identification of thioredoxin h-reducible disulphides in proteomes by differential labelling of...
TRANSCRIPT

REGULAR ARTICLE
Identification of thioredoxin h-reducible disulphides in
proteomes by differential labelling of cysteines: Insight
into recognition and regulation of proteins in barley
seeds by thioredoxin h
Kenji Maeda, Christine Finnie and Birte Svensson
Department of Chemistry, Carlsberg Laboratory, Copenhagen, Denmark
Using thiol-specific fluorescence labelling, over 30 putative target proteins of thioredoxin h withdiverse structures and functions have been identified in seeds of barley and other plants. To gaininsight at the structural level into the specificity of target protein reduction by thioredoxin h,thioredoxin h-reducible disulphide bonds in individual target proteins are identified using anovel strategy based on differential alkylation of cysteine thiol groups by iodoacetamide and4-vinylpyridine. This method enables the accessible cysteine side chains in the thiol form (carba-midomethylated) to be distinguished from those inaccessible or disulphide bound form (pyr-idylethylated) according to the mass difference in the peptide mass maps obtained by matrix-assistend laser desorption/ionisation-time of flight mass spectrometry. Using this approach, invitro reduction of disulphides in recombinant barley a-amylase/subtilisin inhibitor (BASI) bybarley thioredoxin h isoform 1 was analysed. Furthermore, the method was coupled with two-dimensional electrophoresis for convenient thioredoxin h-reducible disulphide identificationin barley seed extracts without the need for protein purification or production of recombinantproteins. Mass shifts of 15 peptides, induced by treatment with thioredoxin h and differentialalkylation, identified specific reduction of nine disulphides in BASI, four a-amylase/trypsin in-hibitors and a protein of unknown function. Two specific disulphides, located structurally closeto the a-amylase binding surfaces of BASI and a-amylase inhibitor BMAI-1 were demonstratedto be reduced to a particularly high extent. For the first time, specificity of thioredoxin h for par-ticular disulphide bonds is demonstrated, providing a basis to study structural aspects of therecognition mechanism and regulation of target proteins.
Received: June 9, 2004Revised: September 30, 2004
Accepted: October 1, 2004
Keywords:
Barley a-amylase/subtilisin inhibitor / Disulphide reduction / Seed germination / Targetrecognition / Thioredoxin h
1634 Proteomics 2005, 5, 1634–1644
1 Introduction
Thioredoxins are small ubiquitous proteins containing aconserved active site sequence WCG(P)PC including tworedox active cysteines essential for the protein disulphidereductase activity [1]. The cysteines form a disulphide bondin the oxidised enzyme but are converted into the catalyti-cally active dithiol form when reduced by electron donorssuch as nicotinamide adenine dinucleotide phosphate(NADPH) or ferredoxin via thioredoxin reductases. Thereduced form of thioredoxin interacts with a variety of target
Correspondence: Dr. Christine Finnie, Biochemistry and NutritionGroup, BioCentrum-DTU, Søltofts Plads, Building 224, TechnicalUniversity of Denmark, Kgs. Lyngby, DenmarkE-mail: [email protected]: 145-4588-6307
Abbreviations: AMY2, barley a-amylase isozyme 2; BASI, barleya-amylase/subtilisin inhibitor; ESP, embryo-specific protein;HvTrxh1, barley thioredoxin h isoform 1; HvTrxh2, barley thiore-doxin h isoform 2; IAA, iodoacetamide; LTP 1, lipid transfer pro-tein 1; NTR, NADP-dependent thioredoxin reductase; TMA,Tenebrio molitor a-amylase
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de
DOI 10.1002/pmic.200401050

Proteomics 2005, 5, 1634–1644 Plant Proteomics 1635
proteins and regulates their activities by reducing disulphidebonds. Uniquely in plants, several thioredoxins coexist thatdiffer in subcellular locations and thus in interaction part-ners [2].
Cytosolic plant thioredoxins, referred to as h-types, areconsidered to play a role in seed germination [3–5]. Recently,various seed proteins from different plant sources have beensystematically identified as putative target proteins of thio-redoxin h from Chlamydomanas reinhardtii [5, 6] and ofEscherichia coli thioredoxin [4] by treatment of proteinextracts with thioredoxins, fluorescence labelling of acces-sible thiol groups, 2-DE and MS. Most recently, the methodwas optimised for sensitivity and resolution and applied in atarget survey of two endogenous thioredoxin h isoforms,barley thioredoxin h isoform 1 (HvTrxh1) and isoform 2(HvTrxh2) [7] in barley seeds [8] for which 2-D gel spot pat-terns have been mapped [9–12]. In addition to a-amylase/trypsin inhibitors and barley a-amylase/subtilisin inhibitor(BASI), already demonstrated as targets of thioredoxin h [13,14], novel putative target proteins such as three chitinaseisozymes, cyclophilin, lipid transfer protein 1 (LTP 1),superoxide dismutase and a protein with unknown function(embryo-specific protein (ESP)) were reduced by both iso-forms. Moreover, several disulphide-containing seed pro-teins previously identified in the 2-D gels apparentlyremained unreduced despite addition of thioredoxins, show-ing that thioredoxin h recognises and reduces disulphidebonds specifically. However, the strategy based on fluores-cence labelling coupled with 2-DE only indicates the loca-tions of target proteins in the 2-D gels and does not specifythe particular disulphides reduced by thioredoxins. In orderto fully understand the mechanisms in target protein recog-nition and regulation by thioredoxin h, identification ofthioredoxin h-reducible disulphides in the individual targetprotein is important, since the majority of those proteins hadmore than one disulphide bond or no available structuraldata. In addition, all putative target proteins identified usingthe 2-D gel based strategy require confirmation as targetproteins because comigration of additional proteins mighthave occurred in 2-DE.
For the well studied chloroplastic thioredoxins, targetdisulphide bonds in target enzymes were mainly identifiedby using strategies based on site directed mutagenesis of thetarget proteins [15–17]. However, to produce recombinanttarget proteins and their mutants would be extremely time-consuming considering the large number of putative targetproteins of thioredoxin h identified to date. On the otherhand, the techniques of proteomics are progressively appliedmore in high-throughput identification of cotranslationalmodification and PTM sites and have the potential to becomeuseful for identification of thioredoxin-catalysed disulphidereduction sites as well. Using MS techniques, peptide frag-ments containing residues with modifications, for examplephosphorylation, are recognised in the peptide mass mapsaccording to an exact mass shift from the expected theoreti-cal values [18]. The thiol and disulphide forms of cysteine
side chains, however, are usually not distinguished in MSbecause complete reduction and alkylation of cysteines areperformed during regular sample preparation.
To allow mass spectrometric detection of thioredoxinh-reducible disulphides, a novel strategy based on differ-ential alkylation of cysteine residues is established in thepresent study. After treatment with thioredoxin h, the acces-sible cysteinyl side chains in thiol form were alkylated usingiodoacetamide (IAA) whereas those inaccessible or formingdisulphides were labelled using 4-vinylpyridine. Accordingto the mass difference of cysteines modified by these tworeagents, conversion of disulphides to thiols catalysed byHvTrxh1 was shown in the peptide mass maps of recombi-nant BASI. Furthermore, the method of differential labellingwas compatible with 2-DE and employed to identify thio-redoxin h-reducible protein disulphide groups in barley seedextract. This introduction of identification of the specific di-sulphide bonds reduced by thioredoxin h provides the basisfor further exploring molecular details important for recog-nition and regulation of target proteins by thioredoxin h.
