (WHO, 2009). Therefore, malaria rapid
diagnostic test (RDT) was introduced as alternative tool for malaria diagnosis in areas
where good quality microscopy is not available or cannot be carried out (McMorrow et
al., 2010; WHO, 2015b).
Univers
ity of
Mala
ya

54
2.3.7 Malaria treatment in Yemen
2.3.7.1 The old strategy (from 1999)
The national antimalarial drug policy in Yemen was formulated in 1999, consisting of
chloroquine (CQ) as first-line, sulphadoxine-pyrimethamine (SP) as a second line and
the third line is mefloquine and primaquine as a gametocytocidal treatment
monotherapy for treating uncomplicated falciparum malaria. However, quinine
intravenous infusion was used for treating complicated and severe falciparum malaria.
In addition, chloroquine and primaquine treatment drug were used for treating non-
falciparum malaria as anti-relapse treatment for Plasmodium vivax and as
gametocytocidal treatment for Plasmodium malariae (NMCP, 2006).
2.3.7.2 The new strategy (from 2005)
In November 2005, following the emergence of chloroquine resistance and the WHO
recommendation, the antimalarial treatment policy shifted to artemisinin-based
combination therapy (ACT) with artesunate (AS) plus sulphadoxine-pyrimethamine
(SP) as the first-line, and artemether-lumefantrine (AL) as the second line therapy for
treating uncomplicated falciparum malaria (Adeel et al., 2015). However, this new
policy was only implemented four years later in 2009 after proper training and
distribution of the national guideline for antimalarial drugs were carried out (Bin
Ghouth, 2013). Artemether or quinine infusion therapy was used for treating
complicated and severe falciparum malaria. In addition, the treatment of non-falciparum
malaria is still chloroquine and primaquine as an anti-relapse treatment for P. vivax
(NMCP, 2010a, 2010b; WHO, 2012; Bin Ghouth, 2013).
Univers
ity of
Mala
ya

55
2.3.7.3 Monitoring anti-malarial drug resistance
A) In vivo studies
Monitoring antimalarial drug efficacy in Yemen started in 2002 to 2005 following the
WHO protocol for in vivo assessment in four sentinel sites that found 39% to 57% of
chloroquine resistance. In 2004, three in vivo studies on the efficacy of SP showed
success rate ranging from 95% to 100%. Four years later, after launching the new
policy, in vivo efficacy trials were conducted in three monitoring sites and they reported
of 97.6–100 % adequate clinical and parasitological response (ACPR) for AS plus SP
(NMCP, 2010b). The efficacy of AS plus SP as first-line treatment for uncomplicated
falciparum malaria was also rated at 97% ACPR in a recent clinical drug efficacy trial
carried out in 2013 (Adeel et al., 2015). It is noteworthy that the currently used routine
clinical efficacy trial is the gold standard for the assessment of the efficiency of the
combined antimalarial drugs, although it does not differentiate between the
effectiveness of AS and its partner drug.
B) Molecular markers based studies
For many years, CQ had been the first line treatment in Yemen. The first case of the
indigenous chloroquine resistance (CQR) in Yemen was reported in 1989 in Taiz
(Mamser, 1989; Alkadi et al., 2006), and then in Hodeidah (Al-Shamahy et al., 2006).
In addition, recent studies have shown high prevalence of CQR marker Pfcrt 76T in
Hodeidah, Dhammar, Rymah and Taiz (Al-Mekhlafi et al., 2011b; Abdul-Ghani et al.,
2013; Al-Hamidhi et al., 2013). Although antimalarial drug policy in Yemen has
changed from CQ to ACT, previous studies conducted in Hadhramout governorate
Univers
ity of
Mala
ya

56
reported that CQ is still commonly prescribed (18 out of 42 prescriptions) and some
clinicians were not aware and had poor knowledge about the new national drug policy
(Bashrahil et al., 2010; Bin Ghouth, 2013). Continued use of CQ sustains the selection
of CQ resistant mutations leading to persistence of mutant parasite. The complete
withdrawal of CQ use may enhance the emergence of CQ sensitive parasite over time
and make CQ possible to be re-introduced for malaria treatment (Kublin et al., 2003;
Laufer et al., 2006). However, the persistence of CQ resistance will be prolonged if the
shift to ACT and the simultaneous withdrawal of CQ are not rigorously implemented.
Molecular markers are practical for monitoring SP resistance. Quintuple mutant of
combined dihydrofolate reductase (dhfr) and dihydropteroate synthase (dhps) genes
(Pfdhfr I51, R59, N108 plus Pfdhps G437, E540) was significantly associated with in
vivo resistance to SP (Picot et al., 2009). Several studies have also been conducted for
the screening of P. falciparum population for molecular markers associated SP
resistance in Yemen. The mutant allele R59 of Pfdhfr was detected in 5 % of P.
falciparum isolates (5/99) in Lahj governorate, southern Yemen (Mubjer et al., 2011).
Double mutant genotype of Pfdhfr (I51/N108) was reported in 54 % of P. falciparum
isolates in Taiz, Dhamar, and Hodeidah governorates in western Yemen (Al-Hamidhi et
al., 2013). Pfdhfr mutant allele (N108) was also reported in 53.2 % of P. falciparum
isolates collected from Hodeidah governorate (Abdul-Ghani et al., 2014). The continued
use of SP in the new policy, availability of this drug in the private sector, and poor
knowledge of the national policy among physicians (Bashrahil et al., 2010) may
increase the monotherapy of SP against P. falciparum, which is likely to compromise
drug efficacy. It is noteworthy that the data on molecular markers associated with CQ
and SP resistance are not available from the Hadhramout governorate, Yemen where
this study is being carried out.
.
Univers
ity of
Mala
ya

57
CHAPTER 3: METHODOLGY
3.1 OVERVIEW OF STUDY METHODS
A cross–sectional household survey was carried out in Hadhramout governorate, which
is located in the southeast of Yemen. A total of 735 participants aged 1-75 years and
genders were enrolled in this study. Participants were from three villages in Hajer
district and four villages in Al-Raydah-Qusyer district. Questionnaire data and blood
samples were collected during transmission seasons from July 2011 to May 2012. These
data were then analysed using the Statistical Package for Social Sciences for Windows
(SPSS) version 23.0. Blood from each individual was smeared on a glass slide as well
as spotted on a Whatman filter paper 3MM (Whatman International Ltd., Maidstone,
England). Blood smears were brought back from the field and stained as soon as
possible with 10% diluted Giemsa stain and screened under a microscope for the
presence of malaria parasites in the laboratory. Then, the positive specimens for
Plasmodium falciparum were used for molecular identification and genotyping studies
(Figure 3.1).
Univers
ity of
Mala
ya

58
Figure 3.1: Schematic diagram of samples and data collection and molecular
marker detections
Univers
ity of
Mala
ya

59
3.6.2 Ethical clearance
The study protocol was approved by the Faculty of Medicine, Hadhramout University
for Science and Technology, the Ministry of Health and Population, Yemen, and the
Malaria National Control Program division in Hadhramout governorate, Yemen.
Informed consent was obtained from each participant, and for children, consent was
obtained from their parents after a clear explanation of the study objectives
(APPENDIX B).
3.3 STUDY AREAS AND STUDY POPULATION
The study was conducted in the Hadhramout governorate in the southeast Yemen, the
largest governorate, accounting for half of the country’s surface area. The population of
this governorate was estimated at 1,028,556 (CSO, 2004). Hadhramout has a humid and
hot climate which is characterised by humidity levels ranging from 18 to 93% and
temperature ranging from 18 to 38°C. In general, Hadhramout climate is unstable and
changes from time to time. For example, Hadhramout governorate faced tropical storm
and flash flood in October of 2008 (Breisinger et al., 2012) and tropical cyclone
Chapala which caused heavy flooding in the southeastern governorates of Hadhramout,
Shabwa and along the coast of Arabian Sea in November of 2015. These events might
affect household health, densities of mosquito vectors, reduce incomes and subsequently
food security levels (Hubálek & Halouzka, 1999; McMichael et al., 2006; Breisinger et
al., 2012).
Malaria transmission in Yemen differs between the regions. Hadhramout has
coastal plain region and mountainous area. The coastal plain region (with an altitude of
0–600 m) is characterised by hot climate throughout the year with irregular rainfall
Univers
ity of
Mala
ya

60
ranging between 50 and 100 mm per annum. A total of 735 participants aged 1-75 years
and genders were enrolled in this study; 221 participants from three villages in Hajer
district and 514 participants from four villages in Al-Raydah-Qusyer district, houses
were selected randomly (Figure 3.2), and some of population in rural area lived in
poorly constructed houses of mud or uncemented brick walls with man-made water tank
located beside the house for water storage (APPENDIX B). These villages were
selected because they are endemic malaria areas with more than 99 % of cases being
caused by P. falciparum and few cases of Plasmodium vivax.
Univers
ity of
Mala
ya

61
Figure 3.2: Map of study area highlighted in the Hadhramout governorate, Yemen
Univers
ity of
Mala
ya

62
3.3 DESIGN OF STUDY
A community-based cross-sectional study was carried out in seven endemic villages,
three in Hajer district and four villages in Al-Raydah-Qusyer district.
3.4 SAMPLE SIZE
Sample size required for this study was estimated according to the previous prevalence
of malaria reported in four recent studies in Yemen (Al-Taiar et al., 2006; Alkadi et al.,
2006; Mohanna et al., 2007; Abdulsalam et al., 2010) which was rated at 18 %. By
using the formula developed by Lwanga and Lemeshow (1991) and according to the
following parameters: 18% as the expected malaria prevalence, 95% confidence level
and P ≤ 0.05, the minimum number of sample size needed for this study was 196-246
subjects. The formula is as the following:
n =z2 𝑃 (1−𝑃)
𝑑2 (Eq. 3.1)
where n= sample size,
Z = Z statistic for a level of confidence,
P = expected prevalence or proportion (in proportion of one; if 20%, P = 0.2), and
d= precision (in proportion of one; if 5%, d = 0.05).
Z statistic (Z): For the level of confidence of 95%, which is conventional, Z value is
1.96.
Univ
ersity
of M
alaya

63
3.5 DESCRIPTION OF VARIABLES
This study has a dependent variable, which takes the form of yes or no responses to
malaria infection and main outcomes such as point mutations of Pfcrt gene at 76, 271,
326, 356 and 371 positions, Pfmdr-1 gene at 86 and 1246 positions and point mutations
of Pfdhfr and Pfdhps genes at different positions. The independent variables such as
age, gender, education level, occupation, socio- economic factors, behaviour factors,
environmental factors, level of parasitemia. The definition of variables are listed in
APPENDIX D.
3.6 DATA AND SAMPLE COLLECTION
3.6.1 Strategy of field work
A household survey from house-to-house was conducted to collect questionnaire data
and blood samples by surveyors who had previous experience with malaria surveys in
the endemic districts in the Hadhramout governorate. Collection was done according to
recommendations from the Malaria National Control Program Unit in Al-Mukalla,
Yemen. All household visits occured during the period from July 2011 to May 2012.
3.6.2 Questionnaire
A pretested standard questionnaire which was developed by the World Health
Organization for malaria prevalence survey was used to collect information about
personal profile, socioeconomic, and environmental background. Firstly, the
questionnaire which was written in English was translated into Arabic, the mother
tongue language of Yemen (APPENDIX E). A checklist was used for malaria clinical
Univers
ity of
Mala
ya

64
signs and symptoms, as observed by a team of physicians, and any history of previous
antimalarial treatment was documented. Knowledge, attitude and practices (KAP) were
investigated using a standard questionnaire. The data were collected from the household
members, or from the parents on behalf of children, via face-to-face interviews
conducted by well-trained interviewers. During the interviews, direct observation was
made for the type of household building, wall, floor; for the availability and the type of
toilet facilities, piped water, clothes-wearing habits, electricity, telephone, mosquito
nets, and finally the presence of nearby pools or rivers. The signs and symptoms
recorded included fever and jaundice.
3.6.3 Blood sampling
Blood samples were collected by the finger prick method and thin and thick blood
smears were made, allowed to air-dry (the thin smears were fixed with methanol within
three hours), and then brought back to the laboratory to be stained with Giemsa
(APPENDIX F). Haemoglobin levels were measured in the field directly from capillary
blood using the HemoCue haemoglobinometer (HemoCue, AB, Angelhom, Sweden).
Haemoglobin levels were considered as normal (>11 g/dl), low anaemia (9-11 g/dl),
moderate anaemia (7-8.9 g/dl), and severe anaemia (<7 g/dl). Three drops of blood were
spotted on Whatman filter paper 3MM (Whatman International Ltd., Maidstone,
England) and kept separately in clean, dry and well-sealed plastic bag with silica gel to
reduce humidity in the bag and stored at room temperature until further use for
molecular analysis. The coordinates of each village were recorded using a global
positioning system (GPS) (Garmin GPSMAP 60CSx, Tonopah, AZ, USA).
Univers
ity of
Mala
ya

65
3.7 DETECTION OF MALARIA PARASITE BY MICROSCOPY METHOD
3.7.1 Staining blood smears
Thin and thick blood smears were stained with 10% Giemsa stain and examined by
three trained malaria microscopists according to standard procedures (APPENDIX F).
3.7.2 Microscopy examination
Species identification was performed in the laboratory of the National Malaria Control
Programme in Hadhramout governorate by three expert microscopists according to the
the key morphological differences between the blood stages of human Plasmodium
species (APPENDIX G). Parasitaemia per µl of blood was calculated from thick smears
by counting the number of asexual parasites per 200 leukocytes using an assumed
leukocyte count of 8000 WBC/µl (Trape, 1985; Singh et al., 1999; Moody & Chiodini,
2000). A negative result was recorded after screening at least 200 fields under the oil
immersion lens of the light microscope. Parasitaemia was expressed as the total number
of Plasmodium asexual forms per microliter of blood. Parasite levels were classified as
low (1 - 999/μL), moderate (1000 - 9999/μL), or high (>10000/μL) (Bouyou-Akotet et
al., 2003). The positive specimens for Plasmodium were used for molecular studies.
Univers
ity of
Mala
ya

66
3.8 MOLECULAR IDENTIFICATION AND GENOTYPING OF MALARIA
SPECIES
Nested PCR was used to detect malaria parasites in blood spot samples. Subsequently,
only falciparum malaria specimens were used for further molecular genotyping
analysis.
3.8.1 DNA extraction
Genomic DNA was extracted from filter paper blood spots. Briefly, by using a sterile-
flamed puncher, a small disc of the filter paper blood spot (approximately 5 mm
diameter) was punched out and transferred into microcentrifuge tubes using a clean and
methanol-flamed forceps. Genomic DNA sample was extracted using Qiagen DNA
Mini Kit for blood and tissue (QIAGEN, DNeasy® blood and tissue kit, Cat. No. 69506,
Germany) by following the manufacturer’s instructions. Extracted DNA was eluted
using 50µL Qiagen AE elution buffer (0.5 mM EDTA, 10 mM Tris-Cl, pH 9.0) and
kept at −20°C until further use (APPENDIX H).
3.8.2 Molecular identification of malaria species
The small subunit ribosomal RNA gene (18SSU rRNA) was targeted in the present
study since it is most commonly used for diagnosis of malaria parasitic infections and
other molecular analysis. The detection of malaria parasites (P. falciparum, P. vivax, P.
malariae, P. ovale and P. knowlesi) was achieved using a nested PCR protocol
developed by Singh et al. (1999). The primary nested PCR reaction was achieved using
genus specific primers sets and yielded an amplicon of about 1.2 kb fragment, whereas,
secondary genus specific nested PCR produced fragment of about 240bp for all
Univers
ity of
Mala
ya

67
Plasmodium parasites. Another secondary nested PCR was run by using species specific
primer sets for the identification of P. falciparum (205 bp), P. vivax (117bp), P.
malariae (144 bp), P. ovale (787 bp) and P. knowlesi (153 bp).
Plasmodium species were identified using nested PCR based on small subunit ribosomal
RNA genes (18SSU rRNA). PCR master mix and thermal cycling conditions were
carried out by using five pairs of oligonucleotide primers as reported previously
(Snounou, 1996; Singh et al., 1999). First amplification was to amplify a fragment of
18SSU rRNA genes of different Plasmodium species by using primary and secondary
nested PCR in which genus specific primers were used. Second amplification was to
identify the five Plasmodium species by using a secondary nested PCR in which five
pairs of species specific primers were used (Table 3.1).
PCR reaction for genus specific amplifications was carried out in a final volume
of 50 µL in a PCR tube containing 4 mM MgCl2, 200mM of each deoxynucleoside
triphosphate (dNTPs), 250 nM of rPLU1 primer, 250 nM of rPLU5 primer and 5 µL of
genomic DNA, 1X buffer of PCR (containing 10 mM Tris-HCI, 50 mM KCl), and 1.25
units of Taq polymerase. PCR reagent and primers were from iNtRON (iNtRON
Biotechnology, Inc., Seoul, Republic of Korea). PCR was performed in the thermal
cycler (MyCycler, BioRad Hercules, USA) with the following cycling conditions: initial
denaturation at 94°C for 10 min, 40 cycles of denaturation at 94°C for 30 sec, annealing
at 55°C for 60 sec, extension at 72°C for 60 sec and final extension at 72°C for 5 min.
About 2 µL of primary PCR product was used as template in the secondary nested PCR-
species specific amplifications. PCR reactions contained the same PCR reagent
concentrations as in the primary PCR except using 1.5 mM MgCl2 and amplification
was set for 25 cycles instead of 45 cycles. The cycling conditions and reagents
Univers
ity of
Mala
ya

68
concentrations were similar to the primary PCR except the annealing and Taq DNA
polymerase were 58°C and 0.5 units, respectively. The PCR products were loaded in a
2% (w/v) agarose gel with 1X TAE buffer (Tris acetate EDTA), stained with SYBER ®
safe DNA gel stain (Invitrogen, USA). Ten µl from each PCR product were mixed with
2 µl of 6X loading dye and loaded into the wells of agarose gel and the electrophoresis
was run for 30 minutes at 100 Volt. The sizes of amplicons were measured against 100
bp DNA ladder (iNtRON Biotechnology, Inc., Seoul, Republic of Korea) through
visualization through UV transilluminator.
Univers
ity of
Mala
ya

69
Table 3.1: Protocol for the detection of Plasmodium malarial species based on 18SSU rRNA gene
C°:degree centigrade, s.: second, m.: minutes, Den.: Denaturation, Ann.: Annealing, Ex.: Extention, Cyc.: Cycle
PCR
reactions
Primer
name Target Sequence (5'-3')
PCR profiles Product
size
Den.
C°/m
Den.
C°/s
Ann.
C°/s
Ex.
C°/s
Final ex.
C°/m
Cyc
.
Primary
PCR
rPLU1 18SSU
rRNA genes
TCA AAG ATT AAG CCA TGC AAG TGA 95/4 94/30 55/60 72/60 72/10 40 1.2 kp
rPLU5 CCT GTT GTT GCC TTA AAC TCC
Genus-
specific
nested PCR
rPLU3 Plasmodium
species
TTT TTA TAA GGA TAA CTA CGG AAA AGC TGT 95/10 94/20 55/20 72/60 72/5 40 240 bp
rPLU4 TAC CCG TCA TAG CCA TGT TAG GCC AAT ACC
Species-
specific
nested PCR
rFAL1 Plasmodium
falciparum
TTA AAC TGG TTT GGG AAA ACC AAA TAT ATT
95/10 94/20 58/20 72/60 72/5 40
205 bp rFAL2 ACA CAA TGA ACT CAA TCA TGA CTA CCC GTC
rVIV1 Plasmodium
vivax
CGC TTC TAG CTT AAT CCA CAT AAC TGA TAC' 117 bp
rVIV2 ACT TCC AAG CCG AAG CAA AGA AAG TCC TTA'
rMAL1 Plasmodium
malariae
ATA ACA TAG TTG TAC GTT AAG AAT AAC CGC 144 bp
rMAL2 AAA ATT CCC ATG CAT AAA AAA TTA TAC AAA
rOVAL1 Plasmodium
ovale
ATC TCT TTT GCT ATC TTT TTT TAG TAT TGG AGA 787 bp
rOVAL2 GGA AAA GGA CAC ATT AAT TGT ATC CTA GTG
Pmk8 Plasmodium
knowlesi
GTT AGC GAG AGC CAC AAAAAA GCG AAT 153 bp
Pmk9 ACT CAA AGT AAC AAA ATCTTC CGT A
Univers
ity of
Mala
ya

70
3.8.3 Molecular detection of mutation in Pfcrt gene at codon K76T
A nested mutation-specific PCR was used to detect mutations in Pfcrt K76T (Djimdé et
al., 2001a). Primary PCR was performed using two sets of primer (CRTP1 and CRTP2)
to amplify 537 bp of Pfcrt gene. The secondary PCR was performed using a common
inner primer (CRTP3) together with either CRTP4m or CRTP4w for mutant and wild
types, respectively, resulting in 366 bp of the two alleles (Table 3.2). PCR reaction was
carried out in a final volume of 25µL in a PCR tube containing 2.5 mM MgCl2, 200mM
of each dNTP, 1.25 units of Taq polymerase, 1mM of CRTP1 primer, 1mM of CRTP2
primer and five microliter of genomic DNA. PCR reagent and primers were from
iNtRON (iNtRON Biotechnology, Inc., Seoul, Republic of Korea). PCR was performed
in a thermal cycler (MyCycler, BioRad Hercules, USA) with the following cycling
conditions: initial denaturation at 95°C for 3 min, 45 cycles of denaturation at 94°C for
30 sec, annealing at 56°C for 30 sec, extension at 60°C for 1 min and final extension at
60°C for 5 min. About 1–2µL of primary PCR product was used as template in the
secondary PCR reaction which contained the same PCR reagent concentrations as in the
primary PCR except using 1.5 mM MgCl2 and amplification cycle of 25 cycles instead
of 45 cycles. The cycling conditions were similar to the primary PCR except that the
annealing and extension temperatures were 47°C and 64°C, respectively. The PCR
products were loaded in a 2% (w/v) agarose gel, stained with SYBER ® safe DNA gel
stain (Invitrogen, USA) for electrophoresis and visualized by UV transilluminator.
Univers
ity of
Mala
ya

