identification of paecilomyces variotii in clinical samples...
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2010, p. 2754–2761 Vol. 48, No. 80095-1137/10/$12.00 doi:10.1128/JCM.00764-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Identification of Paecilomyces variotii in ClinicalSamples and Settings�
Jos Houbraken,1* Paul E. Verweij,2 Anthonius J. M. M. Rijs,2Andrew M. Borman,3 and Robert A. Samson1
CBS-KNAW Fungal Biodiversity Centre, Department of Applied and Industrial Mycology, Utrecht, Netherlands1;Department of Medical Microbiology, Radboud University Nijmegen Medical Centre, Nijmegen, Netherlands2;
and HPA Mycology Reference Laboratory, HPA Southwest, Bristol, United Kingdom3
Received 15 April 2010/Returned for modification 10 May 2010/Accepted 21 May 2010
Paecilomyces variotii is a commonly occurring species in air and food, but it is also associated with many typesof human infections and is among the emerging causative agents of opportunistic mycoses in immunocom-promised hosts. Paecilomyces can cause hyalohyphomycosis, and two species, Paecilomyces lilacinus and P.variotii, are the most frequently encountered organisms. In the present study, a set of 34 clinical isolatesmorphologically identified as P. variotii or P. lilacinus were formally identified by sequencing intergenictranscribed spacer regions 1 and 2 (including 5.8S rDNA) and a part of the �-tubulin gene. Three isolates wereidentified as P. lilacinus, and five of the presumptive P. variotii isolates did not belong to the genus Paecilomycesbut were identified as Talaromyces eburneus (anamorph, Geosmithia argillacea) or Hamigera avellanea (ana-morph, Merimbla ingelheimense). Applying the most recent taxonomy, we found that the clinical P. variotiiisolates could be identified as P. variotii sensu stricto (14 strains), P. formosus (11 strains), and P. dactylethro-morphus (1 strain). These data indicate that P. formosus occurs in clinical samples as commonly as P. variotii.Susceptibility tests showed that the antifungal susceptibility profiles of P. variotii, P. formosus, and P. dac-tylethromorphus are similar and that all strains tested were susceptible to amphotericin B in vitro. P. lilanicus,T. eburneus, and H. avellanea had different susceptibility profiles; and flucytosine and voriconazole were theleast active of the antifungal drugs tested against these species. Our results indicate that correct speciesidentification is important to help guide appropriate antifungal therapy.
Paecilomyces variotii is a commonly occurring species thathas previously been isolated from various substrates, including(pasteurized) foods, soil, indoor air, and wood (23, 36, 41, 42,43). However, it is also associated with many types of humaninfections and is listed among the emerging causative agents ofopportunistic mycoses in immunocompromised hosts. Paecilo-myces can cause hyalohyphomycosis (1), and two species, Pae-cilomyces lilacinus and P. variotii, are the most frequently en-countered (20, 52). Both species are morphologically similarbut can be differentiated on the basis of conidial color andgrowth rates (41). However, small-subunit ribosomal gene se-quences showed that the two species are unrelated: P. variotiibelongs to the order Eurotiales, while P. lilacinus is a memberof the order Hypocreales (27). Although they are uncommon,Paecilomyces infections are associated with almost any organor system of the human body (40). Most cases concern immu-nocompromised patients and are cutaneous or catheter re-lated. However, dissemination, for example, that involving thecentral nervous system, has been observed in a number of cases(15, 24). Ocular infections associated with prolonged contactlens use or ocular surgery have also been reported (40), ashas peritonitis in patients with continuous ambulatory peri-toneal dialysis (24), which apparently responded well to an-tifungal therapy, with cures occurring in 11 of the 13 cases.
However, the majority of these patients were treated for Pae-cilomyces peritonitis and were not severely immunocompro-mised. In vitro amphotericin B was active against clinical P.variotii isolates but not against P. lilacinus (8). Among theazoles, only itraconazole and posaconazole showed clinicallyrelevant activity against P. variotii. The geometric mean MICsof voriconazole and ravuconazole were above 4 mg/liter, indi-cating that these azoles will not be effective in vivo. Interest-ingly, the echinocandins, especially micafungin and anidula-fungin, were highly active against P. variotii, with MIC valuesbeing as low as 0.016 mg/liter. Conversely, P. lilacinus was notinhibited by the echinocandins, underscoring the importanceof correct species identification (8).
Paecilomyces variotii grows rapidly on standard agar andforms velvety olive brown colonies. Conidiophores of P. variotiiare irregularly branched, and the phialides have a broad baseending in a long and slender neck. Samson (41) noted that P.variotii is a morphologically variable species, and the taxonomyof P. variotii and the related Byssochlamys teleomorphs hasrecently been revised (45). This revision is based on morphol-ogy, extrolites, and molecules and shows that P. variotii sensulato comprises five species, namely, Byssochlamys spectabilis(the sexual state of P. variotii), P. brunneolus, P. formosus, P.divaricatus, and P. dactylethromorphus. The last species wasincorrectly named P. saturatus because P. dactylethromorphuswas validly described in 1957 and has priority. The aim of thepresent study was to determine the prevalence of these speciesin clinical samples and settings. A total of 34 isolates originat-ing from various clinical specimens and settings were identifiedby sequencing the intergenic transcribed spacer (ITS) regions,
* Corresponding author. Mailing address: CBS-KNAW Fungal Biodi-versity Centre, Department of Applied and Industrial Mycology, Utrecht3584 CT, Netherlands. Phone: 31 (0)30 2122600. Fax: 31 (0)30 2512097.E-mail: [email protected].
� Published ahead of print on 2 June 2010.
2754
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

including the 5.8S rDNA, and a part of the �-tubulin gene.Furthermore, the antifungal susceptibility profiles of these spe-cies and ex-type strains are reported.
