identification of goat mammary stem/progenitor cells … · still not as well documented compared...
TRANSCRIPT

1
Identification of goat mammary stem/progenitor cells Sonja Prpar1, Eugenio Martignani2, Peter Dovc3, Mario Baratta4
1,3 Department of Animal Science, Biotechnical Faculty, University of Ljubljana, 1230 Domzale, Slovenia, and 2,4 Department of Veterinary Morphophysiology, Faculty of Veterinary Medicine, University of Turin, 10095 Grugliasco (TO), Italy 1 supported by the Slovenian Research Agency (ARRS) (Research programme P4-0220), through the ARRS young researcher scheme and by a grant from Slovene Human Resources Development and Scholarship Fund for funding of cooperation of Slovene graduate students abroad.
2,4 supported by grants from Regione Piemonte and University of Turin.
4 Correspondence: M. Baratta, Department of Veterinary Morphophysiology, Faculty of Veterinary Medicine, University of Turin, 10095 Grugliasco (TO), Italy; e-mail: [email protected] ABSTRACT Goat mammary gland epithelial cells have been used to establish primary and permanent cell lines but to date there is no data on mammary stem cells in this species. The detection and characterization of goat mammary stem cells (gMaSCs) is an important task for a better understanding of the cyclic character of the mammary gland development which will also offer a potential for manipulation of lactation yield and persistency. The objective of the present study was to demonstrate that a subpopulation of MaSCs resides in the goat mammary gland. Mammary tissue from lactating Saanen goats (Capra hircus) was dissociated and processed to a single cell suspension. Using an in vitro colony-forming assay we demonstrated that distinct colony types, which expressed specific lineage markers, arose from unipotent progenitors. Using two different growth media we showed that the frequencies of caprine clonogenic progenitors differed according to growth conditions. Goat epithelial cells were transplanted under the kidney capsule of non-obese diabetic/severe combined immunodeficient (NOD/SCID) mice where they formed organized bilayered structures. Our results indicate the presence of MaSCs in the caprine mammary gland. These data represent the first description of the tissue hierarchy of the goat mammary gland and demonstrate the regenerative potential of goat adult mammary stem cells.
BOR Papers in Press. Published on January 11, 2012 as DOI:10.1095/biolreprod.111.095489
Copyright 2012 by The Society for the Study of Reproduction.

2
INTRODUCTION Adult stem cells are presumed to exist during the entire life cycle of mammary glands, where they are required to expand the cell populations during pregnancy and sustain cell turnover to replace senescent cells. The cellular processes involved in the development of the mammary gland in ruminants are still not as well documented compared to those in humans and rodents. Studies on rodent mammary glands have provided many basic biological insights into the development of this organ, but they do not adequately explain the broad range of specific mechanisms involved in mammary gland development among mammalian species. The morphological development of the mammary gland in humans and ruminants is similar [1], revealing the potential of ruminant species to serve as useful models for a better understanding of human breast development and cancer. Ruminant mammary development is driven by many of the same classic mammogenic hormones active in murine models, but the histological features differ from those in the mouse species. In ruminants, as in humans, stromal fat invasion and ductal branching are concomitant phenomena, while in the mouse extensive branching takes place after primary ducts have invaded the surrounding fat. Goat mammary gland epithelial cells have been used to establish primary and permanent cell lines [2] but to date there is no evidence for the existence of mammary gland stem cells (gMaSCs) in this species. Distinct epithelial subtypes have been isolated from mouse [3, 4], human [5, 6], and bovine [7] mammary glands. In prepubertal bovine mammary glands a population of small light cells (SLCs), that might function as stem cells, was described [8]. These pale-staining cells, also observed in goat mammary glands [9], are characterized by their clear cytoplasm, sparse cytoplasmic organelle distribution, large spherical nuclei and the presence of tight junctions between pale cells and adjacent epithelial cells [10]. Goat MaSCs have not been identified and characterized so far although several reasons make their isolation and characterization very important. From the economic perspective gMaSCs could be exploited in animal production to manipulate milk yield and attain milk persistency in lactation. During the last few decades, milk production per cow has increased steadily, because of genetic improvements achieved by crossbreeding and selection [11]. However the reproductive efficiency of dairy cows has dramatically decreased, mostly due to selection programmes, which did not take into