identification of a substance, previously shown to enhance mitogenesis of human lymphocytes, as the...
TRANSCRIPT

ELSEVIER Biochimica et Biophysics Acta 1214 (1994) 309-316
BiochirnicaL et Biophysics &a
Identification of a substance, previously shown to enhance mitogenesis of human lymphocytes, as the acetamide of p-aminobenzoic acid
Bruno Barbieri a7*, Nikos Papadogiannakis a, Peter Eneroth b, Christian Hansson a, Peter Roepstorff ‘, Lars B. Olding a
a Department of Pathology, Karolinska Institutet, Huddinge University Hospital, S-141 86 Huddinge, Sweden
b Unit for Applied Biochemistry, Karolinska Institutet, Huddinge University Hospital, S-141 86 Huddinge, Sweden ’ Department of Molecular Biology, Odense University, Dk-5230 Odense, Denmark
Received 14 April 1994
Abstract
We characterize here an arachidonic acid (AA)-derived metabolite previously found to have an adjuvant effect in phytohemagglutinin-induced mitogenesis of lymphocytes from mothers of newborn babies and from immunodeficient infants. We named the metabolite ‘compound 4’ due to its position in a thin-layer chromatography system developed for isolation of eicosanoids. The compound was originally found to be produced by peripheral blood mononuclear leukocytes and the T cell leukemia line Jurcat after long-term (18-24 h) incubation with [1-14C]AA. Compound 4 is also produced by lymphocytes, monocytes, platelets, trombocytes, cultured fibroblasts and various types of malignant cell lines. We purified this metabolite by means of high pressure liquid chromatography with synchronous detection of radioactivity and measurement of ultraviolet-light absorption at 278 nm. Proton nuclear
magnetic resonance spectroscopy and mass spectrometry with electron impact techniques demonstrated that compound 4 is not an eicosanoid, but is identical to p-acetamidobenzoic acid (PACBA). The cells synthezise PACBA from p-aminobenzoic acid and a two-carbon residue from AA.
Keywords: Peripheral blood; Mononuclear leukocyte; Arachidonic acid; p-Aminobenzoic acid; p-Acetamidobenzoic acid
1. Introduction
Eicosanoids play a regulatory role both as microenvi- ronmental hormones and intracellular signal mediators. The precursors are the polyunsaturated fatty acids (FA) oleic, linoleic and linolenic acid. These precursors elongate and oxidate to 20-carbon acids, i.e., eicosatrienoic acid
(mead acid), eicosatetraenoic acid (arachidonic acid (AA)) and eicosapentaenoic acid, respectively. Eventually, these become substrates for the formation of eicosanoids such as prostaglandins (PG), leukotrienes, lipoxins and thrombox- anes.
Abbreviations: FA, fatty acid; AA, arachidonic acid; PG, prosta- glandin; PBML, peripheral blood mononuclear leukocyte; PHA, phyto- hemagglutinin; PACBA, p-acetamidobenzoic acid; PABA, p-amino- benzoic acid; FCS, fetal calf serum; TLC, thin-layer chromatography; HPLC, high pressure liquid chromatography; ‘H-NMR, proton nuclear magnetic resonance; Me,SO, dimethylsulfoxide; MS, mass spectrometry.
* Corresponding author. Fax: +46 8 779 55 20.
0005-2760/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved SSDI 0005-2760(94)00110-K
We identified a substance with some of the properties of eicosanoids, while investigating the metabolism of [l- 14C]AA in human neonatal and maternal peripheral blood mononuclear leukocytes (PBML) [l]. We then examined
this substance, ‘compound 4’, to ascertain if it was an eicosanoid. The original investigation was triggered by our discovery that fetal lymphocytes strongly suppress the proliferation of adult lymphocytes [2] and that this suppres- sion is mediated largely by PGE, [3]. Compound 4 was named for its position, among other [1-‘4CjAA-derived metabolites, in a thin-layer radiochromatogram [4].
We found that compound 4 induced an increase in the
incorporation of [ 3H]thymidine in phytohemagglutinin (PI-IA) stimulated PBML from several mothers of new- borns and in PBML from immunodeficient infants [4]. The effect was most prominent among PBML with a low response to PI-IA. In contrast, compound 4 had no effect on the PI-IA-stimulated proliferation of PBML from healthy blood donors.

310 B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316
The purpose of this study was to identify the structure of compound 4 and its formation. Our results clearly indicate that compound 4 is identical to p-acetamidoben- zoic acid (PACBA), a derivative of p-aminobenzoic acid
(PABA) acetylated by two-carbon fragments from AA.
2. Materials and methods
2.1. Sources of cells
Leukocyte-enriched buffy coats originated from the
blood of healthy donors at the Stockholm Slider Hospital. Erythrocytes were kindly provided by the Blood Bank at
Sabbatsbergs Hospital, Stockholm, Sweden. Highly puri- fied monocytes, lymphocytes and platelets were prepared by counterflow centrifugation (elutriation) using a J2-
21M/E Beckman centrifuge (Beckman Instruments, Palo Alto, CA) as previously described [5-71. The following cell lines were obtained from the American Type Culture Collection (ATCC, Rockville, MD); Jurcat (acute T cell leukemia), Molt-4 (peripheral blood, acute lymphoblastic
leukemia), Daudi (Burkitt lymphoma), HeLa (epithelioid carcinoma, cervix), BT-20 (breast carcinoma), MCF-7 (breast adenocarcinoma, pleural effusion), PC 604 (pros-
tate adenocarcinoma), WM 373, WM 9 (melanoma), MRC-5 and WI-38 (lung fibroblasts).