MATERIALS AND METHODS
Strains. This study includes 34 strains isolated from clinical specimens andhospital environments. These strains were identified on the basis of macro- and
microscopic characters and were maintained in various culture collections as P.variotii or P. lilacinus. In additional, ex-type strains and freshly isolated strainswere also included in this study.
Morphological examination. Isolates (Table 1) were grown for 3 days on maltextract agar (MEA) and were incubated in the dark at 25, 30, and 37°C. Fur-thermore, three-point inoculations were made on MEA, Czapek yeast agar(CYA), and creatine agar (CREA); and the isolates were incubated for 7 days at25°C (the medium compositions are described by Samson et al. [43]). After
TABLE 1. Isolates used in this study
Strain no.a Species Source
CBS 100.11NT** Byssochlamys nivea UnknownCBS 146.48NT** Byssochlamys fulva Bottled fruit, UKCBS 373.70T** Byssochlamys lagunculariae Wood of Laguncularia racemosa (mangue), BrazilCBS 605.74HT** Byssochlamys verrucosa Nesting material of Leipoa ocellata, AustraliaCBS 374.70isoT** Byssochlamys zollerniae Wood of Zollernia ilicifolia and Protium heptaphyllum, BrazilUMCN V63-56, DTO 63F6 Hamigera avellanea Human, ear swab; Nijmegen, Netherlands (2007)CBS 295.48isoT* Hamigera avellanea Soil; San Antonio, TXCBS 370.70T** P. brunneolus Nonfat dry milk, CanadaCBS 110430* P. divaricatum Soil, ThailandCBS 284.48T P. divaricatum Mucilage bottle with library paste, USAUMCN V54-40, DTO 63F3 P. formosus Human, sputum; Nijmegen, Netherlands (2006)CBS 296.93 P. formosus Human, bone marrow of patient; Taskent, UzbekistanCBS 297.93 P. formosus Human, blood of patient; Taskent, UzbekistanCBS 298.93 P. formosus Human, breast milk of patient; Taskent, UzbekistanCBS 990.73BT P. formosus UnknownDTO 45H8 P. formosus Pseudo-outbreak in hospital, blood culture, United KingdomUMCN 1274, DTO 63E3 P. formosus (lecythidis type) Human, bronchoalveolar lavage fluid; Nijmegen, NetherlandsUMCN V49-58, DTO 63F1 P. formosus (lecythidis type) Human, sputum; Zwolle, Netherlands (2006)UMCN V56-25, DTO 63F4 P. formosus (lecythidis type) Human, sputum; Nijmegen, Netherlands (2006)CBS 372.70 P. formosus (lecythidis type) Type of P. lecythidis, Lecythis unsitata (Lecythidaceae), wood, BrazilDTO 45I1 P. formosus (lecythidis type) Pseudo-outbreak in hospital, blood culture; UKNCPF 2825, DTO 49D5 P. formosus (lecythidis type) Brain abscess, United Kingdom (1991)NCPF 2837, DTO 49D6 P. formosus (lecythidis type) Brain abscess, same patient as NCPF 2825, United Kingdom (1991)CBS 113247* P. formosus (maximus type) Soil, ThailandCBS 371.70 P. formosus (maximus type) Type of P. maximus, Annona squamosa, BrazilUMCN 1156,* DTO 63E1 P. lilacinus Unknown sourceUMCN 2419,* DTO 63E5 P. lilacinus Human, skin swab; Nijmegen, Netherlands (1994)CBS 284.36T* P. lilacinus Soil; Ithaca, NYV52-21,* DTO 63F2 P. lilacinus Human, sputum; Nijmegen, Netherlands (2006)UMCN V66-47, DTO 63F7 P. dactylethromorphus Human, cornea scrapings from keratomycosis; Nijmegen, Netherlands (2008)CBS 323.34T P. dactylethromorphus Unknown sourceCBS 492.84* P. dactylethromorphus Lepidium sativum, DenmarkCBS 110036, DTO 34C8 P. variotii Cerebrospinal fluid of 60-year-old female with diabetes and cancer;
Istanbul, TurkeyUMCN 1157*, DTO 63E2 P. variotii Unknown sourceUMCN 2266, DTO 63E4 P. variotii Human, feces; Nijmegen, Netherlands (1994)UMCN 3796, DTO 63E6 P. variotii Human, mouthwash; Nijmegen, Netherlands (1995)UMCN 45H9*, DTO 45H9 P. variotii Liver biopsy; London, UKUMCN 577, DTO 63D6 P. variotii Human, mouthwash; Nijmegen, NetherlandsUMCN 654, DTO 63D7 P. variotii Human, feces, Nijmegen; NetherlandsUMCN 730, DTO 63D8 P. variotii Hospital environment, elevator shaft; Nijmegen, NetherlandsUMCN 731, DTO 63D9 P. variotii Human, mouthwash; Nijmegen, NetherlandsUMCN 7845, DTO 63E7 P. variotii Human, cerebrospinal fluid; Nijmegen, Netherlands (1998)UMCN 8490, DTO 63E9 P. variotii Human, mouthwash; Nijmegen, Netherlands (1999)UMCN V57-21, DTO 63F5 P. variotii Human, abscess; Zwolle, Netherlands (2007)CBS 101075 P. variotii Type of B. spectabilis, heat-processed fruit beverage, JapanCBS 102.74T P. variotii Unknown sourceCBS 124.97 P. variotii Human, vitreous tumor left eye, neutropenic leukemic patient with acute
endophthalmitis, Hong KongCBS 339.51 P. variotii Human, sputum, NetherlandsDTO 45I3 Talaromyces eburneus Pseudo-outbreak in hospital, blood culture, UKNCPF 2801, DTO 49D4 Talaromyces eburneus Sputum, cystic fibrosis patient, UK (1991)NCPF 7594, DTO 49D7 Talaromyces eburneus Blood culture, patient with peritonitis, UK (2002)NCPF 7596, DTO 49D9 Talaromyces eburneus Peritoneal dialysis fluid (same patient source as NCPF 7594)
a Strains indicated with one asterisk are not included in the phylogenetic analysis; these strains are used only in the susceptibility tests. The strains labeled with two asterisksare included in the phylogenetic study but not in the susceptibility tests. NT, neotype; HT, holotype; isoT, isotype. CBS, culture collection of the CBS-Fungal BiodiversityCentre, Utrecht, Netherlands; UMCN, culture collection of Radboud University Nijmegen Medical Center; DTO, internal culture collection of CBS-Fungal BiodiversityCentre.