consideration enough traits [12, 13]. In goat such an unwanted development could be prevented through the stimulation of the stem cell compartment which could be manipulated in several ways, with exogenous hormones or paracrine factors to alter cell numbers in order to allow breeders to enhance milk production without losing reproductive efficiency. The aim of our study was to identify through functional assays in vitro and in vivo stem/progenitor cells in goat mammary gland. We wanted to determine if we could identify normal, organ-specific stem cells or stem-like cells that contain the capacity to proliferate and differentiate into all lineages of the mammary gland. This would enable comparison of the tissue cell hierarchy among different mammalian species and lead to the generation of tools to assess the effects of different chemical compounds on ruminant mammary stem cell homeostasis and self-renewal which are key factors in controlling the persistency of lactation. We hypothesize that stem cells play a key role in maintaining high levels of milk production

3
and achieving a higher proportion of these cells in the mammary gland is a reasonable perspective, based on murine models [14]. MATERIALS AND METHODS Animals Female NOD/SCID mice were bred and housed at the animal facility of the Faculty of Veterinary Medicine of the University of Turin according to the procedure and guidelines approved by the Italian Ministry of Health. The mice were used at 8 – 14 weeks of age. Mammary tissue was obtained at the slaughterhouse from three lactating Saanen goats. Goats 1 and 2 were three years old and goat 3 was 7 years old. A 7 month-old Saanen goat (goat 4) was purchased from a breeder and slaughtered at the Department of Animal Science of the Biotechnical Faculty, of the University of Ljubljana. Sample collection was performed with the authorization and under the supervision of representatives of the Veterinary Services of the Slovenian National Health Service branch of the Ministry of Health. Caprine mammary tissue Whole udders were collected from lactating goats of different ages at local abattoirs 30 minutes to 3 hours after slaughter. Sample collection was performed with the authorization and under the supervision of representatives of the Veterinary Services of the Italian National Health Service branch of the Ministry of Health. A piece of tissue was dissected out of the area surrounding the teats and then minced with scalpels or scissors. From each goat udder approximately 10 – 15 grams of tissue was recovered and processed to an epithelium-rich pellet (organoids), which was cryopreserved, as described for human breast samples by Eirew et al. [6]. When needed, an aliquot was processed to a single cell suspension, as described by Eirew [6]. In vitro Colony-Forming Cell (CFC) assay 60 mm tissue culture dishes were coated with rat type 1 collagen solution (BD Bioscience) diluted in phosphate buffered saline (PBS, 1x) and incubated at 37 °C for one hour. Goat mammary cells were mixed with 2x105 mouse 3T3 fibroblasts which were previously treated with 10 µg/ml mitomycin C (Sigma-Aldrich) for 2 hours. Media used were human EpiCult-B (STEMCELL Technologies) supplemented with 5% FBS, 10-6 M hydrocortisone (Sigma-Aldrich), 100 U/ml penicillin, and 100 µg/ml streptomycin and Sf7 (DMEM/F12 supplemented with 5% FBS, 2 mM glutamine, 0.1% w/v bovine serum albumin (BSA), 10 ng/ml EGF, 10 ng/ml cholera toxin, 1 µg/ml insulin, 0.5 µg/ml hydrocortisone, 100 U/ml penicillin, 100 µg/ml streptomycin (all from Sigma-Aldrich)). The assay dishes were incubated at 37°C with 5% CO2 for 24 hours, the medium was then changed to fresh medium with the same composition but without FBS. Cell cultures were incubated for another 6 – 8 days and were then fixed rapidly (30 seconds – 1 minute) with acetone/methanol (1:1 v:v, Fluka) and either stained with crystal violet (50 mg of crystal violet in water solution containing 20% methanol) or used for immunofluorescence staining, as previously described [7]. Primary antibodies were anti-human cytokeratin 14 (KRT14, also known as CK14) (1:500 dilution, polyclonal AF-64, Covance) and cytokeratin 18 (KRT18, also known as CK18) (1:200 dilution, clone KS-B17.2, Sigma-Aldrich). Secondary antibodies were AlexaFluor 488-labelled goat anti-rabbit IgG and AlexaFluor 594-labelled goat anti-mouse IgG (both from Invitrogen). Negatively stained controls were performed for each antigen by replacing the primary antibody with a suitable isotype (normal mouse IgG or normal rabbit IgG from Santa Cruz Biotechnology) at the same dilutions. Xenotransplants

4
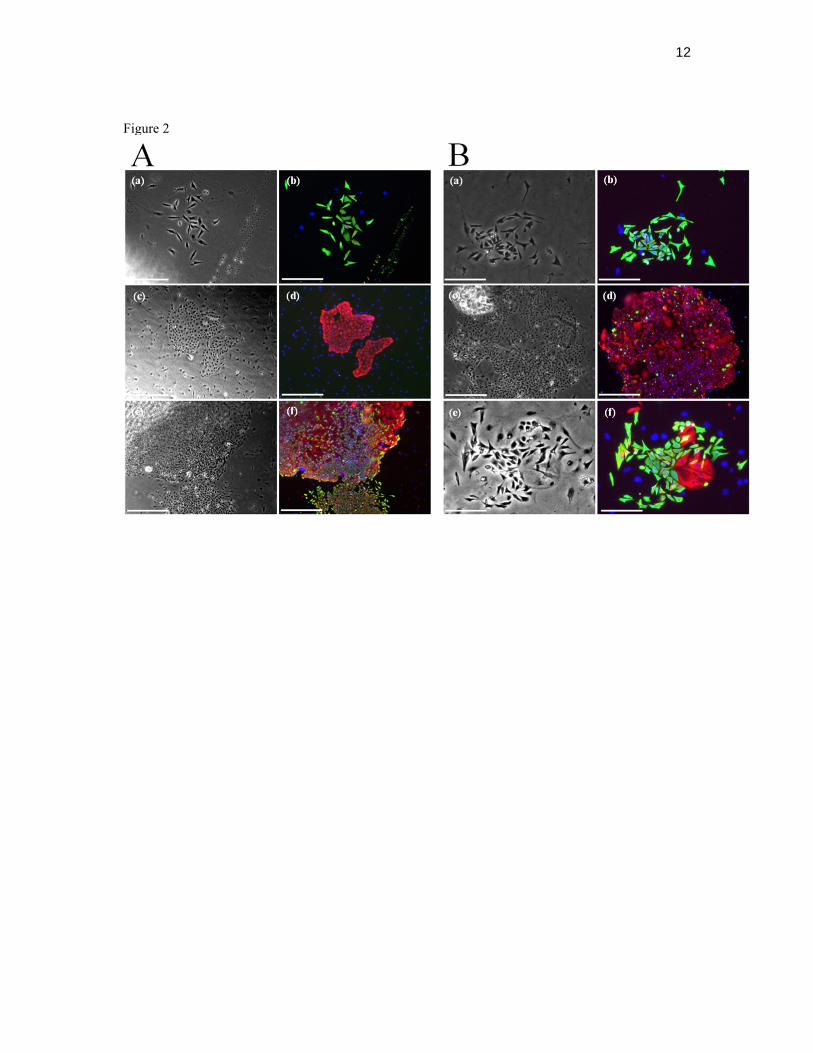