2 Materials and methods
2.1 Materials
Purified Arabidopsis thaliana NADP-dependent thioredoxinreductase (NTR) [19] was the gift of J.-P. Jacquot (INRA,Nancy, France). BASI and HvTrxh1 were produced as describ-ed previously [7, 20]. NADPH was from Fluka (Buchs,Switzerland). IAA and 4-vinylpyridine were from Sigma(St. Louis, MO, USA).
2.2 Reduction and differential labelling of
recombinant BASI
The inhibitory activity of recombinant BASI towards barleya-amylase isozyme 2 (AMY2) was confirmed using the bluestarch assay as described [20]. Purified recombinant BASI(1.1 mg) was incubated with 0.60 mM NADPH, 2.2 mM Arabi-dopsis NTR and 8.0 mM HvTrxh1 (no thioredoxin in control) in5 mL 50 mM Tris-HCl pH 7.5 at room temperature (227C) for1 h. The reaction mixtures were incubated with 50 mM IAAin a total volume of 15 mL 50 mM NH4HCO3 pH 8.0 for30 min to carbamidomethylate (CAM) the accessible thiolgroups. To remove the excess IAA, the resulting mixtureswere separated on SDS-PAGE gels (NuPAGE 4–12% Bis-Trisgels; Invitrogen Leek, The Netherlands) under non-reducingconditions using Xcell SureLock MiniCell (Invitrogen)according to the manufacturer’s recommendations andstained using colloidal Coomassie Brilliant Blue (cCBB) [21].Excised gel pieces containing BASI were incubated with10 mM DTTat 567C for 45 min and subsequently with 50 mM
4-vinylpyridine at room temperature for 30 min in 50 mL50 mM NH4HCO3 pH 8.0 to ensure complete reduction and
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

1636 K. Maeda et al. Proteomics 2005, 5, 1634–1644
S-pyridylethylation of the remaining cysteine residues. Thedifferentially labelled BASI was subjected to in-gel trypsindigestion [22].
2.3 Reduction and differential labelling of barley seed
proteins
The extracts of soluble proteins from mature barley seeds (cv.Barke) were prepared as described earlier [10]. Reactionmixtures containing 0.72 mM NADPH, 0.44 mM ArabidopsisNTR, 3.0 mM HvTrxh1 (no thioredoxin in control) and 150 mLmature seed extract (approximately 150 mg protein) in a totalvolume of 250 mL in 50 mM Tris-HCl pH 7.5 were incubatedat room temperature for 1 h. The reaction mixtures weretreated with 50 mM IAA in 500 mL 50 mM NH4HCO3 pH 8.0at room temperature for 30 min to carbamidomethylateaccessible thiol groups. The proteins were subsequentlydesalted using G25 columns (NAP10; Amersham Bio-sciences, Uppsala, Sweden) to remove excess IAA. Proteinswere precipitated with four volumes of acetone at 2207C for24 h, resuspended in reswelling buffer [8 M urea, 2% w/vCHAPS, 0.5% v/v IPG buffer 3–10 (Amersham Biosciences),20 mM DTT and a trace of bromophenol blue) and loaded on18 cm linear IPG strips, pI 3–10 according to the manu-facturer’s recommendations. The IPG strips were equili-brated in SDS equilibration buffer (50 mM Tris-HCl pH 8.8,6 M urea, 30% v/v glycerol, 2% w/v SDS and 0.01% w/vbromphenol blue) containing 65 mM DTT to ensure com-plete reduction of disulphide bonds and then in the SDSequilibration buffer containing 135 mM 4-vinylpyridine tocompletely S-pyridylethylate the remaining cysteine resi-dues. Second dimension SDS-PAGE gels (12–14%,18 6 24 cm; Amersham Biosciences) were run on a Phar-macia Multiphor II according to the manufacturers recom-mendations and stained using cCBB [21]. Spots were select-
ed based on coverage of cysteine residues in the earlier MSanalysis of the tryptic peptides, excised from the gels andsubjected to in-gel trypsin digestion [22].
The procedure of differential labelling coupled with 2-DEwas also attempted using IAA and 4-vinylpyridine in theopposite order. The procedure was changed to the presentform due to pI shift and solubility problems for some pro-teins treated with 4-vinylpyridine prior to the first dimensionof 2-DE.
2.4 MALDI-TOF MS
Tryptic peptides were micro purified on a home-made 5 mmnanocolumn [23] and analysed using a Bruker Reflex IIIMALDI-TOF mass spectrometer (Bruker-Daltonics, Bremen,Germany) as described earlier [10]. The m/z software (Pro-teometrics, NY, USA) was used to analyse spectra. Spectrawere calibrated using trypsin autolysis products (m/z 842.51and m/z 2211.10) as internal standards. Peaks with massderivations less than 50 ppm from the theoretical valueswere accepted as matches.
3 Results
3.1 Identification of disulphide bonds
reduced by HvTrxh1 in BASI
The structure of BASI contains two disulphide bonds be-tween Cys43 and Cys90, and Cys144 and Cys148 both locatedat the surface, but no cysteines in free form [24] (Fig. 1A).Recombinant BASI was incubated in the presence orabsence of HvTrxh1 and the accessible thiol groups weresubsequently carbamidomethylated by IAA. The treatedBASI was purified by 1-D SDS-PAGE in nonreducing con-
Figure 1. Reduction of disulphide bonds inrecombinant BASI by HvTrxh1. (A) The aminoacid sequence of BASI. The tryptic peptides con-taining cysteine residues (white letters withblack background) are indicated with grey back-ground. Disulphides Cys43-Cys90 and Cys144-Cys148 are drawn with black lines. (B) Selectedsections from the MALDI-TOF MS spectrum ofBASI incubated with (upper spectrum) or with-out (lower spectrum) HvTrxh1, showing reduc-tion of disulphides Cys144-Cys148 (left) andCys43-Cys90 (right). Vertical lines are drawn fororientation. The mass difference of 48 Dabetween carbamidomethylated and pyridyl-ethylated cysteines is indicated (see alsoTable 1).
