identification of proximate toxicant for ethylene glycol developmental toxicity using rat whole...
TRANSCRIPT

TERATOLOGY 53:38-46 (1996)
Identification of Proximate Toxicant for Ethylene Glycol Developmental Toxicity Using Rat Whole Embryo Culture EDWARD W. CARNEY, ASHLEY B. LIBERACKI, MICHAEL J. BARTELS, AND WILLIAM J. BRESLIN Toxicology Research Laboratory, Health and Environmental Sciences, Dow Chemical Company, Midland, Michigan 48674
ABSTRACT The effects of ethylene glycol (EG) and its metabolite, glycolic acid (GA), were compared by culturing day 10.5 rat conceptuses for 46 h in media containing 0.5,2.5, 12.5,25 or 50 mM EG or GA. EG up to 50 mM was essentially without effect, whereas 5 12.5 mM GA inhibited embryo growth and development. Craniofacial dysmorphogenesis was observed in 70% of the 12.5 mM GA embryos (0% in controls). To deter- mine if GA toxicity in vitro was an indirect effect of medium acidification, embryos were cultured in 12.5 mM GA (pH 6.7), 12.5 mM sodium glycolate (pH 7.4), or in control medium (pH 7.4 or 6.7). The percentage of dysmorphic embryos was 67% for the 12.5 mM GA (pH 6.7) group, 58% for the so- dium glycolate (pH 7.4) group, 8% in the pH 6.7 controls, and 0% in the pH 7.4 controls. These re- sults suggest that GA, not parent EG, is the active toxicant for EG-induced developmental toxicity and that acidification of culture medium pH plays only a minor role in GA's effects in vitro. The iden- tification of GA as the active toxicant is important for the risk assessment of EG because GA exhibits dose-rate-dependent, nonlinear kinetics in vivo. 0 1996 Wiley-Liss, Inc.
Ethylene glycol (EG) is a major industrial chemical, with current annual production estimated at almost five billion pounds (U.S. International Trade Commis- sion, June 26,1992). The largest single use for EG is in the production of polyethylene terephthalate for poly- ester fibers, containers, and films. Numerous other ap- plications have capitalized on EGs versatile thermal and solubility characteristics; hence its use in automo- tive coolants, heat transfer fluids, aircraft and runway deicers, and as a solvent in the manufacture of paints, plastics and inks.
EG first became prominent in the toxicology litera- ture in the 1930s following a number of fatal ingestions in humans, and over the years, an extensive data base concerning EG systemic toxicity in animals and hu- mans has been amassed (reviewed by Winek et al., '78; Rowe and Wolf, '82; Jacobsen and McMartin, '86; Hall,
'92). This work has shown that EGs most serious symp- toms are caused primarily by an acidic metabolite of EG, glycolic acid (GA), which at high doses induces a metabolic acidosis leading to renal failure and other toxic sequelae (Jacobsen et al., '84; Hewlett et al., '89).
Although to the best of our knowledge there have been no reports of EG-induced developmental toxicity in humans, studies in rats and mice have shown that EG can induce terata, such as cleft palate, cleft lip, and axial skeleton defects, when given in high doses by the gavage route (Price et al., '85; Yin et al., '86; Neeper- Bradley et al., '95). Based on knowledge about mecha- nisms of EG systemic toxicity, a number of investiga- tions as to EGs mode of action in causing developmental toxicity have ensued. Khera ('91) explored the relation- ship between maternal metabolic acidosis and EG-in- duced teratogenicity by treating gestation day 11 rats with 3,333 mg/kg of EG (injected subcutaneously) and then giving loading and maintenance doses of sodium bicarbonate in an attempt to counteract the acidosis. This treatment significantly, but not completely, main- tained blood pH, PC02 and associated acid/base param- eters and resulted in the amelioration of many, but not all, developmental effects. This suggested that meta- bolic acidosis was responsible for at least some of the developmental effects following high dose EG exposure. Khera ('91) also documented a severe maternal serum hyperosmolality in this study and speculated that this, too, might be a contributing factor to EGs developmen- tal effects.
As opposed to the maternally mediated mechanisms suggested by Khera ('911, Grafton and Hansen ('87) proposed that parent EG acts directly on embryos. This conclusion was based on the results of their whole em- bryo culture study in which gestation day 10.5 embryos exposed to 0.53 or 0.71 molar EG exhibited abnormal development. However, the concentrations of EG tested
Received September 21, 1995; accepted November 10, 1995.
Address reprint requests to Edward W. Carney, Chemical Hazard Evaluation & Communication, 1803 Building, Health and Environ- mental Sciences, The Dow Chemical Co., Midland, MI 48674.
0 1996 WILEY-LISS, INC.

ETHYLENE GLYCOL DEVELOPMENTAL TOXICITY 39
greatly exceeded lethal blood EG concentrations in ro- dents and humans (Hewlett et al., '89; Jacobsen et al., '84), making the relevance of the in vitro exposures uncertain. Finally, the possibility that the glycolate anion exerts a direct chemical toxicity (i.e., not a sec- ondary effect of metabolic acidosis), should not be over- looked (Jacobsen et al., '84; Carney '94).
The present study used a rat gestation day 10.5 whole embryo culture system in an attempt to discrim- inate between the following as the most likely proxi- mate teratogen: (1) parent EG, (2) GA acting indirectly via acidic pH, (3) a direct chemical toxicity of the gly- colate anion, or (4) hyperosmolality. The whole embryo culture system was particularly well suited for this ob- jective as the confounding effects of maternal metabo- lism of parent EG were avoided and the osmolality and pH of the in vitro environment could be easily moni- tored and/or adjusted. However, a deliberate effort was made to choose test material concentrations that re- flected plausible in vivo levels of EG and GA in order to ensure the relevance of the data for risk assessment.