Concentrated rat tail collagen was prepared as previously described [15]. Collagen solution was made by mixing 450 µl of concentrated collagen, 50 µl of 10x DMEM and approximately 0.2 – 0.4% 10 M NaOH to neutralize the pH and kept on ice prior to use. Collagen gels were made as previously described [6]. Each gel contained 1.6x105 10T1/2 fibroblasts previously treated with 2 µg/ml mitomycin C for 16 hours and 7.5x104 caprine primary mammary cells. The gels were kept on ice until surgically implanted as previously described [7].The gels were removed after 4 – 6 weeks and either digested in collagenase solution and seeded in CFC assays or fixed in paraformaldehyde solution (2%) and then embedded in paraffin and immunofluorescently stained. Immunostaining 5 to 7 µm sections were dewaxed and processed for immunofluorescence as previously described [7]. Primary antibodies were against human KRT14, KRT18, TP63 (1:200 dilution, clone 4A4, Thermo Fisher Scientific), α-smooth muscle actin (SMA, 1:500 dilution, A5228, Sigma-Aldrich), ITB1, also known as CD29 (β1-integrin, 1:200 dilution, BD Bioscience), epithelial cell adhesion molecule (EPCAM, 1:200 dilution, E144, Abcam) progesterone receptor (PGR, 1:70 dilution, clone hPRa2, Thermo Fisher Scientific) and milk proteins (RAM/TM, 1:500 dilution, Nordic Immunology). Secondary antibodies and negatively stained controls used were the same as described above. Statistical analyses Data were expressed as the arithmetic mean ± s.e.m. The statistical significance of colony frequencies was determined with the nonparametric test of Friedman. The number of frequency data in each data set (n) is given in Table 1. P values of less than 0.05 were considered to be significant. The statistical significance of percentages of different types of CFCs in 4- and 6-week xenografts was calculated with a Wilcoxon Rank Sum test. P values of less than 0.05 were considered to be significant. RESULTS Histology of goat mammary gland Representative examples of immunocytochemical staining of alveolar structures in goat mammary glands are shown in Figure 1. Myoepithelial cells stained positively for cytokeratin 14 (Fig. 1, A and C), α-smooth muscle actin (Fig. 1D), and ITGB1 (CD29; Fig. 1F) and their nuclei stained positively with p63 antibody (Fig. 1E). Luminal cells were marked by antibodies against cytokeratin 18 (Fig. 1B, C) and progesterone receptor (Fig. 1H), which is located in the nucleus. Epithelial cell adhesion molecule (EPCAM) staining was also located in the luminal compartment, but a lighter staining was also visible in myoepithelial cells (Fig. 1G). In vitro detection of goat mammary epithelial progenitors Distinct large colonies were obtained in vitro after 6 – 8 days when goat mammary cells were cultured at clonal density on a fibroblast feeder layer (Figure 2). The colonies contained cells that expressed cytokeratins typical of differentiated luminal (KRT18) and myoepithelial (KRT14) cells. According to morphological and immunological features at least three types of colonies could be recognized: (i) smaller dispersed or compact tear-drop shaped cells that stained positively for KRT14 (Fig. 2, A: a and b, and B: a and b), (ii) larger, round, compactly packed KRT18+ cells (Fig. 2, A: c and d, and B: c and d), and (iii) a mixture of cells described in the first two morphological types (Fig. 2, A: e and f, and B: e and f). In vitro bipotent progenitors generate mixed colonies that contain both luminal and basal cells. Myoepithelial-restricted progenitors produce colonies that are composed solely of basal-like epithelial cells

5
as defined by their KRT14+ phenotype. Luminal-restricted progenitors generate colonies that are composed only of luminal epithelial KRT18+ cells, and in mixed colonies KRT18+ cells were typically located in the centre of the colony with the KRT14+ cells tending to spread out from the colony boundaries. The in vitro assay to detect the colony-forming ability of mammary cells (CFC) showed that when cells were seeded at clonal density, colonies arose with frequencies ranging from 1 to 8% of the total cells seeded. The three morphologically recognizable types of colonies appeared with different frequencies, depending on the medium used and the sample tested (Table 1). The luminal progenitor compartment is highest in lactating samples (Goat 1 and 2) and during puberty (Goat 4), as shown by luminal type colony frequencies in Sf7 (~80%) and in Epicult-B (from 30 to 70%) medium. Percentage of detected myoepithelial progenitors was highest in sample Goat 3 and was around 80% in both media tested, whereas in the other samples it ranged from 10 to 40% and was consistently higher in Epicult-B. The variation in percentages of specific progenitors detected in the same sample between Sf7 and Epicult-B can be explained with different medium compositions. Therefore the choice of medium to be used in CFC assays must be carefully considered since different media are not equally able to promote the growth of mammary lineage restricted progenitors. In vivo regenerative ability of goat epithelial cells Collagen gels were seeded with a bulk population of caprine epithelial cells obtained from a 7-month-old goat and a 3-year-old lactating goat, mixed with mitomycin C treated 10T1/2 fibroblasts. The initial epithelial cell number was approximately 7.5x104 cells per gel. The gels were placed under the renal capsule of NOD/SCID mice treated with hormones (2 mg