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

Proteomics 2005, 5, 1634–1644 Plant Proteomics 1637
ditions. A gel piece containing BASI was excised and theremaining nonalkylated cysteine residues were completelyreduced and pyridylethylated. Thus, cysteine side chains inaccessible reduced form after thioredoxin h-treatment werecarbamidomethylated causing a 57 Da mass increase,whereas inaccessible or disulphide-bonded cysteines werepyridylethylated resulting in a 105 Da mass increase. The al-kylated BASI was digested with trypsin and the peptidefragments were analysed by MALDI-TOF MS (Fig. 1B,Table 1). In the control peptide map of BASI alkylated with-out prior thioredoxin h-treatment, three peptides containingpyridylethylated cysteines were observed, with [M 1 H]1
2355.1 (residues 42–61 including Cys43), [M 1 H]1 2457.2(residues 86–106 including Cys90) and [M 1 H]1 1939.9(residues 141–155 including Cys144 and Cys148). Togetherthese peptides covered all four cysteine residues (Fig. 1,Table 1). The mass of carbamidomethylated cysteine is48.0 Da lower than that of pyridylethylated cysteine. No pep-tides containing carbamidomethylated cysteines were pres-ent at the expected m/z values in the control peptide map inagreement with all four cysteines forming disulphide bonds(Fig. 1B). When BASI was treated with HvTrxh1, three tryp-tic peptides containing carbamidomethylated cysteineappeared, with [M 1 H]1 2307.1 (residues 42–61 includingCys43), [M 1 H]1 2409.1 (residues 86–106 including Cys90)and [M 1 H]1 1843.8 (residues 141–155 including Cys144and Cys148), showing reduction of both Cys43-Cys90 andCys144-Cys148 (Fig. 1B, Table 1). Although the three pep-tides containing pyridylethylated cysteines were still present,the peak at [M 1 H]1 1939.9 decreased remarkably in rela-tive intensity (Fig. 1B), suggesting almost complete reduc-tion of the disulphide Cys144-Cys148 by HvTrxh1. In con-trast, the relative peak intensities of the two peptides con-taining pyridylethylated Cys43 and Cys90 did not changesignificantly, indicating that the disulphide Cys43-Cys90remained mostly unreduced (Fig. 1B). A small peak at[M 1 H]1 1891.8 matched the theoretical mass of a peptide
containing Cys144 and Cys148 with one carbamidomethyla-tion and one pyridylethylation (Fig. 1B, Table 1). The incom-plete carbamidomethylation of free thiol groups convertedfrom disulphide Cys144-Cys148 either signifies inaccessi-bility after the first is labelled or general incomplete alkyla-tion reaction. Altogether, the result shows that HvTrxh1 iscapable of reducing both disulphide bonds in recombinantBASI, but apparently the disulphide Cys144-Cys148 to amuch greater extent than disulphide Cys43-Cys90.
3.2 Identification of thioredoxin h-reducible
disulphide bonds in extracts of barley seed
proteins
The successful thioredoxin h-reducible disulphide bondidentification in recombinant BASI confirmed the suitabilityof the differential labelling strategy for identification of pro-tein disulphide bonds reduced by thioredoxins. This analyti-cal procedure was then coupled with 2-DE to extend itsapplication to mixtures of proteins as found in extracts ofbiological samples. Proteins in barley seed extracts wereincubated with or without added HvTrxh1 and the accessiblefree thiol groups were carbamidomethylated under nativeconditions. Proteins were desalted to remove excess IAA andsubsequently separated by IEF in denaturating and reducingconditions. The remaining nonalkylated cysteine side chainswere completely pyridylethylated in the IPG strips prior tothe second dimension SDS-PAGE. From the resulting 2-Dgels, proteins for analysis were selected among the putativetarget proteins of HvTrxh1 identified previously [8] based oncoverage of cysteine residues in the prior MS analysis of thetryptic peptides. The spots containing selected proteins wereexcised based on our prior knowledge of the 2-D spot pattern[8–12] and subjected to in-gel digestion. The tryptic digestswere analysed by MALDI-TOF MS. All peaks matching thetheoretical masses of peptides containing modified cysteineresidues are listed in Table 2.
Table 1. Tryptic peptides and modifications of cysteine residues after differential labelling of recombinant barley a-amylase/subtilisin in-hibitor (BASI)
Peptide fragment Covereddisulphide
Cysteinemodificationsa)
Theoreticalmass of observedpeptides (Da)
Change inducedby additionof HvTrxh1b)
42HC43PLFVSQDPNGQHDGFPVR61 Cys43-Cys90 CAM 2307.1 :PE 2355.1 0
86AYTTC90LQSTEWHIDSELAAGR106 Cys43-Cys90 CAM 2409.1 :PE 2457.2 0
141LMSC144GDWC148QDLGVFR155 Cys144-Cys148 26 CAM 1843.8 :CAM, PE 1891.8 :26 PE 1939.9 ;
a) Peptides containing pyridylethylated (PE) or carbamidomethylated (CAM) cysteines (Fig. 1B) after differential labelling are listed.b) Influence of HvTrxh1 addition on appearance and intensity of peaks: :, only appearing in HvTrxh1 treated samples; ;, decreasing sig-
nificantly in intensity after HvTrxh1 addition; and 0, intensity not significantly affected by addition of HvTrxh1.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

1638 K. Maeda et al. Proteomics 2005, 5, 1634–1644
Table 2. Tryptic peptides and modifications of cysteine residues after differential labelling of proteins in barley seed extracts
Spota) Protein Peptide fragment Covereddisulphide
Cysteinemodifi-cationsb)
Theoreti-cal massof ob-servedpeptides(Da)
Changeinducedby addi-tion ofHvTrxh1c)
56 Barley a-amylase/subtilisininhibitor (BASI)
42HC43PLFVSQDPNGQHDGFPVR61 C43-C90 CAM 2307.1 :PE 2355.1 0
86AYTTC90LQSTEWHIDSELAAGR106 C43-C90 CAM 2409.1 :PE 2457.2 0
86AYTTC90LQSTEWHIDSELAAG RR107 C43-C90 CAM 2565.2 :PE 2613.3 0
141LMSC144GDWC148QDLGVFR155 C144-C148 26 CAM 1843.8 :CAM, PE 1891.8 :26 PE 1939.9 ;
111, 112 a-Amylase inhibitor BMAI-1 1SPGEWC6 WPGMGYPVYPFPR19 1 CAM 2283.0 :PE 2331.1 ;
26SQC28AGGQVVESIQK39 3 PE 1538.8 044QIAAIGDEWC53IC55GALGSMR62 1, 5 CAM, PE 2156.0 :
26 PE 2204.0 ;44QIAAIGDEWC53IC55GALGSMR62,
MSO:611, 5 CAM, PE 2172.0 :
26 PE 2220.0 ;68ELGVALADDKATVAEVFPGC87R88 3 PE 2266.2 078ATVAEVFPGC87R88 3 PE 1254.6 0
116, 495 a-Amylase inhibitor BDAI-I 1SGPWMWC7DPEMGHK14 1 PE 1765.7 027LEC29VGNRVPEDVLR40 3 PE 1703.9 041DC42C43QEVANISNEWC54R55 1, 2, 4 CAM, 26 PE 2036.8 :
36 PE 2084.9 090LLVAGVPALC99NVPIPNEAAG TR111 4 CAM 2232.2 :
PE 2280.3 0
28, T2 Trypsin inhibitor CMe 1FGDSC5APGDALPHNPLR17 1 CAM 1823.8 :PE 1871.9 0
21TYVVSQIC28HQGPR33 3 CAM 1544.8 :PE 1592.8 0
42RC43C44DELSAIPAYC54R55 1, 2, 4 36 PE 1914.9 043C43C44DELSAIPAYC54R55 1, 2, 4 36 PE 1758.8 0
113a a-Amylase/trypsin inhibitor,pUP13
26SIPINPLPAC35R36 2 CAM 1237.7 :PE 1285.7 0
43C43AVGDQQVPDVLK55 3 PE 1476.8 043C43AVGDQQVPDVLKQQC58C59R60 2, 3, 4 36 PE 2305.1 061ELSDLPESC69R70 1 PE 1253.6 071C71DALSILVNGVITEDGSR88 5 CAM 1919.0 :
PE 1967.0 0
117 a-Amylase/trypsin inhibitor,CMb
1VGSEDC6TPWTATPITPLPSC20R21 1, 2 26 PE 2441.2 042QQC44C45GELANIPQQC55R56 1, 2, 4 36 PE 2005.9 067KSRPDQSGLMELPGC81PR83 3 PE 1976.0 068SRPDQSGLMELPGC81PR83 3 PE 1847.9 068SRPDQSGLMELPGC81PR83, MSO: 76 3 PE 1863.9 0
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

Proteomics 2005, 5, 1634–1644 Plant Proteomics 1639
Table 2. Continued
Spota) Protein Peptide fragment Covereddisulphide
Cysteinemodifi-cationsb)
Theoreti-cal massof ob-servedpeptides(Da)
Changeinducedby addi-tion ofHvTrxh1c)
115b a-Amylase/trypsin inhibitor,CMd
4AATDC8SPGVAFPTNLLGHC22R23 1, 2 PE 2240.1 060LYC62C63QELAEIPQQC73R714 1, 2, 4 36 PE 2112.0 0120LLVAPGQC127NLATIHNVR136 4 PE 1924.1 0
8 Embryo-specific protein 1MAASC5HDVDVPGK13, MSO: 1 – PE 1450.6 055QVEAHHFC62AHLNEDVR70 – PE 2009.9 071QC72LIFDGPDAGAR83 – PE 1467.7 0169EGQLRQDLADC179VER182 – CAM 1688.8 :
PE 1736.8 ;169EGQLRQDLADC179VERK183 – CAM 1816.9 :
PE 1864.9 ;
a) Spot numbers refer to Fig. 2A [8–12].b) Peptides containing pyridylethylated (PE) or carbamidomethylated (CAM) cysteines (Fig. 1B) after differential labelling are listed.c) Influence of HvTrxh1 addition on appearance and intensity of peaks: :, only appearing in HvTrxh1 treated samples; ;, decreasing sig-
nificantly in intensity after HvTrxh1 addition; and 0, intensity not significantly affected by addition of HvTrxh1.