VOL. 48, 2010 IDENTIFICATION OF PAECILOMYCES VARIOTII 2755
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

incubation, the colony diameters were measured and the reactions on creatineagar recorded.
Phylogeny and molecular identification. Strains were grown on MEA (Oxoid)for 4 to 7 days at 25°C. Genomic DNA was isolated using an Ultraclean microbialDNA isolation kit (MoBio), according to the manufacturer’s instructions. Frag-ments containing ITS region 1 (ITS1) and ITS2, including 5.8S rDNA) and a partof the �-tubulin gene were amplified and subsequently sequenced and analyzedaccording to the procedure described previously (22). For parsimony analyses,PAUP (version 4.0) software was used (48) and Byssochlamys verrucosa CBS605.74 was used as the outgroup.
Antifungal susceptibility tests. The susceptibilities of the majority of thestrains listed in Table 1 were tested; exceptions were the (ex type) strains ofuncommon species in clinical environments, such as Byssochlamys nivea, B. fulva,B. verrucosa, B. lagunculariae, B. zollerniae, and P. brunneolus. Paecilomycesdivaricatus is also uncommon, but it is included to provide a representativeoverview of the susceptibility of the members of the Paecilomyces variotii com-plex. Isolates were revived by subculturing twice on Sabouraud dextrose agartubes for 5 to 7 days at 35°C. Conidial suspensions were adjusted spectrophoto-metrically and were further diluted in RPMI 1640 medium (with L-glutamine andwithout bicarbonate; Gibco BRL, Life Technologies, Woerden, Netherlands).Microtiter plates were inoculated with an initial concentration of 1 � 104 to 5 �104 conidia/ml, as recommended by the CLSI (formerly the NCCLS) for moldtesting (30).
The antifungal activities of amphotericin B (Bristol-Myers Squibb, Woerden,Netherlands), flucytosine (5FC; Valeant, Zoetermeer, Netherlands), itracon-azole (Janssen Pharmaceutica BV, Tilburg, Netherlands), voriconazole (Pfizer,Capelle aan de IJssel, Netherlands), posaconazole (Schering-Plough, Maarssen,Netherlands), terbinafine (Novartis Pharma, Arnhem, Netherlands), and caspo-fungin (Merck, Sharpe, and Dohme, Haarlem, Netherlands) were determined invitro using a broth microdilution method, according to CLSI guidelines (M38-A)(30). The concentration range for amphotericin B, terbinafine, itraconazole,voriconazole, and posaconazole was 0.016 to 16 mg/liter; a range of 0.062 to 64mg/liter was used for 5FC and caspofungin. MICs were determined after 24 and48 h of incubation. For amphotericin B and the azoles itraconazole, voriconazole,and posaconazole, the MIC was defined as the lowest concentration that showedno visible growth. For 5FC and terbinafine, the MIC was defined as the lowestconcentration at which 50% inhibition of growth compared with that of thecontrol was measured (32). For caspofungin, the minimum effective concentra-tion was determined. All susceptibility tests were performed in duplicate.
Nucleotide sequence accession numbers. The sequences newly generated inthe present study are deposited in GenBank under accession numbers GU968650to GU968703.
RESULTS
Identification. Identification of the strains was performed bycombining phenotypic characteristics and the sequences of theITS regions and part of the �-tubulin gene. The investigatedclinical strains were maintained in various collections as P.variotii or P. lilacinus. Critical examination of the culturesshowed that one Hamigera avellanea isolate and four Talaro-myces eburneus isolates were present among the isolates whichhad previously been identified as P. variotii. The ITS sequencesof three T. eburneus isolates (NCPF 7594, NCPF 7596, andDTO 45I3) were identical and had 99.8% homology with thetype strain of T. eburneus (CBS 100538). Isolate DTO 49D4was more divergent and shared 96.4% homology with the typestrain of T. eburneus and 98.8% similarity with the type strainof Geosmithia argillaea (NRRL 5177). Although this strain ismore closely related to G. argillaea, we identified this strain asT. eburneus, since both species are claimed to be conspecific(53). The T. eburneus strains were isolated from patient mate-rial in three separate cases: from the sputum of a patient withcystic fibrosis (NCPF 2801), from a blood culture (DTO 45I3),and from the peritoneal dialysis fluid and blood of a patient(NCPF 7594 and NCPF 7596). P. variotii superficially resem-bles T. eburneus in its olive brown conidial colors and thermo-
philic nature. However, it differs in growing very slowly at 25°C(attaining a diameter of between 10 and 25 mm) and having aGeosmithia anamorph. Geosmithia anamorphs are character-ized by cylindrical phialides, ornamented conidiophores andphialides, and cylindrical conidia (Fig. 1). A further presump-tive isolate of P. variotii was identified as H. avellanea. The ITSsequence of this strain had a similarity of 98.0% with the typestrain of this species (CBS 295.48). This species macroscopi-cally resembles P. variotii in many respects and also formspowdery olive brown colonies, and it has a high growth rate at25°C and 37°C. However, H. avellanea can be distinguishedfrom P. variotii by the presence of a Merimbla-type anamorph(Fig. 1). Three strains were identified as P. lilacinus, and theITS sequences of these strains have 100% homology with thetype strain of P. lilacinus (CBS 284.36).