Other cell lines used were; U1690 (carcinoma, lung),
courtesy Dr. Kenneth Nilsson (Department of Pathology, University of Uppsala, Sweden) and M5 (melanoma), courtesy Dr. Ulrik Ringborg (Department of Oncology, Karolinska Hospital, Stockholm, Sweden). LZ and HL
(fibroblasts, fetal lung) were kindly provided by the Swedish Institute for Infectious Disease Control (Stock- holm, Sweden).
2.2. Biosynthesis of compound 4
PBML were isolated by means of Ficoll-Isopaque (Pharmacia Biotechnology, Uppsala, Sweden) gradient
centrifugation of the leukocyte-enriched buffy coats [3]. The cells were suspended in 50 ml RPM1 1640 medium,
5% fetal calf serum (FCS), and antibiotic (Gentamycin) at a density of lo6 cells/ml. The culture components were obtained from Gibco, Life Technologies, Paisley, Scotland. A mixture of [1-14CjAA (53.0 Ci/mol, NEN Products, Boston, MA) and unlabeled AA (Sigma Chemical, St. Louis, MO) was added to a final concentration of 0.092 &i/ml and 5.0 PM. After 18 h of incubation at 37°C in 5% CO, in humidified air, the cell suspension was cen- trifuged at 430 X g for 15 min, and the supematant was collected. Cell viability was > 95% after incubation ac- cording to the Trypan blue exclusion test. Incubations with Jurcat cells were performed identically to those for PBML. Incubations with other cells and cell lines, done for the purpose of screening, were performed similarly, but media
other than RPM1 were used when recommended by the American Type Culture Collection.
2.3. Purification of compound 4
Supernatants from cell cultures were acidified to pH 3 and subsequently extracted three times with an equal amount of ethyl acetate. The organic phase was washed neutral, evaporated and the residue was then applied to an
open SiO, column, 10 X 200 mm, packed with Silicar cc-4 (Mallinckrodt, Paris, KY). The column was eluted with 35
ml ethyl acetate/hexane (1:9, v/v), 50 ml ethyl acetate/ hexane (2:8, v/v) and 40 ml pure ethyl acetate. The latter fraction containing compound 4 was purified by thin-layer
chromatography (TLC) using the organic phase from ethyl acetate/ water/acetic acid/isooctane (11: 10:2:6, v/v).
[5,6,8,11,12,14,15-3H1PGE, (200 Ci/mmol, NEN) and [1-‘4ClAA were used as standards. The TLC plate was then scanned for the radioactive spot corresponding to compound 4. This material was removed, extracted, and further purified in small portions by high pressure liquid chromatography (HPLC). The column used was a Supelco
C-18, 5 pm, 4.6 X 250 mm (Supelco, Bellefonte, PA), and the mobile phase consisted of a linear gradient between
methanol (20 to 100% in 20 min) and water. Both solvents were buffered with 0.2% acetic acid. The flow rate was constantly 1 ml/min. UV-light absorption was monitored
with a Shimadzu SPD-6A UV detector (Shimadzu Corpo- ration, Kyoto, Japan) at the absorption maxima 278 nm, and radioactivity was measured synchronously with a Flow-One A-200 detector (Packard Instrument Company, Meriden, CT). UV-light absorption and radioactivity were confirmed as arising from a single substance with the retention volume of 11.73 ml. These fractions were col- lected and lyophilized.
2.4. Calculation of carbon atoms derived from AA in
compound 4
These experiments using [1-14C] and [U-‘4C] (1.0
Ci/mmol, NEN) labeled AA were designed in two ways. RPM1 (49.9 ml) with 5% Al3 serum and 50. lo6 PBML was supplemented either with 100 ~1 of ethanol containing 171 nmol unlabeled AA and 3.15 nmol [U-‘4C]AA or with 100 ~1 of ethanol containing 141 nmol of unlabeled AA and 33.0 nmol [l-‘“C&4. The final concentration of AA was 3.48 PM in each experiment. Radioactivity was counted in a LKB 1214 Rackbeta liquid scintillation counter (Wallac Oy, Turku, Finland) after addition of 2 ml Op- tiPhase HiSafe (Wallac) scintillation cocktail per 100 ~1 sample volume. After 24 h of incubation the Trypan blue exclusion test confirmed that > 95% of the cells remained viable. The cell suspensions were centrifuged at 430 X g for 15 min; then the supematants were removed and treated as described above. During HPLC, peaks with the retention volume of 11.7 ml were collected, lyophilized
B. Barbieri et al. / Biochimica et Biophysics Acta 1214 (I 994) 309-316 311
and redissolved in ethyl acetate. Finally the radioactivity in 2.8. Cleavage of the amide bond in compound 4 and in compound 4 was measured and the yield calculated. PACBA
2.5. Proton nuclear magnetic resonance VH-NMR)
‘H-NMR spectra of compound 4 and PACBA were recorded at room temperature in deuterated dimethylsulf- oxide (Me,SO) with JEOL GX-370 and JEOL GX-270 instruments (Jeol, Tokyo, Japan), respectively. Chemical
shifts were reported in ppm (S), using the small amount (0.05%) of undeuterated solvent as an internal reference.