71
Table 3.2: Detection of point mutations in Pfcrt , Pfmdr1, Pfdhfr, and Pfdhps genes at
different codons
Gene codons Analysis
method
Products
Size (N2)
Endo.
Dig.
(R.E)
Post-Dig.
Products
band
Geno-
type
Amino
acids
sub.
(W-M)
Pfcrt K76T MS-Nested
PCR
366 Specific - - Lys-Thr
Pfcrt Q271E Nested
PCR+RFLP
111 XMN1 50+61 Mutant Gln-Glu
Pfcrt N326S Nested
PCR+RFLP
68 Mse1 24+44 Wild Asn-Ser
Pfcrt I356T Nested
PCR+RFLP
100 AlwN1 40+60 Mutant Ile-Thr
Pfcrt R371I Nested
PCR+RFLP
80 Afl II 40+40 Wild Arg-Ile
Pfmdr1 N86Y Nested
PCR+RFLP
291 Afl III 126+165 Mutant Asn-Tyr
Pfmdr1
D1246Y
Nested
PCR+RFLP
203 Bg1 II 113+90 Wild Asp-Tyr
Pfdhfr at
different
codons
Sequencing
and BioEdit
software
700 - - - -
Pfdhps at
different
codons
Sequencing
and BioEdit
software
711 - - - -
MS-Nested PCR: Mutant-Specific Nested Polymerase Chain Reaction.
RFLP: Restriction Fragment Length Polymorphism.
N2: Size of secondary PCR product
Endo. Dig (RE): Endonuclease Digestion Restriction Enzyme.
Post-Dig.: Post Digestion
Amino acids sub. (W-M): Amino acids substitution (wild to mutant)
Univers
ity of
Mala
ya

72
3.8.4 Molecular detection of point mutations in Pfcrt gene (positions Q271E, N326S,
I356T, R371I) and Pfmdr1 gene (positions N86Y and D1246Y)
Nested PCR followed by restriction fragment length polymorphism was performed as
previously described (Djimdé et al., 2001a) (Table 3.3). Briefly, the restriction enzymes
XMN1, MSe1, AlwN1 and AflII digest Pfcrt at codons 271, 326, 356 and 371,
respectively. The enzymes AflIII and Bg1II digest Pfmdr1 at codons 86 and 1246,
respectively. The digestion of PCR product was achieved by Fast Digest restriction
enzymes (New England biolabs Inc., USA) according to the instructions of the
manufacturer. Digestion results were analysed by electrophoresis in a 2.5% (w/v)
agarose gel containing SYBER ® safe DNA gel stain (Invitrogen, USA) and visualized
in a UV transilluminator. Genomic DNA from P. falciparum strains HB3, 3D7 and Dd2
(supplied by Malaria Research and Reference Reagents Resources Centre (MR4,
ATCC, ManassasVA, USA) were used as positive controls for mutant and wild types,
whereas nuclease-free water was used as the negative control (Table 3.2).
3.8.5 Molecular detection of point mutations in Pfdhfr gene at different codons
Genomic DNA of Pfdhfr gene was amplified using nested PCR following the method
described previously (Plowe et al., 1995; Tinto et al., 2007). Briefly, an amplicon of
720 bp was amplified using the primers pair AMP 1: (5'-TTTATATTTTCTCCTTTTT
A-3') and AMP 2: (5'-CATTTTATTATTCGTTTTCT-3') in the primary PCR, and an
amplicon of 700 bp was amplified using the primers SP1: (5'-ATGATGGAACAA
GTCTGCGAC-3') and SP2: (5'-ACATTTTATTATTCGTTTTC-3') in the nested PCR
(Table 3.3). The PCR reaction was carried out in a total of 25μl mixture containing 1x
PCR buffer, 3 mM MgCl2, 0.2 mM of dNTPs, 200 nM of each primer, 1 U of Taq
polymerase and 4 μl of genomic DNA. Cycling condition was as follows; initial
Univers
ity of
Mala
ya

73
denaturation at 94°C for 3 min, followed by 45 cycles of denaturing for 30 sec at 94°C,
annealing for 1 min at 43.5°C and extension at 72°C for 45 sec, and final extension at
72°C for 5 min. The cycling condition for nested PCR was the same except that
annealing was at 55°C for 45 sec and extension at 74°C for 35 sec, the number of cycles
was decreased to 35 cycles (Table 3.3). PCR reagent and primers were obtained from
iNtRON (iNtRON Biotechnology, Inc., Seoul, Republic of Korea). PCR product was
analyzed by electrophoresis in a 2.5% (w/v) agarose gel containing SYBER® safe DNA
gel stain (Invitrogen, USA) and visualized in a UV transilluminator. PCR products were
purified with Presto™ 96 Well PCR Cleanup Kits and then sequenced in both directions
using the inner primers in the ABI 3730xl DNA analyzer (Applied Biosystems).
Mutations were detected by creating consensus sequences and comparing manually with
the sequences in GenBank (GenBank accession number was XM_001351443 for
Pfdhfr) using BioEdit software (Hall, 2011).
3.8.6 Molecular detection of point mutations in Pfdhps gene at different codons
An amplicon of 711 bp of Pfdhps gene was amplified by nested PCR using the outer
primers pair O1: (5'-GATTCTTTTTCAGATGGAGG-3') and O2: (5'-TTCCTCATGT
AATTCATCTGA-3'), and the nested primers N1 (5'-AACCTAAACGTGCTGTTCAA-
3') and N2: (5'-AATTGTGTGATTTGTCCACAA-3') (Pearce et al., 2003) (Table 3.3).
The PCR mixture was as described above. The cycling conditions for primary and
secondary PCR were as follows: initial denaturation at 94°C for 3 min followed by 25
cycles of denaturing for 1 min at 94°C, annealing for 2 min at 52°C and extension at
74°C for 1 min and final extension at 74°C for 5 min. PCR reagent and primers were
obtained from iNtRON (iNtRON Biotechnology, Inc., Seoul, Republic of Korea). PCR
product was analysed by electrophoresis in a 2.5% (w/v) agarose gel containing
Univers
ity of
Mala
ya

74
SYBER® safe DNA gel stain (Invitrogen, USA) and visualized in a UV
transilluminator. PCR products were purified with Presto™ 96 Well PCR Cleanup Kits
and then sequenced in both directions using the inner primers in the ABI 3730xl DNA
analyzer (Applied Biosystems). Mutations were detected by creating consensus
sequences and comparing manually with the sequences in GenBank (GenBank
accession number was Z30654 for Pfdhps) using BioEdit software (Hall, 2011).
Univers
ity of
Mala
ya

75
Table 3. 3: Forward and reverse primers sequences for Pfcrt , Pfmdr1, Pfdhfr, and Pfdhps genes at different codons
Gene
codon
Primers
for
primary
PCR
Sequence (5'-3') Primers for
secondary
PCR
Sequence (5'-3')
Pfcrt 76 CRTP1 CCGTTAATAATAAATACACGCAG
Common
CRTP3 TGACGAGCGTTATAGAG
CRTP2 CGGATGTTACAAAACTATAGTTACC Wild
CRTPw GTTCTTTTAGCAAAAATCT
Mutant
CRTP4m GTTCTTTTAGCAAAAATTG
Pfcrt
271 CRT-2A CCCAAGAATAAACATGCGAAAC CRT271a GGCACATTCATTTATTTATTTTTTCTTTCCTAATTAATGAATACGTT
CRT-2B ACAATTATCTCGGAGCAGTT CRT271b GGCTATGGTATCCTTTTTCC
Pfcrt
300s:
(326,
356 &
371
CRT-3a CCTTGGCATTGTTTTCCT CRT326a CCTTTTTATTCTTACATAGCTGGTTATTGAATTATCAC
CRT-3b CCAAAGTTACGAAATCTAATAATCTTGG
CRT326b TGGCATTGTTTTCCTTCT
CRT356a ATATATATGGCTAAGAATTTAAAGTAATAAGCAGTTGCT
CRT356b AATTATCGACAAATTTTCTACC
CRT371a TATTATTTTTACTTTTTAATTTTATAGGGTGATGTCTTAA
CRT371b AAGTTACGAAATCTAATAATCTTGGTTC
Pfmdr1-
86 MDR-A GCGCGCGTTGAACAAAAAGAGTACCGCTG MDR-D1 TTTACCGTTTAAATGTTTACCTGC
MDR-B GGGCCCTCGTACCAATTCCTGAACTCAC MDR-D2 CCATCTTGATAAAAAACACTTCTT
Pfmdr1-
1246 1246-A GGGGGATGACAAATTTTCAAGATTA 1246-D1 AATGTAAATGAATTTTCAAACC
1246-B GGGGGACTAACACGTTTAACATCTT 1246-D2 CATCTTCTCTTCCAAATTTGATA
Pfdhfr Amp1 TTTATATTTTCTCCTTTTTA SP1 ATGATGGAACAAGTCTGCGAC
Amp2 CATTTTATTATTCGTTTTCT SP2 ACATTTTATTATTCGTTTTC
Pfdhps O1 GATTCTTTTTCAGATGGAGG N1 AACCTAAACGTGCTGTTCAA
O2 TTCCTCATGTAATTCATCTGA N2 AATTGTGTGATTTGTCCACAA Univers
ity of
Mala
ya

76
3.9 STATISTICAL ANALYSES
Data were analyzed using the Statistical Package for Social Sciences (SPSS) version 23
(SPSS Inc., Chicago, IL, USA). The prevalence of a mutant allele or genotype was
calculated as the percentage of the presence of the mutant allele or the genotype in the
examined P. faclciparum isolates. The difference between proportions of variables was
tested using Pearson Chi-Square test or Fisher’s exact test where applicable. 95%
confidence interval (CI) and odd ratios (OR) were computed. A stepwise conditional
logistic regression model was developed for those variables with P value < 0.05 which
was considered significant.
Univers
ity of
Mala
ya

77
CHAPTER 4: RESULTS
4.1 EPIDEMIOLOGICAL RESULTS OF MALARIA IN THE HADHRAMOUT
GOVERNORATE, YEMEN
4.1.1 Characteristic of study population
A total of 735 voluntarily consented individuals from seven villages in two districts of
Hadhramout, Yemen participated in this study. These consisted of 221 participants from
three villages in Hajer district and 514 participants from four villages in Al-Raydah-
Qusyer district, Hadhramout governorate. Of the 735 individuals, 423 (57.6%) were
males and 312 (42.4%) were females. The age of participants ranged from 1 to 75 years
with a median of 16 years and 22 interquartile range. Out of 735, 18.8% (138) were
positive for malaria parasite via microscopy (Figure 4.1). Majority of samples (393;
53.5%) were collected from those above 15 years old (defined as adults), 152 (20.7%)
from those between 5 and 9 years old, 142 (19.3%) from those between 10 – 15 years
old and the least number of samples came from those who were less than five years old
(6.5%). Fourty-seven percent of study population had no formal education and 61.6%
were farmers. Approximately half of the study population had no access to electricity or
communication media such as radio or television. The study population lived in simple
houses of mud or uncemented brick walls and mud or cement floors, with an uncovered
tank located beside the house for water storage (Table 4.1). No association was
observed between the presence of malaria and age and gender of the participants.
Univers
ity of
Mala
ya

78
Figure 4.1: Malaria prevalence in endemic areas of the two districts (i.e, Al-Raydah-
Qusyer and Hajer) of Hadhramout governorate, Yemen
18.8%
81.2%
Positive
Negative
Univers
ity of
Mala
ya

79
Table 4. 1: Demographic characteristics of study populations in Hadhramout
governorate, Yemen
Characteristics
Factors
Number (%)
Gender Male 423 (57.6)
Female 312 (42.4)
Age (years) >15 393 (53.5)
10 – 15 142 (19.3)
5 - 9 152 (20.7)
<5 48 (6.5)
Districts Hajer 221 (30.1)
Al-Raydah and Qusyer 514 (69.9)
Family size >5 members 290 (39.5)
≤5 members
445 (60.5)
Education Not educated 345 (46.9)
Primary 356 (48.4)
Secondary 34 (4.6)
Occupation Not working 180 (24.5)
Farmer 453 (61.6)
Fisherman 26 (3.5)
Government employees
76 (10.3)
Economic status Houses with electricity 379 (51.6)
Availability of TV 295 (40.1)
Availability of telephone 43 (5.9)
Availability of radio 385 (52.4)
Availability of fridge 295 (40.1)
Having motorcycle 148 (20.1)
Having car 212 (28.8)
Univers
ity of
Mala
ya

80
4.1.2 Prevalence of malaria and identify the risk factors associated with malaria in the
Hadhramout governorate, Yemen
Based on microscopy examination, the overall malaria prevalence in the individuals
sampled in the Hadhramout governorate, Yemen was 18.8% (138 of 735), where
majority of cases was due to Plasmodium falciparum as the predominant species
(99.3%; 137 of 138) followed by P. vivax (0.7%; 1 of 138). A majority of positive
individuals (57.7%) aged > 15 years old. The overall prevalence in the districts of Al-
Raydah-Qusyer and Hajer were 21.8% and 11.8%, respectively. In addition, this broad
prevalence variation was also noted between the villages, with Qusyer showing the
highest prevalence (31.8%) and Al-Raydah the lowest (5.6%). There were more positive
samples from male (20.3%) as compared to female (16.7%) and the sex ratio was 1.7
males/females (Table 4.2).
In addition, the number of positive individuals was higher in adult males than in
children and adult females. Persons whose household’s head had primary education
were at higher risk of being infected (OR= 10.1, 95% CI: 1.35 – 74.5), as did fishermen
(OR=11.3, 95%CI: 3.13 – 40.5) and farmers (OR= 4.84, 95%CI: 1.73 – 13.6) (Table
4.3). A number of socioeconomic indicators were also associated with increased
prevalence: living in houses with walls made of uncemented bricks (OR= 2.1, 95% CI:
1.32 – 3.30), no access to toilets (OR= 1.6, 95%CI: 1.05 – 2.32), no fridge (OR=1.6,
95%CI: 1.05 – 2.30), or no TV (OR=1.6, (95%CI: 1.05 – 2.30). Individuals living in
houses with a distance of water collection points less than 200 meters were also at
higher risk of acquiring malaria (OR= 1.6, 95%CI: 1.05 – 2.30). Multivariate analysis
using stepwise forward logistic regression confirmed that the significant risk factors
Univers
ity of
Mala
ya

81
were living in uncemented brick wall houses, or being a fisherman or a farmer or living
in houses with a distance of water collection points less than 200 meters (Table 4.3).
Univers
ity of
Mala
ya

82
Table 4.2: Prevalence and distribution of malaria among population in Hadhramout
governorate, Yemen according to areas
Characteristics
Examined (n)
Infected
(%)
P value
Districts
Hajer 221 26 (11.8) 0.001*
Al-Raydah and Qusyer 514 112 (21.8)
Hajer District Villages
Kunina 83 5 (6) 0.001*
Kinina 47 12 (25.5)
Jol-Bamejah 91 9 (9.9)
Al-Raydah and Qusyer
District Villages
Hadhathim 34 10 (29.4)
Al-Raydah 18 1 (5.6)
Qusyer 22 7 (31.8)
Al-Rahbah 440 94 (21.4)
n: number of subjects
*Significant association at p< 0.05
Univers
ity of
Mala
ya

83
Table 4.3: Risk factors associated with malaria in Hadhramout governorate, Yemen.
Characteristics
Factors
Examined
Infected (%)
OR (95%CI)
Age (years) >15 393 79 (20) 1
10 – 15 142 25 (17.6) 0.85 (0.51 – 1.40)
5 - 9 152 30 (19.7) 0.98 (0.61 – 1.56) <5 48 4 (8.3) 0.36 (0.13 – 1.04)
Gender
Female 312 52 (16.7) 1
Male 423 86 (20.3) 1.04 (0.98 – 1.12)
Education
level of
household’s head
Secondary school &
above
34 1 (2.9) 1
Primary school 356 83 (23.3) 10.1 (1.35 – 74.5)
Not educated 345 54 (15.7) 6.12 (0.82 – 45.7)
Occupation of
household’s head*
Government
employees
76 4 (5.3) 1
Not working 180 28 (15.6) 3.31 (1.12 – 9.80)
Farmer 453 96 (21.2) 4.84 (1.73 – 13.6)
Fisherman 26 10 (38.5) 11.3 (3.13 – 40.5)
Family size
>5 members 290 49 (16.9) 1 ≤5 members 445 89 (20) 1.23 (0.84 – 1.81)
House wall* Mud 221 26 (11.8) 1 Uncemented bricks 514 112 (21.8) 2.1 (1.32 – 3.30)
Material of
house floor
Cement 120 19 (15.8) 1
Mud 615 119 (19.3) 1.27 (0.75 – 2.16)
Availability of toilet
Yes 284 42 (14.8) 1
No 451 96 (21.3) 1.6 (1.05 – 2.32)
Distance to the nearest water
collection
> 200 meters 295 44 (14.9) 1 ≤ 200 meters 440 146 (18.6) 1.6 (1.05 – 2.30)
Availability of electricity
Yes 379 66 (17.4) 1
No 356 72 (20.2) 1.04 (0.97 – 1.11)
Availability of fridge
Yes 295 44 (14.9) 1
No 440 94 (21.4) 1.6 (1.05 – 2.30)
Availability of TV
Yes 295 44 (14.9) 1
No 440 94 (21.4) 1.6 (1.05 – 2.30)
Availability of
radio
Yes 385 70 (18.2) 1
No 350 68 (19.4) 1.02 (0.95 – 1.09)
Availability of
telephone
Yes 43 8 (18.6) 1
No 692 130 (18.8) 1.0 (0.87 – 1.16)
*Variables confirmed as significant factors associated with malaria using stepwise
forward logistic regression.
Univers
ity of
Mala
ya

84
4.1.3 Assessment of knowledge, attitude and practices (KAP) towards malaria in the
Hadhramout governorate, Yemen
A total number of 735 voluntarily participants enrolled for the survey from 130
housholds in seven villages in two districts, namely Hajer and Al-Raydah-Qusyer
districts. Overall, the survey of the villagers’ knowledge, attitude and practices towards
malaria indicated that although they are all aware of malaria, its mode of transmission,
and its clinical symptoms and severity, their knowledge of and attitude towards malaria
prevention were poor.
For the knowledge about transmission, symptoms, and severity; 100% of the
participants (head of the households) mentioned that malaria is transmitted by mosquito
bites. However, out of these, there were approximately 15% of participants who were
unsure and gave more than one answer. More than half of participants, for example 41%
and 59% recognized fever or fever with shivering as symptoms of malaria, respectively.
Responses about knowledge and attitude towards malaria prevention were poor. Most of
participants (91%) mentioned an ineffective preventive measure. Only 7% and 2% of
study participants knew that sleeping under insecticide-treated mosquito nets (ITNs) or
using indoor residual spraying (IRS) were methods for malaria prevention, respectively.
Furthermore, in all cases, the windows in the houses were kept open at night, 11% of
participants reported using ITNs. When asked what they would do first when they get
malaria, 17% (22/130) mentioned they would go to the clinic or public health center
(Table 4.4).
Univers
ity of
Mala
ya

85
Table 4.4: KAPs of malaria in the rural areas of Hadhramout governorate, Yemen
(n=130)
Characteristics
Number (%)
Knowledge and attitudes
Know malaria 130(100)
Malaria can kill
53 (51)
Mode of transmission mentioned
Mosquito bite 130(100)
Lack of sanitation 4(3)
Swamps
19(15)
Causes of malaria mentioned
Flies 42(32)
Sleeping with infected person in the same bed 106(82)
Mosquito bite 130(100)
Drinking or playing in contaminated water 0 (00)
The presence of sewage
9(7)
Symptoms of malaria mentioned
Fever 53(41)
Fever + shivering
77(59)
Serious for adult or children
Children 118(91)
Equally serious
12(9)
Methods of prevention mentioned
Cleaning the house or environment 118(91)
Sleeping under the mosquito net 9(7)
House spraying with insecticides 3(2)
Smoking house
69(53)
Practices
Using insecticide-treated mosquito nets (ITNs)a 14(11)
House spray with insecticide (IRS)# 130(100)
Not closing house windows 130(100)
Closing house doors 130(100)
Going to clinic when having fever 22(17)
Houses with wood roofs 130(100)
Houses with uncemented bricks wall 93(72)
Houses with mud wall 37 (28)
Keeping uncovered water near houses 130 (100)
*KAPs: Knowledge, Attitude and Practices were conducted on the head of the household
# IRS was done by government before one year of the survey a Each house of the 14 houses had one ITNs
Univers
ity of
Mala
ya

86
4.1.4 Clinical manifestations of individuals positive with malaria
Clinical manifestations of infected individuals with malaria showed more than half of
malaria cases detected by microscopy were asymptomatic, where 52% (72/138) of the
patients had no fever during the survey, whereas, the symptomatic cases presented with
fever (48%; 66/138), shivering (27.5%; 38/138), jaundice (10%; 14/138) and most of
cases with low anaemia (67%; 92/138). A positive association between clinical
symptoms and parasitaemia was observed (χ2=422, p <0.001) (Table 4.5).
Parasitaemia were recorded as low, moderate and high in 52%, 35% and 13% of
malaria cases, respectively (Figure 4.2). The median of parasite densities was 960
asexual parasite/µl with interquartile range of 560 – 2333 asexual parasite/µl. Fifty-two
percent of the persons positive for Plasmodium were asymptomatic with low parasite
density.
Univers
ity of
Mala
ya

87
Table 4.5: Clinical manifestations among humans infected with malaria in Hadhramout
governorate, Yemen
* Fisher exact test was used.
Characteristics
Prevalence N (%)
P value
Presence of fever*
Yes 66 (48) <0.05
No
72 (52)
Presence of shivering*
Yes 38 (27.5) <0.05
No
100 (72.5)
Presence of headache*
Yes 21(15) <0.05
No
117 (85)
Presence of jaundice*
Yes 14(10) <0.05
No
124 (90)
Haemoglobin level
Normal 13 (9) <0.05
Low anaemia 92 (67)
Moderate anaemia
33 (24)
Total 138
Univers
ity of
Mala
ya