3.2.1 BASI
The peptide maps of BASI (P07596) from spot 56 [11](Fig. 2A) in the 2-D gels of mature seed proteins were com-parable to the peptide maps of recombinant BASI describedabove (Fig. 1B, Tables 1, 2). As for recombinant BASI, pre-incubation with HvTrxh1 decreased the intensity of the peakat [M 1 H]1 1939.9 covering pyridylethylated Cys144 andCys148 but did not affect the intensity of the peaks coveringthe peptides containing pyridylethylated Cys43 and Cys90(Table 2). Besides the peaks also present in the peptide mapsof recombinant BASI, a peak at [M 1 H]1 2613.3 (residues86–107, covering Cys90, 1 missed cleavage) containing pyr-idylethylated Cys90 was observed in the peptide maps(Table 2). A trace amount of a peak at [M 1 H]1 2565.2 cor-responding to the same peptide containing carbamido-methylated Cys90 emerged after incubation with HvTrxh1.
3.2.2 Æ-Amylase/trypsin inhibitors
Seven a-amylase/trypsin inhibitors belonging to the familyof CM proteins (chloroform-methanol) [25] were previouslyidentified as putative target proteins of HvTrxh1 andHvTrxh2 [8]. The cysteine status in a-amylase inhibitorBMAI-1 (P16968, spots 111 and 112), a-amylase inhibitorBDAI-I (P13691, spots 116 and 495), a-amylase/trypsin in-hibitor CMb (P32936, spot 117), a-amylase/trypsin inhibitorCMd (P11643, spot 115b), trypsin inhibitor CMe (P01086,spot 28, T2) and a-amylase/trypsin inhibitor pUP13(gi/225102, spot 113a) excised from the 2-D gels [8, 12] wereanalysed in the present study (Table 2). They all contain ten
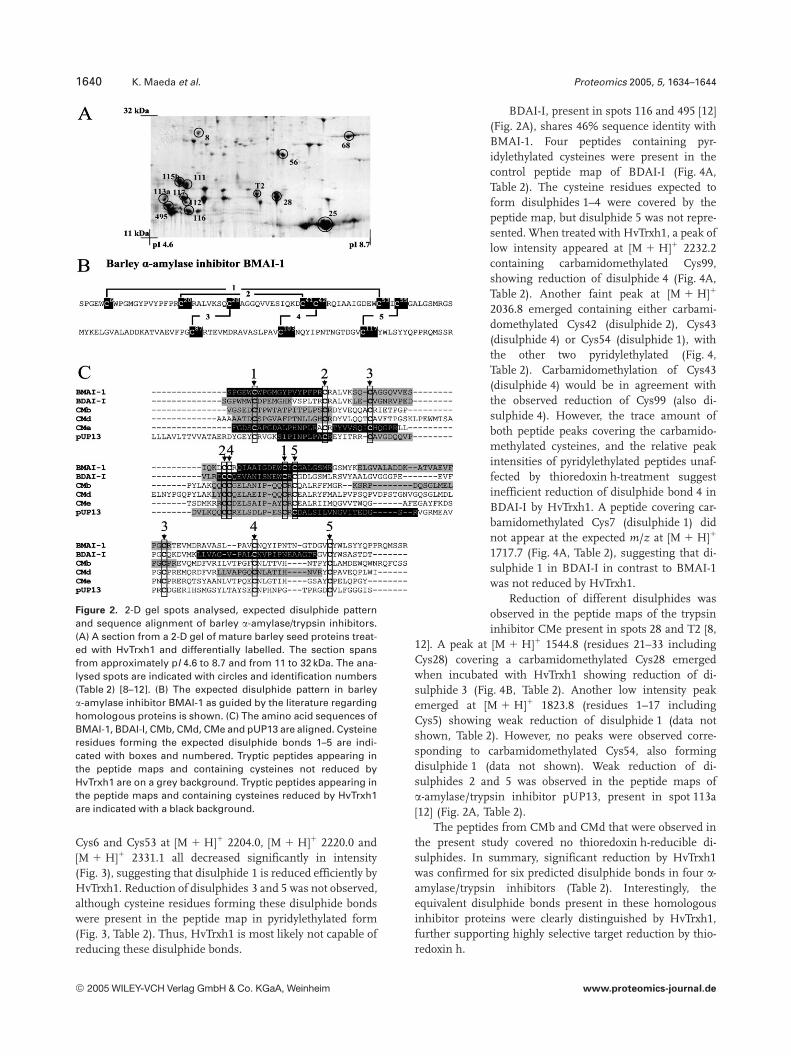
conserved cysteines expected to form five disulphide bonds(Fig. 2B, C) as seen in structures of homologous CM proteins[26, 27]. The expected disulphide profile in BMAI-1 is shownin Fig. 2B. The amino acid sequences of the seven a-amy-lase/trypsin inhibitors are aligned in Fig. 2C and share be-tween 47 and 19% identity.