In the present study, we have adopted the taxonomy ofPaecilomyces variotii and related species proposed by Samsonet al. (45), which uses morphological characteristics, in combi-nation with extrolite data and sequences. Combined molecularand morphological examination of the remaining P. variotiisensu lato isolates showed that three different species werepresent, namely, P. variotii, P. formosus, and P. dactylethromor-phus. P. variotii was the species encountered the most fre-quently (12), 11 isolates were identified as P. formosus, and 1isolate was identified as P. dactylethromorphus. These threespecies can be differentiated on the basis of morphologicalcriteria. P. variotii morphologically resembles P. formosus, butthe latter produces acid components on creatine agar andgrows faster at 30°C than at 37°C. P. dactylethromorphus can bedifferentiated from the other two species by its cylindricalconidia and regular branched conidiophores (Fig. 1). Figure 2shows the results of the phylogenetic analysis of the ITS andpartial tubulin sequences. Both sequenced regions gave similaridentification results, and all the species of the P. variotii com-plex can be differentiated by either their ITS or partial tubulinsequences. A high degree of variation was present in the ITSand tubulin sequences of the P. formosus isolates. Two distinctgroups, with high bootstrap support, were observed. A propor-tion of the isolates (5) formed a group together with the typestrain of P. formosus, and the other group clustered togetherwith the type strain of P. lecythidis. These two groups couldrepresent two cryptic species but were not treated as such sincethey are morphologically similar and produce the same patternof extrolites (45). Three strains (CBS 296.93, CBS 297.93, andCBS 298.93) received as P. variotii var. zaaminella and claimedto be the causal agent of zaaminellosis (11) were identified asP. formosus.
Susceptibility testing. Susceptibility data, i.e., the geometricmean (GM) of the MIC and the MIC range, are shown inTable 2. For those isolates that were not inhibited by thehighest drug concentration, the next higher concentration wasused to calculate the GM MIC; if no growth was observed inthe well with the lowest drug concentration, the next lowerconcentration was used. The activities of most of the antifungalagents differed between the seven different species. Posacon-azole and terbinafine showed good in vitro activity against allthe species tested. Posaconazole showed the lowest MICs, asall isolates except T. eburneus were inhibited by a concentra-tion of 0.25 mg/liter; T. eburneus had slightly higher MIC val-ues (MIC range, 0.25 to 1 mg/liter). All isolates (except one P.
2756 HOUBRAKEN ET AL. J. CLIN. MICROBIOL.
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
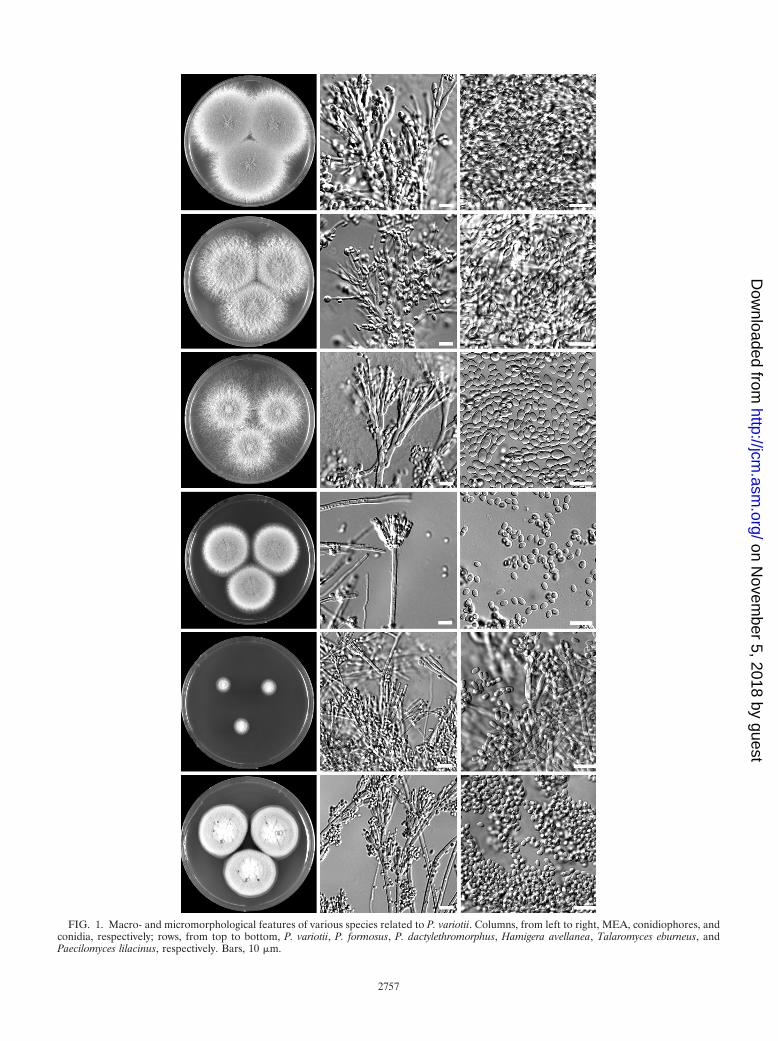
FIG. 1. Macro- and micromorphological features of various species related to P. variotii. Columns, from left to right, MEA, conidiophores, andconidia, respectively; rows, from top to bottom, P. variotii, P. formosus, P. dactylethromorphus, Hamigera avellanea, Talaromyces eburneus, andPaecilomyces lilacinus, respectively. Bars, 10 �m.
2757
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

formosus isolate) were inhibited by terbinafine at a concentra-tion of 1 mg/liter or lower. Itraconazole was the second mostactive azole, having in vitro activity against all Paecilomycesspecies except P. lilanicus and T. eburneus. The P. lilanicusisolates were not inhibited by itraconazole, which was also truefor three of four of T. eburneus isolates. H. avellanea wasmoderately susceptible to itraconazole, but only two isolateswere available for testing. Voriconazole was the least activeazole and had in vitro activity only against P. lilanicus and H.avellanea. Amphotericin B was active in vitro against all speciestested with the exception of P. lilanicus and T. eburneus. Flucy-tosine was also active against most of the species tested; theexceptions were P. lilanicus and H. avellanea. For amphotericinB, itraconazole, posaconazole, voriconazole, and flucytosine,little intraspecific variation in antifungal susceptibility wasnoted. This was not the case for caspofungin, where minimaleffective concentration values varied between 0.063 and 4 mg/liter for different isolates of the same species.