MATERIALS AND METHODS Test materials
EG and GA were obtained from Aldrich Chemical Co. (Milwaukee, WI) and sodium glycolate (NaG) was supplied by Pfaltz and Bauer (Waterbury, CT). Sodium valproate (VPA) was used as a positive control agent and was obtained from Sigma Chemical Co. (St. Louis, MO). All test materials were at least 98% pure, based on information provided by the suppliers.
Animals Adult female time-mated CD (Sprague-Dawley de-
rived) rats were obtained from Charles River Breeding Laboratory (Portage, MI). The day on which a copula- tion plug was observed was designated gestation day 0.5. The rats were housed singly in rooms designed to maintain humidity at - 40-60%, temperature a t - 22"C, photoperiod at 12 L:12 D (lights on at 7:OO a.m.).
Whole embryo culture Dissection and culture of embryos generally were
conducted as described by Cockroft ('90). Briefly, day 10.5 conceptuses (day of sperm-positive vaginal smear or copulation plug designated day 0.5) were dissected free of decidual tissue and Reichert's membrane, leav- ing the visceral yolk sac and ectoplacental cone intact. Day 10.5 was chosen to correspond with previously de- scribed whole embryo culture (Grafton and Hansen, '87) and single-dose teratology/maternal homeostasis (Khera, '91) studies. Early somite stage embryos were then transferred to culture bottles (2 embryos/bottle) containing pre-warmed, pre-gassed test or control cul- ture media. The culture bottles were maintained in a continuous gas flow rotating culture unit (BTC Engi- neering, Cambridge, UK) at 37°C for 46 h.
The culture medium was composed of 75% immedi- ately centrifuged, heat-inactivated serum collected from adult rats (Cockroft, '90) and 25% Dulbecco's Mod- ified Eagle's Medium (D-MEM). Concentrated (100 x ) stock solutions of the test materials were prepared in D-MEM and added directly to the appropriate culture bottles a t a volume of 10 pl/ml to achieve the desired final concentrations.
Experimental design In Experiment 1, groups of 10 embryos were cultured
in media containing 0.5,2.5,12.5,25.0, or 50.0 mM/L of EG or equimolar concentrations of GA for 46 h. These levels were based on rat pharmacokinetic data (Frantz et al., '89; Hewlett et al., '89) and span the range of peak maternal blood concentrations following NOEL, LOEL, and teratogenic gavage doses. An additional group of 10 embryos was cultured with sodium valproate (1.0 mM) and served as positive controls (Kao et al., '81). In Ex- periment 2, groups of 12 embryos were cultured for 46 h in one of four media: (1) control medium at pH 7.41, (2) control medium titrated to pH 6.74 with HC1, (3) medium containing 12.5 mM GA (pH 6.741, or (4) me- dium containing 12.5 mM sodium glycolate (pH 7.42). The 12.5 mM concentration represented the lowest-ob- served-effect concentration (LOEC) from Experiment 1.
To minimize the influence of litter effects, embryos from the same donor rat were distributed as equally as possible amongst the treatment groups. Due to limita- tions in the number of embryos which could be dis- sected on a given day, five replicates of Experiment 1 and two replicates of Experiment 2 were necessary to achieve the desired number of embryos. All experimen- tal treatments were equally represented in each of these replicates.
End points Evaluation of development was essentially as de-
scribed by Brown ('90). Upon completion of the culture period, embryos were evaluated for the presence of a beating heart and an active visceral yolk sac circula- tion. Embryos with a beating heart were considered viable embryos. Morphology was evaluated using the Brown-Fabro scoring system (Brown and Fabro, '81) and was done in an investigator-blind fashion. This was accomplished by random coding of the embryo bot- tles prior to the start of culture by a second investigator not involved in morphological evaluation. The code was revealed only after all embryos had been examined. Growth was assessed by measurement of visceral yolk sac diameter, crown-rump length, and head length with the aid of a calibrated eyepiece reticle. Protein contents of embryo and visceral yolk sac were deter- mined by the Bradford method as described by Brown ('90) and modified for microtiter plate reader as de- scribed by Stark et al. ('87).

40 E.W. CARNEY ET AL. Statistical analysis
Continuous data were evaluated by Bartlett's test for equality of variances. Based on the outcome of Bart- lett's test, a parametric or nonparametric analysis of variance (ANOVA) was performed. If the ANOVA was significant, analysis by Dunnett's test or the Wilcoxon Rank-Sum test with Bonferroni's correction was per- formed, respectively. Nonviable embryos were ex- cluded from the statistical analyses for growth mea- sures and morphological score. Percentage data were analyzed using Fisher's exact test.
Analytical chemistry Representative samples of culture media from each
treatment group taken at the start and end of culture were analyzed for EG and GA to determine (1) if the targeted concentrations were achieved and (2) to verify that EG was not metabolized to GA by the embryos. Culture media samples were prepared for GCIMS anal- ysis of EG and GA as follows: weighed aliquots (15-50 pL) were diluted with 1.0 ml of an internal standard solution (53.6 pg D,-ethylene glycol and 55.2 pg 13C2- glycolic acid in acetone) and sonicated (15 sec) and vor- texed for 10 min at room temperature. Aliquots (50 pL) of 5M aq. NaOH were added to each sample, followed by sonication (10 sec). The samples were then deriva- tized (15 min vortex at 40°C) with 20 pL each of pen- tafluorobenzyl bromide and pentaf luorobenzoyl chlo- ride (Aldrich Chemical Co.). The derivatized samples were then diluted with 1.5 ml distilled water and the analytes extracted into 1 ml toluene (2 min vortex at room temperature). Corresponding matrix standards were prepared from control media, as above.