2.6. Mass spectrometry (MS)
Positive and negative ion plasma desorption mass spec-
tra were obtained on a BioIon 20 mass spectrometer (Applied Biosystems, Foster City, CA) operated at 15 and - 10 KeV acceleration voltage, respectively. The sample
was applied on a nitrocellulose covered target, and spectra were accumulated for lo6 fission events. The spectra were
calibrated by using peaks for H+ and NO+ in the positive mode and H- and CN- in the negative mode. Positive ion
electrospray spectra were acquired on a Vestec 201 single quadrupole mass spectrometer (Vestec Corporation, St. Louis, TS) equipped with a Vector 2 data system (Tecni-
vent). The sample, dissolved in methanol/water (l:l, v/v), was introduced at a flow rate of 0.5 $/min through a syringe pump (Orion Research, Boston, MA).
200 ~1 of 5 M KOH and 100 ~1 methanol were added to 10 nmol portions of PBML-produced 14C-labeled com- pound 4 and to 538 nmol portions of synthetic 14C-labeled
PACBA. The samples were enclosed in ampoules and heated to 110°C for 1 h. The reaction mixtures were then brought to room temperature, supplemented with 3 ml
water, acidified to pH 3 with HCl, and subsequently extracted three times with 2 ml of ethyl acetate. Extracts from each sample were pooled, dried under a helium
stream, left overnight in a desiccator and dissolved in 1 ml of ethyl acetate. The radioactivity was measured and com- pared with that in each original sample. To assure that the
reaction had proceeded as expected, the absorption of UV-light was measured and compared with the one for PACBA.
3. Results
3.1. Biosynthesis and purification of compound 4
Electron impact MS was carried out on a Nermag RlO-10H quadrupole mass analyzer (Delsi Nermag Instru- ments, Ruil-Malmaison, France) consisting of a desorption probe, ionization source, quadrupole filter, and ion detec-
tor. The sample was dissolved in methanol and applied on the tungsten filament of the desorption probe with a Hamilton syringe. Spectra were obtained in the EI mode at 70 eV, at an ion source temperature of 180°C.
After incubation of the human PBML with [l-14C]AA, about 50% of the radioactivity was recovered in the super-
natants. Of this amount, 70-80% remained in the ethyl
acetate extract. After this extract passed through a SiO, column, a typical distribution of the radioactivity among the different eluates was 81% (1:9, ethyl acetate/hexane), 4.2% (2:8, ethyl acetate/hexane) and 9.0% (ethyl acetate). Evaporation of the ethyl acetate fraction, containing possi- ble PGs and compound 4, and separation in the TLC