In the control peptide map obtained from BMAI-1 inboth spots 111 (probably glycosylated form [28]) and 112 [12](Fig. 2A) in the absence of reduction by HvTrxh1, six pep-tides containing pyridylethylated cysteines were observed to-gether covering five of the ten cysteine residues. No peptidescontaining carbamidomethylated cysteines were observed(Fig. 3, Table 2). This pyridylethylation was in agreementwith disulphides 1, 3 and 5, but the peptide map coverednone of the cysteine residues expected to form disulphides 2and 4 (Fig. 2B). When the protein extract was treated withHvTrxh1, BMAI-1 gave an intense peak at [M 1 H]1 2283.0corresponding to the peptide containing carbamidomethy-lated Cys6 in the peptide map unambiguously showing thereduction of disulphide 1 (Fig. 3, Table 2). The appearance oftwo peaks corresponding to Q44-R62 at [M 1 H]1 2156.0 and[M 1 H]1 2172.0, covering Cys53 and Cys55, indicated car-bamidomethylation of either Cys53 or Cys55 (with the otherone pyridylethylated). Identification of the carbamido-methyled cysteine residue is not possible by MALDI-TOFMS in cases where two or more cysteine residues with dif-ferent modifications occur in the same peptide. However,since Cys53 is expected to form disulphide 1 with Cys6(Fig. 2B), the emergence of these peaks further supports theidentification of disulphide 1 as thioredoxin h-reducible. Thepeaks corresponding to peptides covering pyridylethylated
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de
1640 K. Maeda et al. Proteomics 2005, 5, 1634–1644
Figure 2. 2-D gel spots analysed, expected disulphide patternand sequence alignment of barley a-amylase/trypsin inhibitors.(A) A section from a 2-D gel of mature barley seed proteins treat-ed with HvTrxh1 and differentially labelled. The section spansfrom approximately pI 4.6 to 8.7 and from 11 to 32 kDa. The ana-lysed spots are indicated with circles and identification numbers(Table 2) [8–12]. (B) The expected disulphide pattern in barleya-amylase inhibitor BMAI-1 as guided by the literature regardinghomologous proteins is shown. (C) The amino acid sequences ofBMAI-1, BDAI-I, CMb, CMd, CMe and pUP13 are aligned. Cysteineresidues forming the expected disulphide bonds 1–5 are indi-cated with boxes and numbered. Tryptic peptides appearing inthe peptide maps and containing cysteines not reduced byHvTrxh1 are on a grey background. Tryptic peptides appearing inthe peptide maps and containing cysteines reduced by HvTrxh1are indicated with a black background.
Cys6 and Cys53 at [M 1 H]1 2204.0, [M 1 H]1 2220.0 and[M 1 H]1 2331.1 all decreased significantly in intensity(Fig. 3), suggesting that disulphide 1 is reduced efficiently byHvTrxh1. Reduction of disulphides 3 and 5 was not observed,although cysteine residues forming these disulphide bondswere present in the peptide map in pyridylethylated form(Fig. 3, Table 2). Thus, HvTrxh1 is most likely not capable ofreducing these disulphide bonds.
BDAI-I, present in spots 116 and 495 [12](Fig. 2A), shares 46% sequence identity withBMAI-1. Four peptides containing pyr-idylethylated cysteines were present in thecontrol peptide map of BDAI-I (Fig. 4A,Table 2). The cysteine residues expected toform disulphides 1–4 were covered by thepeptide map, but disulphide 5 was not repre-sented. When treated with HvTrxh1, a peak oflow intensity appeared at [M 1 H]1 2232.2containing carbamidomethylated Cys99,showing reduction of disulphide 4 (Fig. 4A,Table 2). Another faint peak at [M 1 H]1
2036.8 emerged containing either carbami-domethylated Cys42 (disulphide 2), Cys43(disulphide 4) or Cys54 (disulphide 1), withthe other two pyridylethylated (Fig. 4,Table 2). Carbamidomethylation of Cys43(disulphide 4) would be in agreement withthe observed reduction of Cys99 (also di-sulphide 4). However, the trace amount ofboth peptide peaks covering the carbamido-methylated cysteines, and the relative peakintensities of pyridylethylated peptides unaf-fected by thioredoxin h-treatment suggestinefficient reduction of disulphide bond 4 inBDAI-I by HvTrxh1. A peptide covering car-bamidomethylated Cys7 (disulphide 1) didnot appear at the expected m/z at [M 1 H]1
1717.7 (Fig. 4A, Table 2), suggesting that di-sulphide 1 in BDAI-I in contrast to BMAI-1was not reduced by HvTrxh1.
Reduction of different disulphides wasobserved in the peptide maps of the trypsininhibitor CMe present in spots 28 and T2 [8,
12]. A peak at [M 1 H]1 1544.8 (residues 21–33 includingCys28) covering a carbamidomethylated Cys28 emergedwhen incubated with HvTrxh1 showing reduction of di-sulphide 3 (Fig. 4B, Table 2). Another low intensity peakemerged at [M 1 H]1 1823.8 (residues 1–17 includingCys5) showing weak reduction of disulphide 1 (data notshown, Table 2). However, no peaks were observed corre-sponding to carbamidomethylated Cys54, also formingdisulphide 1 (data not shown). Weak reduction of di-sulphides 2 and 5 was observed in the peptide maps ofa-amylase/trypsin inhibitor pUP13, present in spot 113a[12] (Fig. 2A, Table 2).
The peptides from CMb and CMd that were observed inthe present study covered no thioredoxin h-reducible di-sulphides. In summary, significant reduction by HvTrxh1was confirmed for six predicted disulphide bonds in four a-amylase/trypsin inhibitors (Table 2). Interestingly, theequivalent disulphide bonds present in these homologousinhibitor proteins were clearly distinguished by HvTrxh1,further supporting highly selective target reduction by thio-redoxin h.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

Proteomics 2005, 5, 1634–1644 Plant Proteomics 1641
Figure 3. The peptide maps of BMAI-1 frommature barley seeds showing specific disulphidereduction by HvTrxh1. Sections of MALDI spectraof BMAI-1 from the 2-D gels of barley seed pro-teins incubated in the presence (upper spectrum)or absence (lower spectrum) of HvTrxh1. Eachsection covers expected m/z values of peptidescontaining pyridylethylated cysteines and corre-sponding peptides containing carbamidomethy-lated cysteines with 48 Da lower mass. The m/zvalues of the peaks are specified and the cysteineresidues and disulphide bonds (in brackets) cov-ered by the peptides are indicated. Peptides
containing pyridylethylated cysteines: [M 1 H]1 1254.6 (residues 78–88 including Cys87 (disulphide 3)); [M 1 H]1 1538.8 (residues 26–39including Cys28 (disulphide 3)); [M 1 H]1 2204.0 (residues 44–62 including Cys53 (disulphide 1) and Cys55 (disulphide 5)); [M 1 H]1 2220.0(residues 44–62 including Cys53 (disulphide 1), Cys55 (disulphide 5) and oxidised Met61); [M 1 H]1 2266.2 (residues 68–88 including Cys87(disulphide 3), with 1 missed cleavage) and [M 1 H]1 2331.1 (residues 1–19 including Cys6 (disulphide1)).