Quantitation of the derivatized analytes was per- formed on a Finnigan SSQ-710 mass spectrometer (Finnigan MAT Corp., San Jose, CAI, equipped with a Hewlett-Packard 5890 gas chromatograph and a 7673A autosampler (Hewlett-Packard Co., Avondale, PA). Separations were achieved with a DB-1701 fused silica capillary GC column (30m x 0.32 mm i.d. x 1.0 pm film; J&W Scientific, Folsom, CA). The gas chro- matographic conditions were as follows: helium carrier gas (15 p.s.i.g.) at a flow rate of - 1 ml min-1; gas chromatograph oven temperature programmed from 50°C (1 min initial hold) to 300" at 25"/min; injector and capillary transfer line at 275°C and 280"C, respec- tively; 1 pL autosampler injection (20 ml split). The mass spectrometer conditions (NCI) were: ion source temperature, 150°C; ionizing current, 0.2 mA; electron energy, 70 eV; methane reagent gas a t 9500 mtorr source pressure; selected ion monitoring at d z 269, 271, 451 and 454 (30 mseclionlscan).
pH stability The pH stability of the test material-containing me-
dia was monitored in the culture system using a Fisher Accumet (Model 815MP) pH meter. The temperature
and gas phase conditions used during culture were maintained during the pH determinations. In Experi- ment 1, pH was measured in mock cultures (without embryos) both at the start and after 46 h of incubation in the culture apparatus. In Experiment 2, the pH of media containing embryos was similarly measured to determine if the presence of embryos influenced pH under the conditions of the experiment.
Osmolality The osmolality of selected culture media samples
was measured using a freezing point depression os- mometer (Osmette A, Precision Systems, Natick, MA).
GLP compliance With the exception of the test material characteriza-
tions, which were conducted by commercial suppliers, all aspects of study conduct were audited and found to be in compliance with Good Laboratory Practice stan- dards (OECD, '82; FDA, '88; EPA, '90).
RESULTS Analytical chemistry
Analysis of medium samples taken at the start of culture verified that the test materiaVmedium prepa- ration methods were sufficient to achieve the targeted concentrations. Analysis of medium samples taken at the end of culture indicated that both test materials were stable in the medium throughout the 46-h culture period and that there was no detectable conversion of EG to GA (detection limit of - 0.16 mM) during this period (data not shown).
Embryo development: Experiment 1 Experiment 1 was designed to compare the effects of
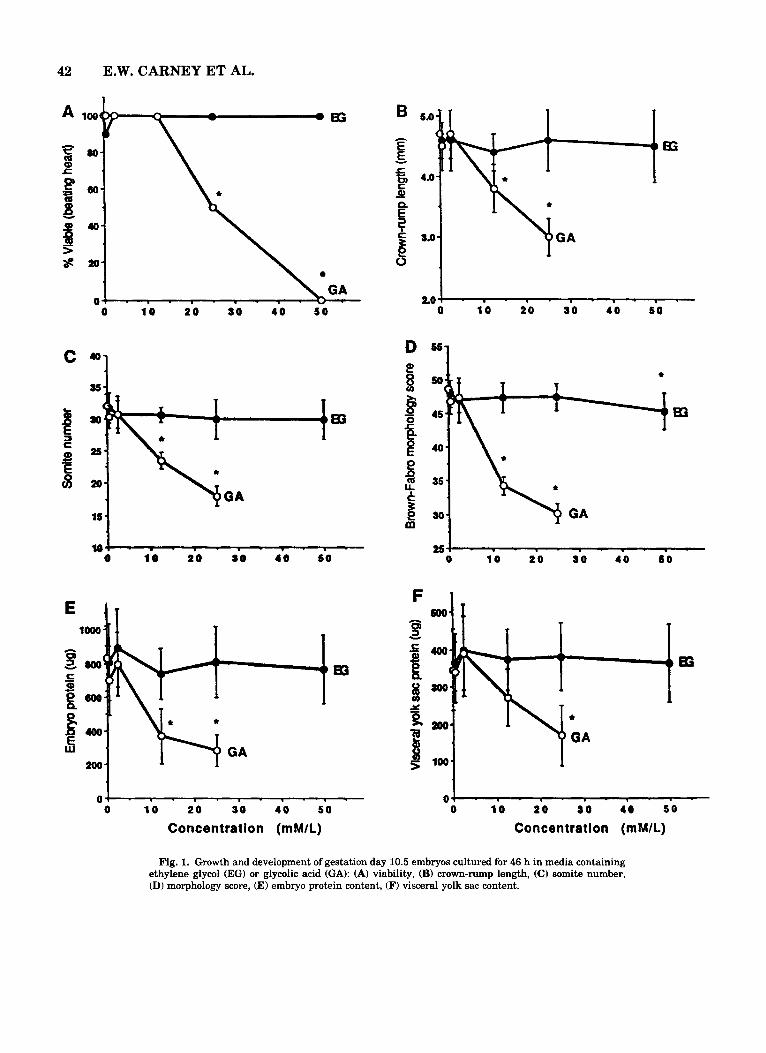
EG versus its metabolite, GA. There were no effects of EG at any concentration tested on the various mea- sures of embryonic growth and development, with the exception of a very slight decrease in morphological score for the 50 mM EG group (Fig. 1). GA at 0.5 and 2.5 mM also appeared to be without effect. However, culture of embryos in media containing 12.5 mM GA resulted in significant decreases in crown-rump length, head length (2.0 -C 0.2 mm vs. 2.4 * 0.2 mm for con- trols), embryo and visceral yolk sac protein content, somite number, and morphology score. Embryo viabil- ity was not significantly affected at this concentration, although higher concentrations were embryolethal. All embryos cultured with the positive control agent, VPA (1 mM), were viable. However, VPA exposure resulted in a significantly decreased mean crown-rump length (4.1 * 0.5 mm), head length (2.0 f 0.3 mm), and em- bryo protein content (506 & 150 pglembryo). Visceral yolk sac diameter (6.4 * 0.8 mm) and protein content were unaffected by VPA treatment, whereas somite numbers and morphology scores could not be accu-

ETHYLENE GLYCOL DEVELOPMENTAL TOXICITY 41
rately assessed due to morphological defects (see be- low).