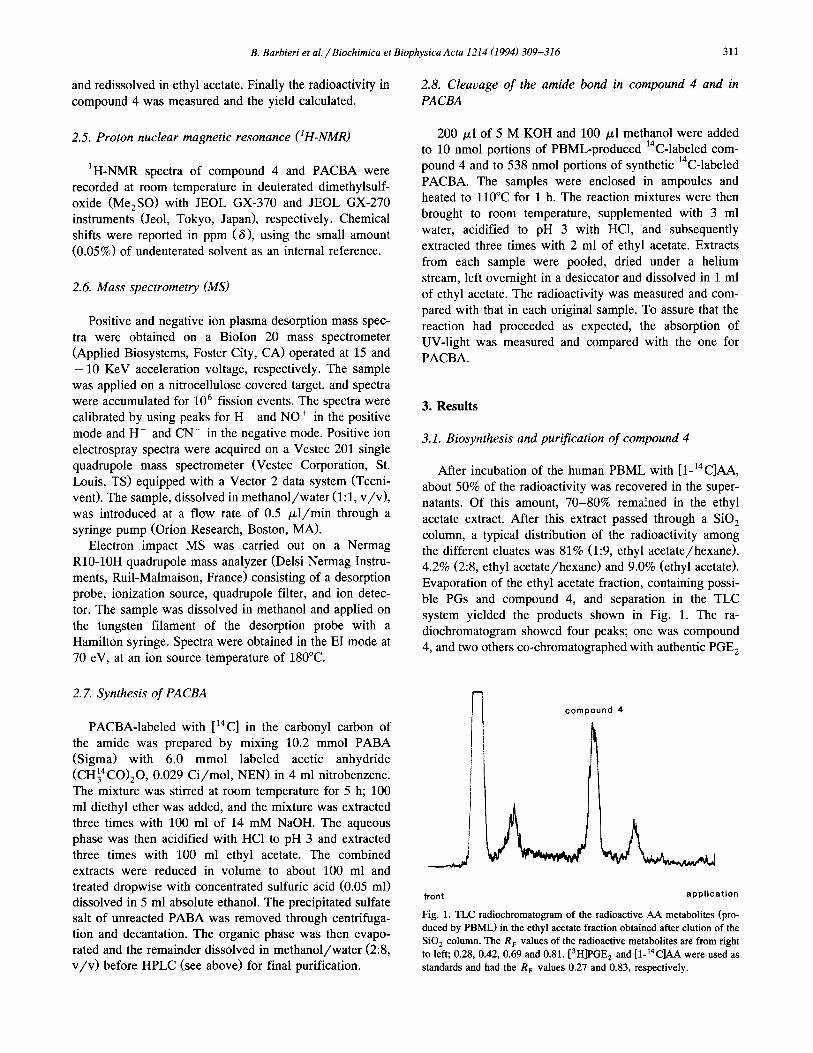
system yielded the products shown in Fig. 1. The ra- diochromatogram showed four peaks; one was compound
4, and two others co-chromatographed with authentic PGE,
2.7. Synthesis of PACBA compound 4
PACBA-labeled with [ 14C] in the carbonyl carbon of the amide was prepared by mixing 10.2 mmol PABA (Sigma) with 6.0 mmol labeled acetic anhydride (CHi4CO),0, 0.029 Ci/mol, NEN) in 4 ml nitrobenzene. The mixture was stirred at room temperature for 5 h; 100 ml diethyl ether was added, and the mixture was extracted three times with 100 ml of 14 mM NaOH. The aqueous phase was then acidified with HCl to pH 3 and extracted three times with 100 ml ethyl acetate. The combined
extracts were reduced in volume to about 100 ml and treated dropwise with concentrated sulfuric acid (0.05 ml) dissolved in 5 ml absolute ethanol. The precipitated sulfate salt of unreacted PABA was removed through centrifuga- tion and decantation. The organic phase was then evapo- rated and the remainder dissolved in methanol/water (2:8, v/v> before HPLC (see above) for final purification.
front application
Fig. 1. TLC radiochromatogram of the radioactive AA metabolites (pro-
duced by PBML) in the ethyl acetate fraction obtained after elution of the
SO, column. The R, values of the radioactive metabolites are from right
to left; 0.28, 0.42, 0.69 and 0.81. [3H]PGE, and [1-14C]AA were used as
standards and had the R, values 0.27 and 0.83, respectively.
312 B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316
and AA. The fourth peak, next to AA, was not analyzed further but might have been a monohydroxy- or a hydro- peroxyeicosatetraenoic acid judging from its position [8].
Compound 4 produced by PBML was on average about 1.5% (0.03% per lo6 cells) of the radioactivity originally added as AA at the start of incubation. The range in
production by PBML from nine different blood donors was only l.l-2.0%. The corresponding yield for Jurcat cells
was about 2.0% (0.04% per lo6 cells), but no PGE,-like peak appeared. However, the intensity of the UV-light absorption at 278 nm of compound 4 in combination with the radioactive yield indicated that substrates other than
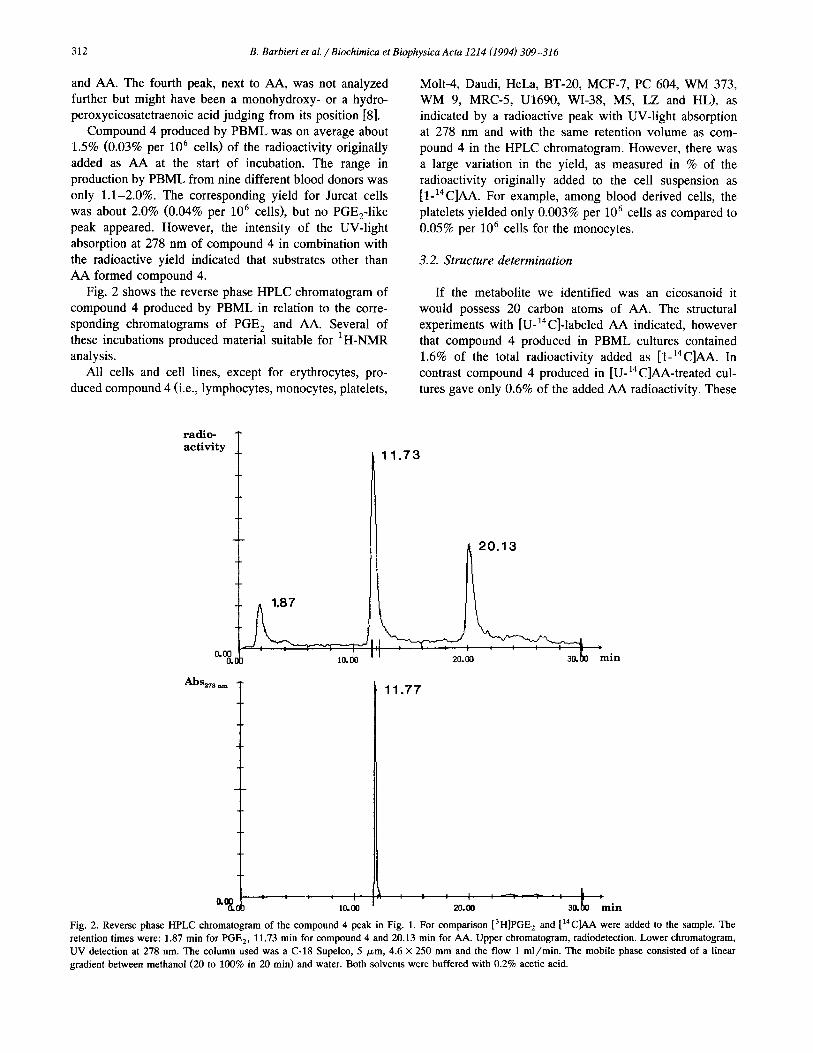
AA formed compound 4. Fig. 2 shows the reverse phase HPLC chromatogram of
compound 4 produced by PBML in relation to the corre-
sponding chromatograms of PGE, and AA. Several of these incubations produced material suitable for ‘H-NMR analysis.