β-estradiol and 4 mg progesterone) administered through a silicon pellet. They were recovered after 4–6 weeks. The hematoxylin and eosin (H&E) stained sections show round (Fig. 3Aa) and slightly elongated (Fig. 3Ab) structures. In the immunostained xenograft sections (Fig. 3A: c – h), the spatial distribution of KRT14 and KRT18 was similar to that seen in caprine mammary tissue (Fig. 1C) and cells were organized as a polarized bilayered epithelium enclosing a lumen. The mammary origin of these cells was confirmed by progesterone receptor expression (Fig. 3Ba) and by presence of milk proteins in the lumen of the outgrowths as shown in Figure 3B (b and c). Both stainings also show that the regenerated structures underwent a functional differentiation and were able to perform as proper mammary alveoli. Mammary progenitors in regenerated structures To test for the presence of mammary progenitors in the regenerated structures, we prepared single-cell suspensions from the removed xenograft gels and plated the cells in CFC assays (Fig. 4A). Since the frequencies of colonies obtained in preliminary CFC assays were higher in Sf7, this was used and the colony morphologies were similar to those observed for CFCs from primary mammary suspensions (Fig. 2). After 6 to 7 days in culture, colonies of all three morphological types were detected, i.e. luminal (KRT18 positive colony in Fig. 4A: a, b), myoepithelial (KRT14 positive colonies in Fig. 4A: a, b, and c, d) and mixed (Fig. 4A: e, f). The average number of colonies per dish from dissociated gels ranged from 2 to 331. For the xenografts seeded with epithelial cells originating from goat 4 there was a clear shift in colony type from those containing only luminal to those containing some myoepithelial cells, when comparing primary and secondary CFCs (Fig.4, B and C ). The average representation of the 3 colony types for CFCs obtained from 4- and 6-week-old xenografts (Fig. 4B) was similar (58±11 and 56±18 for the pure KRT14-positive myoepithelial colonies, 16±11 and 33±15 for

6
the pure KRT18-positive luminal colonies, and 26±15 and 11±13 for the mixed colonies, respectively; Fig. 4B). DISCUSSION The goat mammary gland is a tubulo-alveolar organ composed of two lineages of epithelial cells which include luminal (ductal, alveolar) and myoepithelial cells. The ductal structures and terminal ductal lobular units are similar to those found in the human mammary gland but not to those in rodent glands. In rodents the epithelium is in direct contact with stromal adipose cells, whereas in adult humans and in ruminants adipocytes are not closely associated with the epithelium which normally grows enveloped by a loose connective tissue [16]. Goat mammary tissue has been used in studies of lactation, host pathogen interactions, and establishment of epithelial cell lines [2], however there is no information on gMaSCs. The expression of cytokeratins in goat mammary gland tissue has been demonstrated to vary according to developmental status [9] but no consistent data on the range of antibodies and localization of specific markers has been presented. The distribution of differentiation markers in lactating goat mammary gland presented here is similar to those found in human and rodent mammary glands [3, 17]. KRT14 is localized in the basal compartment of alveoli and ducts, whereas KRT18 marks the secretory epithelial cells that face the lumen of the alveoli. The basal compartment of adult goat mammary epithelium consists of progenitors and differentiated myoepithelial cells that line the epithelial cells and are marked by the ability to bind antibodies against α-smooth muscle actin, ITGB1 (CD29), and the nuclear antigen TP63. ITGB1 (CD29) was first described as a stem cell marker in skin and in mouse mammary gland where high levels of this protein were detected in the cap region of terminal end buds, an area presumably enriched in stem cells [3]. Ablation of b1 integrin (CD29) from the basal cell layer of the mammary epithelium abolished its regenerative potential and altered the orientation of the basal cell division plane in mice [18], and may lead to MaSCs depletion, due to altered niche composition or architecture. TP63 is a p53 homolog that is expressed in nuclei of myoepithelial cells in human mammary tissue and is also an important transcription factor for stem cell function and epithelial stasis [19]. In human mammary gland about one third of the luminal epithelial cells express nuclear hormone receptors, estrogen receptor-α and progesterone receptor [20]. In goat mammary gland the staining also revealed that progesterone receptor is present in the luminal compartment. In vivo expression of EPCAM is related to increased epithelial proliferation and correlates negatively with cell differentiation. Its elevated expression has been associated with the majority of human epithelial neoplasias [21]. In lactating goat mammary gland, EPCAM marks luminal epithelial cells and appears to be located on the plasma membrane of some of the basal epithelial cells. This