DISCUSSION
Of 32 isolates which were identified as P. variotii by theirphenotypic characteristics, 5 were shown here not to belong tothe genus Paecilomyces and instead proved to be Talaromyceseburneus (anamorph, Geosmithia argillacea) or Hamigera avel-lanea (anamorph, Merimbla ingelheimense). These two species
superficially resemble P. variotii, but the micromorphology isdistinct from that of Paecilomyces (35). The occurrence ofthese two species in clinical environments might be more com-mon than has been noted to date. Screening of the StrainInfobioportal (www.straininfo.net) identified two other Hamigeraisolates that have been reported from clinical environments.One isolate (CBS 128.90) originates from continuous ambulatoryperitoneal dialysis liquid of a dialysis patient, and the other(UAMH 2531), maintained under the anamorphic name M. in-gelheimense, was isolated from skin between the toes of a man.Multiple isolates of G. argillacea originating from bronchial wash-ings (UAMH 7717, UAMH 8639, UAMH 9714, UAMH 9854,UAMH 10232), a brain abscess (UAMH 9833), and a dissemi-nated infection in a German shepherd dog (UAMH 10932,UAMH 10933 [21]) are also present in the UAMH culturecollection. In addition, this species has recently been proposedto be a potential new pathogen that colonizes patients withcystic fibrosis lung disease (6, 19). The remaining isolates be-longed to three different Paecilomyces species. P. variotii and P.formosus predominated, although one isolate of P. dactylethro-morphus was also detected. The presence of these species inthe clinical samples might be explained by the fact that thesespecies occur more commonly in food and indoor environ-ments than the other members of the P. variotii complex (J.Houbraken, unpublished results).
FIG. 2. One of the most parsimonious trees from each of the two analyzed loci sequenced. (A) ITS1, ITS2, and 5.8S rDNA (consistency index �0.796; retention index � 0.938, rescaled consistency index � 0.747); (B) partial beta-tubulin data (consistency index � 0.745; retention index �0.909; rescaled consistency index � 0.677).
2758 HOUBRAKEN ET AL. J. CLIN. MICROBIOL.
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

Species identification of fungi in the past has primarily beenbased on morphological features. However, identificationsolely on the basis of morphology appears to be difficult, andtrained staff is required for correct identification. Identificationof fungi from clinical samples might even encounter the prob-lem that isolates grow atypically on inappropriate agars orbecome atypical if antimycotics are used (31). Therefore, mo-lecular-based methods, such as sequencing, appear to be morereliable and are a robust alternative to discriminate fungalspecies (4, 9, 17, 39). Sequencing data are objective and fast,and reliable identification of uncommon species can be ob-tained. The ITS regions are recommended for use for identi-fication of species in a clinical setting, since they are easy toamplify and large data sets are present in various databases,such as GenBank and European Molecular Biology Labora-tory Nucleotide Sequence Database. These databases will ex-pand dramatically in the near future since the ITS region hasbecome the prime bar coding region (5, 46; U. Eberhardt,personal communication). The disadvantage of the ITS regionis that it does not have sufficient discriminatory power invarious genera, for example, the genera Aspergillus, Penicil-lium, and Fusarium (5, 18, 33, 47). In this study, the ITSregions and part of the �-tubulin gene were used, and bothloci were shown to exhibit sufficient interspecific variationfor identification purposes.
Paecilomyces variotii is a commonly occurring species andhas previously been isolated from various substrates. Immuno-suppression is the critical risk factor for infection; and cases ofpneumonia (7), sinusitis (13, 34, 50), endophthalmitis (25, 49),otitis media (12), wound infection in a transplant recipient(26), cutaneous hyalohyphomycoses (3, 29), onychomycosis(2), osteomyelitis in a patient with granulomatous disorder(10), and dialysis-related peritonitis (38) have all been re-ported to be caused by this fungus. This species can be con-sidered extremotolerant and is able to grow at high tempera-tures, on decaying wood, and on creosote treated woodenutility poles (E. de Meyere et al., unpublished data). The abilityto grow on creosote-treated wooden poles suggests that thisspecies is able to break down aromatics and is able to growunder stressful conditions (very hot conditions, dry conditions,conditions very low in micronutrients). Additionally, P. formo-sus has also been isolated from toluene gas biofilters. However,these isolates were misidentified as P. sinensis and P. variotii,and the correct name for these isolates is P. formosus (14, 16,37). The extremotolerant nature is suggested to contribute tothe pathogenic potential of fungi. Prenafeta-Boldu et al. (37)speculated that there might be a link between neurotropismand assimilation of aromatic substrates, and this might be oneof the factors that enable fungi to grow in the human brain,with its unique chemical properties. This suggested link is alsofound in our study, as we have also encountered four strains ofthree independent cases originating from brain or cerebrospi-nal fluid.
Correlations of species identities with susceptibility profiles.Major differences in in vitro antifungal susceptibility profileswere found between the investigated species. In general, vori-conazole is not active against members of the Paecilomycesvariotii complex but is active against P. lilanicus and H. avella-nea. Treatment of infections due to P. lilanicus may be com-plicated, as amphotericin B also showed no activity in vitro.
TA
BL
E2.