88
Figure 4.2: Parasitemia among populations infected with malaria in Hadhramout
governorate, Yemen
13%
35%
52% High
Moderate
Low
Univers
ity of
Mala
ya

89
4.2 MOLECULAR CHARACTERIZATION OF MALARIA IN THE
HADHRAMOUT GOVERNORATE, YEMEN
4.2.1 Malaria parasite identification using nested PCR based on 18SSU rRNA gene
Nested PCR based on the 18SSU rRNA gene was achieved to detect and confirm the
species of Plasmodium, an additional of four other samples which were negative using
microscopy were found positive together with all 138 microscopy positive. Besides that,
three samples which were reported as having mono-infection of P. falciparum via
microscopy, were confirmed as having mixed infections of P. falciparum and P. vivax
(Table 4.6).
Univers
ity of
Mala
ya

90
Table 4.6: Detection of Plasmodium species using nested PCR among populations
infected with malaria in Hadhramout governorate, Yemen
Plasmodium species Microscopy
n (%)
PCR
n (%)
P. falciparum 137 (99.3) 135 (95.07)
P. vivax 1(0.7) 0
Mixed infection (P.f+P.v) 0 3 (2.1)
Non identified by microscopy (P.f) - 4 (2.82)
Total 138 142
Univers
ity of
Mala
ya

91
4.2.2 Prevalence and distribution of mutations in Pfcrt gene at 76, 271, 326, 356 and
371 and Pfmdr1 gene at 86 and 1246 as molecular markers of CQ resistance of
Plasmodium falciparum isolates in Hadhramout governorate, Yemen
The results of the point mutations in Pfcrt and Pfmdr1 genes are shown in Table 4.7. Of
138 Plasmodium falciparum isolates, the prevalence of Pfcrt mutant, wild type, mixed
type alleles were detected in 50.7%, 26.1% and 23.1%, respectively. The prevalence of
the mutant alleles of Pfcrt 271E, Pfcrt 326S and Pfcrt 371I were detected in 58.7%,
54.3% and 44.9% of isolates, respectively. However, all isolates harbored wild type
Pfcrt I356 and Pfmdr1 D1246. The majority of isolates (83.3%) had wild type Pfmdr1
Y86.
The survey showed a significant difference in the distribution of Pfcrt 76 alleles
between Hajer and Al-Raydah–Qusyer districts with the Pfcrt 76T allele being higher in
Al-Raydah–Qusyer (52.7%) (p= 0.006). In contrast, the Pfcrt 371I allele had higher
frequency in Hajer (69.2%) compared to Al-Raydah–Qusyer (39.3%). The distribution
of Pfcrt 371 alleles between the two districts was statistically significant (P= 0.006).
Pfmdr1–86 mutant allele was more prevalent in Hajer than Al Raydah–Qusyer while the
wild type allele was higher in Al-Raydah–Qusyer district (p= 0.03) (Table 4.7).
This survey indicated that there is no significant association between the
distribution of Pfcrt and Pfmdr1 alleles with age and gender. Table 4.8 shows no
significant difference in the frequency of Pfcrt and Pfmdr1 alleles between symptomatic
(n=66) malaria cases and asymptomatic (n=72).
Univers
ity of
Mala
ya

92
Table 4.7: Frequency and distribution of Pfcrt and Pfmdr1 alleles in P. falciparum
isolates from popualtions in Hadhramout governorate, Yemen
Mutation
Codons
Alleles
Districts n (%)
Total
P value
Hajer
Al-Raydah
–Qusyer
Pfcrt 76
Wild
Mutant
Mutant & Wild
3 (11.5)
11 (42.3)
12 (46.2)
33 (29.5)
59 (52.7)
20 (17.9)
36 (26.1)
70 (50.7)
32 (23.1)
0.006*
Pfcrt 271 Wild
Mutant
10 (38.5)
16 (61.5)
47 (42.0)
65 (58.0)
57 (41.3)
81 (58.7)
0.744
Pfcrt 326 Wild
Mutant
9 (34.6)
17 (65.4)
54 (48.2)
58 (51.8)
63 (45.7)
75 (54.3)
0.210
Pfcrt 356 Wild
Mutant
26 (100.0)
0
112 (100.0)
0
138 (100.0)
NA
Pfcrt 371 Wild
Mutant
8 (30.8)
18 (69.2)
68 (60.7)
44 (39.3)
76 (55.1)
62 (44.9)
0.006*
Pfmdr1-86 Wild
Mutant
18 (69.2)
8 (30.8)
97 (86.6)
15 (13.4)
115 (83.3)
23 (16.7)
0.032
Pfmdr-1246# Wild
Mutant
26 (100.0)
0
111 (100.0)
0
137 (100.0)
NA
*Using Pearson Chi-Square
# One sample was missing due to PCR failure for this marker. Univ
ersity
of M
alaya

93
Table 4.8: Frequency and distribution of Pfcrt and Pfmdr1 alleles according to
symptomatology in P. falciparum isolates from populations in
Hadhramout governorate, Yemen
Mutation
Codons
Alleles
Symptoms n (%)
P value Symptomatic*
(n=66)
Asymptomatic
(n=72)
Pfcrt 76 Wild
Mutant
Mutant & Wild
17 (25.8)
36 (55.5)
13 (19.7)
19 (26.4)
34 (47.2)
19 (26.4)
0.596
Pfcrt 271 Wild
Mutant
31 (47.0 (
35 (53.0)
26 )36.1 (
46 (63.9)
0.196
Pfcrt 326 Wild
Mutant
26 (39.4)
40 (60.6)
37 (51.4)
35 (48.6)
0.158
Pfcrt 356 Wild
Mutant
66 (100.0)
0
72 (100.0)
0
NA
Pfcrt 371 Wild
Mutant
37 (56.1)
29 (43.9)
39 (54.2)
33 (45.8)
0.823
Pfmdr 1-86 Wild
Mutant
54 (81.8)
12 (18.2)
61 (84.7)
11 (15.3)
0.647
Pfmdr1-1246# Wild
Mutant
65 (100.0)
0
72 (100.0)
0
NA
*Symptomatic was defined by the presence of fever (>37.5 oC) with or without
shivering and headache. # One sample was missing due to PCR failure for this marker.
Univ
ersity
of M
alaya

94
4.2.3 Prevalence of mutations in Pfdhfr and Pfdhps genes at different codons as
molecular markers of SP resistance of Plasmodium falciparum isolates in
Hadhramout governorate, Yemen
A total of 138 patients with P. falciparum infections based on nested PCR were
included in the analysis of Pfdhfr mutations at codons 51, 59, 108, and 164, as well as
Pfdhps mutations at codons 436, 437 and 540. Of the 138 P. falciparum isolates,
genomic DNAs from 128 and 114 isolates were successfully sequenced for Pfdhfr and
Pdhps genes, respectively.
Mutant alleles are presented in Table 4.9. Pfdhfr mutations were detected in 84%
(107/128) of P. falciparum isolates for codons 51 (I51) and 108 (N108) and in one isolate
for codon 59 (R59). No mutation was identified at codon 164. Pfdhps mutations were
detected in 44.7% (51/114) of P. falciparum isolates for codon 437 (G437). No
significant difference in the distribution of the mutant alleles between Hajer and Al-
Raydah–Qusyer districts was observed.
Subsequently, genotyping analysis based on sequences for Pfdhfr, Pfdhps, and
combined Pfdhfr–Pfdhps genes was conducted. Double (I51C59N108I164) and triple
(I51R59N108I164) mutant genotypes of Pfdhfr were detected in 82.8% (106/128) isolates
and one isolate (0.8%), respectively. For Pfdhps, single mutant genotype (S436G437K540)
was detected in 44.7% (51/114) of the isolates. Genotyping of 107 P. falciparum
isolates for the combined Pfdhfr–Pfdhps genes showed that 5 (4.7%), 46 (43%), 42
(39.3%), and 1 (0.9) isolates had single, double, triple, and quadruple mutant genotypes,
respectively. Although Al-Raydah–Qusyer district had higher prevalence of mutant
Univers
ity of
Mala
ya

95
genotypes than Hajer district, the differences were statistically not significant (Table
4.10).
Univers
ity of
Mala
ya

96
Table 4.9: Prevalence of mutant alleles of Pfdhfr and Pfdhps genes in P. falciparum
isolates from populations in Hadhramout governorate, Yemen
Prevalence n (%)
Mutant alleles* Hajer Al-Raydah–
Qusyer
Total P value
Pfdhfr n = 26 n = 102 n = 128
51I 19 (73.1) 88 (86.3) 107 (84) 0.105
59R 0 (00) 1 (1.0) 1 (0.8) 0.797#
108N 19 (73.1) 88 (86.3) 107 (84) 0.105
164L
0 (00) 0 (00) 0 (00) NA
Pfdhps n = 25 n = 89 n = 114
436A 0 (00) 0 (00) 0 (00) NA
437G 9 (36) 42 (47) 51 (44.7) 0.56
540E 0 (00) 0 (00) 0 (00) NA
n; sample size, NA; not applicable
*Mutant alleles are bold and underlined
#The difference was examined using Fisher exact test.
Univers
ity of
Mala
ya

97
Table 4.10: Prevalence of genotypes of Pfdhfr, Pfdhps and combined Pfdhfr–Pfdhps
genes in P. falciparum isolates from populations in Hadhramout
governorate, Yemen
n; sample size, NA; not applicable
*Mutant alleles are bold and underlined
#The difference was examined using Fisher exact test.
Gene/Genotype* Prevalence n (%)
Hajer Al-Raydah–
Qusyer
Total P value
Pfdhfr
n = 26 n = 102 n = 128
N51C59S108I164 7 (26.9) 14 (13.7) 21 (17) 0.105
I51C59N108I164 19 (73.1) 87 (85.3) 106 (82.8) 0.140
I51R59N108I164
0 (0) 1 (1) 1 (0.8) 0.797#
Pfdhps
n = 25 n = 89 n = 114
S436A437K540 16 (64) 47 (52.8) 63 (55.3)
S436G437K540
9 (36) 42 (47.2) 51 (44.7) 0.56
Pfdhfr–Pfdhps
n = 25 n = 82 n = 107
N51C59S108I164-S436A437K540 5 (20) 8 (9.8) 13 (12.1) 0.170
N51C59S108I164-S436G437K540 1 (4) 4 (4.8) 5 (4.7) 1.000#
I51C59N108I164-S436A437K540 11 (44) 35 (42.7) 46 (43) 0.907
I51C59N108I164-S436G437K540 8 (32) 34 (41.5) 42 (39.3) 0.396
I51R59N108I164-S436G437K540 0 (00) 1 (1.2) 1 (0.9) 1.000#
Univers
ity of
Mala
ya

98
CHAPTER 5: DISCUSSION
5.1 THE EPIDEMIOLOGICAL OF MALARIA IN THE HADHRAMOUT
GOVERNORATE, YEMEN
5.1.1 Prevalence of malaria and identify the risk factors associated with malaria in the
Hadhramout governorate, Yemen
Although Yemen is classified as in the control stage, Hadhramout governorate, located
in the southeast of the country bordering Oman and Saudi Arabia, is considered to be in
the pre-elimination phase and a bilateral collaboration between Yemen and Oman has
been put in place with the aim of making this a malaria-free area (personnel
communication). The purpose of the current study was to evaluate the actual status of
malaria in the Hadhramout community and to investigate associated factors that might
challenge or slow the progress toward malaria elimination.
The overall microscopic prevalence recorded for malaria in the 735 persons
sampled from the seven villages in two districts of Hadhramout governorate was 18.8%.
These high values are inconsistent with a pre-elimination status, and rather placed this
governorate in the control phase. Moreover, the prevalence of malaria in young children
(2-9 years old) exceeded the 10 % level indicative of high to moderate transmission.
This high prevalence could be attributed to several factors including the political
instability in Yemen during the 2011 – 2012 period, which had a direct effect on the
official programs to control and to combat malaria. It was noted that the last IRS was
conducted one year before the field trip. It is also likely that new foci of malaria have
emerged in this area, which was once considered to be of low endemicity. Although the
prevalence of malaria cases is decreasing in Hajer district (NMCP, 2010a), in the
Univers
ity of
Mala
ya

99
traditional malaria endemic area in Hadhramout, an increase of prevalence was recorded
in Al-Raydah-Qusyer districts, areas thought to be of low prevalence. This situation
poses a challenge to control efforts. However, there are many factors related to the
increase of malaria prevalence.
Analysis of the data from the survey presented here have identified some factors
that were associated with increased risk of acquiring malaria. These factors should be
taken into consideration when implementing future malaria control strategies. This
study has highlighted that malaria was more prevalent in adults than in children, who
generally constitute the high-risk group. Multivariate analysis confirmed that people
whose household’s head are fishermen and farmers were at higher risk of being malaria
positive.
Furthermore, observations on the natural work of household that is important to be
taken into account is that household members in Yemen actively contribute to the work
of the head of the household. Such increased risk of malaria linked to occupational
behavior has been noted in other endemic areas such as Malaysia (Trung et al., 2005),
the Philippines (Lansang et al., 1997) and Latin America (Chuquiyauri et al., 2012;
Hiwat et al., 2012). These observations indicated that exposure to the bite of infective
mosquitoes occured outside the home. Consequently, the traditional vector control
interventions (ITNs and IRS) that protect household members would be insufficient, and
control measures should be implemented to reduce mosquito-human contact during
outdoor activities. Occupation-based vector control interventions have been developed
and have shown reduction in malaria cases in Pakistan (Rowland et al., 2004),
Afghanistan (Rowland et al., 1999) and Vietnam (Thang et al., 2009). Such
interventions include topical repellents such as N,N-diethyl-3-methylbenzamide
Univers
ity of
Mala
ya

100
(DEET) (Katz et al., 2008), DEET-based soap (Rowland et al., 2004), plant based
repellant (Hill et al., 2007), long-lasting insecticide-treated hammocks for forest
workers (Sovi et al., 2013) and insecticide-treated personal clothes in refugee areas
(Rowland et al., 1999).
Nonetheless, transmission in and around the house remains significant, as
indicated by the statistical analyses which showed that the type of housing,
unavailability of in-house toilets, and the presence of uncovered water containers close
to the houses are also significant predictors of malaria in Hadhramout. Thus, it will be
important to improve the environment and economic status of the inhabitants if the
government’s efforts to make Hadhramout free of malaria are to be fulfilled. However,
misconceptions, insufficient information, weak or unavailable of health educations
programe about knowledge, attitude and practice against malaria disease especially in
an endemic area will lead to great challenges towards malaria elimination and may
increase malaria transmission in that area.
5.1.2 Assessment of knowledge, attitude and practices towards malaria in the
Hadhramout governorate, Yemen
Communities’ attitudes, beliefs, and knowledge about causes of malaria, well-known
symptom of malaria, proper treatment of malaria, and how to prevent malaria are
helpful and plays an important role in rapid progress toward malaria control and
elimination efforts. In this study, the malaria control strategy in Yemen relies on the
adequate distribution of and use of ITNs, as well as the deployment of the IRS as the
main intervention for vector control. It is, therefore, of concern that in the present study
only 7% of the people expressed the belief that sleeping under ITNs protected them
Univers
ity of
Mala
ya

101
from malaria. This unsatisfactory situation is not unique to this district, as a malaria
indicator survey conducted in Yemen in 2008-2009 revealed that 4.2% of people and
7% of children under 5 years slept under long lasting insecticide-treated net (LLINs)
(WHO, 2011). This represents a major challenge that warrants an urgent action. The fact
that more than half of malaria positive persons identified in the current study were
asymptomatic with low parasite densities suggests that it is likely that these cases would
be missed by passive surveillance and would thus remain as a source of malaria
transmission (Bousema et al., 2004; Okell et al., 2012). Furthermore, ensuring that
people in communities understand and know the proper treatment for malaria disease
may help protect them against the drug resistance development as a result of medication
error such as misuse, overdose or under dosing as reported previously (Abuya et al.,
2007).
Univers
ity of
Mala
ya

102
5.2 MOLECULAR CHARACTERIZATION OF MALARIA IN THE
HADHRAMOUT GOVERNORATE, YEMEN.
5.2.1 Point mutations in Pfcrt gene at 76, 271, 326, 356 and 371 and Pfmdr1 gene at 86
and 1246 as molecular markers of chloroquine resistance of Plasmodium
falciparum isolates in Hadhramout governorate
In the present study, the prevalence of Pfcrt 76T mutation, the highly sensitive marker
for CQ resistance in Hadhramout was 50.7%. This result is lower than the prevalence of
Pfcrt 76T mutation reported from west of Yemen (81–82%) (Al-Mekhlafi et al., 2011b;
Abdul-Ghani et al., 2013) and Lahj in the south of Yemen (98%) (Mubjer et al., 2011).
In contrast, recently published study from Taiz governorate (south of Yemen) revealed
similar findings (50.9%) (Al-Hamidhi et al., 2013). These differences could be
explained by the time period between the implementation of the new drug policy and
screening for CQ resistance markers. The present study was conducted four years after
the launch of the new drug policy while previous reports were carried out either before
or 1–2 years after the initiation of the new antimalarial drug policy. The decline in
prevalence of CQ resistant parasite after the withdrawal of CQ use has been well
documented (Kublin et al., 2003; Laufer et al., 2006). However, the prevalence of Pfcrt
mutations in this survey, is still considered high which means that there is sustained or
an increased CQ usage or genetic adaptations of parasite in this region. The possible
explanation of the sustainability of CQ resistance is the continued unsupervised use of
CQ for the treatment of malaria in Hadhramout governorate, Yemen (Bashrahil et al.,
2010; Bin Ghouth, 2013). Unfortunately, no previous data are available about the
prevalence of Pfcrt 76T gene mutation in Hadhramout which could be used for
comparison to show the trend of CQ resistance in this area over the years. The
prevalence of mutations in Pfcrt at loci 271, 326 and 371 in this survey, was 58.7%,
Univers
ity of
Mala
ya

103
54.3% and 44.9%, respectively. These mutations are more commonly distributed in the
Old World CQ resistant strain and affect the response to CQ in the presence of 76T
mutation (Ibraheem et al., 2014). Findings from this survey necessitate the
implementation of effective control of CQ usage in the Yemeni market especially as CQ
is still necessary for P. vivax infection. However, this has to be done based on accurate
diagnosis.
Although Pfmdr1 plays an important role in modulating levels of antimalarial drug
resistance, CQ resistance has been correlated with the Pfmdr1 86Y (Djimdé et al.,
2001a), while the other point mutations at codons 184, 1034, 1042 and 1246 do not
confer resistance to CQ and are correlated to mefloquine, artesunate, amodiaquine,
halofantrine and quinine resistance (Duraisingh & Cowman, 2005; Danquah et al.,
2010; Gamo, 2014). In the same vein, the Pfmdr1 N86 alleles has been linked to
increased artemether or lumefantrine resistance drugs (Ngo et al., 2003; Lekana-Douki
et al., 2011). In another study, reported lumefantrine usage selected for the Pfmdr1 N86
and 184F alleles (Sisowath et al., 2007; Malmberg et al., 2013). In addition, extensive
use of amodiaquine- artesunate combination therapy selected for the Pfmdr1 N86, 184F
and D1246 alleles (Fröberg et al., 2012). Moreover, many studies reported high
prevalence of wild type Pfmdr1 N86 and D1246 alleles after adopted or increase use of
the artemether/lumefantrine treatment (Zongo et al., 2007; Baliraine & Rosenthal, 2011;
Conrad et al., 2014; Tumwebaze et al., 2015).
However, the mechanism of Pfmdr1 gene mutations in drug resistance is
controversial. In the present study, a prevalence of 16.7% of Pfmdr1 86 mutant type
among P. falciparum isolates was observed, whereas there is no mutation of the Pfmdr1
at codon 1246. This findings are in agreement with recent studies in Yemen and
Univers
ity of
Mala
ya

104
neighboring countries such as Kingdom of Saudi Arabia and Iran (Zakeri et al., 2008;
Dajem et al., 2012; Al-Hamidhi et al., 2013). The low prevalence of Pfmdr1 86Y
mutation has been attributed to almost complete withdrawal of CQ in the community
(Vathsala et al., 2004; Mixson-Hayden et al., 2010). In the same vein, recent studies
suggested that some exert opposite selection by antimalarial drugs on genotypes in
parasite, where the parasite selected the wild type Pfmdr1 N86 and 184F instead of
mutant type 86Y and Y184 due to the change in drug policy to ACTs (Humphreys et al.,
2007; Mungthin et al., 2010; Lekana-Douki et al., 2011; Thomsen et al., 2011).
However, this is not the case in our survey since CQ is still prescribed in Hadhramout
governorate (Bashrahil et al., 2010; Bin Ghouth, 2013).
In the present survey, the association of Pfcrt and Pfmdr1 mutations with gender
and age of participants was not significant. Similar findings have been reported from
Malaysia and Iran (Rastaghi et al., 2008; Atroosh et al., 2012). No significant difference
in the prevalence of Pfcrt and Pfmdr1 mutations between subclinical and clinical
infection of malaria parasites was noted in this survey.
Univers
ity of
Mala
ya