Figure 4. The peptide maps of BDAI-I and CMefrom mature barley seeds showing specific di-sulphide reduction by HvTrxh1. The peptidemaps of BDAI-I (A) and CMe (B) from the 2-D gelsof barley seed proteins incubated in the pres-ence (upper spectrum) or absence (lower spec-trum) of HvTrxh1. Each section covers expectedm/z values of peptides containing pyr-idylethylated cysteines and corresponding pep-tides containing carbamidomethylated cysteineswith 48 Da lower mass. The m/z values of thepeaks are specified and the cysteine residuesand disulphide bonds (in brackets) covered by
the appearing peptides are indicated. For BDAI-I, the peaks from the following tryptic peptides containing pyridylethylated cysteines areshown: [M 1 H]1 1765.7 (residues 1–14 including Cys7 (disulphide 1)); [M 1 H]1 1703.9 (residues 27–40 including Cys29 (disulphide 3));[M 1 H]1 2084.9 (residues 41–55 including Cys42 (disulphide 2), Cys43 (disulphide 4) and Cys54 (disulphide 1)) and [M 1 H]1 2280.3 (resi-dues 90–111 including Cys99 (disulphide 4). For CMe, a peak at m/z value [M 1 H]1 1592.8 (residues 21–33 including Cys28) from a peptidecontaining pyridylethylated cysteine is shown.
3.2.3 Embryo-specific protein (ESP)
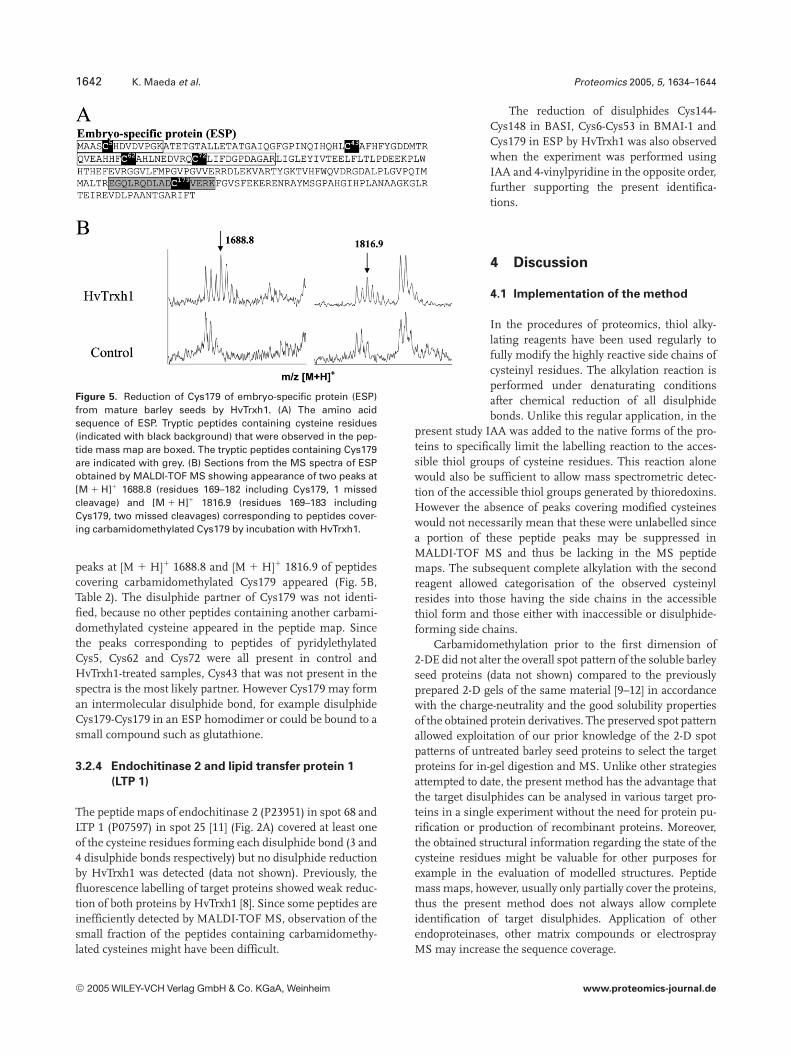
ESP (TC109921) in spot 8 [10] (Fig. 2A) was previously iden-tified as a putative target protein of barley thioredoxin h iso-forms [8]. In contrast to the target proteins examined so farin the present study, no structural information is availablefor ESP or for any homologous proteins, thus the disulphidepattern remains to be identified. The translated tentativeconsensus sequence of ESP contains five cysteine residues,
Cys5, Cys43, Cys62, Cys72 and Cys179 with unknown redoxstate and accessibility (Fig. 5A). In the control peptide map,five peptides containing pyridylethylated cysteines werepresent together covering four (Cys43 was not covered) of thecysteine residues (Fig. 5A, Table 2). The size of the trypticpeptide covering Cys43 was beyond the efficient detectionrange (.4000 Da). Thus, these four cysteine residues areeither in an oxidised form or inaccessible in the native ESP inmature barley seeds. When incubated with HvTrxh1, two
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de
1642 K. Maeda et al. Proteomics 2005, 5, 1634–1644
Figure 5. Reduction of Cys179 of embryo-specific protein (ESP)from mature barley seeds by HvTrxh1. (A) The amino acidsequence of ESP. Tryptic peptides containing cysteine residues(indicated with black background) that were observed in the pep-tide mass map are boxed. The tryptic peptides containing Cys179are indicated with grey. (B) Sections from the MS spectra of ESPobtained by MALDI-TOF MS showing appearance of two peaks at[M 1 H]1 1688.8 (residues 169–182 including Cys179, 1 missedcleavage) and [M 1 H]1 1816.9 (residues 169–183 includingCys179, two missed cleavages) corresponding to peptides cover-ing carbamidomethylated Cys179 by incubation with HvTrxh1.
peaks at [M 1 H]1 1688.8 and [M 1 H]1 1816.9 of peptidescovering carbamidomethylated Cys179 appeared (Fig. 5B,Table 2). The disulphide partner of Cys179 was not identi-fied, because no other peptides containing another carbami-domethylated cysteine appeared in the peptide map. Sincethe peaks corresponding to peptides of pyridylethylatedCys5, Cys62 and Cys72 were all present in control andHvTrxh1-treated samples, Cys43 that was not present in thespectra is the most likely partner. However Cys179 may forman intermolecular disulphide bond, for example disulphideCys179-Cys179 in an ESP homodimer or could be bound to asmall compound such as glutathione.
3.2.4 Endochitinase 2 and lipid transfer protein 1
(LTP 1)
The peptide maps of endochitinase 2 (P23951) in spot 68 andLTP 1 (P07597) in spot 25 [11] (Fig. 2A) covered at least oneof the cysteine residues forming each disulphide bond (3 and4 disulphide bonds respectively) but no disulphide reductionby HvTrxh1 was detected (data not shown). Previously, thefluorescence labelling of target proteins showed weak reduc-tion of both proteins by HvTrxh1 [8]. Since some peptides areinefficiently detected by MALDI-TOF MS, observation of thesmall fraction of the peptides containing carbamidomethy-lated cysteines might have been difficult.
The reduction of disulphides Cys144-Cys148 in BASI, Cys6-Cys53 in BMAI-1 andCys179 in ESP by HvTrxh1 was also observedwhen the experiment was performed usingIAA and 4-vinylpyridine in the opposite order,further supporting the present identifica-tions.