Susceptibilityresults
forP
aecilomyces
species,Ham
igeraavellanea,and
Talarom
cyeseburneus
strains,byspecies
andantifungalagent
SpeciesN
o.ofisolates
Geom
etricm
ean(range)
MIC
(mg/liter) a
AM
B5F
CIT
ZV
CZ
POS
TB
CA
S
P.variotii
160.11
(0.03–0.5)0.03
0.04(0.008–4)
11.3(1–32)
0.02(0.008–0.125)
0.08(0.031–0.5)
0.52(0.063–4)
P.lilanicus
432
1285.7
(0.063–32)0.15
(0.063–0.25)0.2
(0.008–0.25)0.04
(0.031–0.063)0.59
(0.5–1)P
.formosus
140.13
(0.063–0.25)0.04
(0.031–0.25)0.13
(0.063–1)26.25
(16–32)0.08
(0.031–0.25)0.12
(0.031–2)1.16
(0.063–4)P
.dactylethromorphus
30.16
(0.125–0.25)0.05
(0.031–0.125)0.08
(0.031–0.125)32
0.030.31
(0.125–1)0.25
(0.125–0.5)P
.divaricatus2
0.25(0.125–0.5)
0.0080.5
(0.25–1)32
0.18(0.125–0.25)
0.06(0.031–0.125)
0.71(0.25–2)
H.avellanea
20.5
2(1–4)
0.0310.09
(0.063–0.125)0.031
0.04(0.031–0.063)
0.063T
.eburneus4
3.3(1–8)
0.03212.5
(1–32)28
(16–32)0.69
(0.25–1)0.032
0.31(0.25–0.5)
aF
orcaspofungin,the
minim
umeffective
concentrationw
asdeterm
ined.AM
B,am
photericinB
;5FC
,flucytosine;ITZ
,itraconazole;VC
Z,voriconazole;PO
S,posaconazole;TB
,terbinafine;CA
S,caspofungin.
VOL. 48, 2010 IDENTIFICATION OF PAECILOMYCES VARIOTII 2759
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

Posaconazole may be the only appropriate alternative agent,although the lack of an intravenous formulation and limitedpenetration into the cerebrospinal fluid might limit its use.Amphotericin B showed good activity against all other speciestested, as was also the case for flucytosine. The combination ofamphotericin B and flucytosine may therefore be an option incomplicated infections due to Paecilomyces species other thanP. lilanicus. Flucytosine was recently shown to be active in vitroand in vivo against A. fumigatus, with the MIC measured at pH5.0 being found to correlate better with the outcome in amurine model of disseminated aspergillosis than that deter-mined at pH 7.0 (51). It would be of interest to determine theactivity of flucytosine at pH 5.0 against other molds, includingPaecilomyces species. As published previously, terbinafine alsoshows potent activity against all species tested (8). However,the clinical use of this drug for the treatment of invasive fungalinfections remains limited due to its pharmacological proper-ties. The role of the echinocandins remains unclear, as the invitro activity of caspofungin was variable, with the MIC rangeswithin species being broad. This indicates that the activity ofthe drug is not easily predictable, thereby precluding a prom-inent role in the first-line therapy of Paecilomyces infections. Ingeneral, the antifungal susceptibility profiles of P. variotii, P.formosus, P. dactylethromorphus, and P. divaricatus appeared tobe similar, although a limited number of species have beentested. The profiles of P. lilanicus, T. eburneus, and H. avellaneaare different. This is in agreement with the phylogeny, since T.eburneus and H. avellanea are not related to Paecilomyces orByssochlamys (45, 53) and P. lilacinus will shortly be accom-modated in a new genus because it is only distantly related toP. variotii and the other species hitherto placed in the genusPaecilomyces (27, 28). In summary, it is clear that correct spe-cies identification of Paecilomyces isolates is important to helpguide appropriate antifungal therapy. The correlation betweenthe in vitro activity and the in vivo efficacy of these agentsagainst Paecilomyces species remains to be investigated fur-ther.
ACKNOWLEDGMENT
Andrew Borman thanks Elizabeth Johnson for her interest in thestudy and permitting him to collaborate in this study.
REFERENCES
1. Ajello, L. 1986. Hyalohyphomycosis and phaeohyphomycosis: two globaldisease entities of public health importance. Eur. J. Epidemiol. 2:243–251.
2. Arenas, R., M. Arce, H. Munoz, and J. Ruiz-Esmenjaud. 1998. Onychomy-cosis due to Paecilomyces variotii. Case report and review. J. Mycol. Med.8:32–33.
3. Athar, M. A., A. S. Sekhon, J. V. Mcgrath, and R. M. Malone. 1996. Hyalo-hyphomycosis caused by Paecilomyces variotii in an obstetrical patient. Eur.J. Epidemiol. 12:33–35.
4. Balajee, S. A., J. Gribskov, M. Brandt, J. Ito, A. Fothergill, and K. A. Marr.2005. Mistaken identity: Neosartorya pseudofischeri and its anamorph mas-querading as Aspergillus fumigatus. J. Clin. Microbiol. 43:5996–5999.
5. Balajee, S. A., A. M. Borman, M. E. Brandt, J. Cano, M. Cuenca-Estrella, E.Dannaoui, J. Guarro, G. Haase, C. C. Kibbler, W. Meyer, K. O’Donnell,C. A. Petti, J. L. Rodriguez-Tudela, D. Sutton, A. Velegraki, and B. L.Wickes. 2009. Sequence-based identification of Aspergillus, Fusarium andthe Mucorales in the clinical mycology laboratory: where are we and whereshould we go from here? J. Clin. Microbiol. 47:877–884.
6. Barton, R. C., A. M. Borman, E. M. Johnson, J. Houbraken, R. P. Hobson,M. Denton, S. P. Conway, K. G. Brownlee, D. Peckham, and T. W. R. Lee. 26April 2010, posting date. Isolation of the fungus Geosmithia argillacea in thesputum of people with cystic fibrosis. J. Clin. Microbiol. doi:10.1128/JCM.
7. Byrd, R. P., Jr., T. M. Roy, C. L. Fields, and J. A. Lynch. 1992. Paecilomycesvariotii pneumonia in a patient with diabetes mellitus. J. Diabetes Complicat.6:150–153.