105
5.2.2 Point mutations in Pfdhfr and Pfdhps genes at different codons as molecular
markers of sulfadoxine-pyrimethamine resistance of Plasmodium falciparum
isolates in Hadhramout governorate
High prevalence (84%) of Pfdhfr mutant alleles I51 and N108 was found among P.
falciparum isolates in Hadhramout. These findings were higher than those from
previous reports from western governorates of Yemen (Al-Hamidhi et al., 2013; Abdul-
Ghani et al., 2014). Pfdhfr mutant allele R59 was detected in one isolate of P.
falciparum in this study. However, a study conducted in Lahj governorate reported four
samples harboring this mutant allele in 99 P. falciparum isolates (Mubjer et al., 2011).
Mutation at codon 437 of Pfdhps (G437) was also detected for the first time in 44.7% of
isolates in Hadhramout governorate. Increased frequency of mutant alleles of Pfdhfr
gene and emergence of new mutant alleles of Pfdhps gene in Yemen are early alarming
signals of the possibility of decreasing in the efficacy of SP. Accumulation of mutations
in Pfdhfr gene starts at codon 108 from serine to asparagine, resulting in low levels of
pyrimethamine resistance followed by mutations I51 and R59, as well as at codon L164
point mutation which is related to high level of resistance (Mita et al., 2009; Antony &
Parija et al., 2016). Similarly, sulphadoxine resistance is associated with mutations in
the dhps gene at codons 436, 437, 540, 581 and 613 that starts initially with mutation at
codon 437 from alanine to glycine, followed by E540 and G581, as well as other
mutations (Cowman et al., 1988; Sibley et al., 2001; Gregson & Plowe, 2005; Antony
& Parija et al., 2016). Emergence of resistant parasite to antimalarial drugs involves
many factors, such as economic effects, human hosts, drug pattern interactions,
characteristics of the drug itself, parasites, vectors, and environmental factors (Warhurst
& Williams, 1996; Holland & Kiechle, 2005; Bharti et al., 2007; Vo et al., 2007; Africa,
2013).
Univers
ity of
Mala
ya

106
Drug pressure could have driven the emergence and spreading of the mutant genotypes
in this study. SP had been used as second-line monotherapy for treating uncomplicated
malaria for approximately more than five years before the introduction of ACT drug
policy in 2005 (NMCP, 2006, 2011), which theoretically terminated the use of SP
monotherapy. Moreover, SP is not used for intermittent preventive treatment in
pregnant women in Yemen. However, SP is still available in the private sector with poor
knowledge about the new drug policy among physicians (Bashrahil et al., 2010)
emphasizing the possibility of continued use of SP monotherapy, which may result to
the development of SP resistance (Mohanna et al., 2007). Another possible reason could
be the intensity of transmission; Hadhramout has been classified as low malaria
transmission area and the initiation of pre-elimination phase was suggested (NMCP,
2011). The development and spreading of anti-malarial drug resistance in low
transmission area has been well documented (Roper et al., 2004; Anon et al., 2006;
Menegon et al., 2009). Most patients in this area are usually symptomatic and receive
anti-malarial treatment, which increases the chance of selecting the resistant parasite.
Nevertheless, this classification is not supported by recent studies that have reported
high prevalence of malaria in the community setting and among asymptomatic blood
donors in Hadhramout (Othman et al., 2015).
The present study showed high frequency of double mutant genotype
(I51C59N108I164) among P. falciparum isolates. This genotype has been reported in Sudan
(A-Elbasit et al., 2008; Al-Saai et al., 2009), Saudi Arabia (Al-Farsi et al., 2012),
Angola (Lee et al., 2002), Uganda (Sendagire et al., 2005), Gabon (Bouyou-Akotet et
al., 2010), Iran (Zakeri et al., 2010b) and Afghanistan (Awab et al., 2013). In vitro
studies showed a strong association between the Pfdhfr double mutant (I51 and N108) and
pyrimethamine resistance in Kolkata, West Bengal of India, and Purulia (Das et al.,
Univers
ity of
Mala
ya

107
2012; Das et al., 2013). Another study conducted among Colombian children indicated
that double mutant (I51 and N108) is significantly associated with delayed parasite
clearance (Méndez et al., 2002). By contrast, a study in Sudan reported that the presence
of dhfr double mutant I51 and N108 alone is insufficient to induce in vivo pyrimethamine
resistance (Khalil et al., 2002).
In our study, Pfdhfr triple mutant genotype (I51R59N108) was detected in one P.
falciparum isolate in Hadhramout governorate. This genotype has been strongly
associated with in vitro and in vivo SP resistance (Kublin et al., 2002). Mutant genotype
(I51C59N108I164-S436G437K540), which combined Pfdhfr double mutants (I51, N108) and
Pfdhps single mutant (G437), was highly prevalent among P. falciparum isolates in
Hadhramout governorate. Lower frequencies of this mutant genotype compared with the
present study have been reported from southeastern Iran at 2.7% during 2008–2005
(Zakeri et al., 2010b) and again at 1.8% during 2008–2010 (Wongsrichanalai et al.,
2007), as well as in Tanzania at 0.1% (Shiff et al., 1993). Literature review showed that
this genotype is not widely distributed and has not been correlated yet with the efficacy
of SP either in vitro or in vivo. In this study, one isolate of P. falciparum harbored
quadruple mutant genotype combining the triple dhfr mutant (I51R59N108) and single
dhps mutant G437. Significant association between SP resistance and quadruple mutant
genotype has been reported from in vivo studies conducted in Mali (unpublished data)
and in Ghana only after one year of implementation of intermittent preventive treatment
of malaria (Mockenhaupt et al., 2005; Dicko et al., 2010). Low occurrence of this
genotype has been reported from northern Benin (Ogouyèmi-Hounto et al., 2013),
contrary to the high prevalence reported from southern Benin (Bertin et al., 2011),
Ethiopia, (Hailemeskel et al., 2013) and Senegal (Ndiaye et al., 2013). In contrast, high
prevalence of quadruple and quintuple has been reported from Jalpaiguri district, India
Univers
ity of
Mala
ya

108
which is associated with AS + SP treatment failure for falciparum malaria (Saha et al.,
2012), also shown association of quadruple mutant genotype with recrudescence
infection in Congo, suggested this genotype could be induced a low SP resistance
among falciparum malaria (Ndounga et al., 2007). In addition,in vivo and molecular
study in Somalia shown association of dhfr/dhps quintuple and quadruple mutant
genotype with SP resistance drug and association of dhps double mutant genotype with
treatment failure (Warsame et al., 2015), suggested that need to change national drug
policy in somalia from (AS+SP) to another effective drug of ACTs.
Anti-malarial drug policy has been designed to combine SP with longer half-live
partner drug which clears the remaining parasite and prevent or delay the emergence of
resistance to AS (WHO, 2015a). In Yemen, SP has been the partner drug combined with
AS for treating uncomplicated falciparum malaria (NMCP, 2011) therefore the
emergence of SP resistance will expose the parasite to AS, which has the potential to
contribute to the emergence of ACT resistance in this country. In 2004, three in vivo
studies on clinical efficacy trails showed that SP monotherapy was highly efficacies
(95% - 100%) for treating falciparum malaria in three districts included Harad district in
Hajja governorate, Al-Odein district in Ibb governorate and Mesemeer district in Lahj
governorate (NMCP, 2011). From the time when antimalarial drug policy had shifted
from SP monotherapy as second-line to AS+SP as first-line for treating uncomplicated
malaria, all in vivo efficacy trials have assessed the drug combination (AS+SP) as still
being effective (Adeel et al., 2015). However, the inability of the routine therapeutic
trails to distinguish between the efficacy of AS and its partner drug put SP efficacy
under uncertainty particularly with the high prevalence of the double mutant genotype,
which has good correlation with decreasing SP efficacy, indicating the discontinued
usage of the combination therapy of AS + SP due to insufficient dose of AS to clear
Univers
ity of
Mala
ya

109
parasitemia (Méndez et al., 2002; Das et al., 2013). In contrast, the non-emergence of
quintuple dhfr mutant and triple dhps mutant genotypes that have been associated with
the severe failure of SP (Picot et al., 2009) indicates that SP still provides good
therapeutic response in Hadhramout governorate and Yemen.
Univers
ity of
Mala
ya

110
5.3 LIMITATIONS OF STUDY
There are a few limitations were encountrerd in this study. It should be noted that risk
factors of malaria in this study were identified based on a cross – sectional survey which
suggests more robust design to confirm these predictors such as case-control. In
addition, the RLFP- nested PCR was unable to detect a few samples that have low
parasitaemia or non-detected by microscopy method.
Univers
ity of
Mala
ya

111
CHAPTER 6...CONCLUSIONS AND RECOMMENDATIONS
6.1 CONCLUSIONS
In conclusion, malaria remains an important public health concern in the southeast
region of Yemen (ie., Hadhramout governorate), where there seems to be an upward
shift in malaria prevalence with the appearance of new endemic foci and occupational
high risk groups. There are many barriers and factors that are challenging to the success
of malaria control in Yemen that should be identified in order to develop an effective
control strategy. However, there is a scarcity of data on malaria predictors, KAP and
molecular markers associated with antimalarial drugs resistance, especially CQ and SP
resistance in Yemen. In the same vein, no data is available on malaria risk factors, KAP
and molecular from the Hadhramout governorate, Yemen where this study is being
carried out.
In the current study, several environmental, socioeconomic and behavioural issues
were discovered to be the contributing factors to the high prevalence of malaria in
Hadhramout governorate, Yemen. Furthermore, in present study, high prevalence of
mutations was detected in Pfcrt, Pfmdr1, Pfdhfr and Pfdhps mutant alleles at different
codons. This suggests the sustainability of CQ resistance and emergence of SP
resistance despite post four years of implementing ACTs as a new drug policy in
Yemen. These findings warrant for effective and stricter control of CQ usage and its
open availability in the Yemeni market. Also, high prevalence of Pfdhfr double mutant
genotype (I51C59N108I164) and triple Pfdhfr-Pfdhps mutant genotype (I51C59N108I164-
S436G437K540) in P. falciparum population in Hadhramout, Yemen were also discovered.
These results highlight the risk of developing resistance to SP, the partner drug of AS.
Essentially, the findings of this study, provide baseline information on the KAP, risk
Univers
ity of
Mala
ya

112
factors and molecular markers surveillance of drug-resistant P. falciparum in
Hadhramout governorate, Yemen.
A summary of the main findings:-
1. High prevalence of malaria in two endemic districts in Hadhramout governorate
(18.8%), Yemen, Hajer district (11.8%), and Al-Raydah-Qusyer district (21.8%).
2. High prevalence of malaria has been reported in new foci of malaria named Al-
Rahbah area (21.4%).
3. More than 10% of the prevalence in young children aged 2 to 9 years old is shown
to exceed the level that indicative of high to moderate transmission in
Hadhramout governorate.
4. Several environmental factors, socio-demographic and personal behavioral factors
were discovered in this study to be contributing factors to the high prevalence of
malaria in Hadhramout governorate including: the presence of many ponds and
swamps making this region a suitable breeding ground for Anopheles mosquitoes,
also poor sanitary facilities and political instability in Yemen during the 2011–2012
period. Multivariate analysis using stepwise forward logistic regression confirmed
that the significant associated risk factors in the present study include: living in
uncemented brick wall houses (OR= 2.1, 95% CI: 1.32 – 3.30), or being a
fisherman (OR=11.3, 95% CI: 3.13 – 40.5) or a farmer (OR= 4.84, 95% CI: 1.73 –
13.6), had low level of education (i.e., primary school or not educated) (OR= 10.1,
95% CI: 1.35 – 74.5) or living in houses with the distance of water collection points
less than 200 m (OR= 1.6, 95% CI: 1.05 – 2.32).
5. The predominance of falciparum malaria (99.3%) with high prevalence of
anaemia (67%) highlighted the high risk of developing severe malaria in these
communities.
Univers
ity of
Mala
ya

113
6. Only 7% and 2% of study participants were sleeping under the insecticide-treated
mosquito nets (ITNs) or using indoor residual spraying (IRS) as methods of
malaria prevention.
7. Half of malaria positive cases were asymptomatic with low parasite densities
(52%) suggesting a possible miss by passive surveillance and would thus remain
as a source of malaria transmission in the communities.
8. The high prevalence of mutations at Pfcrt K76T, Q271E, N326S and R371I
alleles in the present study suggested the low susceptibility of falciparum malaria
toward CQ after 4 years of shifting to ACTs in Hadhramout governorate. Low
prevalence of Pfmdr1 N86Y mutation has been attributed to susceptibility of
antimalarial drugs other than chloroquine drug toward P. falciparum.
9. High frequencies of markers associated with sulfodoxine pyrimethamine
resistance in the present study include 84% each of Pfdhfr single mutant allele (I51
and N108), 44.7 % of Pfdhps G437, 82.8% of Pfdhfr double mutant genotype
(I51C59N108I164) and 39.3% of triple Pfdhfr-Pfdhps mutant genotype
(I51C59N108I164-S436G437K540).
6.2 RECOMMENDATIONS
Based on the pertinent findings of the current study, it is recommended that the Ministry
of Health programmes in Yemen rectify the misconceptions about malaria prevention,
diagnosis and treatment as well as knowledge about mosquito control by focusing on
health education initiatives. Holistic efforts to improve the environment and economic
status of the inhabitants should also be part of the integral strategic planning if the
government’s efforts to make Hadhramout governorate free of malaria is to be
successful. Occupation-based vector control interventions should be implemented to
reduce mosquito-human contact during outdoor activities. Another crucial aspect is to
Univers
ity of
Mala
ya

114
enforce effective strategies to ensure accurate implementation of malarial drug
treatment policies according to the national control programme in Yemen.
Active case detection (ACD) should also be implemented, as this would be crucial
to identify and treat the substantial reservoir of asymptomatic persons in the
community. Of utmost importance, ITNs and IRS must become a priority for malaria
control policy as these methods not only act as prevention and treatment measures, but
are effective tools to reduce drug resistance. A further study to investigate antimalarial
drugs resistance in Plasmodium falciparum in Hadhramout governorate and Yemen
using the in vivo efficacy trails is highly recommended as a continuous effort in
monitoring the efficacy of the national anti-malarial drugs policy in Yemen. The
utilisation of Pfcrt, Pfmdr1, Pfdhfr and Pfdhps genes as molecular markers of CQ and
SP resistance are highly recommended in Yemen as malaria is highly endemic in this
country.
Univers
ity of
Mala
ya

115
REFERENCES
Abdul-Ghani, R., Farag, H. F., Allam, A. F., & Shawky, S. M. (2014). Prevailing
Plasmodium falciparum dihydrofolate reductase 108-asparagine in Hodeidah,
Yemen: A questionable sulfadoxine–pyrimethamine partner within the
artemisinin-based combination therapy. Acta Tropica, 132, 39-44.
Abdul-Ghani, R., Farag, H. F., Allam, A. F., Shawky, S. M., & Al-Mekhlafi, A. M.
(2013). Mutant Plasmodium falciparum chloroquine resistance transporter in
Hodeidah, Yemen: Association with parasitologic indices and treatment-seeking
behaviors. Acta Tropica, 128(3), 473-478.
Abdulsalam, M. Q. A., Mohammed, A. K. M., Ahmed, A. A., & Fong, M. Y. (2010).
Clinical situation of endemic malaria in Yemen. Tropical Biomedicine, 27(3),
551-558.
Abuya, T. O., Mutemi, W., Karisa, B., Ochola, S. A., Fegan, G., & Marsh, V. (2007).
Use of over-the-counter malaria medicines in children and adults in three
districts in Kenya: implications for private medicine retailer interventions.
Malaria Journal, 6(1), 57.
Adam, I., Khamis, A. H., & Elbashir, M. I. (2005). Prevalence and risk factors for
anaemia in pregnant women of eastern Sudan. Transaction of the Royal Society
of Tropical Medicine & Hygiene, 99(10), 739-743.
Adams, M., Joshi, S. N., Mbambo, G., Mu, A. Z., Roemmich, S. M., Shrestha, B., et al.
(2015). An ultrasensitive reverse transcription polymerase chain reaction assay
to detect asymptomatic low-density Plasmodium falciparum and Plasmodium
vivax infections in small volume blood samples. Malaria Journal, 14(1), 520.
Adeel, A. A., Elnour, F. A. A., Elmardi, K. A., Abd-Elmajid, M. B., Elhelo, M. M., Ali,
M. S., et al. (2016). High efficacy of artemether-lumefantrine and declining
efficacy of artesunate+ sulfadoxine-pyrimethamine against Plasmodium
falciparum in Sudan (2010–2015): evidence from in vivo and molecular marker
studies. Malaria Journal, 15(1), 1.
Adeel, A. A., Saeed, N. A., Aljasari, A., Almohager, A. M., Galab, M. H., AlMahdi, A.,
et al. (2015). High efficacy of two artemisinin-based combinations: artesunate+
sulfadoxine-pyrimethamine and artemether-lumefantrine for falciparum malaria
in Yemen. Malaria Journal, 14(1), 449.
Adiamah, J., Koram, K., Thomson, M., Lindsay, S., Todd, J., & Greenwood, B. (1993).
Entomological risk factors for severe malaria in a peri-urban area of The
Gambia. Annals of Tropical Medicine and Parasitology, 87(5), 491-500.
A-Elbasit, I. E., Khalil, I., Elbashir, M., Masuadi, E., Bygbjerg, I., Alifrangis, M., &
Giha, H. (2008). High frequency of Plasmodium falciparum CICNI/SGEAA and
Univers
ity of
Mala
ya

116
CVIET haplotypes without association with resistance to sulfadoxine /
pyrimethamine and chloroquine combination in the Daraweesh area, in Sudan.
European Journal of Clinical Microbiology & Infectious Diseases, 27(8), 725-
732.
Afsharpad, M., Zakeri, S., Pirahmadi, S., & Djadid, N. D. (2012). Molecular monitoring
of Plasmodium falciparum resistance to antimalarial drugs after adoption of
sulfadoxine–pyrimethamine plus artesunate as the first line treatment in Iran.
Acta Tropica, 121(1), 13-18.
Ahmad, A., Ngui, R., Muhammad Aidil, R., Lim, Y., & Rohela, M. (2014). Current
status of parasitic infections among Pangkor Island community in Peninsular
Malaysia. Tropical Biomedicine, 31(4), 836-843.
Alifrangis, M., Nag, S., Schousboe, M. L., Ishengoma, D., Lusingu, J., Pota, H., et al.
(2014). Independent origin of Plasmodium falciparum antifolate super-
resistance, Uganda, Tanzania, and Ethiopia. Emerging infectious diseases, 20(8),
1280.
Al-Farsi, H. M., Al-Hashami, Z. S., Dajem, S. M. B., Al-Sheikh, A. A. H., Al-Qahtani,
A., Beja-Pereira, A., et al. (2012). Source of drug resistant Plasmodium
falciparum in a potential malaria elimination site in Saudi Arabia. Infection,
Genetics and Evolution, 12(6), 1253-1259.
Al-Hamidhi, S., Mahdy, M., Al-Hashami, Z., Al-Farsi, H., Al-mekhlafi, A., Idris, M., et
al. (2013). Genetic diversity of Plasmodium falciparum and distribution of drug
resistance haplotypes in Yemen. Malaria Journal, 12, 244.
Al-Maktari, M., Bassiouny, H., Al-Hamd, Z., Assabri, A., El-Massry, A., & Shatat, H.
(2003). Malaria status in Al-Hodeidah Governorate, Yemen: malariometric
parasitic survey & chloroquine resistance P. falciparum local strain. Journal of
the Egyptian Society of Parasitology, 33(2), 361-372.
Al-Mekhlafi, A. M., Al-Mekhlafi, H. M., Mahdy, M. A., Azazy, A. A., & Fong, M. Y.
(2011a). Human malaria in the highlands of Yemen. Annals of Tropical
Medicine and Parasitology, 105(3), 187-195.
Al-Mekhlafi, A. M., Mahdy, M., Al-Mekhlafi, H. M., Azazy, A. A., & Fong, M. Y.
(2011b). High frequency of Plasmodium falciparum chloroquine resistance
marker (Pfcrt T76 mutation) in Yemen: An urgent need to re-examine malaria
drug policy. Parasites & Vectors, 4, 94.
Al-Saai, S., Kheir, A., Abdel-Muhsin, A.-M. A., Al-Ghazali, A., Nwakanma, D.,
Swedberg, G., & Babiker, H. A. (2009). Distinct haplotypes of dhfr and dhps
among Plasmodium falciparum isolates in an area of high level of sulfadoxine–
pyrimethamine (SP) resistance in eastern Sudan. Infection, Genetics and
Evolution, 9(5), 778-783.
Univers
ity of
Mala
ya

117
Al-Shamahy, H., Al-Harazy, A. H., Harmal, N. S., & Al-Kabsi, A. M. (2006). The
prevalence and degree of resistance of Plasmodium falciparum to first-line
antimalarial drugs: an in vitro study from a malaria endemic region in Yemen.
Annals of Saudi Medicine, 27(6), 432-436.
Al-Taiar, A., Assabri, A., Al-Habori, M., Azazy, A., Algabri, A., Alganadi, M., et al.
(2009). Socioeconomic and environmental factors important for acquiring non-
severe malaria in children in Yemen: a case-control study. Transaction of the
Royal Society of Tropical Medicine & Hygiene, 103(1), 72-78.
Al-Taiar, A., Jaffar, S., Assabri, A., Al-Habori, M., Azazy, A., Al-Mahdi, N., et al.
(2006). Severe malaria in children in Yemen: two site observational study.
British Medical Journal, 333(7573), 827.
Al‐Taiar, A., Jaffar, S., Assabri, A., Al‐Habori, M., Azazy, A., Al‐Gabri, A., et al.
(2008). Who develops severe malaria? Impact of access to healthcare, socio‐economic and environmental factors on children in Yemen: a case‐control study.
Tropical Medicine & International Health, 13(6), 762-770.
Al Harthi, S. A. (2007). Detection of drug resistance markers for chloroquine and
pyrimethamine-sulfadoxine in Jazan area, Saudi Arabia using PCR and
restriction digestion. Journal of the Egyptian Society of Parasitology, 37(1), 17-
30.
Alemu, A., Tsegaye, W., Golassa, L., & Abebe, G. (2011). Urban malaria and
associated risk factors in Jimma town, south-west Ethiopia. Malaria Journal,
10(173), 24.
Alkadi, H. O., Al-Maktari, M. T., & Nooman, M. A. (2006). Chloroquine-resistant
Plasmodium falciparum local strain in Taiz Governorate, Republic of Yemen.
Chemotherapy, 52(4), 166-170.
Alker, A. P., Mwapasa, V., & Meshnick, S. R. (2004). Rapid real-time PCR genotyping
of mutations associated with sulfadoxine-pyrimethamine resistance in
Plasmodium falciparum. Antimicrobial Agents and Chemotherapy, 48(8), 2924-
2929.
Alonso, P. L., & Tanner, M. (2013). Public health challenges and prospects for malaria
control and elimination. Nature medicine, 19(2), 150-155.Anon, R., Bosca, M.
M., Sanchiz, V., Tosca, J., Almela, P., Amoros, C., & Benages, A. (2006).
Factors predicting poor prognosis in ischemic colitis. World Journal of
Gastroenterol, 12(30), 4875-4878.
Arasu, G. (1991). Risk behavior in malaria in Malaysia. Southeast Asian Journal of
Tropical Medicine and Public Health, 23, 51-56.
Ariey, F., Witkowski, B., Amaratunga, C., Beghain, J., Langlois, A. C., Khim, N., et al.
(2014). A molecular marker of artemisinin-resistant Plasmodium falciparum
malaria. Nature, 505(7481), 50-55.
Univers
ity of
Mala
ya