expression pattern closely resembles what has been described in the human mammary tissue [5]. In vitro proliferation of luminal, bipotent, and myoepithelial restricted progenitors was confirmed for the goat model. The colonies obtained from clonogenic progenitors were morphologically similar to those derived from human breast progenitors [5, 6] and expression of luminal and myoepithelial differentiation markers, KRT18 and KRT14, was confirmed. EpiCult-B was developed commercially from Sf7 medium to better support the growth of mammary epithelial cells. However due to unavailability of EpiCult-B exact composition, it is impossible to speculate which components led to the differences in colony frequencies observed in CFC assays. The medium used in experiments with normal pre-menopausal cells [5] was similar to the Sf7 medium used in our research. The total human colony frequency

7
obtained by Stingl and co-workers [5] was comparable to our results. Only the total frequency of colonies originating from goat kid, in Sf7 medium, was lower, namely 1.15±0.12. The total frequencies of goat colonies in EpiCult-B medium were between 1 and 7%. Eirew and colleagues showed that one in every 63 cells of mouse mammary epithelial cells grown in EpiCult-B produced a colony in a CFC assay [4] which is consistent with our findings for goat samples where 1 in every 14 to 1 in every 95 cells was a progenitor. Although pure myoepithelial-cell colonies are rarely observed in CFC assays seeded with primary human tissue [6], they emerged when primary caprine tissue was seeded. The percentage of myoepithelial colonies was the highest in EpiCult-B medium, i.e. around 40% for samples 1, 2, and 4 and 90% for sample 3, and in CFCs derived from xenografts cultured in Sf7 medium (50 – 60%). Additionally, in Sf7 medium the frequency of myoepithelial progenitors derived from the third lactating mammary gland tissue reached 3.55±0.13 out of total 4.40±0.14 (i.e. 80%). One explanation for this shift in lineage development, when comparing the four samples tested, is that the first two mammary glands were at their peak of lactation where the need for secretory luminal cells is the highest and therefore the development of progenitors shifted towards luminal lineage. In contrast, the third gland might have been at the onset of involution, where secreting cells are no longer needed, which might explain the higher proportion of myoepithelial progenitors detected. Similarly, in xenografts, the myoepithelial progenitors were the most abundant. This indicates that gMaSCs in the xenogel milieu develop towards bipotent and myoepithelial progenitors and that there is less differentiation towards luminal progenitors, or more likely that a myoepithelial-restricted progenitor is a more mature progenitor directly derived from the bipotent progenitor. The higher percentage of luminal progenitors in sample 4, which originated from 7-month old goat, might be due to the pubertal state of the animal, where the epithelium undergoes expansion. Hereby we show that there is a marked difference in CFC frequencies when growing goat epithelial cells in different media. Goat cells might have culture requirements different than human ones for which the media tested were originally developed. Nevertheless, the frequencies were comparable with data obtained for human and mouse mammary samples. Adult stem cell multipotent identity is defined by the ability of a single isolated cell from an adult organ to regenerate an entire organ when transplanted. Recently, populations enriched in stem cells have been isolated from adult mouse mammary glands using the surface markers CD24, β1 or α6 integrin chains [3, 4]. Human mammary stem cells are thought to reside within the epithelial basal layer, can be enriched using EPCAMlowITGA2high phenotype, and show regenerative capacity in vivo [6]. The stem/progenitor potential of goat mammary cell populations was assessed in vivo by transplantation under the renal capsule of NOD/SCID mice. Histological detection of regenerated mammary tissue in the implanted gels revealed bilayered structures, which developed within 4-6 weeks after engraftment. The structures contained both mature cytokeratin 18-expressing luminal cells and cytokeratin 14+ myoepithelial cells. The spatial distribution of differentiation markers in regenerated alveoli was similar to those observed in normal goat mammary tissue. In vitro assays of single cell suspensions prepared from gels with regenerated outgrowths demonstrated that progenitor populations were produced and maintained during the in vivo assay. All types of mammary CFCs were found and their morphology was indistinguishable from those derived from the primary tissue. We also showed that milk proteins are present in the regenerated structures, demonstrating that the cells were able to terminally differentiate into functional alveoli. They also have the capacity to respond to hormonal clues as indicated by the expression of progesterone receptor in the nuclei of the luminal cell compartment.