All cells and cell lines, except for erythrocytes, pro- duced compound 4 (i.e., lymphocytes, monocytes, platelets,
radio- T
Molt-4, Daudi, HeLa, BT-20, MCF-7, PC 604, WM 373, WM 9, MRCJ, U1690, WI-38, M5, LZ and HL), as indicated by a radioactive peak with UV-light absorption at 278 nm and with the same retention volume as com- pound 4 in the HPLC chromatogram. However, there was
a large variation in the yield, as measured in % of the radioactivity originally added to the cell suspension as
[1-‘4ClAA. For example, among blood derived cells, the platelets yielded only 0.003% per lo6 cells as compared to 0.05% per lo6 cells for the monocytes.
3.2. Structure determination
If the metabolite we identified was an eicosanoid it would possess 20 carbon atoms of AA. The structural experiments with [U- l4 Cl-labeled AA indicated, however that compound 4 produced in PBML cultures contained
1.6% of the total radioactivity added as [1-14C]AA. In contrast compound 4 produced in [U- 14C]AA-treated cul- tures gave only 0.6% of the added AA radioactivity. These
am \ : : : : :-:t-: a tam 20. m 30.
Fig. 2. Reverse phase HPLC chromatogram of the compound 4 peak in Fig. 1. For comparison [3~]~~~, and [‘4C]AA were added to the sample. The
retention times were: 1.87 min for PGE,, 11.73 min for compound 4 and 20.13 min for AA. Upper chromatogram, radiodetection. Lower chromatogram,
UV detection at 278 nm. The column used was a C-18 Supelco, 5 pm, 4.6 x 250 mm and the flow 1 ml/min. The mobile phase consisted of a linear gradient between methanol (20 to 100% in 20 min) and water. Both solvents were buffered with 0.2% acetic acid.
B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316 313
results indicated that roughly 7 carbon atoms in compound
4 were derived from the AA substrate, because 0.6/1.6 =
0.375 and 37.5% of 20 carbons is 7.5 carbons assuming
that the molar ratio between substrate and product is 1:l. This number would disqualify compound 4 for considera- tion as an eicosanoid.
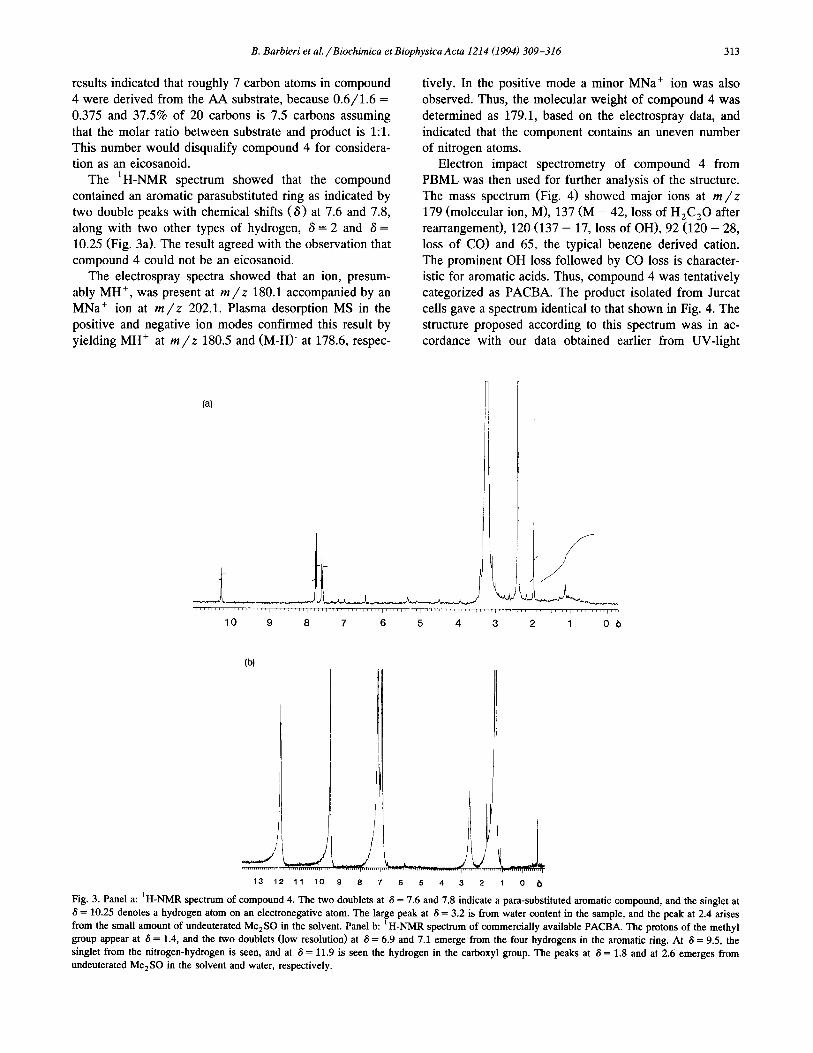
The ‘H-NMR spectrum showed that the compound
contained an aromatic parasubstituted ring as indicated by two double peaks with chemical shifts (6) at 7.6 and 7.8,
along with two other types of hydrogen, S = 2 and S = 10.25 (Fig. 3a). The result agreed with the observation that compound 4 could not be an eicosanoid.