118
Ashley, E. A., Dhorda, M., Fairhurst, R. M., Amaratunga, C., Lim, P., Suon, S., et al.
(2014). Spread of artemisinin resistance in Plasmodium falciparum malaria. New
England Journal of Medicine, 371(5), 411-423.
Atieli, H., Menya, D., Githeko, A., & Scott, T. (2009). House design modifications
reduce indoor resting malaria vector densities in rice irrigation scheme area in
western Kenya. Malaria Journal, 8(1), 108.
Atroosh, W. M., Al-Mekhlafi, H. M., Mahdy, M. A., & Surin, J. (2012). The detection
of Pfcrt and Pfmdr1 point mutations as molecular markers of chloroquine drug
resistance, Pahang, Malaysia. Malaria Journal, 11, 251.
Atta, H., & Zamani, G. (2008). The progress of Roll Back Malaria in the Eastern
Mediterranean Region over the past decade. Eastern Mediterranean Health
Journal, 14(suppl), S82-S89.
Awab, G. R., Imwong, M., Pukrittayakamee, S., Alim, F., Hanpithakpong, W., Tarning,
J., et al. (2016). Clinical trials of artesunate plus sulfadoxine-pyrimethamine for
Plasmodium falciparum malaria in Afghanistan: maintained efficacy a decade
after introduction. Malaria Journal, 15(1), 121.
Awab, G. R., Pukrittayakamee, S., Jamornthanyawat, N., Yamin, F., Dondorp, A. M.,
Day, N. P., et al. (2013). Prevalence of antifolate resistance mutations in
Plasmodium falciparum isolates in Afghanistan. Malaria Journal, 12(1), 96.
Ayele, D. G., Zewotir, T. T., & Mwambi, H. G. (2012). Prevalence and risk factors of
malaria in Ethiopia. Malaria Journal, 11(1), 1.
Azazy, A. A., & Raja'a, Y. A. (2003). Malaria and intestinal parasitosis among children
presenting to the paediatric centre in Sana'a, Yemen. Eastern Mediterranean
Health Journal, 9(5-6), 1048-1053.
Azikiwe, C., Ifezulike, C., Siminialayi, I., Amazu, L., Enye, J., & Nwakwunite, O.
(2012). A comparative laboratory diagnosis of malaria: microscopy versus rapid
diagnostic test kits. Asian Pacific Journal of Tropical Biomedicine, 2(4), 307-
310.
Baird, J. K. (2007). Neglect of Plasmodium vivax malaria. Trends In Parasitology,
23(11), 533-539.
Baliraine, F. N., & Rosenthal, P. J. (2011). Prolonged selection of Pfmdr1
polymorphisms after treatment of falciparum malaria with artemether-
lumefantrine in Uganda. Journal of Infectious Diseases, 204(7), 1120-1124.
Baragatti, M., Fournet, F., Henry, M.-C., Assi, S., Ouedraogo, H., Rogier, C., & Salem,
G. (2009). Social and environmental malaria risk factors in urban areas of
Ouagadougou, Burkina Faso. Malaria Journal, 8(1), 13.
Univers
ity of
Mala
ya

119
Barillas‐Mury, C., & Kumar, S. (2005). Plasmodium–mosquito interactions: a tale of
dangerous liaisons. Cellular Microbiology, 7(11), 1539-1545.
Barry, A. E. (2005). Malaria epidemiology: Insights from the genome of the malaria
parasite. Journal of Molecular and Genetic Medicine, 1(2), 76-86.
Basco, L. K., & Ringwald, P. (2000). Molecular epidemiology of malaria in Yaounde,
Cameroon. VI. Sequence variations in the Plasmodium falciparum dihydrofolate
reductase-thymidylate synthase gene and in vitro resistance to pyrimethamine
and cycloguanil. The American Journal of Tropical Medicine and Hygiene,
62(2), 271-276.
Bashrahil, K. A., Bingouth, A. S., & Baruzaig, A. S. (2010). Antimalarial drugs:
availability and mode of prescribing in Mukalla, Yemen. Eastern Mediterranean
Health Journal, 16(2), 146-150.
Bassiouny, H. K., & Al-Maktari, M. T. (2005). Malaria in late pregnancy in Al
Hodeidah Governorate, Yemen. Eastern Mediterranean Health Journal,11(4),
606-617.
Bates, I., Bekoe, V., & Asamoa-Adu, A. (2004). Improving the accuracy of malaria-
related laboratory tests in Ghana. Malaria Journal, 3(1), 38.
Bejon, P., Warimwe, G., Mackintosh, C. L., Mackinnon, M. J., Kinyanjui, S. M.,
Musyoki, J. N., . . . Marsh, K. (2009). Analysis of immunity to febrile malaria in
children that distinguishes immunity from lack of exposure. Infection and
Immunity, 77(5), 1917-1923.
Bell, D., & Peeling, R. W. (2006). Evaluation of rapid diagnostic tests: malaria. Nature
Reviews Microbiology, 4, S34-S38.
Bertin, G., Briand, V., Bonaventure, D., Carrieu, A., Massougbodji, A., Cot, M., &
Deloron, P. (2011). Molecular markers of resistance to sulphadoxine-
pyrimethamine during intermittent preventive treatment of pregnant women in
Benin. Malaria journal, 10(1), 196.
Beshir, K., Sutherland, C. J., Merinopoulos, I., Durrani, N., Leslie, T., Rowland, M., &
Hallett, R. L. (2010). Amodiaquine resistance in Plasmodium falciparum
malaria in Afghanistan is associated with the Pfcrt SVMNT allele at codons 72
to 76. Antimicrobial Agents and Chemotherapy, 54(9), 3714-3716.
Bharti, A. R., Patra, K. P., Chuquiyauri, R., Kosek, M., Gilman, R. H., Llanos-Cuentas,
A., & Vinetz, J. M. (2007). Polymerase chain reaction detection of Plasmodium
vivax and Plasmodium falciparum DNA from stored serum samples:
implications for retrospective diagnosis of malaria. The American Journal of
Tropical Medicine and Hygiene, 77(3), 444-446.
Univers
ity of
Mala
ya

120
Bin Ghouth, A. S. (2013). Availability and prescription practice of anti-malaria drugs in
the private health sector in Yemen. The Journal of Infection in Developing
Countries, 7(5), 404-412.
Baird, J. K., & Hoffman, S. L. (2004). Primaquine therapy for malaria. Clinical
infectious diseases, 39(9), 1336-1345.
Blanford, J. I., Blanford, S., Crane, R. G., Mann, M. E., Paaijmans, K. P., Schreiber, K.
V., & Thomas, M. B. (2013). Implications of temperature variation for malaria
parasite development across Africa. Scientific Reports, 3, 1300.
Bousema, J. T., Gouagna, L. C., Drakeley, C. J., Meutstege, A. M., Okech, B. A., Akim,
I. N., et al. (2004). Plasmodium falciparum gametocyte carriage in
asymptomatic children in western Kenya. Malaria Journal, 3(1), 18.
Bousema, T., Okell, L., Felger, I., & Drakeley, C. (2014). Asymptomatic malaria
infections: detectability, transmissibility and public health relevance. Nature
Reviews Microbiology, 12(12), 833-840.
Bouyou-Akotet, M. K., Ionete-Collard, D. E., Mabika-Manfoumbi, M., Kendjo, E.,
Matsiegui, P.-B., Mavoungou, E., & Kombila, M. (2003). Prevalence of
Plasmodium falciparum infection in pregnant women in Gabon. Malaria
Journal, 2(1), 18.
Bouyou-Akotet, M. K., Mawili-Mboumba, D. P., de Dieu Tchantchou, T., & Kombila,
M. (2010). High prevalence of sulfadoxine/pyrimethamine-resistant alleles of
Plasmodium falciparum isolates in pregnant women at the time of introduction
of intermittent preventive treatment with sulfadoxine/pyrimethamine in Gabon.
Journal of Antimicrobial Chemotherapy, 65(3), 438-441.
Brabin, B., Maxwell, S., Chimsuku, L., Verhoeff, F., Van der Kaay, H., Broadhead, R., .
. . Thomas, A. (1993). A study of the consequences of malarial infection in
pregnant women and their infants. Parassitologia, 35, 9-11.
Breisinger, C., Ecker, O., Thiele, R., & Wiebelt, M. (2012). The impact of the 2008
Hadramout flash flood in Yemen on economic performance and nutrition: A
simulation analysis (No. 1758): Kiel Working Papers.
Bronner, U., Divis, P. C., Färnert, A., & Singh, B. (2009). Swedish traveller with
Plasmodium knowlesi malaria after visiting Malaysian Borneo. Malaria Journal,
8(1), 15.
Butraporn, P., Sornmani, S., & Hungsapruek, T. (1986). Social, behavioural, housing
factors and their interactive effects associated with malaria occurrence in east
Thailand. Southeast Asian Journal of Tropical Medicine and Public Health,
17(3), 386-392.
Univers
ity of
Mala
ya

121
Butterworth, A. S., Skinner-Adams, T. S., Gardiner, D. L., & Trenholme, K. R. (2013).
Plasmodium falciparum gametocytes: with a view to a kill. Parasitology,
140(14), 1718-1734.
Caminade, C., Kovats, S., Rocklov, J., Tompkins, A. M., Morse, A. P., Colón-González,
F. J., et al. (2014). Impact of climate change on global malaria distribution.
Proceedings of the National Academy of Sciences, 111(9), 3286-3291.
Campaign, A.-M. (1991). Clustering of malaria infections within an endemic
population: risk of malaria associated with the type of housing construction. The
American Journal of Tropical Medicine and Hygiene, 45(1), 77-85.
Carneiro, I., Roca-Feltrer, A., Griffin, J. T., Smith, L., Tanner, M., Schellenberg, J. A.,
et al. (2010). Age-patterns of malaria vary with severity, transmission intensity
and seasonality in sub-Saharan Africa: a systematic review and pooled analysis.
PLoS One, 5(2), e8988.
CDC. (2014). Laboratory identification of parasitic diseases of public health concern.
Centers for Disease Control Prevention.
Chavatte, J., Chiron, F., Chabaud, A., & Landau, I. (2007). Probable speciations by"
host-vector'fidelity'": 14 species of Plasmodium from magpies. Parasite (Paris,
France), 14(1), 21-37.
Cheesbrough, M. (2006). District laboratory practice in tropical countries: Cambridge
university press.
Cholewiński, M., Derda, M., & Hadaś, E. (2015). Parasitic diseases in humans
transmitted by vectors. Annals of Parasitology, 61 (3).
Choudhury, D., & Ghosh, S. (1985). Laboratory diagnosis of malaria. The Indian
Journal of Pediatrics, 52(3), 261-267.
Chuquiyauri, R., Paredes, M., Penataro, P., Torres, S., Marin, S., Tenorio, A., et al.
(2012). Socio-demographics and the development of malaria elimination
strategies in the low transmission setting. Acta Tropica, 121(3), 292-302.
Clarke, S. E., Bøgh, C., Brown, R. C., Walraven, G. E., Thomas, C. J., & Lindsay, S.
W. (2002). Risk of malaria attacks in Gambian children is greater away from
malaria vector breeding sites. Transaction of the Royal Society of Tropical
Medicine & Hygiene, 96(5), 499-506.
Clyde, D., & Shute, G. (1957). Resistance of Plasmodium falciparum in Tanganyika to
pyrimethamine administered at weekly intervals. Transaction of the Royal
Society of Tropical Medicine & Hygiene, 51(6), 505-513.
Coleman, R. E., Maneechai, N., Ponlawat, A., Kumpitak, C., Rachapaew, N., Miller, R.
S., & Sattabongkot, J. (2002). Short report: Failure of the OptiMAL rapid
malaria test as a tool for the detection of asymptomatic malaria in an area of
Univers
ity of
Mala
ya

122
Thailand endemic for Plasmodium falciparum and P. vivax. The American
Journal of Tropical Medicine and Hygiene, 67(6), 563-565.
Coleman, R. E., Sattabongkot, J., Promstaporm, S., Maneechai, N., Tippayachai, B.,
Kengluecha, A., et al. (2006). Comparison of PCR and microscopy for the
detection of asymptomatic malaria in a Plasmodium falciparum/vivax endemic
area in Thailand. Malaria Journal, 5(1), 121.
Corbett, Y., Herrera, L., Gonzalez, J., Cubilla, L., Capson, T. L., Coley, P. D., et al.
(2004). A novel DNA-based microfluorimetric method to evaluate antimalarial
drug activity. The American Journal of Tropical Medicine and Hygiene, 70(2),
119-124.
Cotter, C., Sturrock, H. J., Hsiang, M. S., Liu, J., Phillips, A. A., Hwang, J., et al.
(2013). The changing epidemiology of malaria elimination: new strategies for
new challenges. The Lancet, 382(9895), 900-911.
Coutinho-Abreu, I. V., Zhu, K. Y., & Ramalho-Ortigao, M. (2010). Transgenesis and
paratransgenesis to control insect-borne diseases: current status and future
challenges. Parasitology international, 59(1), 1-8.
Cowman, A., & Foote, S. (1990). Chemotherapy and drug resistance in malaria.
International Journal for Parasitology, 20(4), 503-513.
Cowman, A. F., Morry, M. J., Biggs, B. A., Cross, G., & Foote, S. J. (1988). Amino
acid changes linked to pyrimethamine resistance in the dihydrofolate reductase-
thymidylate synthase gene of Plasmodium falciparum. Proceedings of the
National Academy of Sciences, 85(23), 9109-9113.
Cox-Singh, J., Davis, T. M., Lee, K.-S., Shamsul, S. S., Matusop, A., Ratnam, S., et al.
(2008). Plasmodium knowlesi malaria in humans is widely distributed and
potentially life threatening. Clinical Infectious Diseases, 46(2), 165-171.
Craig, M., Snow, R., & Le Sueur, D. (1999). A climate-based distribution model of
malaria transmission in sub-Saharan Africa. Parasitology Today, 15(3), 105-
111.
Crawley, J., Chu, C., Mtove, G., & Nosten, F. (2010). Malaria in children. The Lancet,
375(9724), 1468-1481.
CSO. (2004). Yemen National Census. Sana’a: Central Statistical Organisation,
Republic of Yemen. http :// www .csoyemen .org/ userimages /Image/ book/
yemen_figures_2004.pdf,
Cunha‐Rodrigues, M., Prudêncio, M., Mota, M. M., & Haas, W. (2006). Antimalarial
drugs–host targets (re) visited. Biotechnology Journal, 1(3), 321-332.
Cunningham, J., & Gatton, M. L. (2014). Malaria rapid diagnostic test performance:
results of WHO product testing of malaria RDTs: round 5 (2013).
Univers
ity of
Mala
ya

123
Dajem, S. B., & Al-Qahtani, A. (2010). Analysis of gene mutations involved in
chloroquine resistance in Plasmodium falciparum parasites isolated from
patients in the southwest of Saudi Arabia. Annals of Saudi medicine, 30(3), 187.
Dajem, S. M. B., Al-Farsi, H. M., Al-Hashami, Z. S., Al-Sheikh, A. A. H., Al-Qahtani,
A., & Babiker, H. A. (2012). Distribution of drug resistance genotypes in
Plasmodium falciparum in an area of limited parasite diversity in Saudi Arabia.
The American Journal of Tropical Medicine and Hygiene, 86(5), 782-788.
Dajem, S. M. B., Al-Sheikh, A. A. H., Bohol, M. F., Alhawi, M., Al-Ahdal, M. N., &
Al-Qahtani, A. (2011). Detecting mutations in Pfcrt and Pfmdr1 genes among
Plasmodium falciparum isolates from Saudi Arabia by pyrosequencing.
Parasitology research, 109(2), 291-296.
Danquah, I., Coulibaly, B., Meissner, P., Petruschke, I., Müller, O., & Mockenhaupt, F.
P. (2010). Selection of Pfmdr1 and Pfcrt alleles in amodiaquine treatment failure
in north-western Burkina Faso. Acta tropica, 114(1), 63-66.
Das, S., Chakraborty, S. P., Hati, A., & Roy, S. (2013). Malaria treatment failure with
novel mutation in the Plasmodium falciparum dihydrofolate reductase (pfdhfr)
gene in Kolkata, West Bengal, India. The International Journal of Antimicrobial
Agents, 41(5), 447-451.
Das, S., Chakraborty, S. P., Tripathy, S., Hati, A. K., & Roy, S. (2012). Association
between prevalence of pyrimethamine resistance and double mutation in pfdhfr
gene in West Bengal, India. Asian Pacific Journal of Tropical Disease, 2(1), 31-
35.
Day, K., Hayward, R., & Dyer, M. (1998). The biology of Plasmodium falciparum
transmission stages. Parasitology, 116(S1), S95-S109.
de Monbrison, F., Raynaud, D., Latour-Fondanaiche, C., Staal, A., Favre, S., Kaiser, K.,
. . . Picot, S. (2003). Real-time PCR for chloroquine sensitivity assay and for
Pfmdr1–Pfcrt single nucleotide polymorphisms in Plasmodium falciparum.
Journal of microbiological methods, 54(3), 391-401.
Dhandapani, A., Kumar, S., & Kadarkarai, M. (2011). larvicidal, pupicidal and smoke
toxicity effect of kaempferia galanga to the malarial vector, anopheles stephensi.
The Bio Scan Journal, 6(2), 329-333.
Dicko, A., Sagara, I., Djimdé, A. A., Touré, S. O., Traore, M., Dama, S., et al. (2010).
Molecular markers of resistance to sulphadoxine-pyrimethamine one year after
implementation of intermittent preventive treatment of malaria in infants in
Mali. Malaria journal, 9(1), 9.
Djimdé, A., Doumbo, O. K., Cortese, J. F., Kayentao, K., Doumbo, S., Diourté, Y., et
al. (2001a). A molecular marker for chloroquine-resistant falciparum malaria.
New England Journal of Medicine, 344(4), 257-263.
Univers
ity of
Mala
ya

124
Djimdé, A., Doumbo, O. K., Steketee, R. W., & Plowe, C. V. (2001b). Application of a
molecular marker for surveillance of chloroquine-resistant falciparum malaria.
The Lancet, 358(9285), 890-891.
Do Manh, C., Beebe, N. W., Le Quang, T., Lein, C. T., Van Nguyen, D., Xuan, T. N., et
al. (2010). Vectors and malaria transmission in deforested, rural communities in
north-central Vietnam. Malaria Journal, 9(1), 1.
Dondorp, A. M., Nosten, F., Yi, P., Das, D., Phyo, A. P., Tarning, J., et al. (2009).
Artemisinin resistance in Plasmodium falciparum malaria. New England Journal
of Medicine, 361(5), 455-467.
Duraisingh, M. T., & Cowman, A. F. (2005). Contribution of the Pfmdr1 gene to
antimalarial drug-resistance. Acta Tropica, 94(3), 181-190.
Durand, R., Eslahpazire, J., Jafari, S., Delabre, J.-F., Marmorat-Khuong, A., di Piazza,
J.-P., & Le Bras, J. (2000). Use of molecular beacons to detect an antifolate
resistance-associated mutation in Plasmodium falciparum. Antimicrobial Agents
and Chemotherapy, 44(12), 3461-3464.
Dyer, M. D., Murali, T. M., & Sobral, B. W. (2007). Computational prediction of host-
pathogen protein-protein interactions. Bioinformatics, 23(13), i159-166.
Ecker, A., Lehane, A. M., Clain, J., & Fidock, D. A. (2012). Pfcrt and its role in
antimalarial drug resistance. Trends in parasitology, 28(11), 504-514.
Ekland, E. H., & Fidock, D. A. (2008). In vitro evaluations of antimalarial drugs and
their relevance to clinical outcomes. International Journal for Parasitology,
38(7), 743-747.
El Samani, F. Z., Willett, W., & Ware, J. (1987). Nutritional and socio-demographic
risk indicators of malaria in children under five: a cross-sectional study in a
Sudanese rural community. The Journal of tropical medicine and hygiene, 90(2),
69-78.
Erhart, A., Thang, N. D., Van Ky, P., Tinh, T. T., Van Overmeir, C., Speybroeck, N., et
al. (2005). Epidemiology of forest malaria in central Vietnam: a large scale
cross-sectional survey. Malaria Journal, 4(1), 58.
Eskandarian, A.-A., Keshavarz, H., Basco, L. K., & Mahboudi, F. (2002). Do mutations
in Plasmodium falciparum dihydropteroate synthase and dihydrofolate reductase
confer resistance to sulfadoxine-pyrimethamine in Iran? Transaction of the
Royal Society of Tropical Medicine & Hygiene, 96(1), 96-98.
Rastaghi, A. E., Nateghpou, M., Assmar, M., Razavi, M. R., Kanbara, H., Uemura, H.,
et al. (2008). Detection of K76T Mutation in pfcrt Gene as an Applicable Ge-
netic Marker for Prediction of Chloroquine Resistant falciparum Malaria in
Isolates from an Endemic District of Iran. Iranian Journal of Parasitology, 3(2),
48-56.
Univers
ity of
Mala
ya