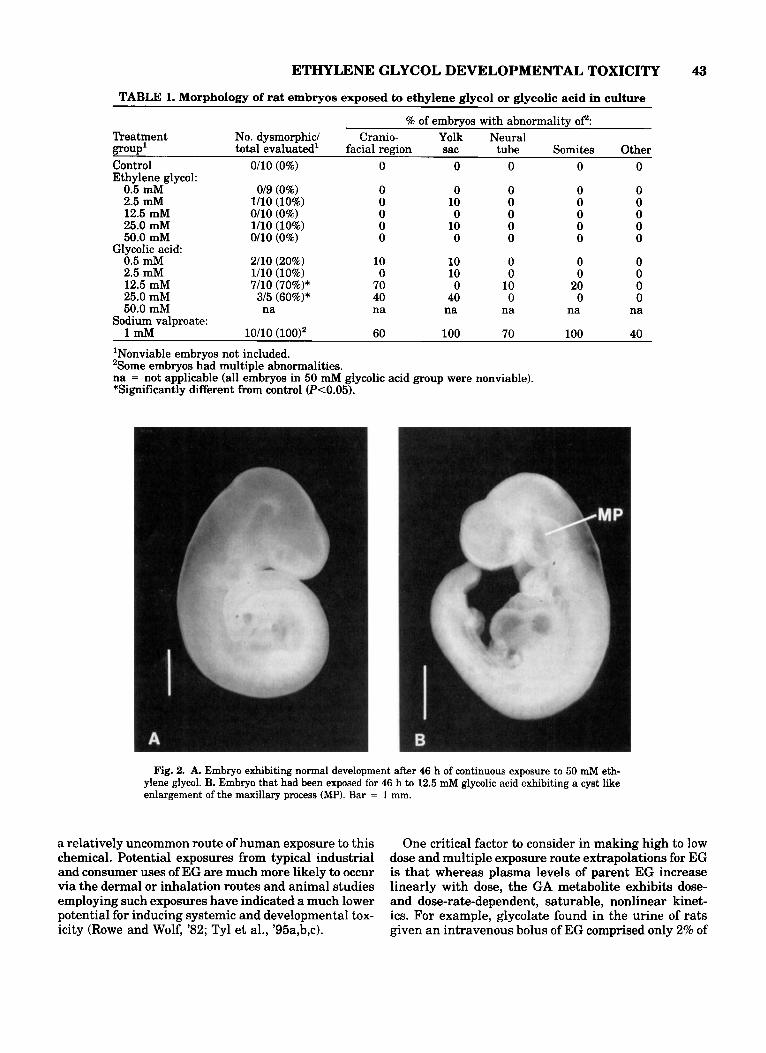
Morphologic examination revealed no evidence of dysmorphogenesis due to EG at concentrations up to and including 50 mM (Table 1). All EG-treated em- bryos appeared morphologically normal (Fig. 2A), with the exception of one conceptus in each of the 2.5 and 25 mM EG groups with a small hemorrhagic area on the surface of the visceral yolk sac (the embryo proper was unaffected). These areas are occasionally observed in control conceptuses in our laboratory. Therefore, the nature of these changes, their low incidence, and the lack of a dose response relationship indicated that these effects were spurious and unrelated to treatment.
In contrast, the percentage of dysmorphic embryos was significantly increased in the GA 12.5 and 25.0 mM groups (Table 1). The most commonly observed morphological alteration was a striking, bilateral, cystlike enlargement of the maxillary process, which was present in seven of 10 embryos in the 12.5 GA group (Fig. 2B). Also affected by exposure to 12.5 mM GA were several rostro-medial structures of the mid- facial region. These included an apparent absence of olfactory rudiments, irregular protrusions extending from an area just above the expected region of the nasal process, and hypoplastic andlor widely spaced telen- cephalic hemispheres. Other effects seen in a smaller number of embryos cultured with GA included dysmor- phogenesis of the optic vesicles and an irregular pat- tern of somite segmentation. Similar effects were ob- served in the few embryos surviving exposure to 25.0 mM GA. In addition, a disorganized pattern of visceral yolk sac vessels was also observed in the latter em- bryos.
Exposure of embryos to the positive control agent, W A , resulted in abnormal somite segmentation, al- tered neural tube development characterized by a wavy or kinked neural suture line, incomplete closure of the otic vesicles, disorganized vitelline (visceral yolk sac) vessels and other abnormalities (Table 1). The effects of VPA on embryo growth and morphology were consis- tent with those reported previously (Kao et al., '81).
Embryo development: Experiment 2 Experiment 2 was designed to determine the roles of
acidic pH and the glycolate anion in GA's toxicity in culture. Embryo viability was not affected by any of the treatments; however, the percentage of embryos with active yolk sac circulation was slightly, but not signif- icantly decreased in both the 12.5 mM GA (pH 6.74) and acidified medium (pH 6.74) groups (Table 2). All embryos in the sodium glycolate at pH 7.42 group had an active visceral yolk sac circulation.
The effects observed on the growth of embryos cul- tured in 12.5 mM GA (pH 6.74) were virtually identical to those observed in Experiment 1, with all parameters significantly different from pH 7.41 controls (Table 2). These same measures were also affected by culture
with 12.5 mM sodium glycolate a t pH 7.42, although to a slightly lesser degree. In contrast, only the protein contents of embryo and visceral yolk sac and embryo head length were significantly affected in embryos cul- tured in control medium titrated to pH 6.74, The de- gree to which the latter parameters was affected was also less in the pH 6.74 control group than in the GA at pH 6.74 or sodium glycolate a t pH 7.42 groups.