8. Castelli, M. V., A. Alastruey-Izquierdo, I. Cuesta, A. Monzon, E. Mellado,J. L. Rodriguez-Tudela, and M. Cuenca-Estrella. 2008. Susceptibility testingand molecular classification of Paecilomyces spp. Antimicrob. Agents Che-mother. 52:2926–2928.
9. Clinical and Laboratory Standards Institute. 2008. Interpretive criteria foridentification of bacteria and fungi by DNA target sequencing: guideline.CLSI document MM18-A. Clinical and Laboratory Standards Institute,Wayne, PA.
10. Cohen-Abbo, A., and K. M. Edwards. 1995. Multifocal osteomyelitis causedby Paecilomyces variotii in a patient with chronic granulomatous disease.Infection 23:55–57.
11. Dekhkan-Khodzhayeva, N. 1992. Newly identified fungal disease zaaminel-losis. Meditsinskaya Gazeta 3:8–9.
12. Dhindsa, M. K., J. Naidu, S. M. Singh, and S. K. Jain. 1995. Chronicsuppurative otitis media caused by Paecilomyces variotii. J. Med. Vet. Mycol.33:59–61.
13. Eloy, P., B. Bertrand, P. Rombeaux, M. Delos, and J. P. Trigaux. 1997.Mycotic sinusitis. Acta Otorhinolaryngol. Belg. 51:339–352.
14. Estevez, E., M. C. Veiga, and C. Kennes. 2005. Biodegradation of toluene bythe new fungal isolates Paecilomyces variotii and Exophiala oligosperma.J. Ind. Microbiol. Biotechnol. 32:33–37.
15. Fagerburg, R., B. Suh, H. R. Buckley, B. Lorber, and J. Karian. 1981.Cerebrospinal fluid shunt colonization and obstruction by Paecilomyces vari-otii. J. Neurosurg. 54:257–260.
16. Garcia-Pena, I., S. Hernandez, R. Auria, and S. Revah. 2005. Correlation ofbiological activity and reactor performance in biofiltration of toluene withthe fungus Paecilomyces variotii CBS 115145. Appl. Environ. Microbiol.71:4280–4285.
17. Geiser, D. M., J. I. Pitt, and J. W. Taylor. 1998. Cryptic speciation andrecombination in the aflatoxin-producing fungus Aspergillus flavus. Proc.Natl. Acad. Sci. U. S. A. 95:388–393.
18. Geiser, D. M., M. A. Klich, J. C. Frisvad, S. W. Peterson, J. Varga, and R. A.Samson. 2007. The current status of species recognition and identification inAspergillus. Stud. Mycol. 59:1–10.
19. Giraud, S., M. Pihet, B. Razafimandimby, J. Carrere, N. Degand, L. Mely, L.Favennec, E. Dannaoui, J.-P. Bouchara, and A. Calenda. 12 May 2010,posting date. Geosmithia argillacea: an emerging pathogen in cystic fibrosispatients? J. Clin. Microbiol. doi:10.1128/JCM.
20. Groll, A. H., and T. J. Walsh. 2001. Uncommon opportunistic fungi: newnosocomial threats. Clin. Microbiol. Infect. 7(Suppl. 2):8–24.
21. Grant, D. C., D. A. Sutton, A. Sandberg, R. D. Tyler, E. H. Thompson, A. M.Romanelli, and B. L. Wickes. 2009. Disseminated Geosmithia argillacea in-fection in a German shepherd dog. Med. Mycol. 47:221–226.
22. Houbraken, J., M. Due, J. Varga, M. Meijer, J. C. Frisvad, and R. A.Samson. 2007. Polyphasic taxonomy of Aspergillus section Usti. Stud. Mycol.59:107–128.
23. Houbraken, J., J. Varga, E. Rico-Munoz, S. Johnson, and R. Samson. 2008.Sexual reproduction as the cause of heat resistance in the food spoilagefungus Byssochlamys spectabilis (anamorph Paecilomyces variotii). Appl. En-viron. Microbiol. 74:1613–1619.
24. Kantarcioglu, A. S., G. Hatemi, A. Yucel, G. S. De Hoog, and N. M. Mandel.2003. Paecilomyces variotii central nervous system infection in a patient withcancer. Mycoses 46:45–50.
25. Lam, D. S., A. P. Koehler, D. S. Fan, W. Cheuk, A. T. Leung, and J. S. Ng.1999. Endogenous fungal endophthalmitis caused by Paecilomyces variotii.Eye 13:113–116.
26. Lee, J., W. W. Yew, C. S. Chiu, P. C. Wong, C. F. Wong, and E. P. J. Wang.2002. Delayed sternotomy wound infection due to Paecilomyces variotii in alung transplant recipient. Heart Lung Transplant. 21:1131–1134.
27. Luangsa-ard, J. J., N. L. Hywel-Jones, and R. A. Samson. 2004. Thepolyphyletic nature of Paecilomyces sensu lato based on 18S-generatedrDNA phylogeny. Mycologia 96:773–780.
28. Luangsa-Ard, J., N. L. Hywel-Jones, L. Manoch, and R. A. Samson. 2005. Onthe relationships of Paecilomyces sect. Isarioidea species. Mycol. Res. 109:581–589.
29. Naidu, J., and S. M. Singh. 1992. Hyalohyphomycosis caused by Paecilomy-ces variotii: a case report, animal pathogenicity and ‘in vitro’ sensitivity.Antonie Van Leeuwenhoek 62:225–230.
30. National Committee for Clinical Laboratory Standards. 2002. Referencemethod for broth dilution antifungal susceptibility testing of filamentousfungi. Approved standard M38-A. National Committee for Clinical Labora-tory Standards, Wayne, PA.