125
Färnert, A., Arez, A., Babiker, H., Beck, H., Benito, A., Björkman, A., et al. (2001).
Genotyping of Plasmodium falciparum infections by PCR: a comparative
multicentre study. Transaction of the Royal Society of Tropical Medicine &
Hygiene, 95(2), 225-232.
Ferdig, M. T., Cooper, R. A., Mu, J., Deng, B., Joy, D. A., Su, X. Z., & Wellems, T. E.
(2004). Dissecting the loci of low‐level quinine resistance in malaria
parasites. Molecular microbiology, 52(4), 985-997.
Ferone, R., Burchall, J., & Hitchings, G. (1969). Plasmodium berghei dihydrofolate
reductase isolation, properties, and inhibition by antifolates. Molecular
Pharmacology, 5(1), 49-59.
Fidock, D. A. (2010). Drug discovery: Priming the antimalarial pipeline. Nature,
465(7296), 297-298.
Fivelman, Q. L., Butcher, G. A., Adagu, I. S., Warhurst, D. C., & Pasvol, G. (2002).
Malarone treatment failure and in vitro confirmation of resistance of
Plasmodium falciparum isolate from Lagos, Nigeria. Malaria journal, 1(1), 1.
Fritsche, T. R., & Selvarangan, R. (2011). Medical parasitology. Henry's Clinical
Diagnosis and Management by Laboratory Methods. 22nd ed. Philadelphia, Pa:
Saunders Elsevier.
Fröberg, G., Jörnhagen, L., Morris, U., Shakely, D., Msellem, M. I., Gil, J. P., et al.
(2012). Decreased prevalence of Plasmodium falciparum resistance markers to
amodiaquine despite its wide scale use as ACT partner drug in Zanzibar.
Malaria journal, 11(1), 321.
Fry, M., & Beesley, J. (1991). Mitochondria of mammalian Plasmodium spp.
Parasitology, 102(01), 17-26.
Fry, M., & Pudney, M. (1992). Site of action of the antimalarial
hydroxynaphthoquinone, 2-[trans-4-(4'-chlorophenyl) cyclohexyl]-3-hydroxy-1,
4-naphthoquinone (566C80). Biochemical Pharmacology, 43(7), 1545-1553.
Fungladda, W., & Sornmani, S. (1986). Health behavior, treatment-seeking patterns,
and cost of treatment for patients visiting malaria clinics in western Thailand.
Southeast Asian Journal of Tropical Medicine and Public Health, 17(3), 379-
385.
Fungladda, W., Sornmani, S., Klongkamnuankarn, K., & Hungsapruek, T. (1987).
Sociodemographic and behavioural factors associated with hospital malaria
patients in Kanchanaburi, Thailand. The Journal of tropical medicine and
hygiene, 90(5), 233-237.
Gadalla, N. B., Abdallah, T. M., Atwal, S., Sutherland, C. J., & Adam, I. (2013).
Selection of pfdhfr/pfdhps alleles and declining artesunate/sulphadoxine-
Univers
ity of
Mala
ya

126
pyrimethamine efficacy against Plasmodium falciparum eight years after
deployment in eastern Sudan. Malaria Journal, 12(1), 1.
Gama, B. E., de Oliveira, N. K. A., Zalis, M. G., de Souza, J. M., Santos, F., Daniel-
Ribeiro, C. T., & de Fátima Ferreira-da-Cruz, M. (2009). Chloroquine and
sulphadoxine-pyrimethamine sensitivity of Plasmodium falciparum parasites in
a Brazilian endemic area. Malaria journal, 8(1), 156.
Gamo, F. J. (2014). Antimalarial drug resistance: new treatments options for
Plasmodium. Drug Discovery Today: Technologies, 11, 81-88.
Ghanchi, N. K., Ursing, J., Beg, M. A., Veiga, M. I., Jafri, S., & Martensson, A. (2011).
Prevalence of resistance associated polymorphisms in Plasmodium falciparum
field isolates from southern Pakistan. Malaria Journal, 10(1), 18.
Ghebreyesus, T. A., Haile, M., Witten, K. H., Getachew, A., Yohannes, M., Lindsay, S.
W., & Byass, P. (2000). Household risk factors for malaria among children in
the Ethiopian highlands. Transaction of the Royal Society of Tropical Medicine
& Hygiene, 94(1), 17-21.
Graves, P. M., Richards, F. O., Ngondi, J., Emerson, P. M., Shargie, E. B., Endeshaw,
T., . . . Hailemariam, A. (2009). Individual, household and environmental risk
factors for malaria infection in Amhara, Oromia and SNNP regions of Ethiopia.
Transaction of the Royal Society of Tropical Medicine & Hygiene, 103(12),
1211-1220.
Gregson, A., & Plowe, C. V. (2005). Mechanisms of resistance of malaria parasites to
antifolates. Pharmacological Reviews, 57(1), 117-145.
Gupta, B., Xu, S., Wang, Z., Sun, L., Miao, J., Cui, L., & Yang, Z. (2014). Plasmodium
falciparum multidrug resistance protein 1 (Pfmrp1) gene and its association with
in vitro drug susceptibility of parasite isolates from north-east
Myanmar. Journal of Antimicrobial Chemotherapy, dku125.
Gupta, S., Gunter, J. T., Novak, R. J., & Regens, J. L. (2009). Patterns of Plasmodium
vivax and Plasmodium falciparum malaria underscore importance of data
collection from private health care facilities in India. Malaria Journal, 8(1), 227.
Habtewold, T., Prior, A., Torr, S., & Gibson, G. (2004). Could insecticide‐treated cattle
reduce Afrotropical malaria transmission? Effects of deltamethrin‐treated Zebu
on Anopheles arabiensis behaviour and survival in Ethiopia. Medical and
Veterinary Entomology, 18(4), 408-417.
Hailemeskel, E., Kassa, M., Taddesse, G., Mohammed, H., Woyessa, A., Tasew, G., et
al. (2013). Prevalence of sulfadoxine–pyrimethamine resistance-associated
mutations in dhfr and dhps genes of Plasmodium falciparum three years after SP
withdrawal in Bahir Dar, Northwest Ethiopia. Acta tropica, 128(3), 636-641.
Univers
ity of
Mala
ya

127
Hall, T. (2011). BioEdit: an important software for molecular biology. Green Earth
Research Foundation Bulletin of Biosciences, 2(1), 60-1.
Hamour, S., Melaku, Y., Keus, K., Wambugu, J., Atkin, S., Montgomery, J., et al.
(2005). Malaria in the Nuba Mountains of Sudan: baseline genotypic resistance
and efficacy of the artesunate plus sulfadoxine–pyrimethamine and artesunate
plus amodiaquine combinations. Transaction of the Royal Society of Tropical
Medicine & Hygiene, 99(7), 548-554.
Haque, U., Sunahara, T., Hashizume, M., Shields, T., Yamamoto, T., Haque, R., &
Glass, G. E. (2011). Malaria prevalence, risk factors and spatial distribution in a
hilly forest area of Bangladesh. PLoS One, 6(4), e18908.
Harbach, R. (2011). Genus anopheles Meigen, 1818. Mosquito Taxonomic Inventory.
Book Genus anopheles Meigen, 1818 Mosquito Taxonomic Inventory City.
Hassan, S. E.-D. H., Okoued, S. I., Mudathir, M. A., & Malik, E. M. (2010). Research
Testing the sensitivity and specificity of the fluorescence microscope
(Cyscope®) for malaria diagnosis. Organization, 25, 8-8.
Hastings, I. M. (2007). Molecular markers as indicators of antimalarial drug failure
rates. Tropical Medicine & International Health, 12(11), 1298-1301.
Hay, S. I., Sinka, M. E., Okara, R. M., Kabaria, C. W., Mbithi, P. M., Tago, C. C., et al.
(2010). Developing global maps of the dominant Anopheles vectors of human
malaria. PLoS Med, 7(2), e1000209.
Heidari, A., Dittrich, S., Jelinek, T., Kheirandish, A., Banihashemi, K., & Keshavarz, H.
(2007). Genotypes and in vivo resistance of Plasmodium falciparum isolates in
an endemic region of Iran. Parasitology Research, 100(3), 589-592.
Hill, N., Lenglet, A., Arnez, A. M., & Carneiro, I. (2007). Plant based insect repellent
and insecticide treated bed nets to protect against malaria in areas of early
evening biting vectors: double blind randomised placebo controlled clinical trial
in the Bolivian Amazon. British Medical Journal, 335(7628), 1023.
Hiwat, H., Hardjopawiro, L. S., Takken, W., & Villegas, L. (2012). Novel strategies
lead to pre-elimination of malaria in previously high-risk areas in Suriname,
South America. Malaria Journal, 11(1), 10.
Holland, C. A., & Kiechle, F. L. (2005). Point-of-care molecular diagnostic systems
past, present and future. Current opinion in microbiology, 8(5), 504-509.
Honrado, E. R., & Fungladda, W. (1994). Social and behavioral risk factors related to
malaria in southeast Asian countries. Philipp. Journal of Microbiology and
Infectious Diseases, 23, 76-80.
Univers
ity of
Mala
ya

128
Howard, N., Durrani, N., Sanda, S., Beshir, K., Hallett, R., & Rowland, M. (2011).
Clinical trial of extended-dose chloroquine for treatment of resistant falciparum
malaria among Afghan refugees in Pakistan. Malaria Journal, 10(171.10), 171.
Hubálek, Z., & Halouzka, J. (1999). West Nile fever--a reemerging mosquito-borne
viral disease in Europe. Emerging Infectious Diseases, 5(5), 643.
Hudson, A. (1993). Atovaquone—a novel broad-spectrum anti-infective drug.
Parasitology Today, 9(2), 66-68.
Humphreys, G. S., Merinopoulos, I., Ahmed, J., Whitty, C. J. M., Mutabingwa, T. K.,
Sutherland, C. J., & Hallett, R. L. (2007). Amodiaquine and artemether-
lumefantrine select distinct alleles of the Plasmodium falciparum mdr1 gene in
Tanzanian children treated for uncomplicated malaria. Antimicrobial agents and
chemotherapy, 51(3), 991-997.
Husain, T., & Chaudhary, J. R. (2008). Human health risk assessment due to global
warming–a case study of the Gulf countries. International Journal of
Environmental Research and Public Health, 5(4), 204-212.
Hustache, S., Nacher, M., Djossou, F., & Carme, B. (2007). Malaria risk factors in
Amerindian children in French Guiana. The American Journal of Tropical
Medicine and Hygiene, 76(4), 619-625.
Ibraheem, Z. O., Abd Majid, R., Noor, S. M., Sedik, H. M., & Basir, R. (2014). Role of
different Pfcrt and Pfmdr-1 mutations in conferring resistance to antimalaria
drugs in Plasmodium falciparum. Malaria Research and Treatment, 2014.
Incardona, S., Vong, S., Chiv, L., Lim, P., Nhem, S., Sem, R., et al. (2007). Large-scale
malaria survey in Cambodia: novel insights on species distribution and risk
factors. Malaria Journal, 6(1), 37.
Inchana, W., Kamchoo, K., & Wetasin, K. (2013). Factors Associated with Malaria
Infection in Vibhavadi District, Surat Thani Province, Southern Thailand.
Journal of Tropical Medicine & Parasitology, 36(2), 49-57.
Jelinek, T., Schelbert, P., Löscher, T., & Eichenlaub, D. (1995). Quinine resistant
falciparum malaria acquired in east Africa. Tropical medicine and parasitology:
official organ of Deutsche Tropenmedizinische Gesellschaft and of Deutsche
Gesellschaft fur Technische Zusammenarbeit (GTZ), 46(1), 38-40.
John, D. T., Petri, W. A., Markell, E. K., & Voge, M. (2006). Markell and Voge's
medical parasitology: Elsevier Health Sciences.
Joubert, F., Harrison, C. M., Koegelenberg, R. J., Odendaal, C. J., & de Beer, T. A.
(2009). Discovery: an interactive resource for the rational selection and
comparison of putative drug target proteins in malaria. Malaria Journal, 8(1),
178.
Univers
ity of
Mala
ya

129
Joy, D. A., Gonzalez-Ceron, L., Carlton, J. M., Gueye, A., Fay, M., McCutchan, T. F.,
& Su, X.-z. (2008). Local adaptation and vector-mediated population structure in
Plasmodium vivax malaria. Molecular Biology and Evolution, 25(6), 1245-1252.
Katz, T. M., Miller, J. H., & Hebert, A. A. (2008). Insect repellents: historical
perspectives and new developments. Journal of the American Academy of
Dermatology, 58(5), 865-871.
Kawamoto, F. (1991). Rapid diagnosis of malaria by fluorescence microscopy with light
microscope and interference filter. The Lancet, 337(8735), 200-202.
Khaireh, B. A., Assefa, A., Guessod, H. H., Basco, L. K., Khaireh, M. A., Pascual, A.,
et al. (2013). Population genetics analysis during the elimination process of
Plasmodium falciparum in Djibouti. Malaria Journal, 12(1), 201.
Khalil, I., Alifrangis, M., Rønn, A. M., Gabar, H. A., Jelinek, T., Satti, G. M., &
Bygbjerg, I. C. (2002). Pyrimethamine/sulfadoxine combination in the treatment
of uncomplicated falciparum malaria: relation between dihydropteroate
synthase/dihydrofolate reductase genotypes, sulfadoxine plasma levels, and
treatment outcome. The American Journal of Tropical Medicine and Hygiene,
67(3), 225-229.
Khatoon, L., Baliraine, F. N., Bonizzoni, M., Malik, S. A., & Yan, G. (2009).
Prevalence of antimalarial drug resistance mutations in Plasmodium vivax and P.
falciparum from a malaria-endemic area of Pakistan. The American Journal of
Tropical Medicine and Hygiene, 81(3), 525-528.
Khatoon, L., Baliraine, F. N., Malik, S. A., & Yan, G. (2013). Sequence analysis of
genes associated with resistance to chloroquine and sulphadoxine
pyrimethamine in P. falciparum and P. vivax isolates from the Bannu district of
Pakistan. The Brazilian Journal of Infectious Diseases, 17(5), 596-600.
Khattak, A. A., Venkatesan, M., Jacob, C. G., Artimovich, E. M., Nadeem, M. F.,
Nighat, F., et al. (2013a). A comprehensive survey of polymorphisms conferring
anti-malarial resistance in Plasmodium falciparum across Pakistan. Malaria
Journal, 12(1), 1-9.
Khattak, A. A., Venkatesan, M., Khatoon, L., Ouattara, A., Kenefic, L. J., Nadeem, M.
F., et al. (2013b). Prevalence and patterns of antifolate and chloroquine drug
resistance markers in Plasmodium vivax across Pakistan. Malaria Journal,
12(1), 310.
Kiszewski, A. E., & Teklehaimanot, A. (2004). A review of the clinical and
epidemiologic burdens of epidemic malaria. The American Journal of Tropical
Medicine and Hygiene, 71(2 suppl), 128-135.
Kitvatanachai, S., Janyapoon, K., Rhongbutsri, P., & Thap, L. C. (2003). A survey on
malaria in mobile Cambodians in Aranyaprathet, Sa Kaeo Province, Thailand.
Southeast Asian Journal of Tropical Medicine and Public Health, 34(1), 48-53.
Univers
ity of
Mala
ya

130
Klein, E. (2013). Antimalarial drug resistance: a review of the biology and strategies to
delay emergence and spread. International Journal of Antimicrobial Agents,
41(4), 311-317.
Klinkenberg, E., McCall, P., Wilson, M. D., Amerasinghe, F. P., & Donnelly, M. J.
(2008). Impact of urban agriculture on malaria vectors in Accra, Ghana. Malaria
Journal, 7(1), 151.
Klinkenberg, E., van der Hoek, W., & Amerasinghe, F. P. (2004). A malaria risk
analysis in an irrigated area in Sri Lanka. Acta Tropica, 89(2), 215-225.
Koenderink, J. B., Kavishe, R. A., Rijpma, S. R., & Russel, F. G. (2010). The ABCs of
multidrug resistance in malaria. Trends in parasitology, 26(9), 440-446.
Konradsen, F., Amerasinghe, P., Van Der Hoek, W., Amerasinghe, F., Perera, D., &
Piyaratne, M. (2003). Strong association between house characteristics and
malaria vectors in Sri Lanka. The American Journal of Tropical Medicine and
Hygiene, 68(2), 177-181.
Koram, K., Bennett, S., Adiamah, J., & Greenwood, B. (1995). Socio-economic risk
factors for malaria in a peri-urban area of The Gambia. Transaction of the Royal
Society of Tropical Medicine & Hygiene, 89(2), 146-150.
Korsinczky, M., Chen, N., Kotecka, B., Saul, A., Rieckmann, K., & Cheng, Q. (2000).
Mutations in Plasmodium falciparum Cytochrome b That Are Associated with
Atovaquone Resistance Are Located at a Putative Drug-Binding
Site. Antimicrobial agents and chemotherapy, 44(8), 2100-2108.
Kouznetsov, R. (1976). Distribution of anophelines in the Yemen Arab Republic and its
relation to malaria: World Health Organization.
Kravchenko, V. (1979). Some aspects of the epidemiology of malaria in the People's
Democratic Republic of Yemen. Meditsinskaya Parazitologiya i Parazitarnye
Bolezni, 48(4), 10-13.
Kublin, J. G., Cortese, J. F., Njunju, E. M., Mukadam, R. A. G., Wirima, J. J., Kazembe,
P. N., et al. (2003). Reemergence of chloroquine-sensitive Plasmodium
falciparum malaria after cessation of chloroquine use in Malawi. Journal of
Infectious Diseases, 187(12), 1870-1875.
Kublin, J. G., Dzinjalamala, F. K., Kamwendo, D. D., Malkin, E. M., Cortese, J. F.,
Martino, L. M., et al. (2002). Molecular markers for failure of sulfadoxine-
pyrimethamine and chlorproguanil-dapsone treatment of Plasmodium
falciparum malaria. Journal of Infectious Diseases, 185(3), 380-388.
Kumar, S., & Bandyopadhyay, U. (2005). Free heme toxicity and its detoxification
systems in human. Toxicology Letters, 157(3), 175-188.
Univers
ity of
Mala
ya

131
Kumar, S., Guha, M., Choubey, V., Maity, P., & Bandyopadhyay, U. (2007).
Antimalarial drugs inhibiting hemozoin (β-hematin) formation: A mechanistic
update. Life sciences, 80(9), 813-828.
Lacroix, R., Mukabana, W. R., Gouagna, L. C., & Koella, J. C. (2005). Malaria
infection increases attractiveness of humans to mosquitoes. PLoS Biology, 3(9),
e298.
Lansang, M. A., Belizario, V. Y., Bustos, M. D., Saul, A., & Aguirre, A. (1997). Risk
factors for infection with malaria in a low endemic community in Bataan, the
Philippines. Acta Trop, 63(4), 257-265.
Laufer, M. K. (2009). Monitoring antimalarial drug efficacy: current challenges.
Current Infectious Disease Reports, 11(1), 59-65.
Laufer, M. K., Thesing, P. C., Eddington, N. D., Masonga, R., Dzinjalamala, F. K.,
Takala, S. L., et al. (2006). Return of chloroquine antimalarial efficacy in
Malawi. New England Journal of Medicine, 355(19), 1959-1966.
Launiala, A., & Kulmala, T. (2006). The importance of understanding the local context:
women's perceptions and knowledge concerning malaria in pregnancy in rural
Malawi. Acta Tropica, 98(2), 111-117.
Lee, K.-S., Divis, P. C., Zakaria, S. K., Matusop, A., Julin, R. A., Conway, D. J., et al.
(2011). Plasmodium knowlesi: reservoir hosts and tracking the emergence in
humans and macaques. PLoS Pathogens, 7(4), e1002015.
Lee, S., Kara, U., Koay, E., Lee, M., Lam, S., & Teo, D. (2002). New strategies for the
diagnosis and screening of malaria. International Journal of Hematology, 76(1),
291-293.
Lekana-Douki, J. B., Boutamba, S. D. D., Zatra, R., Edou, S. E. Z., Ekomy, H.,
Bisvigou, U., & Toure-Ndouo, F. S. (2011). Increased prevalence of the
Plasmodium falciparum Pfmdr1 86N genotype among field isolates from
Franceville, Gabon after replacement of chloroquine by artemether–lumefantrine
and artesunate–mefloquine. Infection, Genetics and Evolution, 11(2), 512-517.
LeRoux, M., Lakshmanan, V., & Daily, J. P. (2009). Plasmodium falciparum biology:
analysis of in vitro versus in vivo growth conditions. Trends in Parasitology,
25(10), 474-481.
Liljander, A. (2010). Malaria: Multiclonal infections and protective immunity.
Liu, J. X., Bousema, T., Zelman, B., Gesase, S., Hashim, R., Maxwell, C., et al. (2014).
Is housing quality associated with malaria incidence among young children and
mosquito vector numbers? Evidence from Korogwe, Tanzania. PLoS One, 9(2),
e87358.
Univers
ity of
Mala
ya