Both treatment with 12.5 mM GA at pH 6.74 and 12.5 mM sodium glycolate at pH 7.42 resulted in a significant increase in the percentage of dysmorphic embryos (Table 3). The type of alterations in the latter two groups were similar and, as seen in Experiment 1, were predominantly craniofacial. Again, the maxillary process and midfacial region were the principle sites affected. Only one embryo in the pH 6.74 control group exhibited dysmorphogenesis, which consisted of exces- sive protrusions of the telencephalic hemispheres and an irregularly shaped otic vesicle.
pH of culture media In Experiment 1, the pH of representative media
samples (without embryos) was monitored at the start and completion of 46 h of culture. Control medium, all EG media and the GA 0.5 and 2.5 mM media all were in the physiological range (pH 7.31-7.52) at the start of culture and remained relatively stable (0-0.17 pH units change) during the 46-h culture period. The pH of GA-containing media at the start of culture was 6.67, 5.90, and 4.31 for the 12.5, 25.0, and 50.0 mM groups, respectively, and was 0.59-0.78 units higher at the end of culture.
In Experiment 2, pH changes were monitored in rep- resentative cultures containing embryos. Relatively slight changes (5 0.24 units) in pH occurred in the 12.5 mM GA (initial pH 6.74), 12.5 mM sodium glycolate (initial pH 7.42), and acidic control groups (initial pH 6.74). However, pH decreased by 0.5 units in the pH 7.41 control group.
Osmolality To gain perspective on the role of hyperosmolality in
EG-induced developmental toxicity, the osmolality of selected media samples was determined. Osmolalities of 299,418,396, and 377 mOsmol/kg H,O were noted for the control, 50 mM EG, 12.5 mM GA, and 12.5 mM sodium glycolate media, respectively. Differences be- tween these values for the serum-based media used in this study vs. theoretical values predicted for dilute aqueous solutions are likely due to interactions among serum components as well as pH influences on the ion- ization of such components.
DISCUSSION Although the potential of EG to induce teratogenic-
ity in rats and mice given high oral doses is not in question, it is important to recognize that ingestion is
42 E.W. CARNEY ET AL.
1
1
2 0 GA
0 0 10 2 0 1 0 4 0 S O 0 10 2 0 90 4 0 5 0
I 0 J . m . I . I - - . - .
0 10 2 0 SO 40 5 0
Concentration (mMIL)
5
254 - . - . - . - . - . . 10 2 0 10 4 0 8 0 0
1 0 4 - - - - . - - . . . .
0 10 2 0 90 4 0 5 0
Con c e n t ra t i on (m MIL)
Fig. 1. Growth and development of gestation day 10.5 embryos cultured for 46 h in media containing ethylene glycol (EG) or glycolic acid (GA): (A) viability, (B) crown-rump length, (C) somite number, (D) morphology score, (E) embryo protein content, (F) visceral yolk sac content.
ETHYLENE GLYCOL DEVELOPMENTAL TOXICITY 43
TABLE 1. Morphology of rat embryos exposed to ethylene glycol or glycolic acid in culture
% of embrvos with abnormalitv OF: Treatment No. dysmorphic/ Cranio- Yolk Neural POUP' total evaluated' facial region sac tube Somites Other Control Ethylene glycol:
0.5 mM 2.5 mM 12.5 mM 25.0 mM 50.0 mM
Glycolic acid: 0.5 mM 2.5 mM 12.5 mM 25.0 mM 50.0 mM
0110 (0%)
019 (0%) 1/10 (10%) 0/10 (0%) 1/10 (10%) 0110 (0%)
2/10 (20%) 1/10 (10%) 7/10 (70%)* 3/5 (60%)* na
0
0 0 0 0 0
10 0 70 40 na
0
0 10 0 10 0
10 10 0 40
na
0
0 0 10 0
na
0
0 0 0 0 0
0 0
20 0
na
0
0 0 0 0
na Sodium valproate:
'Nonviable embryos not included. *Some embryos had multiple abnormalities. na = not applicable (all embryos in 50 mM glycolic acid group were nonviable). *Significantly different from control (k0.05).
1 mM 10/10 (loo)* 60 100 70 100 40
Fig. 2. A. Embryo exhibiting normal development after 46 h of continuous exposure to 50 mM eth- ylene glycol. B. Embryo that had been exposed for 46 h to 12.5 mM glycolic acid exhibiting a cyst like enlargement of the maxillary process (MP). Bar = 1 mm.
a relatively uncommon route of human exposure to this chemical. Potential exposures from typical industrial and consumer uses of EG are much more likely to occur via the dermal or inhalation routes and animal studies employing such exposures have indicated a much lower potential for inducing systemic and developmental tox- icity (Rowe and Wolf, '82; Tyl et al., '95a,b,c).
One critical factor to consider in making high to low dose and multiple exposure route extrapolations for EG is that whereas plasma levels of parent EG increase linearly with dose, the GA metabolite exhibits dose- and dose-rate-dependent, saturable, nonlinear kinet- ics. For example, glycolate found in the urine of rats given an intravenous bolus of EG comprised only 2% of

44 E.W. CARNEY ET AL.
TABLE 2. Role of pH and glycolate anion in glycolic acid-induced inhibition of embryo growth and development
Treatment % VYS' CRL2 HL2 Embmo Somite Morph. diam. protein % VYS VYS
group1 viable circ. (mm) (mm) protein number score (mm) Contro1,pH 7.41 100 92 4.8 f 0.3 2.6 4 0.2 989 2 217 31.8 5 1.7 48.9 t 1.4 6.9 & 0.6 456 f 151 Glvcolic acid.
i2.5 mM, '
pH 6.74 100 58 3.6 f 0.5* 2.1 * 0.2* 399 4 149* 20.2 & 5.9* 41.0? 3.7* 5.8 & 0.9* 301 & 64* Sodium glycolate,
12.5 mM, pH 7.42 100 100 4.3 f 0.3* 2.2 4 0.2* 596 f 127* 25.8* & 4.0* 43.9 2 3.2* 6.1 2 0.6* 330 f 87*
Control, pH 6.74 100 75 4.5 ? 0.3 2.3 f 0.2* 771 4 86* 30.5 f 1.6 47.9 k 2.3 6.6 ? 0.5 345 & 103*
'Twelve embryos per treatment group; pH at start of culture. VYS = visceral yolk sac; CRL = crown-rump length; HL = head length. *Statistically different from control (P<0.05).