8
In conclusion, using in vivo transplantation we show that a subset of goat epithelial cells have regenerative potential. Our in vitro assay confirmed that different progenitor populations exist in the mammary tissue, which are able of clonogenic proliferation. On the basis of our findings, we conclude that a hierarchy in the organization of epithelial cells exists and that the lineage development of gMaSCs through differentiation follows previously described models. These results lay the basis for future studies in ruminants, leading to better characterization of MaSCs, and will include enrichment of stem cells by sorting strategies. By a thorough understanding of the cellular components and mechanisms that control tissue growth and development, the enhancement of the stem/progenitor cell function can be achieved and exploited in the future ACKNOWLEDGMENTS We thank Cristina Cecere for technical assistance and Vesna Mrak for statistical analyses. REFERENCES 1. Hovey RC, Trott JF, Vonderhaar BK. Establishing a framework for the functional mammary gland: from endocrinology to morphology. J Mammary Gland Biol Neoplasia 2002; 7:17-38. 2. Pantschenko AG, Woodcock-Mitchell J, Bushmich SL, Yang TJ. Establishment and characterization of a caprine mammary epithelial cell line (CMEC). In Vitro Cell Dev Biol Anim 2000; 36:26-37. 3. Shackleton M, Vaillant F, Simpson KJ, Stingl J, Smyth GK, Asselin-Labat ML, Wu L, Lindeman GJ, Visvader JE. Generation of a functional mammary gland from a single stem cell. Nature 2006; 439:84-88. 4. Stingl J, Eirew P, Ricketson I, Shackleton M, Vaillant F, Choi D, Li HI, Eaves CJ. Purification and unique properties of mammary epithelial stem cells. Nature 2006; 439:993-997. 5. Stingl J, Eaves CJ, Zandieh I, Emerman JT. Characterization of bipotent mammary epithelial progenitor cells in normal adult human breast tissue. Breast Cancer Res Treat 2001; 67:93-109. 6. Eirew P, Stingl J, Raouf A, Turashvili G, Aparicio S, Emerman JT, Eaves CJ. A method for quantifying normal human mammary epithelial stem cells with in vivo regenerative ability. Nat Med 2008; 14:1384-1389. 7. Martignani E, Eirew P, Accornero P, Eaves CJ, Baratta M. Human milk protein production in xenografts of genetically engineered bovine mammary epithelial stem cells. Plos One 2010; 5:e13372. 8. Ellis S, Capuco AV. Cell proliferation in bovine mammary epithelium: identification of the primary proliferative cell population. Tissue Cell 2002; 34:155-163. 9. Li P, Wilde CJ, Finch LM, Fernig DG, Rudland PS. Identification of cell types in the developing goat mammary gland. Histochem J 1999; 31:379-393. 10. Smith GH, Medina D. A morphologically distinct candidate for an epithelial stem cell in mouse mammary gland. J Cell Sci 1988; 90 ( Pt 1):173-183. 11. Karatzas CN, Turner JD. Toward altering milk composition by genetic manipulation: Current status and challenges. J Dairy Sci 1997; 80:2225-2232. 12. Berglund B. Genetic improvement of dairy cow reproductive performance. Reprod Domest Anim 2008; 43:89-95. 13. Lucy MC. ADSA Foundation Scholar Award - Reproductive loss in high-producing dairy cattle: Where will it end? J Dairy Sci 2001; 84:1277-1293. 14. Joshi PA, Jackson HW, Beristain AG, Di Grappa MA, Mote PA, Clarke CL, Stingl J, Waterhouse PD, Khokha R. Progesterone induces adult mammary stem cell expansion. Nature 2010; 465:803-807. 15. Richards J, Pasco D, Yang J, Guzman R, Nandi S. Comparison of the growth of normal and neoplastic mouse mammary cells on plastic, on collagen gels and in collagen gels. Exp Cell Res 1983; 146:1-14. 16. Hovey RC, McFadden TB, Akers RM. Regulation of mammary gland growth and morphogenesis by the mammary fat pad: a species comparison. J Mammary Gland Biol Neoplasia 1999; 4:53-68. 17. Rudland PS, Hughes CM. Immunocytochemical identification of cell types in human mammary gland: variations in cellular markers are dependent on glandular topography and differentiation. J Histochem Cytochem 1989; 37:1087-1100.