132
Longstreth, J., & Wiseman, J. (1989). The potential impact of climate change on
patterns of infectious disease in the United States. The Potential Effects of
Global Climate Change in the United States.
Lopes, D., Rungsihirunrat, K., Nogueira, F., Seugorn, A., Gil, J. P., Do Rosário, V. E.,
& Cravo, P. (2002). Molecular characterisation of drug-resistant Plasmodium
falciparum from Thailand. Malaria Journal, 1(1), 12.
Lumb, V., Das, M. K., Singh, N., Dev, V., Khan, W., & Sharma, Y. D. (2011). Multiple
origins of Plasmodium falciparum dihydropteroate synthetase mutant alleles
associated with sulfadoxine resistance in India. Antimicrobial agents and
chemotherapy, 55(6), 2813-2817.
Luxemburger, C., Ricci, F., Nosten, F., Raimond, D., Bathet, S., & White, N. J. (1997).
The epidemiology of severe malaria in an area of low transmission in Thailand.
Transaction of the Royal Society of Tropical Medicine & Hygiene, 91(3), 256-
262.
Luxemburger, C., Thwai, K. L., White, N., Webster, H., Kyle, D., Maelankirri, L., et al.
(1996). The epidemiology of malaria in a Karen population on the western
border of Thailand. Transaction of the Royal Society of Tropical Medicine &
Hygiene, 90(2), 105-111.
Lvova, M., Zhukova, M., Kiseleva, E., Mayboroda, O., Hensbergen, P., Kizilova, E., et
al. (2016). Hemozoin is a product of heme detoxification in the gut of the most
medically important species of the family Opisthorchiidae. International Journal
for parasitology, 46(3), 147-156.
Lwanga, S. K., & Lemeshow, S., & World Health Organization. (1991). Sample size
determination in health studies: a practical manual:.
Lwin, M., Vijaykumar, S., Lim, G., Theng, Y., & Foo, S. (2014). ‘It's effective but
should I bother?’A study of personal protection measures against Malaria in
urban India. Public Health, 128(7), 654-664.
Mackintosh, C. L., Beeson, J. G., & Marsh, K. (2004). Clinical features and
pathogenesis of severe malaria. Trends in Parasitology, 20(12), 597-603.
Mali, S., Steele, S., Slutsker, L., Arguin, P. M.., & Centers for Disease Control and
Prevention (CDC). (2010). Malaria surveillance--United States, 2008.
Department of Health and Human Services, Centers for Disease Control and
Prevention.
Malmberg, M., Ferreira, P. E., Tarning, J., Ursing, J., Ngasala, B., Björkman, A., et al.
(2013). Plasmodium falciparum drug resistance phenotype as assessed by patient
antimalarial drug levels and its association with Pfmdr1 polymorphisms. Journal
of Infectious Diseases, 207(5), 842-847.
Univers
ity of
Mala
ya

133
Mamser, A. (1989). Report on a visit to the Yemen Arab Republic from 14–2 to 13-3-
1989 (pp. 1-5). WHO/RA/MAL/005. Geneva, EMRO: World Health
Organization.
Marques, A. C. (1987). Human migration and the spread of malaria in Brazil.
Parasitology Today, 3(6), 166-170.
Martens, P., & Hall, L. (2000). Malaria on the move: human population movement and
malaria transmission. Emerging Infectious Diseases, 6(2), 103.
Martínez-Espinosa, F. E., Daniel-Ribeiro, C. T., & Alecrim, W. D. (2004). Malaria
during pregnancy in a reference centre from the Brazilian Amazon: unexpected
increase in the frequency of Plasmodium falciparum infections. Memórias do
Instituto Oswaldo Cruz, 99(1), 19-21.
McMichael, A. J., Woodruff, R. E., & Hales, S. (2006). Climate change and human
health: present and future risks. The Lancet, 367(9513), 859-869.
McMorrow, M. L., Masanja, M. I., Kahigwa, E., Abdulla, S. M., & Kachur, S. P.
(2010). Quality assurance of rapid diagnostic tests for malaria in routine patient
care in rural Tanzania. The American Journal of Tropical Medicine and
Hygiene, 82(1), 151-155.
Mendez, F., Carrasquilla, G., & Muñoz, A. (2000). Risk factors associated with malaria
infection in an urban setting. Transactions of the Royal Society of Tropical
Medicine and Hygiene, 94(4), 367-371.
Menegon, M., Pearce, R. J., Inojosa, W. O., Pisani, V., Abel, P. M., Matondo, A., et al.
(2009). Monitoring for multidrug-resistant Plasmodium falciparum isolates and
analysis of pyrimethamine resistance evolution in Uige province, Angola.
Tropical Medicine & International Health, 14(10), 1251-1257.
Miller, J. M., Shallcross, L., Ringwald, P., Seiber, E., & Organization, W. H. (2005).
Susceptibility of Plasmodium falciparum to antimalarial drugs: Report on global
monitoring 1996-2004.
Miller, L. H., Baruch, D. I., Marsh, K., & Doumbo, O. K. (2002). The pathogenic basis
of malaria. Nature, 415(6872), 673-679.
Miller, L. H., Good, M. F., & Milon, G. (1994). Malaria pathogenesis. Science,
264(5167), 1878-1883.
Milne, L., Kyi, M., Chiodini, P., & Warhurst, D. (1994). Accuracy of routine laboratory
diagnosis of malaria in the United Kingdom. Journal of clinical pathology,
47(8), 740-742.
Minakawa, N., Seda, P., & Yan, G. (2002). Influence of host and larval habitat
distribution on the abundance of African malaria vectors in western Kenya. The
The American Journal of Tropical Medicine and Hygiene, 67(1), 32-38.
Univers
ity of
Mala
ya

134
Mita, T., Tanabe, K., & Kita, K. (2009). Spread and evolution of Plasmodium
falciparum drug resistance. Parasitology international, 58(3), 201-209.
Mitiku, K., Mengistu, G., & Gelaw, B. (2004). The reliability of blood film examination
for malaria at the peripheral health unit. Ethiopian Journal of Health
Development, 17(3), 197-204.
Mixson-Hayden, T., Jain, V., McCollum, A. M., Poe, A., Nagpal, A. C., Dash, A. P., et
al. (2010). Evidence of selective sweeps in genes conferring resistance to
chloroquine and pyrimethamine in Plasmodium falciparum isolates in
India. Antimicrobial agents and chemotherapy, 54(3), 997-1006.
Mockenhaupt, F. P., Teun Bousema, J., Eggelte, T. A., Schreiber, J., Ehrhardt, S.,
Wassilew, N., et al. (2005). Plasmodium falciparum dhfr but not dhps mutations
associated with sulphadoxine‐pyrimethamine treatment failure and gametocyte
carriage in northern Ghana. Tropical Medicine & International Health, 10(9),
901-908.
Mohanna, B., Bin Ghouth, A., & Raja'a, Y. (2007). Malaria signs and infection rate
among asymptomatic schoolchildren in Hajr Valley, Yemen: World Health
Organization.
Moody, A. (2002). Rapid diagnostic tests for malaria parasites. Clinical Microbiology
Reviews, 15(1), 66-78.
Moody, A., & Chiodini, P. (2000). Methods for the detection of blood parasites.
Clinical & Laboratory Haematology, 22(4), 189-201.
Moore, S. J., Min, X., Hill, N., Jones, C., Zaixing, Z., & Cameron, M. M. (2008).
Border malaria in China: knowledge and use of personal protection by minority
populations and implications for malaria control: a questionnaire-based survey.
BMC Public Health, 8(1), 344.
Mu, J., Ferdig, M. T., Feng, X., Joy, D. A., Duan, J., Furuya, T., et al. (2003). Multiple
transporters associated with malaria parasite responses to chloroquine and
quinine. Molecular microbiology, 49(4), 977-989.
Mubjer, R. A., Adeel, A. A., Chance, M. L., & Hassan, A. A. (2011). Molecular
markers of anti-malarial drug resistance in Lahj Governorate, Yemen: baseline
data and implications. Malaria Journal, 10(1), 245.
Mueller, I., Zimmerman, P. A., & Reeder, J. C. (2007). Plasmodium malariae and
Plasmodium ovale–the ‘bashful’malaria parasites. Trends in Parasitology, 23(6),
278-283.
Mugittu, K., Adjuik, M., Snounou, G., Ntoumi, F., Taylor, W., Mshinda, H., et al.
(2006). Molecular genotyping to distinguish between recrudescents and new
infections in treatment trials of Plasmodium falciparum malaria conducted in
Sub-Saharan Africa: adjustment of parasitological outcomes and assessment of
Univers
ity of
Mala
ya

135
genotyping effectiveness. Tropical Medicine and International Health, 11(9),
1350-1359.
Mungthin, M., Suwandittakul, N., Chaijaroenkul, W., Rungsrihirunrat, K.,
Harnyuttanakorn, P., Seugorn, A., & Bangchang, K. N. (2010). The patterns of
mutation and amplification of Plasmodium falciparum Pfcrt and Pfmdr1 genes
in Thailand during the year 1988 to 2003. Parasitology research, 107(3), 539-
545.
Murphy, G. S., Basri, H., Andersen, E. M., Bangs, M. J., Gorden, J., Sorensen, K., et al.
(1993). Vivax malaria resistant to treatment and prophylaxis with chloroquine.
The Lancet, 341(8837), 96-100.
Mutero, C., Blank, H., Konradsen, F., & van der Hoek, W. (2000). Water management
for controlling the breeding of Anopheles mosquitoes in rice irrigation schemes
in Kenya. Acta Tropica, 76(3), 253-263.
Nair, S., Brockman, A., Paiphun, L., Nosten, F., & Anderson, T. J. (2002). Rapid
genotyping of loci involved in antifolate drug resistance in Plasmodium
falciparum by primer extension. International Journal for Parasitology, 32(7),
852-858.
Nair, S., Williams, J. T., Brockman, A., Paiphun, L., Mayxay, M., Newton, P. N., et al.
(2003). A selective sweep driven by pyrimethamine treatment in southeast asian
malaria parasites. Molecular Biology and Evolution, 20(9), 1526-1536.
Nawaz, F., Nsobya, S. L., Kiggundu, M., Joloba, M., & Rosenthal, P. J. (2009).
Selection of parasites with diminished drug susceptibility by amodiaquine-
containing antimalarial regimens in Uganda. Journal of Infectious Diseases
, 200(11), 1650-1657.
Ndiaye, D., Dieye, B., Ndiaye, Y. D., Van Tyne, D., Daniels, R., Bei, A. K., et al.
(2013). Polymorphism in dhfr/dhps genes, parasite density and ex vivo response
to pyrimethamine in Plasmodium falciparum malaria parasites in Thies, Senegal.
International Journal for Parasitology: Drugs and Drug Resistance, 3(1), 135-
142.
Ndounga, M., Tahar, R., Basco, L. K., Casimiro, P. N., Malonga, D. A., & Ntoumi, F.
(2007). Therapeutic efficacy of sulfadoxine–pyrimethamine and the prevalence
of molecular markers of resistance in under 5‐year olds in Brazzaville, Congo.
Tropical Medicine & International Health, 12(10), 1164-1171.
Ngo, T., Duraisingh, M., Reed, M., Hipgrave, D., Biggs, B., & Cowman, A. F. (2003).
Analysis of Pfcrt, Pfmdr1, dhfr, and dhps mutations and drug sensitivities in
Plasmodium falciparum isolates from patients in Vietnam before and after
treatment with artemisinin. The American journal of tropical medicine and
hygiene, 68(3), 350-356.
NMCP. (2002). Malaria Report.Yemen: National Malaria Control Programme.
Univers
ity of
Mala
ya

136
NMCP. (2006). Yemen’s National Malaria Control and Elimination Strategic Plan
2006–2010. Ministry and Public Health and Population,primary Health Care
Sector: National Malaria Control Programme.
NMCP. (2010a). Malaria Report.Hadhramout Region.Yemen. National Malaria Control
Programme.
NMCP. (2010b). National Malaria Control and Elimination Strategic Plan (2011-
2015), Yemen-Sana’a.: National Malaria Control Programme and Ministry of
health and Population.
NMCP. (2011). Yemen’s National Malaria Control and Elimination Strategic Plan
2011–2015. Ministry and Public Health and Population, primary Health Care
Sector: National Malaria Control Programme.
Noedl, H., Wernsdorfer, W. H., Miller, R. S., & Wongsrichanalai, C. (2002). Histidine-
rich protein II: a novel approach to malaria drug sensitivity testing.
Antimicrobial Agents and Chemotherapy, 46(6), 1658-1664.
Noedl, H., Wongsrichanalai, C., & Wernsdorfer, W. H. (2003). Malaria drug-sensitivity
testing: new assays, new perspectives. Trends in Parasitology, 19(4), 175-181.
Noedl, H., Se, Y., Schaecher, K., Smith, B. L., Socheat, D., & Fukuda, M. M. (2008).
Evidence of artemisinin-resistant malaria in western Cambodia. New England
Journal of Medicine, 359(24), 2619-2620.
Noor, A. (2009). Yemen National Malaria Indicator Survey 2008–2009 Submitted to the
World Health Organization Cairo: Eastern Mediterranean Regional Office.
Nosten, F., Ter Kuile, F., Maelankirri, L., Decludt, B., & White, N. (1991). Malaria
during pregnancy in an area of unstable endemicity. Transactions of the Royal
Society of Tropical Medicine and Hygiene, 85(4), 424-429.
Oemijati, S. (1992). Risk behavior in malaria transmission in Indonesia. Southeast
Asian Journal of Tropical Medicine and Public Health, 23, 47-50.
Ogouyèmi-Hounto, A., Ndam, N. T., Fadégnon, G., Azagnandji, C., Bello, M.,
Moussiliou, A., et al. (2013). Low prevalence of the molecular markers of
Plasmodium falciparum resistance to chloroquine and sulphadoxine/
pyrimethamine in asymptomatic children in Northern Benin. Malaria journal,
12(1), 413.
Ohrt, C., O'Meara, W. P., Remich, S., McEvoy, P., Ogutu, B., Mtalib, R., & Odera, J. S.
(2008). Pilot assessment of the sensitivity of the malaria thin film. Malaria
Journal, 7(1), 22.
Okell, L. C., Bousema, T., Griffin, J. T., Ouédraogo, A. L., Ghani, A. C., & Drakeley,
C. J. (2012). Factors determining the occurrence of submicroscopic malaria
infections and their relevance for control. Nature Communications, 3, 1237.
Univers
ity of
Mala
ya

137
Olliaro, P. (2001). Mode of action and mechanisms of resistance for antimalarial drugs.
Pharmacology & Therapeutics, 89(2), 207-219.
Olliaro, P. (2005). Drug resistance hampers our capacity to roll back malaria. Clinical
Infectious Diseases, 41(Supplement 4), S247-S257.
Othman, R., Eldeek, H., Almatary, A., Sayed, A., & Alsakaf, A. (2015). Detection of
malaria in healthy blood donors using PCR in an endemic area in Yemen.
Journal of Advances in Parasitology, 2, 40-47.
Paaijmans, K. P., Read, A. F., & Thomas, M. B. (2009). Understanding the link between
malaria risk and climate. Proceedings of the National Academy of Sciences,
106(33), 13844-13849.
Pagola, S., Stephens, P. W., Bohle, D. S., Kosar, A. D., & Madsen, S. K. (2000). The
structure of malaria pigment β-haematin. Nature, 404(6775), 307-310.
Pandey, A. V., Bisht, H., Babbarwal, V. K., Srivastava, J., Pandey, K. C., & Chauhan,
V. S. (2001). Mechanism of malarial haem detoxification inhibition by
chloroquine. Biochemical Journal, 355(2), 333-338.
Pearce, R. J., Drakeley, C., Chandramohan, D., Mosha, F., & Roper, C. (2003).
Molecular determination of point mutation haplotypes in the dihydrofolate
reductase and dihydropteroate synthase of Plasmodium falciparum in three
districts of northern Tanzania. Antimicrobial agents and chemotherapy, 47(4),
1347-1354.
Peterson, I., Borrell, L. N., El-Sadr, W., & Teklehaimanot, A. (2009). A temporal-
spatial analysis of malaria transmission in Adama, Ethiopia. The The American
Journal of Tropical Medicine and Hygiene, 81(6), 944-949.
Phillips, A., Bassett, P., Szeki, S., Newman, S., & Pasvol, G. (2009). Risk factors for
severe disease in adults with falciparum malaria. Clinical Infectious Diseases,
48(7), 871-878.
Pichainarong, N., & Chaveepojnkamjorn, W. (2004). Malaria infection and life-style
factors among hilltribes along the Thai-Myanmar border area, northern
Thailand. The Southeast Asian Journal of Tropical Medicine and Public Health,
35(4), 834-839.
Picot, S., Olliaro, P., de Monbrison, F., Bienvenu, A.-L., Price, R. N., & Ringwald, P.
(2009). A systematic review and meta-analysis of evidence for correlation
between molecular markers of parasite resistance and treatment outcome in
falciparum malaria. Malaria Journal, 8(1), 89.
Piekarski, G. (2012). Medical parasitology: Springer Science & Business Media.
Pirahmadi, S., Talha, B. A., Nour, B. Y., & Zakeri, S. (2014). Prevalence of mutations
in the antifolates resistance-associated genes (dhfr and dhps) in Plasmodium
Univers
ity of
Mala
ya

138
vivax parasites from Eastern and Central Sudan. Infection, Genetics and
Evolution, 26, 153-159.
Plowe, C. V., Cortese, J. F., Djimde, A., Nwanyanwu, O. C., Watkins, W. M.,
Winstanley, P. A., et al. (1997). Mutations in Plasmodium falciparum
dihydrofolate reductase and dihydropteroate synthase and epidemiologic
patterns of pyrimethamine-sulfadoxine use and resistance. Journal of Infectious
Diseases, 176(6), 1590-1596.
Plowe, C. V., Djimde, A., Bouare, M., Doumbo, O., & Wellems, T. E. (1995).
Pyrimethamine and proguanil resistance-conferring mutations in Plasmodium
falciparum dihydrofolate reductase: polymerase chain reaction methods for
surveillance in Africa. The American Journal of Tropical Medicine and Hygiene,
52(6), 565-568.
Plowe, C. V., Roper, C., Barnwell, J. W., Happi, C. T., Joshi, H. H., Mbacham, W., et
al. (2007). World Antimalarial Resistance Network (WARN) III: molecular
markers for drug resistant malaria. Malaria Journal, 6(1), 121.
Preechapornkul, P., Imwong, M., Chotivanich, K., Pongtavornpinyo, W., Dondorp, A.
M., Day, N. P., et al. (2009). Plasmodium falciparum Pfmdr1 amplification,
mefloquine resistance, and parasite fitness. Antimicrobial agents and
chemotherapy, 53(4), 1509-1515.
Price, R. N., Uhlemann, A. C., Brockman, A., McGready, R., Ashley, E., Phaipun, L., et
al. (2004). Mefloquine resistance in Plasmodium falciparum and increased
Pfmdr1 gene copy number. The Lancet, 364(9432), 438-447.
Prothero, R. M. (1965). Migrants and Malaria. Migrants and Malaria: World Health
Organization.
Pukrittayakamee, S., Supanaranond, W., Looareesuwan, S., Vanijanonta, S., & White,
N. J. (1994). Quinine in severe falciparum malaria: evidence of declining
efficacy in Thailand. Transactions of the Royal Society of Tropical Medicine and
Hygiene, 88(3), 324-327.
Raghavendra, K., Barik, T. K., Reddy, B. N., Sharma, P., & Dash, A. P. (2011). Malaria
vector control: from past to future. Parasitology Research, 108(4), 757-779.
Ranford-Cartwright, L., Johnston, K., Abdel-Muhsin, A., Khan, B., & Babiker, H.
(2002). Critical comparison of molecular genotyping methods for detection of
drug-resistant Plasmodium falciparum. Transactions of the Royal Society of
Tropical Medicine and Hygiene, 96(5), 568-572.
Ranson, H., & Lissenden, N. (2016). Insecticide Resistance in African Anopheles
Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain
Malaria Control. Trends in Parasitology.
Univers
ity of
Mala
ya

139
Rayah, E., Amin, E., ME, A. S., HA, G., & EA, E. A. (2016). Knowledge, practices and
perceptions which affect acquiring malaria in man-made malarious area in
Khartoum State, Sudan. Sudanese Journal of Public Health, 4 (1), 199-209.
Raza, A., Ghanchi, N. K., Khan, M. S., & Beg, M. A. (2013). Prevalence of drug
resistance associated mutations in Plasmodium vivax against sulphadoxine-
pyrimethamine in southern Pakistan. Malaria Journal, 12(1), 261.
Reyburn, H. (2010). New WHO guidelines for the treatment of malaria. BMJ, 340,
c2637.
Rogier, C., Pradines, B., Bogreau, H., Koeck, J.-L., Kamil, M.-A., & Mercereau-
Puijalon, O. (2005). Malaria epidemic and drug resistance, Djibouti. Emerging
Infectious Diseases Journal, 11(2), 317-321.
Roper, C., Pearce, R., Bredenkamp, B., Gumede, J., Drakeley, C., Mosha, F., et al.
(2003). Antifolate antimalarial resistance in southeast Africa: a population-based
analysis. The Lancet, 361(9364), 1174-1181.
Roper, C., Pearce, R., Nair, S., Sharp, B., Nosten, F., & Anderson, T. (2004).
Intercontinental spread of pyrimethamine-resistant malaria. Science, 305(5687),
1124-1124.
Rouhani, M., Zakeri, S., Pirahmadi, S., Raeisi, A., & Djadid, N. D. (2015). High
prevalence of pfdhfr–pfdhps triple mutations associated with anti-malarial drugs
resistance in Plasmodium falciparum isolates seven years after the adoption of
sulfadoxine–pyrimethamine in combination with artesunate as first-line
treatment in Iran. Infection, Genetics and Evolution, 31, 183-189.
Rowland, M., Downey, G., Rab, A., Freeman, T., Mohammad, N., Rehman, H., et al.
(2004). DEET mosquito repellent provides personal protection against malaria: a
household randomized trial in an Afghan refugee camp in Pakistan. Tropical
Medicine & International Health, 9(3), 335-342.
Rowland, M., Durrani, N., Hewitt, S., Mohammed, N., Bouma, M., Carneiro, I., et al.
(1999). Permethrin-treated chaddars and top-sheets: appropriate technology for
protection against malaria in Afghanistan and other complex emergencies.
Transactions of the Royal Society of Tropical Medicine and Hygiene, 93(5),
465-472.
Sabitinelli, G. (2002). Determinants of malaria in WHO European region. The
Contextual Determinants of Malaria, 66-92.
Saha, P., Guha, S. K., Das, S., Mullick, S., Ganguly, S., Biswas, A., et al. (2012).
Comparative efficacy of Artemisinin Combination Therapies (ACTs) in P.
falciparum malaria and polymorphism of PfATPase6, Pfcrt, Pfdhfr and Pfdhps
genes in tea gardens of Jalpaiguri district, India. Antimicrobial agents and
chemotherapy, AAC-05388.
Univers
ity of
Mala
ya