TABLE 3. Role of pH and prlscolate anion in glycolic acid-induced dysmorphogenesis ~
% of embryos with abnormality of?
Threatment group' no. evaluated (%) facial region sac Other
Glycolic acid, 12.5 mM, pH 6.74 8/12 (67%)* 50 33 25
No. dysmorphicl Cranio- Yolk
Control, pH 7.41 0/12 (0%) 0 0 0
Sodium glycolate, 12.5 mM, pH 7.42 7/12 (58%)* 58 8 0 Control, pH 6.74 1/12 (8%) 8 0 0
'Twelve embryos per treatment group; pH at start of culture. 'Some embryos had multiple abnormalities. *Statistically different from control (P<0.05).
a 20 or 200 mg/kg dose, but 20% of a 1,000 or 2,000 mgkg dose (Marshall, '82). This shift occurs because oxidation of GA by glycolic acid oxidase is a rate-lim- iting step in the EG metabolism pathway (Jacobsen and McMartin, '86; Lenk et al., '89) and exhibits satu- ration kinetics at high substrate concentrations. Also, renal elimination of GA is likely to be saturable. In contrast, no evidence of metabolic saturation was found in mice and rats receiving 1,000 mglkg of EG applied to the skin or after exposure to EG aerosols at concentra- tions up to 184 mg/m3 (Marshall and Cheng, '83; Frantz et al., '89, '91). It should also be noted that whereas the percentage of EG converted to GA in- creases with dose, other EG metabolites (glycoalde- hyde, glyoxylate, oxalate) remain at a very low ((1%) and constant proportion across the 20-2,000 m g k g EG dose range (Marshall, '82).
A secondary consequence of metabolic saturation and GA accumulation is metabolic acidosis. That GA is the primary cause of this acidosis is indicated by the abundance of this metabolite and the inverse relation- ship between blood GA concentration and blood bicar- bonate levels following high dose EG exposure. Also, treatments that inhibit GA formation (e.g., 4-meth- ylpyrazole) are effective in preventing metabolic acido- sis and consequently most symptoms of acute toxicity (Clay and Murphy, '77; Jacobsen et al., '84; Hewlett et al., '89; Karlson-Stiber and Persson, '92). Clinical cases of severe human EG poisoning also show this relation-
ship between GA and metabolic acidosis, with blood glycolate concentrations of 17-29 mML, base excesses of -15--31 mM/L, and blood pHs as low as 6.7 (Ja- cobsen et al., '84). In contrast, there was no evidence of any aciabase disturbances in human male volunteers exposed continuously for 30 days to EG aerosols at con- centrations up to 75 mg/m3 (Wills et al., '74).
Because GA accumulation and metabolic acidosis are likely to occur only in high dose exposure scenarios, it is critically important for human risk assessment to determine if they are responsible for EG's developmen- tal toxicity. If such were the case, then a nonlinear, physiologically based risk assessment model would be most appropriate. However, a causal role for parent EG would imply that more standard, linear extrapolations would be most appropriate in making risk assessments for this chemical.
Whole embryo culture was particularly well suited to evaluate the direct toxicity of parent EG due to the fact that isolated rat conceptuses (prior to gestation day 15) do not possess significant activities of alcohol dehydrogenase (ADH) and acetaldehyde dehydroge- nase (ALDH), the enzymes responsible for the oxida- tion of EG to GA. This lack of significant ADWALDH activity has been shown previously through enzymatic activity assay of ra t fetal liver and placenta (Sjoblom et al., '78), in rat whole embryo culture experiments com- paring the effects of parent glycol ether compounds vs. their oxidized metabolites (Yonemoto et al., '84) and

ETHYLENE GLYCOL DEVELOPMENTAL TOXICITY 45
was confirmed in this study using GCMS analysis for GA formed during culture. Thus the whole embryo cul- ture approach enabled the assessment of the direct ef- fects of EG without the confounding factor of parent compound biotransformation.
The results of this study clearly showed that parent EG has little effect on embryo development, even after 46 h of continuous exposure to extremely high EG con- centrations. The only treatment-related effect seen in embryos exposed to EG occurred at the highest concen- tration tested (50 mM) and consisted of a very slight decrease in morphology score. Based on published data describing morphology scores for in vivo developed Sprague-Dawley rat embryos (Brown, 'go), this de- creased score is equivalent to a developmental delay of - 2.5 hours relative to controls. This relatively small delay is considered insignificant in light of the very high concentration of EG to which the embryos were exposed. To put this concentration into perspective, it should be noted that rats given a teratogenic and ma- ternally toxic dose of 2,000 mg/kg of EG had a peak blood EG concentration of 31 mM (Hewlett et al., '89). In severely intoxicated human patients, the highest blood EG concentration observed was 41 mM (Jacobsen et al., '84).
The lack of direct EG toxicity refutes suggestions made in a previous rat whole embryo culture study (Grafton and Hansen, '87) that parent EG may be di- rectly embryotoxic. In that study, day 10.5 embryos were exposed to 530 or 710 mM EG, concentrations greatly exceeding those associated with even lethal doses of EG (Jacobsen et al., '84). Not surprisingly, these in vitro exposures adversely affected embryo growth and development, possibly by an indirect sol- vent effect on the embryos. However, many of the ef- fects seen in that study, such as absent hind limb bud, do not correspond with EGs effects in vivo and proba- bly reflect delayed development rather than a frank teratogenic effect. Thus it is highly unlikely that par- ent EG is responsible for any of EGs developmental effects under plausible exposure scenarios.