9
18. Taddei I, Deugnier MA, Faraldo MM, Petit V, Bouvard D, Medina D, Fassler R, Thiery JP, Glukhova MA. Beta1 integrin deletion from the basal compartment of the mammary epithelium affects stem cells. Nat Cell Biol 2008; 10:716-722. 19. Yalcin-Ozuysal O, Fiche M, Guitierrez M, Wagner KU, Raffoul W, Brisken C. Antagonistic roles of Notch and p63 in controlling mammary epithelial cell fates. Cell Death Differ 2010; 17:1600-1612. 20. Clarke RB, Howell A, Potten CS, Anderson E. Dissociation between steroid receptor expression and cell proliferation in the human breast. Cancer Res 1997; 57:4987-4991. 21. Balzar M, Winter MJ, de Boer CJ, Litvinov SV. The biology of the 17-1A antigen (Ep-CAM). J Mol Med 1999; 77:699-712. FIGURE LEGENDS FIG.1. Expression and spatial localization of immunofluorescently detected markers in lactating caprine mammary tissue. Expression of cytokeratins 14 (A – green) and 18 (B – red). Overlay of A and B showing spatial localization of the two markers (C), α-smooth muscle actin (D), TP63 (E), ITGB1 (F), epithelial cell adhesion molecule (G) and progesterone receptor (H) in lactating goat mammary gland. Arrows indicate myoepithelial cells (A, D, E, F, G) and arrowheads luminal cells (B, G, H). Arrows in image E and arrowheads in H depict nuclear staining. Bars = 50 µm. Nuclei were counterstained with DAPI (blue). FIG. 2. Morphology and expression of differentiation markers in caprine mammary colonies grown in two different media. Expression of lineage restricted cytokeratins (KRT14 – green, KRT18 – red) in colonies grown in either Sf7 medium (A) or EpiCult-B medium (B). Representative bright field and immunofluorescence images of a myoepithelial-like colony (A: a, b), a luminal-like colony (A: c, d) and a mixed colony (A: e, f) grown in Sf7 medium, and a myoepithelial-like colony (B: a, b), a luminal-like colony (B: c,d) and a mixed colony (B: e, f) grown in EpiCult-B medium. Bars = 250 µm (A: e, f); bars = 500 µm (other images). Nuclei were counterstained with DAPI (blue). FIG.3. Expression and spatial localization of immunofluorescently detected markers in regenerated structures. (A) H&E staining and cytokeratin 14 and 18 expression in xenografts formed by cells originating from 7-month-old goat (a-d) and cytokeratin 14 and 18 expression in xenografts formed by cells originating from 3-year-old lactating goat (e-h). KRT14 marks myoepithelial cells whereas KRT18 luminal cells. (B) Cytokeratin 14 and progesteron receptor (a; green and red) and milk proteins expression (b,c; green) in regenerated structures formed with epithelial cells of 3-year-old lactating goat, and milk protein expression in lactating mammary tissue (d). Bars = 50 µm (A: a, c, e, g), 100 µm (A: b, f), 62 µm (Ad), 20 µm (Ah and Bc), 15 µm (Bb), 9 µm (Bc), 18 µm (Bd). Nuclei were counterstained with DAPI. FIG.4. A) Morphology and differentiation markers expression in colony forming assays performed on cells from 4 week xenografts. Representative bright field and immunofluorescence images of a luminal colony (a, b), myoepithelial colonies (a,b, and c,d), and a mixed colony (e, f) grown in Sf7 medium. Bars = 500 µm. Nuclei were counterstained with DAPI (blue). B) Percentages of different types of CFCs in freshly thawed lactating (samples Goat 1, Goat 2, Goat 3) and juvenile caprine mammary tissue (Goat 4) in Sf7 and EpiCult-B medium. Data are expressed as the arithmetic mean ± s.e.m. of the number of colonies of specific type over total number of colonies per dish. Brackets on top of each data set marked with an asterisk indicate that there is a statistically significant difference among luminal, myoepithelial and bipotent colony frequencies as calculated with a Friedman test. Statistical significance has been set at p<0.05. C) Percentages of different types of CFCs in 4- and 6-week xenografts (n=3 and n=5) in Sf7 medium generated with epithelial fraction from a 7-month-old goat (Goat 4). Data are expressed as the arithmetic mean ± s.e.m. of the number of colonies of specific type over total number of colonies per dish. Differences in colony frequencies were not found to be statistically significant (p>0.05) as calculated with a Wilcoxon Rank Sum test.