140
Sangster, N., Batterham, P., Chapman, H. D., Duraisingh, M., Le Jambre, L., Shirley,
M., et al. (2002). Resistance to antiparasitic drugs: the role of molecular
diagnosis. International Journal for Parasitology, 32(5), 637-653.
Satoskar, A. R., Simon, G. L., Hotez, P. J., & Tsuji, M. (2009). Medical Parasitology:
Landes Bioscience.
Segurado, A. A. C., SANTI, S. M. D., & Shiroma, M. (1997). In vivo and in vitro
Plasmodium falciparum resistance to chloroquine, amodiaquine and quinine in
the Brazilian Amazon. Revista do Instituto de Medicina Tropical de São Paulo,
39, 85-90.
Sendagire, H., Kaddumukasa, M., Ndagire, D., Aguttu, C., Nassejje, M., Pettersson, M.,
. . . Kironde, F. (2005). Rapid increase in resistance of Plasmodium falciparum
to chloroquine-Fansidar in Uganda and the potential of amodiaquine-Fansidar as
a better alternative. Acta Tropica, 95(3), 172-182.
Senescau, A., Berry, A., Benoit-Vical, F., Landt, O., Fabre, R., Lelièvre, J., et al.
(2005). Use of a locked-nucleic-acid oligomer in the clamped-probe assay for
detection of a minority Pfcrt K76T mutant population of Plasmodium
falciparum. Journal of Clinical Microbiology, 43(7), 3304-3308.
Shah, N. K., Dhillon, G. P., Dash, A. P., Arora, U., Meshnick, S. R., & Valecha, N.
(2011). Antimalarial drug resistance of Plasmodium falciparum in India:
changes over time and space. The Lancet infectious diseases, 11(1), 57-64.
Sharma, J., Dutta, P., Khan, S. A., Soni, M., Dey, D., & Mahanta, J. (2015b). Genetic
polymorphisms associated with sulphadoxine-pyrimethamine drug resistance
among Plasmodium falciparum field isolates in malaria endemic areas of
Assam. Journal of postgraduate medicine, 61(1), 9.
Sharma, R. K., Singh, M. P., Saha, K. B., Bharti, P. K., Jain, V., Singh, P., et al.
(2015a). Socio-economic & household risk factors of malaria in tribal areas of
Madhya Pradesh, central India. The Indian Journal of Medical Research, 141(5),
567.
Shiff, C., Premji, Z., & Minjas, J. (1993). The rapid manual ParaSight®-F test. A new
diagnostic tool for Plasmodium falciparum infection. Transactions of the Royal
Society of Tropical Medicine and Hygiene, 87(6), 646-648.
Sibley, C. H., Hyde, J. E., Sims, P. F., Plowe, C. V., Kublin, J. G., Mberu, E. K., et al.
(2001). Pyrimethamine–sulfadoxine resistance in Plasmodium falciparum: what
next? Trends in Parasitology, 17(12), 570-571.
Sidhu, A. B. S., Uhlemann, A. C., Valderramos, S. G., Valderramos, J. C., Krishna, S.,
& Fidock, D. A. (2006). Decreasing Pfmdr1 copy number in Plasmodium
falciparum malaria heightens susceptibility to mefloquine, lumefantrine,
halofantrine, quinine, and artemisinin. Journal of Infectious Diseases, 194(4),
528-535.
Univers
ity of
Mala
ya

141
Sidhu, A. B. S., Verdier-Pinard, D., & Fidock, D. A. (2002). Chloroquine resistance in
Plasmodium falciparum malaria parasites conferred by Pfcrt mutations. Science,
298(5591), 210-213.
Singh, B., Bobogare, A., Cox-Singh, J., Snounou, G., Abdullah, M. S., & Rahman, H.
A. (1999). A genus- and species-specific nested polymerase chain reaction
malaria detection assay for epidemiologic studies. The American Journal of
Tropical Medicine and Hygiene, 60(4), 687-692.
Singh, B., Sung, L. K., Matusop, A., Radhakrishnan, A., Shamsul, S. S., Cox-Singh, J.,
et al. (2004a). A large focus of naturally acquired Plasmodium knowlesi
infections in human beings. The Lancet, 363(9414), 1017-1024.
Singh, N., Chand, S., Mishra, A., & Nagpal, A. (2004b). Migration malaria associated
with forest economy in central India. Current Science, 87(12), 1396-1399.
Singhanetra-Renard, A. (1986). Population movement, socio-economic behavior and the
transmission of malaria in northern Thailand. Southeast Asian Journal of
Tropical Medicine and Public Health, 17(3), 396-405.
Singhanetra-Renard, A. (1993). Malaria and mobility in Thailand. Social Science &
Medicine Abbreviation, 37(9), 1147-1154.
Sinka, M. E., Bangs, M. J., Manguin, S., Coetzee, M., Mbogo, C. M., Hemingway, J., et
al. (2010). The dominant Anopheles vectors of human malaria in Africa, Europe
and the Middle East: occurrence data, distribution maps and bionomic précis.
Parasites & Vectors, 3(1), 1.
Sintasath, D. M., Ghebremeskel, T., Lynch, M., Kleinau, E., Bretas, G., Shililu, J., et al.
(2005). Malaria prevalence and associated risk factors in Eritrea. The American
Journal of Tropical Medicine and Hygiene, 72(6), 682-687.
Sisowath, C., Ferreira, P. E., Bustamante, L. Y., Dahlström, S., Mårtensson, A.,
Björkman, A., et al. (2007). The role of Pfmdr1 in Plasmodium falciparum
tolerance to artemether‐lumefantrine in Africa. Tropical Medicine &
International Health, 12(6), 736-742.
Snounou, G. (1996). Detection and identification of the four malaria parasite species
infecting humans by PCR amplification Species Diagnostics Protocols (pp. 263-
291): Springer.
Snow, R., Amratia, P., Zamani, G., Mundia, C. W., Noor, A. M., Memish, Z. A., et al.
(2013). The malaria transition on the Arabian Peninsula: progress toward a
malaria-free region between 1960–2010. Advances in Parasitology, 82, 205.
Soucy, A. (2011). Mixed migration from the Horn of Africa to Yemen: Protection risks
and challenges. Danish Refugee Council of Horn of Africa and Yemen, Kenya
Univers
ity of
Mala
ya

142
Sovi, A., Azondékon, R., Aïkpon, R. Y., Govoétchan, R., Tokponnon, F., Agossa, F., et
al. (2013). Impact of operational effectiveness of long-lasting insecticidal nets
(LLINs) on malaria transmission in pyrethroid-resistant areas. Parasites &
Vectors, 6(1), 319.
Srivastava, I. K., Rottenberg, H., & Vaidya, A. B. (1997). Atovaquone, a broad
spectrum antiparasitic drug, collapses mitochondrial membrane potential in a
malarial parasite. Journal of Biological Chemistry, 272(7), 3961-3966.
Staedke, S. G., Nottingham, E. W., Cox, J., Kamya, M. R., Rosenthal, P. J., & Dorsey,
G. (2003). Short report: proximity to mosquito breeding sites as a risk factor for
clinical malaria episodes in an urban cohort of Ugandan children. The American
Journal of Tropical Medicine and Hygiene, 69(3), 244-246.
Stepniewska, K., Taylor, W. R., Mayxay, M., Price, R., Smithuis, F., Guthmann, J.-P.,
et al. (2004). In vivo assessment of drug efficacy against Plasmodium
falciparum malaria: duration of follow-up. Antimicrobial Agents and
Chemotherapy, 48(11), 4271-4280.
Su, X.-z., Kirkman, L. A., Fujioka, H., & Wellems, T. E. (1997). Complex
polymorphisms in an∼ 330 kDa protein are linked to chloroquine-resistant P.
falciparum in Southeast Asia and Africa. Cell, 91(5), 593-603.
Subbarao, S., & Sharma, V. (1997). Anopheline species complexes & malaria control.
The Indian Journal of Medical Research, 106, 164-173.
Sullivan, D., & Krishna, S. (2006). Malaria: Drugs, disease and post-genomic biology
(Vol. 295): Springer Science & Business Media.
Sullivan, D. J., Gluzman, I. Y., Russell, D. G., & Goldberg, D. E. (1996). On the
molecular mechanism of chloroquine's antimalarial action. Proceedings of the
National Academy of Sciences, 93(21), 11865-11870.
Takala-Harrison, S., Jacob, C. G., Arze, C., Cummings, M. P., Silva, J. C., Dondorp, A.
M., et al. (2015). Independent emergence of artemisinin resistance mutations
among Plasmodium falciparum in Southeast Asia. Journal of Infectious
Diseases, 211(5), 670-679.
Tangpukdee, N., Duangdee, C., Wilairatana, P., & Krudsood, S. (2009). Malaria
diagnosis: a brief review. The Korean Journal of Parasitology, 47(2), 93-102.
Tatem, A. J., Smith, D. L., Gething, P. W., Kabaria, C. W., Snow, R. W., & Hay, S. I.
(2010). Ranking of elimination feasibility between malaria-endemic countries.
The Lancet, 376(9752), 1579-1591.
Teklehaimanot, H. D., Lipsitch, M., Teklehaimanot, A., & Schwartz, J. (2004).
Weather-based prediction of Plasmodium falciparum malaria in epidemic-prone
regions of Ethiopia I. Patterns of lagged weather effects reflect biological
mechanisms. Malaria Journal, 3(1), 41.
Univers
ity of
Mala
ya

143
Terkuile, F., White, N., Holloway, P., Pasvol, G., & Krishna, S. (1993). Plasmodium
falciparum: in vitro studies of the pharmacodynamic properties of drugs used for
the treatment of severe malaria. Experimental Parasitology, 76(1), 85-95.
Thang, N. D., Erhart, A., Speybroeck, N., Xa, N. X., Thanh, N. N., Ky, P. V., et al.
(2009). Long-Lasting Insecticidal Hammocks for controlling forest malaria: a
community-based trial in a rural area of central Vietnam. PLoS One, 4(10),
e7369.
Thomas, D., Tazerouni, H., Sundararaj, K. G. S., & Cooper, J. C. (2016). Therapeutic
failure of primaquine and need for new medicines in radical cure of Plasmodium
vivax. Acta tropica, 160, 35-38.
Tinto, H., Ouedraogo, J. B., Zongo, I., Van Overmeir, C., Van Marck, E., Guiguemde,
T. R., & D’ALESSANDRO, U. (2007). Sulfadoxine–pyrimethamine efficacy
and selection of Plasmodium falciparum DHFR mutations in Burkina Faso
before its introduction as intermittent preventive treatment for pregnant women.
The American journal of tropical medicine and hygiene, 76(4), 608-613.
Tipmontree, R., Fungladda, W., Kaewkungwal, J., Tempongko, M., & Schelp, F.-P.
(2009). Migrants and malaria risk factors: a study of the Thai-Myanmar border.
Southeast Asian Journal of Tropical Medicine and Public Health, 40(6), 1148.
Trampuz, A., Jereb, M., Muzlovic, I., & Prabhu, R. M. (2003). Clinical review: Severe
malaria. Critical Care-London, 7(4), 315-323.
Trape, J.-F. (1985). Rapid evaluation of malaria parasite density and standardization of
thick smear examination for epidemiological investigations. Transactions of the
Royal Society of Tropical Medicine and Hygiene, 79(2), 181-184.
Trung, H. D., Bortel, W. V., Sochantha, T., Keokenchanh, K., Briet, O. J., &
Coosemans, M. (2005). Behavioural heterogeneity of Anopheles species in
ecologically different localities in Southeast Asia: a challenge for vector control.
Tropical Medicine & International Health, 10(3), 251-262.
Tumwebaze, P., Conrad, M. D., Walakira, A., LeClair, N., Byaruhanga, O., Nakazibwe,
C., et al. (2015). Impact of antimalarial treatment and chemoprevention on the
drug sensitivity of malaria parasites isolated from ugandan children.
Antimicrobial agents and chemotherapy, 59(6), 3018-3030.
Van Der Hoek, W., Konradsen, F., Amerasinghe, P. H., Perera, D., Piyaratne, M., &
Amerasinghe, F. P. (2003). Towards a risk map of malaria for Sri Lanka: the
importance of house location relative to vector breeding sites. International
Journal of Epidemiology, 32(2), 280-285.
van der Hoek, W., Konradsen, F., Dijkstra, D., Amerasinghe, P., & Amerasinghe, F.
(1998). Risk factors for malaria: a microepidemiological study in a village in Sri
Lanka. Transactions of the Royal Society of Tropical Medicine and Hygiene,
92(3), 265-269.
Univers
ity of
Mala
ya

144
van Eijk, A. M., & Terlouw, D. J. (2011). Azithromycin for treating uncomplicated
malaria. The Cochrane Database of Systematic Reviews, 2(2).
Vathsala, P. G., Pramanik, A., Dhanasekaran, S., Devi, C. U., Pillai, C. R., Subbarao, S.
K., et al. (2004). Widespread occurrence of the Plasmodium falciparum
chloroquine resistance transporter (Pfcrt) gene haplotype SVMNT in P.
falciparum malaria in India. The American journal of tropical medicine and
hygiene, 70(3), 256-259.
Vo, T. K. D., Bigot, P., Gazin, P., Sinou, V., De Pina, J. J., Huynh, D. C., et al. (2007).
Evaluation of a real-time PCR assay for malaria diagnosis in patients from
Vietnam and in returned travellers. Transactions of the Royal Society of Tropical
Medicine and Hygiene, 101(5), 422-428.
Waheed, A. A., Ghanchi, N. K., Rehman, K. A., Raza, A., Mahmood, S. F., & Beg, M.
A. (2015). Vivax malaria and chloroquine resistance: a neglected disease as an
emerging threat. Malaria Journal, 14(1), 146.
Waitumbi, J. N., Opollo, M. O., Muga, R. O., Misore, A. O., & Stoute, J. A. (2000).
Red cell surface changes and erythrophagocytosis in children with severe
Plasmodium falciparum anemia. Blood, 95(4), 1481-1486.
Warhurst, D., & Williams, J. (1996). ACP Broadsheet no 148. July 1996. Laboratory
diagnosis of malaria. Journal of clinical pathology, 49(7), 533.
Warrell, D. A., & Gilles, H. M. (2002). Essential malariology (No. Ed. 4). Arnold.
Warsame, M., Hassan, A. M., Barrette, A., Jibril, A. M., Elmi, H. H., Arale, A. M., et al.
(2015). Treatment of uncomplicated malaria with artesunate plus sulfadoxine–
pyrimethamine is failing in Somalia: evidence from therapeutic efficacy studies
and Pfdhfr and Pfdhps mutant alleles. Tropical Medicine & International
Health, 20(4), 510-517.
White, N. (2008a). Plasmodium knowlesi: the fifth human malaria parasite. Clinical
Infectious Diseases, 46(2), 172-173.
White, N., Pukrittayakamee, S., Hien, T., Faiz, M., Mokuolu, O., & Dondorp, A.
(2014). Malaria (vol 383, pg 723, 2014). Lancet, 383(9918), 696-696.
White, N. J. (1996). The treatment of malaria. New England Journal of Medicine,
335(11), 800-806.
White, N. J. (2002). The assessment of antimalarial drug efficacy. Trends in
Parasitology, 18(10), 458-464.
White, N. J. (2008b). Qinghaosu (artemisinin): the price of success. Science, 320(5874),
330-334.
Univers
ity of
Mala
ya

145
WHO. (1996). Assessment of therapeutic efficacy of antimalarial drugs: for
uncomplicated falciparum malaria in areas with intense transmission. Geneva.
WHO. (2001). World Malaria Report 2001. Geneva.
WHO. (2002). Monitoring antimalarial drug resistance: report of a WHO consultation,
Geneva.
WHO. (2003). Assessment and monitoring of antimalarial drug efficacy for the
treatment of uncomplicated falciparum malaria. Geneva.
WHO. (2008a). World malaria report 2008. Geneva.
WHO. (2008b). Methods and techniques for clinical trials on antimalarial drug efficacy:
genotyping to identify parasite populations: informal consultation organized by
the Medicines for Malaria Venture and cosponsored by the World Health
Organization, 29-31 May 2007, Amsterdam, The Netherlands.
WHO. (2009). Malaria microscopy quality assurance manual. Geneva.
WHO. (2010). Global report on antimalarial drug efficacy and drug resistance: 2000-
2010. Geneva.
WHO. (2011). World malaria report 2011. Geneva.
WHO. (2012). World malaria report 2012. Geneva.
WHO. (2013). World Malaria Report 2013. Geneva.
WHO. (2014a). Status report on artemisinin resistance. Geneva.
WHO. (2014b). World malaria report 2014. Geneva.
WHO. (2015a). Guidelines for the treatment of malaria. ( 3rd ed.). Geneva.
WHO. (2015b). Malaria rapid diagnostic test performance: results of WHO product
testing of malaria RDTs: round 6 (2014-2015). Geneva.
Winskill, P., Rowland, M., Mtove, G., Malima, R. C., & Kirby, M. J. (2011). Malaria
risk factors in north-east Tanzania. Malaria Journal, 10(98), 10.1186.
Wongsrichanalai, C., Barcus, M. J., Muth, S., Sutamihardja, A., & Wernsdorfer, W. H.
(2007). A review of malaria diagnostic tools: microscopy and rapid diagnostic
test (RDT). The American Journal of Tropical Medicine and Hygiene, 77(6
Suppl), 119-127.
Univers
ity of
Mala
ya

146
Worrall, E., Basu, S., & Hanson, K. (2003). The relationship between socio-economic
status and malaria: a review of the literature. Background paper for Ensuring
that malaria control interventions reach the poor, London, 56.
Yahia, A. R. (2005). Estimation of malaria burden in Hadramout mesoendemic coastal
districts, Yemen. WHO Regional Office for the Eastern Mediterranean.
Yapabandara, A., Curtis, C., Wickramasinghe, M., & Fernando, W. (2001). Control of
malaria vectors with the insect growth regulator pyriproxyfen in a gem-mining
area in Sri Lanka. Acta Tropica, 80(3), 265-276.
Yé, Y., Hoshen, M., Louis, V., Séraphin, S., Traoré, I., & Sauerborn, R. (2006).
Housing conditions and Plasmodium falciparum infection: protective effect of
iron-sheet roofed houses. Malaria Journal, 5(1), 8.
Yewhalaw, D., Legesse, W., Van Bortel, W., Gebre-Selassie, S., Kloos, H., Duchateau,
L., & Speybroeck, N. (2009). Malaria and water resource development: the case
of Gilgel-Gibe hydroelectric dam in Ethiopia. Malaria Journal, 8(1), 21.
Yuvaniyama, J., Chitnumsub, P., Kamchonwongpaisan, S., Vanichtanankul, J.,
Sirawaraporn, W., Taylor, P., et al. (2003). Insights into antifolate resistance
from malarial DHFR-TS structures. Nature Structural & Molecular Biology,
10(5), 357-365.
Zahar, A. (1974). Review of the ecology of malaria vectors in the WHO Eastern
Mediterranean Region. Bulletin of the World Health Organization, 50(5), 427.
Zakeri, S., Gil, J. P., Bereckzy, S., Djadid, N. D., & Bjorkman, A. (2003). High
prevalence of double Plasmodium falciparum dhfr mutations at codons 108 and
59 in the Sistan-Baluchistan province, Iran. Journal of Infectious Diseases,
187(11), 1828-1829.
Zakeri, S., Afsharpad, M., Raeisi, A., & Djadid, N. D. (2007). Prevalence of mutations
associated with antimalarial drugs in Plasmodium falciparum isolates prior to
the introduction of sulphadoxine-pyrimethamine as first-line treatment in Iran.
Malaria Journal, 6(1), 148.
Zakeri, S., Afsharpad, M., Kazemzadeh, T., Mehdizadeh, K., Shabani, A., & Djadid, N.
D. (2008). Association of Pfcrt but not Pfmdr1 alleles with chloroquine
resistance in Iranian isolates of Plasmodium falciparum. The American Journal
of Tropical Medicine and Hygiene, 78(4), 633-640.
Zakeri, S., Afsharpad, M., Ghasemi, F., Raeisi, A., Safi, N., Butt, W., et al. (2010a).
Molecular surveillance of Plasmodium vivax dhfr and dhps mutations in isolates
from Afghanistan. Malaria Journal, 9(1), 1-8.
Zakeri, S., Farahani, M. S., Afsharpad, M., Salehi, M., Raeisi, A., & Djadid, N. D.
(2010b). High prevalence of the 437G mutation associated with sulfadoxine
resistance among Plasmodium falciparum clinical isolates from Iran, three years
Univers
ity of
Mala
ya

147
after the introduction of sulfadoxine–pyrimethamine. International Journal of
Infectious Diseases, 14, e123-e128.
Zakeri, S., Afsharpad, M., Ghasemi, F., Raeisi, A., Kakar, Q., Atta, H., & Djadid, N. D.
(2011). Plasmodium vivax: prevalence of mutations associated with
sulfadoxine–pyrimethamine resistance in Plasmodium vivax clinical isolates
from Pakistan. Experimental Parasitology, 127(1), 167-172.
Zakai, H. A., Khan, W., & Asma, U. (2013). Prevalence of mutation and phenotypic
expression associated with sulfadoxine-pyrimethamine resistance in Plasmodium
falciparum and Plasmodium vivax. Folia Parasitologica, 60(4), 372.
Zhang, Y., & Meshnick, S. R. (1991). Inhibition of Plasmodium falciparum
dihydropteroate synthetase and growth in vitro by sulfa drugs. Antimicrobial
Agents and Chemotherapy, 35(2), 267-271.
Zongo, I., Dorsey, G., Rouamba, N., Tinto, H., Dokomajilar, C., Guiguemde, R. T., et
al. (2007). Artemether-lumefantrine versus amodiaquine plus sulfadoxine-
pyrimethamine for uncomplicated falciparum malaria in Burkina Faso: a
randomised non-inferiority trial. The Lancet, 369(9560), 491-498.
Univers
ity of
Mala
ya