4 Discussion
4.1 Implementation of the method
In the procedures of proteomics, thiol alky-lating reagents have been used regularly tofully modify the highly reactive side chains ofcysteinyl residues. The alkylation reaction isperformed under denaturating conditionsafter chemical reduction of all disulphidebonds. Unlike this regular application, in the
present study IAA was added to the native forms of the pro-teins to specifically limit the labelling reaction to the acces-sible thiol groups of cysteine residues. This reaction alonewould also be sufficient to allow mass spectrometric detec-tion of the accessible thiol groups generated by thioredoxins.However the absence of peaks covering modified cysteineswould not necessarily mean that these were unlabelled sincea portion of these peptide peaks may be suppressed inMALDI-TOF MS and thus be lacking in the MS peptidemaps. The subsequent complete alkylation with the secondreagent allowed categorisation of the observed cysteinylresides into those having the side chains in the accessiblethiol form and those either with inaccessible or disulphide-forming side chains.
Carbamidomethylation prior to the first dimension of2-DE did not alter the overall spot pattern of the soluble barleyseed proteins (data not shown) compared to the previouslyprepared 2-D gels of the same material [9–12] in accordancewith the charge-neutrality and the good solubility propertiesof the obtained protein derivatives. The preserved spot patternallowed exploitation of our prior knowledge of the 2-D spotpatterns of untreated barley seed proteins to select the targetproteins for in-gel digestion and MS. Unlike other strategiesattempted to date, the present method has the advantage thatthe target disulphides can be analysed in various target pro-teins in a single experiment without the need for protein pu-rification or production of recombinant proteins. Moreover,the obtained structural information regarding the state of thecysteine residues might be valuable for other purposes forexample in the evaluation of modelled structures. Peptidemass maps, however, usually only partially cover the proteins,thus the present method does not always allow completeidentification of target disulphides. Application of otherendoproteinases, other matrix compounds or electrosprayMS may increase the sequence coverage.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

Proteomics 2005, 5, 1634–1644 Plant Proteomics 1643
4.2 Identified target disulphide bonds
BASI is suggested to be implicated in regulation of the starchdegrading activity of AMY2 in prematurely sprouting seeds[29]. A study of transgenic barley seeds overexpressing wheatthioredoxin h showed increase of a-amylase activity earlier inthe germination process compared to normal seeds [4]. It hasbeen demonstrated that incubation with wheat thioredoxin hdecreases the inhibitory activity of BASI towards AMY2 andsubtilisin [14], but the inhibition kinetic parameters of thereduced BASI have not been determined. Thioredoxin h alsorenders BASI more susceptible towards proteolytic degrada-tion, and the regulatory importance of this loss of AMY2inhibition for the germination process was suggested [14]. Thecapability of the endogenous thioredoxin h isoforms, HvTrxh1and HvTrxh2, to reduce BASI was also shown [7, 8]. Di-sulphide Cys144-Cys148, shown here to be efficiently reducedby HvTrxh1, is located on the structure surface close to Lys140and Asp150 in the AMY2 binding site according to the crystalstructure of the complex between BASI and AMY2 [24].Mutational analysis of BASI has shown that point mutationsof Lys140 or Asp150 highly reduced the inhibitory activity ofBASI towards AMY2 [30, 38]. Lys140 and Asp150 are posi-tioned in two different b-strands flanking loop Gly145-Cys148and it is likely that the structural changes around this loopcaused by reduction of the disulphide Cys144-Cys148 lowerthe affinity of BASI towards AMY2. Determination of bindingkinetics towards AMY2 of BASI mutants lacking this di-sulphide bond is one possible approach to confirm the sug-gested effects of the disulphide reduction.
HvTrxh1 is demonstrated here to reduce the disulphideCys43-Cys90 of BASI much less efficiently as judged by theremaining high relative peak intensities of the peptides con-taining pyridylethylated Cys43 and Cys90 after incubationwith HvTrxh1. This disulphide bond is located on the oppositeside of BASI to disulphide Cys144-Cys148, and close to thebinding site for proteases. Mutations of residues near Cys43-Cys90 such as Tyr87 are shown to have some effect on bindingto savinase of the subtilisin family of proteases (Bønsager,Nielsen, Fukuda, Prætorius-Ibba, Svensson, manuscript inpreparation). The cysteine residues corresponding to Cys144-Cys148 shown here to be targeted by thioredoxin h are con-served in the homologous proteins RASI from rice and WASIfrom wheat. However, the disulphide Cys43-Cys90 in BASIthat is less favoured by thioredoxin is not conserved in WASI.
Considering target recognition by thioredoxin h, thepresence of a bulky side chain of Trp147 adjacent to di-sulphide Cys144-Cys148 is notable (Fig. 1). Several con-served hydrophobic residues are assembled on the putativeinteraction surface in thioredoxins, thus the importance ofhydrophobic interaction for target recognition has been sug-gested [31]. The charge distribution differs around the twodisulphides in BASI and this may also influence recognitionby thioredoxins. The influence of surface charge distributionon recognition of some chloroplastic target proteins by thio-redoxins has been reported [32].
In the present study, HvTrxh1 was shown to be capableof efficiently reducing the disulphide Cys6-Cys53 (disul-phide 1) in BMAI-1, which inhibits a-amylase (TMA)from the yellow meal worm Tenebrio molitor [28]. The struc-ture of TMA in complex with a homologous inhibitor pro-tein from another plant source, ragi bifunctional inhibitor,shows that several residues adjacent to the disulphide bondcorresponding to disulphide 1 of BMAI-1 are located in theinteraction surface and participate in the binding [26]. Theimportance of some of these residues for the binding,including the conserved N-terminal serine, is also con-firmed by a mutational study of wheat monomerica-amylase inhibitor WMAI-1 [33]. In fact, wheat a-amylaseinhibitor 0.53 has a proline at the position of Cys53 ofBMAI-1 and inhibits TMA and bovine pancreas a-amylaseless efficiently than wheat a-amylase inhibitor 0.19 that hasall conserved cysteines, in spite of 94% sequence identity[34, 35].
Since HvTrxh1 was not observed to reduce the dis-ulphide counterparts of Cys6-Cys53 (disulphide 1) ofBMAI-1 in other barley a-amylase/trypsin inhibitors,BMAI-1 should have unique structural features around thisdisulphide bond required for efficient recognition byHvTrxh1. Assembly of bulky hydrophobic groups such asthe side chains of Trp5, Trp7 and Trp52 are adjacent to thedisulphide Cys6-Cys53 in the amino acid sequence ofBMAI-1 (Fig. 2B, C), further indicating the possible role ofhydrophobic interaction for the target recognition. Thequaternary structures may also affect the accessibility ofsome disulphide bonds in a-amylase/trypsin inhibitors.BMAI-1 is monomeric [31], while BDAI-I forms a homo-dimer [36] and CMa, CMb and CMd are found as hetero-tetramers [37].
The identification of Cys179 of ESP as a cysteine resi-due forming a target disulphide bond with an unidentifiedpartner confirms the previous identification of ESP as atarget protein [8]. Moreover, the structural data obtainedregarding cysteine residues is novel information becauseno 3-D structures are available for ESP or any homologueproteins. In addition to Cys179, which is shown to form adisulphide bond, Cys5, Cys62 and Cys72 are shown to beeither structurally inaccessible or forming disulphidebonds that are not targeted by thioredoxin h.