The electrospray spectra showed that an ion, presum-
ably MH+, was present at m/z 180.1 accompanied by an MNa+ ion at m/z 202.1. Plasma desorption MS in the positive and negative ion modes confirmed this result by
yielding MH+ at m/t 180.5 and (M-H)‘ at 178.6, respec-
tively. In the positive mode a minor MNa+ ion was also
observed. Thus, the molecular weight of compound 4 was
determined as 179.1, based on the electrospray data, and
indicated that the component contains an uneven number of nitrogen atoms.
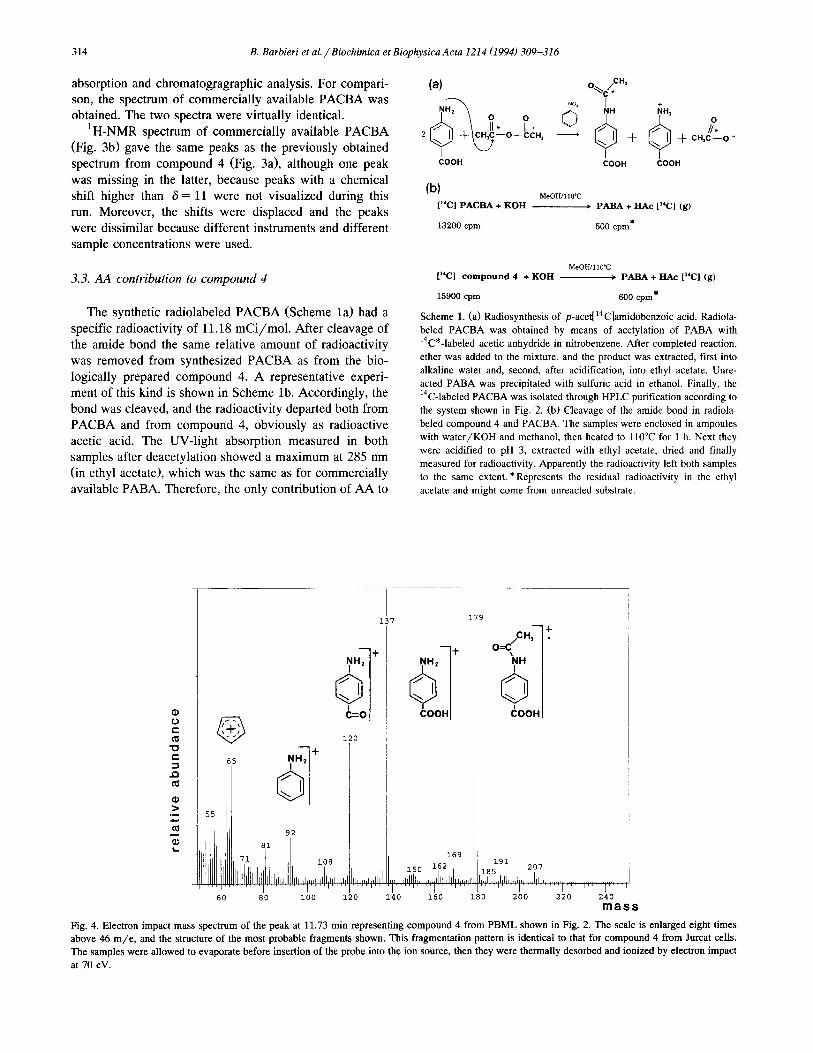
Electron impact spectrometry of compound 4 from
PBML was then used for further analysis of the structure. The mass spectrum (Fig. 4) showed major ions at m/z 179 (molecular ion, M), 137 (M - 42, loss of H,C,O after rearrangement), 120 (137 - 17, loss of OH), 92 (120 - 28,
loss of CO) and 65, the typical benzene derived cation. The prominent OH loss followed by CO loss is character- istic for aromatic acids. Thus, compound 4 was tentatively categorized as PACBA. The product isolated from Jurcat
cells gave a spectrum identical to that shown in Fig. 4. The structure proposed according to this spectrum was in ac- cordance with our data obtained earlier from UV-light
(4
,““~““,‘“‘,“‘,“,,1”,‘,““i”,‘,“,,l’,~,,’,”,’,~1”,,ll,lr/l,,,,,,,,,,,,‘,,‘~,,,~
10 9 8 7 6 5 4 3 2 1 Ob
13 12 11 10 9 8 7 6 5 4 3 2 1 0 (,
Fig. 3. Panel a: ‘H-NMR spectrum of compound 4. The two doublets at 6 = 7.6 and 7.8 indicate a para-substituted aromatic compound, and the singlet at
6 = 10.25 denotes a hydrogen atom on an electronegative atom. The large peak at S = 3.2 is from water content in the sample, and the peak at 2.4 arises
from the small amount of undeuterated Me,SO in the solvent. Panel b: ‘H-NMR spectrum of commercially available PACBA. The protons of the methyl
group appear at 6 = 1.4, and the two doublets (low resolution) at 6 = 6.9 and 7.1 emerge from the four hydrogens in the aromatic ring. At 6 = 9.5, the
singlet from the nitrogen-hydrogen is seen, and at 6 = 11.9 is seen the hydrogen in the carboxyl group. The peaks at 6 = 1.8 and at 2.6 emerges from
undeuterated Me,SO in the solvent and water, respectively.