10
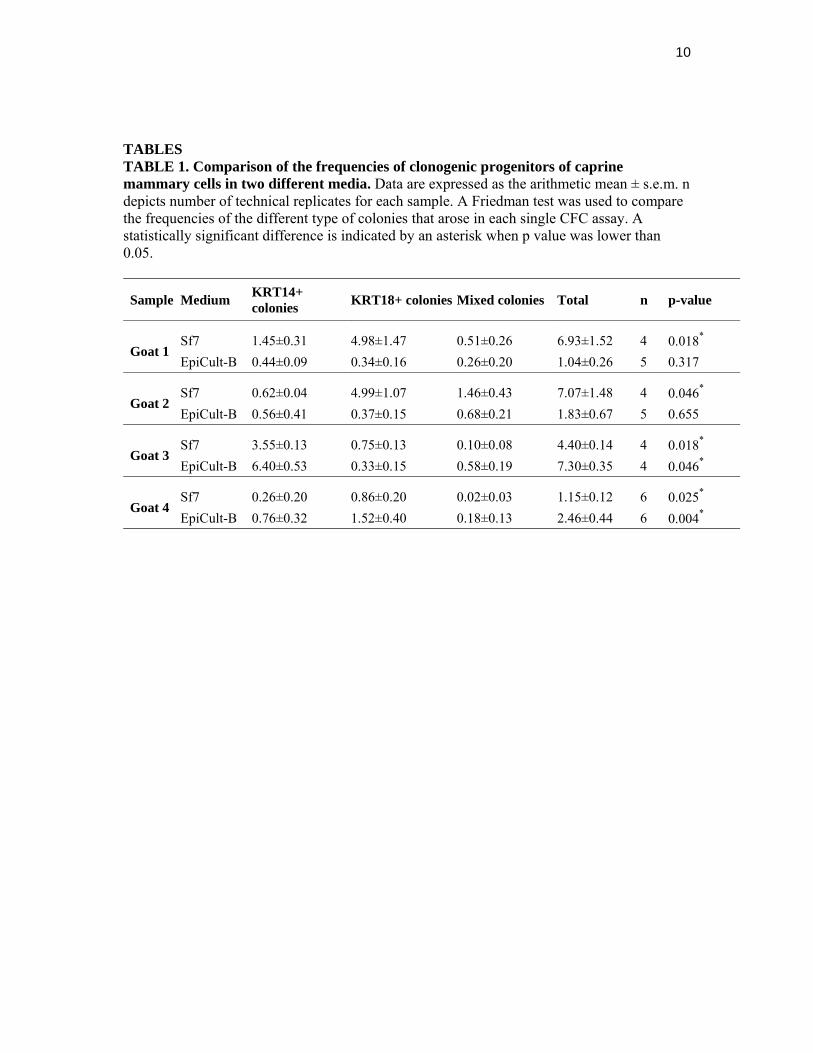
TABLES TABLE 1. Comparison of the frequencies of clonogenic progenitors of caprine mammary cells in two different media. Data are expressed as the arithmetic mean ± s.e.m. n depicts number of technical replicates for each sample. A Friedman test was used to compare the frequencies of the different type of colonies that arose in each single CFC assay. A statistically significant difference is indicated by an asterisk when p value was lower than 0.05.
Sample Medium KRT14+ colonies
KRT18+ colonies Mixed colonies Total n p-value
Goat 1 Sf7 1.45±0.31 4.98±1.47 0.51±0.26 6.93±1.52 4 0.018*
EpiCult-B 0.44±0.09 0.34±0.16 0.26±0.20 1.04±0.26 5 0.317
Goat 2 Sf7 0.62±0.04 4.99±1.07 1.46±0.43 7.07±1.48 4 0.046*
EpiCult-B 0.56±0.41 0.37±0.15 0.68±0.21 1.83±0.67 5 0.655
Goat 3 Sf7 3.55±0.13 0.75±0.13 0.10±0.08 4.40±0.14 4 0.018*
EpiCult-B 6.40±0.53 0.33±0.15 0.58±0.19 7.30±0.35 4 0.046*
Goat 4 Sf7 0.26±0.20 0.86±0.20 0.02±0.03 1.15±0.12 6 0.025*
EpiCult-B 0.76±0.32 1.52±0.40 0.18±0.13 2.46±0.44 6 0.004*

11
FIGURES Figure 1
12
Figure 2

13
Figure 3

14
Figure 4