31. Nosanchuk, J. D., W. Cleare, S. P. Franzot, and A. Casadevall. 1999. Am-photericin B and fluconazole affect cellular charge, macrophage phagocyto-sis, and cellular morphology of Cryptococcus neoformans at subinhibitoryconcentrations. Antimicrob. Agents Chemother. 43:233–239.
32. Odds, F. C., F. van Gerven, A. Espinel-Ingroff, M. S. Bartlett, M. A. Ghan-noum, M. V. Lancaster, M. A. Pfaller, J. H. Rex, M. G. Rinaldi, and T. J.Walsh. 1998. Evaluation of possible correlations between antifungal suscep-tibilities of filamentous fungi in vitro and antifungal treatment outcomes inanimal infection models. Antimicrob. Agents Chemother. 42:282–288.
33. O’Donnell, K., D. A. Sutton, A. Fothergill, D. McCarthy, M. G. Rinaldi,
2760 HOUBRAKEN ET AL. J. CLIN. MICROBIOL.
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

M. E. Brandt, N. Zhang, and D. M. Geiser. 2008. Molecular phylogeneticdiversity, multilocus haplotype nomenclature, and in vitro antifungal resis-tance within the Fusarium solani species complex. J. Clin. Microbiol. 46:2477–2490.
34. Otcenasek, M., Z. Jirousek, Z. Nozicka, and K. Mencl. 1984. Paecilomycosisof the maxillary sinus. Mykosen 27:242–251.
35. Pitt, J. I., and A. D. Hocking. 1979. Merimbla gen. nov. for the anamorphicstate of Talaromyces avellaneus. Can. J. Bot. 57:2394–2398.
36. Pitt, J. I., and A. D. Hocking. 2009. Fungi and food spoilage, 3rd ed.Springer, Berlin, Germany.
37. Prenafeta-Boldu, F. X., R. Summerbell, and G. Sybren de Hoog. 2006. Fungigrowing on aromatic hydrocarbons: biotechnology’s unexpected encounterwith biohazard? FEMS Microbiol. Rev. 30:109–130.
38. Rinaldi, S., E. Fiscarelli, and G. Rizzoni. 2000. Paecilomyces variotii perito-nitis in an infant on automated peritoneal dialysis. Pediatr. Nephrol. 14:365–366.
39. Rinyu, E., J. Varga, and L. Ferenczy. 1995. Phenotypic and genotypic analysisof variability in Aspergillus fumigatus. J. Clin. Microbiol. 33:2567–2575.
40. Salle, V., E. Lecuyer, T. Chouaki, F. X. Lescure, A. Smail, A. Vaidie, C.Dayen, J. L. Schmit, J. P. Ducroix, and Y. Douadi. 2005. Paecilomyces variotiifungemia in a patient with multiple myeloma: case report and literaturereview. J. Infect. 51:e93–e95.
41. Samson, R. A. 1974. Paecilomyces and some allied hyphomycetes. Stud.Mycol. 6:1–119.
42. Samson, R. A., J. Houbraken, R. C. Summerbell, B. Flannigan, and J. D.Miller. 2001. Common and important species of fungi and actinomycetes inindoor environment, p. 287–474. In B. Flannigan, R. A. Samson, and J.Miller (ed.), Microorganisms in home and indoor work environments. CRCPress LLC, Boca Raton, FL.
43. Samson, R. A., E. S. Hoekstra, and J. C. Frisvad. 2004. Introduction to food-
and airborne fungi, 7th ed. Centraalbureau voor Schimmelcultures, Utrecht,Netherlands.
44. Reference deleted.45. Samson, R. A., J. Houbraken, J. Varga, and J. C. Frisvad. 2009. Polyphasic
taxonomy of the heat resistant ascomycete genus Byssochlamys and its Pae-cilomyces anamorphs. Persoonia 22:14–27.
46. Seifert, K. A. 2009. Barcoding fungi. Progress towards DNA barcoding offungi. Mol. Ecol. Resource 9:83–89.
47. Skouboe, P., J. C. Frisvad, J. W. Taylor, D. Lauritsen, M. Boysen, and L.Rossen. 1999. Phylogenetic analysis of nucleotide sequences from the ITSregion of terverticillate Penicillium species. Mycol. Res. 103:873–881.
48. Swofford, D. L. 2000. PAUP*: phylogenetic analysis using parsimony (*andother methods), version 4.0b4a. Sinauer Associates, Sunderland, MA.
49. Tarkkanen, A., V. Raivio, V. J. Anttila, P. Tommila, R. Ralli, L. Merenmies,and I. Immonen. 2004. Fungal endophthalmitis caused by Paecilomyces vari-otii following cataract surgery: a presumed operating room air-conditioningsystem contamination. Acta Ophthalmol. Scand. 82:232–235.
50. Thompson, R. F., R. B. Bode, J. C. Rhodes, and J. L. Gluckman. 1988.Paecilomyces variotii. An unusual cause of isolated sphenoid sinusitis. Arch.Otolaryngol. Head Neck Surg. 114:567–569.
51. Verweij, P. E., D. T. A. Te Dorsthorst, W. H. P. Janssen, J. F. G. M. Meis,and J. W. Mouton. 2008. In vitro activity at pH 5.0 and pH 7.0 and in vivoefficacy of flucytosine against Aspergillus fumigatus. Antimicrob. Agents Che-mother. 52:4483–4485.
52. Walsh, T. J., A. Groll, J. Hiemenz, R. Fleming, E. Roilides, and E. Anaissie.2004. Infections due to emerging and uncommon medically important fungalpathogens. Clin. Microbiol. Infect. 10:48–66.
53. Yaghuchi, T., S. Udagawa, and K. Nishimura. 2005. Geosmithia argillacea isthe anamorph of Talaromyces eburneus as a heat resistant fungus. Crypto-gamie Mycol. 26:133–141.
VOL. 48, 2010 IDENTIFICATION OF PAECILOMYCES VARIOTII 2761
on Novem
ber 5, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from