314 B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316
absorption and chromatogragraphic analysis. For compari- son, the spectrum of commercially available PACBA was obtained. The two spectra were virtually identical.
’ H-NMR spectrum of commercially available PACBA (Fig. 3b) gave the same peaks as the previously obtained
spectrum from compound 4 (Fig. 3a), although one peak
was missing in the latter, because peaks with a chemical shift higher than 6 = 11 were not visualized during this run. Moreover, the shifts were displaced and the peaks were dissimilar because different instruments and different sample concentrations were used.
3.3. AA contribution to compound 4
The synthetic radiolabeled PACBA (Scheme la) had a specific radioactivity of 11.18 mCi/mol. After cleavage of the amide bond the same relative amount of radioactivity
was removed from synthesized PACBA as from the bio- logically prepared compound 4. A representative experi- ment of this kind is shown in Scheme lb. Accordingly, the
bond was cleaved, and the radioactivity departed both from PACBA and from compound 4, obviously as radioactive acetic acid. The UV-light absorption measured in both samples after deacetylation showed a maximum at 285 nm
(in ethyl acetate), which was the same as for commercially available PABA. Therefore, the only contribution of AA to
tOOH 600” Coon
MeOWl 10°C
[“Cl PACBA + KOH - PABA + HAc [“Cl (g)
13200 cpm 500 cpm*
MeOH.'llO"C
[“Cl compound 4 + KOH ____, PABA + HAc [“Cl (g)
15900 cpm 600 cpm*
Scheme 1. (a) Radiosynthesis of p-acet[‘4C]amidobenzoic acid. Radiola-
beled PACBA was obtained by means of acetylation of PABA with
r4C*-labeled acetic anhydride in nitrobenzene. After completed reaction,
ether was added to the mixture, and the product was extracted, first into
alkaline water and, second, after acidification, into ethyl acetate. Unre-
acted PABA was precipitated with sulfuric acid in ethanol. Finally, the
r4C-labeled PACBA was isolated through HPLC purification according to
the system shown in Fig. 2. (b) Cleavage of the amide bond in radiola-
beled compound 4 and PACBA. The samples were enclosed in ampoules
with water/KOH and methanol, then heated to 110°C for 1 h. Next they
were acidified to pH 3, extracted with ethyl acetate, dried and finally
measured for radioactivity. Apparently the radioactivity left both samples
to the same extent. *Represents the residual radioactivity in the ethyl
acetate and might come from unreacted substrate.
179
,h- o=c
t
mass
Fig. 4. Electron impact mass spectrum of the peak at 11.73 min representing compound 4 from PBML shown in Fig. 2. The scale is enlarged eight times
above 46 m/e, and the structure of the most probable fragments shown. This fragmentation pattern is identical to that for compound 4 from Jurcat cells.
The samples were allowed to evaporate before insertion of the probe into the ion source, then they were thermally desorbed and ionized by electron impact
at 70 eV.

B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316 315
the formation of PACBA was the acetyl groups that acety- lated PABA. The main source of PABA for the cells in the
cultures was the media because PABA is present as a
constituent of RPM1 1640 at a consentration of 7.30 PM. The molar ratio between AA and PACBA must theoreti-
cally be l:lO, but the results from the experiments with [U-14C]AA indicated that the virtual molar ratio is about
1:4, i.e, that on average only 4 acetyl fragments from AA were used for the acetylation of PABA.
4. Discussion
In the present investigation, the combined data from
chromatography as well as MS and ‘H-NMR spectroscopy identified compound 4, derived from human cells and cell lines, as the acetamide of PABA. Our data also showed
that the amine in PABA is acetylated by acetyl groups derived from AA.
N-Acetylation is a common reaction for amines in most
cells including lymphocytes [9]. According to our results, the N-acetylation capacity of
PABA is a metabolic pathway shown to be common in all eucaryotic blood cells and platelets investigated, but not in
erythrocytes. It is also preserved in cultured fibroblasts and many different malignant cell lines. According to the known mechanism for acetylation of amines every kind of
molecule giving acetyl-CoA during metabolism could serve as acetyl donor for acetylation of PABA. Thus, it is apparent that AA contributes to a small extent to the total acetylation of PABA.
Enzymatic N-acetylation proceeds according to a ‘ ping-pong’ reaction mechanism [lo]. This is a double displacement reaction in which the enzyme, N-acetyltrans-
ferase, oscillates between free and acetylated forms. The enzyme acetylated by acetyl-CoA acetylates the actual amine. This means that, in the system we analyzed, AA
must have undergone P-oxidation to form acetyl-CoA before acetylation of PABA. The lack of mitochondria is
probably the reason why we could not detect any acetyla- tion of PABA by erythrocytes.