To the contrary, the weak acid metabolite, GA, pro- duced dysmorphogenesis in a pattern corresponding to malformations in vivo and at concentrations reflecting blood levels associated with teratogenic doses of EG (Frantz et al., '89; Hewlett et al., '89). Specifically, GA at B 12.5 mM/L caused abnormalities of the developing maxillary process, midfacial region, optic vesicles, and somites (Fig. 2B), effects that appear to be embryolog- ically consistent with cleft palate and lip, micro- gnathia, anopthalmia, and axial skeleton malforma- tions observed following large oral EG doses in vivo (Price et al., '85). The maxillary process lesion was par- ticularly striking and was the most commonly observed morphological abnormality in this group.
The results of Experiment 2 indicated that the effects of GA in culture were due in large part to an inherent toxicity of the glycolate anion, rather than an indirect
effect due to acidification of pH. This was shown by the fact that sodium glycolate (12.5 mM) at pH 7.4 was nearly as potent and caused similar morphological al- terations as did 12.5 mM GA at pH 6.7. A direct effect of the glycolate anion could explain why treatment of metabolic acidosis with sodium bicarbonate infusion was only partially effective in ameliorating EG-in- duced developmental toxicity (Khera, '91), as plasma glycolate levels would be expected to remain high even with this trcatment.
Lesser, but statistically significant effects of control medium titrated with HC1 to pH 6.7 indicated a small, additive contribution of medium acidity. The relatively minor effects of acidic culture medium are consistent with the results of a similar rat whole embryo culture study examining pH/formate interactions (Andrews et al., '93). Although both studies showed that acidic pH per se does not have a major direct effect on embryos, they do not necessarily contradict a causal role for met- abolic acidosis in vivo. Metabolic acidosis is a complex condition involving large decreases in PCOz and bicar- bonate and significant increases in glucose and lactate among others (Khera, '91). Because it is not possible to replicate all of these dynamic changes in an embryo culture system, simply acidifying the medium with HC1 should not be considered an adequate representation of metabolic acidosis in vivo. Metabolic acidosis could still adversely affect development through changes in PC02, bicarbonate, glucose, etc. Alternatively, metabolic ac- idosis could be acting as a modulator of toxicity by shifting the ionization and consequent distribution of glycolate to the embryo.
It is also interesting to note that culture medium containing the highest concentration of EG (50 mM) was severely hyperosmotic (418 mOsmol/kg H20), yet this did not adversely affect embryo development. In vivo, serum osmolality rose from a normal value of 285 mosmolkg H20 to a maximum of 359 mosmolkg H20 following a subcutaneous dose of 3,333 mgkg of EG (Khera, '91). The resistance of cultured embryos to hy- perosmotic serum suggests that maternal hyperosmo- lality does not have a direct effect on the conceptus. This also implies that the visceral yolk sac enclosing the cultured embryos can maintain a substantial os- motic gradient and protect embryos from osmotic chal- lenge.
In conclusion, this study provided direct evidence that GA is the proximate toxicant for EG-induced de- velopment toxicity in rats and that parent EG is essen- tially nonembryotoxic even at concentrations exceed- ing those that would be lethal to the mother. The identification of GA as the developmentally toxic me- tabolite of EG is critically important for risk assess- ment because the kinetics of this metabolite are satu- ration-dependent and nonlinear. Thus relatively high doses and/or dose-rates would appear to be required to induce developmental toxicity. Integration of the present results with previous in vivo homeostasis find-

46 E.W. CARNEY ET AL.
ings (Khera, '91) indicate that following high doses of EG, the GA metabolite can act both through an inher- ent toxicity of the glycolate anion as well as via induc- tion of metabolic acidosis. However, the relative con- tributions of each of these factors during in vivo exposures to EG remains to be elucidated.
ACKNOWLEDGMENTS Technical assistance was provided by J. Hammond,
S. Close, P. Bush, and M. Matthews. A. Hardy assisted in the preparation of this manuscript. The authors are also grateful to T. McNutt and C. Harris (Univ. of Michigan) for providing details of their whole embryo culture methods. Scientific input and financial support for this study were provided by the Chemical Manufac- turer's Association (Ethylene Glycol Toxicology Re- search Task Group and Ethylene Glycol Panel).
LITERATURE CITED Andrews, J.E., M. Ebron-McCoy, R.J. Kavlock, and J.M. Rogers
(1993) Lowering pH increases embryonic sensitivity to formate in whole embryo culture. Toxicol. In Vitro 7:757-762.
Brown, N.A. and Fabro, S. (1981) Quantitation of rat embryonic de- velopment in vitro: A morphological scoring system. Teratology 24:
Brown, N.A. (1990) Routine assessment of morphology and growth: scoring systems and measurements of size. In: Postimplantation Mammalian Embryos: A Practical Approach. A.J. Copp and D.L. Cockroft, eds. Oxford University Press, New York, pp. 93-108.
Carney, E.W. (1994) An integrated perspective on the developmental toxicity of ethylene glycol. Reprod. Toxicol. 8:99-113.
Clay, K.L. and R.C. Murphy (1977) On the metabolic acidosis of eth- ylene glycol intoxication. Toxicol. Appl. Pharmacol. 39:39-49.
Coekroft, D.L. (1990) Dissection and culture of postimplantation em- bryos. In: Postimplantation Mammalian Embryos: A Practical Ap- proach. A.J. Copp and D.L. Cockroft, ed~. Oxford University Press, New York, pp. 15-40.
EPA (1990) Environmental Protection Agency Toxic Substances Con- trol Ad; Good Laboratory Practice Standards, 40 CFR Part 792.
FDA (1988) United States Food and Drug Administration: Good Lab- oratory Practice Regulations for NonClinical Studies.
Frantz, S.W., J.L. Beskitt, C.M. Grosse, C.B. Jensen, and M.J. Tallant (1989) Ethylene glycol: Comparison of pharmacokinetica and mate- rial balance following single intravenous, oral and cutaneous ad- ministrations to male and female Sprague-Dawley Rats. Chemical Manufacturers Association Project Report 51-543.