Specific thioredoxin h-reducible disulphides were iden-tified for first time in any protein in the present study.This knowledge enables further investigation of the activityregulation of the target proteins. Production of mutantslacking these identified target disulphide bonds, character-isation and comparison of their activities with wild-typeswould reveal the regulatory importance of thioredoxin h.Furthermore, the present study has clearly shown at thestructural level that thioredoxin h reduces disulphidesselectively. This is clear evidence that thioredoxin h recog-nises specific disulphide bonds and it provides a basis fordiscovering the mechanisms of target recognition. Cate-gorisation of a larger number of disulphide bonds into
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de

1644 K. Maeda et al. Proteomics 2005, 5, 1634–1644
targets and nontargets of thioredoxin h would provide atool to characterise the structural features required for tar-get recognition.
Mette Hersom Bien is gratefully acknowledged for technicalassistance, Sejet Plantbreeding (Sejet, Denmark) for seed mate-rial. We thank Prof. J.-P. Jacquot (INRA, Nancy) for purifiedA. thaliana NTR, and Karen Skriver (University of Copenha-gen), Birgit Bønsager, Kristian Sass Bak-Jensen and TorbenSteenholdt (Carlsberg Laboratory) for helpful discussions. K.M.was supported by a scholarship from Carlsbergs Mindelegat forBrygger J.C. Jacobsen. The project was supported by the DanishResearch Agency’s SUE programme (samarbejde mellem sektor-forskning, universitet og erhverv) grant no. 9901194.
5 References
[1] Holmgren, A., Ann. Rev. Biochem. 1985, 54, 237–271.
[2] Schürmann, P., Jacquot, J. P., Annu. Rev. Plant Biol. 2000, 51,317–400.
[3] Kobrehel, K., Wong, J. H., Balogh, A., Kiss, F. et al., PlantPhysiol. 1992, 99, 919–924.
[4] Wong, J. H., Kim, Y. B., Ren, P. H., Cai, N. et al., Proc. Natl.Acad. Sci. USA 2002, 99, 16325–16330.
[5] Marx, C., Wong, J. H., Buchanan, B. B., Planta 2003, 216, 454–460.
[6] Yano, H., Wong, J. H., Lee, Y. M., Cho, M. J., Buchanan, B. B.,Proc. Natl. Acad. Sci. USA 2001, 98, 4797–4799.
[7] Maeda, K., Finnie, C., Østergaard, O., Svensson, B., Eur. J.Biochem. 2003, 270, 2633–2643.
[8] Maeda, K., Finnie, C., Svensson, B., Biochem. J. 2004, 378,497–507.
[9] Østergaard, O., Melchior, S., Roepstorff, P., Svensson, B.,Proteomics 2002, 2, 733–739.
[10] Finnie, C., Melchior, S., Roepstorff, P., Svensson, B., PlantPhysiol. 2002, 129, 1308–1319.
[11] Bak-Jensen, K. S., Laugesen, S., Roepstorff, P., Svensson, B.,Proteomics 2004, 4, 728–742.
[12] Østergaard, O., Finnie, C., Laugesen, S., Roepstorff, P.,Svensson, B., Proteomics 2004, 4, 2437–2447.
[13] Kobrehel, K., Yee, B. C., Buchanan, B. B., J. Biol. Chem. 1991,266, 16135–16140.
[14] Jiao, J., Yee, B. C., Wong, J. H., Kobrehel, K., Buchanan, B.B., Plant Physiol. Biochem. 1993, 31, 799–804.
[15] Issakidis, E., Miginiac-Mascow, M., Decottignies, P., JacquotJ. P. et al., J. Biol. Chem. 1992, 267, 21577–21583.
[16] Jacquot, J. P., Lopez-Jaramillo, J., Miginiac-Maslow, M.,Lemaire, S. et al., FEBS Lett. 1997, 401, 43–47.
[17] Motohashi, K., Koyama, F., Nakanishi, Y., Ueoka-Nakanishi,H., Hisabori, T., J. Biol. Chem. 2003, 278, 31848–31852.
[18] Yip, T. T., Hunchens, T. W., FEBS Lett. 1992, 308, 149–153.
[19] Jacquot, J. P., Rivera-Madrid, R., Marinho, P., Kollarova, M.et al., J. Mol. Biol. 1994, 235, 1357–1363.
[20] Bønsager, B. C., Prætorius-Ibba, M., Nielsen, P. K., Svensson,B., Protein Expr. Purif. 2003, 30, 185–193.
[21] Rabilloud, T., Charmont, S., in: Rabilloud, T. (Ed.), ProteomeResearch: Two-Dimensional Gel Electrophoresis and Identi-fication Methods, Springer Verlag, Berlin, Heidelberg 2000,pp. 107–126.
[22] Shevchenko, A., Wilm, M., Vorm, O., Mann, M., Anal. Chem.1996, 68, 850–858.
[23] Gobom, J., Nordhoff, E., Mirgorodskaya, E., Ekman, R.,Roepstorff, P., J. Mass Spectrom. 1999, 34, 105–116.
[24] Vallee, F., Kadziola, A., Bourne, Y., Juy, M. et al., Structure1998, 6, 649–659.
[25] Barber, D., Sanchez-Monge, R., Mendez, E., Lazaro, A. et al.,Biochem. Biophys. Acta 1986, 869, 115–118.
[26] Strobl, S., Muhlhahn, P., Bernstein, R., Wiltscheck, R. et al.,Biochemistry 1995, 34, 8281–8293.
[27] Oda, Y., Matusnaga, T., Fukuyama, K., Miyazaki, T., Mor-imoto, T., Biochemistry 1997, 44, 13503–13511.
[28] Mena, M., Sanchez-Monge, R., Gomez, L., Salcedo, G., Car-bonero, P., Plant Mol. Biol. 1992, 20, 451–458.
[29] Mundy, J., Svendsen, I., Hejgaard, J., Carlsberg Res. Com-mun. 1983, 48, 81–91.
[30] Nielsen, P. K., Bønsager, B. C., Berland, C. R., Sigurskjold, B.W., Svensson, B., Biochemistry 2003, 42, 1478–1487.
[31] Eklund, H., Gleason, F. K., Holmgren, A., Proteins 1991, 11,13–28.
[32] Mora-Garcia, S., Rodriguez-Suarez, R., Wolosiuk, R. A., J.Biol. Chem. 1998, 273, 16273–16280.
[33] Garcia-Maroto, F., Carbonero, P., Garcia-Olmedo, F., PlantMol. Biol. 1991, 17, 1005–1111.
[34] Takase, K., Biochemistry 1994, 33, 7925–7930.
[35] Franco, O. L., Rigden, D. J., Melo, F. R., Block, C. Jr. et al., Eur.J. Biochem. 2000, 267, 2166–2173.
[36] Lazaro, A., Sanchez-Monge, R., Salcedo, G., Paz-Ares, J. etal., Eur. J. Biochem. 1988, 172, 129–134.
[37] Sanchez-Monge, R., Gomez, L., Garcia-Olmedo, F., Salcedo,G. A., FEBS Lett. 1986, 207, 105–109.
[38] Bønsager, B. C., Nielson, P. K., Abou Hachem, M., Fukada, K.et al., J. Biol. Chem. 2005, in press.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.de