The incorporation of radioactivity, originating from AA, into PACBA was about 1.5% for PBML and 2.0% for Jurcat cells. This amount is large considering that the AA carbons 1 and 2 must have made up a rather small proportion of the acetate pool. No specific features are known for the P-oxidation of AA. However, some might be suspected from the evidence that AA is a precursor of messenger substances or is used as a specific building
block in cell membranes as a constituent of phospholipids, predominantly phosphatidylcholine and phosphatidyl- ethanolamine [ll]. AA is not considered simply as a fuel, especially not when other common fuels are available in excess. It has been suggested that its slower turnover than other FA reflects the essential character of AA [12]. A marked tendency for M to enter the phospholipid pool
[13] would offer an explanation for the low catabolic rate
of this FA. Phospholipids are important to the structural
integrity of cells and are known to turn over more slowly
than, for example, triacylglycerols [12]. Although little is known about PABA’s biological ef-
fects per se, one report infers that PABA interferes with the incorporation of acetate into human blood lipids 1141. Our previous data indicated that the acetylated form of PABA may function in connection with cell proliferation.
Humans can be classified as either fast or slow acetyla- tors on the basis of their ability to metabolize isoniazide or
sulphametazine [9]. These are termed polymorphic sub-
strates. The acetylation of other compounds, e.g., PABA, does not vary much among individuals and such com-
pounds are termed monomorphic substrates [15]. Similarly, in our experiments the acetylation of PABA by PBML did
not vary much from one healthy blood donor to another. Experiments in which PABA was administrated orally
(800 mg) showed that about 12% was excreted as PACBA in the urine within 24 h [16]. However, the major metabo-
lite excreted was p-aminohippuric acid, which represented about 50% of the administrated PABA. According to our
preliminary results (not shown) the appearance of PACBA in urine from individuals on an average mixed diet, with- out any supplementation of PABA, is in the range of 0.5 to
1.5 mg/l.
Acknowledgments
This work was supported by grants from the Karolinska Institutet, the Swedish Cancer Society and Magn. Berg- wall’s Foundation, Stockholm. We are indebted to Ag- netha Beinhoff and Agneta Soderstedt for their invaluable
laboratory assistance, to Kerstin Svanborg for methodolog- ical advice, to Gunvor Alvelius for skillful assistance with
mass spectrometry and to Phyllis Minick for editorial assistance.
References
[l] Johnsen, S.A., Olding, L.B. and Green, K. (1983) Immunol. Lett. 6,
213-218.
[2] Olding, L.B. and Oldstone, M.B.A. (1974) Nature Lond. 249,
161-162.
[3] Papadogiannakis, N., Johnsen, S.A. and Olding, L.B. (1990) in The
immunology of the fetus (Chaouat, G., ed.), pp. 21.5-227, CRC
Press, Boca Raton, Florida.
141 Johnsen, S.A., Green, K., Papadogiannakis, N. and Olding, L.B.
(1987) Stand. J. Immunol. 25, 425-428.
[51 De Mulder, P.H.M., Wessles, J.M.C., Rosenbrand, D.A., Smeulders, J.B.J.M., Wagener, D.J.T. and Haanen, C. (1981) J. Immunol.
Methods 47, 31-38.
[6] Mackler, B.F., O’Neil, P.A. and Meistrich, M. (1977) Eur. J. Immunol. 7, 55-61.
[7] Jakubowski, J.A., Thompson, C.B., Vaillancourt, R., Valeri, C.R.
and Deykin, D. (1983) Br. J. Haematol. 53, 503-511.

316 B. Barbieri et al. /Biochimica et Biophysics Acta 1214 (1994) 309-316
[8] Hurst, J.S., Flatman, S. and McDonald-Gibson, R.G. (1987) in
Prostaglandins and related substances (Benedetto, C., McDonald-
Gibson, R.G., Nigam, S. and Slater, T.F., ed.), pp. 53-73, IRL
Press, Oxford, England.
[9] Coroneos, E., Gordon, J.W., Kelly, S.L., Wang, P.D. and Sim, E.
(1991) Biochim. Biophys. Acta 1073, 593-599.
lo] Weber, W.W. and Hein, D.W. (1985) Pharmacol. Rev. 37, 25-79.
111 Hong, S.L. (1988) in Arachidonate metabolism in immunologic
systems (Levine, L., ed.), pp. 99-139, Karger, Base], Switzerland.
[12] Coots, R.H. (1965) J. Lipid. Res. 6, 494-497.
[13] Stoffel, W. und Scheid, A. (1967) Hoppe-Seyler’s Z. Physiol. Chem.
348, 205-226.
[14] Mandelbaum-Shavit, F. and Blondheim, S.H. (1981) Biochem. Phar-
macol. 30, 65-69.
(151 Cribb, A.E., Grant, D.M., Miller, M.A. and Spielberg, S.P. (1991) J.
Pharmacol. Exp. Therap. 3, 1241-1246.
[16] Chan, K., Miners, J.O. and Birkett, D.J. (1988) J. Chromatog. 426,
103-109.