Frantz, S.W., J.P. Van Miller, and F.R. Frank (1991) Ethylene glycol: comparisons of pharmacokinetic and material balance studies fol- lowing single intravenous, peroral and percutaneous administra- tions to female CD (Rbl mice. Chemical Manufacturers Association Project Report 53-550.
Grafton, T.F. and D.K. Hansen (1987) In vitro embryotoxic effects of ethylene glycol in rats. Teratogenesis Carcinog Mutagen 7.483- 489.
Hall, A. (1992) Acute ethylene glycol poisoning. Emergency Medicine Reports. 13:29-38.
Hewlett, T.P., D. Jacobsen, T.D. Collins, and K.E. McMartin (1989) Ethylene glycol and glycolate kinetics in rats and dogs. Vet. Human Toxicol. 31:116-120.
Jacobsen, D. and K.E. McMartin (1986) Methanol and ethylene glycol poisonings: mechanism of toxicity, clinical course. diagnosis and
65-78.
Jacobeen, D., S. Ovrebo, J. Ostborg, and O.M. Serjersted (1984) Gly- colate causes the acidosis in ethylene glycol poisoning and is effec- tively removed by hemodialysis. Acta. Med. Scand. 21 13409-416.
Kao, J., N.A. Brown, B. Schmid, E.H. Goulding, and S. Fabro (1981) Teratogenicity of valproic acid in vivo and in vitro investigations. Teratogenesis Carcinogen. Mutagen 1 :367-382.
Karlson-Stiber, C. and H. Persson (1992) Ethylene glycol poisoning: Experiences from an epidemic in Sweden. Clin. Toxicol. 30:565- 574.
Khera, K.S. (1991) Chemically induced alterations in maternal ho- meostasis and histology of conceptus: Their etiologic significance in rat fetal anomalies. Teratology 44:259-297.
Lenk, W., D. hh r , and J . Sonnenbichler (1989) Pharmacokinetics and biotransformation of diethylene glycol and ethylene glycol in the rat. Xenbiotica 19361-979.
Marshall, T.C. (1982) Dose-dependent disposition of ethylene glycol in the rat after intravenous administration. J. Toxicol. Environ. Hlth.
Marshall, T.C. and Y.S. Cheng (1983) Deposition and fate of inhaled ethylene glycol vapor and condensation aerosol in the rat. Fundam. Appl. Toxicol. 3:175-181.
Neeper-Bradley, T.L., R.W. 5 1 , L.C. Fisher, M.F. Kubena, M.A. Vr- banic, and P.E. h o (1990) Determination of a no-observed-effect- level for developmental toxicity of ethylene glycol administered by gavage to CD rats abd CD-1 mice. Fundam. Appl. Toxicol. 27:121- 130.
OECD (1982) Organization for Economic Co-Operation and Develop ment: Principles of Good Laboratory Practice. Paris: ISBN 92-64-
Price, C.J., C.A. Kimmel, R.W. 5 1 , and M.C. Marr (1985) The devel- opmental toxicity of ethylene glycol in rats and mice. Toxicol. Appl. Pharmacol. 81: 113 -127.
Rowe, V.K. and M.A. Wolf (1982) Glycols. In: Industrial Hygiene and Toxicology, Vol. 2c, 3rd ed. F.C. Patty and G.D. Clayton, eds. John Wiley & Sons, New York, pp. 3817-3908.
Sjoblom, M., L. Pilstrom, and J. Morland (1978) Activity of alcohol dehydrogenase and acetaldehyde dehydrogenases in the liver and placenta during the development of the rat. Enzyme 23:108-115.
Stark, K.L., C. Harris, and M.R. Juchau (1987) Embryotoxicity elic- ited by inhibition of gamma-glutamyltransferase by acivicin and transferase antibodies in cultured rat embryos. Toxicol. Appl. Phar- macol. 97:548-560.
5 1 , R.W., B. Ballantyne, L.C. Fisher, D.L. Fait, T.A. Savine, D.E. Dodd, D.R. Klonne, and I.M. pritts (1995a) Evaluation of the de- velopmental toxicity af ethylene glycol aerosol in the CD rat and CD-1 mouse by whole-body exposure. Fundam. Appl. Toxicol. 24: 57-75.
5 1 , R.W., B. Ballantyne, L.C. Fisher, D.L. Fait, D.E. Dodd, D.R. Klonne, LM. Pritts, and P.E. Losco (1995b3 Evaluation of the de- velopmental toxicity of ethylene glycol aerosol in CD-1 mice by nose-only exposure. Fundam. Appl. Toxicol. 27:49-62.
5 1 , R.W., L.C. Fisher, M.F. Kubena, M.A. Vrbanic, and P.E. Losco (1995~) Assessment of the developmental toxicity of ethylene glycol applied cutaneously to CD-1 mice. Fundam. Appl. Toxicol. (in press).
Wills, J.H., F. Coulston, E.S. Hams, E.W. McChesney, J.C. Russell, and D.W. Serrone (1974) Inhalation of aerosolized ethylene glycol by man. Clin. Toxicol. 7:463-476.
Winek, C.L., D.P. Shingleton, and S.P. Shanor (1978) Ethylene and diethylene glycol toxicity. Clin. Toxicol. 13:297-324.
Yin, L., C. Liu, L. Shih and K. Po (1986) A study of the teratogenic action of ethylene glycol in rats. Chinese J. Prev. Med. 20:289-290.
Yonemoto, J., N.A. Brown, and M. Webb (1984) Effects of dimethoxy- ethyl phthalate, monomethoxyethyl phthalate, 2-methoxyethanol, and methoxyacetic acid on postimplantation rat embryos in culture.
10:397-409.
12367-9.
. I
ireatmeit. Med. Toxicol. 13309-334. Tox. Lett. 21 :97-102.